
Análisis metabolómico del efecto de la GnRH sobre la tasa de preñez de ovejas con esquema de sincronización del celo basado en progesterona

Análisis metabolómico del efecto de la GnRH sobre la tasa de preñez de ovejas con esquema de sincronización del celo basado en progesterona

 Sol de Shuyuan1
Sol de Shuyuan1 Xinyu Bai2Nana Yang1Yiyong Liu3
Xinyu Bai2Nana Yang1Yiyong Liu3 Xinglong Wu1*Xiangyun Li1*
Xinglong Wu1*Xiangyun Li1*- 1Facultad de Ciencia y Tecnología Animal, Centro de Innovación Tecnológica de Embriones de Ganado Vacuno y Ovino de Hebei, Universidad Agrícola de Hebei, Baoding, Hebei, China
- número arábigoFacultad de Ciencia Animal, Universidad de Tarim, Alear, Xinjiang, China
- 3Instituto de Ciencia Animal Yili de Xinjiang, Yining, Xinjiang, China
Introducción: La hormona liberadora de gonadotropina (GnRH) se utiliza ampliamente en el protocolo de inseminación artificial programada para ovejas. Sin embargo, sigue existiendo un debate sobre su impacto en las tasas de embarazo durante la inseminación artificial. Este estudio tiene como objetivo evaluar el efecto de la GnRH en las tasas de preñez en ovejas Huyang, analizar los cambios en los metabolitos preimplantacionales causados por la GnRH utilizando la metabolómica y dilucidar el efecto del mecanismo en las tasas de preñez.
Métodos: A todas las ovejas se les administró una esponja vaginal de progesterona que contenía 45 mg de acetato de flurogestona durante 12 días y recibieron 330 unidades de gonadotropina coriónica equina (eCG) por vía intramuscular después de la extracción de la esponja. El grupo experimental (n = 69) recibió un tratamiento intramuscular de 17 μg de triptorelina agonista de la GnRH 48 h después de la retirada de la esponja el día 0, mientras que el grupo control (n = 41) recibió 1 mL de solución salina estéril. Todas las ovejas se sometieron a una única inseminación vaginal 58 h después de la retirada de la esponja de progesterona. Se calculó la diferencia en las tasas de embarazo entre los dos grupos. El análisis metabolómico se realizó en muestras de plasma recogidas el día 7 después del tratamiento con el agonista de la GnRH.
Resultados: El tratamiento con hormona liberadora de gonadotropina (GnRH) redujo significativamente la tasa de embarazo en el grupo experimental en comparación con el grupo control (72,2 vs. 82,9%, p < 0,05). El análisis metabolómico indicó que el tratamiento con GnRH afectó a los metabolitos implicados en la síntesis de colágeno y la síntesis de prostaglandinas en el tejido endometrial, lo que incluye una marcada disminución del contenido de aminoácidos hidroxiprolina y un aumento significativo de los lípidos y ácidos grasos insaturados de corticosterona y prostaglandina D2.
Conclusión: En resumen, la inyección del agonista de la GnRH triptorelina 48 h después de la retirada de las esponjas de progesterona reduce la tasa de preñez de la oveja Huyang después de la inseminación artificial. También afecta a los niveles de metabolitos relacionados con la síntesis endometrial de colágeno y prostaglandinas, perjudicando la implantación embrionaria.
1 Introducción
La promoción de la sincronización del celo y la tecnología de inseminación artificial en la industria ovina puede aumentar la producción de corderos, mejorar la eficiencia reproductiva y mejorar el manejo del rebaño y el mejoramiento genético (1). La cría sincronizada da como resultado un parto sincronizado, lo que simplifica la gestión centralizada y la venta de corderos (2). En la práctica de la producción, un programa de sincronización del celo comúnmente utilizado para las ovejas que se someten a inseminación artificial o que sirven como receptoras en el proceso de Ovulación Múltiple y Transferencia de Embriones (MOET) implica el uso de insertos vaginales o esponjas que contienen acetato de fluorogestona o acetato de medroxiprogesterona y eCG (3, 4). Numerosos estudios han demostrado la eficacia del uso de la hormona liberadora de gonadotropina (GnRH) para sincronizar e inducir la ovulación en la reproducción de los animales de granja (5-7). Menchaca et al. (2) han demostrado que la administración de una dosis única de GnRH 24-36 h después de retirar el dispositivo intravaginal liberador de progesterona da lugar a un pico de hormona luteinizante aproximadamente 40 h después. Además, el 90% de las ovejas ovulan dentro de las 72 horas posteriores a la retirada del dispositivo.
Aunque la GnRH se ha utilizado ampliamente en proyectos de Inseminación Artificial a Tiempo Fijo (FTAI) para ovinos (8), lo que ha dado lugar a una cierta mejora del beneficio económico en la producción ganadera (9), su efecto sobre las tasas de preñez sigue siendo objeto de debate (2). Algunos estudios indicaron que la administración de una dosis única de GnRH después de la retirada del pesario aumenta las tasas de embarazo (10, 11), en contraste con los hallazgos de ningún efecto (6, 12, 13) o una disminución en las tasas de embarazo en otros estudios (14). De manera similar, en vacas que experimentan sincronización de celo, la administración de GnRH durante la inseminación conduce a la ovulación de folículos fisiológicamente inmaduros, lo que afecta la tasa de preñez de las vacas y la supervivencia fetal en gestación posterior (15). La exposición del folículo preovulatorio al aumento de gonadotropina desencadenado por la GnRH exógena puede reducir la calidad de los ovocitos, disminuyendo la tasa de embarazo (16). Además, se ha inferido que la GnRH podría dificultar la implantación embrionaria, afectando consecuentemente las tasas de embarazo en los protocolos FTAI (17).
La metabolómica es una tecnología importante para el estudio de las redes metabólicas de los sistemas biológicos. Implica el análisis cuantitativo de todos los metabolitos en los organismos y la exploración de las relaciones relativas entre los metabolitos y los cambios fisiopatológicos (18). Esta técnica se aplica ampliamente en la investigación de plantas (19), el desarrollo de fármacos (20) y el estudio de enfermedades (21). Varios estudios han utilizado la metabolómica para detectar la calidad de la carne ovina (22), la alimentación y la nutrición (23) y las enfermedades (24). Estudios relevantes también han utilizado la metabolómica para analizar la composición del moco cervical de la oveja y el impacto negativo de las respuestas inflamatorias en el transporte de semen en el cuello uterino (25). Sin embargo, la investigación sobre el entorno de implantación durante el período periimplantacional sigue siendo limitada.
En el estudio, las ovejas Huyang fueron tratadas con sincronización de celo, se inyectó un agonista de GnRH 48 h después de la retirada de las esponjas de progesterona y se comparó la diferencia en la tasa de preñez 40 días después de la inseminación. Se recolectaron muestras de sangre en el día 7 después del tratamiento con GnRH para el análisis metabolómico para identificar las vías metabólicas y los metabolitos que afectan las tasas de preñez en las ovejas Huyang causadas por GnRH. El objetivo del presente estudio fue evaluar el efecto del tratamiento con GnRH sobre las tasas de preñez y los metabolitos preimplantacionales en ovejas Huyang.
2 Materiales y métodos
2.1 Animales y ubicación
El estudio se llevó a cabo en enero de 2023 en Hulunbeir, Mongolia Interior. Se encuentra a 120°28′E de longitud y 47°5′N de latitud. Esta región experimenta un clima continental templado y recibe precipitaciones anuales que oscilan entre 300 y 500 mm. Los inviernos son fríos y áridos, mientras que los veranos son calurosos y lluviosos, lo que provoca fluctuaciones significativas en las temperaturas anuales y diarias. El experimento seleccionó 23 ovejas Huyang sanas y multíparas (de 1,5 a 4 años, con un peso de 40 a 45 kg, BSC ≥ 3, que van desde la demacración hasta la obesidad). Todas las ovejas que participan en el experimento poseen tractos reproductivos sanos y capacidades de lactancia, con un período promedio de destete superior a 3 meses, lo que las califica para el tratamiento de sincronización de celo. Durante el período experimental, las ovejas se mantuvieron alejadas de los carneros para evitar el apareamiento voluntario. Las ovejas se mantuvieron en el interior por la noche y se les permitió pastar en los pastos naturales de la granja durante todo el día. Cuando las ovejas se mantenían en el interior, todas las ovejas recibían TMR, incluyendo heno de alfalfa, cebada y concentrado mixto de acuerdo con la recomendación de la NRC, y tenían libre acceso al agua y al bloque mineral. Este estudio fue aprobado por el Comité de Cuidado y Uso de Animales de la Universidad Agrícola de Hebei, Baoding, China (Número: 2021011). Se hizo todo lo posible para minimizar el sufrimiento de los animales. Los autores declaran que todos los procedimientos del experimento se llevaron a cabo de manera consistente con los preceptos del bienestar animal, y que el personal involucrado en el cuidado y manejo de los animales eran veterinarios autorizados.
2.2 Sincronización del celo y tratamiento hormonal
Todas las ovejas experimentales fueron ovejas Huyang autóparas y multíparas, que se sincronizaron mediante un protocolo de combinación progestágeno-eCG. Cada oveja recibió una esponja intravaginal de poliuretano impregnada con 45 mg de acetato de flurogestona (Muqimuye Sci-Tech Co., Ltd., Shanghai, China) durante 12 días, seguida de un tratamiento con 330 UI de eCG i.m. (Sansheng Biological Technology Co., Ltd., Ningbo, China) en el momento de la extracción de la esponja. El grupo experimental (B, n = 69) fue sometido a 48 h (Día 0) después de la retirada de la esponja a la administración intramuscular de 17 μg del agonista de la GnRH triptorelina (Sansheng Biological Technology Co., Ltd., Ningbo, China), y el grupo control (Y, n = 41) con 1 mL de suero fisiológico estéril. Todas las ovejas se sometieron a una única inseminación vaginal 58 h después de la retirada de las esponjas de progesterona.
2.3 Inseminación artificial a tiempo fijo
Se recolectaron espermatozoides frescos de seis carneros Huyang sanos, y se evaluó la motilidad y densidad de los espermatozoides utilizando un hemocitómetro y un microscopio óptico. Los espermatozoides (motilidad progresiva superior al 50% y densidad superior a 1 × 109 espermatozoides/ml) se diluyeron utilizando 5-7 veces el volumen de leche desnatada (leche desnatada Yili, Mongolia Interior Yili Industrial Group Limited by Share Ltd), asegurando que la motilidad de los espermatozoides después de la dilución se mantuviera por encima de 0,5, y la densidad se ajustó a 4 × 108 espermatozoides/mL. A continuación, los espermatozoides diluidos se almacenaron en una tetera al baño de agua a 30 °C. Las ovejas experimentales fueron inmovilizadas boca abajo y su moco vaginal se limpió con solución salina normal tibia para garantizar la higiene del entorno vaginal, reduciendo así el impacto en la viabilidad de los espermatozoides. Se insertó un espéculo vaginal para abrir la vagina y se observó y localizó el orificio cervical. Se utilizó un inseminador específico para ovejas (producido por Baoding Zhengmu Biotechnology Co., Ltd.) para aspirar 0,3 ml de semen e inyectarlo en el orificio cervical.
2.4 Toma de muestras de sangre
Las ovejas fueron seleccionadas aleatoriamente del grupo experimental (n = 13) y del grupo control (n = 10). En el grupo experimental (n = 13) y en el grupo control (n = 10), se recolectaron 5 mL de sangre venosa yugular utilizando tubos de extracción de sangre al vacío que contenían heparina sódica en el día 7 después del tratamiento con el agonista de la GnRH. Posteriormente, las muestras de sangre se sometieron a centrifugación a 3.000 × g durante 15 min a 4°C. El plasma sobrenadante resultante se separó cuidadosamente y se transfirió a tubos EP de 1,5 ml libres de enzimas utilizando una pipeta de precisión. Las muestras de plasma se almacenaron inmediatamente a -80 °C para preservar su integridad para su posterior análisis metabolómico.
2.5 Detección por cromatografía líquida sérica-espectrometría de masas en tándem
Mezcle 100 μL de la muestra con 400 μL de metanol preenfriado, asegurando una mezcla completa por vórtice. Incubar la mezcla en hielo durante 5 minutos, centrifugarla a 15.000 × g y a 4 °C durante 20 minutos. Diluya una porción del sobrenadante resultante con agua de grado LC-MS para lograr una concentración final de metanol del 53%. A continuación, transfiera la muestra diluida a un nuevo tubo de centrífuga utilizando un filtro de 0,22 μm y vuelva a centrifugarla a 15.000 × g y 4 °C durante 20 min. Finalmente, la muestra filtrada se inyecta en el sistema LC-MS/MS para su análisis.
El análisis LC-MS/MS se llevó a cabo utilizando el sistema UHPLC Vanquish (ThermoFisher, Alemania) junto con un espectrómetro de masas Orbitrap Q ExactiveTMHF-X (ThermoFisher, Alemania). Las muestras se inyectaron en una columna cromatográfica Hypersil Gold (100 mm × 2,1 mm, 1,9 μm) a un caudal de 0,2 ml/min utilizando un gradiente lineal de 17 min. Para el modo de polaridad positiva, las fases móviles consistieron en el eluyente A (0,1% de ácido fórmico en agua) y el eluyente B (metanol). Para el modo de polaridad negativa, las fases móviles fueron el eluyente A (acetato de amonio de 5 mM, pH 9,0) y el eluyente B (metanol). El gradiente de disolvente se estableció de la siguiente manera: 2%B, 1,5 min; 2–85%B, 3 min; 85–100%B, 10 min; 100–2%B, 10,1 min; 2%B, 12 min. El espectrómetro de masas Q ExactiveTMHF-X funcionó tanto en modo positivo como negativo con una tensión de pulverización de 3,5 kV, una temperatura capilar de 320 °C, un caudal de gas de vaina de 35 psi y un caudal de gas auxiliar de 10 L/min, un nivel de RF de lente S de 60, una temperatura del calentador de gas auxiliar de 350 °C.
2.6 Prueba de embarazo
Cuarenta días después de la inseminación, un técnico fijo utilizó un probador de ultrasonido B (DP-30Vet, Shenzhen Mindray Biomedical Electronics Co., Ltd.) para realizar una prueba de embarazo en las ovejas inseminadas a través del abdomen. La ecografía B mostró un saco gestacional claro, que se identificó como embarazo. Las tasas de preñez se calcularon como el porcentaje de ovejas preñadas entre las ovejas inseminadas.
2.7 Análisis de datos
El análisis estadístico se realizó con el software GraphPad Prism 8.0. Las tasas de embarazo en los grupos experimental y control se analizaron mediante una prueba de Chi-cuadrado. Se aceptó como significativo si el valor de p calculado <0,05. Utilice las bases de datos de la Base de Datos del Metaboloma Humano (HMDB, http://www.hmdb.ca/), la Enciclopedia de Genes y Genomas de Kioto (KEGG, http://www.genome.jp/kegg/) y los mapas de lípidos (Programa de Investigación de la Vía del Metabolismo de los Lípidos, https://lipidmaps.org/) para anotar los metabolitos. Utilice metaX para el análisis discriminante de mínimos cuadrados parciales (PLS-DA) y emplee el análisis univariante (prueba T) para calcular la significación estadística (valor p). Se realizó PLS-DA para calcular las puntuaciones de Importancia de la Variable en la Proyección (VIP). Los metabolitos que cumplen con los criterios de Importancia Variable en Proyección (VIP) > 1, valor de p <0.05 y Cambio de Pliegue (FC) > 1.2 o FC < 0.833 se consideran metabolitos diferenciales. Las funciones de estos metabolitos y las vías metabólicas se estudiaron utilizando la base de datos KEGG. Se realizó el enriquecimiento de metabolitos diferenciales por vías metabólicas y se consideró enriquecimiento por vías metabólicas. Cuando el valor de p de la vía metabólica <0,05, las vías metabólicas se consideraron enriquecimiento estadísticamente significativo.
3 Resultados
3.1 Efecto de la GnRH sobre las tasas de embarazo a los 40 días después de la inseminación
El embarazo se detectó mediante ecografía a los 40 días de la inseminación, siendo el resultado el que se muestra en la Tabla 1. La tasa de embarazo del grupo experimental fue significativamente menor que la del grupo control (72,2 vs. 82,9%, p < 0,05). Los resultados mostraron que el tratamiento con GnRH durante la inseminación artificial disminuyó las tasas de preñez en las ovejas Huyang.

3.2 Clasificación de los metabolitos en ovejas
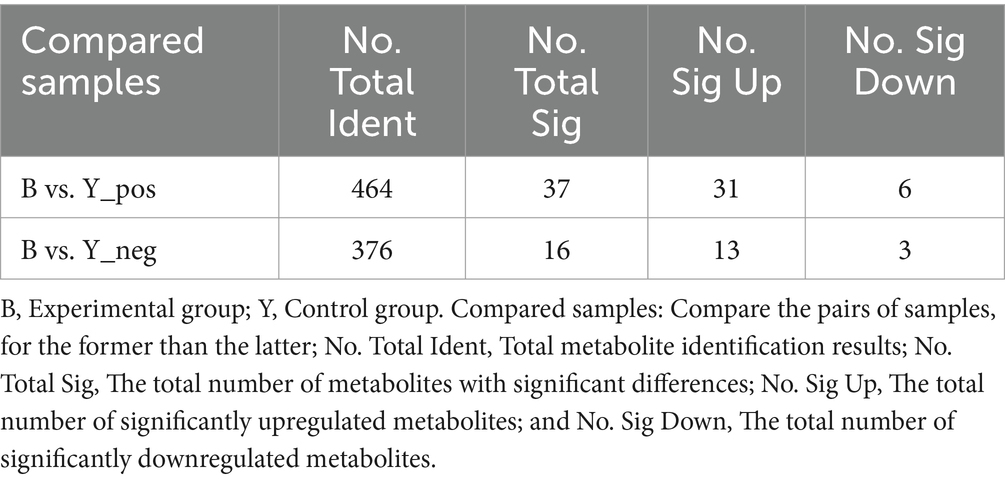
Para investigar los factores que contribuyen a la reducción de las tasas de preñez después del tratamiento con GnRH en ovejas Huyang, se recolectaron muestras de plasma de los grupos experimental y control 7 días después de la inseminación artificial para metabolómica. Las 23 muestras identificaron 464 metabolitos en modo de iones positivos (POS) y 376 en modo de iones negativos (NEG). El análisis del modo iónico positivo reveló una variedad de metabolitos como lípidos y moléculas similares a los lípidos (57,38%), ácidos orgánicos y derivados (14,48%), compuestos organoheterocíclicos (10,86%), bencenoides, (5,01%), fenilpropanoides y policétidos (3,90%), compuestos orgánicos de nitrógeno (3,34%), nucleósidos, nucleótidos y análogos (2,79%), compuestos orgánicos de oxígeno (1,95%) y alcaloides y derivados (0,28%) (Figura 1A). El análisis en modo iónico negativo indicó la existencia de lípidos y moléculas similares a los lípidos (63,93%), ácidos orgánicos y derivados (13,44%), bencenoides, (6,89%), compuestos orgánicos de oxígeno (4,26%), compuestos organoheterocíclicos (3,93%), fenilpropanoides y policétidos (3,93%), nucleósidos, nucleótidos y análogos (2,95%), ninguno (0,33%) y compuestos orgánicos nitrogenados (0,33%) (Figura 1B).
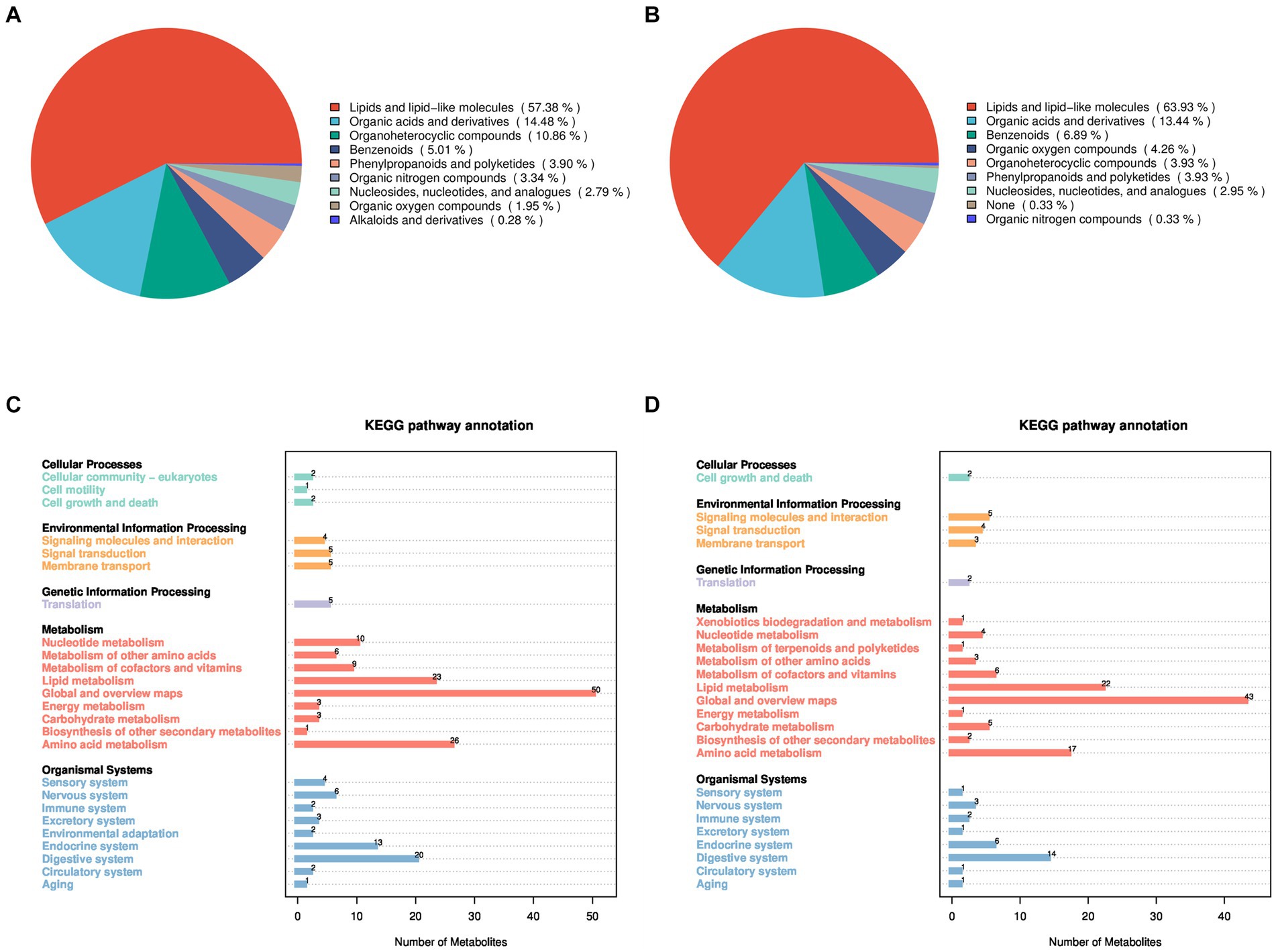
Además, se utilizó la base de datos KEGG para la anotación funcional de los metabolitos identificados con el fin de comprender sus rasgos funcionales y clasificaciones (Figuras 1C, D). En los modos positivo y negativo, los metabolitos identificados participan en procesos de vía relacionados con procesos celulares, procesamiento de información ambiental, procesamiento de información genética, metabolismo y sistemas orgánicos. Específicamente, bajo el modo de iones positivos, los metabolitos participan principalmente en procesos biológicos como el crecimiento y la muerte celular, el transporte de membrana, el metabolismo de los lípidos, el metabolismo energético y el metabolismo de los aminoácidos. En el modo de iones negativos, los metabolitos involucran principalmente procesos biológicos, incluido el crecimiento y la muerte celular, las moléculas de señalización y la interacción, el metabolismo de los lípidos, el metabolismo de los aminoácidos y el sistema inmunológico.
3.3 Cribado y análisis de metabolitos diferenciales
El Análisis Discriminante de Mínimos Cuadrados Parciales estableció un modelo de relación entre la expresión de metabolitos y la categoría de la muestra, facilitando la predicción de la categoría de la muestra. Para discriminar los metabolitos diferenciales entre los dos grupos, se realizó PLS-DA supervisado para encontrar metabolitos diferentes. Como se muestra en la Figura 2, los parámetros de predicción del modelo de evaluación PLS-DA exhiben valores R2Y de 0,84 y 0,78 en los modos de iones positivos y negativos (Figuras 2A, B), con los valores Q2Y de 0,09 y − 0,03, respectivamente. Los valores más altos de R2Y en comparación con Q2Y en ambos modos indican la fiabilidad del modelo utilizado en este estudio.
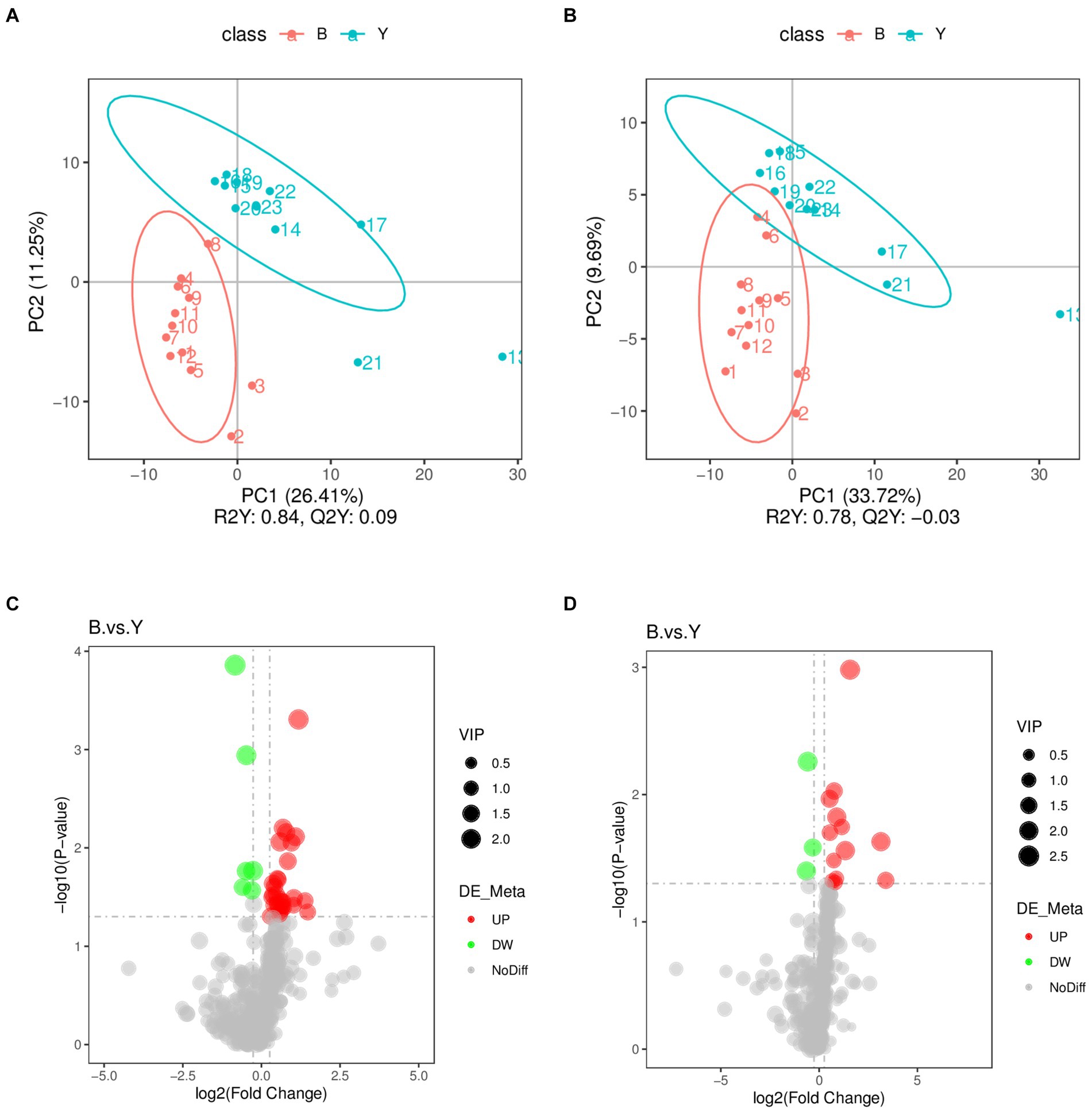
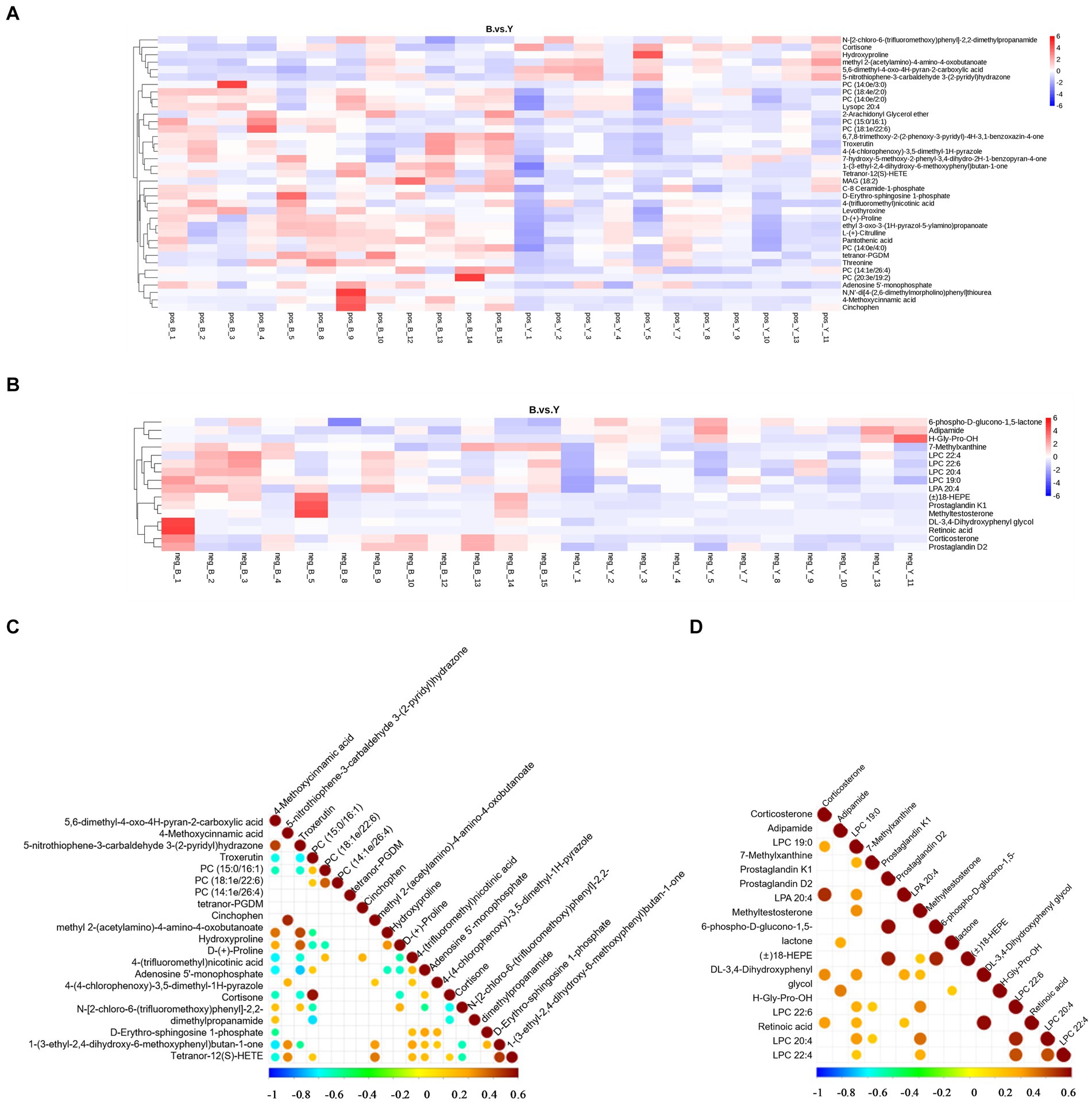
Los datos metabolómicos se analizaron mediante FC, valor de p y VIP en análisis univariado. Posteriormente, se identificaron metabolitos diferenciales basados en VIP > 1,0, FC > 1,2 o FC < 0,833, y p < 0,05. Se identificaron un total de 53 metabolitos que presentaban diferencias significativas: 37 en modo iónico positivo, que comprenden 31 metabolitos regulados al alza y seis regulados a la baja, y 16 en modo de iones negativos, con 13 regulados al alza y tres regulados a la baja (Tabla 2; Figuras 2C y D). Como se muestra en las Figuras 3A, B, el mapa de calor mostró todos los metabolitos diferenciales en los modos de iones positivos y negativos. Los 10 principales metabolitos diferenciales en modo de iones positivos son N,N′-di[4-(2,6-dimetilmorfolino)fenil]tiourea, fosfatidilcolina (20:3e/19:2), ácido 4-metoxicinámico, fosfatidilcolina (18:1e/22:6), éter de 2-araquidonil glicerol, fosfatidilcolina (14:0e/3:0), tetranor-PGDM, cinchofeno, fosfatidilcolina (15:0/16:1) y monoacilglicerol (18:2). Los 10 principales metabolitos diferenciales en modo de iones negativos son el ácido retinoico, la metiltestosterona, la corticosterona, el ácido (±) 18-hidroxieicosa-5,8,11,14,16-pentaenoico (18-HEPE), la prostaglandina D2, la prostaglandina K1, la lisofosfatidilcolina (LPC) 22:6, la LPC 22:4, la LPC 19:0 y la DL-3,4-dihidroxifenilglicol. Entre ellos, los niveles metabólicos de cortisona e hidroxiprolina disminuyeron significativamente, y el nivel de ácido pantoténico aumentó significativamente en el grupo experimental en modo iónico positivo. En el modo de iones negativos, los niveles metabólicos de corticosterona y prostaglandina D2 del grupo experimental aumentaron significativamente (Figura 3B). El análisis de correlación de metabolitos diferenciales se realizó mediante análisis de correlación de Pearson (Figuras 3C, D). En el modo iónico positivo, los niveles metabólicos de hidroxiprolina tuvieron una correlación positiva significativa con la cortisona y el 5-nitrotiofeno-3-carbaldehído 3-(2-piridilo) hidrazona (Figura 3C, p < 0,05). Los niveles metabólicos de corticosterona y prostaglandina D2 se correlacionaron significativamente positivamente en modo iónico negativo (Figura 3D, p < 0,05).
3.4 El impacto de la GnRH en las vías metabólicas
Para explorar las funciones fisiológicas de los metabolitos afectados por el tratamiento con GnRH, se realizó un análisis de enriquecimiento de la vía KEGG en 53 metabolitos diferenciales en modos iónicos positivos y negativos. El análisis de enriquecimiento de la vía KEGG reveló 22 vías significativamente enriquecidas en los modos de iones positivos y 19 en los negativos. Estas vías pertenecían a cinco módulos: metabolismo, procesamiento de información genética, procesos celulares, procesamiento de información ambiental y sistemas de organismos. Las vías enriquecidas de metabolitos diferenciales en modo de iones positivos incluyeron principalmente la biosíntesis de pantotenato y CoA, la vía de señalización Foxo, la vía de señalización mTOR, la vía de señalización PI3K-Akt, la vía de señalización cGMP-PKG, la reabsorción de sodio regulada por aldosterona y la vía de señalización de AMPc (Figura 4A). La hidroxiprolina regulada significativamente a la baja se enriqueció en la vía del metabolismo de la arginina y la prolina (Figura 4B).
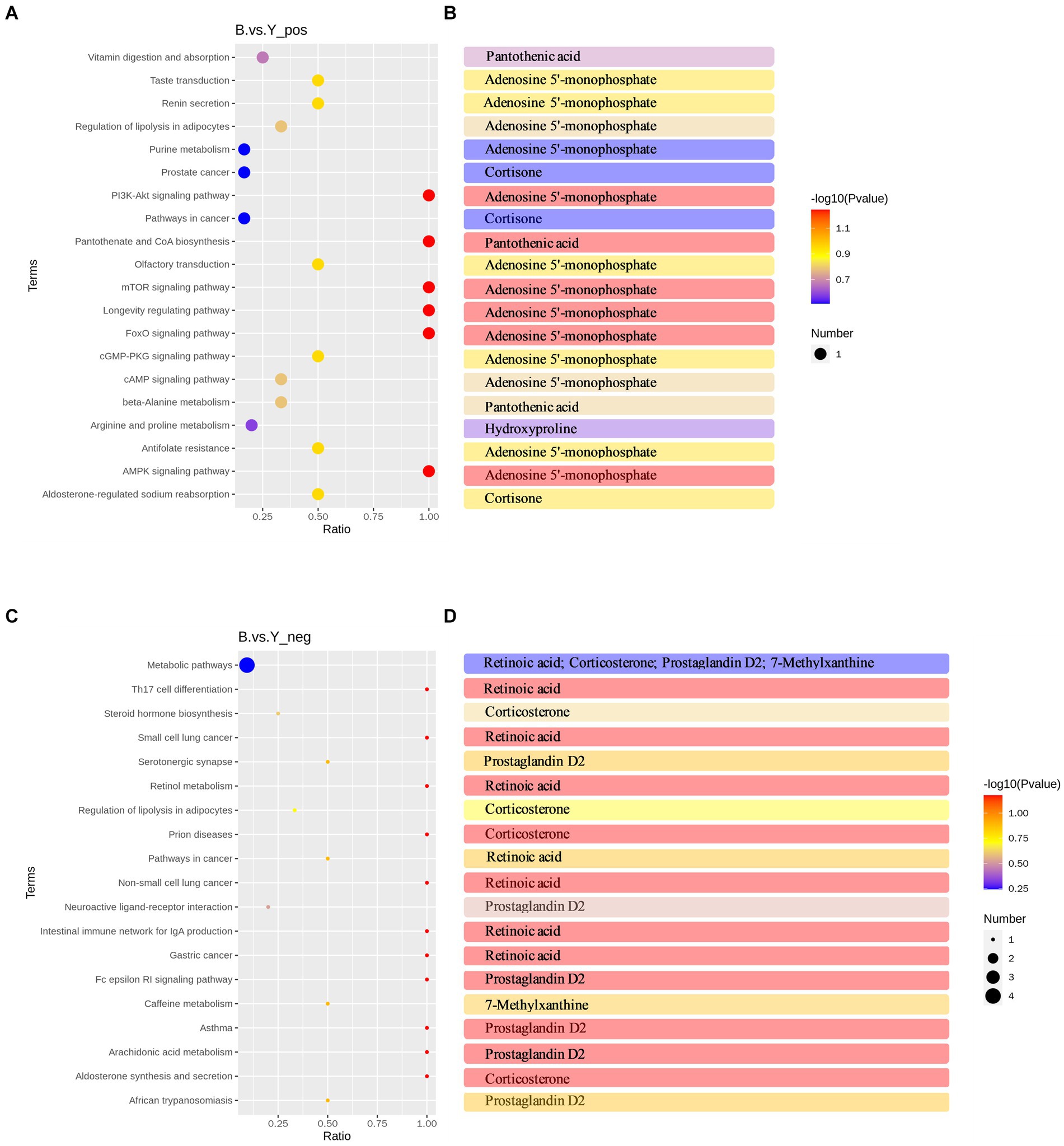
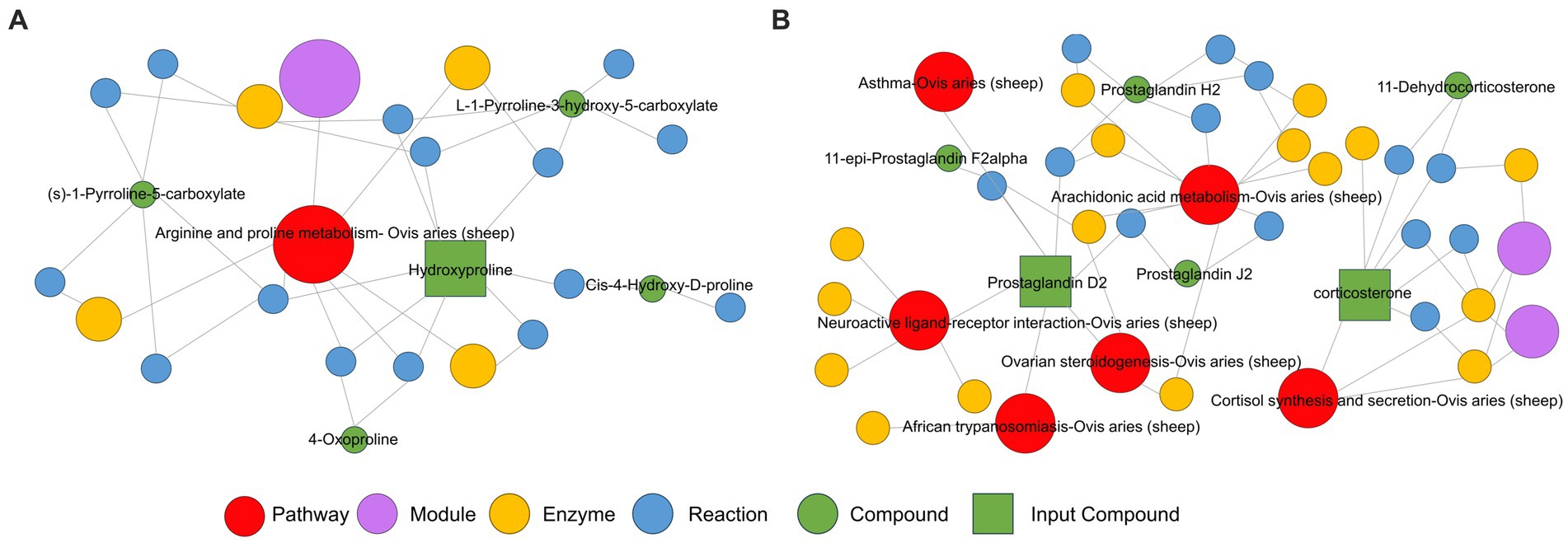
Por el contrario, en el modo de iones negativos, los metabolitos diferenciales se enriquecieron principalmente en la síntesis y secreción de aldosterona, el metabolismo del ácido araquidónico, la interacción ligando-receptor neuroactivo, la biosíntesis de hormonas esteroides y la diferenciación celular Th17, entre otras vías metabólicas (Figura 4C). La corticosterona significativamente regulada al alza se enriqueció principalmente en la vía de síntesis y secreción de aldosterona y en la vía de biosíntesis de hormonas esteroideas (Figura 4D). Además, hay un aumento significativo en el contenido metabólico de prostaglandina D2 en la vía del metabolismo del ácido araquidónico y la interacción de los ligandos neuroactivos con los receptores (Figura 4D). Además, se construyó una red reguladora KEGG utilizando los metabolitos diferenciales (Figuras 5A, B). La figura representa visualmente los cambios en metabolitos como la hidroxiprolina, la corticosterona y la prostaglandina D2. También muestra las intersecciones y los procesos biológicos relacionados con estos metabolitos y sus correspondientes vías.
4 Discusión
Las técnicas de sincronización e inseminación artificial se han utilizado ampliamente en la reproducción ovina, y el tratamiento con GnRH durante la inseminación se ha convertido en una práctica relativamente común en la industria ovina (26). Sin embargo, el impacto de la GnRH en el desarrollo embrionario y la implantación sigue siendo objeto de debate entre varios estudios. Nuestro estudio comparó las tasas de preñez a los 40 días entre las ovejas tratadas con GnRH y las de control. Los resultados indicaron que la tasa de embarazo en el grupo experimental (72,5%, n = 69) fue significativamente menor que la del grupo control (82,9%, n = 41) (p < 0,05).
Los resultados de embarazo obtenidos en nuestro grupo control son aceptables, lo que coincide con los resultados del tratamiento convencional de sincronización progesterona + eCG y la inseminación artificial (27). Estudios previos han demostrado que el tratamiento con GnRH 24-48 h después de la retirada de la progesterona en ovejas no mejora la tasa de preñez (11, 14). Las ovejas de Corriedale fueron tratadas con dos prostaglandinas para inducir la sincronización y FTAI con GnRH 24 h después de que la segunda prostaglandina utilizada disminuyera la tasa de preñez después de la inseminación artificial. Sin embargo, el tratamiento con GnRH 36 h después no tuvo un efecto significativo sobre la tasa de embarazo (14). Además, la administración de GnRH simultáneamente con la inseminación a tiempo único puede disminuir potencialmente la tasa de embarazo (28). Del mismo modo, el tratamiento con GnRH 36 h después de la retirada de la progesterona y la realización de la inseminación 48 h después no condujo a una mejora significativa en la tasa de preñez en cabras (6). En el proceso MOET, los receptores tratados con el análogo de GnRH Folligon también lograron una baja tasa de embarazo del 43,2% (29). Los resultados de estos estudios anteriores son consistentes con los resultados de este estudio.
Además, se realizó un análisis metabolómico para investigar las posibles razones detrás de la menor tasa de preñez observada en las ovejas tratadas con GnRH. La metabolómica emplea un análisis exhaustivo a través de los modos de iones positivos y negativos, con el objetivo de capturar los diversos metabolitos dentro del organismo. El modo de iones positivos tiende a detectar metabolitos alcalinos, mientras que el modo de iones negativos se centra en sustancias ácidas (30). En este estudio se llevó a cabo el cribado y la discusión de metabolitos por separado en dos modos. Significativamente, se analizaron metabolitos diferenciales, incluida la hidroxiprolina en modo iónico positivo y la corticosterona y la prostaglandina D2 en modo iónico negativo. La hidroxiprolina, un aminoácido distintivo que se encuentra en el colágeno, es un marcador de la degradación del colágeno (31, 32). Comprende aproximadamente el 13% del contenido total de aminoácidos del colágeno, por lo que es crucial para mantener la estructura y función del colágeno. Durante el embarazo, el contenido de colágeno uterino aumenta progresivamente, lo que conduce a un aumento correspondiente en los niveles de hidroxiprolina (33). En la actualidad, la matriz extracelular del útero y del tracto genital contiene abundante colágeno, esencial para soportar la creciente carga fetal y mantener la gestación (34). Los resultados de Metabonomics revelaron que el metabolito de la hidroxiprolina estaba significativamente regulado a la baja en la vía del metabolismo de la arginina y la prolina (p < 0,05) en ovejas Huyang tratadas con GnRH durante el período periimplantacional. Esta regulación a la baja puede disminuir la síntesis de colágeno, afectando la estabilidad y la resistencia mecánica del tejido endometrial y potencialmente causando daño o disfunción en el tejido uterino. Tales condiciones son desfavorables para establecer un ambiente adecuado para la implantación embrionaria. Además, los estudios han demostrado que el colágeno se degrada rápidamente después de que las vacas dan a luz, disminuyendo el tamaño y el peso uterino como parte del proceso de involución uterina (35). En conclusión, una reducción significativa en el nivel de hidroxiprolina durante el período periimplantacional es perjudicial para la implantación del embrión en el útero, lo que en última instancia disminuye la tasa de preñez en las ovejas Huyang.
Los resultados de este estudio indican que el metabolismo de la corticosterona en la vía de síntesis y secreción de aldosterona aumentó significativamente (p = 0,01) en ovejas Huyang tratadas con GnRH. La corticosterona, producida principalmente por la corteza suprarrenal, desempeña un papel crucial en la respuesta al estrés (36). Los estudios han demostrado que los niveles de corticosterona regulados al alza pueden inhibir el desarrollo embrionario y disminuir el número de células de masa celular interna, comprometiendo la calidad del embrión y afectando las tasas de embarazo (37). Zheng et al. utilizan continuamente corticosterona exógena en ratones durante los días 1 a 4 de gestación. En comparación con el grupo control, el uso de corticosterona resultó en una fuerte disminución de Stat3 fosforilado, afectando la receptividad endometrial y reduciendo significativamente el número de sitios de implantación en el día 5 (38). Del mismo modo, los estudios en un modelo de ratón de exceso de corticosterona han demostrado que los niveles elevados de corticosterona afectan a la fertilidad femenina al influir en el útero en lugar de en los ovocitos (39). Estos hallazgos son consistentes con nuestros resultados. Por lo tanto, pensamos que el tratamiento con GnRH durante la inseminación aumenta significativamente los niveles de corticosterona en las ovejas Huyang antes de la implantación, lo que afecta el entorno uterino antes de la implantación, interrumpe la receptividad endometrial y, en última instancia, disminuye la tasa de embarazo.
La prostaglandina D2 (PGD2) pertenece a la familia de las prostaglandinas y se sintetiza a partir del ácido araquidónico (AA) a través de la acción de la ciclooxigenasa (COX) (40). Los análisis metabolómicos indicaron un aumento significativo de la prostaglandina D2 dentro de la vía del metabolismo del ácido araquidónico (p = 0,017). Aunque el PGD2 promueve la regresión lútea, su efecto es menos pronunciado que el de la prostaglandina F2 alfa (PGF2α). No obstante, PGF2α y PGD2 exhiben efectos sinérgicos en la promoción de la regresión lútea mediante la expansión efectiva de los vasos sanguíneos y el aumento de la entrada al tejido lúteo, lo que facilita la afluencia de PGF2α adicional sintetizada en el endometrio y acelera el proceso de regresión lútea (41). Además, los PG sirven como mediadores clave en el estudio de la inflamación (42). Bajo condiciones inflamatorias u otros estímulos, la expresión de COX-2 se regula significativamente al alza, promoviendo aún más la síntesis de PG en un grado notable (43). La inducción de inflamación intrauterina puede tener un impacto negativo en la implantación y el desarrollo de embriones dentro del útero (44). Los estudios han revelado la presencia de PGD2 en los tejidos del endometrio humano y del músculo liso uterino, donde destaca como la sustancia más potente dentro de la familia de los PG para aumentar el flujo sanguíneo al endometrio y a los músculos uterinos (45). La elevación significativa de PGD2 puede desencadenar y acelerar la frecuencia de contracción del músculo liso uterino, disminuyendo en última instancia la tasa de supervivencia embrionaria (46). En consecuencia, el desequilibrio de los niveles de PGD2 en las ovejas afecta negativamente a la implantación embrionaria, lo que resulta en una disminución de la tasa de preñez en el grupo experimental.
5 Conclusión
En resumen, la administración de una dosis única del agonista de la GnRH triptorelina 48 h después de la retirada de la esponja reduce la tasa de preñez en ovejas Huyang después de la inseminación. El análisis metabolómico reveló una disminución significativa en los niveles de hidroxiprolina y un aumento significativo en los niveles de corticosterona y prostaglandina D2, que están relacionados con una disminución de la tasa de preñez en las ovejas Huyang.
Declaración de disponibilidad de datos
Los datos presentados en el estudio están depositados en el repositorio MetaboLights, número de acceso MTBLS10318.
Declaración ética
El estudio en animales fue aprobado por el Comité de Cuidado y Uso de Animales de la Universidad Agrícola de Hebei. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
JZ: Curación de datos, Análisis formal, Investigación, Metodología, Administración de proyectos, Software, Validación, Redacción – borrador original, Redacción – revisión y edición. SS: Curación de datos, Análisis formal, Investigación, Metodología, Recursos, Redacción – borrador original. XB: Curación de datos, Análisis formal, Investigación, Metodología, Visualización, Redacción – borrador original. NY: Curación de datos, metodología, recursos, redacción, revisión y edición. YL: Curación de datos, metodología, recursos, redacción, revisión y edición. XW: Conceptualización, Análisis formal, Obtención de fondos, Metodología, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. XL: Conceptualización, Curación de datos, Obtención de fondos, Metodología, Recursos, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declaran/n que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio contó con el apoyo del Programa Clave de Investigación y Desarrollo de la Prefectura Autónoma de Ili Kazak (YZ2023A04) y el Fondo Especial de Talentos de la Universidad Agrícola de Hebei (YJ201952).
Reconocimientos
Los autores agradecen al personal de Hulunbeir Longxingchuan Animal Husbandry Technology Co., Ltd. por su ayuda durante este estudio. Los autores agradecen a Novogene (Novogene Co., Ltd., Pekín, China) por sus servicios de análisis metabolómico.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Gibbons, AE, Fernandez, J, Bruno-Galarraga, MM, Spinelli, MV y Cueto, MI. Recomendaciones técnicas para la inseminación artificial en ovejas. Anim Reprod. (2019) 16:803–9. doi: 10.21451/1984-3143-AR2018-0129
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Menchaca, A, y Rubianes, E. Nuevos tratamientos asociados a la inseminación artificial temporizada en pequeños rumiantes. Reprod Fertil Dev. (2004) 16:403–13. doi: 10.1071/RD04037
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Titi, HH, Kridli, RT y Alnimer, MA. Sincronización del estro en ovejas y cabras mediante combinaciones de GnRH, progestágeno y prostaglandina F2alfa. Reprod Domest Anim. (2010) 45:594–9. doi: 10.1111/j.1439-0531.2008.01309.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Ciornei, ŞG, Drugociu, D, Ciornei, L, y Roşca, P. Respuesta ovárica al tratamiento con P4-PGF-FSH en ovejas de Suffolk y sincronización de P4-PGF-PMSG en ovejas cruzadas, para el protocolo IVD y ET. Vet Med Sci. (2022) 8:726–34. doi: 10.1002/vms3.705
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Gonźalez-Bulnes, A, Baird, DT, Campbell, BK, Cocero, MJ, García-García, RM, Inskeep, EK, et al. Múltiples factores que afectan la eficiencia de la ovulación múltiple y la transferencia embrionaria en ovejas y cabras. Reprod Fertil Dev. (2004) 16:421–35. doi: 10.1071/RD04033
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Reyna, J, Thomson, PC, Evans, G, y Maxwell, WM. Sincronía de la ovulación y dinámica folicular en ovejas merinas tratadas con GnRH en las temporadas de cría y no reproducción. Reprod Domest Anim. (2007) 42:410–7. doi: 10.1111/j.1439-0531.2006.00800.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Balaro, MF, Fonseca, JF, Barbosa, TG, Souza-Fabjan, JM, Figueira, LM, Teixeira, TA, et al. Papel potencial de la GnRH en la sincronización de la emergencia folicular antes del protocolo de día 0 superovulatorio. Domest Anim Endocrinol. (2016) 54:10–4. doi: 10.1016/j.domaniend.2015.07.002
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Hameed, N, Khan, MI, Zubair, M, y Andrabi, SMH. Enfoques de la sincronización estral en ovejas: desarrollos durante las últimas dos décadas: una revisión. Trop Anim Salud Prod. (2021) 53:485. doi: 10.1007/s11250-021-02932-8
9. Berean, D, Bogdan, LM, Opris, P, y Cimpean, R. Implicaciones económicas y el impacto de la administración de la hormona liberadora de gonadotropina en el momento de la inseminación artificial en vacas criadas en el sistema extensivo en el norte de Rumanía. Front Vet Sci. (2023) 10:1167387. doi: 10.3389/fvets.2023.1167387
10. Biehl, MV, Ferraz Junior, MVC, Ferreira, EM, Polizel, DM, Miszura, AA, Barroso, JPR, et al. Efecto de los métodos reproductivos y la administración de GnRH en el protocolo a largo plazo en ovejas Santa Inés. Trop Anim Salud Prod. (2017) 49:1303–8. doi: 10.1007/s11250-017-1329-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Kutlu, M, y Dinç, DA. El efecto de las inyecciones de dosis dobles de GnRH sobre los parámetros de rendimiento reproductivo después de la administración de progestágenos a corto plazo en ovejas Awassi lactadas durante la temporada no reproductiva. Trop Anim Salud Prod. (2021) 53:277. doi: 10.1007/s11250-021-02735-x
12. Olivera-Muzante, J, Fierro, S, López, V, y Gil, J. Comparación de protocolos basados en prostaglandinas y progesterona para la inseminación artificial programada en ovejas. Teriogenología. (2011) 75:1232–8. doi: 10.1016/j.theriogenology.2010.11.036
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Ayaseh, M, Mirzaei, A, Boostani, A, y Mehrvarz, M. El efecto de la inyección de prostaglandinas y gonadotrofinas (GnRH y hCG) combinado con el efecto carnero sobre las concentraciones de progesterona y el rendimiento reproductivo de las ovejas karakul durante la temporada no reproductiva. Vet Med Sci. (2021) 7:148–55. doi: 10.1002/vms3.353
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Olivera-Muzante, J, Gil, J, Viñoles, C, y Fierro, S. Resultado reproductivo con inclusión de GnRH a las 24 o 36 h siguiendo un protocolo basado en prostaglandina F2α para IA temporizada en ovejas. Anim Reprod Sci. (2013) 138:175–9. doi: 10.1016/j.anireprosci.2013.02.013
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Perry, GA, Smith, MF, Lucy, MC, Green, JA, Parks, TE, MacNeil, MD, et al. Relación entre el tamaño del folículo en el momento de la inseminación y el éxito del embarazo. Proc Natl Acad Sci USA. (2005) 102:5268–73. doi: 10.1073/pnas.0501700102
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Moorey, SE, Monnig, JM, Smith, MF, Ortega, MS, Green, JA, Pohler, KG, et al. Perfiles de transcripción diferenciales en complejos cúmulo-ovocito originados en folículos preovulatorios de madurez fisiológica variada en vacas de carne. Genes (Basilea). (2021) 12:893. doi: 10.3390/genes12060893
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Fernández, J, Bruno-Galarraga, MM, Soto, AT, de la Sota, RL, Cueto, MI, Lacau-Mengido, IM, et al. Efecto de la administración de GnRH o hCG en el día 4 post inseminación sobre el rendimiento reproductivo en ovejas merinas de la Patagonia Norte. Teriogenología. (2019) 126:63–7. doi: 10.1016/j.theriogenology.2018.12.008
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Fiehn, O. Metabolómica: el vínculo entre genotipos y fenotipos. Planta Mol Biol. (2002) 48:155–71. doi: 10.1023/A:1013713905833
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Hong, J, Yang, L, Zhang, D y Shi, J. Metabolómica de plantas: una herramienta indispensable de biología de sistemas para la ciencia de las plantas. Int J Mol Sci. (2016) 17:767. doi: 10.3390/ijms17060767
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Wishart, DS. Aplicaciones emergentes de la metabolómica en el descubrimiento de fármacos y la medicina de precisión. Nat Rev Droga Discov. (2016) 15:473–84. doi: 10.1038/nrd.2016.32
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Johnson, CH, Ivanisevic, J y Siuzdak, G. Metabolómica: más allá de los biomarcadores y hacia los mecanismos. Nat Rev Mol Cell Biol. (2016) 17:451–9. doi: 10.1038/nrm.2016.25
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Zhang, X, Han, L, Hou, S, Raza, SHA, Gui, L, Sun, S, et al. El enfoque metabolómico revela que la dieta alta en energía mejora la calidad y realza el sabor de la carne de oveja tibetana negra al alterar la composición de la microbiota ruminal. Nutr delantero. (2022) 9:915558. doi: 10.3389/fnut.2022.915558
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Zhang, Z, Shahzad, K, Shen, S, Dai, R, Lu, Y, Lu, Z, et al. La alteración de los niveles de proteína soluble en la dieta con la disminución de la proteína bruta puede ser una estrategia potencial para mejorar la eficiencia del nitrógeno en ovejas Hu basada en el microbioma ruminal y la metabolómica. Nutr delantero. (2022) 8:815358. doi: 10.3389/fnut.2021.815358
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. de Sousa, EB, Dos Santos, GC Jr, Duarte, MEL, Moura, VN, y Aguiar, DP. La metabolómica como herramienta prometedora para el diagnóstico precoz de la artrosis. Braz J Med Biol Res. (2017) 50:e6485. doi: 10.1590/1414-431×20176485
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Abril-Parreño, L, Krogenæs, A, y Fair, S. El perfil lipidómico del moco cervical revela el papel potencial de los metabolitos derivados proinflamatorios en el transporte de espermatozoides a través del cuello uterino ovino. Animal. (2024) 18:101136. doi: 10.1016/j.animal.2024.101136
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Türk, G, Gür, S, Sönmez, M, Bozkurt, T, Aksu, EH y Aksoy, H. Efecto de la GnRH exógena en el momento de la inseminación artificial sobre el rendimiento reproductivo de ovejas Awassi sincronizadas con la combinación progestágeno-PMSG-PGF2alfa. Reprod Domest Anim. (2008) 43:308–13. doi: 10.1111/j.1439-0531.2007.00896.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Berean, D, Ergene, O, Blaga-Petrean, A, Bogdan, I, Ciupe, S, Cenariu, M, et al. Datos comparativos sobre la inducción del celo y la tasa de preñez en ovejas lacaunes en temporada no reproductiva después de implantes de melatonina y progestágeno intravaginal. Ind J Anim Res. (2021) 55:517–21. doi: 10.18805/ijar. B-1282
28. Yilmazbas-Mecitoglu, G, Karakaya, E, Keskin, A, Alkan, A, y Gumen, A. La reducción de la duración entre el tratamiento con la hormona liberadora de gonadotropina (GnRH) y la prostaglandina F₂α en el protocolo Ovsynch a 6 días mejoró la ovulación al segundo tratamiento con GnRH, pero se inclinó a reducir la fertilidad. J Dairy Sci. (2013) 96:3817–24. doi: 10.3168/jds.2012-6496
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Aybazov, M, Selionova, M, Trukhachev, V, Malorodov, V, Yuldashbayev, Y, y Easa, AA. Producción y trasplante embrionario en temporada no reproductiva de razas ovinas de carne mediante la estimulación de la superovulación con diferentes preparaciones de hormonas folículo-estimulantes. Reprod Domest Anim. (2023) 58:230–7. doi: 10.1111/rda.14279
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Jeppesen, MJ y Powers, R. Metabolómica no dirigida multiplataforma. Magn Reson Chem. (2023) 61:628–53. doi: 10.1002/mrc.5350
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Ghartey, J, Anglim, L, Romero, J, Brown, A, y Elovitz, MA. Las mujeres con parto prematuro sintomático tienen un metaboloma cervicovaginal distinto. Am J Perinatol. (2017) 34:1078–83. doi: 10.1055/s-0037-1603817
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Woessner, JF Jr. Catabolismo del colágeno y la proteína no colágena en el útero de rata durante la involución posparto. Biochem J. (1962) 83:304–14. doi: 10.1042/bj0830304
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Shimizu, K, y Maekawa, K. Degradación del colágeno en el útero de ratón durante la involución posparto: vía extracelular. Acta Anat (Basilea). (1983) 117:257–60. doi: 10.1159/000145795
34. Belstra, BA, Flowers, WL, Croom, WJ, Degroot, J, y See, MT. Excreción urinaria de marcadores de degradación de colágeno por cerdas durante la involución uterina posparto. Anim Reprod Sci. (2005) 85:131–45. doi: 10.1016/j.anireprosci.2004.04.035
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Rizzo, A, Gazza, C, Silvestre, A, Maresca, L, y Sciorsci, RL. Escopolamina para la involución uterina de vacas lecheras. Teriogenología. (2018) 122:35–40. doi: 10.1016/j.theriogenology.2018.08.025
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Austin, SH, Harris, RM, Booth, AM, Lang, AS, Farrar, VS, Krause, JS, et al. Aislar el papel de la corticosterona en la respuesta al estrés transcriptómico hipotálamo-hipofisario-gonadal. Endocrinol frontal. (2021) 12:632060. doi: 10.3389/fendo.2021.632060
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Zheng, HT, Fu, T, Zhang, HY, Yang, ZS, Zheng, ZH y Yang, ZM. Hsd11b2 regulada por progesterona como barrera para equilibrar la corticosterona uterina de ratón. J Endocrinol. (2020) 244:177–87. doi: 10.1530/JOE-19-0349
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Zheng, HT, Zhang, HY, Chen, ST, Li, MY, Fu, T y Yang, ZM. Los efectos perjudiciales de la exposición a glucocorticoides inducida por el estrés sobre la receptividad uterina y la decidualización de ratones. FASEB J. (2020) 34:14200–16. doi: 10.1096/fj.201902911RR
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Diao, H, Aplin, JD, Xiao, S, Chun, J, Li, Z, Chen, S, et al. Expresión espacio-temporal alterada de los tipos de colágeno I, III, IV y VI en útero de ratón periimplantacional deficiente en Lpar3. Biol Reprod. (2011) 84:255–65. doi: 10.1095/biolreprod.110.086942
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Yue, L, Durand, M, Lebeau, Jacob, MC, Hogan, P, McManus, S, Roux, S, et al. La prostaglandina D2 induce la apoptosis de los osteoclastos humanos mediante la activación del receptor CRTH2 y la vía de apoptosis intrínseca. Hueso. (2012) 51:338–46. doi: 10.1016/j.bone.2012.06.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Mori, A, Saito, M, Sakamoto, K, Nakahara, T e Ishii, K. Las prostaglandinas vasodilatadoras administradas por vía intravenosa aumentan el flujo sanguíneo de la retina y la coroides en ratas. J Pharmacol Sci. (2007) 103:103–12. doi: 10.1254/jphs. FP0061061
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Malhotra, S, Deshmukh, SS y Dastidar, SG. Inhibidores de la COX para la inflamación de las vías respiratorias. Objetivos de Opin Ther expertos. (2012) 16:195–207. doi: 10.1517/14728222.2012.661416
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Burdan, F, Chałas, A, y Szumiło, J. Cyklooksygenaza i prostanoidy–znaczenie biologiczne [Ciclooxigenasa y prostanoides–implicaciones biológicas]. Postepy Hig Med Dosw. (2006) 60:129–41.
44. Robertson, S.A., Care, A.S., y Moldenhauer, L.L. Células T reguladoras en la implantación embrionaria y la respuesta inmune al embarazo. J Clin Invest. (2018) 128:4224–35. doi: 10.1172/JCI122182
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Schramm, W, Einer-Jensen, N, Brown, MB y McCracken, JA. Efecto de cuatro prostaglandinas primarias y relaxina sobre el flujo sanguíneo en el endometrio y miometrio ovino. Biol Reprod. (1984) 30:523–31. doi: 10.1095/biolreprod30.3.523
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Michimata, T, Tsuda, H, Sakai, M, Fujimura, M, Nagata, K, Nakamura, M, et al. Acumulación de células T helper 2 y T-citotóxicas 2 positivas para CRTH2 en sitios de implantación de decidua humana de manera mediada por prostaglandina D(2). Mol Hum Reprod. (2002) 8:181–7. doi: 10.1093/molehr/8.2.181
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: hormona liberadora de gonadotropina, tasa de preñez, metabolómica, hidroxiprolina, prostaglandina D2, corticosterona, ovejas
Cita: Zhang J, Sun S, Bai X, Yang N, Liu Y, Wu X y Li X (2024) Análisis metabolómico del efecto de la GnRH en la tasa de preñez de ovejas con esquema de sincronización de celo basado en progesterona. Frente. Vet. Sci. 11:1442931. doi: 10.3389/fvets.2024.1442931
Editado por:
Stefan Gregore Ciornei, Universidad de Ciencias de la Vida de Iasi (IULS), Rumanía
Revisado por:
Daniel Berean, Universidad de Ciencias Agrarias y Medicina Veterinaria de Cluj-Napoca, Rumanía
Laura Abril-Parreño, Universidad de Murcia, España
Derechos de autor © 2024 Zhang, Sun, Bai, Yang, Liu, Wu y Li. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Xinglong Wu, wuxl32@hebau.edu.cn; Xiangyun Li, Lxyun@hebau.edu.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía