
Cambios en el rendimiento del crecimiento, la bioquímica sérica, la fermentación ruminal, la comunidad de la microbiota ruminal en cabras
Cambios en el rendimiento del crecimiento, la bioquímica sérica, la fermentación ruminal, la comunidad de la microbiota ruminal y el desarrollo intestinal en cabras destetadas durante el tratamiento con metionina protegida del rumen
 Youli Wang1,2,3
Youli Wang1,2,3 Wei Liu1,2,3
Wei Liu1,2,3 An Li1,2,3Wuqie Qubi1,2,3Chengsi Gong1,2,3Xuening Li1,2,3Jiani Xing1,2,3Haitao Shi1,2,3
An Li1,2,3Wuqie Qubi1,2,3Chengsi Gong1,2,3Xuening Li1,2,3Jiani Xing1,2,3Haitao Shi1,2,3 Yanyan Li1,2,3Yong Wang1,2,3
Yanyan Li1,2,3Yong Wang1,2,3 Yaqiu Lin1,2,3*
Yaqiu Lin1,2,3*- 1Facultad de Ciencia Animal y Veterinaria, Universidad del Suroeste de Minzu, Chengdu, China
- número arábigoLaboratorio Clave de Reserva y Utilización de Recursos Genéticos Animales de la Meseta Qinghai-Tibetana, Ministerio de Educación, Universidad del Sudoeste de Minzu, Chengdu, China
- 3Laboratorio clave de la provincia de Sichuan para la reserva y explotación de recursos genéticos animales de la meseta Qinghai-Tibetana, Universidad del Sudoeste de Minzu, Chengdu, China
La metionina (RPM) protegida contra el rumen, como la metionina (CM) recubierta y el éster isopropílico del ácido 2-hidroxi-4-(metiltio)-butanoico (HMBi), se utilizaba generalmente en las vacas lecheras, pero aún no está claro cómo afecta el RPM a las cabras de carne. En este estudio, treinta cabras machos Jianzhou Da’er destetados fueron asignados aleatoriamente a uno de tres tratamientos: alimentados con dieta basal o dieta basal suplementada con 0,12% de CM o 0,22% de HMBi, con el objetivo de examinar su impacto en el rendimiento del crecimiento, la bioquímica sérica, la fermentación ruminal, la microbiota ruminal y el desarrollo intestinal en cabras de carne. Los resultados indican que la suplementación con HMBi condujo a un aumento en el peso corporal, la ingesta de alimento y la relación alimento-ganancia, mientras que la MC solo resultó en un aumento en la ingesta de alimento (todos p < 0,05). Tanto la MC como la HMBi dieron lugar a un aumento del colesterol total (CT) sérico, el nitrógeno ureico en sangre (BUN), la fosfatasa alcalina (ALP) y la aspartato aminotransferasa (AST), aunque con una disminución de los triglicéridos séricos (TG) y del ácido β-hidroxibutírico (BHB, todos p < 0,05). Tanto la suplementación con CM como con HMBi disminuyeron la concentración ruminal de ácido butírico (ambos p < 0,05). La secuenciación del ARNr 16S mostró que la suplementación con HMBi aumentó significativamente la abundancia total de Bacteroidetes y Firmicutes. Los suplementos de CM y HMBi aumentaron la abundancia de Rikenella y Proteiniphilum, pero disminuyeron la abundancia de Eisenbergiella, Enterocloster, Massilioclostridium, Eubacterium, Angelakisella, Blastopirellula, Christensenella y Pseudoruminococcus. La suplementación con MC aumentó específicamente la abundancia de Desulfobulbus, Sodaliphilus y Coprococcus, al tiempo que disminuyó la prevalencia de Anaerocella, Mogibacterium y Collinsella. La suplementación con HMBi aumentó significativamente la abundancia de Paraprevotella, Bacilliculturomica, Lachnoclostridium, Dysosmobacter, Barnesiella y Paludibacter, mientras que disminuyó la abundancia de Butyrivibrio y Pirellula. Además, la administración de suplementos de CM y HMBi dio lugar a un aumento de las bacterias productoras de amoníaco y reductoras de sulfato, mientras que se observó una disminución de las bacterias oxidantes de amoníaco, asociadas a la salud y asociadas a la enfermedad. El análisis correlacional reveló que TG y BHB tenían una correlación positiva con las bacterias asociadas a enfermedades y oxidantes de amoníaco, mientras que tenían una correlación negativa con las bacterias productoras de amoníaco. El BUN, la ALP y la AST séricos se correlacionaron positivamente con las bacterias productoras de amoníaco, pero se correlacionaron negativamente con las bacterias oxidantes de amoníaco. Además, tanto la suplementación con CM como con HMBi mejora el desarrollo del intestino delgado, y el HMBi tiene un mejor efecto. En resumen, este estudio indica que tanto la suplementación con CM como con HMBi mejora el metabolismo de los lípidos, la utilización del nitrógeno y el desarrollo intestinal. El efecto de promoción del crecimiento de la suplementación con HMBi puede atribuirse a la mayor abundancia de bacterias productoras de ácidos grasos volátiles y que utilizan nitrógeno y a un mejor desarrollo intestinal.
1 Introducción
La metionina es un aminoácido limitado para los rumiantes que puede ser degradado por los microbios del rumen. Con el fin de evitar la degradación del microbio ruminal, se ha desarrollado bien la metionina (RPM) protegida del rumen, como la metionina (CM) recubierta y el éster isopropílico del ácido 2-hidroxi-4-(metiltio)-butanoico (HMBi). La MC puede evitar en gran medida ser degradada por el microbio ruminal y luego ser absorbida por el intestino posterior. El HMBi ingerido puede ser absorbido por la pared ruminal y el intestino posterior o degradado por el microbio ruminal. El HMBi puede transportarse como sangre a casi todos los tejidos y órganos del cuerpo, como el rumen, el omaso, el riñón y el hígado, y luego convertirse en metionina y acetona (1, 2). Aunque las RPM tienen una mayor eficiencia en el suministro de metionina (2), una parte de ellas todavía sufren degradación en el rumen, un órgano digestivo crucial en los rumiantes. Esto puede influir en la fermentación ruminal y en los microbios ruminales. Además, el intestino delgado es un sitio importante para absorber las proteínas microbianas que se producen en el rumen (3). Un estudio ha demostrado que la metionina beneficia el desarrollo de organoides intestinales (4). Sin embargo, los efectos de la suplementación con RPM sobre la fermentación ruminal, las poblaciones microbianas del rumen y el desarrollo intestinal siguen sin estar claros.
En la actualidad, las RPM han sido ampliamente estudiadas en el ganado lechero y se ha demostrado que tienen un impacto positivo sobre la grasa y la proteína de la leche (5). Sin embargo, se han realizado pocos estudios sobre rumiantes cárnicos. En bovinos, Qin et al. (6) informaron que una dieta suplementada con HMBi en aproximadamente un 0,3% aumentó el peso corporal en el ganado de engorde. Sin embargo, la suplementación con CM no tuvo impacto en el rendimiento del crecimiento en los toros Fleckvieh alimentados con CM (7). Hasta ahora, el período de alimentación de los estudios de RPM en cabras y ovejas ha variado de 30 a 80 días, el nivel de HMBi ha variado de 0,05 a 0,2% MS y el nivel de CM ha sido de 0,1 ~ 1,15% (8-15). Sin embargo, no se observó mejoría en las cabras. El período de alimentación puede tener un impacto en la eficacia de las RPM. Por lo tanto, es de mayor importancia investigar las alteraciones en la fermentación ruminal, los microbios ruminales, la bioquímica sérica y el desarrollo intestinal cuando aparece un mejor rendimiento del crecimiento durante las RPM de la alimentación.
La cabra Jianzhou Da’er es una nueva raza formada por el cruce de cabras nubias con cabras autóctonas de la ciudad de Jianyang, provincia de Sichuan, en China. La carne es conocida por su textura tierna y su ligero olor, que ha ganado cada vez más atención por parte de los consumidores. Sin embargo, hay información limitada sobre las cabras Jianzhou Da’er. El objetivo de este estudio fue investigar los efectos de diferentes fuentes de RPM sobre el rendimiento del crecimiento, la bioquímica sérica, los parámetros de fermentación del rumen, los microbios del rumen y el desarrollo intestinal de cabras Jianzhou Da’er destetadas cuando apareció un mejor rendimiento de crecimiento.
2 Materiales y métodos
Este estudio se llevó a cabo en Sichuan Tiandi Goat Biological Engineering Co., Ltd. (Chengdu, China). La aprobación ética para el ensayo en animales en este estudio fue otorgada por el Comité de Ética y Cuidado Animal de la Universidad del Suroeste de Minzu (código de aprobación: SMU-202401047).
2.1 Diseño experimental y dieta
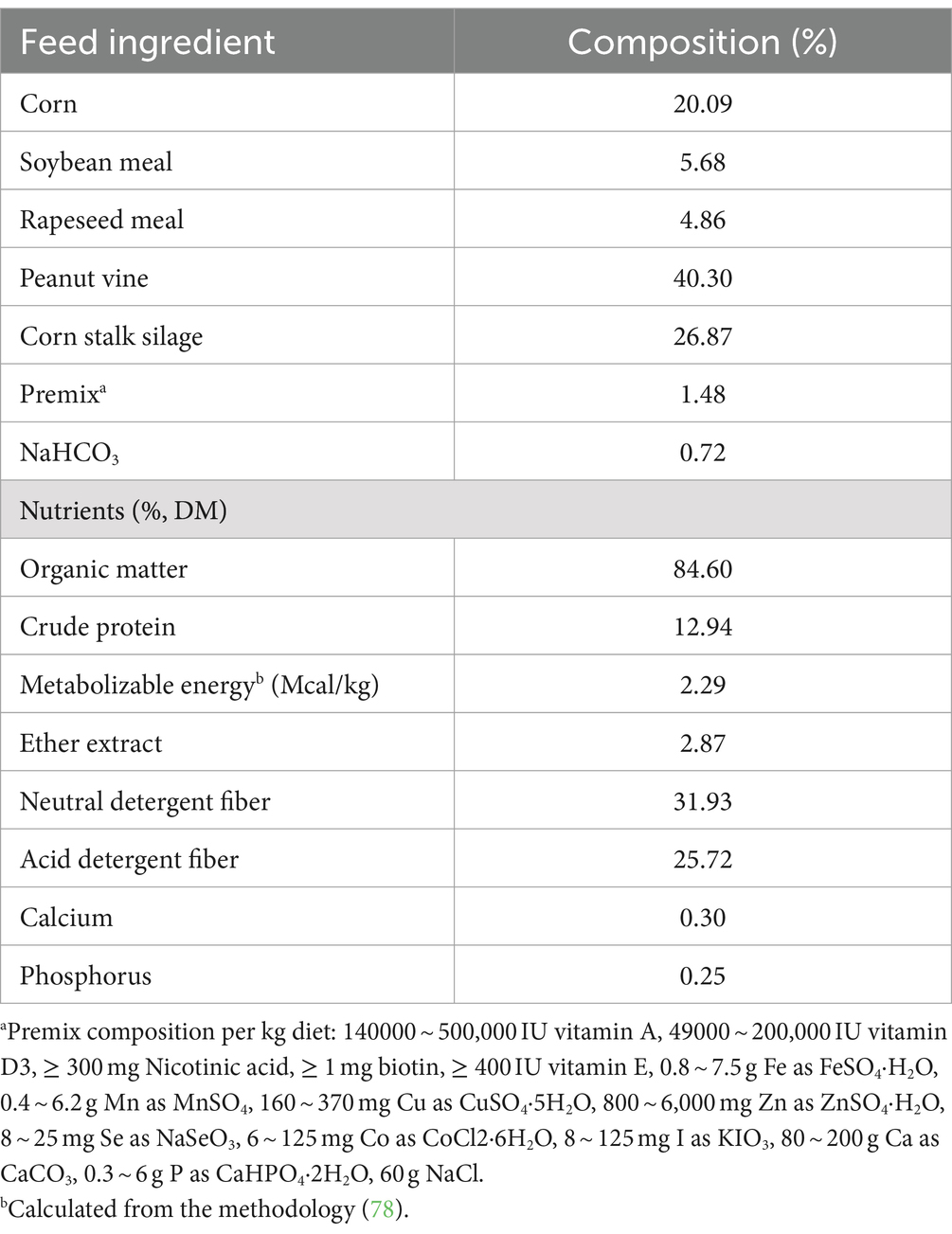
Un total de treinta cabras machos Jianzhou Da’er destetados (de aproximadamente 2,5 meses de edad) con peso corporal similar (15,7 ± 0,19 kg) fueron asignados aleatoriamente a uno de tres tratamientos: alimentados con dieta básica (TC), materia seca dietética (MS) con 0,12% de CM (DL-metionina disponible ≥85%; Mepron, Evonik Industries, Frankfort, Alemania), o MS dietética con 0,22% de HMBi (contenido disponible ≥46%; Metasmart, Adisseo, París, Francia). Cada tratamiento contenía 10 cabras, cada una de las cuales fue alojada individualmente y se le proporcionó acceso gratuito a agua dulce. El nivel de suplementación con metionina recubierta se basa en los hallazgos de un estudio previo (16). Se determinó que la inclusión de CM y HMBi proporciona un 0,1% adicional de metionina metabolizable en la DM dietética. La dieta se formuló de acuerdo con las recomendaciones del Consejo Nacional de Investigación (17), y la formulación y los nutrientes se muestran en la Tabla 1. La alimentación se proporcionó tres veces al día (08:00, 14:00 y 18:00). La alimentación de concentración se proporcionó a las 18:00 con un RPM suplementado. El experimento duró 4,5 meses, y los primeros 7 días se utilizaron para la preadaptación. El peso corporal se registró mensualmente. La ingesta de alimento se registró todos los días.
2.2 Sistemas de toma de muestras
Después de la alimentación durante 4,5 meses, se recogieron muestras de sangre utilizando un tubo de extracción de sangre al vacío después de ayunar durante 12 h. En cada tratamiento, el líquido ruminal de 10 cabras fue filtrado por 4 capas de gasa esterilizada. Una parte del fluido ruminal se congeló con nitrógeno líquido y se almacenó a -80 °C con el fin de determinar los AGV y la microbiota ruminal. Se utilizó una porción del líquido ruminal para determinar el pH.
2.3 Análisis bioquímico sérico
La sangre se colocó a temperatura ambiente durante 4 h, seguida de centrifugación a 3000 g durante 10 min a 4°C. Se detectaron colesterol total sérico (CT), triglicéridos (TG), nitrógeno ureico en sangre (BUN), ácido β-hidroxibutírico (BHB), proteína total (TP), albúmina (ALB), globulina (GLB), fosfatasa alcalina (ALP), alanina transaminasa (ALT), aspartato aminotransferasa (AST) y deshidrogenasa láctica (LDH) de acuerdo con las instrucciones proporcionadas por los kits comerciales (Mindray, Shenzhen, China).
2.4 Parámetros de fermentación ruminal
El líquido ruminal recién recolectado se examinó para determinar el pH utilizando un medidor de pH portátil (Testo 206 pH2, Schwarzwald, Alemania). Se utilizó fluido Remen almacenado a -80 °C para analizar la concentración de AGV mediante un cromatógrafo de gases (Agilent Technologies 7890B, California, Estados Unidos). La columna utilizada fue una columna capilar HP-INNOWax (30 m de longitud, 0,32 mm de diámetro y 0,5 μm de espesor de película; Agilent, California, Estados Unidos). La temperatura del horno de columna fue de 120 °C, el tiempo de equilibrio fue de 3 minutos y luego aumentó 10 °C cada 10 minutos para estabilizarse a 180 °C. La temperatura del puerto de inyección frontal era de 220 °C y la del detector frontal de 250 °C. El patrón interno fue ácido 2-etilbutanoico. Cada muestra de líquido ruminal se examinó dos veces.
2.5 Secuenciación de ARNr 16S
Para cada tratamiento, se utilizaron 10 muestras en cada tratamiento para la secuenciación del ARNr 16S. Según lo descrito por He et al. (17, 18), el ADN microbiano de muestras de fluido ruminal fue extraído por un kit de ADN E.Z.N.A. ® (Norcross, Georgia, Estados Unidos) siguiendo las instrucciones del fabricante. Los genes de ARN ribosómico 16S bacteriano se amplificaron a través de la reacción en cadena de la polimerasa (PCR). Todas las secuenciaciones de amplicones fueron realizadas por Shanghai Biozeron Biotechnology Co., Ltd. (Shanghai, China). Las lecturas en bruto de PacBio se procesaron utilizando el software SMRT Link Analysis versión 9.0 para obtener lecturas de secuencia de consenso circular demultiplexadas con los siguientes ajustes: número mínimo de pasadas = 3, precisión mínima predicha = 0,99. Las unidades taxonómicas operacionales (OTUs) se agruparon con un corte de similitud del 98,65% utilizando UPARSE (versión 7.1), y las secuencias quiméricas se identificaron y eliminaron utilizando UCHIME.
2.6 Tinción de hematoxilina-eosina
Después de 48 h de fijación con paraformaldehído al 4%, los íleon y yeyuno se deshidrataron con concentraciones crecientes de etanol (75, 85, 95 y 100%), se limpiaron con xileno y se incluyeron en parafina. A continuación, se tiñeron secciones de parafina del intestino delgado (5 μm) con los kits de hematoxilina y eosina. La altura de las vellosidades (VH) y la profundidad de la cripta (CD) se determinaron bajo un microscopio (Olympus, Tokio, Japón), y se fotografió el campo de visión típico. Cada corte se examinó en busca de aproximadamente 10 vellosidades y criptas, y el promedio de VH y CD de cada corte se utilizó para el análisis estadístico.
2.7 Análisis estadístico
Los datos se analizaron con el programa SPSS (Versión 25.0, SPSS Inc., Chicago, IL). Las diferencias estadísticas entre los diferentes tratamientos se determinaron mediante ANOVA de un factor, y las diferencias entre los diferentes tratamientos se determinaron mediante la prueba de rangos múltiples de Duncan. Se utilizó la prueba HSD de Tukey para analizar la diversidad α y β de la microbiota ruminal. La varianza en las medias etiquetadas sin una letra común es significativamente diferente (p ≤ 0,05).
3 Resultados
3.1 Impacto de la suplementación dietética con diferentes fuentes de RPM ruminales en el rendimiento del crecimiento de las cabras
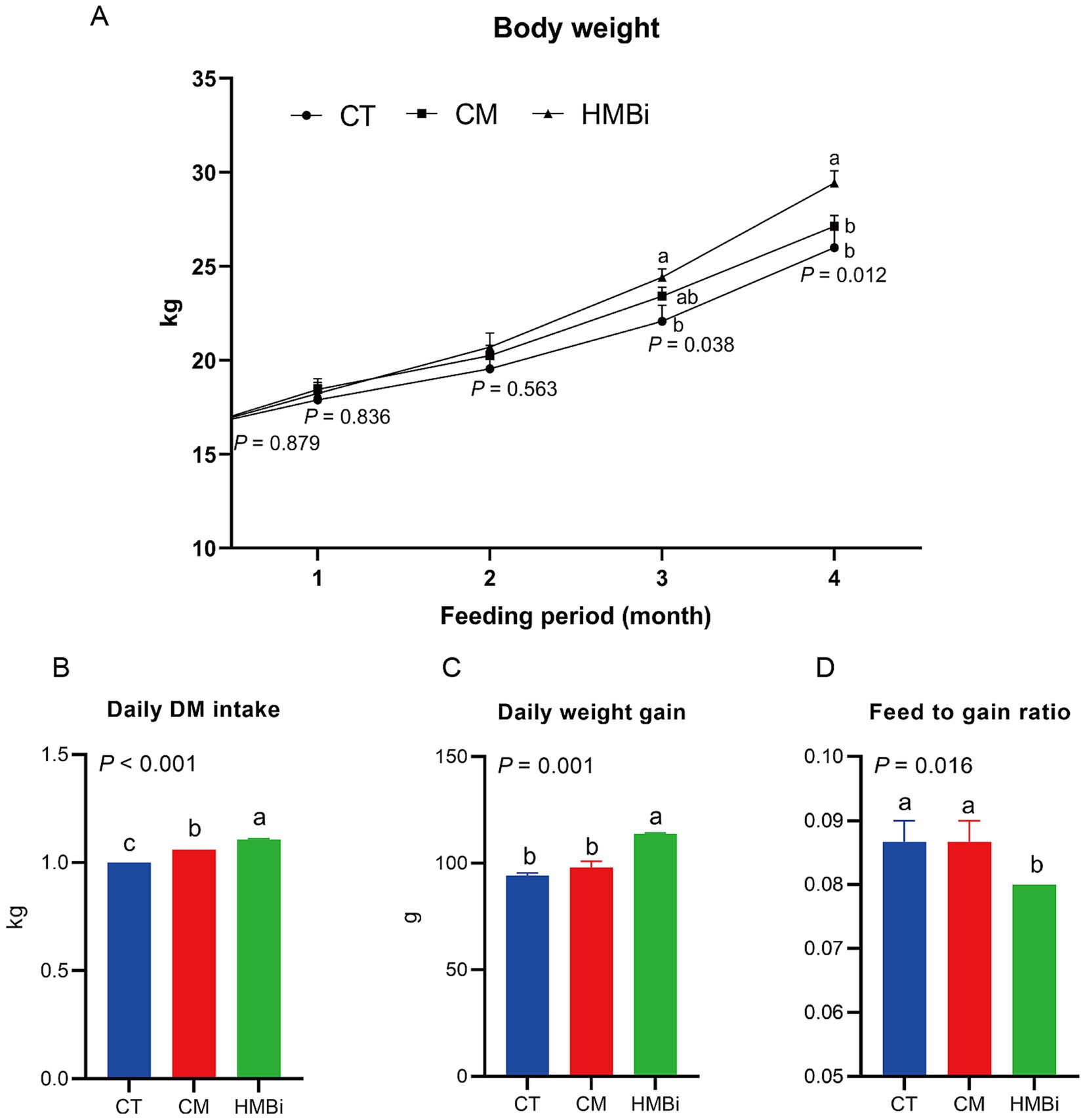
En este estudio, la suplementación con HMBi aumentó significativamente el peso corporal de las cabras después de 3 y 4 meses de alimentación en comparación con la TC (p < 0,05, Figura 1A). A los 3 y 4 meses de alimentación con HMBi, el peso corporal aumentó en un 10,55 y un 13,19%, respectivamente. A pesar de que la suplementación con MC mostró un aumento en el número de peso corporal, no se encontraron diferencias significativas en comparación con la TC (p > 0,05). Además, observamos que una dieta suplementada con HMBi aumentó significativamente la ingesta diaria de MS (p < 0,05, Figura 1B) y la ganancia diaria de peso (p < 0,05, Figura 1C), mientras que la suplementación con MC aumentó significativamente la ingesta diaria de MS (p < 0,05, Figura 1B). Además, la administración de suplementos de HMBi dio lugar a la relación alimento-ganancia (p < 0,05, Figura 1D).
3.2 Efectos de la suplementación dietética con diferentes fuentes de RPM sobre la bioquímica sérica de cabras
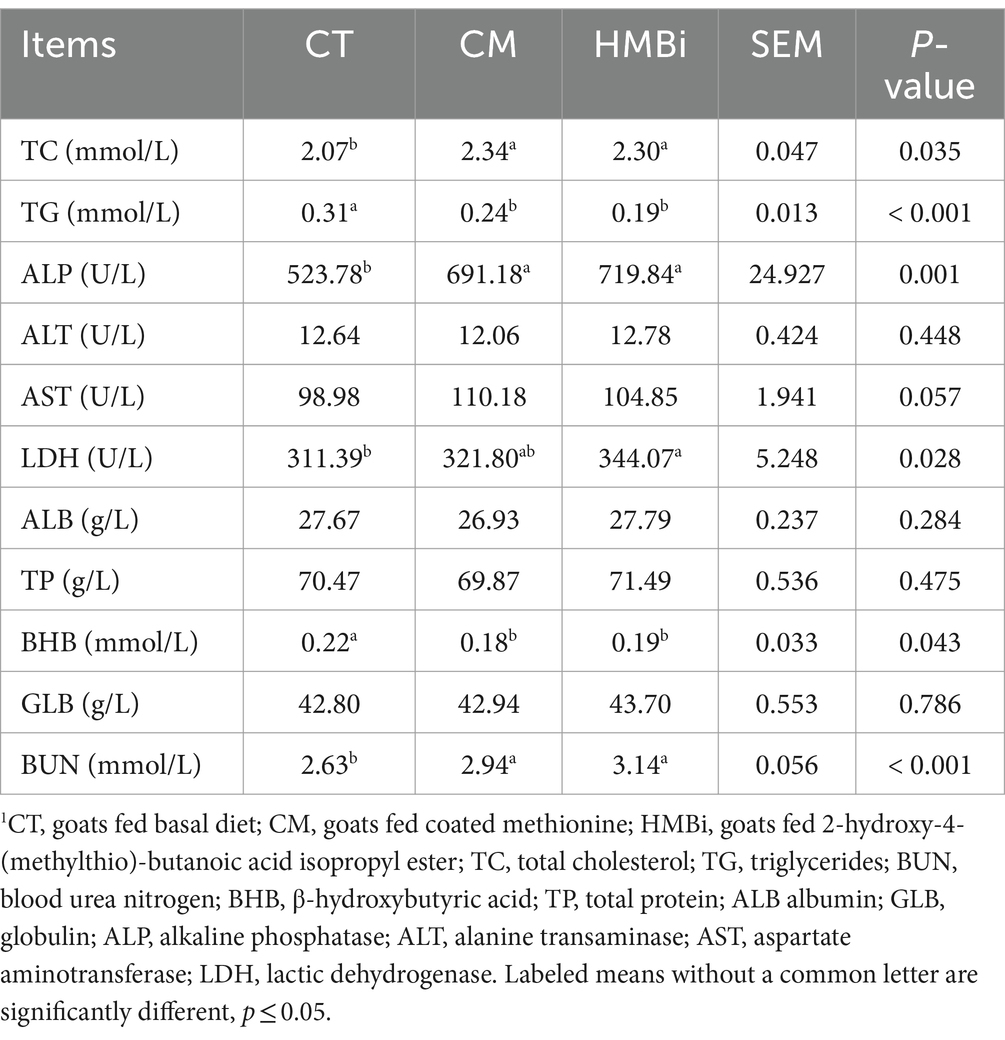
La bioquímica sérica se incluyó en la Tabla 2. Los resultados mostraron que la dieta suplementada con CM o HMBi aumentó significativamente el contenido sérico de TC, BUN, ALP y AST (todos p < 0,05) y disminuyó el contenido de TG y BHB (todos p < 0,05). No se encontraron diferencias significativas para el contenido sérico de TP, ALB, GLB y ALT y LDH (todos p > 0,05).
3.3 Impacto de la suplementación dietética con diferentes fuentes de RPM sobre los parámetros de fermentación ruminal
El parámetro de fermentación ruminal se mostró en la Tabla 3. Encontramos que la dieta suplementada con MC y HMBi aumentó significativamente el pH del líquido ruminal (p = 0,011). Además, tanto la suplementación con CM como con HMBi redujo la concentración de ácido butírico (p = 0,007). Sin embargo, ambas fuentes de aditivos de metionina no influyeron en la concentración de ácido acético, ácido propiónico, ácido acético/ácido propiónico, ácido isobutírico, ácido isovalérico, ácido pentanoico o AGV totales (todos p > 0,05).
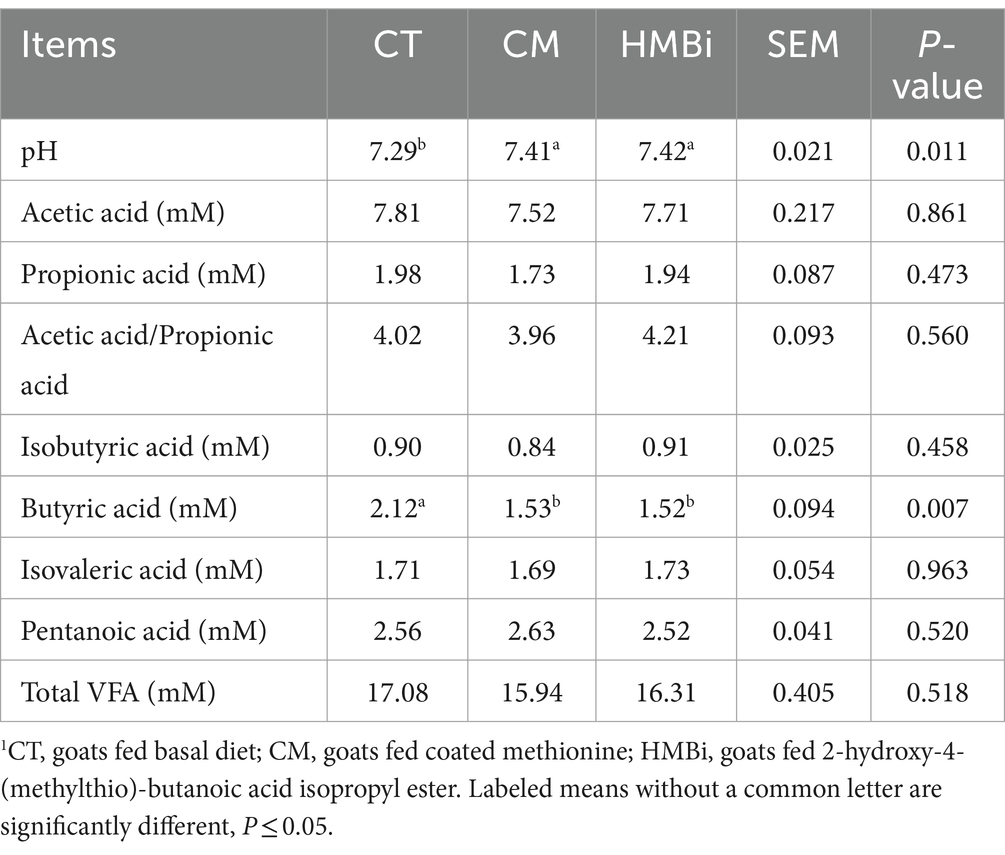
3.4 Impacto de la suplementación dietética con diferentes fuentes de RPM sobre la riqueza y diversidad de la comunidad bacteriana ruminal
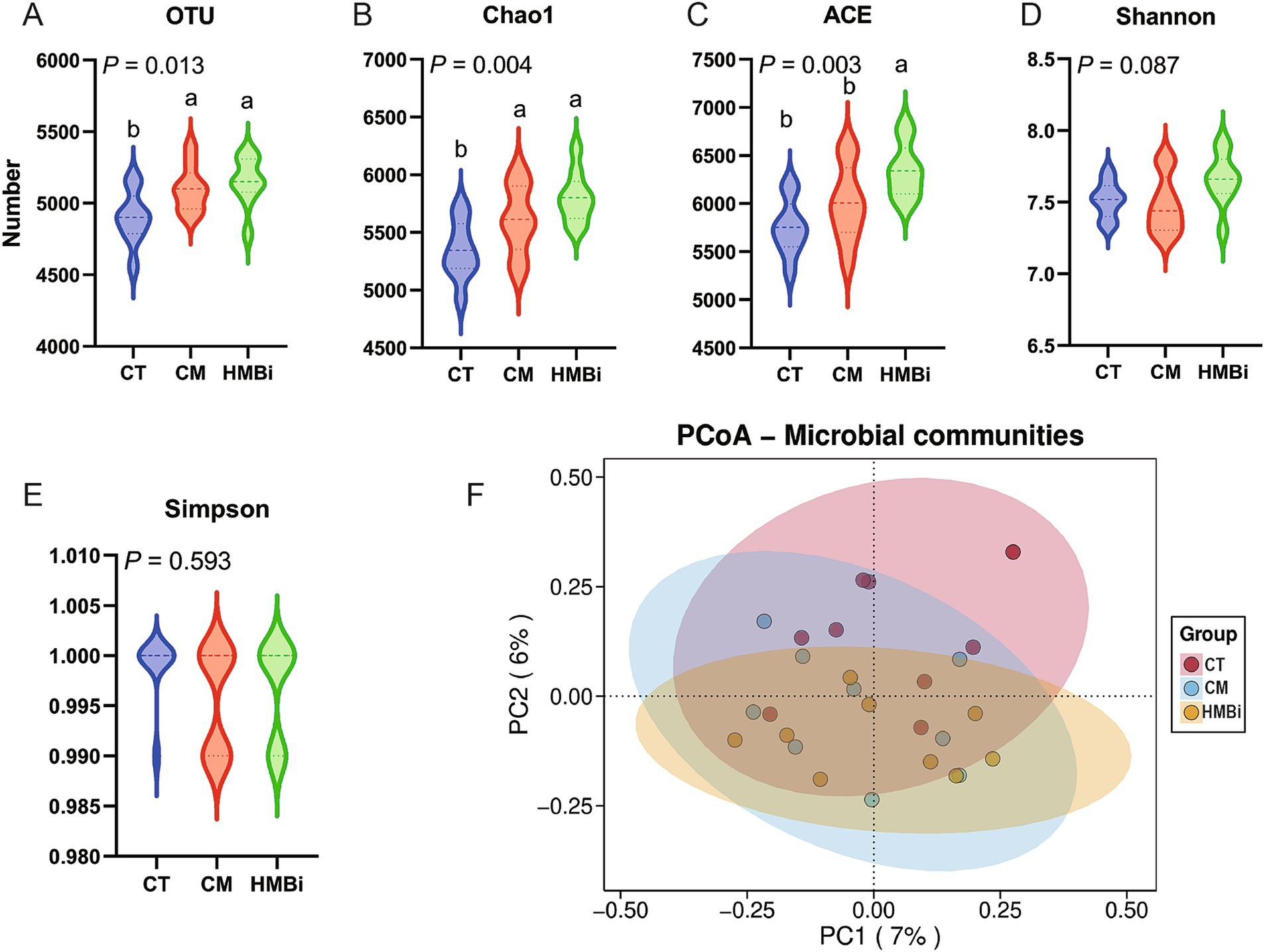
Después de fusionar y filtrar, el análisis de la secuencia del gen ARNr 16S de las muestras de contenido ruminal de cabra en diferentes tratamientos (30 muestras) generó un total de 128.664 secuencias de alta calidad, y las OTU se clasificaron en 31 filos, 67 clases, 129 órdenes, 252 familias y 678 géneros. En la Figura 2 se muestran los índices de diversidad α, incluidos los índices de riqueza comunitaria (OUT, Chao1 y ACE) y los índices de diversidad comunitaria (Shannon y Simpson). Los resultados indican que tanto la suplementación con MC como con HMBi condujeron a un aumento en los niveles de OUT y Chao1 (p < 0,05, Figuras 2A,B). Sin embargo, solo se observó un aumento de la ECA en la suplementación con HMBi (p < 0,01, Figura 2C). No se observaron diferencias entre Shannon y Simpson (p > 0,05, Figuras 2D,E). El PCoA de las OTUs basado en métricas UniFrac ponderadas reveló una clara diferencia entre los tres grupos (Figura 2F).
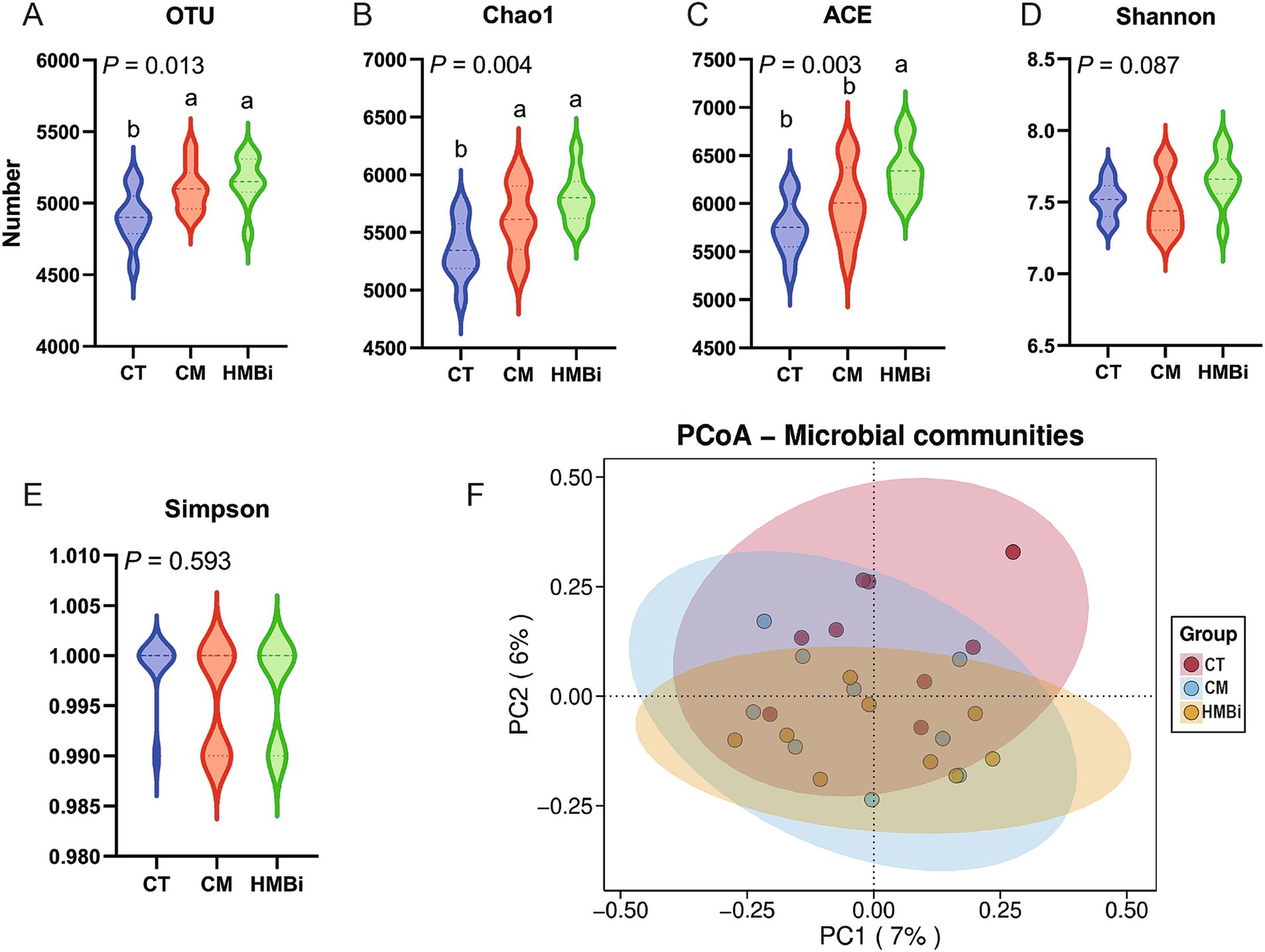
3.5 Impacto de la suplementación dietética con diferentes fuentes de RPM en la composición de las comunidades microbianas y predicción de la función
Las 10 bacterias más abundantes a nivel de filos del líquido ruminal se presentan en la Figura 3A. Encontramos que los filos microbianos dominantes fueron Bacteroidetes (44,66%) y Firmicutes (43,76%), los cuales fermentan fibra dietética y producen AGV (19, 20). Los 10 géneros con mayor abundancia relativa se muestran en la Figura 3B. El género con mayor abundancia fue Prevotella, que representa el 22,01%, seguido de Selenomonas (4,73%), Sodaliphilus (3,86%), Succiniclasticum (3,28%) y Lentimicrobium (3,01%).
Con el fin de identificar biomarcadores microbianos específicos en los tres grupos, los datos se analizaron mediante la selección de LEfSe con puntuaciones de LDA superiores a 2,0 y p < 0,05 entre los tres grupos, y se generó un cladograma (Figuras 3C,D) mediante el análisis LEfSe de la comunidad de microbiota ruminal. Los taxones bacterianos más abundantes en el grupo CT fueron Actinobacteria, Eisenbergiella, Christensenellaceae, Christensenella, Clostridiaceae, Massilioclostridium y Enterocloster. Durante el tratamiento con MC, los taxones bacterianos más diferencialmente abundantes fueron Flavisolibacter, Ruminiclostridium, Desulfobulbus, Desulfobulbaceae y Desulfobacterales. Los taxones bacterianos más prevalentes en el tratamiento con HMBi fueron Paraprevotella, Dysosmobacter, Bacilliculturomica, Denitratisoma y Lachnoclostridium.
3.6 Impacto de la suplementación dietética con diferentes fuentes de RPM en las diferencias taxonómicas de la microbiota ruminal
Como se muestra en la Tabla 4, se ha observado que tanto CM como HMBi resultaron en un aumento significativo en la abundancia de Bacteroidetes y Proteobacterias (ambos p < 0.05); sin embargo, solo el HMBi aumentó significativamente la abundancia de la suma de Bacteroidetes y Firmicutes (p < 0.05). Además, la suplementación con CM y HMBi resultó en una disminución en la abundancia de Planctomycetes y Actinobacteria (p < 0.05). Además, el HMBi dietético aumentó la abundancia de Synergistete (p < 0,05).
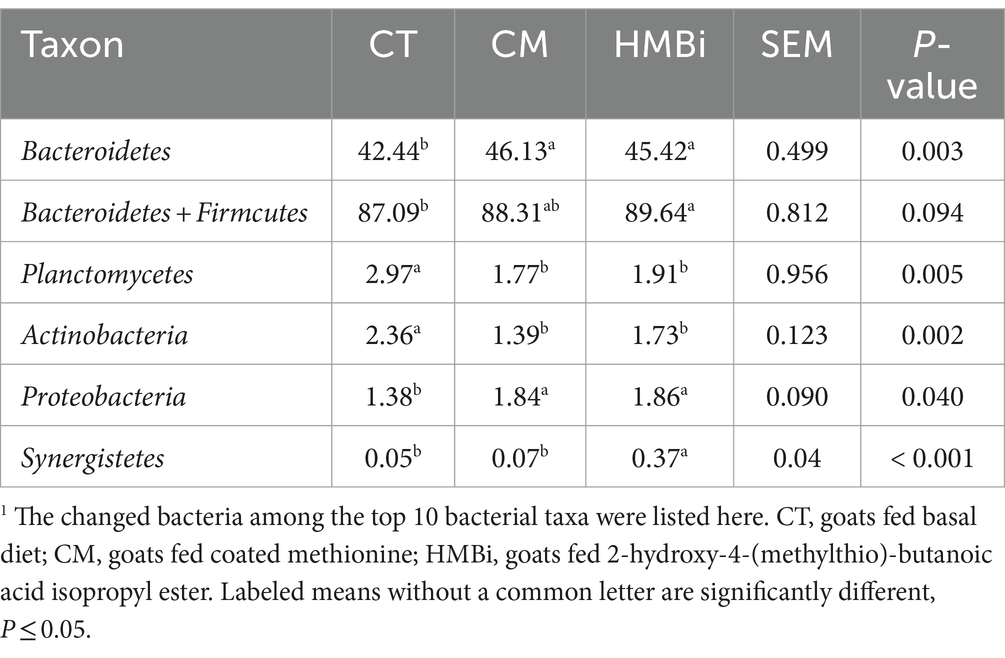
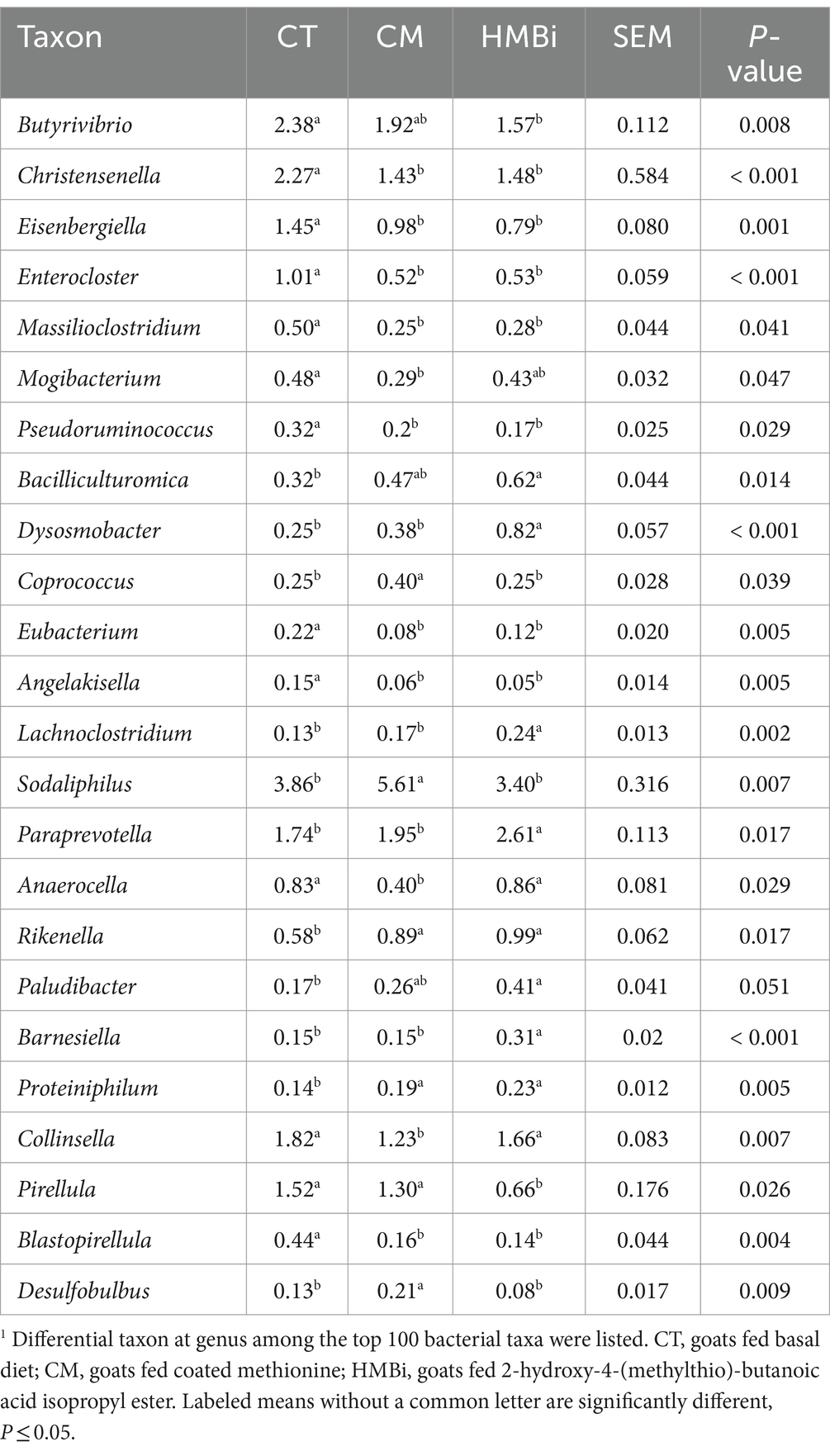
A continuación, examinamos las 100 bacterias principales (abundancia relativa ≥0,05%) y descubrimos que la abundancia relativa de 24 bacterias cambiaba cuando las cabras eran alimentadas con CM o HMBi (Tabla 5). Como se muestra en el diagrama de Venn (Figura 5), se encontró que tanto la suplementación con CM como con HMBi aumentó la abundancia de Rikenella y Proteiniphilum, pero disminuyó la abundancia de Eisenbergiella, Enterocloster, Massilioclostridium, Eubacterium y Angelakisella, Blastopirellula, Christensenella y Pseudoruminococcus (todos p < 0,05). La suplementación con MC aumentó especialmente la abundancia de Desulfobulbus, Sodalifilo y Coprococcus, al tiempo que disminuyó la prevalencia de Anaerocella, Mogibacterium y Collinsella (todos p < 0,05). La suplementación con HMBi aumentó significativamente la abundancia de Paraprevotella, Bacilliculturomica, Lachnoclostridium, Dysosmobacter, Barnesiella y Paludibacter, pero disminuyó la abundancia de Butyrivibrio y Pirellula (todos p < 0,05).
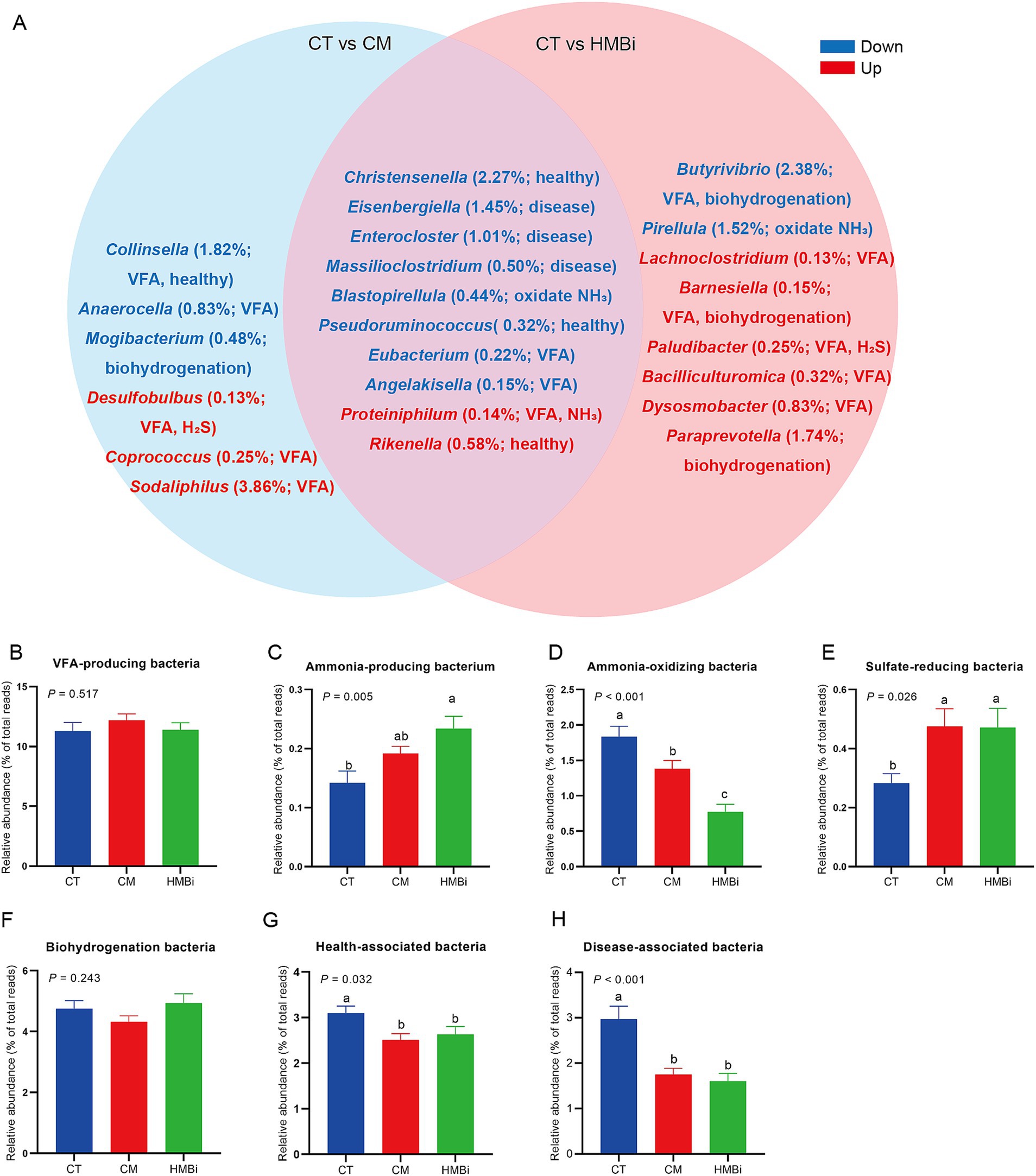
De acuerdo con la función de cada género (Figura 5A), encontramos que tanto CM como HMBi aumentaron la abundancia de bacterias reductoras de sulfato (p < 0.05, Figura 5E) y bacterias productoras de amoníaco (p < 0.05, Figura 5C), pero disminuyeron la abundancia de bacterias oxidantes de amoníaco (p < 0.05, Figura 5D), bacterias asociadas a la salud (p < 0.05, Figura 5G) y bacterias asociadas a enfermedades ( p < 0,05, Figura 5H). Sin embargo, tanto el CM como el HMBi no influyeron en la abundancia de bacterias productoras de AGV (p > 0.05, Figura 5B) y bacterias de biohidrogenación (p > 0.05, Figura 5F).
3.7 Correlaciones entre las bacterias ruminales y los metabolitos séricos
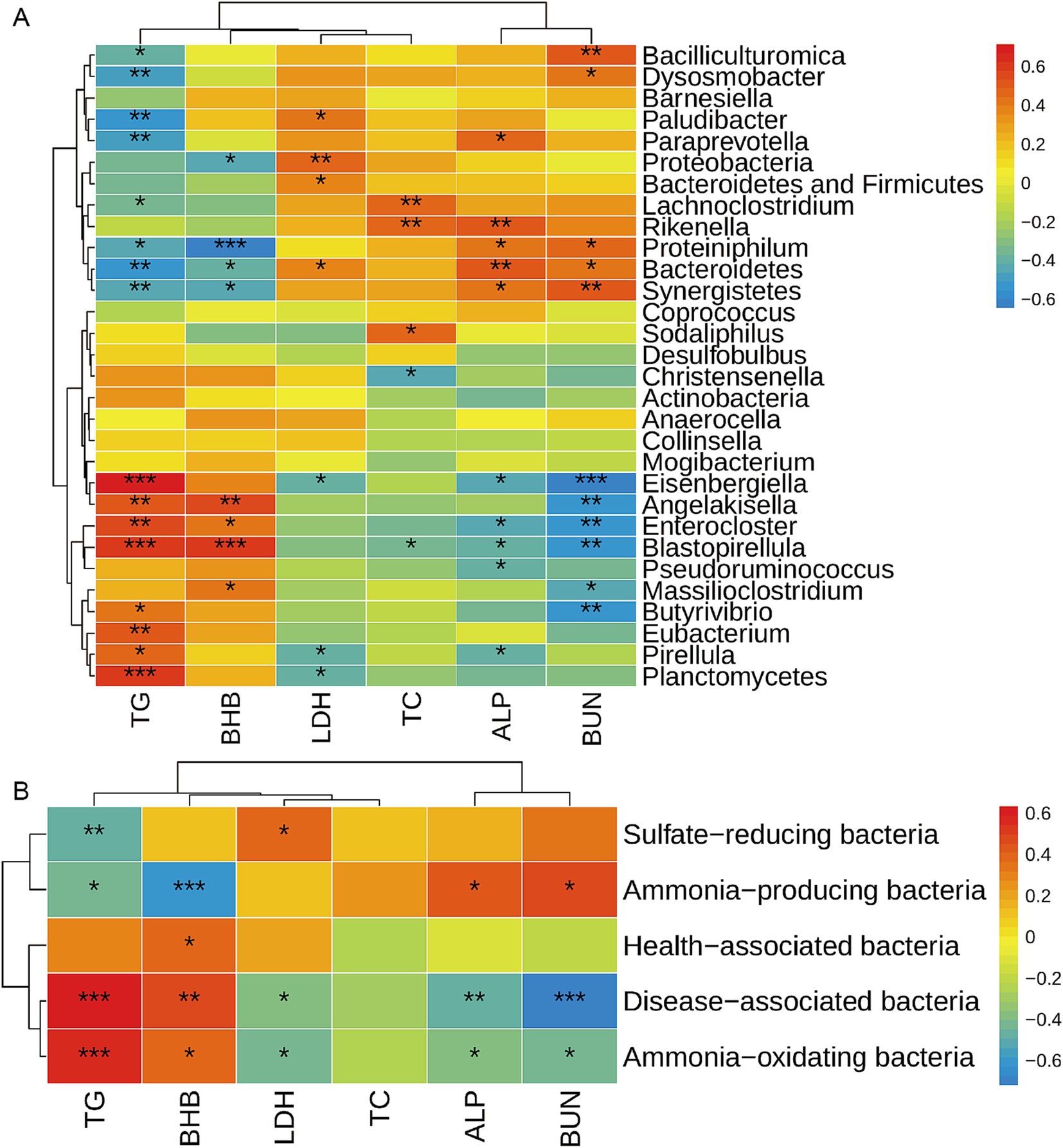
Las bacterias intestinales son complejas, y el trastorno de la comunidad bacteriana suele causar enfermedades como enfermedades neurodegenerativas, enfermedades cardiovasculares, enfermedades metabólicas y enfermedades gastrointestinales (9, 10, 21). Los metabolitos sanguíneos siempre se utilizan como índice para reflejar las enfermedades más comunes. Por lo tanto, analizamos más a fondo las correlaciones entre las bacterias del rumen y los metabolitos séricos. La correlación entre la microbiota ruminal afectada (a nivel de filo y género) y los metabolitos séricos se analizó mediante el análisis de correlación de Spearman (Figura 4A). El TG sérico mostró una correlación positiva con Planctomycetes, Eubacterium, Pirellula, Angelakisella, Butyrivibrio, Blastopirellula, Enterocloster y Eisenbergiella, mientras que se correlacionó negativamente con Bacteroidetes, Synergistetes, Proteiniphilum, Paludibacter, Lachnoclostridium, Paraprevotella, Bacilliculturomica, y Dysosmobacter. El BHB se correlacionó positivamente con Massilioclostridium, Angelakisella, Blastopirelula y Enterocloster, mientras que se correlacionó negativamente con Bacteroidetes, Synergistetes, Proteobacteria y Proteiniphilum. El BUN se correlacionó positivamente con Bacteroidetes, Synergistetes, Proteiniphilum, Bacilliculturomica y Dysosmobacter, mientras que se correlacionó negativamente con Massilioclostridium, Angelakisella, Butyrivibrio, Blastopirellula, Enterocloster y Eisenbergiella. El CT sérico se correlacionó positivamente con Lachnoclostridium, Rikenella y Sodaliphilus, mientras que se correlacionó negativamente con Christensenella y Blastopirellula. La LDH sérica se correlacionó positivamente con Proteobacteria, la suma de Bacterioidetes y Firmicutes, Bacterioidetes y Plaudibacter, mientras que se correlacionó negativamente con Planctomycetes, Pirellilla y Eisenbergiella. La FA sérica se correlacionó positivamente con Bacteroidetes, Synergistetes, Proteiniphilum, Rikenella y Paraprevotella, mientras que se correlacionó negativamente con Pseudoruminococcus, Pirellula, Blastopirellula, Enterocloster y Eisenbergiella.
Además, se analizó la correlación de los metabolitos séricos y las bacterias funcionales mediante análisis Sperson (Figura 4B). Observamos que la TG sérica se correlacionó positivamente con las bacterias asociadas a enfermedades y las bacterias oxidantes de amoníaco, y negativamente con las bacterias reductoras de sulfato y las bacterias productoras de amoníaco. El BHB sérico se correlacionó positivamente con las bacterias asociadas a enfermedades, las bacterias asociadas a la salud y las bacterias oxidantes de amoníaco, mientras que se correlacionó negativamente con las bacterias productoras de amoníaco. La FA sérica se correlacionó negativamente con las bacterias asociadas a enfermedades y las bacterias oxidantes de amoníaco, y se correlacionó positivamente con las bacterias productoras de amoníaco. El BUN se correlacionó negativamente con las bacterias asociadas a enfermedades, las bacterias oxidantes de amoníaco y se correlacionó positivamente con las bacterias productoras de amoníaco.
3.8 Impacto de la suplementación dietética con diferentes fuentes de RPM sobre la morfología del intestino delgado
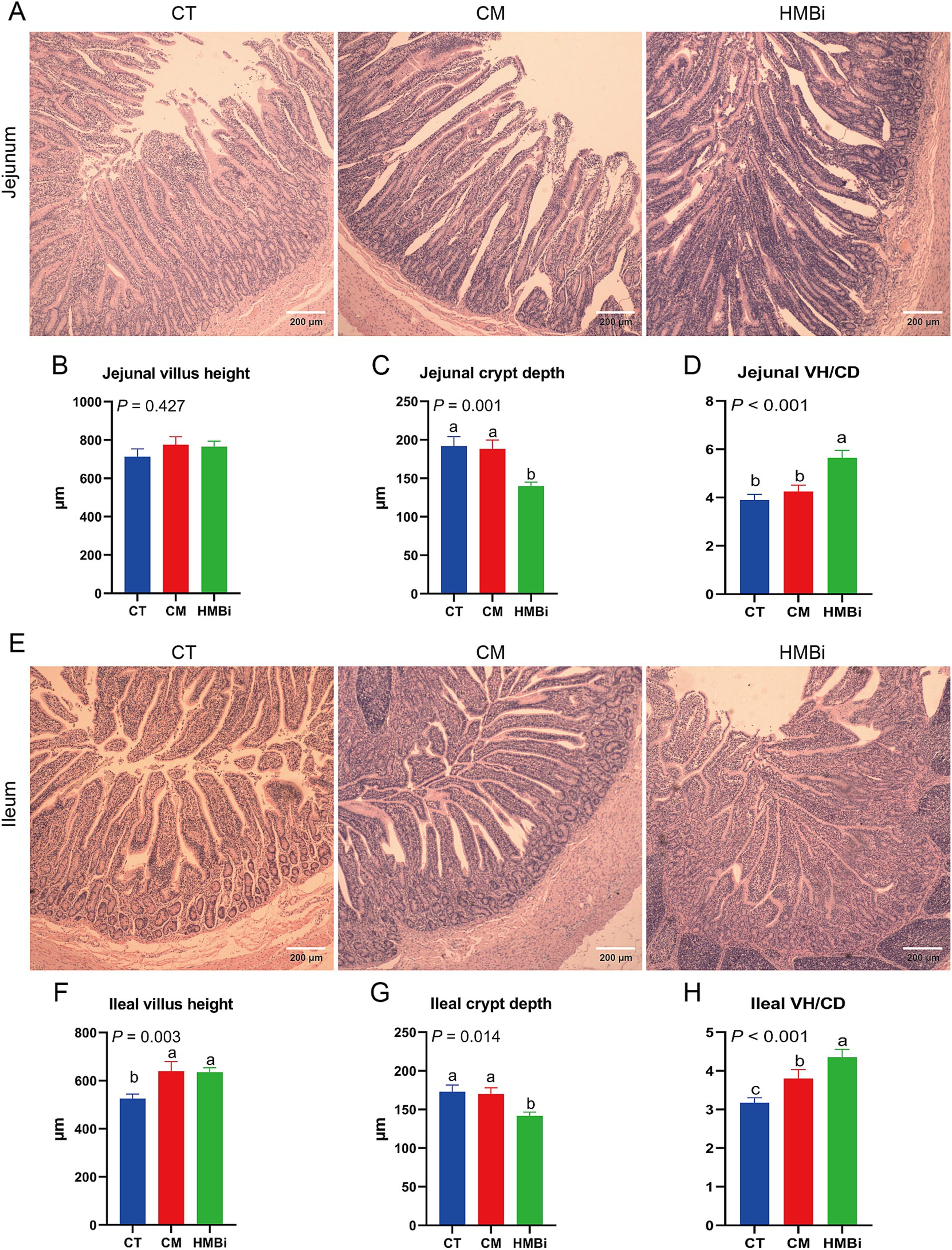
El intestino es un sitio importante para la absorción de nutrientes, como las microproteínas sintetizadas por los microbios del rumen (22). Por lo tanto, investigamos más a fondo la influencia de diferentes fuentes de RPM en el desarrollo del intestino delgado (Figura 6). Los resultados mostraron que la suplementación con HMBi disminuyó la CD (p < 0,05) y aumentó la relación VH/CD (VH/CD, p < 0,05) en yeyuno en comparación con la TC. En íleon, observamos que tanto la suplementación con MC como con HMBi aumentaron la VH y la VH/CD (ambas p < 0,05), pero solo la suplementación con HMBi disminuyó significativamente la CD (p < 0,05).
4 Discusión
Las RPM se utilizan ampliamente en animales rumiantes lecheros, como vacas lecheras y cabras lecheras. Los estudios han demostrado que la suplementación dietética con RPM mejora la grasa y la proteína de la leche (5). Sin embargo, el efecto de las RPM en las cabras de carne no está claro. El rumen contiene una gran diversidad de microbiota y es un sitio importante para la digestión de la celulosa y las hemicelulosas de la dieta. También produce AGV y proteínas microbianas en rumiantes. Los AGV son absorbidos principalmente por el rumen, y las proteínas microbianas suelen ser absorbidas por el abomaso y el intestino. Por lo tanto, este estudio encontró que tanto el CM como el HMBi mejoraron el metabolismo de los lípidos séricos, la composición de la comunidad de microbios del rumen y el desarrollo intestinal, pero el HMBi tuvo un mejor desempeño que el MC. Esto puede explicar la mejora del rendimiento del crecimiento en HMBi.
El peso corporal es el índice clave para la evaluación de la eficiencia. Estudios recientes realizados en cabras en crecimiento han demostrado que la suplementación con MC a una concentración entre 0,1 y 1,15% no tiene impacto sobre el peso corporal (8-15). En este estudio, la dieta suplementada con MC al 0,12% no tuvo ningún efecto sobre el peso corporal, incluso después de un período de alimentación más largo. Parece que la suplementación con MC no influye en el peso corporal, incluso si se alimenta durante mucho tiempo. No se ha reportado aumento de peso en cabras que comen HMBi (8). En este estudio, los períodos de alimentación dentro de los 2 meses no mostraron diferencias entre las cabras alimentadas con HMBi y las dietas basales. Sin embargo, se observó una diferencia significativa después de 3 meses de período de alimentación, y la tasa de crecimiento aumentó gradualmente con un período de alimentación prolongado. Además, encontramos que la ganancia diaria de peso, la ingesta diaria de MS y la relación entre el alimento y la ganancia de peso también mejoraron en las cabras alimentadas con HMBi, lo cual es similar al informe en cabras (8) y terneros Holstein (23). Se ha demostrado que la restricción de metionina en la dieta prolonga la vida útil, y un alto nivel de ingesta de metionina muestra efectos adversos como hiperhomocisteinemia, reducción de peso corporal y aumento de los niveles de colesterol (24). Sin embargo, en este estudio, una dieta suplementada con CM o HMBi no redujo el peso corporal, pero aumentó el nivel de CT sérico. Por lo tanto, teniendo en cuenta que no se observó ningún efecto negativo en el peso corporal cuando las cabras fueron alimentadas con CM o HMBi, especulamos que la dosis de RPM en este estudio fue soportable para las cabras. En resumen, estos resultados indican que 3 meses de alimentación con HMBi al 0,22% es necesaria para mejorar el rendimiento del crecimiento y que la suplementación con MC no influye en el rendimiento del crecimiento.
Los metabolitos séricos suelen reflejar la salud del organismo. Por lo general, el CT y el TG, los principales lípidos séricos, se consideran causas de enfermedad cardiovascular (25, 26). En este estudio, tanto la suplementación con CM como con HMBi disminuyeron el TG sérico, lo que es similar a los resultados de ovejas adultas alimentadas con HMBi (16) y vacas lecheras lactantes (27). Esto implica que tanto la suplementación con CM como con HMBi mejoran el metabolismo de los lípidos séricos. ALP, ALT, AST y LDH son índices séricos de la función hepática (28). La ALT y la AST existen principalmente en las células hepáticas parenquimatosas, con un nivel bajo en la sangre, mientras que la ALP y la LDH aumentaron en la excreción biliar alterada y la hepatitis, respectivamente. En este estudio, tanto la suplementación con CM como con HMBi aumentaron la ALP y la LDH séricas, lo que es similar a los estudios en yaks (29) y ratas (30). Además, la FA sérica se correlaciona positivamente con el efecto promotor del crecimiento en yaks alimentados con HMBi (29). Del mismo modo, se observó un aumento del peso corporal en las cabras alimentadas con HMBi. Por lo tanto, la mejora del rendimiento del crecimiento puede ser a expensas del daño hepático. El BHB es un indicador de cetonemia. Los niveles más altos de BHB suelen estar causados por la gluconia, la hiperlipidemia y la proteólisis durante el balance energético negativo (31). En este estudio, encontramos que tanto la suplementación con CM como con HMBi disminuyeron el BHB sérico, lo que sugiere que la capacidad anticetonemia se fortalece durante la suplementación con MC o HMBi. Además, se observó que tanto el CM como el HMBi aumentaron el BUN, lo cual es similar a lo reportado en el ganado bovino (6). El BUN refleja la producción de amoníaco en el rumen. En el rumen, los llamados Planctomycetes ‘anammox’ tienen una capacidad única para oxidar el amonio a nitrógeno (32), y Proteiniphilum utiliza el nitrógeno para sintetizar amoníaco (33). En este estudio, la suplementación con CM y HMBi disminuyó la abundancia de Planctomycetes, pero aumentó la abundancia de Proteiniphilum, lo que significa que se produjo más amoníaco. Además, un informe ha demostrado que tanto la suplementación con CM como con HMBi aumentan la utilización de nitrógeno (34). Por lo tanto, el aumento del BUN durante la suplementación con RPM implica una mejor utilización del nitrógeno en el rumen. En resumen, la suplementación con CM y HMBi mejora el metabolismo de los lípidos séricos y la utilización del nitrógeno y disminuye el riesgo de cetonemia, pero aumenta el riesgo de daño hepático.
Los AGV producidos por los microbios del rumen contienen principalmente ácido acético, ácido propiónico, ácido isobutírico, ácido butírico, ácido isovalérico y ácido pentanoico (35). Los AGV fueron absorbidos principalmente por el rumen, y el resto fue absorbido por el omaso y el abomaso. Por lo tanto, la concentración de AGV ruminales está determinada por la fermentación microbiana y la absorción de rumen, omaso y abomaso. En este estudio, se encontraron concentraciones disminuidas de ácido butírico en CM y HMBi en la dieta tanto en la suplementación con CM como con HMBi, lo cual es similar al estudio realizado en ratones (36). Por el contrario, el estudio en vacas ha demostrado que la alimentación con MC aumentó la concentración de butirato ruminal (11, 12). La disminución de la concentración de ácido butírico puede ser el resultado de la disminución de la abundancia de Butyrivibrio, una bacteria clave para la producción de ácido butírico (37). A pesar de que en este estudio se observó un aumento en la ingesta de alimento y bacterias degradadoras de fibra (suma de Bacteroidetes y Firmicutes) en HMBi alimentados con cabras, no se encontraron diferencias en la concentración de AGV ruminales. Del mismo modo, tampoco se encontraron diferencias en las vacas lactantes (38, 39). Un estudio reciente ha demostrado que la metionina beneficia el crecimiento de las células epiteliales en el rumen y el intestino (4). Aquí, creemos que la suplementación con HMBi aumenta las bacterias productoras de AGV, acompañadas de una mayor tasa de absorción de AGV en el rumen.
Una microbiota ruminal muy diversa se considera generalmente beneficiosa para la salud del huésped y se considera un signo de una microbiota ruminal madura (40). En este estudio, los índices de diversidad alfa (OUT y Chao1) de la microbiota ruminal aumentaron significativamente en las cabras alimentadas con CM o HMBi. Además, se ha reportado que un pH alto se acompaña de una alta diversidad bacteriana (41). De manera similar, se observó un aumento del pH ruminal en cabras alimentadas con CM o HMBi en este estudio.
A nivel de filo, Bacteroidetes (44,66%) y Firmicutes (43,76%) fueron los filos dominantes en el rumen y representaron la mayoría de la comunidad microbiana en las cabras Jianzhou Da’er, que es similar a las ovejas Hu, las ovejas Tan, las ovejas Dorper y las cabras Bengala (42, 43). Bacteroidetes y Firmicutes son reconocidos por su capacidad para degradar una amplia gama de carbohidratos complejos para producir AGV (19, 20). En este estudio, las cabras alimentadas con HMBi mostraron un mayor nivel de la suma de Bacteroidetes y Firmicutes, lo que significa que el HMBi es beneficioso para la descomposición de los carbohidratos y la producción de AGV. Esto explica la alta ingesta diaria de alimento en el grupo HMBi. Los planctomycetes pueden oxidar el amoníaco en nitrógeno (32) y Proteobacteria es una bacteria fijadora de nitrógeno (44). En este estudio, tanto la suplementación con HMBi como con CM disminuyó la abundancia de la bacteria oxidante de amoníaco Planctomycetes y aumentó la bacteria fijadora de nitrógeno Proteobacteria, lo que significa que tanto la suplementación con HMBi como con CM se benefician de un alto nivel de amoníaco ruminal. Esto explica el alto BUN en CM y HMBi alimentados con cabras. Varios géneros de Actinobacteria (45) y Proteobacteria (46) tienen la capacidad de combatir bacterias patógenas. En este estudio, el aumento de patógenos contra la bacteria Proteobacteria compensó la disminución de la abundancia de Actinobacterias. Los sinergistas pueden usar aminoácidos para producir AGV (47) y también tienen genes para la biosíntesis de lipopolisacáridos (48). En resumen, a nivel de filo, estos hallazgos sugieren que tanto la suplementación con CM como con HMBi tienen un impacto positivo en la producción de amoníaco, con HMBi mostrando un beneficio particular en la descomposición de los carbohidratos.
La mayoría de las bacterias ruminales en Firmicutes y Bacteroidetes pueden hidrolizar la fibra dietética y producir AGV (19, 20). En este estudio, a pesar de que la mayoría de las bacterias diferenciales eran bacterias productoras de AGV, como Butyrivibrio (49), Bacilliculturomica (50), Dysosmobacter (51), Coprococcus (14), Eubacterium (52), Angelakisella (53), Lachnoclostridium (54, 55), Sodaliphilus (56), Anaerocella (57), Barnesiella (58) y Collinsella (17, 18), la suma de estas bacterias mostró que tanto la CM como la HMBi no influyeron en la abundancia de las bacterias productoras de AGV, lo que explica el resultado de AGV totales en el líquido ruminal. Además, la dieta de los rumiantes contiene principalmente ácidos grasos insaturados de 18 carbonos, como el ácido α-linolénico, el ácido linoleico y el ácido oleico. Bajo la biohidrogenación de bacterias como Paraprevotella (59), Barnesiella (58) y Mogibacterium (60), el ácido graso insaturado se convirtió en ácido graso saturado, seguido de la hidrogenación de los dobles enlaces (61). A pesar de que se justifica una baja biohidrogenación para un producto rico en ácidos grasos insaturados (62), no se encontraron diferencias durante la suplementación con MC y HMBi. Por lo tanto, la suplementación con CM y HMBi no mostró influencia sobre la biohidrogenación ruminal. Además, las bacterias del rumen fermentan las sustancias nitrogenadas en amoníaco, y el amoníaco se utiliza para la síntesis de proteínas microbianas por parte de las bacterias productoras de amoníaco y luego fluye hacia el intestino posterior para ser absorbido (22). Además, una parte del amoníaco fue oxidado en nitrógeno por bacterias oxidantes de amoníaco (33). En este estudio, la bacteria productora de amoníaco Proteiniphilum (33) aumentó cuando la dieta se suplementó con HMBi. Además, tanto CM como HMBi disminuyeron las bacterias oxidantes de amoníaco, incluidas Pirellula (63) y Blastopirellula (64). Del mismo modo, los estudios en vacas han demostrado que la suplementación con HMBi aumenta la eficiencia del nitrógeno (34) y de la proteína de la leche (5). Estos resultados implican que CM y HMBi tienen un impacto positivo en la utilización del nitrógeno, lo cual es similar a los resultados del análisis en los filos y explica también el aumento del BUN. El azufre de la dieta puede ser reducido a sulfuro de hidrógeno (H2S) en el rumen por bacterias reductoras de sulfato, como Desulfobulbus (65) y Paludibacter (66). ElH2Ses un gas tóxico que puede ser absorbido rápidamente por la pared del rumen, y la acumulación excesiva deH2Sen el rumen puede plantear riesgos de toxicidad para los animales rumiantes (67). Se ha demostrado que el nivel de azufre en la dieta se correlacionó positivamente con la producción deH2S(68). La metionina es un aminoácido que contiene azufre. Además, nuestro estudio reveló distintos mecanismos por los cuales CM y HMBi mejoran la producción deH2S, con CM promoviendo un aumento de Desulfobulbus y HMBi facilitando un aumento de Paludibacter. En este estudio, se observó que tanto el CM como el HMBi condujeron a un aumento de las bacterias reductoras de sulfato, lo que significa un aumento de la producción deH2S. Además, descubrimos que tanto la suplementación con CM como con HMBi disminuyó la abundancia de bacterias asociadas a la enfermedad Eisenbergiella (69), Enterocloster (70) y Massilioclostridium (71), y las bacterias asociadas a la salud Christensenella (72), Pseudoruminococcus (54, 55) y Rikenella (73-75). La disminución de las bacterias asociadas a la salud puede ser un efecto receptivo de la disminución de las bacterias asociadas a la enfermedad. Por lo tanto, la suplementación con CM y HMBi beneficia la salud del cuerpo. En resumen, se ha encontrado que la suplementación con RPM tiene efectos positivos sobre la producción de amoníaco yH2Sy disminuye el riesgo de enfermedad.
El intestino delgado desempeña un papel importante en la absorción de las proteínas microbianas que se producen en el rumen (3). En general, una VH más alta significa una mayor área de absorción, mientras que una CD poco profunda significa una tasa de crecimiento rápido de las vellosidades (76). Un estudio en cerdas demostró que la suplementación con metionina o HMBi aumentó el VH y la relación VH/CD de los lechones (77). En este estudio, encontramos que tanto la CM como la HMBi aumentaron la VH ileal y la suplementación con HMBi disminuyó la CD, con HMBi mejorando. Trabajos anteriores han demostrado que la metionina beneficia el desarrollo de criptas aisladas (4), por lo que el mejor desarrollo del intestino aquí puede ser causado por el desarrollo de criptas inducido por la metionina. En resumen, estos resultados implican que tanto la suplementación con MC como con HMBi mejoran la morfología intestinal, especialmente la suplementación con HMBi.
5 Conclusión
Este estudio indica que tanto la suplementación con CM como con HMBi mejora el metabolismo de los lípidos, la utilización del nitrógeno y el desarrollo intestinal. En contraste con la suplementación con MC, el efecto promotor del crecimiento de la suplementación con HMBi puede atribuirse a la mayor abundancia de bacterias productoras de AGV en el rumen, bacterias que utilizan nitrógeno y un mayor desarrollo intestinal. Este estudio sugiere que el HMBi es un buen aditivo de metionina para las cabras de carne.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio se incluyen en el artículo/material complementario, las consultas posteriores pueden dirigirse al autor correspondiente.
Declaración ética
Los estudios en animales fueron aprobados por el Comité de Ética y Cuidado de Animales de la Universidad del Suroeste de Minzu. Los estudios se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
YouW: Conceptualización, Curación de datos, Análisis formal, Obtención de fondos, Metodología, Software, Visualización, Redacción – borrador original. WL: Administración de proyectos, redacción, revisión y edición. AL: Administración de proyectos, redacción, revisión y edición. WQ: Administración de proyectos, redacción, revisión y edición. CG: Análisis formal, Redacción – revisión y edición. XL: Análisis formal, Redacción – revisión y edición. JX: Escritura – revisión y edición. HS: Conceptualización, Curación de datos, Análisis formal, Adquisición de fondos, Metodología, Software, Visualización, Redacción – borrador original. YanL: Redacción – revisión y edición. YonW: Metodología, Supervisión, Redacción – revisión y edición. YaqL: Conceptualización, Curación de datos, Análisis formal, Adquisición de fondos, Metodología, Software, Visualización, Redacción – borrador original.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este trabajo fue apoyado por los Fondos de Investigación de la Universidad del Suroeste de Minzu (subvención no. RQD2022037), la Fundación Nacional de Ciencias Naturales de China (subvención n.º 32372857), el Proyecto de Desarrollo de la Construcción del Equipo de Innovación de Sichuan del Sistema Nacional de Tecnología de la Industria Agrícola Moderna (SCCXTD-2024-14) y el Programa de Ciencia y Tecnología de Sichuan (subvención n.º 2024NSFSC1171).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Graulet, B, Richard, C, y Robert, JC. Disponibilidad de metionina en plasma de vacas lecheras suplementadas con éster isopropílico análogo de hidroxi metionina. J Lácteos Sci. (2005) 88:3640–9. DOI: 10.3168/JDS. S0022-0302(05)73049-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Lobley, GE, Wester, TJ, Holtrop, G, Dibner, JJ, Parker, DS y Vázquez-Anon, M. Absorción y metabolismo del tracto digestivo del ácido 2-hidroxi-4-metiltiobutanoico en corderos. J Lácteos Sci. (2006) 89:3508–21. DOI: 10.3168/JDS. S0022-0302(06)72391-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Stein, HH, Sève, B, Fuller, MF, Moughan, PJ, de Lange, CFM, y Comité, OTTR. Revisión invitada: biodisponibilidad y digestibilidad de aminoácidos en ingredientes de alimentos para cerdos: terminología y aplicación. J Anim Sci. (2007) 85:172–80. doi: 10.2527/jas.2005-742
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Wang, Y, Hou, Q, Wu, Y, Xu, Y, Liu, Y, Chen, J, et al. La deficiencia de metionina y su análogo hidroxilo influyen en el desarrollo de organoides tridimensionales intestinales de pollos. Anim Nutr. (2022) 8:38–51. doi: 10.1016/j.aninu.2021.06.001
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Wei, C, He, T, Wan, X, Liu, S, Dong, Y y Qu, Y. Metaanálisis de la metionina protegida del rumen en la producción de leche y la composición de las vacas lecheras. Animales. (2022) 12:1505. doi: 10.3390/ani12121505
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Qin, X, Zhang, D, Qiu, X, Zhao, K, Zhang, S, Liu, C, et al. La suplementación con éster isopropílico del ácido 2-hidroxi-4-(metiltio) butanoico alteró la composición bacteriana ruminal y cecal y mejoró el rendimiento del crecimiento del ganado de carne de engorde. Nuerca delantera. (2022) 9:833881. doi: 10.3389/fnut.2022.833881
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Inhuber, V, Windisch, W, Bächler, B, Schuster, M, Spiekers, H y Ettle, T. Efectos de complementar una dieta reducida en PC con metionina protegida del rumen en el engorde de toros fleckvieh. Animal. (2021) 15:100366. doi: 10.1016/j.animal.2021.100366
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Chen, D, Yan, J, Shen, W, Song, Y, Lan, X, Yi, K, et al. Efecto de la inclusión de HMBi en la ración de cabras sobre la ingesta de alimento, la digestibilidad de los nutrientes, la comunidad de bacterias ruminales y los parámetros del suero sanguíneo. J Anim Physiol Anim Nutr. (2020) 104:987–97. doi: 10.1111/jpn.13270
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Chen, J, Niu, X, Li, F, Li, F y Guo, L. Sustitución de la harina de soja por granos secos de destilería con solubles más lisina y metionina protegidas en el rumen: efectos sobre el rendimiento del crecimiento, la digestión de nutrientes, la fermentación ruminal y los parámetros séricos en ovejas Hu. Animales. (2021) 11:2428. doi: 10.3390/ani11082428
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Chen, Y, Zhou, J y Wang, L. Papel y mecanismo de la microbiota intestinal en las enfermedades humanas. Las células frontales infectan el microbiol. (2021) 11:625913. doi: 10.3389/fcimb.2021.625913
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Li, S, Zeng, H, Wang, C y Han, Z. Efecto de los suplementos alimenticios análogos de hidroxi metionina: alteración significativa y enriquecimiento de la microbiota y el metaboloma del rumen en ovejas Hu. Frente Vet Sci. (2022) 9:999726. doi: 10.3389/fvets.2022.999726
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Li, Y, Wei, J, Dou, M, Liu, S, Yan, B, Li, C, et al. Efectos de la suplementación con metionina protegida del rumen sobre el rendimiento de la producción, la digestibilidad aparente, los parámetros sanguíneos y la fermentación ruminal de vacas lecheras Holstein lactantes. Frente Vet Sci. (2022) 9:981757. doi: 10.3389/fvets.2022.981757
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Montout, L, Bahloul, L, Feuillet, D, Jean-Bart, M, Archimède, H y Bambou, J. La suplementación con metionina protegida en el rumen redujo el efecto parasitario de Haemonchus contortus en cabras. Veterinario Sci. (2023) 10:559. doi: 10.3390/vetsci10090559
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Wang, S, Kong, F, Liu, J, Xia, J, Du, W, Li, S, et al. Análisis comparativo de la composición de la microbiota ruminal en vacas lecheras con indigestión simple y vacas sanas. Microorganismos. (2023) 11:2673. doi: 10.3390/microorganismos11112673
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Wang, W, Ye, L, Dou, X, Liu, H y Han, D. Efectos de la suplementación con metionina protegida en el rumen sobre el rendimiento del crecimiento, la digestión de nutrientes, la utilización de nitrógeno y los perfiles de aminoácidos plasmáticos de cabras de cachemira de Liaoning. Animales. (2023) 13:2995. doi: 10.3390/ani13192995
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Baghbanzadeh-Nobari, B, Taghizadeh, A, Khorvash, M, Parnian-Khajehdizaj, F, Maloney, SK, Hashemzadeh-Cigari, F, et al. Digestibilidad, fermentación ruminal, metabolitos sanguíneos y estado antioxidante en ovejas suplementadas con dl-metionina o éster isopropílico de ácido hidroxi-4 (metiltio) butanoico. J Anim Physiol Anim Nutr. (2017) 101:E266-77. doi: 10.1111/jpn.12600
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Consejo Nacional de Investigación de las Academias Nacionales, División de Estudios de la Tierra y la Vida, Junta de Agricultura y Recursos Naturales, Comité de Necesidades de Nutrientes de los Pequeños Rumiantes. Editores. (2007). Requerimientos de nutrientes de los pequeños rumiantes. Editorial de las Academias Nacionales. 271.
18. He, Q, Gao, H, Chang, Y, Wu, X, Lin, R, Li, G, et al. ETS-1 facilita la inflamación de la mucosa mediada por células th1 en enfermedades inflamatorias intestinales a través de la regulación positiva de CIRBP. J Autoimmun. (2022) 132:102872. doi: 10.1016/j.jaut.2022.102872
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Él, Z, Ma, Y, Yang, S, Zhang, S, Liu, S, Xiao, J, et al. El ácido ursodesoxicólico derivado de la microbiota intestinal procedente de terneros lecheros recién nacidos mejora la homeostasis intestinal y la colitis para atenuar la infección por Escherichia coli enteroagregativa productora de β-lactamasas de espectro extendido. Microbioma. (2022) 10:79. doi: 10.1186/s40168-022-01269-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. McKee, LS, La Rosa, SL, Westereng, B, Eijsink, VG, Pope, PB y Larsbrink, J. Degradación de polisacáridos por los Bacteroidetes: mecanismos y nomenclatura. Environ Microbiol Rep. (2021) 13:559–81. doi: 10.1111/1758-2229.12980
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Sun, Y, Zhang, S, Nie, Q, He, H, Tan, H, Geng, F, et al. Gut Firmicutes: relación con la fibra dietética y papel en la homeostasis del huésped. Crit Rev Comida Sci Nutr. (2023) 63:12073–88. doi: 10.1080/10408398.2022.2098249
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Witkowski, M, Weeks, TL y Hazen, SL. Microbiota intestinal y enfermedades cardiovasculares. Circ Res. (2020) 127:553–70. doi: 10.1161/CIRCRESAHA.120.316242
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Ansia, I, Stein, HH, Brøkner, C, Hayes, CA, y Drackley, JK. Digestibilidad de nutrientes y pérdidas de proteínas endógenas en el intestino anterior y delgado de terneros lecheros destetados alimentados con iniciadores de terneros con harina de soja convencional o tratada con enzimas. J Lácteos Sci. (2021) 104:2979–95. doi: 10.3168/jds.2020-18776
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Molano, RA, Saito, A, Luchini, DN, y Van Amburgh, ME. Efectos de la metionina protegida ruminal o los análogos de la metionina en el iniciador sobre los metabolitos plasmáticos, el crecimiento y la eficiencia de los terneros Holstein de 14 a 91 días de edad. J Lácteos Sci. (2020) 103:10136–51. doi: 10.3168/jds.2020-18630
25. Navik, U, Sheth, VG, Khurana, A, Jawalekar, SS, Allawadhi, P, Gaddam, RR, et al. La metionina como arma de doble filo en salud y enfermedad: perspectiva actual y retos futuros. Envejecimiento Res Rev. (2021) 72:101500. doi: 10.1016/j.arr.2021.101500
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Jung, E, Kong, SY, Ro, YS, Ryu, HH y Shin, SD. Niveles de colesterol sérico y riesgo de muerte cardiovascular: una revisión sistemática y un metanálisis dosis-respuesta de estudios de cohortes prospectivos. Int J Environ Res Salud Pública. (2022) 19:8272. doi: 10.3390/ijerph19148272
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Tao, LC, Xu, JN, Wang, TT, Hua, F y Li, JJ. Índice de triglicéridos-glucosa como marcador en enfermedades cardiovasculares: panorama y limitaciones. Cardiovasc Diabetol. (2022) 21:68. doi: 10.1186/s12933-022-01511-x
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Giallongo, F, Harper, MT, Oh, J, Lopes, JC, Lapierre, H, Patton, RA, et al. Efectos de la metionina, lisina e histidina protegidas del rumen en el rendimiento de la lactancia de las vacas lecheras. J Lácteos Sci. (2016) 99:4437–52. doi: 10.3168/jds.2015-10822
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Tamber, SS, Bansal, P, Sharma, S, Singh, RB y Sharma, R. Biomarcadores de enfermedades hepáticas. Mol Biol Rep. (2023) 50:7815–23. doi: 10.1007/s11033-023-08666-0
30. Zhang, X, Zuo, Z, Liu, Y, Wang, C, Peng, Z, Zhong, J, et al. Efecto de los análogos de metionina sobre el rendimiento del crecimiento, los parámetros bioquímicos séricos, los aminoácidos libres séricos y la fermentación ruminal de los yaks. Animales. (2022) 12:3175. doi: 10.3390/ani12223175
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Derouiche, F, Djemil, R, Sebihi, FZ, Douaouya, L, Maamar, H y Benjemana, K. El estrés oxidativo mediado por la dieta con alto contenido de metionina y el deterioro del proteasoma causan toxicidad en el hígado. Sci Rep. (2024) 14:5555. doi: 10.1038/s41598-024-55857-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Overton, TR y Waldron, MR. Manejo nutricional de vacas lecheras en transición: estrategias para optimizar la salud metabólica. J Lácteos Sci. (2004) 87:E105-19. DOI: 10.3168/JDS. S0022-0302(04)70066-1
33. Fuerst, JA, y Sagulenko, E. Más allá de la bacteria: los planctomycetes desafían nuestros conceptos de estructura y función microbiana. Nat Rev Microbiol. (2011) 9:403–13. doi: 10.1038/nrmicro2578
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. He, Q, Li, L, Zhao, X, Qu, L, Wu, D y Peng, X. Investigación de las causas de la formación de espuma en tres digestores de residuos de alimentos mesófilos: rendimiento del reactor y análisis microbiano. Sci Rep. (2017) 7:13701. doi: 10.1038/s41598-017-14258-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Chen, ZH, Broderick, GA, Luchini, ND, Sloan, BK y Devillard, E. Efecto de la alimentación con diferentes fuentes de metionina protegida por el rumen en la producción de leche y la utilización de N en vacas lecheras lactantes. J Lácteos Sci. (2011) 94:1978–88. doi: 10.3168/jds.2010-3578
36. Beckett, L, Gleason, CB, Bedford, A, Liebe, D, Yohe, TT, Hall, MB, et al. Proporciones molares de ácidos grasos volátiles ruminales, expresión génica epitelial ruminal y respuestas de concentración de metabolitos sanguíneos a los suministros de almidón y fibra degradables ruminalmente. J Lácteos Sci. (2021) 104:8857–69. doi: 10.3168/jds.2020-19622
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Xu, Y, Yang, Y, Li, B, Xie, Y, Shi, Y y Le, G. La restricción de metionina en la dieta mejora la composición de la microbiota intestinal y previene el deterioro cognitivo en ratones envejecidos inducidos por d-galactosa. Funct alimentario. (2022) 13:12896–914. doi: 10.1039/d2fo03366f
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Kopecny, J, Zorec, M, Mrazek, J, Kobayashi, Y, y Marinsek-Logar, R. Butyrivibrio hungatei sp. Nov. y Pseudobutyrivibrio xylanivorans sp. Nov., bacterias productoras de butirato del rumen. Int J Syst Evol Microbiol. (2003) 53:201–9. doi: 10.1099/ijs.0.02345-0
39. Baldin, M, García, D, Zanton, GI, Hao, F, Patterson, AD, y Harvatine, KJ. Efecto del 2-hidroxi-4-(metiltio)butanoato (HMTBa) sobre la grasa láctea, el ambiente ruminal y la biohidrogenación, y los protozoos ruminales en vacas lactantes alimentadas con dietas con mayor riesgo de depresión de la grasa láctea. J Lácteos Sci. (2022) 105:7446–61. doi: 10.3168/jds.2022-21910
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Noftsger, S, St-Pierre, NR y Sylvester, JT. Determinación de la degradabilidad ruminal y los efectos ruminales de tres fuentes de metionina en vacas lactantes. J Lácteos Sci. (2005) 88:223–37. DOI: 10.3168/JDS. S0022-0302(05)72680-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Verhaar, BJH, Prodan, A, Nieuwdorp, M y Muller, M. Microbiota intestinal en la hipertensión y la aterosclerosis: una revisión. Nutrientes. (2020) 12:2982. doi: 10.3390/nu12102982
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Liang, J, Zhang, P, Zhang, R, Chang, J, Chen, L, Wang, G, et al. Respuesta de los microorganismos ruminales al ph durante la hidrólisis anaerobia y la acidogénesis de la biomasa lignocelulósica. Gestión de residuos. (2024) 174:476–86. doi: 10.1016/j.wasman.2023.12.035
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Cheng, J, Zhang, X, Xu, D, Zhang, D, Zhang, Y, Song, Q, et al. Relación entre las diferencias y rasgos microbianos del rumen entre ovejas Hu, ovejas bronceadas y ovejas dorper. J Anim Sci. (2022) 100:skac261. doi: 10.1093/jas/skac261
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Suryawanshi, PR, Badapanda, C, Singh, KM y Rathore, A. Exploración de la diversidad microbiana del rumen y el perfil de enzimas activas de carbohidratos de la cabra bengala negra utilizando un enfoque metagenómico. Anim Biotechnol. (2023) 34:761–74. doi: 10.1080/10495398.2019.1609489
45. Delmont, TO, Quince, C, Shaiber, A, Esen, OC, Lee, ST, Rappe, MS, et al. Las poblaciones fijadoras de nitrógeno de Planctomycetes y Proteobacteria son abundantes en los metagenomas de la superficie de los océanos. Nat Microbiol. (2018) 3:804–13. doi: 10.1038/s41564-018-0176-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Azman, AS, Mawang, CI, Khairat, JE y AbuBakar, S. Actinobacterias: una fuente natural prometedora de agentes anti-biopelícula. Int Microbiol. (2019) 22:403–9. doi: 10.1007/s10123-019-00066-4
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Desriac, F, Jegou, C, Balnois, E, Brillet, B, Le Chevalier, P y Fleury, Y. Péptidos antimicrobianos de Proteobacterias marinas. Mar Drogas. (2013) 11:3632–60. doi: 10.3390/md11103632
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Davis, CK, Webb, RI, Sly, LI, Denman, SE y McSweeney, CS. Aislamiento y estudio de nuevas bacterias degradadoras de fluoroacetato pertenecientes al filo Synergistetes. FEMS Microbiol Ecol. (2012) 80:671–84. doi: 10.1111/j.1574-6941.2012.01338.x
49. Hugenholtz, P, Hooper, SD, y Kyrpides, Carolina del Norte. Enfoque: Sinergistas. Environ Microbiol. (2009) 11:1327–9. doi: 10.1111/j.1462-2920.2009.01949.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Carbone, V, Reilly, K, Sang, C, Schofield, LR, Ronimus, RS, Kelly, WJ, et al. Estructuras cristalinas de las pectinas metilesterasas bacterianas Pme8a y Pmec2 del rumen Butyrivibrio. Int J Mol Sci. (2023) 24:13738. doi: 10.3390/ijms241813738
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Traore, SI, Azhar, EI, Yasir, M, Bibi, F, Fournier, PE, Jiman-Fatani, AA, et al. Descripción de ‘Arabia massiliensis’ gen. Nov., Sp. Nov., ‘Gordonibacter massiliensis’ sp. Nov., y ‘Bacilliculturomica massiliensis’ gen. Nov., Sp. Nov., aislado de un espécimen fecal de una mujer beduina saudita de 50 años. Nuevos microbios, nuevos infectos. (2017) 19:87–90. doi: 10.1016/j.nmni.2017.05.011
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Le Roy, T, Moens, DHE, Van Hul, M, Paquot, A, Pelicaen, R, Regnier, M, et al. Dysosmobacter welbionis es una bacteria comensal humana recientemente aislada que previene la obesidad inducida por la dieta y los trastornos metabólicos en ratones. Tripa. (2022) 71:534–43. doi: 10.1136/gutjnl-2020-323778
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Belanche, A, Arturo-Schaan, M, Leboeuf, L, Yáñez-Ruiz, D, y Martín-García, I. Suplementación temprana con una mezcla natural que contiene cúrcuma, timol y componentes de la pared celular de levadura para optimizar el desarrollo anatómico y microbiológico del rumen y la productividad en cabras lecheras. J Lácteos Sci. (2023) 106:4634–49. doi: 10.3168/jds.2022-22621
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Yi, L, Zhu, J, Li, Q, Guan, X, Cheng, W, Xie, Y, et al. Los tallos y las hojas de Panax notoginseng afectan a la comunidad microbiana y a la función en el ciego de los cerdos duzang. Transl Anim Sci. (2024) 8:txad142. doi: 10.1093/tas/txad142
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Zhang, J, Shi, B, Lu, S, Wang, S, Ren, X, Liu, R, et al. Análisis metagenómico para explorar el potencial de Lactobacillus yoelii FYL1 para mitigar la diarrea bacteriana y los cambios en la microbiota intestinal de yaks juveniles. Microb Pathog. (2024) 186:106496. doi: 10.1016/j.micpath.2023.106496
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Zhang, J, Zhang, C, Yang, X, Li, L, Cao, Y, Zhang, X, et al. Efectos a corto y largo plazo de diferentes tipos de forraje suplementados en terneros lecheros predestete sobre el rendimiento y la producción de leche hasta la primera lactancia. J Lácteos Sci. (2024) 107:7405–22. doi: 10.3168/jds.2023-24244
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. Jiao, J, Wu, J, Zhou, C, He, Z, Tan, Z y Wang, M. Nichos ecológicos y dinámica de ensamblaje de diversos consorcios microbianos en el gastrointestino de cabritos de cabra. ISME J. (2024) 18:WRAE002. doi: 10.1093/ismejo/wrae002
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Abe, K., Ueki, A., Ohtaki, Y., Kaku, N., Watanabe, K. y Ueki, K. (2012). Anaerocella delicata gen. Nov., Sp. Nov., una bacteria estrictamente anaeróbica del filo Bacteroidetes aislada de un reactor metanogénico de granjas ganaderas. J Gen Appl Microbiol. 58, 405–412. doi: 10.2323/jgam.58.405
59. Jiang, B, Qin, C, Xu, Y, Song, X, Fu, Y, Li, R, et al. La multiómica revela el mecanismo del microbioma ruminal y su metaboloma, junto con el metaboloma del huésped, que participan en la regulación de los rasgos de producción de leche en las búfalas lecheras. Microbiol frontal. (2024) 15:1301292. doi: 10.3389/fmicb.2024.1301292
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Carreño, D, Toral, PG, Pinloche, E, Belenguer, A, Yáñez-Ruiz, DR, Hervás, G, et al. Respuestas de la comunidad bacteriana ruminal a DPA, EPA y DHA en ganado vacuno y ovino: un estudio comparativo in vitro. Sci Rep. (2019) 9:11857. doi: 10.1038/s41598-019-48294-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Mi, L, Yang, B, Hu, X, Luo, Y, Liu, J, Yu, Z, et al. El análisis comparativo de la microbiota entre el rumen de oveja y el ciego de conejo proporciona nuevos conocimientos sobre su producción diferencial de metano. Microbiol frontal. (2018) 9:575. doi: 10.3389/fmicb.2018.00575
Resumen de PubMed | Texto completo de Crossref | Google Académico
62. Dewanckele, L, Toral, PG, Vlaeminck, B y Fievez, V. Revisión invitada: papel de los intermedios de biohidrogenación ruminal y los microbios ruminales en la depresión de la grasa láctea inducida por la dieta: una actualización. J Lácteos Sci. (2020) 103:7655–81. doi: 10.3168/jds.2019-17662
Resumen de PubMed | Texto completo de Crossref | Google Académico
63. Yanza, YR, Szumacher-Strabel, M, Lechniak, D, Slusarczyk, S, Kolodziejski, P, Patra, AK, et al. Dieta Coleus amboinicus lour. Disminuye la metanogénesis y la biohidrogenación ruminal, y mejora la calidad de la carne y la composición de ácidos grasos en el músculo longissimus thoracis de corderos. J Anim Sci Biotechnol. (2022) 13:5. doi: 10.1186/s40104-021-00654-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
64. Mohamed, NM, Saito, K, Tal, Y y Hill, RT. Diversidad de bacterias oxidantes de amoníaco aeróbicas y anaeróbicas en esponjas marinas. ISME J. (2010) 4:38–48. doi: 10.1038/ismej.2009.84
Resumen de PubMed | Texto completo de Crossref | Google Académico
65. Sreya, PK, Gaurav, K, Ahmed, S, Sasikala, C, y Ramana, CV. Blastopirellula sediminis sp. Un nuevo miembro de Pirellulaceae aislado de las islas Andamán y Nicobar. Antonie Van Leeuwenhoek. (2023) 116:463–75. doi: 10.1007/s10482-023-01818-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
66. Anderson, CJ, Koester, LR y Schmitz-Esser, S. Las comunidades epiteliales del rumen comparten una microbiota bacteriana central: un metaanálisis de conjuntos de datos de secuenciación del gen de ARNr 16s illumina miSeq. Microbiol frontal. (2021) 12:625400. doi: 10.3389/fmicb.2021.625400
Resumen de PubMed | Texto completo de Crossref | Google Académico
67. Li, L, Wu, L, Yang, L, Liu, C, Li, J y Li, N. Impacto combinado de la materia orgánica, el fósforo, el nitrato y el nitrógeno amoniacal en el proceso de Blackwater. Environ Sci Pollut Res. (2021) 28:32831–43. doi: 10.1007/s11356-021-13050-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
68. Qian, K, Xu, J, Zu, HC y Cong, YY. Avances de la investigación sobre la producción de sulfuro de hidrógeno ruminal en rumiantes. Anim Sci J. (2020) 91:E13349. doi: 10.1111/asj.13349
Resumen de PubMed | Texto completo de Crossref | Google Académico
69. Sarturi, JO, Erickson, GE, Klopfenstein, TJ, Rolfe, KM, Buckner, CD y Luebbe, MK. Impacto de la fuente de azufre en el sulfuro de hidrógeno ruminal y lógica para el concepto de azufre ruminal disponible para la reducción1. J Anim Sci. (2013) 91:3352–9. doi: 10.2527/jas.2012-5626
70. Bailén, M, Bressa, C, Martínez-López, S, González-Soltero, R, Montalvo, LM, San, JC, et al. Características de la microbiota asociadas a una dieta alta en grasas y baja en fibra en adultos sanos. Nuerca delantera. (2020) 7:583608. doi: 10.3389/fnut.2020.583608
Resumen de PubMed | Texto completo de Crossref | Google Académico
71. Chen, H, Ou, R, Tang, N, Su, W, Yang, R, Yu, X, et al. Alternancia de la microbiota intestinal en el síndrome del intestino irritable: un análisis integrado basado en datos de secuenciación de amplicones multicéntricos. J Transl Med. (2023) 21:117. doi: 10.1186/s12967-023-03953-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
72. Lo, CI, Mailhe, M, Ricaboni, D, Vitton, V, Benezech, A, Michelle, C, et al. Massilioclostridium coli gen. Nov., Sp. Nov., un nuevo miembro de la familia Clostridiaceae aislado del colon izquierdo de una mujer de 27 años. Nuevos microbios, nuevos infectos. (2017) 16:63–72. doi: 10.1016/j.nmni.2017.01.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
73. Ignatyeva, O, Tolyneva, D, Kovalyov, A, Matkava, L, Terekhov, M, Kashtanova, D, et al. Christensenella minuta, un nuevo candidato a probiótico de próxima generación: evidencia actual y trayectorias futuras. Microbiol frontal. (2023) 14:1241259. doi: 10.3389/fmicb.2023.1241259
Resumen de PubMed | Texto completo de Crossref | Google Académico
74. Liu, X, Zeng, B, Zhang, J, Li, W, Mou, F, Wang, H, et al. Papel del microbioma intestinal en la modulación de la progresión de la artritis en ratones. Sci Rep. (2016) 6:30594. doi: 10.1038/srep30594
Resumen de PubMed | Texto completo de Crossref | Google Académico
75. Lopetuso, LR, Petito, V, Graziani, C, Schiavoni, E, Paroni, SF, Poscia, A, et al. Microbiota intestinal en la salud, enfermedad diverticular, síndrome del intestino irritable y enfermedades inflamatorias intestinales: tiempo de marcadores microbianos de trastornos gastrointestinales. Dig Dis. (2018) 36:56–65. doi: 10.1159/000477205
Resumen de PubMed | Texto completo de Crossref | Google Académico
76. Xu, B, Yan, Y, Huang, J, Yin, B, Pan, Y y Ma, L. Efecto antidiarreico del extracto de phellodendri de Cortex en ratones relacionado con su modificación de la microbiota intestinal. Farmacia Biomédica. (2020) 123:109720. doi: 10.1016/j.biopha.2019.109720
Resumen de PubMed | Texto completo de Crossref | Google Académico
77. Pluske, JR, Thompson, MJ, Atwood, CS, Bird, PH, Williams, IH y Hartmann, PE. Mantenimiento de la altura de las vellosidades y la profundidad de las criptas, y mejora de la digestión de disacáridos y la absorción de monosacáridos, en lechones alimentados con leche entera de vaca después del destete. Hno. J Nutr. (1996) 76:409–22. doi: 10.1079/bjn19960046
Resumen de PubMed | Texto completo de Crossref | Google Académico
78. Zhong, H, Li, H, Liu, G, Wan, H, Mercier, Y, Zhang, X, et al. El aumento del consumo materno de metionina como análogo hidroxilo promovió el crecimiento intestinal neonatal sin comprometer la homeostasis energética materna. J Anim Sci Biotechnol. (2016) 7:46. doi: 10.1186/s40104-016-0103-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: cabra, metionina ruminal, rendimiento del crecimiento, microbiota ruminal, bioquímica sérica, intestino delgado
Cita: Wang Y, Liu W, Li A, Qubi W, Gong C, Li X, Xing J, Shi H, Li Y, Wang Y y Lin Y (2024) Cambios en el rendimiento del crecimiento, la bioquímica sérica, la fermentación ruminal, la comunidad de microbiota ruminal y el desarrollo intestinal en cabras destetadas durante el tratamiento con metionina protegida del rumen. Frente. Vet. Sci. 11:1482235. doi: 10.3389/fvets.2024.1482235
Editado por:
Shuai Du, Universidad Agrícola de Mongolia Interior, China
Revisado por:
Hengbo Shi, Universidad de Zhejiang, China
Bo Wang, Universidad Agrícola de China, China
Derechos de autor © 2024 Wang, Liu, Li, Qubi, Gong, Li, Xing, Shi, Li, Wang y Lin. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Yaqiu Lin, linyq1999@163.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía