
Efecto de la suplementación dietética del cultivo de levadura Saccharomyces cerevisiae en cabras lactantes

Efecto de la suplementación dietética del cultivo de levadura Saccharomyces cerevisiae en cabras lactantes
 Li Zhang†‡Ge Qin†‡Jiaxue Guo‡Mingding Zhang‘
Li Zhang†‡Ge Qin†‡Jiaxue Guo‡Mingding Zhang‘ Guangxin E
Guangxin E Yongfu Huang
Yongfu Huang Yanguo Han*
Yanguo Han*- Laboratorio Clave de Forraje y Herbrívoro de Chongqing, Centro de Investigación de Ingeniería de Chongqing para la Protección y Utilización de Recursos de Herbívoros, Facultad de Ciencia y Tecnología Animal, Universidad del Suroeste, Chongqing, China
Este estudio fue diseñado para investigar los efectos de la adición de un nuevo cultivo de levadura, Saccharomyces cerevisiae granos secos de sorgo refermentado con solubles (SSDDGS), a las dietas de cabras hembras lactantes sobre el rendimiento de la lactancia y el rendimiento del crecimiento de los corderos. Dividimos 10 cabras negras Dazu lactantes de edad, peso y descendencia similares en dos grupos: uno alimentado con una dieta granulada con 50 g/día de SSDDGS (ET) y el otro sin SSDDGS como control (EC) durante 7 semanas. Monitoreamos los cambios de peso de cada cabra y recolectamos muestras de sangre y leche de ovejas experimentales en momentos específicos para la determinación de la composición hormonal y de la leche. Utilizamos cromatografía líquida de ultra alta resolución y espectrometría de masas en tándem (UPLC-MS/MS) para detectar metabolitos en el suero de ovejas lactantes. Nuestros resultados mostraron que la suplementación con SSDDGS redujo significativamente la pérdida de peso diaria promedio de las cabras durante las semanas 2-4 de lactancia y aumentó los niveles séricos de IGF-1 y prolactina en la semana 4 (p < 0,05). La suplementación con SSDDGS al principio de la lactancia aumentó significativamente el contenido de proteína de leche, lactosa y cenizas (p < 0,05). El análisis UPLC-MS/MS mostró que SSDDGS cambió los niveles de 58 metabolitos en el suero de cabras lactantes. Estos metabolitos estuvieron principalmente involucrados en la vía de señalización de sohingolípidos y en el metabolismo de la cisteína, la metionina y los esfingolípidos. En resumen, los SSDDGS de cultivo de levadura redujeron la pérdida de peso, mejoraron la calidad de la leche y modificaron los perfiles metabólicos en cabras en lactancia temprana, proporcionando información sobre el papel regulador potencial y el mecanismo del cultivo de levadura en cabras hembras lactantes.
1 Introducción
La baja tasa de fertilidad de las cabras plantea un gran desafío para el desarrollo de la industria caprina. Además del número de corderos, la tasa de supervivencia de los corderos también es un rasgo reproductivo extremadamente importante, mientras que la leche materna es la principal fuente de nutrición para los corderos. El sistema digestivo y circulatorio de las cabras es débil en las primeras etapas de la lactancia y experimenta cambios metabólicos significativos y cambios en la asignación de energía, incluida la pérdida de peso, el aumento de la ingesta de glucosa y la inmunidad comprometida (1). La ingesta insuficiente o desequilibrada de nutrientes de los piensos conducirá fácilmente a un deterioro del estado de salud de las cabras, que no puede satisfacer las necesidades de crecimiento y desarrollo rápidos de los corderos, lo que provoca un crecimiento lento e incluso la muerte del cordero (2). Por lo tanto, es fundamental diseñar un alimento adecuado para cabras lactantes para mejorar el rendimiento de la lactancia en las primeras etapas de la lactancia.
El cultivo de levadura (YC) consiste principalmente en metabolitos extracelulares de levadura, medio modificado después de la fermentación y una pequeña cantidad de células de levadura inactivas. La suplementación con YC en la dieta de los rumiantes puede mejorar el entorno del pH del rumen, la eficiencia de la digestibilidad de la fibra bruta y el rendimiento del crecimiento (3, 4). El YC puede aumentar la puntuación corporal, la producción y la calidad de la leche en la etapa temprana de lactancia de las vacas lecheras y las cerdas (5-8). Los granos secos de destilería de sorgo refermentado con solubles (SSDDGS) de Saccharomyces cerevisiae son un nuevo YC que se produce por refermentación del sustrato de los granos de destilería, lo que aumentó en gran medida el rendimiento y redujo el costo de la producción de cultivo de levadura. La suplementación con SSDDGS en la dieta de los cerdos mejoró el rendimiento de la lactancia de las cerdas (9). Sin embargo, el estudio sobre el efecto de los nuevos YC SSDDGS en el rendimiento de la lactancia de las cabras sigue siendo incierto.
Por lo tanto, este estudio tiene como objetivo determinar el efecto de la suplementación de un nuevo cultivo de levadura SSDDGS en la dieta de cabras lactantes sobre la pérdida de peso de cabras hembras, el rendimiento de la lactancia, las hormonas y los metabolitos.
2 Materiales y métodos
2.1 Manipulación de animales y recogida de muestras
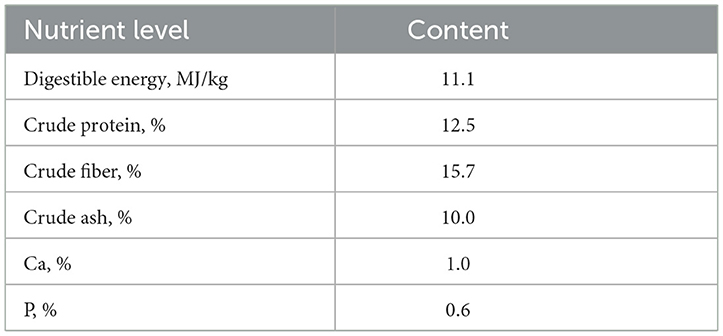
Todas las pruebas y el manejo de animales se adhirieron a las regulaciones establecidas por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Southwestern (IACUC-20210515-05). El experimento de alimentación se llevó a cabo en Tengda Animal Husbandry, Inc. en Chongqing, China. Diez cabras lactantes que acababan de dar a luz con pesos comparables se dividieron aleatoriamente en dos grupos, y cada cabra hembra con corderos gemelos. Un grupo sirvió como grupo EC y fue alimentado con una dieta granulada de valor completo (906) obtenida de Pizhou Xiaohe Technology Development Ltd, y el nivel nutricional de las dietas se muestra en la Tabla 1. El otro grupo constituido por el grupo ET recibió una dieta peletizada de valor completo (906) suplementada con 50 g/d de levadura nueva, cultivo de Saccharomyces cerevisiae, granos secos de destilería de sorgo refermentado con solubles (SSDDGS). Ambos grupos de cabras hembras lactantes recibieron el mismo peso de alimento dos veces al día a las 7 a.m. y a las 3 p.m.
El período experimental inició después de 1 semana de prealimentación, y el período de alimentación formal fue de 7 semanas, de las semanas 2 a 8 de lactancia. Se recogieron muestras de sangre periférica y leche de cabras lactantes en las semanas 4 y 8 de lactancia. Las muestras de sangre se colocaron durante la noche a 4 °C y luego se centrifugaron a 3.000 rpm durante 10 min, y se recolectó la muestra de suero. El peso de las cabras lactantes y de los corderos descendientes se registró a las 4 y 8 semanas de lactancia, y el ADL (pérdida de peso diaria media) de las cabras hembras y el GMD de los corderos descendientes a las 2-4 y 2-8 semanas de lactancia.
2.2 Determinación de las hormonas y de la composición de la leche
Estas concentraciones séricas de hormona liberadora de hormona de crecimiento (GHRH), somatostatina (SS), factor de crecimiento similar a la insulina-1 (IGF-1), prolactina (PRL), hormona de crecimiento (GH) en cabras lactantes se cuantificaron en las semanas 4 y 8 de lactancia mediante radioinmunoensayo en el Instituto Chino-Británico de Biotecnología en Beijing, China. Además, estos componentes de la leche, como la grasa láctea, la proteína, la lactosa, el contenido de cenizas y la materia seca, se probaron utilizando un analizador automático de la composición de la leche (LACTOSCAN MCC 50, Milkotronic, Bulgaria).
2.3 Análisis metabolómico ampliamente específico del suero de cabra
Los metabolitos del suero de cabra lactante se determinaron mediante UPLC-MS/MS, según los estudios previos (10-12). En resumen, 50 μL de la muestra y 300 μL de una solución de extracción (que comprende CAN y metanol en una proporción de 1:4; V/V) que contenían patrones internos se centrifugaron a 12.000 rpm a 4 °C y luego se utilizaron los sobrenadantes para el análisis UPLC-MS. Las muestras se analizaron utilizando un sistema UPLC que presentaba una columna UPLC (2,1 mm × 100 mm, 1,8 μm; Aguas ACQUITY UPLC HSS T3 C18). La temperatura de la columna se mantuvo a 40 °C y el caudal se fijó en 0,4 mL/min. El sistema de disolventes estaba formado por A (agua con ácido fórmico al 0,1%) y B (acetonitrilo con ácido fórmico al 0,1%). El programa de gradiente del sistema fue el siguiente: 95:5 V/V a 0 min; 10:90 V/V a 11,0 min, 10:90 V/V a 12,0 min; 95:5 V/V a 12,1 min; y 95:5 V/V a 14.0 min. El volumen de inyección fue de 2 μL. Para garantizar la estabilidad del sistema, se introdujo una muestra de control de calidad cada 10 muestras de prueba.
El análisis de las muestras se llevó a cabo utilizando un sistema MS/MS equipado con una interfaz ESI Turbo Ion-Spray, que funcionaba tanto en modo de iones positivos como negativos. Los parámetros de operación de la fuente ESI fueron los siguientes: temperatura de la fuente, 500 °C; voltaje de pulverización de iones (IS), 5.500 V (positivo) y -4.500 V (negativo); y los gases de fuente iónica I, II y gas de cortina se mantuvieron a 55, 60 y 25.0 psi, respectivamente.
2.4 Análisis estadístico
Los datos de ADL de cabra hembra, GMD de corderos, concentraciones de hormonas, composición de la leche se analizaron utilizando un modelo lineal general en el software GraphPad Prism8.0.1 (San Diego, CA, EE. UU.), con el tratamiento y el tiempo como factores fijos, y las muestras individuales como factor aleatorio. La interacción entre el tratamiento y el tiempo se incluyó en el modelo. Estas diferencias entre las medias se analizaron mediante comparaciones múltiples de Duncan.
El análisis estadístico multivariado incluyó análisis de componentes principales (PCA) no supervisado y análisis discriminante de mínimos cuadrados parciales ortogonales supervisado (OPLS-DA). La importancia de la variable p ≤ 0,05, proyección (VIP) ≥ 1 y log2FC (fold-change) ≤ 0,67 o log2FC ≥ 1,5 representó una diferencia significativa en los metabolitos entre el grupo ET y el grupo EC. Los metabolitos identificados se anotaron utilizando la base de datos de compuestos KEGG, y las vías KEGG se utilizaron para el análisis del enriquecimiento del conjunto de metabolitos. Un valor de p obtenido de la prueba hipergeométrica de < 0,05 indicó significación biológica. P < 0,05 se consideró estadísticamente significativo. Los resultados se expresan como la media ± el error estándar (SEM).
3 Resultados
3.1 Cambios en el peso de las cabras lactantes y de los corderos descendientes
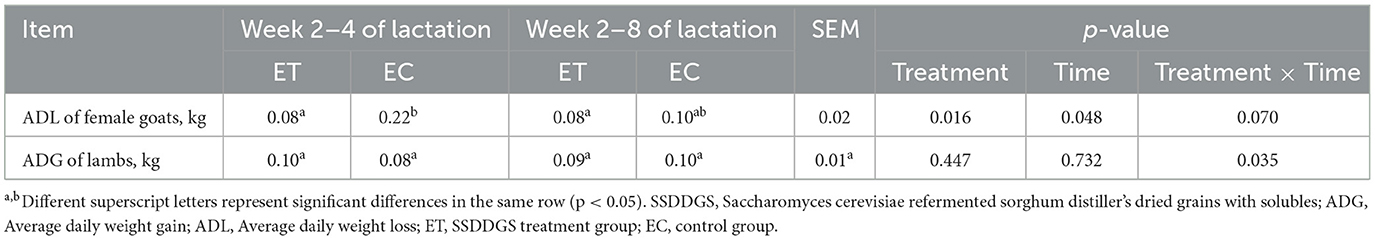
La pérdida de peso diaria promedio (AVD) de las cabras lactantes en el grupo de tratamiento experimental (ET) fue significativamente menor durante la semana 2-4 de lactancia que en el grupo de control experimental (EC) (Tabla 2, p < 0,05). No hubo diferencia significativa en la ganancia de peso diaria promedio (GMD) de los corderos descendientes entre los grupos ET y EC durante las semanas 2-4 o 2-8 de lactancia (Tabla 2, p > 0,05).
3.2 Cambios en las hormonas séricas en cabras lactantes
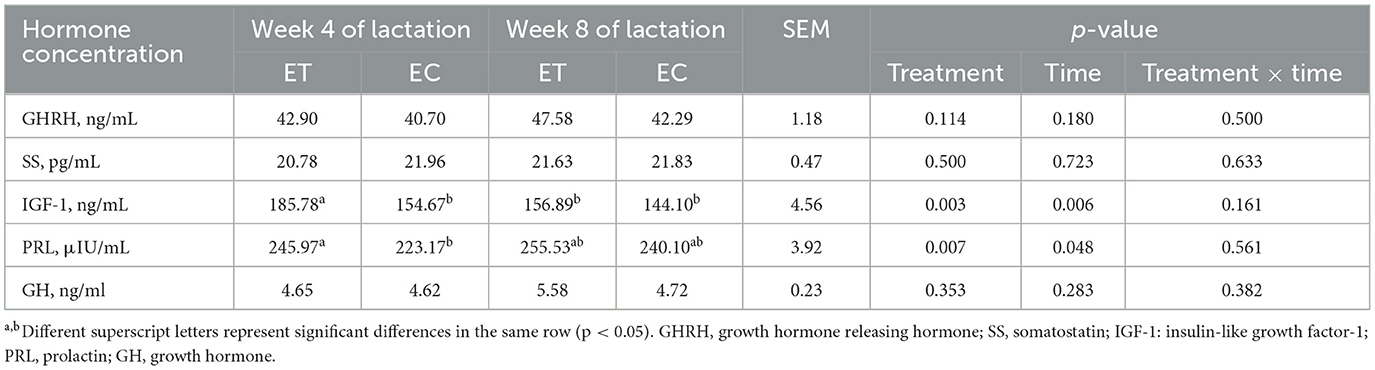
La concentración sérica de factor de crecimiento similar a la insulina-1 (IGF-1) o prolactina (PRL) en el grupo ET fue significativamente mayor que en el grupo EC en la semana 4 de lactancia (Tabla 3, p < 0,05). Además, en el grupo ET, la concentración de IGF-1 en la semana 8 de lactancia fue significativamente menor que en la semana 4 de lactancia (Tabla 3, p < 0,05).
3.3 Efectos de las SSDDGS en la calidad de la leche en cabras lactantes
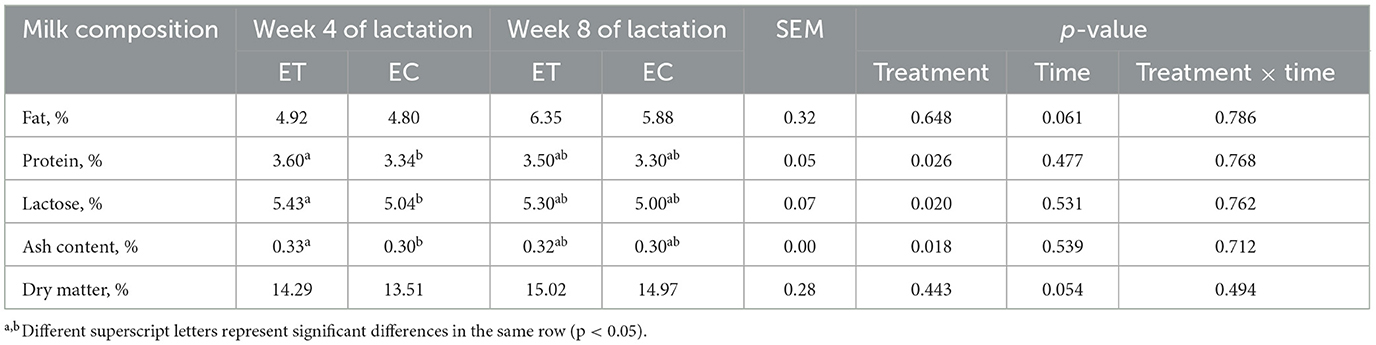
La composición de la leche fue más abundante en el grupo ET en comparación con el grupo EC en la semana 4 de lactancia. El contenido de proteína láctea, lactosa láctea o cenizas en el grupo ET fue significativamente mayor que el del grupo EC en la semana 4 de lactancia (Tabla 4, p < 0,05).
3.4 Efectos de las SSDDGS sobre los metabolitos de las cabras
Los efectos de SSDDGS sobre los metabolitos de las cabras se analizaron mediante cromatografía líquida de ultra alta resolución con espectrometría de masas en tándem (UPLC-MS/MS). El gráfico de puntuación del análisis de componentes principales (PCA) (Figura 1A) reveló que las tasas de interpretación del modelo para PC1 y PC2 fueron del 24,96% y del 16,03%, respectivamente, con una clara separación de muestras entre los dos grupos. Los resultados de OPLS-DA reflejaron los obtenidos de PCA, con un componente principal de predicción en los grupos de ET y EC que representó el 17,3% de la tasa de interpretación del modelo (Figura 1B). Además, el modelo OPLS-DA mostró los parámetros predictivos de Q2 = 0,62 y R2Y = 0,991, lo que indica la estabilidad y fiabilidad del modelo (Figura 1C).
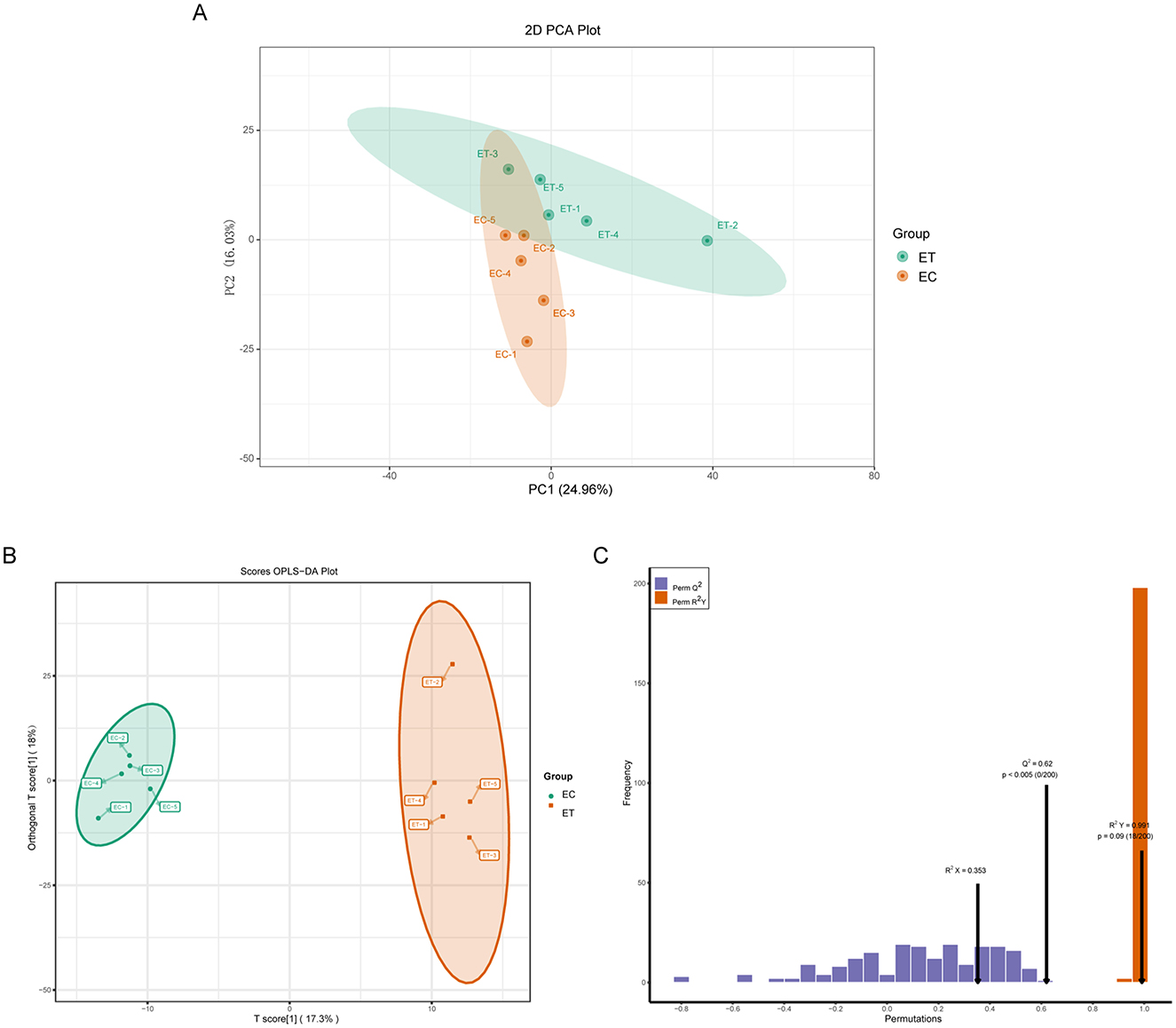
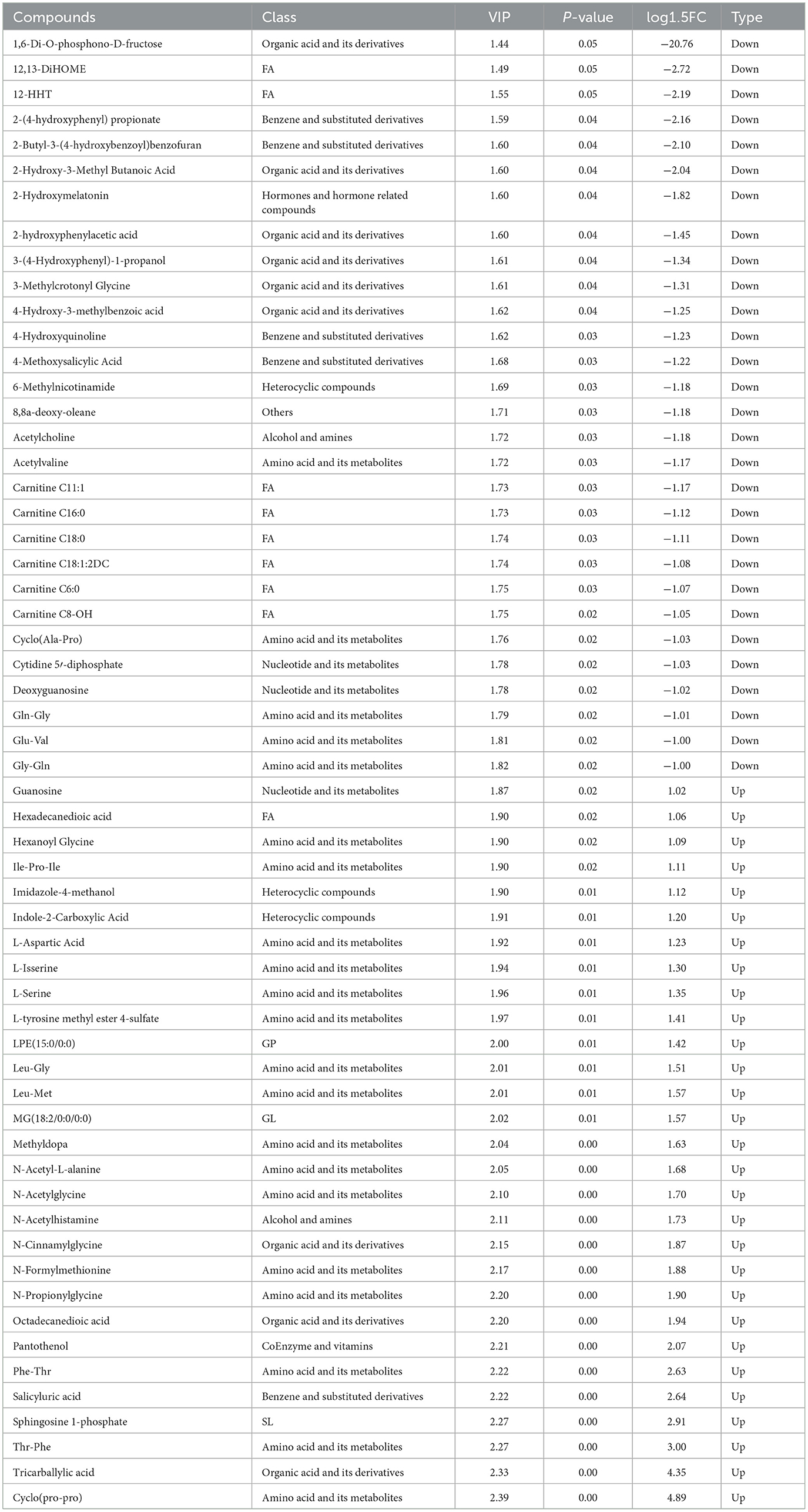
La identificación de 60 metabolitos significativamente diferentes en el grupo ET en comparación con el grupo EC (Tabla 5, Figura 2A). Entre estos, 31 metabolitos fueron regulados a la baja, mientras que 29 fueron regulados al alza (Figura 2B). Los diversos tipos de metabolitos incluyen L-leucilglicina, ácido aspártico, esfingosina 1-fosfato, cinamil glicina, serina y hexanoil glicina.
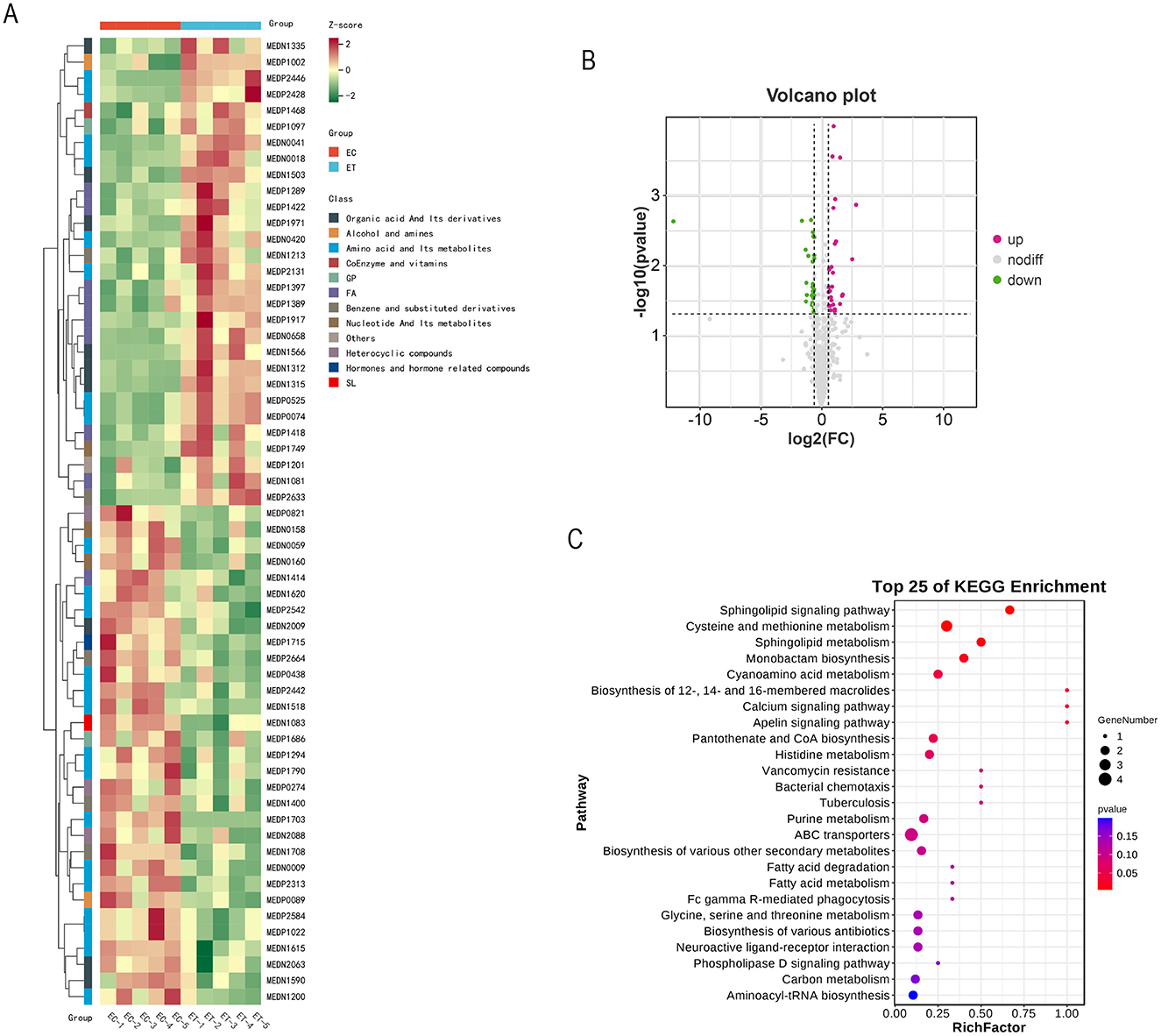
Las vías metabólicas de estos 60 metabolitos diferentes se analizaron mediante análisis de enriquecimiento KEGG. Los resultados revelaron la implicación de 25 vías metabólicas, entre las que se encuentran la con el valor p más cercano a 0 y el mayor número de metabolitos diferencialmente significativos como la vía de soligolípidos sianaling, el metabolismo de cisteína y metionina y el metabolismo de esfingolípidos (Figura 2C).
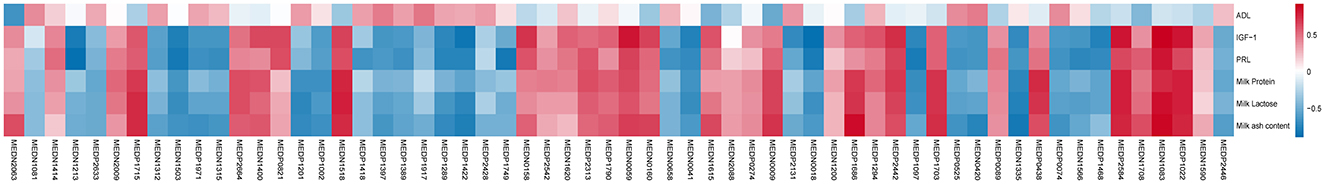
Mientras tanto, para analizar más a fondo las posibles correlaciones entre los datos fenotípicos de la cabra hembra y los metabolitos diferenciales en suero, realizamos un análisis de correlación de Pearson (Figura 3). La L-leucilglicina, el ácido aspártico, la esfingosina y el 1-fosfato mostraron correlaciones positivas con el índice bioquímico sérico (IGF-1, PRL) y la composición de la leche (contenido de proteínas, lactosa y cenizas) de cabras hembras (P < 0,05). Además, los monoglicéridos, monoglicéridos y aminoácidos (AA) como la cinamil glicina, la serina y la hexanoil glicina tuvieron correlaciones significativas negativas con el índice bioquímico sérico (IGF-1, PRL) y la composición de la leche (contenido de proteínas, lactosa y cenizas) de las cabras hembras (P < 0,05).
4 Discusión
El cultivo de levadura contiene una gran cantidad de microorganismos beneficiosos y moléculas pequeñas de fácil digestión, lo que es muy beneficioso para mejorar la salud animal y el rendimiento de la producción. Durante la lactancia temprana, las cabras hembras experimentan un mayor gasto de energía, una mayor producción de ácidos grasos volátiles y una disminución del peso corporal (1). La suplementación con YC en la dieta puede mejorar la digestibilidad de los nutrientes y promover el crecimiento animal al aumentar la diversidad y abundancia de microorganismos ruminales (13-15).
En este estudio, la suplementación con SSDDGS en la dieta redujo significativamente las AVD de las cabras lactantes durante la lactancia temprana. De manera similar, Song et al. informaron que la incorporación de YC en la dieta mejoró la comunidad microbiana del rumen y aumentó la GMD de los corderos (16). Además, en este estudio, la suplementación con SSDDGS en la dieta aumentó significativamente la concentración sérica de IGF-1 durante la lactancia temprana. El IGF-1 es una hormona endocrina producida principalmente por el hígado y promueve el crecimiento y desarrollo animal, que está involucrado en el metabolismo de los lípidos, la secreción de insulina y la absorción de glucosa (17). Estos resultados indican que la suplementación dietética de SSDDGS redujo la pérdida de peso de las cabras lactantes durante la lactancia temprana a través del papel del IGF-1.
La suplementación de YC en la dieta puede mejorar eficazmente el rendimiento de la lactancia animal. En este estudio, la suplementación con SSDDGS en la dieta aumentó significativamente el contenido de proteína de la leche, lactosa láctea y cenizas durante la lactancia temprana, aunque la suplementación con SSDDGS en la dieta no aumentó la GMD de los corderos en lactancia, que fue un indicador importante de la producción de leche en cabras hembras. Estos estudios son consistentes con estudios previos. La suplementación con YC aumentó el contenido de proteína láctea y redujo la incidencia de mastitis en las vacas (18). De manera similar, el contenido de proteína y grasa de la leche en las vacas aumentó después de la suplementación con YC, particularmente durante la lactancia temprana, sin afectar la producción total de leche (19). La suplementación con YC o Saccharomyces cerevisiae en la dieta aumentó la producción de leche y la producción de grasa en cabras lactantes (20, 21). Además, en este estudio, la suplementación con SSDDGS en la dieta aumentó significativamente los niveles séricos de PRL durante la lactancia temprana. La PRL es vital para mantener la función de la glándula mamaria, la producción de leche y la calidad de la leche en rumiantes. Chen et al. encontraron que la PRL aumentaba la síntesis de proteínas de la leche al estimular el metabolismo de las células epiteliales mamarias (22). Estos resultados sugieren que la suplementación con YC en la dieta mejoró la calidad de la leche de las cabras lactantes durante la lactancia temprana a través del papel de la PRL.
La suplementación con SSDDGS en la dieta jugó un papel crucial en la inflamación y el sistema inmunológico de los organismos según el análisis del metaboloma. La suplementación con postbióticos de fermentación de Saccharomyces cerevisiae en terneros aumentó la resistencia a la enfermedad respiratoria bovina a través de las respuestas inmunes sistémicas y mucosas (23). El cultivo de Saccharomyces cerevisiae puede reducir el número de células somáticas en la leche y mejorar la capacidad antioxidante en vacas sometidas a estrés térmico (24). Durante la lactancia, hay una gran demanda de glucosa para la síntesis de leche y la producción de ácidos grasos. Sin embargo, el consumo excesivo de energía puede provocar mastitis, y la producción excesiva de ácidos grasos libres da lugar a la formación de cuerpos cetónicos, lo que puede reducir la inmunidad de las cabras hembras (25). A partir de los resultados de este estudio, observamos una regulación positiva significativa de las carnitina C11:1 y carnitina C18:1:2DC en el grupo SSDDGS, que están involucradas en el metabolismo de los ácidos grasos y las vías de degradación. La abundancia de expresión de esfingosina 1-fosfato se reguló significativamente a la baja e involucró en la fagocitosis mediada por Fc gamma R y la vía de la tuberculosis. Informes previos han demostrado que los suplementos de carnitina aumentan el flujo de metabolitos a través de la piruvato carboxilasa, aumentando así la secreción de insulina y la producción de glucosa hepática (26). En las mitocondrias aisladas de hígado de cerdo alimentado con carnitina, se encontró que la cantidad de piruvato carboxilasa mitocondrial se triplicó. Los cerdos alimentados con carnitina son más capaces de utilizar la grasa para obtener energía, transferir carbono a la síntesis de aminoácidos y ahorrar aminoácidos de cadena ramificada utilizados para la síntesis de proteínas (27). De manera similar, otro estudio encontró que los altos niveles de carnitina pueden mejorar el metabolismo de los lípidos en las ovejas, alterando así el rendimiento de la lactancia (28). Sin embargo, no ha habido investigaciones sobre la suplementación de carnitina en la dieta de las cabras, y estos informes sugieren que la carnitina puede ser la principal sustancia que mejora el rendimiento de la lactancia de las cabras.
Además, la desoxiguanosina, Gly Leu, Phe Thr y esfingosina 1-fosfato se correlacionaron significativamente positivamente con los niveles de IGF-1 y PRL. En relación con esto, el metabolismo de las purinas, los transportadores ABC, la vía de señalización del calcio y la vía de señalización de la apelina se enriquecieron significativamente. Luo et al. investigaron los efectos de la harina de soja fermentada (FSBM) rica en ligandos glicosídicos de isoflavonas a diferentes niveles en ovejas desde el final de la gestación hasta el principio de la lactancia. La alimentación de ovejas con FSBM6 redujo las concentraciones de peróxido de hidrógeno y desoxiguanosina en la placenta, mejoró la capacidad antioxidante tanto de la madre como de la placenta, y mejoró las hormonas séricas y la calidad de la leche (29). La esfingosina quinasa (SK) cataliza la formación de esfingosina-1-fosfato (S1P), que desempeña un papel esencial en el crecimiento y la supervivencia celular (30). Döll et al. descubrieron que la prolactina (PRL) activa las isoformas SK-1 pero no las SK-2 en una línea celular de cáncer de mama humano (MCF7). El retraso en la activación de SK-1 es el resultado de la regulación positiva de la expresión de ARNm y proteínas y se debe al aumento de la actividad del promotor de SK-1 por un mecanismo que involucra la activación de STAT5, así como la proteína quinasa C y las proteínas quinasas clásicas activadas por mitógenos. El retraso en la activación de SK-1 es el resultado de la regulación al alza de la expresión de ARNm y proteínas debido al aumento de la actividad del promotor de SK-1 e implica la activación de STAT5, así como la proteína quinasa C y la proteína quinasa clásica activada por mitógenos (31). Esto sugiere que es probable que la esfingosina-1-fosfato desempeñe un papel importante en el rendimiento de la lactancia.
5 Conclusion
Este estudio demuestra que la suplementación dietética de cultivo de levadura SSDDGS redujo la pérdida de peso y mejoró la calidad de la leche de las cabras hembras en la etapa temprana de la lactancia. Además, las SSDDGS pueden desempeñar un papel vital en el metabolismo energético y las respuestas inmunitarias, principalmente a través de la modulación de las vías metabólicas de los ácidos biliares y la cafeína. Este estudio nos ayudará a comprender mejor los efectos y el mecanismo de un nuevo cultivo de levadura SSDDGS en cabras lactantes.
Declaración de disponibilidad de datos
Los conjuntos de datos presentados en este estudio se pueden encontrar en repositorios en línea. Los nombres del repositorio o repositorios y el número de acceso se pueden encontrar en el artículo/material complementario.
Declaración ética
Los estudios en animales fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Southwestern. Los estudios se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
LZ: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. GQ: Conceptualización, Análisis formal, Supervisión, Redacción – revisión y edición. JG: Análisis formal, Supervisión, Redacción – revisión y edición. MZ: Supervisión, Redacción – revisión y edición. GE: Supervisión, Redacción – revisión y edición. YHa: Supervisión, Redacción – revisión y edición. YHu: Conceptualización, Análisis formal, Obtención de fondos, Supervisión, Redacción – revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue apoyada por el Proyecto Especial de Innovación Científica y Tecnológica de Chongqing (No. CSTB2022TIAD-CUX0013; cstc2019jscx-gksbX0135) y el Proyecto de Colaboración Estratégica del Gobierno Popular Municipal de Chongqing y la Academia China de Ciencias Agrícolas, y Fondos de Investigación Fundamental para las Universidades Centrales (SWU-XDJH202301).
Reconocimientos
El equipo de investigación desea agradecer a Tengda Animal Husbandry, Inc. por la alimentación de los animales y la recolección de muestras durante esta investigación.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Zamuner F, Cameron AWN, Carpenter EK, Leury BJ, DiGiacomo K. Respuestas endocrinas y metabólicas a las infusiones de glucosa, insulina y adrenocorticotropina en cabras lecheras de lactancia temprana de alto y bajo rendimiento de leche. J Lácteos Sci. (2020) 103:12045–58. doi: 10.3168/jds.2020-18625
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Ataollahi F, McGrath S, Friend M, Dutton G, Peters A, Bhanugopan M. Evaluación del efecto de la suplementación con calcio, magnesio y sodio de las ovejas merinas en el crecimiento de sus corderos. Aust Vet J. (2023) 101:391–96. doi: 10.1111/avj.13274
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Razzaghi A, Malekkhahi M, Brito AF. Rendimiento de la lactancia, producción de grasa láctea y respuestas de digestibilidad de nutrientes a la adición de melaza líquida o cultivo de levadura en vacas lecheras alimentadas con maíz superacondicionado. J Lácteos Sci. (2023) 106:6080–93. doi: 10.3168/jds.2022-22768
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Zhang X, Dong X, Wanapat M, Shah AM, Luo X, Peng Q, et al. Patrón de pH ruminal, características de fermentación y bacterias relacionadas en respuesta a la suplementación con levadura viva (Saccharomyces cerevisiae) en ganado vacuno. Anim Biosci. (2022) 35:184–95. doi: 10.5713/ab.21.0200
5. Chen H, Liu S, Li S, Li D, Li X, Xu Z, et al. Effects of yeast culture on growth performance, immune function, antioxidant capacity and hormonal profile in Mongolian ram lambs. Front Vet Sci. (2024) 11:1424073. doi: 10.3389/fvets.2024.1424073
6. Kim SW, Duarte ME. Saccharomyces yeast postbiotics supplemented in feeds for sows and growing pigs for its impact on growth performance of offspring and growing pigs in commercial farm environments. Anim Biosci. (2024) 37:1463–73. doi: 10.5713/ab.23.0467
7. Verdier-Metz I, Delbès C, Bouchon M, Rifa E, Theil S, Chaucheyras-Durand F, et al. Dietary Live Yeast Supplementation Influence on Cow’s Milk, Teat and Bedding Microbiota in a Grass-Diet Dairy System. Microorganisms. (2023) 11:673. doi: 10.3390/microorganisms11030673
8. Zhao Y, Wang Q, Zhou P, Li Z, Zhong W, Zhuo Y, et al. Effects of yeast culture supplementation from late gestation to weaning on performance of lactating sows and growth of nursing piglets. Animal. (2022) 16:100526. doi: 10.1016/j.animal.2022.100526
9. Song D, Li X, Cheng Y, Wu G, Xiao X, Wang F, et al. Effects of supplementing sow diets with Saccharomyces cerevisiae refermented sorghum dried distiller’s grains with solubles from late gestation to weaning on the performance of sows and progeny. J Anim Sci. (2017) 95:2025–31. doi: 10.2527/jas.2017.1438
10. Qin G, Zhang L, Guo J, Fang S, Zeng Y, Huang Y, et al. Combined proteomic and metabolomic analysis reveals comprehensive regulation of somatostatin DNA vaccine in goats. Int J Mol Sci. (2024) 25:6888. doi: 10.3390/ijms25136888
11. Wang H, Li B, Li A, An C, Liu S, Zhuang Z. Integrative metabolomics, enzymatic activity, and gene expression analysis provide insights into the metabolic profile differences between the slow-twitch muscle and fast-twitch muscle of Pseudocaranx dentex. Int J Mol Sci. (2024) 25:6131. doi: 10.3390/ijms25116131
12. Xiao Y, Li X, Zeng X, Wang H, Mai Q, Cheng Y, et al. Low ω-6/ω-3 ratio high-fat diet improves rat metabolism via purine and tryptophan metabolism in the intestinal tract, while reversed by inulin. J Agric Food Chem. (2019) 67:7315–24. doi: 10.1021/acs.jafc.9b02110
13. Liu S, Yang L, Zhang Y, Chen H, Li X, Xu Z, et al. Review of yeast culture concerning the interactions between gut microbiota and young ruminant animals. Front Vet Sci. (2024) 11:1335765. doi: 10.3389/fvets.2024.1335765
14. Ogbuewu IP, Mbajiorgu CA. Meta-analysis of Saccharomyces cerevisiae on enhancement of growth performance, rumen fermentation and haemato-biochemical characteristics of growing goats. Heliyon. (2023) 9:e14178. doi: 10.1016/j.heliyon.2023.e14178
15. Zhang X., Liu T., Hou X., Hu C., Zhang L., Wang S., et al. Multi-channel metabolomics analysis identifies novel metabolite biomarkers for the early detection of fatty liver disease in dairy cows. Cells. (2022) 11:2883. doi: 10.3390/cells11182883
16. Song B, Wu T, You P, Wang H, Burke JL, Kang K, et al. Dietary supplementation of yeast culture into pelleted total mixed rations improves the growth performance of fattening lambs. Front Vet Sci. (2021) 8:657816. doi: 10.3389/fvets.2021.657816
17. Kasprzak A. Insulin-like growth factor 1 (IGF-1) signaling in glucose metabolism in colorectal cancer. Int J Mol Sci. (2021) 22:6434. doi: 10.3390/ijms22126434
18. Nocek JE, Holt MG, Oppy J. Effects of supplementation with yeast culture and enzymatically hydrolyzed yeast on performance of early lactation dairy cattle. J Dairy Sci. (2011) 94:4046–56. doi: 10.3168/jds.2011-4277
19. Kalmus P, Orro T, Waldmann A, Lindjärv R, Kask K. Effect of yeast culture on milk production and metabolic and reproductive performance of early lactation dairy cows. Acta Vet Scand. (2009) 51:32. doi: 10.1186/1751-0147-51-32
20. Mavrommatis A, Mitsiopoulou C, Christodoulou C, Karabinas D, Nenov V, Zervas G., et al. Dietary supplementation of a live yeast product on dairy sheep milk performance, oxidative and immune status in peripartum period. J Fungi (Basel). (2020) 6:334. doi: 10.3390/jof6040334
21. Ogbuewu IP, Mbajiorgu CA. Corrigendum: Meta-analytic effect of Saccharomyces cerevisiae on dry matter intake, milk yield and components of lactating goats. Front Vet Sci. (2023) 10:1144334. doi: 10.3389/fvets.2023.1144334
22. Chen Z, Luo J, Zhang C, Ma Y, Sun S, Zhang T, et al. Mechanism of prolactin inhibition of miR-135b via methylation in goat mammary epithelial cells. J Cell Physiol. (2018) 233:651–62. doi: 10.1002/jcp.25925
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Maina TW, McDonald PO, Rani Samuel BE, Sardi MI, Yoon I, Rogers A, et al. La alimentación con productos postbióticos de fermentación de Saccharomyces cerevisiae altera la función inmune y el transcriptoma pulmonar de terneros antes del destete con una coinfección viral-bacteriana experimental. J Lácteos Sci. (2024) 107:2253–67. doi: 10.3168/jds.2023-23866
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Du D, Feng L, Chen P, Jiang W, Zhang Y, Liu W, et al. Efectos de los cultivos de Saccharomyces cerevisiae sobre el rendimiento y el rendimiento inmunológico de las vacas lecheras durante el estrés térmico. Frente Vet Sci. (2022) 9:851184. doi: 10.3389/fvets.2022.851184
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Bouvier-Muller J, Allain C, Enjalbert F, Tabouret G, Portes D, Caubet C, et al. Respuesta a la restricción energética inducida por la dieta en ovejas lecheras seleccionadas de manera divergente por resistencia o susceptibilidad a la mastitis. J Lácteos Sci. (2016) 99:480–92. doi: 10.3168/jds.2015-9785
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Carlson DB, McFadden JW, D’Angelo A, Woodworth JC, Drackley JK. La L-carnitina dietética afecta el metabolismo de los nutrientes periparto y la lactancia en vacas multíparas. J Lácteos Sci. (2007) 90:3422–41. doi: 10.3168/jds.2006-811
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Owen KQ, Ji H, Maxwell CV, Nelssen JL, Goodband RD, Tokach MD, et al. La L-carnitina dietética suprime la actividad deshidrogenasa del ácido cetogénico de cadena ramificada mitocondrial y mejora la acumulación de proteínas y las características de la canal de los cerdos. J Anim Sci. (2001) 79:3104–12. doi: 10.2527/2001.79123104x
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Bai U, Su X, Zheng Z, Zhang L, Ma Y, Dou Y, et al. Análisis metabolómico comparativo de la leche ovina de han de cola pequeña y DairyMeade. Eur Food Res Technol. (2021) 247:2869–76. doi: 10.1007/s00217-021-03840-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Luo Z, Yu S, Zhu Y, Zhang J, Xu W, Xu J. Efecto de varios niveles de harina de soja fermentada enriquecida con aglicona con isoflavonas sobre el estado redox, las hormonas séricas y la calidad de la leche en ovejas. S Afr J Anim Sci. (2018) 48:673–82. doi: 10.4314/sajas.v48i4.9
30. Glueck M, Lucaciu A, Subburayalu J, Kestner RI, Pfeilschifter W, Vutukuri R, et al. Metabolitos atípicos de esfingosina-1-fosfato: implicaciones biológicas de la longitud de la cadena de alquilo. Pflugers Archiv-Eur J Physiol. (2024). doi: 10.1007/s00424-024-03018-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Döll F, Pfeilschifter J, Huwiler A. La prolactina regula al alza la expresión y la actividad de la esfingosina quinasa-1 en la línea celular de cáncer de mama humano MCF7 y desencadena una mayor proliferación y migración. Endocr Relat Cáncer. (2007) 14:325–35. doi: 10.1677/ERC-06-0050
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: cultivo de levaduras, lactancia, peso, metabolismo, cabras
Cita: Zhang L, Qin G, Guo J, Zhang M, E G, Huang Y y Han Y (2024) Efecto de la suplementación dietética del cultivo de levadura Saccharomyces cerevisiae en cabras lactantes. Frente. Vet. Sci. 11:1482800. doi: 10.3389/fvets.2024.1482800
Recibido: 18 de agosto de 2024; Aceptado: 25 de octubre de 2024;
Publicado: 12 de noviembre de 2024.
Editado por:
Junshi Shen, Universidad Agrícola de Nanjing, China
Revisado por:
Yanfeng Xue, Universidad Agrícola de Anhui, China
Bahram Chachar, Universidad de Agricultura, Agua y Ciencias Marinas de Lasbela, Pakistán
Derechos de autor © 2024 Zhang, Qin, Guo, Zhang, E, Huang y Han. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Yanguo Han, hyg2015@swu.edu.cn
†Estos autores han contribuido igualmente a este trabajo
‡ORCID: Li Zhang orcid.org/0009-0009-5526-5376
Ge Qin orcid.org/0009-0007-2810-2358
Jiaxue Guo orcid.org/0009-0000-9722-5262
Mingding Zhang orcid.org/0009-0006-4474-0395
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía