
El eritritol altera la expresión génica de la fosfotransferasa e inhibe el crecimiento in vitro de Staphylococcus coagulans

El eritritol altera la expresión génica de la fosfotransferasa e inhibe el crecimiento in vitro de Staphylococcus coagulans aislados de caninos con pioderma
 Saki Onishi-Sakamoto1†
Saki Onishi-Sakamoto1†  Tadashi Fujii2,3 Keito Watanabe4 Reina Makida5
Tadashi Fujii2,3 Keito Watanabe4 Reina Makida5  Keita Iyori6† Yoichi Toyoda6 Takumi Tochio2,3†
Keita Iyori6† Yoichi Toyoda6 Takumi Tochio2,3†  Koji Nishifuji7*†
Koji Nishifuji7*†- 1División Cooperativa de Ciencias Veterinarias, Escuela de Posgrado de Agricultura, Universidad de Agricultura y Tecnología de Tokio, Fuchu, Tokio, Japón
- 2Departamento de Gastroenterología y Hepatología, Fujita Health University, Toyoake, Japón
- 3Departamento de Investigación Médica sobre Prebióticos y Probióticos, Universidad de Salud de Fujita, Toyoake, Japón
- 4Departamento Cooperativo de Medicina Veterinaria, Facultad de Agricultura, Universidad de Agricultura y Tecnología de Tokio, Fuchu, Tokio, Japón
- 5Centro de Investigación y Desarrollo, B Food Science Co., Ltd., Chita, Japón
- 6Servicio Dermatológico y de Laboratorio para Animales, Vet Derm Tokyo, Fujisawa, Japón
- 7División de Ciencias de la Vida Animal, Instituto de Agricultura, Universidad de Agricultura y Tecnología de Tokio, Fuchu, Tokio, Japón
Staphylococcus coagulans (SC) pertenece a un grupo de estafilococos coagulasa positivos ocasionalmente aislados de las lesiones cutáneas de perros con pioderma. Recientemente revelamos que el eritritol, un alcohol de azúcar, inhibe el crecimiento de la cepa SC JCM7470. En este estudio se investigaron los mecanismos moleculares implicados en esta inhibición del crecimiento de JCM7470 por eritritol, y se determinó si el eritritol inhibe el crecimiento de SC aislado de la piel de perros con pioderma. El análisis exhaustivo de la expresión génica de JCM7470 en presencia de eritritol reveló que el eritritol reguló al alza la expresión de los genes glcB y ptsG, los cuales codifican los dominios C, B y A de permeasa específica de glucosa del sistema fosfotransferasa (PTS) y permeasa específica de glucosa (EIICBA), respectivamente, asociados con la absorción de azúcar. Además, el eritritol suprimió el crecimiento in vitro de las 27 cepas SC aisladas de las lesiones cutáneas de pioderma canino, incluidas 13 cepas del gen mecA positivo y 14 cepas del gen mecA negativas. Finalmente, se restableció la inhibición del crecimiento de los aislados clínicos SC por eritritol mediante la adición de glucosa. En resumen, revelamos que el eritritol promueve la expresión génica del PTS y suprime el crecimiento in vitro de aislados clínicos de SC de perros con pioderma. La restauración de la inhibición del crecimiento inducida por el eritritol por la glucosa sugirió que la inanición de la glucosa puede contribuir a la inhibición del crecimiento del SC.
Introducción
Staphylococcus coagulans (SC), anteriormente clasificado como S. schleiferi subsp. coagulans, es un estafilococo coagulasa positivo aislado principalmente de la piel y los canales auditivos de los perros (1-6). Entre los estafilococos, el SC es la segunda causa más prevalente de lesión cutánea en perros con pioderma después de Staphylococcus pseudintermedius (SP) (7). Además de las infecciones en perros, se ha reportado que el SC causa infecciones oportunistas en humanos (8, 9). Al igual que en otras especies de estafilococos (10-13), se ha informado de la aparición de SC resistentes a la meticilina y a múltiples fármacos (14-17), lo que representa un problema para la medicina humana y veterinaria. En este contexto, se prevé el desarrollo de nuevas estrategias para la prevención del pioderma canino utilizando sustratos bacteriostáticos.
El eritritol (1,2,3,4-butanetetrol) es un poliol que se utiliza ampliamente en los alimentos como edulcorante artificial (18). Se demostró que tanto el eritritol como el xilitol, otro tipo de poliol, inhiben el crecimiento de Streptococcus mutans, Streptococcus gordonii y Porphyromonas gingivalis, las principales bacterias comensales orales humanas (18-20). Recientemente, los estudios también han informado que el eritritol inhibió el crecimiento de las bacterias comensales de la piel humana Corynebacterium minutissimum, Corynebacterium striatum, Staphylococcus epidermidis y Cutibacterium acnes, así como las principales bacterias comensales orales caninas Porphyromonas gulae y Porphyromonas macacae (21-23).
Los mecanismos por los cuales el xilitol inhibe S. mutans han sido bien estudiados (24). S. mutans incorpora xilitol a través del sistema fosfotransferasa dependiente de fosfoenolpiruvato (PEP-PTS) y lo fosforila a xilitol 5-fosfato. El xilitol 5-fosfato acumulado en las bacterias inhibe directamente la actividad enzimática relacionada con la glucólisis y compite con la proteína estable al fósforo (24) para inhibir indirectamente la absorción de azúcar. Tales mecanismos pueden resultar en la inanición de glucosa en S. mutans y la inhibición del crecimiento bacteriano.
Muy recientemente, revelamos que el eritritol suprimió el crecimiento in vitro de las cepas SP y SC JCM17571 y JCM7470, respectivamente (25). Además, el eritritol reguló al alza los genes relacionados con PEP-PTS (ptsG, ppdK y ppdkR) en SP JCM17571 (26). Sin embargo, no se ha dilucidado el mecanismo molecular exacto por el cual el eritritol suprime el crecimiento de SC. En este estudio, nuestro objetivo fue identificar los grupos de genes SC cuya expresión fue alterada por el eritritol. Además, investigamos si el eritritol suprime el crecimiento de aislados clínicos de SC in vitro.
Materiales y métodos
Cepas bacterianas
La cepa SC JCM7470 (idéntica a ATCC 49545) fue proporcionada por la Colección de Microorganismos de Japón (JCM) y utilizada como cepa de referencia (2). Un estudio previo reveló que esta cepa era susceptible a la oxacilina y a la cefoxitina mediante pruebas de difusión en disco (27). Un total de 27 aislados cutáneos SC de 27 perros con pioderma en diferentes consultorios privados y enviados a Vet Derm Tokyo Co., Ltd., para pruebas de susceptibilidad a los antibióticos también se utilizaron como aislamientos clínicos. La identificación de SC se confirmó de la siguiente manera: El ADN extraído de las 27 cepas bacterianas se sometió a PCR multiplex para la identificación de cepas estafilocócicas coagulasas positivas (28). Si el tamaño de la banda de los amplicones era idéntico al de S. schleiferi, las cepas se sometieron a una prueba de coagulasa utilizando plasma de conejo (Eiken Chemical Co. Ltd., Tokio, Japón) para identificar S. schleiferi a nivel de subespecie (28). El gen mecA en los aislados clínicos SC se identificó mediante PCR con pares de cebadores utilizados para identificar este gen en S. aureus, SP y S. schleiferi aislados de perros (29). La prueba de sensibilidad a los antimicrobianos se realizó mediante una prueba de difusión en disco utilizando KB Disk™ (Eiken Chemical Co. Ltd.), como se describió anteriormente (30). Para la prueba de susceptibilidad se utilizaron los siguientes antimicrobianos: amoxicilina-clavulanato (AMPC/ACV; 20 ó 10 μg/disco), cefalexina (CEX; 30 μg/disco), cefpodoxima (CPDX; 10 μg/disco), enrofloxacino (ERFX; 5 μg/disco), gentamicina (GM; 10 μg/disco), sulfametoxazol-trimetoprima (ST; 23,75-1,25 μg/disco), clindamicina (CLDM; 2 μg/disco), doxiciclina (DOXY; 30 μg/disco), minociclina (MINO; 30 μg/disco), cloranfenicol (PC; 30 μg/disco) y fosfomicina (FOM; 50 μg/disco). En la Tabla Suplementaria 1 se enumeran los cebadores de PCR utilizados en este estudio, y en la Tabla Suplementaria 2 se muestran los resultados de las pruebas de susceptibilidad a la difusión en disco.
Cultivo bacteriano
Los siguientes experimentos se realizaron de acuerdo con la metodología utilizada en un estudio previo (26). Se inoculó una sola colonia de JCM7470 en 3 ml de medio #802 del Centro de Recursos Biológicos del Instituto Nacional de Tecnología y Evaluación (NRBC) que contenía 1% de hipoliptón (Fujifilm Wako, Osaka, Japón), 0,2% de extracto de levadura (Nacalai Tesque Inc., Kyoto, Japón) y 0,1% de MgSO4·7H2O (Fujifilm Wako, pH 7.0), e incubado con rotación a 210 rpm hasta la densidad óptica a 600 nm (OD600) alcanzó el 3,4. Las suspensiones bacterianas se diluyeron 100 veces en medio NRBC #802 con o sin eritritol al 5% (p/p) (B Food Sciences Co. Ltd., Tokio, Japón) y se incubaron a 30 °C hasta OD600 alcanzó entre 0,8 y 1,0. Estos experimentos se realizaron por triplicado. Elegimos una concentración de eritritol del 5% en este estudio, ya que habíamos observado que el eritritol en concentraciones más altas inhibía significativamente el crecimiento de JCM7470 en un estudio anterior (25).
Secuenciación de ARN (RNA-seq)
Se utilizó RNeasy Mini Kit (Qiagen, Venlo, Países Bajos) para extraer el ARN total de las muestras bacterianas. El total de muestras de ARN se envió al Laboratorio de Bioingeniería (Sagamihara, Japón). Después de la eliminación del ARN ribosómico utilizando riboPOOLS (siTOOLs Biotech, Planegg, Alemania), se generó una biblioteca de ADNc para el análisis de secuenciación de ARN utilizando MGIEasy RNA Directional Library Prep Set (MGI Tech, Shenzhen, China). La biblioteca de ADNc se utilizó para construir una biblioteca de ADN circular utilizando el kit de circularización MGIEasy (MGI Tech). La biblioteca de ADNc anclada por DNA Nanoball (DNA) se sometió a un análisis de secuenciación utilizando DNBSEC-G400 (MGI Tech). Recurso de Análisis de Secuencias de Ácidos Nucleicos (NASQAR; https://nasqar.abudhabi.nyu.edu) se utilizó para crear gráficos de análisis de componentes principales (PCA) de las muestras por triplicado y un mapa de calor para visualizar los resultados de RNA-seq. Se creó un gráfico de volcán utilizando ggVolcanoR (https://ggvolcanor.erc.monash.edu). Proteína ANNotación con Z-scoRE (PANNZER2; http://ekhidna2.biocenter.helsinki.fi/sanspanz) se utilizó para el análisis de ontología génica (GO) para predecir los genes regulados al alza y a la baja en respuesta al eritritol.
Reacción en cadena de la polimerasa cuantitativa con transcripción inversa (RT-qPCR)
Se utilizó el kit de síntesis de ADNc de primera cadena de transcriptores (Roche Diagnostics, Rotkreuz, Suiza) con cebadores aleatorios para sintetizar ADN complementario a partir de ARN total extraído de JCM7470 con o sin eritritol. En la Tabla Suplementaria 1 se enumeran los cebadores utilizados en este estudio. Los juegos de cebadores y TB Green Fast qPCR Mix (Takara Bio, Kusatsu, Japón) se utilizaron para RT-qPCR en un termociclador Dice Real-Time System III (Takara Bio) con 45 ciclos de 95 °C durante 5 s y 55 °C durante 60 s. El gen recA, que ha sido validado como un gen de referencia apropiado para la qPCR en SP (31), se utilizó como referencia para evaluar los niveles relativos de expresión génica de los otros genes. El RNA-seq realizado en este estudio reveló que la expresión del gen recA en SC fue consistente independientemente de la presencia de eritritol (log®®2FC = −0,19, p = 0,111), lo que sugiere la validez del gen recA como gen de referencia para la qPCR en SC. Este experimento se realizó por triplicado y se compararon los valores medios entre los grupos.
Ensayo de turbidez in vitro de aislados clínicos SC
Los aislados clínicos de SC se precultivaron en medio Luria-Bertani (LB) (Kanto Chemical Co., Inc., Tokio, Japón) y se diluyeron cinco veces en este medio. Luego, se mezclaron 30 μL de la suspensión bacteriana diluida con 900 μL de medio NRBC #802 con eritritol en concentraciones de 0%, 5%, 10% y 15% [p/p] en microplacas de fondo en U de 96 pocillos (Watson Corporation, Tokio, Japón). El OD600 se midió a lo largo del tiempo utilizando EpochTM2 (Agilent Technologies, Inc., Santa Clara, CA, EE. UU.) de 0 a 6 h.
El efecto de la glucosa sobre la inhibición del crecimiento inducida por eritritol de aislados clínicos de SC se analizó de la siguiente manera. Los aislados de SC se incubaron en medio NRBC #802 durante 2 h y se diluyeron cinco veces en medio NRBC #802. Luego, se mezclaron 30 μL de la suspensión bacteriana diluida con 900 μL de medio NRBC #802 que contenía 0% de eritritol y 0% de glucosa, 0% de eritritol y 0,1% de glucosa, 10% de eritritol y 0,1% de glucosa, 10% de eritritol y 1% de glucosa, o 10% de eritritol y 0% de glucosa, y se inocularon 200 μL en cada placa de 96 pocillos. El OD600 se midió a lo largo del tiempo durante un máximo de 6 h. Los experimentos de análisis de crecimiento se realizaron por triplicado y se compararon los valores medios entre los grupos.
Análisis estadístico
Se utilizó el análisis empírico de datos digitales de expresión génica en R (edgeR) exactTest para comparar los niveles de expresión génica analizados por RNA-seq. Para el siguiente análisis estadístico se utilizó el software GraphPad Prism 9 (GraphPad Software Inc., San Diego, CA, EE. UU.). La prueba t de Welch se utilizó para comparar los niveles de transcripción de los genes glcB y ptsG entre los grupos, y el efecto del eritritol en las cepas SC positivas y negativas para el gen mecA. La prueba de Dunnett se utilizó para comparar la turbidez entre cepas SC incubadas en presencia o ausencia de eritritol y/o glucosa. Un valor de p inferior a 0,05 se consideró estadísticamente significativo.
Resultados
Análisis exhaustivo de la expresión génica de la JCM7470 SC en respuesta al eritritol
Primero realizamos un análisis de RNA-seq para investigar el mecanismo molecular detrás de la inhibición del crecimiento de JCM7470 por eritritol. Después de filtrar las lecturas de secuenciación en bruto, obtuvimos 16.589.804, 16.962.572 y 27.123.064 lecturas limpias del transcriptoma en muestras de control. Por el contrario, hubo 17.294.425, 18.110.786 y 15.713.381 lecturas limpias en las muestras tratadas con eritritol.
Los niveles de expresión génica calculados [|log2 veces cambian (log2FC)| > 1, p < 0,05] identificó un total de 162 genes expresados diferencialmente, incluidos 60 genes regulados al alza y 102 genes regulados a la baja, en JCM7470 después del tratamiento con eritritol. El gráfico de PCA con el 80% de la varianza explicada por PC1 mostró una clara división entre las muestras control y las tratadas con eritritol (Figura 1A). El mapa de calor se muestra en la Figura complementaria 1.
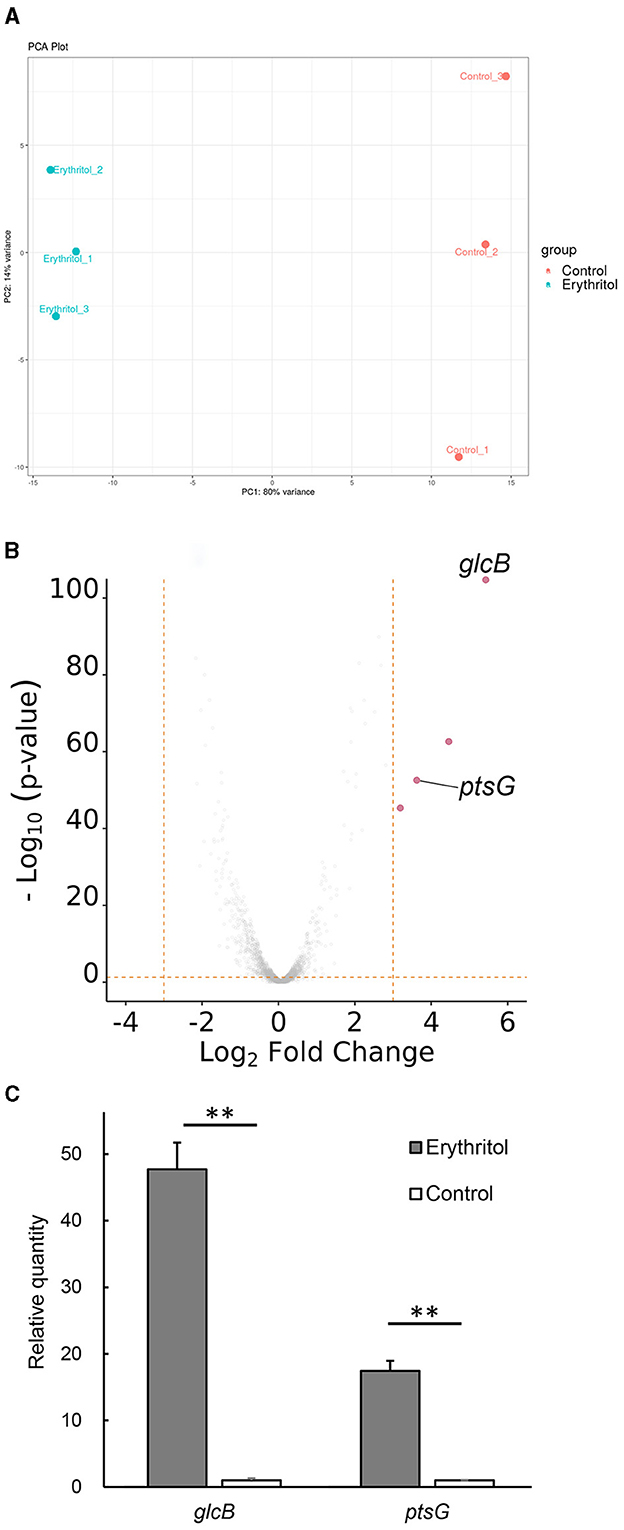 Figura 1. Análisis de expresión génica. (A) El gráfico de PCA muestra una clara división entre el control y el Staphylococcus coagulans tratado con eritritol JCM7470, con un 80% de la varianza explicada por el PC1. (B) El diagrama del volcán muestra diferencias en los niveles de expresión de genes regulados al alza y a la baja en JCM7470 tratados con eritritol. Los genes se regulan al alza >10 veces (log2FC > 3,322, p < 0,05) en respuesta al eritritol se resaltan en rojo. (C) Comparación de los niveles de expresión de los genes glcB y ptsG en JCM7470 tratados con eritritol o medio control, según lo determinado por RT-qPCR. p < 0,01.**
Figura 1. Análisis de expresión génica. (A) El gráfico de PCA muestra una clara división entre el control y el Staphylococcus coagulans tratado con eritritol JCM7470, con un 80% de la varianza explicada por el PC1. (B) El diagrama del volcán muestra diferencias en los niveles de expresión de genes regulados al alza y a la baja en JCM7470 tratados con eritritol. Los genes se regulan al alza >10 veces (log2FC > 3,322, p < 0,05) en respuesta al eritritol se resaltan en rojo. (C) Comparación de los niveles de expresión de los genes glcB y ptsG en JCM7470 tratados con eritritol o medio control, según lo determinado por RT-qPCR. p < 0,01.**
El eritritol reguló al alza la expresión de los genes del sistema fosfotransferasa específicos de la glucosa en JCM7470
De los 162 genes en JCM7470 expresados diferencialmente en respuesta al eritritol, solo cuatro estaban regulados al alza más de 10 veces (log2FC > 3.322) en presencia de eritritol. Por el contrario, ningún gen se reguló a la baja más de 10 veces en presencia de eritritol.
La Tabla Suplementaria 3 muestra las descripciones funcionales predichas y los procesos biológicos de GO para los 15 genes principales regulados al alza y a la baja en respuesta al eritritol. El análisis de GO reveló que dos de los cuatro genes más regulados al alza eran glcB [log2FC = 5,459, p < 0,001, valor predictivo positivo (VPP) = 0,71] y ptsG (log2FC = 3,727, p < 0,001, PPV = 0,71), los cuales codifican la subunidad transportadora PTS IIBC (Figura 1B). Mientras tanto, los otros dos genes más regulados al alza codifican proteínas hipotéticas para las que se desconocen los procesos biológicos asociados (Figura 1B). La RT-qPCR reveló que los niveles de transcripción de los genes glcB y ptsG en el grupo tratado con eritritol fueron significativamente más altos que los del grupo control (glcB, p = 0,0024; ptsG, p = 0,0028, Figura 1C). El registro2 Los cambios en los pliegues de glcB y ptsG en el grupo tratado con eritritol en relación con los niveles en el grupo control fueron de 5,57 ± 0,06 y 4,12 ± 0,06 (media ± EE), respectivamente.
La suplementación con glucosa restauró la inhibición del crecimiento inducida por eritritol de aislados clínicos de SC
A continuación, investigamos si el eritritol suprime el crecimiento de SC aislado de lesiones clínicas de pioderma canino. El ensayo de turbidez in vitro reveló que la turbidez aumentó con el tiempo hasta 6 h tanto en el grupo de control como en los grupos suplementados con eritritol, independientemente de si las cepas portaban el gen mecA (Figura suplementaria 2). La turbidez de 27 aislamientos clínicos de SC incubados durante 6 h en presencia de 5% (0,728 ± 0,172, p < 0,0001), 10% (0,460 ± 0,130, p < 0,0001) y 15% de eritritol (0,283 ± 0,070, p < 0,0001) fue significativamente menor que el nivel en la incubación en ausencia de eritritol (1,023 ± 0,193). Además, el eritritol suprimió el crecimiento de los aislados clínicos de SC de forma dependiente de la concentración (p < 0,0001). Por el contrario, no hubo diferencias significativas en la turbidez bacteriana basal en los grupos SC entre los grupos suplementados con eritritol y el grupo control (p > 0,05) (Figura 2A). Además, no hubo diferencias significativas en la turbidez entre las cepas con gen mecA positivo (n = 13) y las cepas con gen mecA negativo (n = 14) suplementadas con 0% (p = 0,295), 5% (p = 0,332), 10% (p = 0,703) y 15% de eritritol (p = 0,709) (Figura 2B).
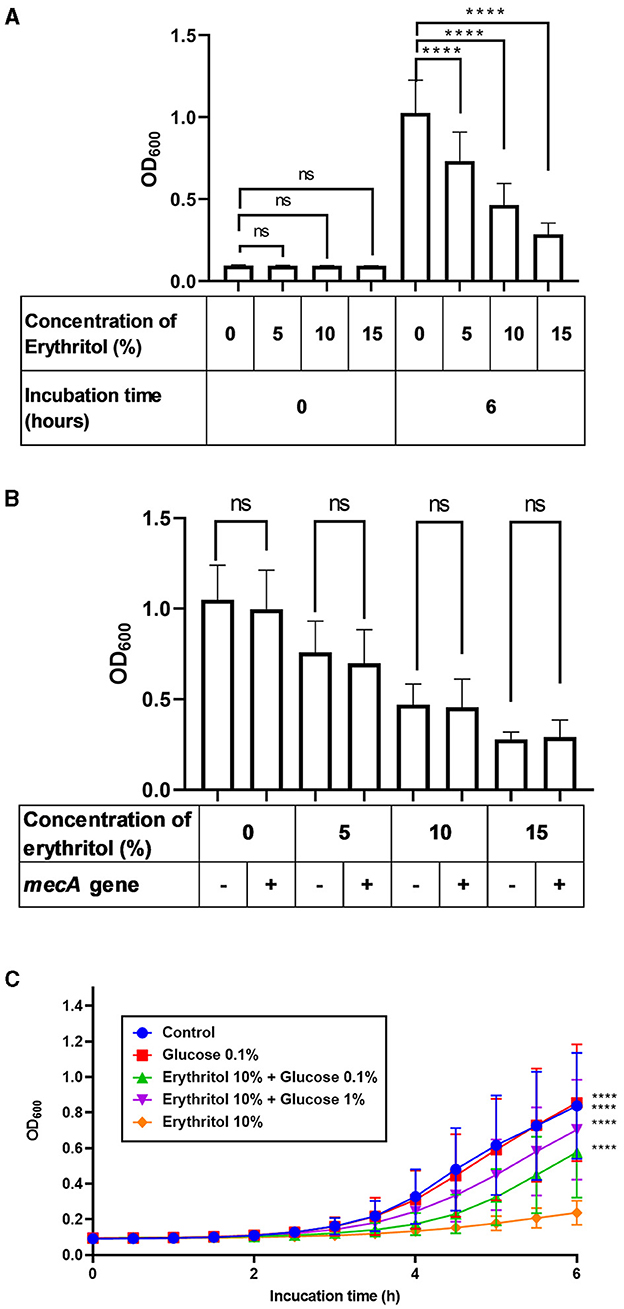 Figura 2. Restauración de la inhibición del crecimiento inducida por eritritol de aislados clínicos de SC mediante suplementación con glucosa. (A) Turbidez bacteriana de los aislados clínicos SC (n = 27) inmediatamente o 6 h después de la incubación con diferentes dosis de eritritol. (B) Comparación de la turbidez bacteriana entre cepas SC positivas y negativas para el gen mecA 6 h después de la incubación con diferentes dosis de eritritol. (C) Fluctuación de la turbidez bacteriana de las cepas SC incubadas con diferentes dosis de eritritol y glucosa. ns, no significativo; p < 0,0001.
Figura 2. Restauración de la inhibición del crecimiento inducida por eritritol de aislados clínicos de SC mediante suplementación con glucosa. (A) Turbidez bacteriana de los aislados clínicos SC (n = 27) inmediatamente o 6 h después de la incubación con diferentes dosis de eritritol. (B) Comparación de la turbidez bacteriana entre cepas SC positivas y negativas para el gen mecA 6 h después de la incubación con diferentes dosis de eritritol. (C) Fluctuación de la turbidez bacteriana de las cepas SC incubadas con diferentes dosis de eritritol y glucosa. ns, no significativo; p < 0,0001.
Teniendo en cuenta la regulación positiva de los genes relacionados con el SPT y la inhibición del crecimiento, nos preguntamos si la inanición de carbohidratos en respuesta al eritritol causaba la inhibición del crecimiento de los aislados clínicos SC. Por lo tanto, investigamos si la suplementación con glucosa de los medios de cultivo de los aislados clínicos SC superaría la inhibición del crecimiento inducida por el eritritol. Encontramos que la turbidez de los aislados clínicos de SC suplementados con eritritol al 10% y al 0,1% (0,577 ± 0,250, p < 0,0001) o al 1% de glucosa (0,704 ± 0,275, p < 0,0001) fue significativamente mayor que la de la suplementación con eritritol al 10% solo (Figura 2C).
Discusión
El eritritol puede inducir la inanición de glucosa en el SC
El presente estudio reveló que el eritritol aumentó significativamente la expresión de glcB y ptsG que codifican la subunidad transportadora IIBC de PTS en la cepa SC. El análisis de GO predijo que las transcripciones de estos dos genes funcionan como componentes específicos de la enzima II de glucósidos y glucosa EIICBA, respectivamente. La regulación positiva de la expresión génica ptsG en respuesta al eritritol en SC estuvo de acuerdo con los datos obtenidos en nuestro reciente estudio utilizando una cepa SP (26). Los EIICBA son permeados de membrana que desempeñan un papel importante en la absorción de carbohidratos en el citoplasma bacteriano (32, 33). EIIA, EIIB y EIIC generalmente incorporan específicamente un sustrato o carbohidratos estrechamente relacionados en el citoplasma bacteriano (34).
Además, la restauración de la supresión del crecimiento inducida por el eritritol mediante la suplementación con glucosa implica que la inanición de glucosa en respuesta al eritritol puede causar la regulación positiva de la expresión génica relacionada con el SPT y dar lugar a la inhibición del crecimiento del SC. Se informó que, en S. mutans, el xilitol 5-fosfato, un metabolito derivado del xilitol, inhibe directamente las enzimas glucolíticas y compite con la glucosa 6-fosfato, un metabolito de la glucosa incorporado a la glucólisis (24). Especulamos que el eritritol o sus metabolitos también compiten por la enzima glucolítica en SC, aunque aún no se han determinado la absorción exacta de eritritol y las vías metabólicas en los estafilococos.
Estudios anteriores revelaron que el operón ptsG de S. carnosus consta de dos genes adyacentes, glcA y glcB, que codifican IICBAGlc1 y IICBAGlc2, respectivamente (32, 33, 35). El análisis de la secuencia de aminoácidos deducida sugirió que el gen ptsG en SC (NCBI WP_ 050331035.1) consta de dos componentes, PTS-II-BC-glcB (componente IIBC específico de glucosa) y PTS-EIIA-1, mientras que el gen glcB en SC (NCBI WP_ 0503356536.1) consta de dos componentes, PTS-II-BC-glcB y NagE (componente IIA). Se deben realizar esfuerzos para garantizar la coherencia en la nomenclatura génica entre S. carnosus y SC. Sin embargo, el análisis de la secuencia sugirió que estos dos genes codifican componentes de la enzima II cruciales en la absorción de carbohidratos en SC.
Diferencias en la alteración de los perfiles de expresión génica inducidos por eritritol en SC y SP
En la cepa SP, el eritritol reguló al alza ppdK y ppdkR, que son otros genes relacionados con el PTS que se predice que codifican la piruvato fosfato diquinasa (PPDK) y la proteína reguladora de PPDK (PPDKR), respectivamente (26). El mismo estudio también reveló que el eritritol reguló a la baja la expresión de los genes de operones pur implicados en la síntesis de ácido inosínico (IMP), lo que conduce a la biosíntesis de purinas en la cepa SP. Sin embargo, tales cambios no fueron reconocidos en el presente estudio utilizando las cepas SC. Las posibles razones de esta discrepancia incluyen diferencias en las especies bacterianas o que tales cambios son un fenómeno tardío que ocurre en respuesta a la inanición de glucosa. De hecho, PPDK y PPDKR contribuyen a la regeneración de PEP necesaria para reactivar PTS (36, 37), y el pirofosfato de fosforribosilo, precursor de IMP, es un metabolito derivado de la glucosa 6-fosfato a través de la vía de las pentosas fosfato (38). La expresión de genes que codifican vraTSR, que se asocian con la resistencia a β-lactámicos y glicopéptidos en S. aureus (39-43), y sgtB, que está involucrado en la biosíntesis de proteoglicanos en S. aureus (44), también se reguló al alza en la cepa SP. El presente estudio reveló ligeros aumentos en la expresión de los genes vraS, sgtB y vraR (|log 2FC| < 1.9) (Cuadro complementario 3). Sin embargo, en este estudio no se identificó la importancia biológica de estos cambios inducidos por el eritritol en el CS porque no se evaluaron los cambios en la susceptibilidad del SC a los antibióticos.
Perspectivas futuras sobre la aplicación clínica del eritritol
También revelamos que el eritritol inhibió el crecimiento de SC aislado de las lesiones cutáneas de caninos con pioderma. Un estudio previo reveló que el eritritol tiene un efecto bacteriostático sobre las bacterias asociadas a la enfermedad periodontal canina (45). Sobre la base de estos hallazgos, asumimos que las cepas SC cuyo crecimiento se inhibió en presencia de eritritol pueden crecer en placas de agar para el recuento de colonias y, por lo tanto, no analizamos el efecto del eritritol en las unidades formadoras de colonias. Los estudios han informado que el eritritol fue más eficiente en la inhibición del crecimiento de estreptococos comensales orales humanos que el xilitol (18), mientras que dicha eficacia del eritritol contra los aislados clínicos de SP y SC fue similar a la del xilitol (25).
A diferencia de los humanos, el rango de dosis segura de xilitol en perros es estrecho y existe un riesgo de toxicidad por xilitol, como hipoglucemia e insuficiencia hepática aguda, lo que plantea problemas de seguridad (46, 47). Por el contrario, el eritritol era menos probable que causara hipoglucemia y parecía ser seguro para los perros, según lo determinado por los estudios de toxicidad oral (48, 49). Por lo tanto, se espera que la aplicación tópica de eritritol tenga el potencial de aliviar la gravedad clínica de la pioderma canina causada por SC o prevenir su recurrencia. Se esperan futuros ensayos clínicos con eritritol aplicado tópicamente para la pioderma canina. También se anticipa que el eritritol puede prevenir las infecciones por SC a través de equipos médicos contaminados en humanos y animales.
Conclusión
Nuestros hallazgos sugieren que la inanición de glucosa en respuesta al eritritol contribuye a la inhibición del crecimiento en el SC. Nuestros hallazgos también sugieren el potencial del eritritol en la prevención de infecciones cutáneas asociadas a SC en perros y la contaminación de equipos médicos.
Declaración de disponibilidad de datos
Los datos de RNA-seq están disponibles en Gene Expression Omnibus (GEO) en el Centro Nacional de Información Biotecnológica con número de acceso GEO GSE245057. Todos los demás datos están disponibles a través del autor corresponsal previa solicitud razonable.
Contribuciones de los autores
SO-S: Análisis formal, Investigación, Redacción – borrador original. TF: Análisis formal, Conceptualización, Metodología, Redacción – revisión y edición. KW: Escritura – revisión y edición, investigación. RM: Investigación, Escritura, revisión y edición. KI: Investigación, Redacción – revisión y edición, Análisis formal. YT: Redacción – revisión y edición, Metodología. TT: Escritura – revisión y edición, conceptualización. KN: Conceptualización, Escritura – revisión y edición.
Financiación
El/los autor/es declaran/n que no se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo.
Reconocimientos
Los autores agradecen a todos los miembros del Centro de Investigación y Desarrollo de B Food Science Co., Ltd., por apoyar este estudio. Los autores también agradecen a Edanz (https://jp.edanz.com/ac) por editar un borrador de este manuscrito.
Conflicto de intereses
RM es un empleado de B Food Science Co., Ltd. TF y TT eran empleados de la misma empresa en el momento en que se inició este estudio. KI fue empleado de Vet Derm Tokyo Ltd., donde YT era el director representante.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2023.1272595/full#supplementary-material
Referencias
1. Madhaiyan M, Wirth JS, Saravanan VS. Los análisis filogenómicos de la familia Staphylococcaceae sugieren la reclasificación de cinco especies dentro del género Staphylococcus como sinónimos heterotípicos, la promoción de cinco subespecies a especies nuevas, la reasignación taxonómica de cinco especies de Staphylococcus a Mammaliicoccus gen. nov, y la asignación formal de Nosocomiicoccus a la familia Staphylococcaceae. Int J Syst Evol Microbiol. (2020) 70:5926–36. doi: 10.1099/ijsem.0.004498
2. Igimi S, Takahashi E, Mitsuoka T. Staphylococcus schleiferi subsp. coagulans subsp nov, aislado del meato auditivo externo de perros con otitis del oído externo. Int J Syst Bacteriol. (1990) 40:409–11. doi: 10.1099/00207713-40-4-409
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Bes M, Guerin-Faublee V, Freney J, Etienne J. Aislamiento de Staphylococcus schleiferi subspecies coagulans a partir de dos casos de pioderma canino. Rec. Veterinario (2002) 150:487–8. doi: 10.1136/vr.150.15.487
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Frank LA, Kania SA, Hnilica KA, Wilkes RP, Bemis DA. Aislamiento de Staphylococcus schleiferi de perros con pioderma. J Am Vet Med Assoc. (2003) 222:451–4. doi: 10.2460/javma.2003.222.451
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. May ER, Hnilica KA, Frank LA, Jones RD, Bemis DA. Aislamiento de Staphylococcus schleiferi de perros sanos y perros con otitis, pioderma o ambos. J Am Vet Med Assoc. (2005) 227:928–31. doi: 10.2460/javma.2005.227.928
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Naing SY, Duim B, Broens EM, Schweitzer V, Zomer A, van der Graaf-van Bloois L, et al. Caracterización molecular y relevancia clínica de la reasignación taxonómica de la subespecie Staphylococcus schleiferi en dos especies separadas, Staphylococcus schleiferi y Staphylococcus coagulans. Microbiol Spectr. (2023):E0467022. doi: 10.1128/spectrum.04670-22
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Loeffler A, Lloyd DH. ¿Qué ha cambiado en la pioderma canina? Una revisión narrativa. Veterinario J. (2018) 235:73–82. doi: 10.1016/j.tvjl.2018.04.002
8. Thibodeau E, Boucher H, Denofrio D, Pham DT, Snydman D. Primer reporte de una infección por dispositivo de asistencia ventricular izquierda causada por Staphylococcus schleiferi subespecie coagulans: un microorganismo coagulasa positivo. Diagn Microbiol Infect Dis. (2012) 74:68–9. doi: 10.1016/j.diagmicrobio.2012.05.027
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Kobayashi T, Ikeda M, Ohama Y, Murono K, Ikeuchi K, Kitaura S, et al. Primer caso humano de infección del torrente sanguíneo relacionada con el catéter causada por Staphylococcus schleiferi subespecie coagulans: informe de un caso y revisión de la literatura. Ann Clin Microbiol Antimicrob. (2021) 20:68. doi: 10.1186/s12941-021-00474-3
10. Papá RK, Avais M, Khan JA, Anjum AA. Evaluación de la efectividad de las vacunas multirresistentes contra la mastitis en ganado lechero. Pak Vet J. (2022) 42:300–7. doi: 10.29261/pakvetj/2022.038
11. Aljahani AH, Alarjani KM, Hassan ZK, Elkhadragy MF, Ismail EA, Al-Masoud AH, et al. Detección molecular de cepas de Staphylococcus aureus resistentes al calor con meticilina en leche de camella pasteurizada en Arabia Saudita. Biosci Rep. (2020) 40:BSR20193470. doi: 10.1042/BSR20193470
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Zehra A, Singh R, Kaur S, Gill JPS. Caracterización molecular de Staphylococcus aureus resistente a antibióticos en ganado bovino (bovino y porcino). Mundo Veterinario. (2017) 10:598–604. doi: 10.14202/vetworld.2017.598-604
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Abd-Elfatah EB, Fawzi EM, Elsheikh HA, Shehata AA. Prevalencia, genes de virulencia y patrón de susceptibilidad al antibiograma de Staphylococcus aureus y Streptococcus agalactiae aislados de ovejas mastíticas. Int J Vet Sci. (2023) 12:152–60. doi: 10.47278/journal.ijvs/2022.176
14. Cain CL, Morris DO, O’Shea K, Rankin SC. Relación genotípica y caracterización fenotípica de la subespecie Staphylococcus schleiferi en muestras clínicas de perros. Am J Vet Res. (2011) 72:96–102. doi: 10.2460/ajvr.72.1.96
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Cain CL, Morris DO, Rankin SC. Caracterización clínica de las infecciones por Staphylococcus schleiferi e identificación de factores de riesgo para la adquisición de cepas resistentes a oxacilina en perros: 225 casos (2003-2009). J Am Vet Med Assoc. (2011) 239:1566–73. doi: 10.2460/javma.239.12.1566
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Huse HK, Miller SA, Chandrasekaran S, Hindler JA, Lawhon SD, Bemis DA, et al. Evaluación de la difusión en disco de oxacilina y cefoxitina y de los puntos de ruptura de la micro establecidos por el instituto de estándares clínicos y de laboratorio para la detección de la resistencia a la oxacilina mediada por meca en Staphylococcus schleiferi. J Clin Microbiol. (2018) 56:e01653–17. doi: 10.1128/JCM.01653-17
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Kawakami T, Shibata S, Murayama N, Nagata M, Nishifuji K, Iwasaki T, et al. Susceptibilidad antimicrobiana y resistencia a la meticilina en Staphylococcus pseudintermedius y Staphylococcus schleiferi subsp. coagulans aislados de perros con pioderma en Japón. J Vet Med Sci. (2010) 72:1615–9. doi: 10.1292/jvms.10-0172
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. de Cock P, Makinen K, Honkala E, Saag M, Kennepohl E, Eapen A. El eritritol es más eficaz que el xilitol y el sorbitol en el manejo de los criterios de valoración de la salud bucal. Int J Dent. (2016) 2016:9868421. doi: 10.1155/2016/9868421
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Hashino E, Kuboniwa M, Alghamdi SA, Yamaguchi M, Yamamoto R, Cho H, et al. El eritritol altera la microestructura y los perfiles metabolómicos de la biopelícula compuesta por Streptococcus gordonii y Porphyromonas gingivalis. Mol Microbiol Oral. (2013) 28:435–51. doi: 10.1111/omi.12037
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Park Y-N, Jeong S-S, Zeng J, Kim S-H, Hong S-J, Ohk S-H, et al. Efectos anticariogénicos del eritritol sobre el crecimiento y la adhesión de Streptococcus mutans. Biotecnología de la ciencia de los alimentos. (2014) 23:1587–91. doi: 10.1007/s10068-014-0215-0
21. Shimizu M, Miyawaki S, Kuroda T, Umeta M, Kawabe M, Watanabe K. El eritritol inhibe el crecimiento de bacterias asociadas a la enfermedad periodontal aisladas de la cavidad oral canina. Heliyon. (2022) 8:E10224. doi: 10.1016/j.heliyon.2022.e10224
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Fujii T, Inoue S, Kawai Y, Tochio T, Takahashi K. Supresión del olor axilar y control de la flora bacteriana axilar por eritritol. J Cosmet Dermatol. (2022) 21:1224–33. doi: 10.1111/jocd.14201
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Fujii T, Tochio T, Endo A. Inhibición y promoción del crecimiento dependiente del ribotipo por eritritol en Cutibacterium acnes. J Cosmet Dermatol. (2022) 21:5049–57. doi: 10.1111/jocd.14958
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Miyasawa-Hori H, Aizawa S, Takahashi N. Diferencia en la sensibilidad al xilitol de la producción de ácido entre las cepas de Streptococcus mutans y el mecanismo bioquímico. Microbiol Oral Immunol. (2006) 21:201–5. doi: 10.1111/j.1399-302X.2006.00273.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Tochio T, Kawano K, Iyori K, Makida R, Kadota Y, Fujii T, et al. El eritritol tópico combinado con L-ascorbil-2-fosfato inhibe el crecimiento estafilocócico y alivia el sobrecrecimiento estafilocócico en lesiones cutáneas de pioderma superficial canino. Pol J Vet Sci. (2023) 26:647–55. doi: 10.24425/pjvs.2023.148284
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Fujii T, Tochio T, Nishifuji K. El eritritol altera las firmas del transcriptoma génico, el crecimiento celular y la formación de biopelículas en Staphylococcus pseudintermedius. BMC Vet Res. (2023) 19:146. doi: 10.1186/s12917-023-03711-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Schissler JR, Hillier A, Daniels JB, Cole LK, Gebreyes WA. Evaluación de los criterios interpretativos del Instituto de Estándares de Laboratorio Clínico para Staphylococcus pseudintermedius resistente a la meticilina aislado de perros. J Vet Diagn Invest. (2009) 21:684–8. doi: 10.1177/104063870902100514
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Sasaki T, Tsubakishita S, Tanaka Y, Sakusabe A, Ohtsuka M, Hirotaki S, et al. Método de PCR múltiple para la identificación de especies de estafilococos coagulasa positivos. J Clin Microbiol. (2010) 48:765–9. doi: 10.1128/JCM.01232-09
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. González-Domínguez MS, Carvajal HD, Calle-Echeverri DA, Chinchilla-Cárdenas D. Detección molecular y caracterización de los genes mecA y nuc de especies de Staphylococcus (S. aureus, S. pseudintermedius y S. schleiferi) aisladas de perros con pioderma superficial y sus perfiles de resistencia a los antimicrobianos. Front Vet Sci. (2020) 7:376. doi: 10.3389/fvets.2020.00376
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Iyori K, Shishikura T, Shimoike K, Minoshima K, Imanishi I, Toyoda Y. Influencia del tamaño del hospital en la resistencia a los antimicrobianos y ventajas de restringir el uso de antimicrobianos en función de los antibiogramas acumulativos en perros con infecciones por Staphylococcus pseudintermedius en Japón. Veterinario Dermatol. (2021) 32:668-E178. doi: 10.1111/vde.12975
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Crawford EC, Singh A, Metcalf D, Gibson TW, Weese SJ. Identificación de genes de referencia apropiados para estudios de qPCR en Staphylococcus pseudintermedius y evaluación preliminar de la expresión génica icaA en bacterias incluidas en biopelículas. Notas de BMC Res. (2014) 7:451. doi: 10.1186/1756-0500-7-451
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Christiansen I, Hengstenberg W. Sistema fosfotransferasa dependiente de fosfoenolpiruvato estafilocócico: dos permeasas de glucosa muy similares en Staphylococcus carnosus con diferente especificidad de glucósidos: ¿ingeniería de proteínas in vivo? Microbiología (Lectura). (1999) 145:2881–9. doi: 10.1099/00221287-145-10-2881
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Knezevic I, Bachem S, Sickmann A, Meyer HE, Stulke J, Hengstenberg W. Regulación del sistema fosfotransferasa específico de glucosa (PTS) de Staphylococcus carnosus por la proteína antiterminadora GlcT. Microbiología (Lectura). (2000) 146:2333–42. doi: 10.1099/00221287-146-9-2333
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Deutscher J, Ake FM, Derkaoui M, Zebre AC, Cao TN, Bouraoui H, et al. El sistema fosfoenolpiruvato bacteriano: carbohidrato fosfotransferasa: regulación por fosforilación de proteínas e interacciones proteína-proteína dependientes de fosforilación. Microbiol Mol Biol Rev. (2014) 78:231–56. doi: 10.1128/MMBR.00001-14
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Christiansen I, Hengstenberg W. Clonación y secuenciación de dos genes de Staphylococcus carnosus que codifican para PTS específico de glucosa y su expresión en Escherichia coli K-12. Mol Gen Genet. (1996) 250:375–9. doi: 10.1007/BF02174396
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Cui J, Maloney MI, Olson DG, Lynd LR. Conversión de fosfoenolpiruvato en piruvato en Thermoanaerobacterium saccharolyticum. Metab Eng Commun. (2020) 10:e00122. doi: 10.1016/j.mec.2020.e00122
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Chastain CJ, Xu W, Perejil K, Sarath G, Hibberd JM, Chollet R. Las proteínas reguladoras de piruvato y ortofosfato dicinasa de Arabidopsis poseen una estructura primaria de proteína quinasa Ser/Thr novedosa y sin precedentes. Planta J. (2008) 53:854–63. doi: 10.1111/j.1365-313X.2007.03366.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. García Sánchez R, Hahn-Hagerdal B, Gorwa-Grauslund MF. Reacciones cruzadas entre las vías de la xilosa y la galactosa modificadas en Saccharomyces cerevisiae recombinante. Biotecnol Biocombustibles. (2010) 3:19. doi: 10.1186/1754-6834-3-19
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Boyle-Vavra S, Yin S, Jo DS, Montgomery CP, Daum RS. VraT/YvqF es necesario para la resistencia a la meticilina y la activación del regulón VraSR en Staphylococcus aureus. Agentes antimicrobianos quimiotera. (2013) 57:83–95. doi: 10.1128/AAC.01651-12
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Kuroda M, Kuroda H, Oshima T, Takeuchi F, Mori H, Hiramatsu K. El sistema de dos componentes VraSR modula positivamente la regulación de la vía de biosíntesis de la pared celular en Staphylococcus aureus. Mol Microbiol. (2003) 49:807–21. doi: 10.1046/j.1365-2958.2003.03599.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Gardete S, Wu SW, Gill S, Tomasz A. Papel de VraSR en la resistencia a los antibióticos y la respuesta al estrés inducida por antibióticos en Staphylococcus aureus. Agentes antimicrobianos quimiotera. (2006) 50:3424–34. doi: 10.1128/AAC.00356-06
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Boyle-Vavra S, Yin S, Daum RS. El sistema regulador de dos componentes VraS/VraR requerido para la resistencia a la oxacilina en Staphylococcus aureus resistente a la meticilina adquirida en la comunidad. FEMS Microbiol Lett. (2006) 262:163–71. doi: 10.1111/j.1574-6968.2006.00384.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Mehta S, Cuirolo AX, Plata KB, Riosa S, Silverman JA, Rubio A, et al. El sistema regulador de dos componentes VraSR contribuye a la disminución de la susceptibilidad a la daptomicina mediada por mprF en cepas clínicas seleccionadas in vivo de Staphylococcus aureus resistente a la meticilina. Agentes antimicrobianos quimiotera. (2012) 56:92–102. doi: 10.1128/AAC.00432-10
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Wu S, Lin K, Liu Y, Zhang H, Lei L. Las vías de señalización de dos componentes modulan la resistencia a los medicamentos de Staphylococcus aureus (Revisión). Biomed Rep. (2020) 13:5. doi: 10.3892/br.2020.1312
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Tochio T, Makida R, Fujii T, Kadota Y, Takahashi M, Watanabe A, et al. El efecto bacteriostático del eritritol sobre las bacterias relacionadas con la enfermedad periodontal canina. Pol J Vet Sci. (2022) 25:75–82. doi: 10.24425/pjvs.2022.140843
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Dunayer EK, Gwaltney-Brant SM. Insuficiencia hepática aguda y coagulopatía asociada a la ingestión de xilitol en ocho perros. J Am Vet Med Assoc. (2006) 229:1113–7. doi: 10.2460/javma.229.7.1113
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Schmid RD, Hovda LR. Insuficiencia hepática aguda en un perro después de la ingestión de xilitol. J Med Toxicol. (2016) 12:201–5. doi: 10.1007/s13181-015-0531-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Decano I, Jackson F, Greenough RJ. Estudio de toxicidad oral crónica (1 año) de eritritol en perros. Regul Toxicol Pharmacol. (1996) 24:S254-60. doi: 10.1006/rtph.1996.0106
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Eapen AK, de Cock P, Crincoli CM, Means C, Wismer T, Pappas C. Estudios de toxicidad oral aguda y subcrónica del eritritol en perros Beagle. Toxicol químico alimentario. (2017) 105:448–55. doi: 10.1016/j.fct.2017.04.049
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: eritritol, Staphylococcus coagulans, perro, pioderma, glucosa, sistema fosfotransferasa
Cita: Onishi-Sakamoto S, Fujii T, Watanabe K, Makida R, Iyori K, Toyoda Y, Tochio T y Nishifuji K (2024) El eritritol altera la expresión génica de la fosfotransferasa e inhibe el crecimiento in vitro de Staphylococcus coagulans aislados de caninos con pioderma. Frente. Vet. Sci. 10:1272595. doi: 10.3389/fvets.2023.1272595
Recibido: 04 de agosto de 2023; Aceptado: 11 de diciembre de 2023;
Publicado: 04 Enero 2024.
Editado por:
Kun Li, Universidad Agrícola de Nanjing, China
Revisado por:
Meera Surendran Nair, Universidad Estatal de Pensilvania (PSU), Estados
Unidos Amjad Islam Aqib, Universidad de Ciencias Veterinarias y Animales de Cholistán, Pakistán
Derechos de autor © 2024 Onishi-Sakamoto, Fujii, Watanabe, Makida, Iyori, Toyoda, Tochio y Nishifuji. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Koji Nishifuji, kojimail@cc.tuat.ac.jp
†ORCID: Saki Onishi-Sakamoto orcid.org/0009-0009-9303-283X
Keita Iyori orcid.org/0000-0003-1977-4201
Takumi Tochio orcid.org/0000-0003-3206-4116
Koji Nishifuji orcid.org/0000-0003-4583-5485
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía