
La inyección intramuscular de vitamina A en corderos recién nacidos aumenta la capacidad antioxidante y mejora la calidad de la carne

La inyección intramuscular de vitamina A en corderos recién nacidos aumenta la capacidad antioxidante y mejora la calidad de la carne
 Canción de Pengkang
Canción de Pengkang  Guoqiang Huo Jinxin Feng Weipeng Zhang Xuying Li
Guoqiang Huo Jinxin Feng Weipeng Zhang Xuying Li  Junxing Zhao*
Junxing Zhao*- Facultad de Ciencia Animal, Universidad Agrícola de Shanxi, Taigu, Shanxi, China
Introducción: La vitamina A (VA) y su metabolito, el ácido retinoico (AR) poseen varias funciones biológicas. Este informe investigó si la inyección intramuscular neonatal de VA afectaba la actividad antioxidante y la calidad de la carne en el músculo longissimus dorsi (LD) de corderos.
Métodos: A los corderos se les inyectó 0 (control) o 7.500 UI de palmitato de VA en el músculo bíceps femoral el día 2 después del nacimiento. A las 3, 12 y 32 semanas de edad, se recogieron muestras de sangre en la vena yugular para determinar los niveles séricos de AR y se recogieron muestras musculares en el bíceps femoral para analizar la expresión relativa del ARNm de las enzimas que contribuyen al metabolismo de los retinoides. Todos los animales fueron cosechados a las 32 semanas de edad y se recolectaron muestras musculares para explorar el papel del AV en la calidad de la carne y la capacidad antioxidante de los corderos.
Resultados y discusión: Nuestros resultados indicaron que el AV aumentó el enrojecimiento, la proteína cruda y la grasa bruta (p < 0,05), sin afectar la composición de humedad, cenizas y aminoácidos en el músculo LD (p > 0,05). Además, el AV aumentó la actividad de la catalasa (CAT) y disminuyó los niveles de malondialdehído (MDA) en el músculo LD (p < 0,05). Por su parte, se observaron mayores niveles de ARNm y contenido proteico de CAT y NRF2 con el tratamiento con AV en el músculo LD (p < 0,05), explicado en parte por el aumento del nivel de AR (p < 0,05). En conjunto, nuestros hallazgos indicaron que la inyección de VA al nacer podría mejorar la calidad de la carne de cordero al elevar el enrojecimiento, la proteína cruda, la grasa bruta y la capacidad antioxidante en el músculo LD de los corderos.
Introducción
La carne y sus productos relacionados son fuentes importantes de nutrición para los seres humanos. La calidad de la carne es importante para garantizar la satisfacción del consumidor, que se ve afectada por factores genéticos, métodos de alimentación y el estado nutricional del animal (1). Tradicionalmente, la percepción sensorial humana se utiliza como un factor para evaluar la calidad de la carne, incluida la apariencia, el color, el sabor, la textura (especialmente la ternura), la jugosidad / poder de retención y el olor. Además, la frescura o la salud también es un importante indicador de calidad que se relaciona con la inocuidad de la carne para el consumo (2). En los últimos años, la demanda de carne de cordero ha aumentado constantemente en varias regiones del mundo (3). Dado el mayor contenido de hierro y zinc (3, 4), el deterioro y el cambio de color son más fáciles de producir en la carne de cordero (5). La oxidación de lípidos y proteínas es la razón principal de la disminución del valor nutricional y el deterioro de las propiedades sensoriales y fisicoquímicas (como el color, el sabor o la terneza) de la carne (6, 7). En particular, los corderos mantenidos en cautividad durante largos períodos de tiempo y alimentados con una sola variedad de alimento exacerban aún más el estrés oxidativo en los animales y, en última instancia, conducen a una mala calidad de la carne de cordero (8). Por lo tanto, mejorar la capacidad antioxidante de los corderos y la calidad de la carne se ha convertido en una prioridad en la ganadería ovina de carne.
Numerosos estudios han investigado las funciones biológicas del AV como suplemento nutricional natural, donde la capacidad antioxidante (9), la adipogénesis (10), la miogénesis (11), la función mitocondrial (12) y la regulación de la conversión del tipo de fibra muscular (13) pueden asociarse con una mejor calidad de la carne. Cuando el VA entra en la célula, produce AR bajo la acción de la deshidrogenasa. En el núcleo, la AR se une al heterodímero del receptor de ácido retinoico (RAR) y al receptor X retinoide (RXR) para realizar su función biológica (14).
De hecho, ya en 1932 se informó del AV como antioxidante natural, y su informe indicaba el potencial antioxidante del AV y los carotenoides, los cuales protegen a los lípidos del enranciamiento (9). El retinol es un potente eliminador de radicales peroxi al inhibir la peroxidación en una solución homogénea de linoleato de metilo y en liposomas modelo de fosfatidilcolina (15). Además, en estudios previos, hemos demostrado que la inyección intramuscular de AV neonatal promovió eficazmente el crecimiento muscular de los corderos (16). Sin embargo, no existen estudios relevantes para aclarar si existe una posible relación entre el AV y la calidad de la carne de cordero. Tradicionalmente, el AV se ha añadido a las dietas de los animales como suplemento nutricional (17). Sin embargo, el VA no es completamente absorbido por el ganado y no realiza sus funciones biológicas con este enfoque. En este experimento, inyectamos AV en el músculo para asegurarnos de que el músculo recibe suficiente AV y el efecto es más directo. Por lo tanto, el objetivo de esta investigación fue explorar los efectos de la inyección de AV neonatal sobre la calidad de la carne de cordero y la actividad antioxidante.
Materiales y métodos
Manejo de animales
Todos los procedimientos experimentales fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Agrícola de Shanxi (sxnd202028). Seleccionadas al azar 80 ovejas Hu de raza pura con estados corporales similares, todas ellas estaban en su tercera preñez. Para evitar la influencia del padre, todas las ovejas fueron inseminadas artificialmente con semen de un carnero Dorper después del celo simultáneo. Se debe prestar atención a la operación y la higiene durante la inseminación. Después de la preñez de la oveja, cada 3 ovejas preñadas fueron colocadas en un establo. Las ovejas eran alimentadas en los establos donde se les daba agua limpia para beber, y eran libres de moverse después de alimentarse. La dieta se formuló de acuerdo con los requisitos del Consejo Nacional de Investigación (18) para la nutrición de las ovejas. Durante el período experimental, el manejo de la alimentación y el ambiente de las ovejas de cada grupo se mantuvieron al mismo nivel y se esterilizaron regularmente para mantener el establo seco e higiénico. A los 35 d de gestación, se determinó el número fetal mediante un monitor ecográfico. Para el estudio de seguimiento, solo se utilizaron ovejas con 2 fetos. Después del nacimiento, seleccionamos corderos gemelos, ambos machos, con un peso de 3,5 ± 0,5 kg, y los distribuimos a los grupos control y VA para garantizar la misma condición corporal de los corderos (n = 8 en cada grupo).
Con base en estudios previos (13, 19), determinamos inyectar 7.500 UI de palmitato de VA (producto no. PHR1235, Sigma, Milwaukee, EE. UU.) o un volumen equivalente de aceite de maíz (producto n.º c8267, Sigma, Milwaukee, EE. UU.) en el músculo bíceps femoral el día 2 después del nacimiento. A los corderos se les administraron inyecciones semanales en un momento fijo durante 3 semanas y se emparejaron con ovejas para su manejo. Los corderos fueron destetados a las 12 semanas de edad y alimentados con una dieta de fondo durante 55 días, seguida de una dieta de acabado con libre acceso a agua limpia y bloques de sal. Además, se agregó heno de pasto (plántulas de maní) para promover el crecimiento de los corderos durante el período de engorde. La composición de nutrientes de los piensos concentrados y del heno de hierba ha sido reportada en otro manuscrito (16).
Para las plántulas de maní, se utilizó el método AOAC (20) para determinar el contenido de MS, y el método de Van Soest para determinar los contenidos de FDN y FDA (21). Se utilizaron los métodos de extracción Kjeldahl y Soxhlet para medir el contenido de proteína bruta y grasa bruta, respectivamente (22). El contenido de cenizas totales en las muestras se midió después de 40 min de carbonización en un crisol a temperatura constante a 600°C. Todos los animales fueron cosechados a las 32 semanas de edad.
Obtención y tratamiento de biopsias séricas y musculares
Se recogieron muestras de sangre de corderos de la vena yugular en la 3ª, 12ª y 32ª semana. El suero se separó y almacenó a -80 °C para evitar la degradación de los componentes del suero. Después de determinar el método de detección, la AR en suero se detectó mediante cromatografía líquida de alta resolución (HPLC) con una columna de fase reversa. (Atlantis dC18, 5 μm, 100 A, Estados Unidos), y su contenido se determinó a partir de la curva patrón de la muestra patrón (#B21287, Shanghái, China). El método fue el siguiente: se utilizó metanol como fase móvil con un caudal de 1,0 mL/min, longitud de onda de detección de 325 nm, temperatura de columna de 35 °C, inyección de 20 μL (23). Además, el contenido de hexoquinasa (HK) y lactato deshidrogenasa (LDH) en suero y las actividades de capacidad antioxidante total (T-AOC), superóxido dismutasa total (T-SOD), catalasa (CAT), glutatión peroxidasa 4 (GPx4) y el contenido de malondialdehído (MDA) en el músculo LD fueron detectados por kits. A continuación se proporciona información sobre los kits: HK (#A077-3-1), LDH (#A020-2-2), T-AOC (#A015-2-1), catalasa (#A007-1-1), Gpx4 (#H545-1-1), SOD (#A001-3-2) y MDA (#A003-1-2) se adquirieron del Instituto de Bioingeniería Nanjing Jiancheng (Nanjing, Jiangsu, China).®
Se tomaron muestras de músculo esquelético de corderos a las 3 y 12 semanas de edad utilizando una aguja de biopsia. Se extrajo una pequeña área de lana del músculo bíceps femoral de la pata trasera derecha del cordero con una afeitadora y se limpió con yodoforo. Los corderos se fijaron y el músculo se extirpó insertando una aguja de biopsia en el bíceps femoral y se colocó en nitrógeno líquido para la extracción de ARN (q-RT-PCR).
Análisis de la calidad de la carne
Después de la cosecha, se extrajo el músculo LD de los corderos y se analizó el pH, el color de la carne, la conductividad, la fuerza de cizallamiento, la pérdida por cocción y la pérdida por goteo. El pH se midió 45 min después de que el cordero dejara de respirar (pH 45 min) y de nuevo 24 h después de dejar de respirar (pH 24 h) utilizando un medidor de pH-STAT (SFK-Technology, Dinamarca). El color de la carne (L*, a* y b*) y la conductividad se midieron simultáneamente con el pH, utilizando un medidor de conductividad de la canal y un medidor de color de la carne LAB (Matthäus, Klausa, Alemania). Para el análisis de la pérdida por goteo, el músculo al que se le había quitado la fascia se recortó en tiras, se pesó y se colgó en ganchos. Las tiras se colocaron en un tubo centrífugo de 50 mL, evitando el contacto entre las muestras y la pared del tubo, y se almacenaron a 4°C durante 48 h; Se retiraron las tiras, se absorbió el agua de la superficie de la tira con papel de filtro y se pesó cada tira. Para la determinación de la fuerza de cizallamiento, la muestra de músculo se envejeció a 4 °C durante 72 h, luego se expuso a un baño de agua termostático a 80 °C y se calentó hasta que la temperatura central del músculo alcanzó los 70 °C. Muestras musculares de 3 cm de longitud y 1 cm2 en la sección transversal se cortaron a partir de muestras enfriadas a temperatura ambiente en la dirección de las fibras musculares y se midió la fuerza de cizallamiento, expresada en «N», utilizando un medidor de cizallamiento (Mecmesin, West Sussex, Reino Unido). Se pesaron aproximadamente 100 g de músculo para medir la pérdida de cocción. El método fue el siguiente: la muestra se despojó de su membrana externa y se le adhirió grasa, se pesó y se cocinó al vapor en agua hirviendo durante 30 min. Las muestras de músculo cocido se colgaron en un lugar fresco durante 30 minutos y se pesaron.
Análisis de nutrientes cárnicos
Se utilizaron métodos reconocidos de AOAC (24) para analizar la humedad, la proteína bruta, el extracto de éter y el contenido total de cenizas del músculo LD. Específicamente, el contenido de humedad se calculó horneando muestras de peso constante a 105 ° C. Se utilizaron los métodos de extracción Kjeldahl y Soxhlet para medir el contenido de proteína cruda y extracto de éter, respectivamente (22). El contenido de cenizas totales en las muestras se midió después de 40 min de carbonización en un crisol a temperatura constante a 600°C.
Tinción de ácido peryódico muscular-Schiff y análisis de glucógeno
Las muestras de tejido muscular se fijaron en PFA al 4% durante 2 días, se deshidrataron en serie mediante etanol de gradiente y xileno, luego se sumergieron en parafina durante 9 h y se incluyeron en parafina. A continuación, se utilizó un micrótomo (Leica, Alemania) para cortar las muestras a un grosor de 5 μm y las secciones se desparafinaron secuencialmente, se rehidrataron y se tiñeron en una solución de tinción de ácido peryódico-Schiff (PAS) (n.º de cat. G1008, Servicebio, Wuhan, China), luego se enjuagaron con agua corriente, se deshidrataron, se transparentaron, se sellaron y, finalmente, se observaron con un microscopio (microscopio DMi8, Leica, Alemania) para la adquisición y análisis de imágenes (todas las muestras de tejido muscular se cortaron a intervalos de 50 μm, con al menos 3 repeticiones en cada muestra). Para medir el contenido de glucógeno, las muestras de músculo LD se trituraron y homogeneizaron en solución salina al 0,9% mediante un homogeneizador, se centrifugaron (2.500 × g, 10 min, 4°C) y se extrajo el sobrenadante para medir la concentración de glucógeno con un kit (#A043-1-1, Instituto de Bioingeniería Nanjing Jiancheng, Jiangsu, China).
Tinción de Masson Muscular y análisis de colágeno
Las muestras de músculo se incrustaron en un compuesto de temperatura óptima de corte (OCT) y se cortaron en secciones de 10 μm de espesor con un micrótomo de criostato (Leica, Alemania). Las secciones congeladas se desparafinaron secuencialmente, se rehidrataron y se tiñeron en una solución de tinción de Masson (n.º de cat. G1006, Servicebio, Wuhan, China), luego se enjuagaron, se deshidrataron, se sellaron y finalmente se observaron con un microscopio (microscopio DMi8, Leica, Alemania) para la adquisición y análisis de imágenes (todas las muestras de tejido muscular se cortaron a intervalos de 50 μm, con al menos 3 réplicas en cada muestra). Además, se analizó la expresión relativa de las fibrillas de colágeno tipo I, tipo III y fibronectina.
Composición de aminoácidos
Determinación del contenido de aminoácidos en músculo LD de corderos mediante cromatografía líquida de alta resolución (HPLC). El método fue el siguiente: fase móvil (A: CH₃COONa; B: CH3OH, H2O), caudal 1,0 mL/min, longitud de onda de detección 360 nm, temperatura de columna 40 °C, inyección 20 μL. El contenido de aminoácidos en la LD se determinó a partir de la curva estándar basada en la muestra estándar (AAS18-10ML, Sigma, Milwaukee, Estados Unidos).
PCR cuantitativa en tiempo real
El ARN total se extrajo del tejido muscular utilizando el reactivo Trizol (Sigma, Saint Louis, MO) y se sintetizó en ADNc utilizando un kit de transcripción inversa (TAKARA Co, Ltd., Dalian, China). Para la q-RT-PCR se utilizaron el sistema de detección CFX RT-PCR (Bio-Rad, Hercules, CA) y el kit SYBR Green RT-PCR (TAKARA Co, Ltd.). El procedimiento fue el siguiente: 95°C, 10 min; 45 ciclos de 2 pasos de 95 °C, 15 s, 60 °C, 30 s, con al menos 3 repeticiones por serie. Las secuencias de cebadores se muestran en la Tabla 1. Los cambios relativos de la expresión génica se calcularon mediante 2−ΔΔCt (8), y se utilizó β-actina como gen de limpieza. La expresión génica en el grupo VA mostró un cambio de pliegue en comparación con el grupo control.
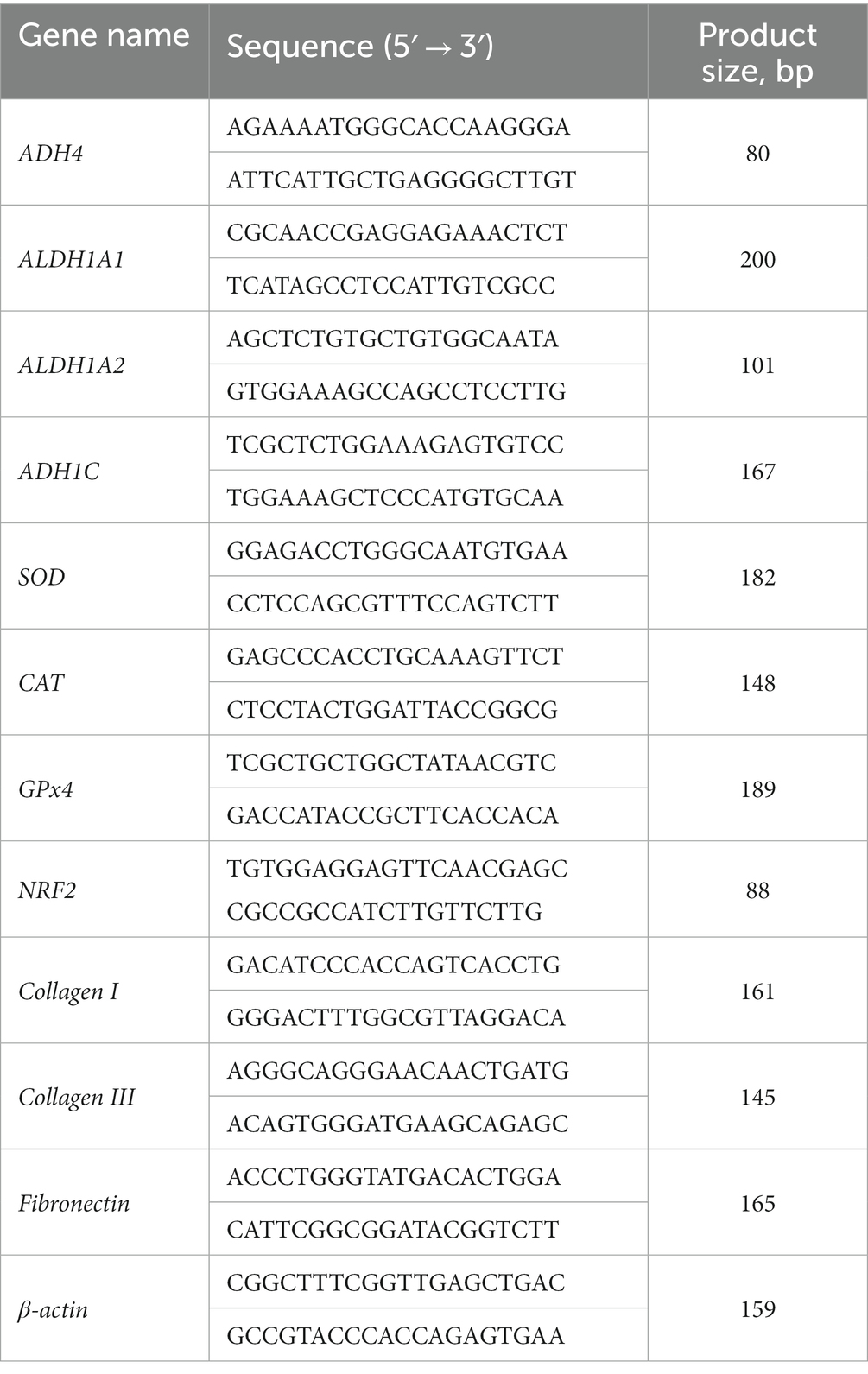 Tabla 1. Secuencias de cebadores para PCR en tiempo real.
Tabla 1. Secuencias de cebadores para PCR en tiempo real.
Western blot
Las muestras de músculo se molieron en nitrógeno líquido y se lisaron con lisado de RIPA (1% NaF, 1% Na3VO4, PMSF al 1%, β-mercaptoetanol al 2%, inhibidor de la proteasa al 0,1%, volumen constante del tampón de carga de 1 × a 10 ml) durante 30 min, luego hervido a 100 °C durante 10 min, centrifugado a 12.000 × g durante 8 min a 4 °C y el sobrenadante eliminado como proteína aislada. Las proteínas extraídas se separaron mediante SDS-PAGE (temperatura ambiente, 80 V durante 0,5 h, 120 V durante 1,5 h) y luego se transfirieron a membranas de nitrocelulosa (4 °C, 100 V durante 2 h), bloqueadas con leche desnatada en polvo al 5% (Shanghai Sanger Biotechnology Co., Ltd., Shanghai, China) durante 1 h. Finalmente, las membranas de nitrocelulosa se incubaron con el anticuerpo primario (4°C, durante la noche) y el anticuerpo secundario correspondiente (temperatura ambiente, 1 h). Se utilizó el sistema de imágenes infrarrojas Odyssey para visualizar las bandas de proteínas, y la densidad de bandas se estandarizó al contenido de β-tubulina.
Los anticuerpos contra SOD (n.º sc-8637), GPx4 (n.º sc-50497) y catalasa (n.º sc-34281) se compraron a Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, Estados Unidos). NRF2 (n.º bs-1074R) y β-tubulina (bsm-33034 M) procedían de Biosynthesis Biotechnology Co., Ltd. (Pekín, China). Un anticuerpo secundario anti-conejo de cabra (926-32.211) fue de LI-COR Biosciences (Lincoln, NE) y un anticuerpo secundario anti-cabra de burro (no. D110120) fue comprado de Sangon Biotech Co., Ltd. (Shanghai, China).
Análisis estadístico
Para el análisis estadístico se utilizó el software Graphpad Prism 9 (Monrovia, CA, Estados Unidos). Los corderos utilizados en el experimento no fueron asignados completamente al azar, por lo que utilizamos la prueba t emparejada para analizar los datos. Los resultados se mostraron como la media ± SEM. p < 0,05 se consideró significativa entre los grupos control y AV.
Resultados
Contenido de ácido retinoico en la expresión sérica y de ARNm de las enzimas metabolizadoras de retinol en el músculo longissimus dorsi
Como se muestra en la Figura 1A, la inyección de AV elevó significativamente el contenido de AR en el suero de corderos en la 3ª (Figura 1A, p < 0,01) y en la 12ª (Figura 1A, p < 0,05) semanas. Consistentemente, el nivel de ARNm de las enzimas metabolizadoras de retinol en el grupo VA mostró la misma tendencia en el músculo LD de los corderos. En concreto, no hubo diferencia en la ADH1C entre el grupo AV y el grupo control (Figura 1B, p > 0,05), mientras que la expresión de ADH4 en el grupo VA fue mayor que la del grupo control a la edad de 3 semanas (Figura 1C, p < 0,05). Además, el AV aumentó el nivel de ALTH1A1 a los 3 años (Figura 1D, p < 0,01) y a las 12 semanas (Figura 1D, p < 0,05), y el AV aumentó el nivel de ALTH1A2 a los 3, 12 semanas y cosecha (Figura 1E, p < 0,05).
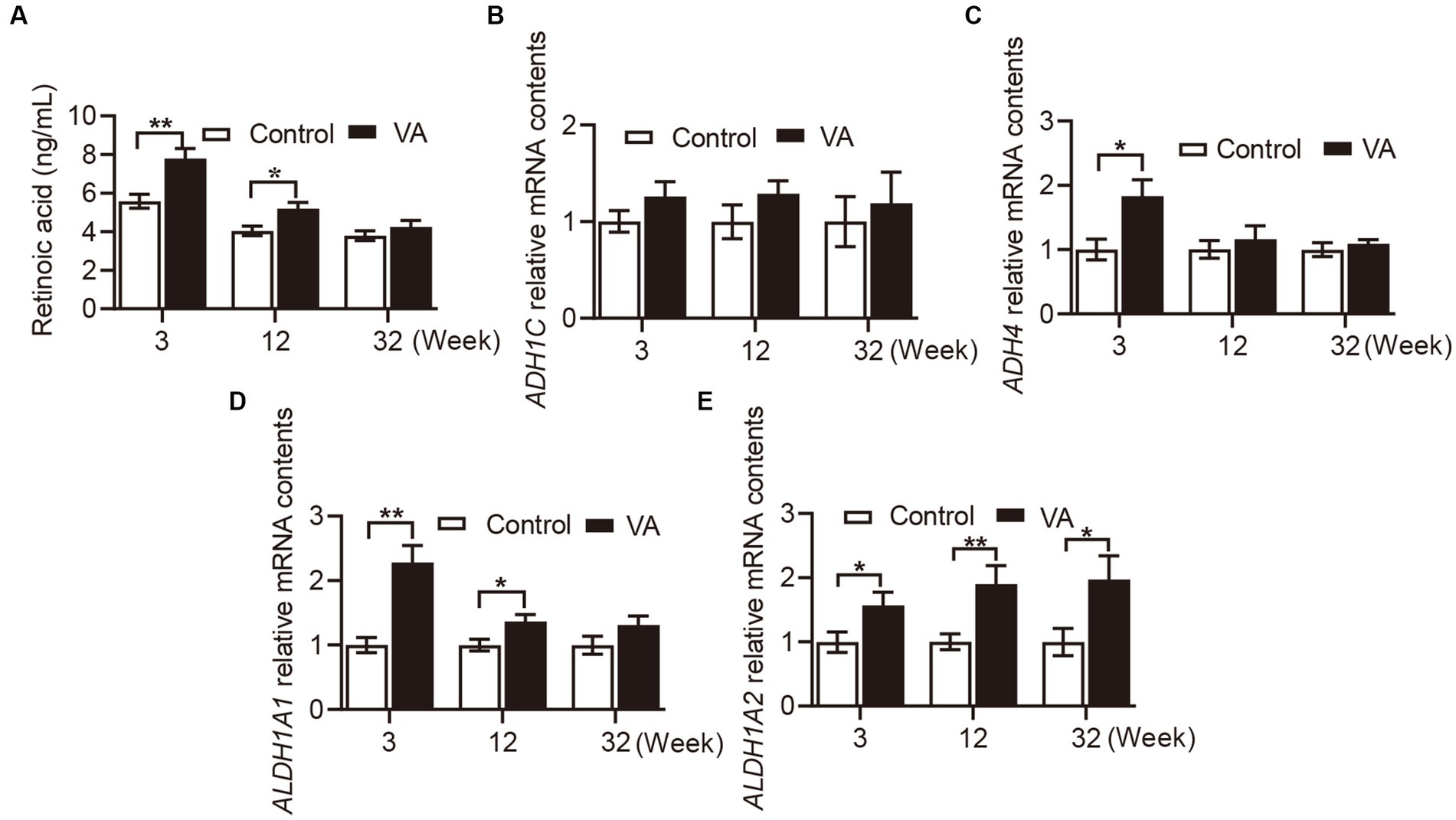 Figura 1. Efectos de la inyección de vitamina A sobre el contenido de AR en suero y la expresión de ARNm de enzimas metabolizadoras de retinol en el músculo LD de corderos. (A) Contenido de AR en suero. (B) ARNm relativo de ADH1C. (C) ARNm relativo de ADH4. (D) ARNm relativo de ALDH1A1. (E) ARNm relativo de ALDH1A2. (Media ± SEM; n = 8 en cada grupo, *p < 0,05 y **p < 0,01).
Figura 1. Efectos de la inyección de vitamina A sobre el contenido de AR en suero y la expresión de ARNm de enzimas metabolizadoras de retinol en el músculo LD de corderos. (A) Contenido de AR en suero. (B) ARNm relativo de ADH1C. (C) ARNm relativo de ADH4. (D) ARNm relativo de ALDH1A1. (E) ARNm relativo de ALDH1A2. (Media ± SEM; n = 8 en cada grupo, *p < 0,05 y **p < 0,01).
Calidad de la carne y composición nutricional en el músculo longissimus dorsi
No hubo alteraciones en el pH, el color de la carne (valores de L* y b*), la conductividad, la fuerza de cizallamiento, la pérdida por cocción y la pérdida por goteo en el músculo LD entre el AV y el grupo control de corderos (Tabla 2, p > 0,05). Sin embargo, los corderos del grupo VA exhibieron una mayor a*24h valor superior al del grupo control (Tabla 2, p < 0,05). Además, la inyección de AV no tuvo efectos sobre el contenido de humedad y cenizas del músculo LD (Tabla 3, p > 0,05), pero elevó el contenido de proteína bruta y grasa bruta (Tabla 3, p < 0,05).
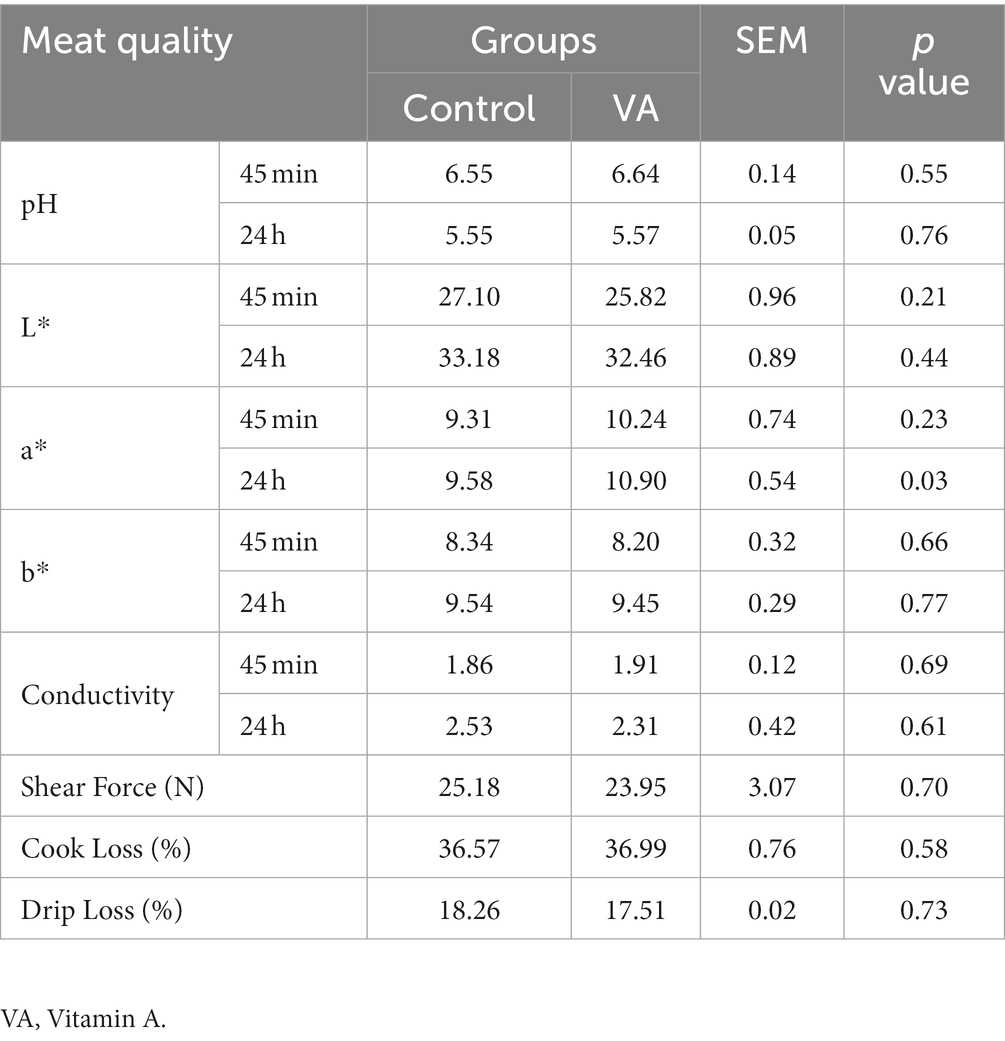 Tabla 2. Efectos de la inyección de vitamina A en la calidad de la carne de cordero.
Tabla 2. Efectos de la inyección de vitamina A en la calidad de la carne de cordero.
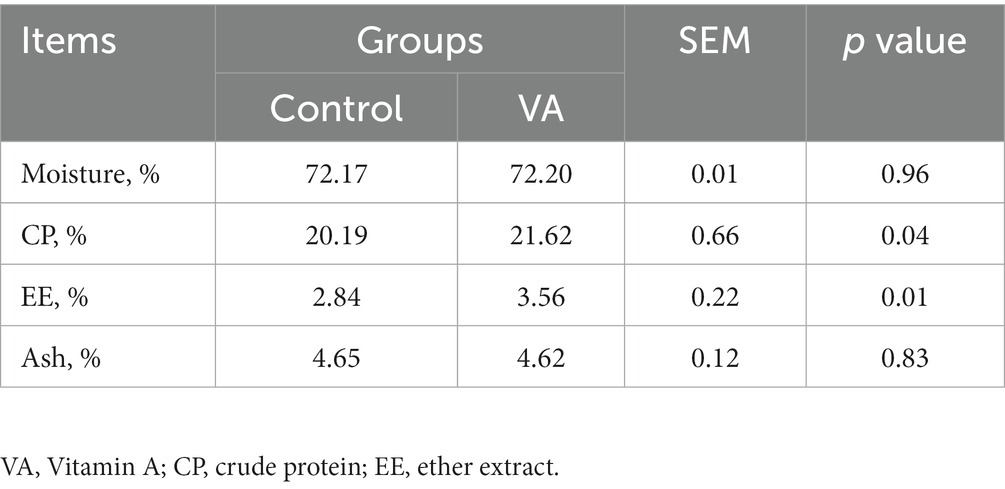 Tabla 3. Composición nutricional en el músculo longissimus dorsi de corderos.
Tabla 3. Composición nutricional en el músculo longissimus dorsi de corderos.
Abundancia de glucógeno y colágeno en el músculo longissimus dorsi y contenido de hexoquinasa, lactato deshidrogenasa en suero
Como se muestra en la Figura 2 y Tabla 4, la AV no afectó el contenido de glucógeno en el músculo LD (Figura 2 y Tabla 4, p > 0,05) y el contenido de HK y LDH en suero (Tabla 4, p > 0,05) de los corderos. En cuanto al contenido de colágeno en el músculo LD de los corderos, tampoco hubo diferencia entre los grupos control y AV (Figuras 3A, B, p > 0,05).
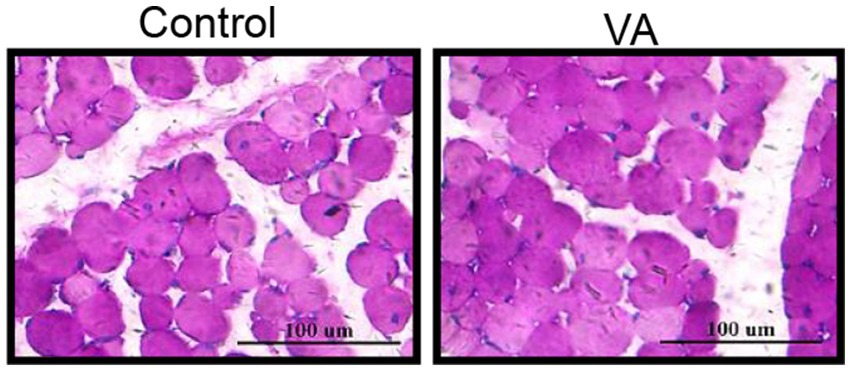 Figura 2. Efectos de la inyección de vitamina A sobre el contenido de glucógeno en el músculo LD de corderos. Tinción de glucógeno PAS del músculo LD. (Media ± SEM; n = 8 en cada grupo).
Figura 2. Efectos de la inyección de vitamina A sobre el contenido de glucógeno en el músculo LD de corderos. Tinción de glucógeno PAS del músculo LD. (Media ± SEM; n = 8 en cada grupo).
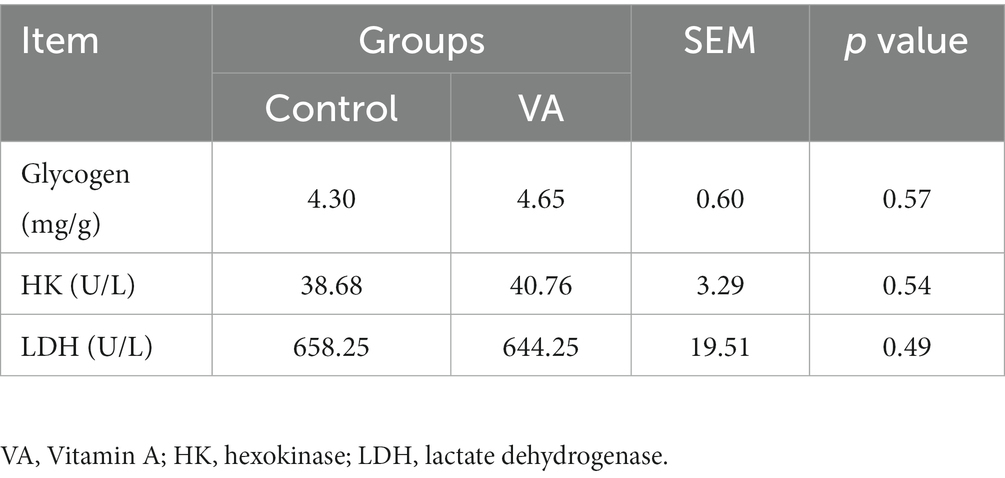 Tabla 4. El contenido de glucógeno en el músculo longissimus dorsi, la hexoquinasa y el contenido de lactato deshidrogenasa en suero de corderos.
Tabla 4. El contenido de glucógeno en el músculo longissimus dorsi, la hexoquinasa y el contenido de lactato deshidrogenasa en suero de corderos.
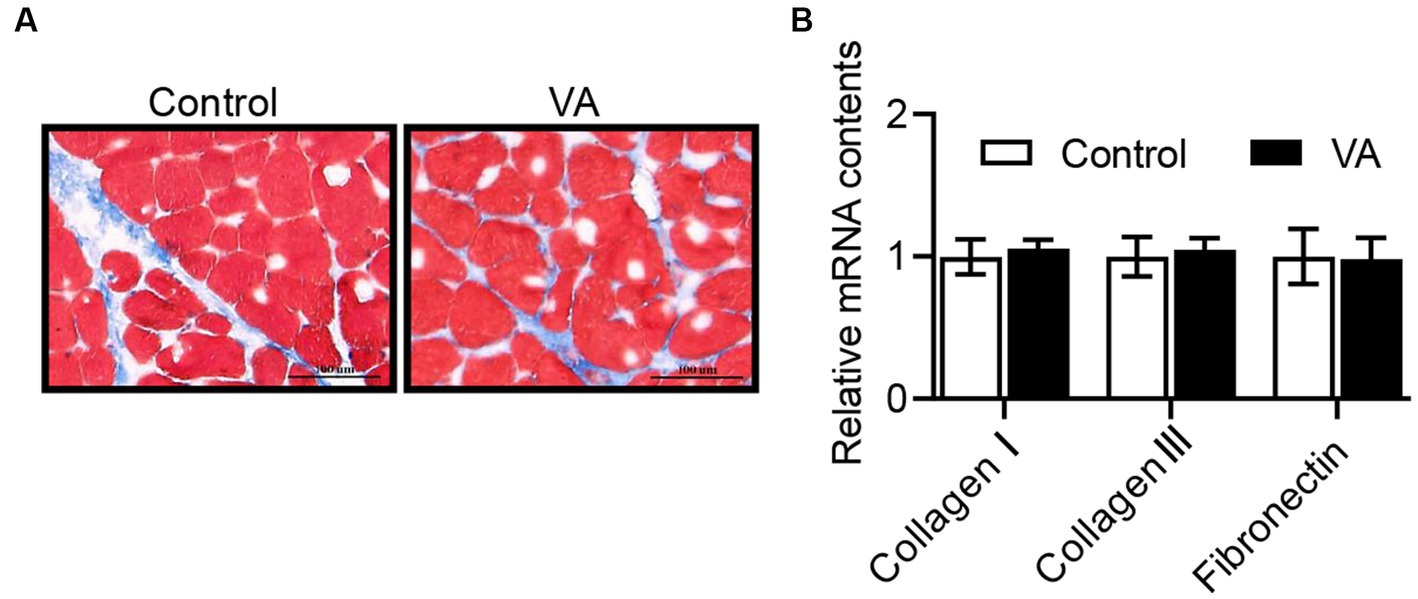
Figura 3. Efectos de la inyección de vitamina A sobre el contenido de colágeno en el músculo LD de corderos. (A) Tinción de Masson del músculo LD. (B) ARNm relativos de colágeno I, colágeno III y fibronectina. (Media ± SEM; n = 8 en cada grupo).
Composición de aminoácidos en el músculo longissimus dorsi
En comparación con el grupo control, la inyección de AV no tuvo efectos sobre la composición de aminoácidos y el total de aminoácidos en el músculo LD de los corderos (Tabla 5, p > 0,05).
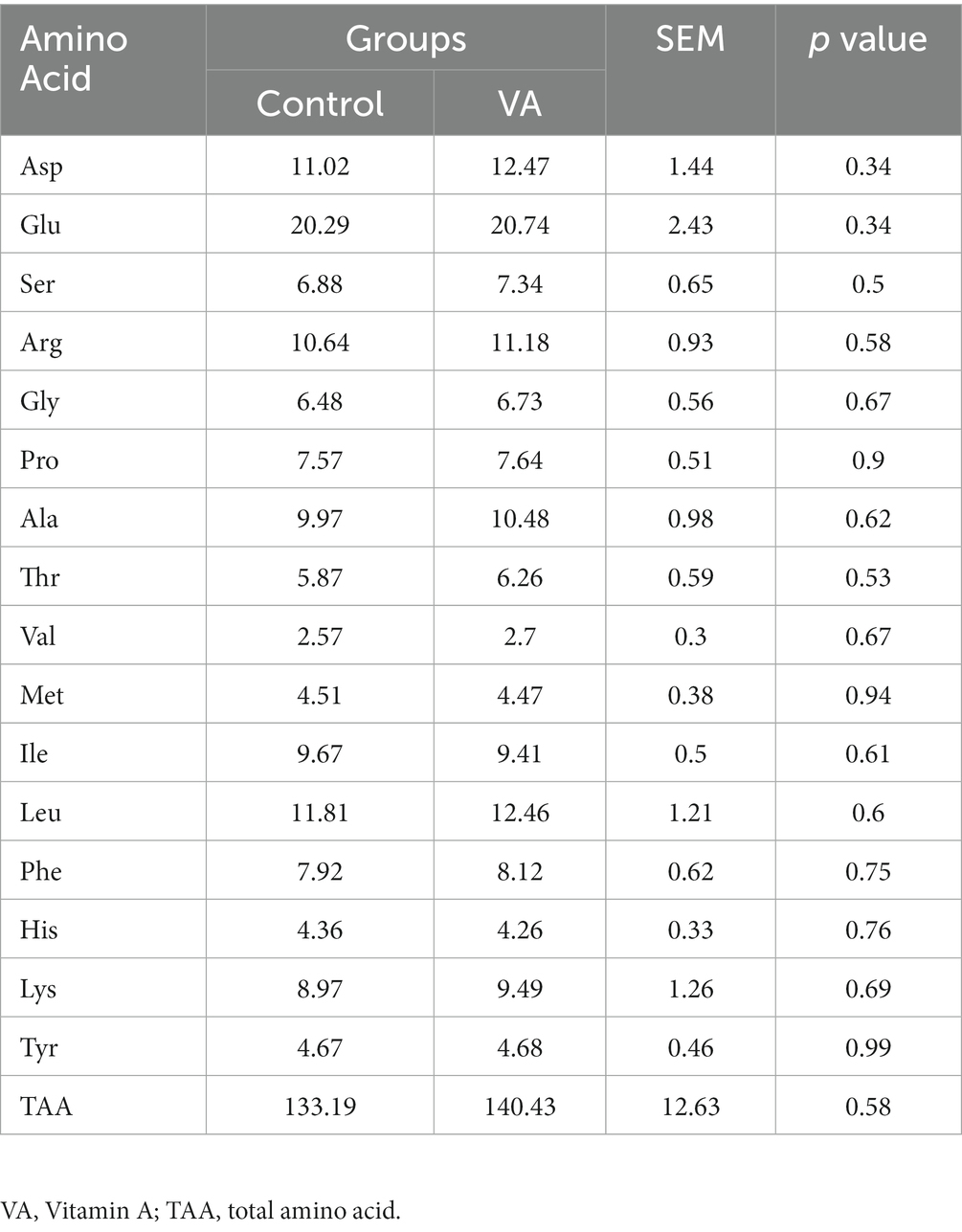
Tabla 5. Composición de aminoácidos en el músculo longissimus dorsi de corderos (mg/g).
Actividades enzimáticas antioxidantes y contenido de malondialdehído
En comparación con el grupo control, la AV aumentó la actividad de CAT y redujo el contenido de MDA en el músculo LD de los corderos (Tabla 6, p < 0,05). Además, los datos revelaron que la expresión de ARNm de CAT y NRF2 en el músculo LD fue mayor en el grupo VA, y la abundancia de proteínas también fue significativamente elevada (Figuras 4A-E, p < 0,05).
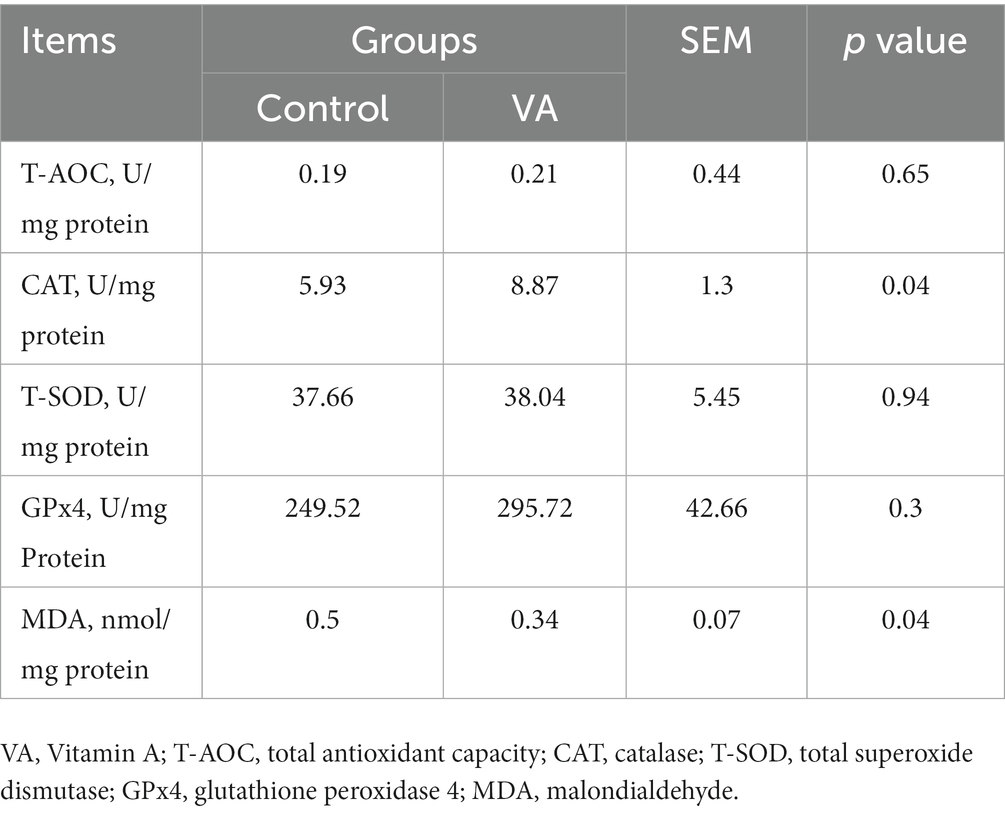 Tabla 6. Actividad de enzimas antioxidantes y contenido de malondialdehído en el músculo longissimus dorsi de corderos.
Tabla 6. Actividad de enzimas antioxidantes y contenido de malondialdehído en el músculo longissimus dorsi de corderos.
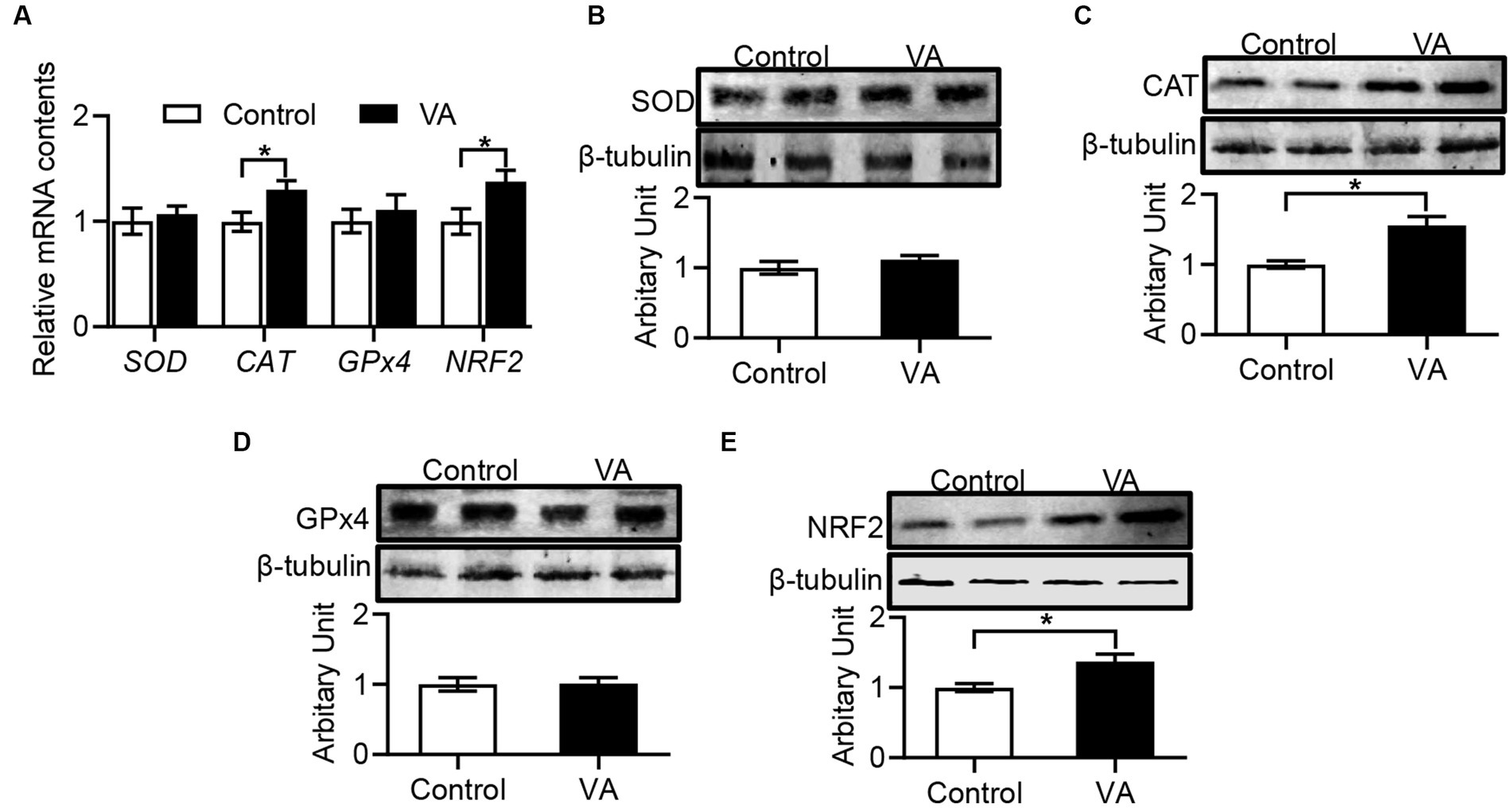
Figura 4. Efectos de la inyección de vitamina A sobre la actividad antioxidante en el músculo LD de corderos. (A) ARNm relativos de SOD, CAT, GPx4 y NRF2. (B) Abundancia de proteína SOD. (C) Abundancia de proteína CAT. (D) Abundancia de proteína GPx4. (E) Abundancia de proteína NRF2. (Media ± SEM; n = 8 en cada grupo, *p < 0,05).
Discusión
El AV se ha utilizado ampliamente como suplemento nutricional en la cría de animales. En el presente estudio, encontramos que la inyección intramuscular de AV aumentó el contenido de AR en suero de corderos a las 3 semanas de edad y al destete. Se sabe que la artritis reumatoide es el metabolito más activo de la AV, que media las funciones metabólicas de la AV, excepto la visión. Para explicar este fenómeno, examinamos la expresión de ARNm de enzimas que catalizan la síntesis de AR, ya que la síntesis de AR depende principalmente de la acción catalítica de la alcohol deshidrogenasa (ADHS) y la aldehído deshidrogenasa (ALDH). Sin embargo, los datos mostraron que la expresión de las enzimas no parecía estar claramente relacionada con el contenido de AR, ya que exhibían diferentes niveles en diferentes estadios entre los grupos control y AV, lo que fue consistente con los hallazgos de Harris et al. (19). De hecho, se ha demostrado que la AR disminuye la sensibilidad de las neuronas embrionarias de pollo (25, 26), las células PC12 (27) y las células mesangiales (28) al estrés oxidativo. Por lo tanto, estos resultados sugirieron que parte del efecto del AV sobre la calidad de la carne podría atribuirse a la síntesis de altos niveles de AR.
El pH de la canal después de la cosecha depende del metabolismo del glucógeno en el músculo esquelético, y el contenido de glucógeno se correlaciona positivamente con el valor del pH de la carne (29). Además, la LDH es una oxidorreductasa clave en la vía glucolítica de los organismos que cataliza la formación de ácido lactato, y su actividad se correlaciona negativamente con el pH. En el presente estudio, la inyección de AV no alteró ni el glucógeno en el músculo LD ni el HK y LDH en suero en este ensayo, lo que indica que la AV no afectó el pH final de la carne de cordero. Teniendo en cuenta que el pH final es un factor importante que afecta a la calidad de la carne, que está fuertemente asociado con la capacidad de retención de agua y la terneza (30-32), estos datos podrían explicar por qué la inyección intramuscular de AV no afectó a la capacidad de retención y a la terneza. De hecho, la suplementación dietética de antioxidantes naturales en animales no afecta el pH de la carne. Por ejemplo, la adición de extracto de semilla de uva a la dieta no tuvo ningún efecto sobre el pH final del músculo porcino (33). En corderos, la adición dietética de extracto de trigo sarraceno tampoco afectó el pH del músculo Longissimus thoracis et lumborum (8). Además, la inyección intramuscular de AV no afectó la conductividad, la pérdida de cocción y la pérdida por goteo del músculo LD, lo que puede atribuirse al valor de pH constante.
El color de la carne se considera uno de los indicadores para juzgar la salud de la carne fresca, y el enrojecimiento de la carne juega un papel clave en la determinación de las decisiones de los consumidores (34). En el estudio actual, la inyección de VA aumentó significativamente el enrojecimiento muscular LD de los corderos, como lo demuestra un aumento en el valor de a*. Del mismo modo, estudios anteriores han demostrado que la suplementación con antioxidantes naturales aumenta el enrojecimiento de la carne. Por ejemplo, la alimentación con pastos abundantes en antioxidantes naturales mejoró el enrojecimiento de la carne (35). La suplementación de los cerdos de engorde con resveratrol o extracto de proantocianidina de semilla de uva mejoró el enrojecimiento de la carne de cerdo (36, 37). De hecho, el color de la carne está estrechamente relacionado con el nivel de mioglobina. Kim et al. informaron que el aumento del enrojecimiento en la carne de cerdo fue causado por la alta concentración de mioglobina en el músculo (38). Cuando la carne fresca se expuso al aire, la mioglobina en el músculo LD se oxidó a oximioglobina, lo que le dio a la carne un color rojo cereza brillante, que es lo que los consumidores perciben como el color de la frescura (39). Además, el color de la carne está altamente correlacionado con la composición del tipo de fibra muscular (37). Las fibras musculares en mamíferos adultos se dividen en cuatro tipos principales: MyHC I, MyHC IIa, MyHC IIb y MyHC IIx (40, 41). Se ha sugerido que el mayor valor de a* siempre se atribuyó a una alta proporción de miofibra tipo I (38). Consistentemente, la miofibra tipo I es fibra muscular oxidada con un color de carne más rojo y tiende a tener concentraciones de mioglobina más altas. Sin embargo, es necesario explorar más a fondo si la inyección de VA afectó la composición del tipo de fibra muscular.
En el ganado vacuno, el contenido de grasa intramuscular afectó en gran medida la ternura y el sabor de la carne de res (42, 43), mientras que la inyección intramuscular de AV aumentó el contenido de grasa intramuscular a través de la regulación positiva de la expresión de Zfp423, un factor de transcripción clave que regula la adipogénesis (19). Consistentemente, se observó una mayor EE en el músculo LD de los corderos inyectados con VA. Sin embargo, no observamos la alteración en la fuerza de cizallamiento muscular, lo que implica que la AV no afectó la terneza de la carne de cordero. De hecho, la ternura de la carne está determinada por numerosos factores, como la raza y la edad del animal (44), el contenido de colágeno intramuscular y la longitud del sarcómero (45), el contenido de grasa intramuscular y el pH final (46). Curiosamente, observamos que el AV aumentaba el contenido de proteína bruta en el músculo LD, en línea con un estudio previo, que mostraba que el AV promovía el crecimiento del músculo esquelético (16).
La oxidación lipídica es una de las principales causas de deterioro y deterioro de la carne y los productos cárnicos (47). El sistema antioxidante es esencial para proporcionar protección contra el daño oxidativo y puede ser activado por una variedad de sustancias biológicamente activas y genes relacionados con los antioxidantes (48). Varios estudios han demostrado que los antioxidantes naturales pueden mejorar la calidad de la carne al reducir la peroxidación lipídica y mejorar el estado antioxidante (49, 50). Por ejemplo, la adición dietética de isoflavonas de soja mejora la calidad de la carne al aumentar la capacidad antioxidante en pollos de engorde machos (51). En los cerdos, la suplementación con resveratrol en la dieta también mejora la calidad de la carne de cerdo al aumentar la capacidad antioxidante (37). En este ensayo, el tratamiento con AV aumentó significativamente la actividad de la CAT y disminuyó el contenido de MDA en el músculo LD de los corderos. La catalasa existe en casi todos los organismos expuestos al oxígeno (por ejemplo, bacterias, plantas y animales) y cataliza la descomposición del peróxido de hidrógeno en agua y oxígeno (52). De hecho, la catalasa se ha utilizado para fabricar papel de envolver alimentos con el fin de evitar la oxidación de los alimentos en la industria alimentaria (53). Una investigación previa ha demostrado que la sobreexpresión de catalasa redujo el estrés oxidativo en ratones envejecidos (54). La producción de MDA también se ha utilizado como biomarcador para evaluar el estrés oxidativo de los organismos (55). Además, la señalización de Keap1-NRF2 también es importante para regular el sistema antioxidante, y Keap1 desempeña un papel regulador negativo en la activación de la expresión de genes antioxidantes como CAT, SOD1 y NRF2 (48, 56). Zhao et al., demostraron que la suplementación dietética de extracto de trigo sarraceno tártaro aumentó significativamente la abundancia de proteínas de Gpx4 y Nrf2 y disminuyó el contenido de malondialdehído en el músculo esquelético, mejorando así la capacidad antioxidante del músculo de cordero (8). Xu et al., encontraron que la inclusión de extracto de proantocianidina de semilla de uva en el alimento de cerdos de engorde afectó la expresión de ARNm de genes relacionados con antioxidantes, incluidos SOD1, CAT, GPX1, GST, Keap1 y NRF2 en el músculo LD (36). Como era de esperar, nuestros datos mostraron que la inyección de AV mejoró la abundancia de NRF2 y CAT, lo que sugiere que AV activa el sistema antioxidante y protege al cuerpo del daño oxidativo. Teniendo en cuenta que la vida útil es una limitación económica importante para la industria de la carne de cordero (57), la mejora del potencial antioxidante de los corderos inyectados en AV puede prolongar la vida útil de la carne de cordero.
Conclusión
En resumen, la inyección intramuscular de AV en la etapa neonatal mejoró la calidad de la carne de los corderos, incluido el valor a* postmortem, el contenido de proteína bruta y grasa bruta, y mejoró la capacidad antioxidante en el músculo LD.
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán puestos a disposición por los autores, sin reservas indebidas.
Declaración ética
Los estudios en animales fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Agrícola de Shanxi (sxnd202028). Los estudios se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
PD: Conceptualización, Software, Validación, Visualización, Escritura – borrador original. GH: Curación de datos, Análisis formal, Redacción – borrador original. JF: Investigación, Escritura – borrador original. WZ: Metodología, Redacción – borrador original. XL: Metodología, Redacción – borrador original. JZ: Obtención de fondos, administración de proyectos, recursos, supervisión, redacción, revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue financiada por la Fundación Nacional de Ciencias Naturales de China (31972559) y el Proyecto de Cultivo de Jóvenes Académicos Distinguidos y Excelentes de la Universidad Agrícola de Shanxi (2022JQPYGC01).
Reconocimientos
Los autores agradecen el apoyo y la asistencia brindados por el personal de la Pastoral Taigu Paulson.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Molinero, RK . Factores que afectan a la calidad de la carne cruda. Procesamiento de carne: mejora de la calidad, Woodhead Publishing: Sawston, Reino Unido, (2002) 27–63
2. Purslow, PP . Nuevos aspectos de la calidad de la carne: de los genes a la ética Editorial Woodhead (2017). 744p pág.
3. Ponnampalam, EN, Holman, B, y Scollan, ND. Ovejas: Carne. Enciclopedia de alimentación y salud (2016), 750–757. Prensa Académica
4. Pannier, L, Pethick, DW, Boyce, M, Ball, A, Jacob, RH y Gardner, GE. Asociaciones de factores genéticos y no genéticos con concentraciones de hierro y zinc en el músculo longissimus de cordero. Ciencia de la carne. (2014) 96:1111–9. doi: 10.1016/j.meatsci.2013.08.013
5. Ponnampalam, EN, Butler, KL, McDonagh, MB, Jacobs, JL y Hopkins, DL. Relación entre el estado antioxidante muscular, las formas de hierro, los ácidos grasos poliinsaturados y la funcionalidad (colorante minorista) de la carne en corderos. Ciencia de la carne. (2012) 90:297–303. doi: 10.1016/j.meatsci.2011.07.014
6. Contini, C, Álvarez, R, O’sullivan, M, Dowling, DP, Gargan, SÓ, y Monahan, FJ. Efecto de un envase activo con extracto de cítricos sobre la oxidación lipídica y la calidad sensorial de la carne de pavo cocida. Ciencia de la carne. (2014) 96:1171–6. doi: 10.1016/j.meatsci.2013.11.007
7. Moczkowska, M, Półtorak, A, Montowska, M, Pospiech, E, y Wierzbicka, A. Efecto del sistema de envasado y el tiempo de almacenamiento en el proceso de degradación y oxidación de proteínas miofibrilares en relación con la terneza de la carne de vacuno. Ciencia de la carne. (2017) 130:7–15. doi: 10.1016/j.meatsci.2017.03.008
8. Zhao, J, Li, K, Su, R, Liu, W, Ren, Y, Zhang, C, et al. Efecto de la suplementación dietética con extracto de trigo sarraceno tártaro sobre el rendimiento del crecimiento, la calidad de la carne y la actividad antioxidante en corderas. Ciencia de la carne. (2017) 134:79–85. doi: 10.1016/j.meatsci.2017.07.016
9. Monaghan, BR, y Schmitt, FO. Los efectos del caroteno y de la vitamina A sobre la oxidación del ácido linoleico. J Biol Chem. (1932) 96:387–95. doi: 10.1016/S0021-9258(18)76278-9
10. Schwarz, EJ, Reginato, MJ, Shao, D, Cracovia, SL, y Lazar, MA. El ácido retinoico bloquea la adipogénesis mediante la inhibición de la transcripción mediada por C/EBPbeta. Mol Cell Biol. (1997) 17:1552–61. doi: 10.1128/MCB.17.3.1552
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Ryan, T, Liu, J, Chu, A, Wang, L, Blais, A, y Skerjanc, IS. El ácido retinoico mejora la miogénesis esquelética en las células madre embrionarias humanas al expandir la población de progenitores premiogénicos. Representante de Células Madre Rev. (2012) 8:482–93. doi: 10.1007/s12015-011-9284-0
12. Chidipi, B, Shah, SI, Reiser, M, Kanithi, M, Garcés, A, Cha, BJ, et al. El ácido retinoico totalmente trans aumenta los niveles de DRP1 y promueve la fisión mitocondrial. Células. (2021) 10:1202. doi: 10.3390/cells10051202
13. Wang, B, Nie, W, Fu, X, de Ávila, JM, Ma, Y, Zhu, MJ, et al. La inyección neonatal de vitamina A promueve el crecimiento muscular del ganado y aumenta las fibras musculares oxidativas. J Animal Sci Biotechnol. (2018) 9:1–10. doi: 10.1186/s40104-018-0296-3
14. Lara-Ramírez, R, Zieger, E y Schubert, M. Señalización del ácido retinoico en el desarrollo de la médula espinal. Int J Biochem Cell Biol. (2013) 45:1302–13. doi: 10.1016/j.biocel.2013.04.002
15. Tesoriere, L, Ciaccio, M, Bongiorno, A, Riccio, A, Pintaudi, AM, y Livrea, MA. Actividad antioxidante del todo-trans-retinol en solución homogénea y en liposomas de fosfatidilcolina. Arch Biochem Biophys. (1993) 307:217–23. doi: 10.1006/abbi.1993.1581
16. Song, P, Chen, X, Zhao, J, Li, Q, Li, X, Wang, Y, et al. La inyección de vitamina A al nacer mejora el crecimiento muscular en corderos. Animal Nutr. (2023) 14:204–12. doi: 10.1016/j.aninu.2023.05.011
17. Bryant, T, Wagner, J, Tatum, J, Galyean, M, Anthony, R y Engle, T. Efecto de la concentración de vitamina A suplementaria dietética sobre el rendimiento, el mérito de la canal, los metabolitos séricos y la actividad enzimática lipogénica en novillos de carne de un año. J Anim Sci. (2010) 88:1463–78. doi: 10.2527/jas.2009-2313
18. Consejo Nacional de Investigación . Requerimientos de nutrientes de pequeños rumiantes: ovejas, cabras, cérvidos y camélidos del Nuevo Mundo. Washington, D.C., EE.UU.: The National Academies Press (2007).
19. Harris, CL, Wang, B, Deavila, JM, Busboom, JR, Maquivar, M, Parish, SM, et al. La administración de vitamina A al nacer promueve el crecimiento de los terneros y el desarrollo de grasa intramuscular en el ganado vacuno Angus. J Animal Sci Biotechnol. (2018) 9:55. doi: 10.1186/s40104-018-0268-7
20. Asociación de Químicos Analíticos Oficiales (AOAC) . Internacional a: Métodos oficiales de análisis de la AOAC internacional. La Asociación del Condado de Arlington, VA, EE.UU.: (2000).
21. Van Soest, PV, Robertson, JB y Lewis, BA. Métodos para la fibra dietética, la fibra detergente neutra y los polisacáridos sin almidón en relación con la nutrición animal. J Lácteos Sci. (1991) 74:3583–97. doi: 10.3168/jds. S0022-0302(91)78551-2
22. Li, X, Liu, X, Song, P, Zhao, J, Zhang, J y Zhao, J. Masa muscular esquelética, calidad de la carne y estado antioxidante en corderos en crecimiento suplementados con ácido guanidinoacético. Ciencia de la carne. (2022) 192:108906. doi: 10.1016/j.meatsci.2022.108906
23. Kim, YK y Quadro, L. Análisis de cromatografía líquida de alta resolución (HPLC) de fase inversa de retinol y ésteres de retinilo en suero y tejidos de ratón. Retinoides. (2010):263–75. doi: 10.1007/978-1-60327-325-1_15
24. Asociación de Químicos Analíticos Oficiales (AOAC) . Métodos oficiales de análisis 18ª Asociación de Químicos Analíticos Oficiales, Gaithersburg, MD, EE.UU. (2005).
25. Ahlemeyer, B y Krieglstein, J. La inhibición del agotamiento del glutatión por el ácido retinoico y el tocoferol protege a las neuronas cultivadas del estrés oxidativo y la apoptosis inducidos por estaurosporina. Neurochem Int. (2000) 36:1–5. doi: 10.1016/S0197-0186(99)00101-1
26. Ahlemeyer, B y Krieglstein, J. El ácido retinoico reduce el daño apoptótico inducido por estaurosporina en neuronas embrionarias de pollo mediante la supresión de la producción de especies reactivas de oxígeno. Neurosci Lett. (1998) 246:93–6. doi: 10.1016/S0304-3940(98)00242-0
27. Jackson, GR, Morgan, BC, Werrbach-Pérez, K y Pérez-Polo, JR. Efecto antioxidante del ácido retinoico en el feocromocitoma de rata PC12. Int J Dev Neurosci. (1991) 9:161–3. doi: 10.1016/0736-5748(91)90007-9
28. Moreno-Manzano, V, Ishikawa, Y, Lucio-Cazana, J y Kitamura, M. Supresión de la apoptosis por ácido trans-retinoico: intervención dual en la vía c-Jun N-terminal quinasa-AP-1. J Biol Chem. (1999) 274:20251–8. doi: 10.1074/jbc.274.29.20251
29. Munekata, PE, Pateiro, M, López-Pedrouso, M, Gagaoua, M, y Lorenzo, JM. Foodomics en la calidad de la carne. Curr Opin Food Sci. (2021) 38:79–85. doi: 10.1016/j.cofs.2020.10.003
30. Devine, CE, Graafhuis, AE, Muir, PD, y Chrystall, BB. El efecto de la tasa de crecimiento y el pH final en la calidad de la carne de los corderos. Ciencia de la carne. (1993) 35:63–77. doi: 10.1016/0309-1740(93)90070-X
31. Watanabe, A, Daly, C, y Devine, CE. Los efectos del pH final de la carne sobre la terneza cambian durante el envejecimiento. Ciencia de la carne. (1996) 42:67–78. doi: 10.1016/0309-1740(95)00012-7
32. Zhao, J, Li, Q, Zhang, R, Liu, W, Ren, Y, Zhang, C, et al. Efecto del orujo de uva dietético sobre el rendimiento del crecimiento, la calidad de la carne y la actividad antioxidante en corderos de carnero. Anim Feed Sci Technol. (2018) 236:76–85. doi: 10.1016/j.anifeedsci.2017.12.004
33. O’Grady, M, Carpenter, R, Lynch, P, O’brien, N, y Kerry, JP. Adición de extracto de semilla de uva y gayuba a las dietas porcinas: influencia en los atributos de calidad de la carne de cerdo cruda y cocida. Ciencia de la carne. (2008) 78:438–46. doi: 10.1016/j.meatsci.2007.07.011
34. Suman, SP y Joseph, P. Química de la mioglobina y color de la carne. Annu Rev Food Sci Technol. (2013) 4:79–99. doi: 10.1146/annurev-food-030212-182623
35. Ponnampalam, EN, Warner, R, Kitessa, S, McDonagh, M, Pethick, D, Allen, D, et al. Influencia de los sistemas de engorde y del lugar de muestreo en la composición de ácidos grasos y la vida útil minorista del cordero. Anim Prod Sci. (2010) 50:775–81. doi: 10.1071/AN10025
36. Xu, M, Chen, X, Huang, Z, Chen, D, Li, M, He, J, et al. Efectos de la suplementación dietética con extracto de proantocianidina de semilla de uva sobre la calidad de la carne, las características de la fibra muscular y la capacidad antioxidante de los cerdos de engorde. Químico de los alimentos. (2022) 367:130781. doi: 10.1016/j.foodchem.2021.130781
37. Zhang, C, Luo, J, Yu, B, Zheng, P, Huang, Z, Mao, X, et al. La suplementación con resveratrol en la dieta mejora la calidad de la carne de los cerdos de engorde a través del cambio de las características de la fibra muscular y el estado antioxidante. Ciencia de la carne. (2015) 102:15–21. doi: 10.1016/j.meatsci.2014.11.014
38. Kim, GD, Jeong, JY, Hur, SJ, Yang, HS, Jeon, JT y Joo, ST. La relación entre el color de la carne (CIE L* y a*), el contenido de mioglobina y su influencia en las características de la fibra muscular y la calidad de la carne de cerdo. Fuente de alimentos Sci Animal. (2010) 30:626–33. doi: 10.5851/kosfa.2010.30.4.626
39. Boles, JA, y Pegg, R. Color de la carne. Universidad Estatal de Montana y Programa de Innovación de Productos Alimenticios de Saskatchewan, Universidad de Saskatchewan (2010).
40. Brooke, MH, y Kaiser, KK. Tipos de fibras musculares: ¿cuántas y de qué tipo? Arch Neurol. (1970) 23:369–79. doi: 10.1001/archneur.1970.00480280083010
41. Lefaucheur, L, Hoffman, R, Gerrard, D, Okamura, C, Rubinstein, N y Kelly, A. Evidencia de tres isoformas adultas de cadena pesada de miosina rápida en fibras musculares esqueléticas tipo II en cerdos. J Anim Sci. (1998) 76:1584–93. doi: 10.2527/1998.7661584x
42. Shiranita, K, Hayashi, K, Otsubo, A, Miyajima, T y Takiyama, R. Clasificación de la calidad de la carne mediante el procesamiento de imágenes. Reconocimiento de patrones. (2000) 33:97–104. doi: 10.1016/S0031-3203(99)00035-7
43. Wood, JD, Enser, M, Fisher, AV, Nute, GR, Sheard, PR, Richardson, RI, et al. Deposición de grasa, composición de ácidos grasos y calidad de la carne: una revisión. Ciencia de la carne. (2008) 78:343–58. doi: 10.1016/j.meatsci.2007.07.019
44. Hopkins, D, y Mortimer, S. Efecto del genotipo, el sexo y la edad en la calidad de la carne ovina y un estudio de caso que ilustra la integración del conocimiento. Ciencia de la carne. (2014) 98:544–55. doi: 10.1016/j.meatsci.2014.05.012
45. Veiseth, E, Shackelford, S, Wheeler, T y Koohmaraie, M. Los factores que regulan la terneza del cordero longissimus se ven afectados por la edad en el momento del sacrificio. Ciencia de la carne. (2004) 68:635–40. doi: 10.1016/j.meatsci.2004.05.015
46. Van Laack, R, Stevens, S, y Stalder, KJ. La influencia del pH final y el contenido de grasa intramuscular en la terneza y el ablandamiento de la carne de cerdo. J Anim Sci. (2001) 79:392–7. doi: 10.2527/2001.792392x
47. Addis, M . Principales causas del deterioro de la carne y técnicas de conservación: a. Rev Food Sci Qual Manag. (2015) 41:101–14.
48. Wakabayashi, N, Itoh, K, Wakabayashi, J, Motohashi, H, Noda, S, Takahashi, S, et al. La mutación Keap1-nula conduce a la letalidad postnatal debido a la activación constitutiva de Nrf2. Nat Genet. (2003) 35:238–45. doi: 10.1038/ng1248
49. Ponnampalam, EN, Norng, S, Burnett, VF, Dunshea, FR, Jacobs, JL, y Hopkins, DL. Sinergismo de los componentes bioquímicos que controlan la oxidación lipídica en el músculo de cordero. Lípidos. (2014) 49:757–66. doi: 10.1007/s11745-014-3916-5
50. Salami, S, Guinguina, A, Agboola, J, Omede, A, Agbonlahor, E, y Tayyab, U. Efectos in vivo y post mortem de los antioxidantes para piensos en ganado: una revisión de las implicaciones en la autorización de aditivos antioxidantes para piensos. Animal. (2016) 10:1375–90. doi: 10.1017/S1751731115002967
51. Jiang, Z, Jiang, S, Lin, Y, Xi, P, Yu, D y Wu, T. Efectos de la isoflavona de soja en el rendimiento del crecimiento, la calidad de la carne y la antioxidación en pollos de engorde machos. Poult Sci. (2007) 86:1356–62. doi: 10.1093/ps/86.7.1356
52. Chelikani, P, Fita, I, y Loewen, P. Diversidad de estructuras y propiedades entre catalasas. Cell Mol Life Sci. (2004) 61:192–208. doi: 10.1007/s00018-003-3206-5
53. Hengge, AJ . Re: ¿cómo se usa la catalasa en la industria? MadSci Network: Biología General (1999).
54. Selvaratnam, J y Robaire, B. La sobreexpresión de catalasa en ratones reduce el estrés oxidativo relacionado con la edad y mantiene la producción de espermatozoides. Exp Gerontol. (2016) 84:12–20. doi: 10.1016/j.exger.2016.08.012
55. Del Río, D, Stewart, AJ, y Pellegrini, NJ. Una revisión de estudios recientes sobre el malondialdehído como molécula tóxica y marcador biológico de estrés oxidativo. Nutr Metab Cardiovasc Dis. (2005) 15:316–28. doi: 10.1016/j.numecd.2005.05.003
56. Keum, YS, y Choi, BY. Regulación molecular y química de la vía de señalización Keap1-Nrf2. Moléculas. (2014) 19:10074–89. doi: 10.3390/moléculas190710074
Palabras clave: vitamina A, ácido retinoico, actividad antioxidante, calidad de la carne, longissimus dorsi, cordero
Cita: Song P, Huo G, Feng J, Zhang W, Li X y Zhao J (2023) La inyección intramuscular de vitamina A en corderos recién nacidos aumenta la capacidad antioxidante y mejora la calidad de la carne. Frente. Vet. Sci. 10:1272874. doi: 10.3389/fvets.2023.1272874
Editado por:
RuiHu, Universidad Agrícola de Sichuan, China
Revisado por:
Tiande Zou, Universidad Agrícola de Jiangxi, China Yuanqing Xu, Universidad Agrícola de Mongolia Interior, China
Derechos de autor © 2023 Song, Huo, Feng, Zhang, Li y Zhao. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Junxing Zhao, Junxzh@sxau.edu.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía