
Aprovechar al máximo la vida: la elección del medio ambiente durante la cría mejora la capacidad de las gallinas ponedoras para aprovechar las oportunidades

Aprovechar al máximo la vida: la elección del medio ambiente durante la cría mejora la capacidad de las gallinas ponedoras para aprovechar las oportunidades
 Lena Skånberg1*
Lena Skånberg1*  Regine V. Holt2
Regine V. Holt2  Ruth C. Newberry2
Ruth C. Newberry2  Inma Estévez3,4 Kirste McCrea1
Inma Estévez3,4 Kirste McCrea1  Linda J. Keeling1
Linda J. Keeling1- 1Departamento de Ciencia Aplicada y Bienestar Animal, Universidad Sueca de Ciencias Agrícolas, Uppsala, Suecia
- 2Departamento de Ciencias Animales y Acuícolas, Facultad de Biociencias, Universidad Noruega de Ciencias de la Vida, Ås, Noruega
- 3Neiker Instituto Vasco de Investigación Agraria, Alianza Vasca de Investigación y Tecnología, Vitoria, España
- 4IKERBASQUE Fundación Vasca para la Ciencia, Bilbao, España
Introducción: El potencial del alojamiento en aviario para mejorar el bienestar de las gallinas ponedoras (Gallus gallus domesticus) se verá limitado si las condiciones de cría limitan la capacidad conductual de las gallinas para aprovechar las oportunidades. Incorporando teorías sobre la plasticidad del desarrollo y la agencia animal, este estudio tuvo como objetivo determinar: (1) si la elección de los tipos de camada y percha durante la cría promovería cambios duraderos en el uso de nuevos lugares y recursos, y (2) la influencia de la provisión de tiempo de elección.
Métodos: Los polluelos de gallinas ponedoras fueron asignados a un ambiente de «opción única» (un tipo de camada y percha) o «opción múltiple» (cuatro tipos de camada y percha) durante la cría «temprana» (día 1-semana 4) y «tardía» (semana 5-15). Los ambientes se cambiaron en la mitad de los 16 corrales en la semana 5, lo que resultó en un diseño factorial de 2 × 2 con cuatro combinaciones de ambiente por período. La asignación de espacio para perchas y camadas fue la misma en todas las combinaciones de tratamientos. En la semana 16, todos los grupos fueron trasladados a corrales de puesta de aviarios estándar (período de puesta, semanas 16-27).
Resultados: Cuando se trasladaron por primera vez a los corrales de ponedora, las gallinas con opción múltiple en uno o ambos períodos de cría se extendieron más rápido en su corral que las gallinas con opción única durante la cría. La opción múltiple en la cría temprana también redujo la latencia para usar nuevas estructuras elevadas (perchas y nidos) en los corrales de puesta. La opción múltiple durante la cría tardía aumentó el éxito en la búsqueda y el consumo de gusanos de la harina ocultos (probado en las semanas 9-17) y aumentó la proporción de huevos puestos en bandejas de anidación elevadas. Numéricamente, las gallinas que cambiaron de opción múltiple a opción única en la semana 5 utilizaron el rango al aire libre menos que las gallinas que cambiaron de opción única a opción múltiple.
Discusión: Estos resultados apoyan la hipótesis de que ofrecer múltiples opciones de recursos durante la cría mejora la capacidad de las gallinas para aprovechar al máximo las nuevas oportunidades al ser más proactivas en la exploración y explotación de los nuevos recursos disponibles. En diferentes desafíos de oportunidad, las gallinas mostraron resultados positivos en respuesta a la elección durante las etapas temprana, tardía o ambas de la cría, lo que sugiere que se pueden obtener mejores resultados ofreciendo opciones ambientales durante la cría.
1 Introducción
En la naturaleza, una alta motivación para explorar y un bajo miedo a la novedad durante los primeros años de vida pueden ser esenciales para aprender a identificar los alimentos adecuados y localizar otros recursos esenciales (1-3). Se propone que la exploración, que implica el acercamiento a las fuentes de estimulación en el entorno, está motivada por el sistema emocional primario de «búsqueda» (4) y conduce a experimentar recompensas que promueven el aprendizaje asociativo (5). La exploración puede ir acompañada de neofilia, la preferencia o atracción por la novedad, que se sugiere como un rasgo independiente de la neofobia, la evitación de estímulos desconocidos (6). Una mayor neofilia, especialmente hacia los posibles alimentos, puede ser un buen indicador de éxito en un hábitat nuevo (7, 8). Sin embargo, la búsqueda y explotación de recursos novedosos conlleva riesgos. Por ejemplo, buscar alimento en un lugar alejado del resto del grupo puede aumentar el riesgo de depredación (9), y existe el riesgo de que un nuevo alimento pueda ser venenoso. La relación costo-beneficio entre la exploración y la neofobia en las poblaciones silvestres está influenciada no solo por el riesgo del medio ambiente (por ejemplo, la presión de los depredadores), sino también por la complejidad del hábitat, siendo la recopilación de información probablemente más importante en entornos más variados (2, 10, 11).
Una mayor exploración cuando es joven puede promover la plasticidad del desarrollo adaptativo del estado somático (12) y conducir a una mayor motivación para recopilar información a lo largo de la vida, promoviendo tanto la acción impulsada como el desarrollo de competencias (11). Se cree que la flexibilidad conductual resultante es adaptativa al permitir que el individuo joven se adapte mejor a su entorno adulto (12, 13). Si bien los riesgos de depredación y consumo de alimentos venenosos suelen ser menores para los animales domésticos y la fauna silvestre en cautividad que para los animales salvajes, el desarrollo de la capacidad de acción también podría ser importante para ellos, especialmente si están destinados a ser mantenidos en complejos sistemas de alojamiento para adultos o se les da acceso al aire libre. Los animales mantenidos para uso humano a menudo están expuestos a diferentes fuentes de novedad a lo largo de su vida, como nuevos alimentos, rutinas y entornos de alojamiento. Deben estar motivados para explorar, atraídos por recursos novedosos y ser flexibles desde el punto de vista conductual para aprovechar al máximo las nuevas oportunidades (14, 15).
Un entorno de cría más complejo para los animales de granja, como las gallinas ponedoras (Gallus gallus domesticus), podría fomentar la recopilación de información y promover el desarrollo de habilidades de navegación espacial (16-18), así como el aprendizaje espacial y la resolución de problemas en general (19-21). En otras palabras, la cría en un entorno complejo podría aumentar la probabilidad de que las gallinas ponedoras sean lo suficientemente flexibles en cuanto a su comportamiento como para aprovechar al máximo las oportunidades que se les ofrecen más adelante en la vida, contribuyendo así a un bienestar positivo a largo plazo. En las configuraciones experimentales, la complejidad a menudo aumenta al agregar recursos y comparar el resultado con el de un entorno más estéril que carece de estos recursos [(p. ej., 20-22)]. En el caso de las gallinas ponedoras, esto puede implicar proporcionar cama, perchas, refugios, objetos y estímulos sensoriales (23). Tales entradas durante la cría pueden dar lugar a que las gallinas ponedoras exhiban una mayor exploración de objetos nuevos (24) y un mayor uso del rango (23). En la naturaleza, sin embargo, un entorno más complejo también implica una mayor variación dentro del mismo tipo de recurso general, como tener acceso a múltiples tipos de alimentos y múltiples opciones de sustratos para alimentarse, posarse y anidar, en lugar de un solo tipo de cada uno. Proporcionar a las gallinas ponedoras una variación en los recursos relevantes cuando son jóvenes podría aumentar la relevancia biológica del medio ambiente, estimulando así la aprovechamiento de oportunidades más adelante en la vida más allá de lo que se observa típicamente en los entornos de producción actuales.
Los polluelos prefieren diferentes formas de recursos, como la cama y las perchas, para diferentes tipos de comportamiento desde su primera semana después de la eclosión (25). Por lo tanto, la presentación de variantes de recursos en el entorno doméstico tiene el potencial de conducir a una mayor activación del sistema emocional de búsqueda (26), promoviendo la formación de asociaciones positivas a través de la elección y el uso de diferentes variantes de recursos, o microhábitats a lo largo de gradientes ambientales, para diferentes propósitos (5). De hecho, cuando se les proporcionaron cuatro tipos diferentes de camada y percha durante la cría temprana, los polluelos fueron más capaces de localizar un nuevo recurso alimenticio a las 3 semanas de edad (27), mostraron un mayor nivel de locomoción en un entorno desconocido, fueron mejores para resolver una tarea espacial a las 5 semanas de edad (19) y también mostraron una mayor prevalencia de comportamientos de valencia positiva (28), en comparación con los pollitos de un entorno de cría estándar que proporciona solo un tipo de camada y un tipo de percha. Parece que la promoción de la «búsqueda» y la experiencia de tener opciones puede aumentar el uso de recursos novedosos en comparación con lo observado con la misma cantidad de recursos pero sin la variedad.
Las primeras semanas de vida parecen ser críticas para el desarrollo de un repertorio conductual que funcione bien (p. ej., buscar comida, bañarse en el polvo, posarse) y pueden afectar rasgos como el miedo y la capacidad espacial a largo plazo (29-32). Sin embargo, las pollitas jóvenes destinadas a sistemas de varios niveles a menudo se encierran en las filas de aviarios de la instalación de cría durante estas primeras semanas de vida, antes de que se les dé acceso completo al complejo espacio del aviario, incluido el piso de la cama y los múltiples niveles (24, 33). Esta práctica garantiza que puedan encontrar fácilmente recursos esenciales como alimentos y agua que solo se proporcionan en los niveles de aviario. Sin embargo, la falta de experiencia con otros recursos podría suprimir el desarrollo conductual durante una ventana sensible (34), con repercusiones a largo plazo para el uso posterior de nuevos recursos, como alimentos, tipos de camada, diseños de perchas, nidos y áreas de distribución al aire libre.
El objetivo del presente estudio fue investigar los efectos a largo plazo de la cría de gallinas ponedoras en un ambiente con mayor variación, en este caso un ambiente con una variedad de tipos de cama y percas. Basándonos en el valor adaptativo de la plasticidad del desarrollo (12) y la agencia (11), planteamos la hipótesis de que el acceso a un entorno que ofrezca múltiples opciones durante la cría mejoraría la flexibilidad conductual de las gallinas para aprovechar al máximo las nuevas oportunidades. Por lo tanto, predijimos que el comportamiento exploratorio y el uso de nuevos recursos se promoverían al experimentar una mayor elección ambiental durante la cría. Además, basándonos en un hipotético período sensible temprano, predijimos que las gallinas que tenían múltiples opciones ambientales durante las primeras 4 semanas de cría mostrarían respuestas a nuevas oportunidades indicativas de tener una mayor motivación exploratoria, mayor neofilia, menor neofobia y una mayor flexibilidad de comportamiento que las gallinas que solo estaban expuestas a múltiples opciones ambientales más adelante durante la cría. Dado que las respuestas a nuevos alimentos, nuevos objetos y entornos novedosos no siempre están correlacionadas (6, 7, 35), evaluamos las respuestas de las gallinas ponedoras a diferentes fuentes de novedad y a las oportunidades que es probable que encuentren en condiciones comerciales a edades relevantes.
2 Materiales y métodos
2.1 Ética
Este estudio fue aprobado por el Comité Nacional de Ética para Experimentos con Animales en Uppsala, Suecia (código de protocolo 5.8.18-11,549/2017, 28 de julio de 2017).
2.2 Animales y alojamiento
Los pollitos de un día (n = 364) del híbrido de capa blanca Bovans Robust fueron llevados desde una planta de incubación comercial a las instalaciones de cría en el Centro Sueco de Investigación Ganadera en Uppsala. Se marcaron con anillos de patas numerados individualmente, se pesaron y se asignaron a uno de los 16 corrales (240 × 120 × 180 cm) en grupos iniciales de 22 o 23 de manera que se asegurara un peso promedio similar del ave y uniformidad en cada corral. Los anillos de las piernas fueron reemplazados por etiquetas de ala numeradas individualmente (Jiffy bands, National Band and Tag Company, Newport, KY, Estados Unidos) el día 17. Las aves se mantuvieron en el mismo corral desde el día 1 hasta la semana 16, cuando cada grupo fue reubicado en un corral de puesta en otro edificio. Cinco polluelos murieron o fueron sacrificados por diversas razones antes de comenzar las observaciones de comportamiento, lo que llevó a 23 aves en 8 corrales, 22 aves en 7 corrales y 21 aves en un corral durante todo el estudio. Se proporcionó ad libitum alimento comercial estándar (iniciador desde el día 1, engorde a partir de la semana 7, ponedora a partir de la semana 16) y agua. La temperatura, el ciclo de iluminación y la intensidad de la luz siguieron las recomendaciones de los criadores [Hendrix Genetics (36)]. Todos los días incluían un período de 15 minutos al amanecer y al atardecer durante el ciclo diario de luz: oscuridad. En los corrales de cría, la intensidad media de la luz fue de 18 lux a nivel de las aves (rango: 7-37 lux), mientras que en los corrales de puesta fue de 5 lux a nivel de las aves (rango: 3-8 lux). Las paredes de los corrales, tanto en el corral de cría como en el de puesta, se cubrieron para bloquear visualmente a las aves vecinas. Las aves fueron revisadas diariamente durante el cuidado rutinario de los animales.
Durante las primeras 4 semanas (período de cría temprana), la cama se presentó en bandejas de plástico poco profundas (71 × 35 × 3,5 cm) así como en el piso de concreto. Se colocó una lámpara de calor en el centro del corral (centrada sobre las bandejas). Las perchas (120 cm de largo) tenían inicialmente 15 cm de altura y luego se elevaron a las 3 semanas a 45 cm y en la semana 5 a 55 cm. A partir de la semana 5 (período de cría tardía), la cama se proporcionó solo en cajas de plástico (55 litros, 78 × 56 × 18 cm) en los mismos lugares que las bandejas de plástico anteriores. En la semana 16, todos los grupos fueron trasladados a una nave de puesta (período de puesta) y mantenidos en 16 corrales (362 × 356 × 297 cm) con un área de cama (132 × 356 cm) y un área de listones de plástico elevada (32 cm) (230 × 356 cm). Todos los corrales de puesta tenían la misma cama (gránulos de paja triturada), cuatro perchas de plástico en forma de hongo (alturas 43, 96, 149 y 205 cm), una sola percha de madera de forma cuadrada (altura 187 cm), tres bandejas de nido de plástico abiertas elevadas (71 × 35 × 3,5 cm, a las alturas 70, 155 y 220 cm), y dos nidos de colonia cerrados con una puerta de cortina de vinilo y una alfombra de césped de plástico (115 × 46 × 30 cm) ubicados en el área del piso de listones. El experimento terminó a las 27 semanas, cuando las aves fueron donadas a criadores de aves de corral aficionados. El experimento se llevó a cabo entre los 1c de octubre de 2019 y el 5ésimo de abril de 2020.
2.3 Tratamientos de elección ambiental
Comparamos dos ambientes (de elección única y de opción múltiple) durante dos períodos: crianza temprana (día 1-semana 4) y tardía (semana 5-15). En todo momento, la mitad de los corrales de cría fueron de opción única y la otra mitad de opción múltiple. En la semana 5, la mitad de los corrales se cambiaron al otro entorno, lo que dio como resultado cuatro combinaciones de tratamiento de elección por período: Simple*Simple, Simple*Multi, Multi*Single y Multi*Multi (Figura 1).
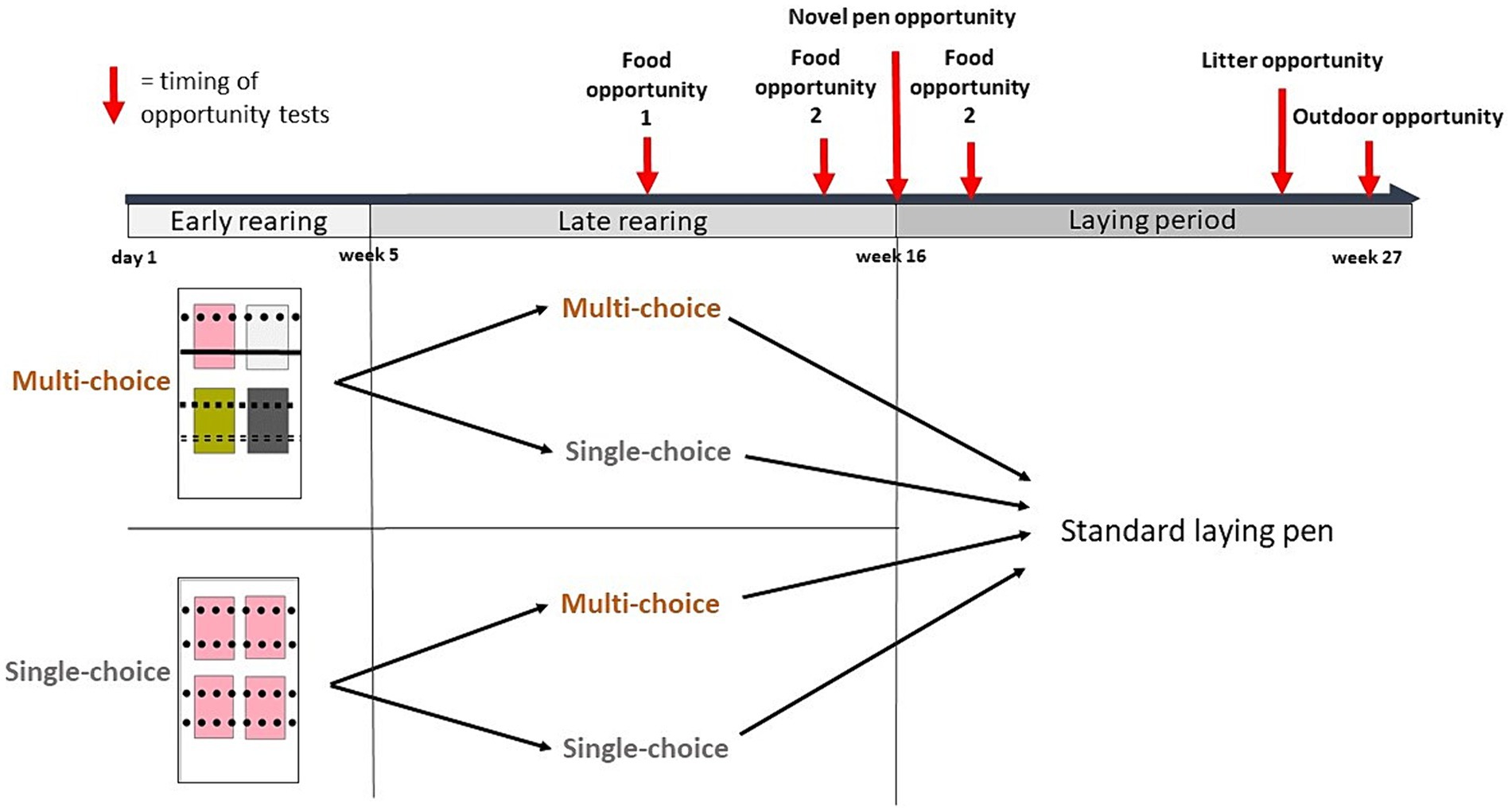 Figura 1. Se proporcionaron dos ambientes, de elección única y de opción múltiple, durante dos períodos de cría, temprano (día 1 a la semana 4) y tardío (semana 5-15), lo que resultó en cuatro tratamientos de elección durante la cría. Los rectángulos ilustran un corral de muestra de cada entorno, con cuadrados de colores que representan los tipos de camada y diferentes estilos de línea que representan los tipos de percas. En la semana 16, las aves de cada corral de cría se trasladaron a un corral de puesta estándar. Se investigaron las respuestas conductuales a diferentes oportunidades de recursos en diferentes pruebas o contextos: oportunidades de comida (en un campo de prueba durante las semanas 9-10 y en los corrales domésticos en las semanas 14 y 17, respectivamente), una nueva oportunidad de corral (la 1 hora siguiente a la reubicación en los corrales de puesta en la semana 16), una oportunidad de camada (en la semana 26), y una oportunidad al aire libre cuando se dio acceso a las aves al aire libre en la última semana del estudio (semana 27). El tiempo de las diferentes observaciones de comportamiento se ilustra con flechas rojas. También se registraron las ubicaciones de los huevos puestos durante todo el período de puesta (semanas 16-27).
Figura 1. Se proporcionaron dos ambientes, de elección única y de opción múltiple, durante dos períodos de cría, temprano (día 1 a la semana 4) y tardío (semana 5-15), lo que resultó en cuatro tratamientos de elección durante la cría. Los rectángulos ilustran un corral de muestra de cada entorno, con cuadrados de colores que representan los tipos de camada y diferentes estilos de línea que representan los tipos de percas. En la semana 16, las aves de cada corral de cría se trasladaron a un corral de puesta estándar. Se investigaron las respuestas conductuales a diferentes oportunidades de recursos en diferentes pruebas o contextos: oportunidades de comida (en un campo de prueba durante las semanas 9-10 y en los corrales domésticos en las semanas 14 y 17, respectivamente), una nueva oportunidad de corral (la 1 hora siguiente a la reubicación en los corrales de puesta en la semana 16), una oportunidad de camada (en la semana 26), y una oportunidad al aire libre cuando se dio acceso a las aves al aire libre en la última semana del estudio (semana 27). El tiempo de las diferentes observaciones de comportamiento se ilustra con flechas rojas. También se registraron las ubicaciones de los huevos puestos durante todo el período de puesta (semanas 16-27).
En el entorno de opción múltiple, había cuatro tipos diferentes de arena y cuatro tipos diferentes de perchas en cada corral. Los tipos de cama fueron arena de grano fino (máximo 0,3 mm Ø), virutas de madera (sin polvo), paja (trigo, corte largo) y turba (100% musgo Sphagnum). Los diseños de las perchas eran de goma redonda con un núcleo de madera (3,5 cm Ø), un tablón plano de madera (9,5 × 2 cm), una cuerda trenzada oscilante (4 × 3 cm) y una malla de alambre plana en un marco de madera (marco de madera de 2 × 1 cm, malla de 9,5 cm, aberturas de malla de 1 cm2). La ubicación de los diferentes tipos de camadas y perchas se equilibró en los corrales. En el entorno de elección única, el tipo de camada y el tipo de percha fueron los mismos en las cuatro ubicaciones de los corrales. Equilibramos los efectos potenciales de tipos específicos de perchas y camadas al proporcionar todos los tipos utilizados en el entorno de opción múltiple en los corrales de elección única. Colocamos los tipos de cama y percha en cada corral de elección única para que, según los resultados de un estudio piloto, los tipos de camada preferidos se emparejaran con los tipos de percha menos preferidos y viceversa. Esto se hizo para evitar que los corrales ofrecieran únicamente tipos de cama y perchas no preferidos o preferidos. Los emparejamientos fueron: virutas de madera-cuerda, arena-tablón de madera, paja-caucho y malla de alambre de turba. La misma configuración ambiental se utilizó en Nazar et al. (27).
2.4 Oportunidad de comida
Llevamos a cabo dos conjuntos de pruebas de oportunidad alimentaria que consistieron en localizar y consumir una recompensa alimenticia inicialmente novedosa. Las primeras pruebas (oportunidad alimenticia 1) se llevaron a cabo en condiciones experimentales en un campo de pruebas inicialmente novedoso, que es una práctica típica para la evaluación del comportamiento exploratorio [(p. ej., 37)]. También realizamos una segunda serie de pruebas (oportunidad de alimento 2) en los corrales domésticos, siguiendo la recomendación de Réale et al. (38), quienes propusieron que la exploración de objetos o alimentos nuevos debería evaluarse en condiciones familiares y ecológicamente relevantes, que para los animales de producción serían su entorno de origen. Este método evita la influencia del miedo debido a la novedad del campo de prueba (39).
2.4.1 Campo de pruebas (oportunidad de comida 1)
El primer conjunto de pruebas de oportunidad de alimento se llevó a cabo en las semanas 9-10, después de que las aves se habían mantenido aproximadamente iguales períodos de tiempo en los ambientes de cría temprana y tardía, a una edad en la que las aves rojas de la selva (G. gallus) se alimentan independientemente de su madre (40). Las pruebas se realizaron durante un período de 12 días, con un máximo de dos repeticiones/día y cuatro días de prueba por ave. La arena estaba situada en el galpón de cría, de modo que la intensidad de la luz, la temperatura y los sonidos eran similares a las condiciones de los corrales de cría. La arena tenía un piso de concreto (248 × 218 cm), techo de malla metálica y paredes de cartón (70 cm de altura). Se pegaron nueve tazas de café de plástico al suelo, cada una cebada con un gusano de la harina vivo (larva de Tenebrio molitor) (Figura 2A). Las mismas tres aves por corral fueron probadas repetidamente. Se seleccionaron tres aves en cada corral de una manera estandarizada y semialeatoria para abarcar la variación entre las aves dentro de cada corral. Un ave era fácil de atrapar, a menudo se acercaba directamente al investigador, un ave era levantada de una percha y un ave era más difícil de atrapar debido a más intentos de evitar al investigador. Los tres sujetos de prueba fueron marcados con un color específico del anillo de la pierna para facilitar su identificación durante los siguientes días de prueba.
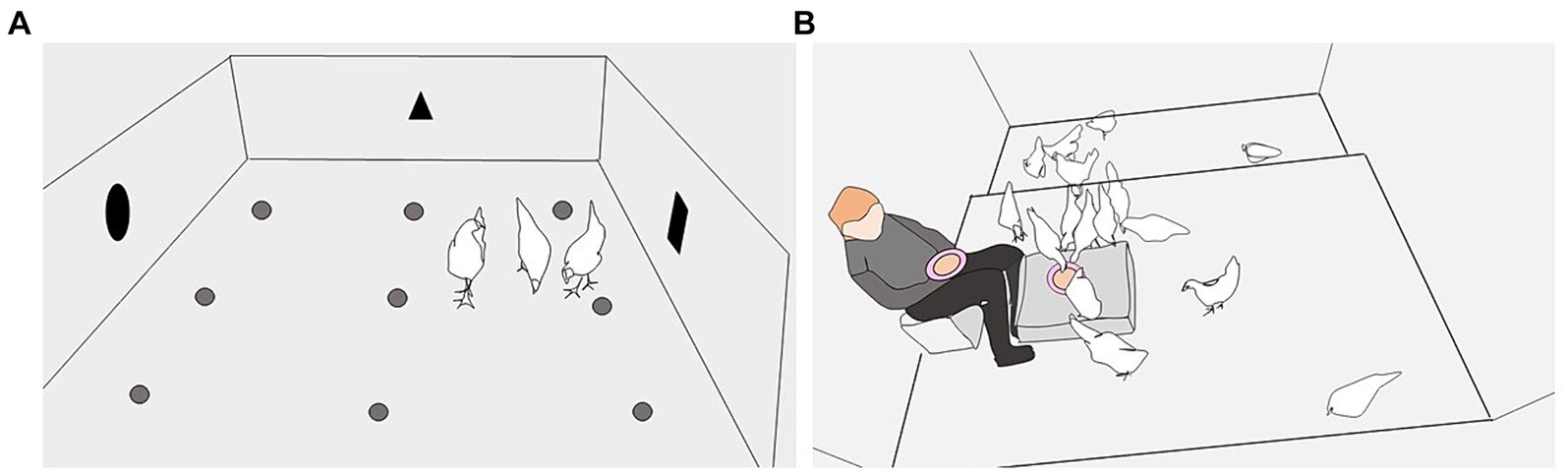 Figura 2. Los dos tipos de oportunidades alimentarias. (A) Las pruebas de oportunidad alimenticia 1 se llevaron a cabo en una arena inicialmente novedosa instalada en la nave de cría y que contenía nueve tazas de café de plástico en el suelo, cada una cebada con un gusano de la harina vivo. Las aves fueron evaluadas individualmente o en tríos y se les dio 5 minutos para encontrar y comer los gusanos de la harina. La ilustración es de una sesión de trío. (B) Se realizaron pruebas de oportunidad de alimentación 2 en el corral de la casa y se les dio a las aves 1.5 minutos para localizar y consumir gusanos de la harina escondidos debajo de gránulos de paja triturada en un recipiente «molido» en el piso de la cama o en una caja, y un tazón sostenido en el regazo de un investigador familiar. La ilustración es de una prueba en el corral de ponedora, con el cuenco «molido» en una caja.
Figura 2. Los dos tipos de oportunidades alimentarias. (A) Las pruebas de oportunidad alimenticia 1 se llevaron a cabo en una arena inicialmente novedosa instalada en la nave de cría y que contenía nueve tazas de café de plástico en el suelo, cada una cebada con un gusano de la harina vivo. Las aves fueron evaluadas individualmente o en tríos y se les dio 5 minutos para encontrar y comer los gusanos de la harina. La ilustración es de una sesión de trío. (B) Se realizaron pruebas de oportunidad de alimentación 2 en el corral de la casa y se les dio a las aves 1.5 minutos para localizar y consumir gusanos de la harina escondidos debajo de gránulos de paja triturada en un recipiente «molido» en el piso de la cama o en una caja, y un tazón sostenido en el regazo de un investigador familiar. La ilustración es de una prueba en el corral de ponedora, con el cuenco «molido» en una caja.
Las tres aves de prueba por corral se probaron en dos condiciones sociales: juntas (tríos) en las repeticiones 1-4 y 6, y cada ave individualmente en las repeticiones 5 y 7. Para las pruebas, los tres sujetos por corral fueron capturados y llevados juntos a la arena en una caja de transporte cubierta. En sesiones individuales, cada ave se colocó sola en una esquina de la arena de prueba y se le dio 5 minutos para encontrar 9 gusanos de la harina. Los gusanos de la harina que se comían se reemplazaban antes de que la segunda ave se colocara sola en la arena de prueba, y así sucesivamente. En sesiones de trío, las tres aves se colocaron juntas en la misma esquina de la arena de prueba y se les dio un máximo de 5 minutos para encontrar 9 gusanos de la harina. Un investigador se sentó fuera de la arena y registró cuándo se comió cada gusano de la harina y por qué individuo. Si todos los gusanos de la harina se comieron antes de que transcurrieran 5 minutos, la prueba se consideró finalizada. Después de las pruebas, las tres aves fueron devueltas a su corral de origen en la caja de transporte.
Se registró el número de gusanos de la harina consumidos del número proporcionado para cada repetición, y se calculó una puntuación de pluma para la proporción de gusanos de la harina consumidos. Por ejemplo, si el pájaro 1 comió tres gusanos de la harina, el pájaro 2 comió un gusano de la harina y el pájaro 3 comió cinco gusanos de la harina, la puntuación del lápiz sería de 0,33 (9 de 27 posibles) si se prueba individualmente, pero de 1,0 (9 de 9 posibles) si se prueba como trío.
2.4.2 Corral casero (oportunidad de comida 2)
El segundo conjunto de pruebas de oportunidad alimentaria se llevó a cabo a nivel de grupo, en los corrales domésticos, durante la semana 14 (13 días antes de la transición a la nave de puesta) y la semana 17 (13 días después de la transición). La prueba consistió en tres repeticiones de una oportunidad de 1,5 minutos para que las aves en el corral localizaran y consumieran gusanos de la harina escondidos debajo de gránulos de paja triturada en dos cuencos cebados (porcelana rosa, 22 cm Ø x 6 cm de profundidad), cada uno de los cuales contenía 10 gusanos de la harina. En el corral de cría, un cuenco se colocaba en el suelo y el otro estaba en el regazo de un investigador sentado en el corral [(véase 27) para una ilustración]. En el corral de ponedora, aumentamos el nivel de desafío colocando el «cuenco molido» en una novedosa caja de plástico (78 × 56 × 18 cm; Figura 2B). Las dos primeras repeticiones se realizaron con 3 horas de diferencia el mismo día, mientras que la tercera tuvo lugar 24 horas después de la primera repetición. Antes de cada repetición en el corral de cría, las tres aves probadas previamente en la arena fueron retiradas del corral, ya que tenían experiencia previa con gusanos de la harina. Esto significaba que los gránulos de paja triturada, los gusanos de la harina y los cuencos eran inicialmente nuevos para todas las aves del corral. En el período de puesta, todas las aves tenían experiencia previa con los gusanos de la harina, por lo que todas permanecieron en el corral durante la prueba. Se registró el número de gusanos de la harina consumidos de los 20 proporcionados para cada repetición, y los datos se expresaron como proporciones.
2.5 Oportunidad de pluma novedosa
Las aves fueron grabadas en video durante la primera hora después de haber sido reubicadas de los corrales de cría a los corrales de puesta de adultos (Figura 3). Los corrales de puesta tenían un diseño estándar (ver sección 2.2) y los comederos, el bebedero, el área de listones, las perchas y los nidos de colonia cerrados eran todos novedosos, mientras que las bandejas de anidación elevadas y la cama eran parcialmente novedosas. Las bandejas de anidación se habían utilizado como bandejas de arena durante el período de cría temprana, pero ahora estaban sin cama y colocadas en un marco de madera elevado unido a la pared, lo que les daba una apariencia novedosa. El tipo de cama en el corral de puesta (gránulos de paja triturada) se había utilizado en la prueba de oportunidad alimentaria 2 durante un total de 4,5 minutos de posible exposición previa.
 Figura 3. La oportunidad de la pluma novedosa. Después de que las aves de un corral de cría fueron liberadas en una esquina de su nuevo corral de puesta, su comportamiento se registró durante su primera hora en este nuevo entorno. El corral contenía un piso de cama y un orificio para pop (cerrado) en la parte posterior, un piso de listones en la parte delantera, dos comederos colgantes, un bebedero de campana, perchas de plástico y madera, bandejas de anidación elevadas (dos de tres son visibles en la pared izquierda) y dos nidos de colonia en la parte delantera (no visibles). Se registraron las latencias de la primera y quinta ave en utilizar estos nuevos recursos, así como los recuentos de aves en la hojarasca y los listones dentro del área rectangular roja.
Figura 3. La oportunidad de la pluma novedosa. Después de que las aves de un corral de cría fueron liberadas en una esquina de su nuevo corral de puesta, su comportamiento se registró durante su primera hora en este nuevo entorno. El corral contenía un piso de cama y un orificio para pop (cerrado) en la parte posterior, un piso de listones en la parte delantera, dos comederos colgantes, un bebedero de campana, perchas de plástico y madera, bandejas de anidación elevadas (dos de tres son visibles en la pared izquierda) y dos nidos de colonia en la parte delantera (no visibles). Se registraron las latencias de la primera y quinta ave en utilizar estos nuevos recursos, así como los recuentos de aves en la hojarasca y los listones dentro del área rectangular roja.
Cuando se les presentó su nuevo corral, las aves se colocaron en grupo en los listones. Para cada grupo, se determinaron las latencias para la primera y quinta ave que se observará alimentándose, en la cama y en una estructura elevada (percha o nido) a partir de las grabaciones de video y, por lo tanto, se utilizaron dos latencias por corral para el análisis. Si ningún pájaro utilizaba estos recursos, se asignaba una latencia máxima de 60 min para ambas latencias, mientras que si menos de cinco pájaros los utilizaban, se asignaba 60 min como latencia del quinto pájaro. Como solo dos corrales tenían cinco aves que usaban una estructura elevada durante el período de observación de 60 minutos, solo se analizó la latencia de la primera ave para esta variable.
Para evaluar la exploración a nivel de grupo del nuevo corral, se registró el número de aves en la cama y los listones en la mitad opuesta del corral desde la ubicación inicial utilizando un muestreo instantáneo de barrido a intervalos de 2 minutos durante la primera hora inmediatamente después de su introducción. Para el análisis de los datos, el período de observación de 60 minutos se dividió en tres fases de 20 minutos y el recuento promedio por exploración por fase se expresó como proporción del número total de aves en el corral.
2.6 Oportunidad de anidación
Los huevos se recolectaron una vez al día, y sus ubicaciones y pesos (con una precisión de 0,1 g) se registraron para cada grupo desde el primer huevo hasta el final del experimento (excluyendo los huevos agrietados o rotos). Como medida de cómo las aves utilizaron el espacio del corral para la oviposición durante el período de puesta, contamos el número de huevos puestos en las bandejas de anidación elevadas, en los nidos de la colonia y en el suelo («huevos en el suelo» situados en los listones o en la hojarasca). Para el análisis, se calculó la proporción de huevos puestos en cada lugar (nidos elevados, nidos de colonias, en el suelo).
Para evaluar los cambios en la producción y el peso del huevo a lo largo del tiempo, el período de puesta de 72 días (semana 16-27) se dividió en tres períodos de 24 días. Para cada corral, se calculó el número promedio de huevos producidos por ave por período de 24 días y el peso promedio de los huevos por período de 24 días.
2.7 Oportunidad de la camada
Esta prueba se realizó en el corral casero para adultos en la semana 26. A las aves se les dio 1 h de acceso a seis cajas (55 litros, 78 × 56 × 18 cm), cada una de las cuales contenía un tipo diferente de cama fresca (Figura 4): pellets de paja triturada (tipo de cama actual en los corrales de puesta), paja, turba, arena y virutas de madera (los cuatro tipos de cama utilizados durante la cría) y virutas de cáñamo (un nuevo tipo de cama para todas las aves). La paja, la turba, la arena y las virutas de madera eran familiares para las aves expuestas a la opción múltiple, mientras que tres tipos eran nuevos para las aves expuestas solo a la opción única, y los nuevos tipos variaban entre los grupos. La ubicación de los tipos de camada dentro de las cajas se equilibró en los corrales. En cada corral, se registró la latencia de la primera ave que se vio en cada ubicación de la caja de arena mediante la observación continua de una grabación de video de la prueba (Figura 4A). Además, se registró el número de aves que realizaban forrajeo y baños de polvo por ubicación de la caja de arena utilizando un muestreo instantáneo de escaneo del video a intervalos de un minuto durante 1 h (Figura 4B). Para el análisis de cada comportamiento, el conteo de comportamiento por caja por escaneo se expresó como una proporción de las aves en el corral. El número de tipos de camadas familiares y novedosas varió según los tratamientos. Por lo tanto, para cada variable latencia y proporción se calcularon tres valores promedio de corral, uno para cada uno de los tres grados de familiaridad con la camada (actual, familiar de la crianza y novedoso, respectivamente) para su uso en el análisis estadístico.
 Figura 4. En la prueba de oportunidad de camada, (A) se registró la latencia de la primera gallina en entrar en cada caja que contenía un tipo de camada diferente, y (B) se registró el número de gallinas que realizaban forrajeo, baños de polvo u otro comportamiento en cada caja de arena a intervalos de 1 minuto durante 1 h. Los tipos de camada se clasificaron como actuales, familiares de la crianza o novedosos.
Figura 4. En la prueba de oportunidad de camada, (A) se registró la latencia de la primera gallina en entrar en cada caja que contenía un tipo de camada diferente, y (B) se registró el número de gallinas que realizaban forrajeo, baños de polvo u otro comportamiento en cada caja de arena a intervalos de 1 minuto durante 1 h. Los tipos de camada se clasificaron como actuales, familiares de la crianza o novedosos.
2.8 Oportunidad al aire libre
En la semana 27, se abrió un orificio en cada corral de puesta que le dio a cada grupo acceso a un campo de tiro al aire libre separado (3,5 × 16 m) conectado al corral. Esto ocurrió en abril durante un período de tiempo soleado sin lluvias. Cada campo tenía los lados y el techo de malla de alambre, grava fina con algo de vegetación corta (principalmente pasto) y estaba dividida en cuatro áreas iguales (3,5 × 4 m) que representaban la sección más cercana a la más lejana de la pared de la casa. En los primeros 3 días, los orificios de popa estuvieron abiertos durante un total de 12,5 h, durante los cuales se realizó un muestreo instantáneo de barrido cada 30 min mediante observación directa. La latencia para el primer pájaro por corral que se vio al aire libre se registró en el intervalo de observación más cercano de 30 minutos, expresado como un valor de 0 a 12,5. También se registró el número de aves en cada sección de distribución por escaneo. Sin embargo, pocas aves utilizaron el rango durante los 3 días de observación, por lo que estos datos se combinaron y analizaron como el número promedio de aves al aire libre por escaneo.
2.9 Análisis estadístico
Los análisis se realizaron en R Studio (versión 1.3.959). En la Tabla Suplementaria S1 se ofrece una descripción del modelo estadístico para cada variable. Los modelos estadísticos siempre incluyeron el ambiente de cría temprana (opción única o opción múltiple), el entorno de cría tardía (opción única o opción múltiple) y su interacción (Soltero temprano * Soltero tardío, Soltero temprano * Múltiple tardío, Múltiple temprano * Único tardío y Multi temprano * Multi tardío, para evaluar si los efectos de la elección cambiaron a lo largo de los períodos de cría). Para las variables con mediciones repetidas, se incluyó el lápiz como efecto aleatorio. Se incluyeron factores de efecto fijo adicionales en los modelos para variables específicas. La condición social (Individual o Trío) se incluyó en el modelo para la oportunidad de alimentación 1, y el período de prueba (Cría o Puesta) para la oportunidad de alimentación 2. El orden (primera o quinta ave) se incluyó al examinar las latencias para usar los recursos (cama y alimento) en el nuevo corral. Los modelos que evaluaron las variables de producción de huevos incluyeron un período de 24 días (1º, 2º o 3º), mientras que el modelo que examinó la proporción de huevos puestos en lugares específicos también incluyó la ubicación de los huevos (nidos de colonia, nidos elevados o suelo). El modelo de oportunidad de camada incluyó el grado de familiaridad con los tipos de camada (Actual, Familiar de la crianza o Novedoso). Los modelos con un factor adicional también incluían la interacción de ese factor con el ambiente de elección, dentro y a través de los períodos de cría, pero si estas interacciones tenían p ≥ 0.10, se eliminaban del modelo final.
Todas las variables se consideraron como valores continuos si cumplían los supuestos de normalidad y homocedasticidad. Las medidas repetidas se analizaron en modelos lineales mixtos con máxima verosimilitud restringida utilizando los paquetes lme4 (41), pbkrtest (42) y lmerTest (43). La significación se evaluó mediante pruebas de Wald F tipo III con la aproximación de Kenward-Roger de grados de libertad. Cuando solo hubo un valor por corral, se emplearon modelos lineales (función lm), con significación evaluada mediante pruebas de Tipo III y la función de contradicción. Los supuestos para los modelos ajustados se verificaron visualmente en gráficos de probabilidad de residuos (gráfico QQ) y gráficos de residuales frente a valores ajustados. Cuando no se cumplieron los supuestos para un modelo lineal, los datos calculados como promedios por corral se analizaron utilizando pruebas de Kruskal Wallis. Pocos corrales tuvieron aves bañándose en el polvo durante la prueba de oportunidad de cama y solo se presentan datos descriptivos para esta variable.
Los resultados se presentan como medias marginales estimadas con error estándar y se grafican utilizando el paquete ggplot2 (44). Los valores transformados se presentan después de la transformación inversa a la escala original. El nivel de significación se fijó en 0,05. Los resultados significativos se presentan con comparaciones planificadas por pares realizadas utilizando el paquete de medios (45). Para todas las variables, se presentan las medias globales con EE para cada combinación de tratamiento (Opción única o Opción múltiple en los períodos de recría temprana y tardía, respectivamente; Cuadro complementario S2).
3 Resultados
En los dos conjuntos de pruebas de oportunidad alimentaria, la proporción de gusanos de la harina consumidos se utilizó como indicador de la capacidad de las aves para explotar un alimento inicialmente nuevo en el campo de prueba y en el corral doméstico, respectivamente. Se encontró que el ambiente de cría tardía tuvo un efecto principal sobre el consumo de gusanos de la harina, por lo que el consumo fue mayor en las aves expuestas al ambiente de opción múltiple durante el período de cría tardía en ambos conjuntos de pruebas (oportunidad alimenticia 1: opción única, 0,20 ± 0,04; Opción múltiple, 0,41 ± 0,06; F1,12 = 10,4, p = 0,007; oportunidad alimentaria 2: opción única, 0,33 ± 0,05; Opción múltiple, 0,48 ± 0,05; F1,12 = 5,11, p = 0,043, Figura 5). Además, se consumió una mayor proporción de gusanos de la harina en la oportunidad alimentaria 1 cuando las aves se probaron en grupos de tres (0,40 ± 0,04) que individualmente (0,21 ± 0,03; F1,12 = 33,64, p < 0,001), y en la oportunidad de alimentación 2 después de la reubicación en los corrales de puesta (corral de cría, 0,30 ± 0,04; corral de ponedora, 0,51 ± 0,04, F1,12 = 17,87, p < 0,001).
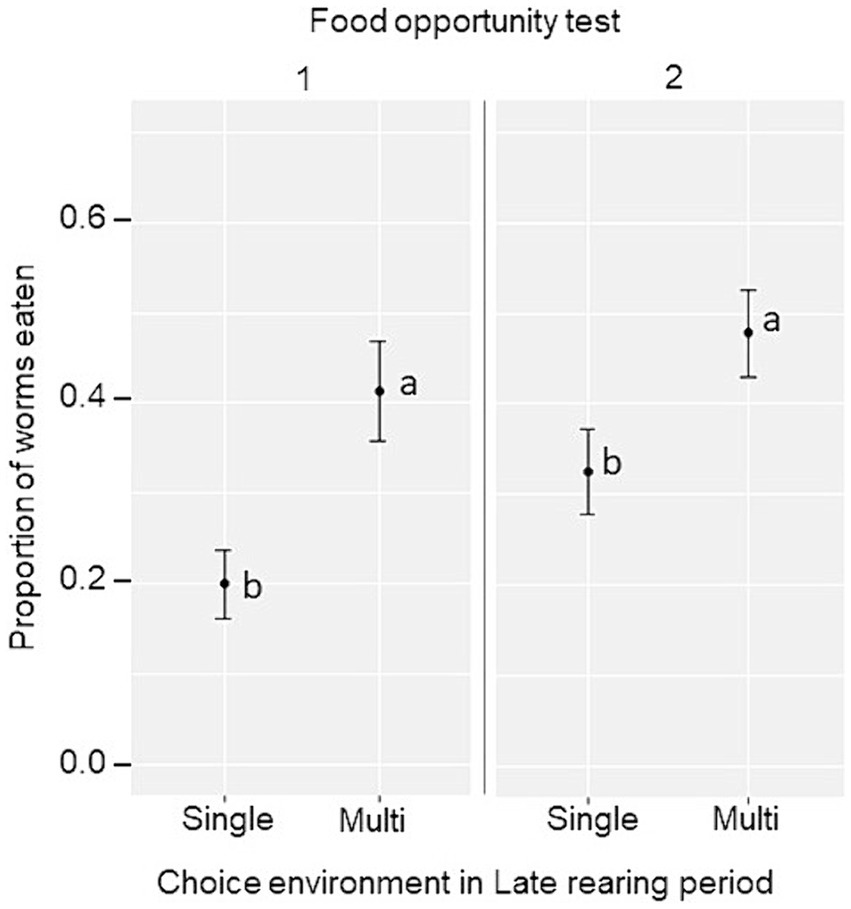 Figura 5. Proporción marginal media estimada ± SE de gusanos de la harina consumidos por grupo en la prueba de oportunidad alimentaria 1 (campo de prueba) y 2 (corral doméstico) por aves mantenidas en un ambiente de opción única o múltiple durante el período de cría tardía. Diferentes letras (a, b) indican diferencias significativas dentro de cada prueba de oportunidad alimentaria (p < 0.05).
Figura 5. Proporción marginal media estimada ± SE de gusanos de la harina consumidos por grupo en la prueba de oportunidad alimentaria 1 (campo de prueba) y 2 (corral doméstico) por aves mantenidas en un ambiente de opción única o múltiple durante el período de cría tardía. Diferentes letras (a, b) indican diferencias significativas dentro de cada prueba de oportunidad alimentaria (p < 0.05).
Las latencias para utilizar los nuevos recursos en el corral de puesta se determinaron durante la primera hora después de la transferencia desde el corral de cría. No se encontraron efectos significativos del ambiente de cría temprana o tardía, ni de su interacción, con respecto a las latencias de la primera y quinta ave para alimentarse o ser vista en la camada (p ≥ 0.10; ver Tabla Suplementaria S2 para los promedios de tratamiento). Sin embargo, hubo un efecto principal del ambiente de cría temprana sobre la latencia para la primera ave que usó estructuras elevadas, siendo la latencia más corta para los grupos criados en un ambiente de opción múltiple (F1,12 = 5,49, p = 0,037; Figura 6A). En cuanto a la proporción de aves que exploran la cama y los listones situados en el lado opuesto del corral de puesta a aquel en el que fueron introducidos, se observa una tendencia a una interacción tripartita entre el ambiente de cría temprana, el ambiente de cría tardía y la fase de observación (1ª, 2ª y 3ª 20 min; F2,425 = 2,37, p = 0,094). Las comparaciones por pares mostraron que, en los grupos de todas las combinaciones de tratamientos, excepto el tratamiento Single*Single, hubo un aumento en la proporción de aves en el lado opuesto del corral de la primera a la segunda fase (Single*Multi, t = 3,75 p < 0,001; Multi*Simple, t = 2,49 p = 0,036), y/o de la primera a la tercera fase (Simple*Multi, t = 3,5 p = 0,002; Multi*Multi, t = 3,11 p = 0,006; Figura 6B).
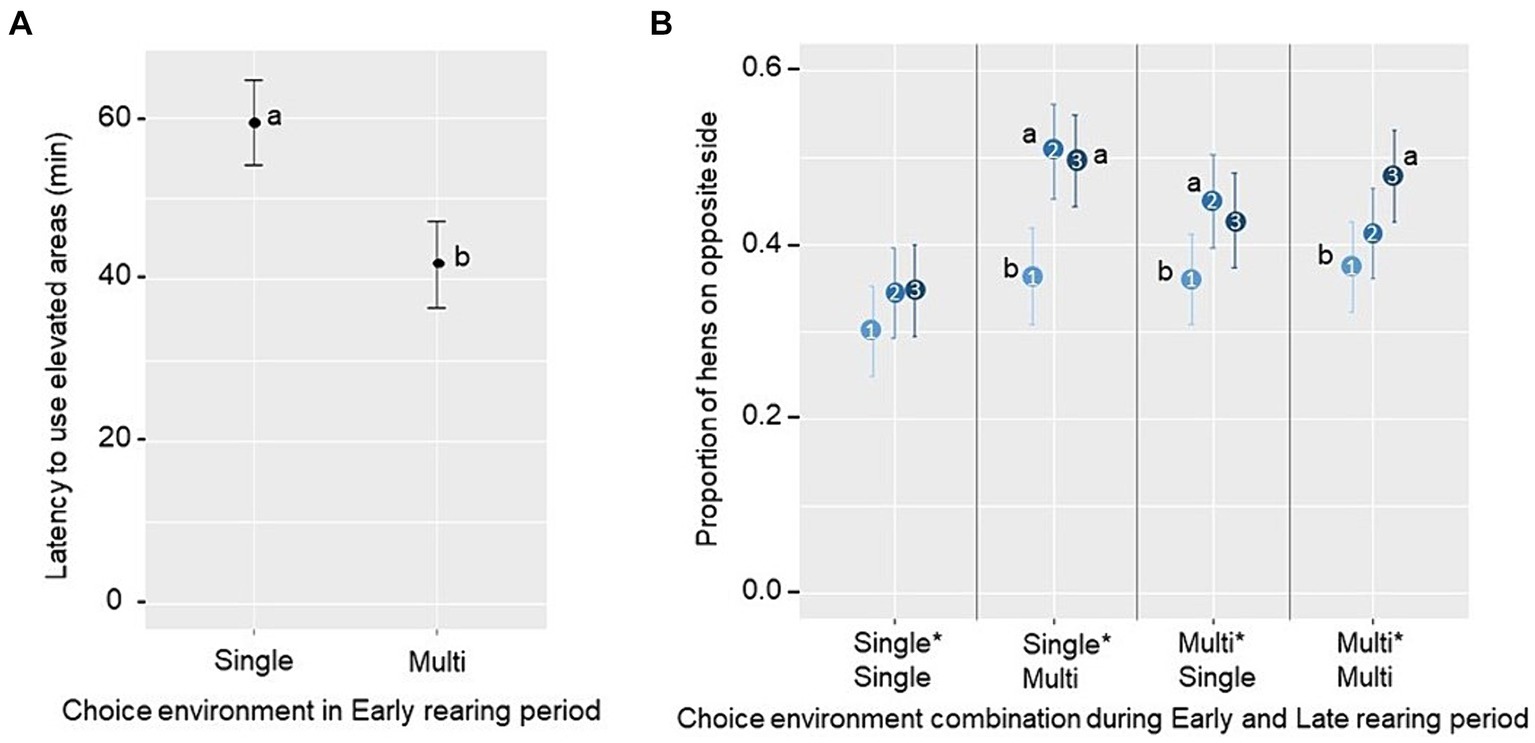 Figura 6. Media marginal estimada ± SE de (A) latencia para la primera gallina que utiliza una estructura elevada (percha o nido) en la primera hora después de la reubicación en el corral de puesta (nueva oportunidad de corral) en grupos criados en un entorno de opción única o de opción múltiple durante la cría temprana y (B) la proporción de gallinas ubicadas en el lado opuesto a la entrada durante las tres primeras fases de 20 minutos (1, 2 y 3, que van de azul claro a azul oscuro, respectivamente) en grupos de diferentes combinaciones de ambientes de elección durante el período de cría temprana y tardía. Letras diferentes (a, b) indican una diferencia significativa (A) entre o (B) dentro de un tratamiento (p < 0,05).
Figura 6. Media marginal estimada ± SE de (A) latencia para la primera gallina que utiliza una estructura elevada (percha o nido) en la primera hora después de la reubicación en el corral de puesta (nueva oportunidad de corral) en grupos criados en un entorno de opción única o de opción múltiple durante la cría temprana y (B) la proporción de gallinas ubicadas en el lado opuesto a la entrada durante las tres primeras fases de 20 minutos (1, 2 y 3, que van de azul claro a azul oscuro, respectivamente) en grupos de diferentes combinaciones de ambientes de elección durante el período de cría temprana y tardía. Letras diferentes (a, b) indican una diferencia significativa (A) entre o (B) dentro de un tratamiento (p < 0,05).
Si bien la mayoría de los huevos fueron puestos en los nidos de la colonia en todos los grupos, se encontró un efecto principal del ambiente de cría tardía para el uso del nido (F2,114 = 5,64, p = 0,005). Las aves del ambiente de opción múltiple pusieron una menor proporción de huevos en los nidos de la colonia (t = −2.59, p = 0.012), y una mayor proporción en las bandejas de anidación elevadas (t = 2.08, p = 0.041), en comparación con las aves del ambiente de elección única durante el período de cría tardía (Figura 7). No se encontraron diferencias de tratamiento en cuanto a la proporción de huevos puestos en el suelo, el número promedio de huevos por ave o el peso promedio de los huevos por período de 24 días (p ≥ 0.10, ver Tabla Suplementaria S2 para los promedios de tratamiento).
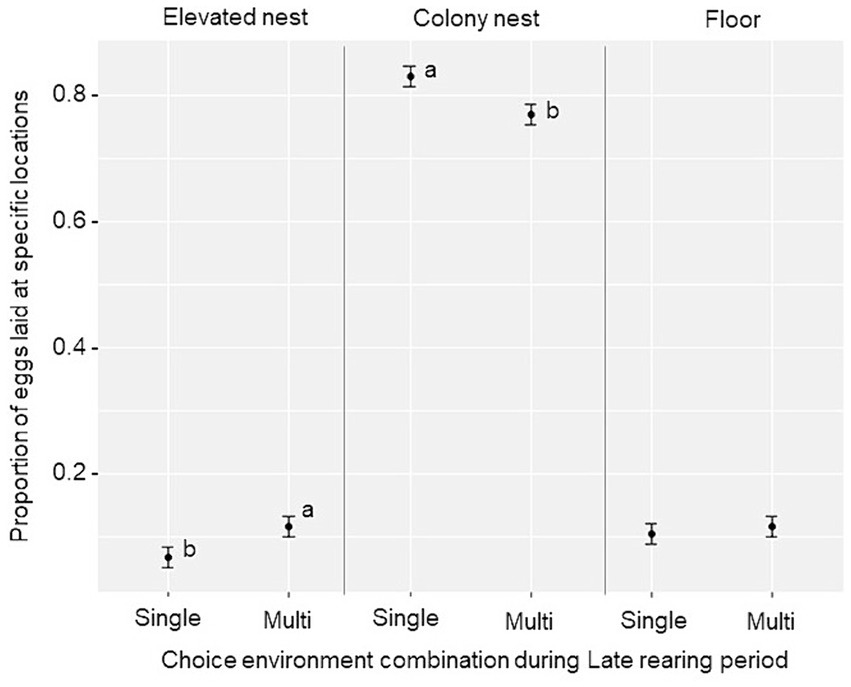 Figura 7. Uso del sitio de anidación (media marginal estimada ± SE) durante el período de puesta (semanas 16-27) para la proporción de huevos puestos en las bandejas de anidación elevadas, nidos de colonia y en el suelo por aves criadas en los ambientes de «elección única» y «opción múltiple» durante el período de cría tardía. Las diferentes letras (a, b) muestran una diferencia significativa en el tratamiento (p < 0,05) dentro de cada ubicación del huevo.
Figura 7. Uso del sitio de anidación (media marginal estimada ± SE) durante el período de puesta (semanas 16-27) para la proporción de huevos puestos en las bandejas de anidación elevadas, nidos de colonia y en el suelo por aves criadas en los ambientes de «elección única» y «opción múltiple» durante el período de cría tardía. Las diferentes letras (a, b) muestran una diferencia significativa en el tratamiento (p < 0,05) dentro de cada ubicación del huevo.
La latencia para que la primera ave entrara en una caja de arena durante la prueba de oportunidad de camada se vio afectada por la interacción de tres vías entre el entorno de cría temprana, el entorno de cría tardía y el grado de familiaridad con el tipo de camada en la caja (F2,24 = 5,25, p = 0,013). Las comparaciones por pares revelaron que, en los corrales del tratamiento Single*Single, la latencia para el primer pájaro en explorar un nuevo tipo de camada fue mayor que la latencia para el primer pájaro en explorar un tipo de camada que era familiar de la cría (p = 0.018). En los corrales del tratamiento Multi*Single, la latencia para que el primer pájaro entrara en una caja que contenía una camada familiar de la cría fue mayor que la latencia para entrar en una caja que contenía una camada nueva o del tipo de camada actual (p < 0,01; Figura 8). Las latencias observadas en los grupos de tratamiento Single*Multi y Multi*Multi no aumentaron para ninguno de los tres grados de familiaridad con la camada (p > 0.05). No se encontraron efectos significativos del ambiente de elección en ninguno de los períodos de cría para el forrajeo en las cajas de arena (p > 0.05). Sin embargo, el forrajeo se vio afectado por el grado de familiaridad con la camada (F2,30 = 7,91, p = 0,002), con una menor proporción media de aves por escaneo que se alimentan en un nuevo tipo de camada (0,021 ± 0,004) que en el tipo de camada actual (0,040 ± 0,005; t = −3,06, p = 0,005) o un tipo de camada familiar de la cría (0,045 ± 0,006; t = −3,73, p < 0,001). Se observaron baños de polvo en cinco corrales (un Single*Single-pen con 5 observaciones, dos Single*Multi-pens con 47 observaciones y dos Multi*Multi-pens, con 84 observaciones en total). Todas estas observaciones de baños de polvo se observaron en la turba cuando la turba era un tipo de cama familiar de la cría.
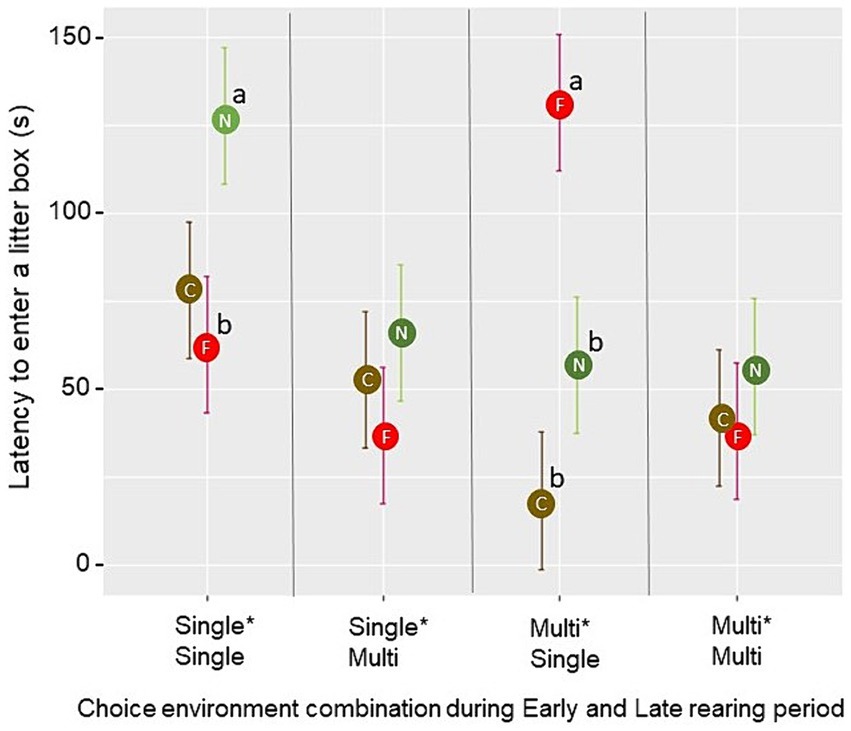 Figura 8. En la prueba de oportunidad de cama, la latencia media ± SE para la primera ave que entró en una caja de arena de prueba estuvo influenciada por el grado de familiaridad con los tipos de camada en las cajas (C = actual (marrón), F = familiar de la cría (rojo), N = nuevo (verde)) y el tratamiento de crianza. Diferentes letras (a, b) indican una diferencia significativa (p < 0.05) dentro de la combinación de ambientes de elección durante el período de cría temprana y tardía entre cajas de arena de diversa familiaridad.
Figura 8. En la prueba de oportunidad de cama, la latencia media ± SE para la primera ave que entró en una caja de arena de prueba estuvo influenciada por el grado de familiaridad con los tipos de camada en las cajas (C = actual (marrón), F = familiar de la cría (rojo), N = nuevo (verde)) y el tratamiento de crianza. Diferentes letras (a, b) indican una diferencia significativa (p < 0.05) dentro de la combinación de ambientes de elección durante el período de cría temprana y tardía entre cajas de arena de diversa familiaridad.
El ambiente de elección durante el período de cría temprana o tardía no afectó la latencia hasta que se vio la primera ave por corral en el campo de tiro al aire libre después de abrir los orificios de salida (p ≥ 0.10), o el número promedio de aves que usaron el campo de distribución al aire libre por escaneo (p ≥ 0.10, ver Tabla Suplementaria S2 para los promedios de tratamiento). Sobre la base de los datos descriptivos, las aves de los cuatro corrales Single*Multi visitaron el campo de distribución al aire libre en algún momento durante las observaciones. Se observaron aves de solo uno de los corrales de tratamiento Multi*Single y de solo dos de los corrales de tratamiento Multi*Multi y Single*Single, respectivamente, en el exterior. Además, solo en dos corrales (ambos corrales de tratamiento simple y múltiple) se observó que las aves se alejaban más de la nave que la primera sección de distribución (es decir, más allá del 25% más cercano del área de distribución total disponible).
4 Discusión
Los resultados de este estudio respaldaron nuestra primera predicción de que proporcionar a las gallinas ponedoras jóvenes acceso a una variedad de tipos de cama y percha (es decir, ofrecer múltiples opciones) durante la cría, tendría efectos a largo plazo en las habilidades de las aves para capitalizar las nuevas oportunidades que se les presentan. Esto fue evidente por el mayor uso de recursos como nuevos alimentos, y nidos elevados, y una exploración más rápida de los nuevos corrales de ponedora. Además, influyó la edad de las aves en el momento en que se proporcionó esta variedad durante la cría. La exposición a una selección de tipos de cama y perchas durante las primeras 4 semanas aumentó el uso de un nuevo espacio tridimensional para corrales durante la primera hora después de la transferencia a los corrales de ponedora. Sin embargo, se observaron efectos potencialmente beneficiosos del ambiente de elección sobre los resultados en las pruebas de oportunidad de alimento y cama y el uso elevado del nido entre las aves que habían estado expuestas a esta variación durante las semanas 5-15, contrariamente a nuestra predicción con respecto a la importancia de la elección del entorno durante las primeras 4 semanas después de la eclosión. Discutimos estos resultados, pero también cómo la experiencia de diferentes variantes del mismo recurso puede actuar de una manera más general sobre el comportamiento de las aves.
Independientemente de las características del ambiente de cría tardío, las elecciones ambientales durante las primeras 4 semanas de edad (opción múltiple temprana), aumentaron la preparación de las aves para posarse después de la reubicación en el nuevo corral de puesta (nueva oportunidad de corral), como lo demuestra la latencia reducida para usar estructuras elevadas. Hay varias explicaciones posibles para este hallazgo. Los tipos de perchas durante la cría fueron visualmente diferentes, lo que podría haber mejorado la capacidad de las aves para detectar e identificar una estructura elevada en el nuevo corral como un posible lugar de posadero (46). La experiencia temprana con diferentes tipos de perchas, y la consiguiente mejora del equilibrio (25), pueden haber facilitado el movimiento en el espacio tridimensional en mayor medida que proporcionar un solo tipo de percha durante la cría (30, 47). La reducción de la latencia para el uso de estructuras elevadas también puede haber sido influenciada por un mejor desarrollo óseo y muscular, que se ha reportado en aves criadas con perchas (48, 49), y puede haber mejorado aún más al balancearse en diferentes tipos de perchas en el entorno de opción múltiple. La mejora de las habilidades de navegación espacial puede haberse asociado con el período sensible para la formación de sinapsis cerebrales en las 3 semanas posteriores a la eclosión (50, 51). En general, nuestros resultados actuales pueden interpretarse como un apoyo a la existencia de una ventana corta en las primeras etapas de la vida para que las aves adquieran habilidades fundamentales para posarse, y agregamos que la adquisición de estas habilidades para posarse parece mejorar aún más con la provisión de diferentes tipos de perchas durante esta ventana.
Gunnarsson et al. (52) informaron que el acceso temprano a las perchas estaba relacionado con el uso posterior del nido, ya que las gallinas sin perchas dentro de las primeras 4 semanas de vida ponían un mayor porcentaje de huevos en el suelo que en los nidos. En el presente estudio, no hubo diferencias de tratamiento en el porcentaje de huevos de piso, tal vez porque todas las aves tuvieron acceso temprano a las perchas. Pero sí encontramos que las aves de los corrales de opción múltiple tardía (criadas con cuatro tipos de camada y percha en las semanas 5 a 15), pusieron más huevos en las bandejas de anidación elevadas y menos en los nidos de la colonia en comparación con las aves criadas con un solo tipo de camada y percha durante el mismo período de cría (elección única tardía). Este hallazgo sugiere que, incluso en ausencia de una elección temprana, la experiencia de cría posterior en el uso de estructuras elevadas variadas puede afectar la forma en que las aves explotan el espacio tridimensional como adultas. La selección del nido está probablemente determinada por una combinación de preferencias por la altura (53), el material de anidación (54) y la familiaridad con el nido (55), entre otros factores. En las gallinas salvajes, el aumento de la incubabilidad y la supervivencia inicial de las crías se ha asociado con sitios de anidación elevados (56). Sería necesario investigar más a fondo el uso de nidos para evaluar la posibilidad de que las aves tardías de opción múltiple sean más propensas a utilizar nidos elevados que las aves tardías de elección única en momentos en que los nidos de las colonias están ocupados, reduciendo así la competencia por el espacio de nido preferido (57).
Las aves criadas con acceso a una variedad de materiales de cama y perchas entre las 5 y las 15 semanas (opción múltiple tardía) consumieron más gusanos de la harina en comparación con las aves criadas con un solo tipo de cama y percha durante este período (elección única tardía). Se encontró un mayor éxito en la localización y el consumo de gusanos de la harina en todas las pruebas de oportunidad alimentaria, independientemente de si se realizaron durante el período de cría tardía o durante el período de puesta. No probamos el efecto del acceso a una variedad de camarascas y perchas sobre el éxito en el consumo de gusanos de la harina durante el período de cría temprana, ya que esto ya fue demostrado por Nazar et al. (27). Las aves del mismo tratamiento (Late Multi-choice) mostraron una latencia consistente y corta para ingresar a diferentes cajas de arena que contenían arena fresca de diversos grados de familiaridad cuando se les dio acceso brevemente. Por el contrario, las aves de los corrales de elección única tardía mostraron latencias aumentadas. En combinación, estos resultados apoyan la opinión de que un entorno de opción múltiple estimula la motivación para explorar, facilitando así la recopilación de información (3, 7) y el compromiso afectivo positivo (58). Estos resultados presumiblemente promovieron un sentido de agencia (11), mejorando el potencial de las aves para encontrar y hacer uso de un recurso alimenticio oculto, así como sitios de anidación y un recurso de forrajeo y baños de polvo (basura). Se ha demostrado que un entorno espacialmente complejo mejora el aprendizaje y la memoria de trabajo en tareas espaciales (20, 59, 60), potencialmente por una mayor activación neuronal en las regiones cerebrales relacionadas. Incluso si las sinapsis cerebrales ya están formadas, la maduración del cerebro continúa hasta la décima semana después de la eclosión (es decir, hasta bien entrado nuestro período de elección múltiple tardía) cuando los circuitos sinápticos están afinados y son más sensibles a las entradas ambientales (50), lo que respalda los grandes efectos que vemos en el entorno de cría tardía.
Dado que los efectos del entorno de cría temprana no anularon los efectos del entorno de cría tardía para la mayoría de nuestras variables medidas, sugerimos que la experiencia de perder opciones a través de una reducción en la variedad de tipos de perchas y camadas en la semana 5 (opción múltiple temprana y opción única tardía) suprimió la motivación y/o la capacidad de las aves para aprovechar las oportunidades. mientras que un aumento en el número de opciones en la semana 5 (Opción Única Temprana y Opción Múltiple Tardía) mejoró la motivación y/o la capacidad de las aves para aprovechar las oportunidades. Los efectos de contraste encontrados en nuestro estudio están en línea con las trayectorias afectivas, en las que los estados afectivos de los individuos están influenciados por sus expectativas del entorno y una pérdida de recursos o mercancías puede conducir a estados afectivos negativos y viceversa (61). Aunque Paul et al. (62) no encontraron evidencia que apoye un cambio en los estados afectivos generales en gallinas ponedoras adultas que ganaron o perdieron un ambiente más rico, es posible que una reducción o un aumento en la complejidad ambiental en la semana 5 tenga mayores consecuencias para la pollita joven altamente activa que un cambio más adelante en la vida. Dado que la impronta ocurre durante los primeros días después de la eclosión y los pollitos permanecen cerca de su madre durante al menos las primeras 4 semanas después de la eclosión (63, 64), los pollitos pueden haber desarrollado un apego a las características del entorno de elección múltiple durante el período de cría temprana. Además, basándose en investigaciones en humanos, Atkinson et al. (50) sugieren que las alteraciones durante la fase de maduración cerebral (que en las gallinas ponedoras se produce entre las semanas 3 y 10), pueden conducir a trastornos del neurodesarrollo. Es posible que la pérdida experimentada del entorno de opción múltiple cuando se cambió al entorno de opción única en la semana 5 haya tenido una influencia negativa en el desarrollo del cerebro, con consecuencias relacionadas con sus habilidades y motivación para participar en nuevas oportunidades.
El único resultado claramente influenciado por el ambiente de opción múltiple, independientemente de cuándo se proporcionó durante la cría (es decir, durante el período temprano, el período tardío o ambos) fue la velocidad a la que las aves se extendieron por el suelo cuando se colocaron por primera vez en el corral de puesta. Las aves con alguna experiencia previa de variación en la cama o perchas fueron más rápidas para explorar toda el área del nuevo corral de puesta que las aves que experimentaron un solo tipo de cama y percha. Las aves criadas en el ambiente de elección única durante la cría (elección única temprana y tardía) tuvieron los peores resultados. El nivel de exploración mostrado por un individuo (medido como locomoción en un nuevo escenario) como un polluelo está relacionado con el nivel mostrado como adulto en el Pájaro rojo de la selva (37). Por lo tanto, es probable que estimular la exploración dando a las aves la experiencia de diferentes variantes de recursos en algún momento durante la cría tenga efectos durante un tiempo aún más largo que el estudiado en nuestro experimento.
En este estudio adoptamos el enfoque novedoso de centrarnos en la medida en que las aves aprovechaban las oportunidades cuando se les ofrecían, lo cual es diferente a lo que se ha hecho comúnmente cuando se investiga la exploración o la flexibilidad conductual [(por ejemplo, 6, 7, 35)]. La razón de esto fue explorar el potencial de utilizar la agencia de un ave como una forma de mejorar su propio bienestar (11, 65) y tal vez promover el bienestar animal positivo. Proponemos que las diferencias en la forma en que las aves utilizaron las oportunidades dadas en este estudio, incluida la búsqueda rápida de un lugar seguro para descansar (un área elevada), la exploración del espacio y la búsqueda de nuevas fuentes de alimento, reflejaron diferencias en su motivación para estas actividades específicas, así como en su capacidad general para explotar la oportunidad a través del aumento de las habilidades cognitivas y motoras. La originalidad de este estudio es la propuesta de que sin motivación o la capacidad de aprovechar una oportunidad, la reducción del miedo y la neofobia por sí sola no aumentará el uso de recursos novedosos. Un ave que carece de habilidades para moverse en un espacio tridimensional, por ejemplo, es menos capaz de usar una percha segura incluso si está motivada para hacerlo, y un ave con poco miedo y neofobia no explorará un nuevo área de distribución al aire libre a menos que esté motivada para hacerlo.
En conclusión, un entorno de cría que ofrezca opciones de variantes de recursos relevantes puede conducir potencialmente a un bienestar animal positivo al promover la agencia animal y el compromiso afectivo positivo. Proponemos que dicha experiencia aumenta la motivación y la capacidad de las aves para explotar situaciones novedosas, construyendo una agencia que hace más probable que puedan aprovechar al máximo las oportunidades cuando se trasladen a un entorno diferente o se encuentren con nuevos recursos. Los resultados de nuestro estudio sugieren que es importante que las aves jóvenes experimenten la elección de ciertos recursos biológicamente relevantes, como los tipos de percha, durante las primeras 4 semanas de vida. Proporcionar opciones dentro de los tipos de recursos más adelante durante la cría también muestra beneficios. Por lo tanto, proporcionar un entorno de elección múltiple durante todo el período de cría puede sacar provecho de todos los efectos positivos de plasticidad en el desarrollo observados en este estudio.
Declaración de disponibilidad de datos
Los conjuntos de datos generados y los códigos R utilizados para este estudio se pueden encontrar en el repositorio de datos de Mendeley, doi: 10.17632/wczdpy85k3.1.
Declaración ética
El estudio en animales fue aprobado por el Comité Nacional de Ética para Experimentos con Animales en Uppsala, Suecia. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
LS: Redacción – revisión y edición, Redacción – borrador original, Visualización, Supervisión, Metodología, Investigación, Análisis formal, Conceptualización. RH: Metodología, Redacción – revisión y edición, Investigación, Conceptualización. RN: Redacción – revisión y edición, Supervisión, Recursos, Obtención de fondos, Conceptualización. IE: Redacción – revisión y edición, Obtención de fondos, Conceptualización. KM: Redacción – revisión y edición, Metodología, Investigación. LK: Redacción, revisión y edición, Supervisión, Recursos, Administración de proyectos, Metodología, Investigación, Obtención de fondos, Conceptualización.
Financiación
El/los autor/es declaran/n que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. El estudio fue financiado a través de la subvención número 2016-01761 de Formas, el Consejo de Investigación del Gobierno Sueco para el Desarrollo Sostenible. RH contó con el apoyo de una beca de doctorado de la Universidad Noruega de Ciencias de la Vida y la beca de viaje Erasmus+ N AS03.
Reconocimientos
Agradecemos a Yezica Norling, Helen Zobrist y Kai Anna Braun por su ayuda con las observaciones de comportamiento y al personal del Centro Sueco de Investigación Ganadera por cuidar de las aves.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
El/los autor/es declararon, en el momento de la presentación, ser miembro del consejo editorial de Frontiers. Esto no tuvo ningún impacto en el proceso de revisión por pares ni en la decisión final.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1425851/full#supplementary-material
Referencias
1. Grünberger, S, y Leisler, B. Componentes innatos y aprendidos en la selección del hábitat de carboneros carboneros (Parus ater). J Ornithol. (1990) 131:460–4. doi: 10.1007/BF01639825
2. Mettke-Hofmann, C . Movimientos aviares en un mundo moderno: desafíos cognitivos. Anim Cogn. (2017) 20:77–86. doi: 10.1007/s10071-016-1006-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Miller, R, Bugnyar, T, Pölzl, K y Schwab, C. Diferencias en el comportamiento de exploración en cuervos comunes y cuervos carroñeros durante el desarrollo y en el contexto social. Behav Ecol Sociobiol. (2015) 69:1209–20. doi: 10.1007/s00265-015-1935-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Panksepp, J . Conciencia afectiva: sentimientos emocionales centrales en animales y humanos. Cogn. Consciente. (2005) 14:30–80. doi: 10.1016/j.concog.2004.10.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Alcaro, A, Huber, R, y Panksepp, J. Funciones conductuales del sistema dopaminérgico mesolímbico: una perspectiva neuroetológica afectiva. Brain Res Rev. (2007) 56:283–321. doi: 10.1016/j.brainresrev.2007.07.014
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Kimball, MG y Lattin, CR. La exploración de un entorno nuevo no se correlaciona con la neofobia a los objetos en gorriones domésticos capturados en la naturaleza (Passer domesticus). Proceso de comportamiento. (2023) 210:104913. doi: 10.1016/j.beproc.2023.104913
7. Martin, LB, y Fitzgerald, L. Gusto por la novedad en los gorriones domésticos invasores, Passer domesticus. Compórtate Ecol. (2005) 16:702–7. doi: 10.1093/beheco/ari044
8. Sol, D, y Lefebvre, L. La flexibilidad conductual predice el éxito de la invasión en aves introducidas en Nueva Zelanda. Oikos. (2000) 90:599–605. doi: 10.1034/j.1600-0706.2000.900317.x
9. Aplin, LM, Farine, DR, Mann, RP y Sheldon, BC. La personalidad a nivel individual influye en el forrajeo social y el comportamiento colectivo en las aves silvestres. Proc R Soc B Biol Sci. (2014) 281:20141016. doi: 10.1098/rspb.2014.1016
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Mettke-Hofmann, C, Winkler, H, y Leisler, B. La importancia de los factores ecológicos para la exploración y la neofobia en loros. Etología. (2002) 108:249–72. doi: 10.1046/j.1439-0310.2002.00773.x
11. Špinka, M . Agencia animal, conciencia animal y bienestar animal. Anim Welf. (2019) 28:11–20. doi: 10.7120/09627286.28.1.011
12. Nettle, D, y Bateson, M. Plasticidad adaptativa del desarrollo: ¿qué es, cómo podemos reconocerla y cuándo puede evolucionar? Proc R Soc B Biol Sci. (2015) 282:20151005. doi: 10.1098/rspb.2015.1005
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Sellos, JA . Diferencias individuales en plasticidades conductuales. Biol Rev. (2016) 91:534–67. doi: 10.1111/brv.12186
14. Wechsler, B, y Lea, SEG. Adaptación por aprendizaje: su importancia para la ganadería de granja. Appl Anim Behav Sci. (2007) 108:197–214. doi: 10.1016/j.applanim.2007.03.012
15. Widowski, T, y Torrey, S. Cría de aves jóvenes para la adaptabilidad En: T Widowski editor. Bienestar Avícola Avanzado. Cambridge, Reino Unido: Woodhead Publishing (2018)
16. Mayer, U, Bhushan, R, Vallortigara, G, y Lee, SA. Representación de la forma ambiental en el hipocampo de polluelos domésticos (Gallus gallus). Función de estructura cerebral. (2018) 223:941–53. doi: 10.1007/s00429-017-1537-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Morandi-Raikova, A, y Mayer, U. Efecto de la oclusión monocular sobre la expresión de c-Fos en el hipocampo en polluelos domésticos (Gallus gallus). Sci Rep. (2020) 10:7205. doi: 10.1038/s41598-020-64224-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Morandi-Raikova, A, y Mayer, U. La exploración activa de un entorno impulsa la activación del complejo hipocampo-amígdala de los polluelos domésticos. J Exp Biol. (2022) 225:15. doi: 10.1242/jeb.244190
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Skånberg, L, Newberry, RC, Estevez, I, y Keeling, LJ. El cambio o elección del entorno durante la cría temprana mejora la adaptabilidad del comportamiento en los pollitos de gallinas ponedoras. Sci Rep. (2023) 13:6178. doi: 10.1038/s41598-023-33212-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Tahamtani, FM, Nordgreen, J, Nordquist, RE, y Janczak, AM. La vida temprana en un ambiente estéril afecta negativamente la cognición espacial en las gallinas ponedoras (Gallus gallus domesticus). Frente. Vet. Sci. (2015) 2:1–12. doi: 10.3389/fvets.2015.00003
21. Zidar, J, Campderrich, I, Jansson, E, Wichman, A, Winberg, S, Keeling, L, et al. La complejidad ambiental amortigua el sesgo de juicio negativo inducido por el estrés en las gallinas. Sci Rep. (2018) 8:1–14. doi: 10.1038/s41598-018-23545-6
22. Campderrich, I, Nazar, FN, Wichman, A, Marin, RH, Estevez, I, y Keeling, LJ. Complejidad ambiental: un amortiguador contra el estrés en el pollito doméstico. PLoS Uno. (2019) 14:e0210270. doi: 10.1371/journal.pone.0210270
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Campbell, DLM, Hinch, GN, Downing, JA, y Lee, C. Enriquecimiento precoz en gallinas ponedoras criadas en libertad: efectos sobre el comportamiento de pastoreo, el bienestar y la respuesta a los factores estresantes. Animal. (2018) 12:575–84. doi: 10.1017/S1751731117001859
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Brantsæter, M, Tahamtani, FM, Nordgreen, J, Sandberg, E, Hansen, TB, Rodenburg, TB, et al. El acceso a la cama durante la cría y el enriquecimiento ambiental durante la producción reducen el miedo en las gallinas ponedoras adultas. Appl Anim Behav Sci. (2017) 189:49–56. doi: 10.1016/j.applanim.2017.01.008
25. Skånberg, L, Kjærsgaard Nielsen, CB, y Keeling, LJ. El tipo de camada y percha ya es importante desde el principio: explorar las preferencias y el equilibrio de la percha en los polluelos de gallinas ponedoras. Poult Sci. (2021) 100:431–40. doi: 10.1016/j.psj.2020.11.041
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Alcaro, A, y Panksepp, J. La mente BUSCADORA: sustratos neuroafectivos primarios para los estados de incentivo apetitivos y su dinámica patológica en adicciones y depresión. Neurosci Biobehav Rev. (2011) 35:1805–20. doi: 10.1016/j.neubiorev.2011.03.002
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Nazar, FN, Skånberg, L, McCrea, K, y Keeling, LJ. El aumento de la complejidad ambiental al proporcionar diferentes tipos de cama y perchas durante la cría temprana aumenta la capacidad de afrontamiento de los polluelos de aves domésticas. Animales. (2022) 12:1969. doi: 10.3390/ani12151969
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Holt, RV, Skånberg, L, Keeling, LJ, Estevez, I, y Newberry, RC. La elección de los recursos durante la ontogenia mejora el bienestar a corto y largo plazo de las pollitas ponedoras. Sci Rep. (2024) 14:3360. doi: 10.1038/s41598-024-53039-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Aerni, V, Brinkhof, MWG, Wechsler, B, Oester, H y Fröhlich, E. Productividad y mortalidad de gallinas ponedoras en aviarios: una revisión sistemática. Worlds Poult Sci J. (2005) 61:130–42. doi: 10.1079/WPS200450
30. Gunnarsson, S, Yngvesson, J, Keeling, LJ y Forkman, B. La cría sin acceso temprano a las perchas perjudica las habilidades espaciales de las gallinas ponedoras. Appl Anim Behav Sci. (2000) 67:217–28. doi: 10.1016/S0168-1591(99)00125-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Johnsen, PF, Vestergaard, KS y Nørgaard-Nielsen, G. Influencia de las condiciones de cría temprana en el desarrollo del picoteo de plumas y el canibalismo en aves domésticas. Appl Anim Behav Sci. (1998) 60:25–41. doi: 10.1016/S0168-1591(98)00149-X
32. Norman, KI, Adriaense, JEC, y Nicol, CJ. El impacto del enriquecimiento estructural temprano en la cognición espacial en pollos ponedoras. Proceso de comportamiento. (2019) 164:167–74. doi: 10.1016/j.beproc.2019.05.008
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. de Haas, EN, Bolhuis, JE, Kemp, B, Groothuis, TGG, y Rodenburg, TB. Los padres y el entorno de los primeros años de vida afectan el desarrollo conductual de las gallinas ponedoras. PLoS Uno. (2014) 9:577. doi: 10.1371/journal.pone.0090577
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Dixon, LM, Sparks, NHC, y Rutherford, KMD. Las experiencias tempranas importan: una revisión de los efectos del ambiente prenatal en las características de la descendencia en las aves de corral. Poult Sci. (2016) 95:489–99. doi: 10.3382/ps/pev343
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Carter, AJ, Feeney, WE, Marshall, HH, Cowlishaw, G, y Heinsohn, R. Personalidad animal: ¿qué miden los ecólogos conductuales? Biol Rev. (2013) 88:465–75. doi: 10.1111/brv.12007
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Genética Hendrix. Guía de gestión comercial de sistemas alternativos de vivienda. (2020). Disponible en: https://layinghens.hendrix-genetics.com/documents/827/Management_Guide_Alternative_Housing_Systems_L0204-4.pdf.
37. Favati, A, Zidar, J, Thorpe, H, Jensen, P, y Løvlie, H. La ontogenia de los rasgos de personalidad en el gallo selvático rojo, Gallus gallus. Compórtate Ecol. (2016) 27:484–93. doi: 10.1093/beheco/arv177
38. Réale, D, Reader, SM, Sol, C, McDougall, PT, y Dingemanse, NJ. Integrar el temperamento animal dentro de la ecología y la evolución. Biol Rev. (2007) 82:291–318. doi: 10.1111/j.1469-185X.2007.00010.x
39. Forkman, B, Boissy, A, Meunier-Salaün, MC, Canali, E, y Jones, RB. Una revisión crítica de las pruebas de miedo utilizadas en bovinos, porcinos, ovinos, aves de corral y caballos. Physiol Behave. (2007) 92:340–74. doi: 10.1016/j.physbeh.2007.03.016
40. Collias, N, y Jennrich, RI. Las gallinas rojas dominantes de gallinas selváticas (Gallus gallus) en una bandada no confinada crían a la mayor cantidad de crías a lo largo de su vida. Alca. (1994) 111:863–72. doi: 10.2307/4088818
41. Bates, D, Mächler, M, Bolker, BM y Walker, SC. Ajuste de modelos lineales de efectos mixtos utilizando lme4. J Stat Softw. (2015) 67:1–48. doi: 10.18637/jss.v067.i01
42. Halekoh, U, y Højsgaard, S. Una aproximación de Kenward-Roger y métodos de arranque paramétricos para pruebas en modelos mixtos lineales: el paquete de R pbkrtest. J Stat Softw. (2014) 59:1–30. doi: 10.18637/jss.v059.i09
43. Kuznetsova, A, Brockhoff, PB, y Christensen, RHB. Paquete lmerTest: ensayos en modelos lineales de efectos mixtos. J Stat Softw. (2017) 82:1–26. doi: 10.18637/jss.v082.i13
44. Wickham, H . Ggplot2: Gráficos elegantes para el análisis de datos. Nueva York: Springer-Verlag (2016).
45. Lente, R . Medios: medias marginales estimadas, también conocidas como medias de mínimos cuadrados. Versión 1. 4. 1. (2022). Disponible en: https://cran.r-project.org/web/packages/emmeans.
46. EFSA. Dictamen científico sobre los aspectos de bienestar del uso de perchas para gallinas ponedoras. EFSA J. (2015) 13:4131. doi: 10.2903/j.efsa.2015.4131
47. Jones, CT, Pullin, AN, Blatchford, RA, Makagon, MM y Horback, K. Efectos de la cría con estructuras verticales en la ontogenia de la percepción de profundidad en gallinas ponedoras. Appl Anim Behav Sci. (2023) 259:105837. doi: 10.1016/j.applanim.2023.105837
48. Hester, PY, Garner, JP, Enneking, SA, Cheng, HW y Einstein, ME. El efecto de la disponibilidad de perchas durante la cría de pollitas y la puesta de huevos en el comportamiento de las gallinas Leghorn blancas enjauladas. Poult Sci. (2014) 92:1972–80. doi: 10.3382/ps.2013-03008
49. Yan, FF, Hester, PY, y Cheng, HW. El efecto del acceso a la perca durante la cría de pollitas y la puesta de huevos sobre las medidas fisiológicas de estrés en leghorns blancos a las 71 semanas de edad. Poult Sci. (2014) 93:1318–26. doi: 10.3382/ps.2013-03572
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Atkinson, R, Migues, PV, Cammarota, M, Kavanagh, JM, Hunter, M, y Rostas, JAP. Investigaciones bioquímicas, conductuales y electrofisiológicas de la maduración cerebral en pollos. Cerebro Res Bull. (2008) 76:217–23. doi: 10.1016/j.brainresbull.2008.02.036
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Rogers, LJ . Relevancia de la lateralización cerebral y conductual para el bienestar animal. Appl Anim Behav Sci. (2010) 127:1–11. doi: 10.1016/j.applanim.2010.06.008
52. Gunnarsson, S, Keeling, LJ y Svedberg, J. Efecto de los factores de cría en la prevalencia de huevos de suelo, canibalismo cloacal y picoteo de plumas en bandadas comerciales de gallinas ponedoras de alojamiento suelto. Hno. Poult Sci. (1999) 40:12–8. doi: 10.1080/00071669987773
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Lundberg, A, y Keeling, LJ. Impacto de los factores sociales en la anidación en gallinas ponedoras (Gallus gallus domesticus). Appl Anim Behav Sci. (1999) 64:57–69. doi: 10.1016/S0168-1591(99)00020-9
54. Clausen, T, y Riber, AB. Efecto de la heterogeneidad de las cajas nido en la ocurrencia de anidación gregaria en gallinas ponedoras. Appl Anim Behav Sci. (2012) 142:168–75. doi: 10.1016/j.applanim.2012.10.005
55. Krause, ET, y Schrader, L. ¿Alto, bajo o familiar? Preferencias de sitio de anidación de gallinas ponedoras experimentadas. Hno. Poult Sci. (2018) 59:359–64. doi: 10.1080/00071668.2018.1470318
56. Wood-Gush, DGM, y Duncan, IJH. Algunas observaciones sobre el comportamiento de las aves domésticas en estado silvestre. Appl Anim Ethol. (1976) 2:255–60. doi: 10.1016/0304-3762(76)90057-2
57. Cronin, GM, Barnett, TLJL y Hemsworth, PH. La importancia del comportamiento previo a la puesta y de las cajas nido para el bienestar de las gallinas ponedoras: una revisión. Anim Prod Sci. (2012) 52:398–405. doi: 10.1071/AN11258
58. Lawrence, AB, Vigors, B, y Sandøe, P. ¿Qué tiene de positivo el bienestar animal positivo? Una revisión crítica de la literatura. Animales. (2019) 9:783. doi: 10.3390/ani9100783
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Salvanes, AGV, Moberg, O, Ebbesson, LOE, Nilsen, TO, Jensen, KH, y Braithwaite, VA. El enriquecimiento ambiental promueve la plasticidad neuronal y la capacidad cognitiva en los peces. Proc R Soc B Biol Sci. (2013) 280:20131331. doi: 10.1098/rspb.2013.1331
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Zentall, TR . Efecto del enriquecimiento ambiental en el cerebro y en el aprendizaje y la cognición de los animales. Animales. (2021) 11:973. doi: 10.3390/ani11040973
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Cunningham, WA, Dunfield, KA, y Stillman, PE. Estados emocionales a partir de dinámicas afectivas. Emot Rev. (2013) 5:344–55. doi: 10.1177/1754073913489749
62. Paul, ES, Browne, W, Mendl, MT, Caplen, G, Held, S, Trevarthen, A, et al. Trayectorias afectivas: ¿las gallinas están influenciadas por cambios positivos y negativos en sus condiciones de vida? Appl Anim Behav Sci. (2023) 261:105883. doi: 10.1016/j.applanim.2023.105883
63. Collias, NE, y Collias, EC. Un estudio de campo de la gallina roja de la selva en el centro-norte de la India. Cóndor. (1967) 69:360–86. doi: 10.2307/1366199
64. McBride, G, Parer, IP, y Foenander, F. La organización social y el comportamiento de las aves domésticas asilvestradas. Anim Behav Monogr. (1969) 2:125–81. doi: 10.1016/S0066-1856(69)80003-8
65. Englund, MD, y Cronin, KA. Elección, control y bienestar animal: definiciones e indagaciones esenciales para avanzar en la ciencia del bienestar animal. Front Vet Sci. (2023) 10:251. doi: 10.3389/fvets.2023.1250251
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: pollitas, desarrollo conductual, complejidad ambiental, enriquecimiento, camada, perchas, bienestar animal positivo, agencia
Cita: Skånberg L, Holt RV, Newberry RC, Estevez I, McCrea K y Keeling LJ (2024) Aprovechar al máximo la vida: la elección del medio ambiente durante la cría mejora la capacidad de las gallinas ponedoras para aprovechar las oportunidades. Frente. Vet. Sci. 11:1425851. doi: 10.3389/fvets.2024.1425851
Editado por:
Marcella Guarino, Universidad de Milán, Italia
Revisado por:
Dana L. M. Campbell, Organización de Investigación Científica e Industrial de la Commonwealth (CSIRO), Australia
James Edward Brereton, Sparsholt College, Reino Unido
Derechos de autor © 2024 Skånberg, Holt, Newberry, Estevez, McCrea y Keeling. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Lena Skånberg, lena.skanberg@slu.se
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía