
Las dietas bajas en proteínas suplementadas con glicina mejoran el rendimiento del crecimiento de los cerdos

Las dietas bajas en proteínas suplementadas con glicina mejoran el rendimiento del crecimiento de los cerdos y la calidad de la carne: un análisis metabolómico no dirigido
 Shengwang Jiang1,2
Shengwang Jiang1,2 Wei Quan1
Wei Quan1 Jie Luo1Aihua Lou1
Jie Luo1Aihua Lou1 Xihong Zhou3
Xihong Zhou3 Fengna Li3*Qingwu W. Shen1*
Fengna Li3*Qingwu W. Shen1*- 1Facultad de Ciencia y Tecnología de los Alimentos, Universidad Agrícola de Hunan, Changsha, Hunan, China
- número arábigoFacultad de Ciencia Animal, Universidad de Xichang, Xichang, Sichuan, China
- 3Instituto de Agricultura Subtropical, Academia China de Ciencias, Changsha, Hunan, China
Con el fin de mejorar la calidad de la carne, los cerdos fueron alimentados con una dieta normal (ND), una dieta baja en proteínas (LPD) y una LPD suplementada con glicina (LPDG). Los análisis químicos y metabolómicos mostraron que la LPD aumentó la deposición de IMF y las actividades de GPa y PK, pero disminuyó el contenido de glucógeno, las actividades de CS y CcO, y la abundancia de acetil-CoA, tirosina y sus metabolitos en el músculo. El LPDG promovió la transición de la fibra muscular del tipo II al tipo I, aumentó la síntesis de múltiples aminoácidos no esenciales y ácido pantoténico en el músculo, lo que debería contribuir a mejorar la calidad de la carne y la tasa de crecimiento. Este estudio proporciona una nueva visión sobre el mecanismo de la alteración inducida por la dieta del rendimiento del crecimiento animal y la calidad de la carne. Además, el estudio muestra que la suplementación dietética de glicina con LPD podría utilizarse para mejorar la calidad de la carne sin deteriorar el crecimiento animal.
1. Introducción
China es el mayor productor y consumidor de carne de cerdo del mundo, cuya producción y consumo alcanzaron 54,04 y 55,95 millones de toneladas en 2018 según las estadísticas de la Oficina Nacional,1 respectivamente. Sin embargo, con la expansión de la cría de cruces comerciales importados, la calidad de la carne está siendo cada vez más criticada por los consumidores. Obviamente, esto se debe principalmente a la reducción de la grasa intramuscular (IMF, comúnmente por debajo del 2,5%) como resultado de la selección genética para el crecimiento magro (1, 2), ya que el IMF afecta en gran medida la calidad de la carne y algunos estudios preliminares mostraron que los rasgos sensoriales de la carne de cerdo, como la jugosidad, la ternura y la aceptabilidad general, se ven afectados negativamente cuando el contenido de IMF es inferior a 2 ~ 2,5% (3). Por el contrario, las razas de cerdos autóctonas y rústicas suelen presentar un mayor contenido subcutáneo y de IMF, que se puntúan más alto en los paneles sensoriales (4, 5). Por lo tanto, se deben desarrollar estrategias para mejorar la calidad de la carne.
Las dietas reducidas en proteínas (DPR), solas o combinadas con algunos componentes, como los aminoácidos funcionales, parecen ser un enfoque prometedor para satisfacer las necesidades de los consumidores en cuanto a la calidad de la carne sin aumentar o incluso reducir los costes de producción para el sector ganadero (1). Varios estudios han demostrado que la reducción de proteínas en la dieta al 15-20% de NRC aumenta el contenido de IMF en los cerdos de engorde (1). La restricción de proteínas en la dieta (11,9, 13,3, 14,8, 16,2 y 17,6%) aumenta el contenido de marmoleado y IMF del músculo longissimus dorsi de los cerdos de engorde, y las concentraciones más bajas de proteínas en la dieta muestran el mayor contenido de marmoleado e IMF (6). Además, la reducción de las concentraciones de proteína bruta de 19.0 a 10.0% en una dieta alimentada con cerdos de 30 a 110 kg de peso corporal aumenta el contenido de músculo longissimus dorsi de 3.8 a 9.4% (7). Al mismo tiempo, Kerr et al. (8) informaron que la restricción de proteínas en la dieta desde el destete hasta el peso en el mercado aumentó el contenido de IMF en el músculo longissimus dorsi de los cerdos a un nivel dos veces mayor que el de los cerdos de control. También se han estudiado los efectos de la RPD combinada con aminoácidos sobre el rendimiento animal y la calidad de la carne. La literatura previa informa que el suplemento dietético de arginina promueve el crecimiento muscular y la deposición de IMF, pero disminuye el contenido de grasa corporal total en cerdos (9, 10), mostrando una mejor partición de la grasa. En comparación con la dieta proteica normal, la RPD y la RPD suplementadas con arginina aumentaron la IMF de los cerdos de 1,34 a 1,85 y 2,30 g/100 g de músculo, respectivamente, y en consecuencia mejoraron la terneza de la carne y la aceptación general (11). Aunque la RPD con suplementos de leucina no tuvo ningún efecto sobre el contenido de IMF (12), aumentó la jugosidad de la carne (11). Todos estos estudios demuestran que la reducción de proteínas en la dieta con o sin suplementos de aminoácidos es eficaz para mejorar la calidad de la carne. Sin embargo, al mismo tiempo, se observaron algunos efectos adversos de las dietas bajas en proteínas, como un menor rendimiento del crecimiento, una menor superficie del lomo y un mayor grosor de la grasa dorsal (7, 8). Por lo tanto, es necesario desarrollar estrategias para promover la deposición de IMF sin un aumento de la grasa subcutánea (mejor partición de la grasa) y una reducción del crecimiento magro para la producción porcina.
La glicina suele ser deficiente en las dietas bajas en proteínas (LPD). Algunos estudios en pollos han revelado que la suplementación con glicina o sus precursores de LPD supera la deficiencia y mejora el rendimiento del crecimiento animal (13, 14), lo que no se ha realizado en cerdos. En consecuencia, el objetivo del presente estudio fue explorar los efectos de la suplementación con glicina a LPD sobre el rendimiento del crecimiento y la calidad de la carne, así como los mecanismos subyacentes en cerdos de engorde y engorde.
2. Materiales y métodos
Todos los procedimientos con animales fueron revisados y aprobados por el Comité de Revisión y Gestión del Protocolo del Instituto de Agricultura Subtropical de la Academia China de Ciencias (Nº 8420190045).
2.1. Tratamiento animal y dietético
Se seleccionaron un total de 96 cerdos cruzados (Duroc × Landrace × Yorkshire, DLY) con un peso corporal inicial similar (peso corporal, 60,00 ± 2,00 kg) y se asignaron aleatoriamente a 3 tratamientos dietéticos (8 corrales por tratamiento y 4 cerdos por corral): (1) dieta normal (ND): los cerdos fueron alimentados con una dieta basal con 16% de proteína bruta (PC) para satisfacer las necesidades nutricionales de los cerdos en crecimiento y engorde según el Consejo Nacional de Investigación (NRC, 2012); (2) dieta baja en proteínas (LPD): los cerdos fueron alimentados con una dieta baja en proteínas con 12% de PC al disminuir el contenido de harina de soja; (3): LPD suplementado con glicina (LPDG): los cerdos fueron alimentados con un LPD suplementado con glicina al 0,57%. Todas las dietas fueron isoenergéticas y la composición química y los niveles de nutrientes de las dietas se enumeran en la Tabla Suplementaria S1. Los cerdos tenían acceso ad libitum a alimento y agua. El ensayo de alimentación se llevó a cabo durante 43 días.
2.2. Recogida de muestras
Al final del ensayo de alimentación, se seleccionaron aleatoriamente 8 cerdos (un cerdo por corral) de cada tratamiento dietético, se ayunaron durante 12 h y se sacrificaron de acuerdo con el procedimiento de sacrificio aplicado regularmente en una planta de sacrificio comercial (Hunan New Wellful Co., LTD, Changsha, China). A las 0, 0,75, 4 y 24 h postmortem (PM), se diseccionó un trozo de músculo Longissimus thoracis (LT) entre las vértebras torácicas 10 y 11 en el lado izquierdo de la canal, se cortó rápidamente en trozos pequeños, se mezcló y se congeló en nitrógeno líquido para su análisis químico, enzimático y metabonómico. Al mismo tiempo, se fijó un trozo de músculo a las 0 h PM para la tipificación de las fibras musculares mediante análisis histoquímico. Una chuleta de lomo de cerdo deshuesada se diseccionó a las 24 h PM desde la misma ubicación en el lado derecho de la canal para su evaluación sensorial.
2.3. Análisis químico
El contenido porcentual (p/p) de humedad, cenizas, proteínas y lípidos en el músculo (0 h PM) se analizó de acuerdo con GB5009.3-2016 Norma Nacional de Seguridad Alimentaria-Determinación de la Humedad en los Alimentos, GB5009.4-2016 Norma Nacional de Seguridad Alimentaria-Determinación de Cenizas en los Alimentos, GB5009.5-2016 Norma Nacional de Seguridad Alimentaria-Determinación de Proteínas en los Alimentos y GB5009.6-2016 Norma Nacional de Seguridad Alimentaria-Determinación de Lípidos en Alimentos, respectivamente.
2.4. Ensayo glucolítico
El glucógeno, la glucosa y el lactato en el músculo se determinaron utilizando kits de ensayo disponibles en el mercado (Solarbio, Pekín, China) de acuerdo con las instrucciones del fabricante. La glucosa-6-fosfato muscular se midió utilizando un kit de ensayo (Sigma, St. Louis, MO, Estados Unidos). El potencial glucolítico (GP) se calculó como potencial glucolítico = 2 × ([glucógeno] + [glucosa] + [glucosa-6-fosfato]) + [lactato] (15). El contenido de glucógeno en el músculo se expresó como μmol de glucosa/g de músculo. La GP se expresó como μmol de lactato/g de músculo. El contenido de glucosa, glucosa-6-fosfato y lactato en el músculo se expresó como μ mol/g de músculo.
2.5. Análisis de la actividad enzimática
Las actividades enzimáticas de glucógeno fosforilasa a (GPa), piruvato quinasa (PK), lactato deshidrogenasa (LDH) y citrato sintasa (CS) se determinaron utilizando kits comerciales (Solarbio, Beijing, China) de acuerdo con el procedimiento del fabricante. Las actividades de la citocromo c oxidasa (CcO) se midieron utilizando un kit de BioVision (BioVision, Palo Alto, CA, Estados Unidos). La actividad de GPa se expresó como nanomoles de NADPH producidos a partir de glucosa-6-fosfato por glucosa-6-fosfato deshidrogenasa por minuto por miligramo de proteína muscular. La actividad PK se calculó como nanomoles de NADH oxidados por minuto por miligramo de proteína muscular. La actividad de LDH se expresó como nanomoles de piruvato producidos a partir de lactato por minuto por miligramo de proteína muscular. La actividad de CS se calculó como nanomoles de 5′-tionitrobenzoato (TNB) producidos a partir de 5,5′-ditiobis-(2-ácido nitrobenzoico) (DTNB) por minuto por miligramo de proteína muscular. La actividad de CcO se calculó como nanomoles de citocromo C oxidados por minuto por miligramo de proteína muscular.
2.6. Análisis histoquímico de los tipos de fibras musculares
Un trozo de músculo LT se fijó en fijador (Servicebio, Pekín, China) durante 24 h, se deshidrató sucesivamente en soluciones de sacarosa al 15 y al 30% a 4°C, y se embró en el compuesto Tissue-Tek O.C.T. (Sakura, Japón). Los bloques de tejido se seccionaron en rodajas de 8 a 10 μm utilizando un criostato (CryoStar NX50, Thermo, Estados Unidos) a -20 °C.
La tinción histoquímica de la miosina adenosina trifosfatasa (m-ATPasa) se realizó como en la literatura (16). Brevemente, las secciones musculares se preincubaron primero en solución (0,17 M Tris-base, 20 mM CaCl2, pH 10,4) durante 5 min, luego se incubaron en solución (0,17 M Tris-base, 18 mM CaCl2, 2,7 mM ATP, pH 9,4) durante 30 min. Las secciones de tejido se tiñeron con cloruro de calcio al 2% durante 3 veces cada una durante 2 min, luego se tiñeron con nitrato de cobalto al 2% durante 5 min y se lavaron, y finalmente se tiñeron con sulfuro de amonio al 1% durante 30 s y se lavaron. Las secciones de tejido se deshidrataron sucesivamente en alcohol al 70, 80, 95 y 100%, se diafanizaron en xileno y se sellaron con goma neutra.
Los portaobjetos histológicos se escanearon con un escáner digital de portaobjetos (Pannoramic DESK, 3DHistech, Hungría) y las fibras musculares se contaron con el software Image-Pro Plus 6.0 (Media Cybernetics, Inc., Estados Unidos). Se calcularon los porcentajes de fibras musculares tipo I y tipo II.
2.7. Evaluación sensorial
La evaluación sensorial fue realizada por 10 panelistas al igual que en la literatura (17). Brevemente, las chuletas de lomo deshuesadas (2 cm de grosor) se asaron en una sartén a una temperatura interna de 71 ° C, se cortaron encubos de 2 cm 3, se envolvieron en papel de aluminio preetiquetado y se colocaron en una incubadora calentada antes de entregarlas a los evaluadores. Se entregaron a cada panelista en una sesión cuatro muestras, de las cuales tres eran muestras de prueba (una muestra de cada tratamiento dietético) y una era una muestra de referencia (chuletas de lomo de cerdo comerciales). Las muestras de prueba se puntuaron en una escala de 1 a 10 puntos en comparación con la muestra de referencia descrita anteriormente.
2.8. Análisis metabolómico no dirigido
Los tejidos musculares se pulverizaron en nitrógeno líquido. Se suspendieron 100 mg de muestra en polvo en 500 μl de metanol preenfriado al 80% que contenía ácido fórmico al 0,1% y se homogeneizaron para la extracción de metabolitos. Las muestras se centrifugaron a 9000 rpm, 4°C durante 5 min. Los sobrenadantes se recolectaron y diluyeron con agua de grado LC-MS hasta una concentración que contenía 60% de metanol. Los extractos se pasaron a través de un filtro de 0,22 μm y se utilizaron para el análisis LC-MS/MS.
Los análisis LC-MS/MS se realizaron utilizando un sistema UHPLC Vanquish (Thermo Fisher) junto con un espectrómetro de masas Orbitrap Q Exactive HF-X (Thermo Fisher). Las muestras se inyectaron en una columna de Hyperil Gold (1,9 μm, 100 × 2,1 mm, Thermo Fisher) y se eluyeron utilizando un gradiente lineal de 16 min a un caudal de 0,2 ml/min. Las fases móviles para el modo de iones positivos fueron ácido fórmico al 0,1% en agua (A) y metanol (B). Las fases móviles para el modo de iones negativos fueron 5 mM de acetato de amonio (pH 9,0) (A) y metanol (B). El gradiente de elución fue el siguiente: 0 min, 2% B, 1,5 min, 2% B; 12 min, 100% B; 14.0 min, 100% B; 14,1 min, 2% B; 16 min, 2% B. Q El espectrómetro de masas HF-X exactivo funcionó en modo de polaridad positiva/negativa con voltaje de pulverización de 3,2 kV, temperatura capilar de 320 °C, caudal de gas de la vaina de 35 arb y caudal de gas auxiliar de 10 arb.
Los datos brutos generados por UHPLC–MS/MS se procesaron utilizando el Compound Discoverer (versión 3.0, Thermo Fisher) para realizar la alineación de picos, la selección de picos y la cuantificación de cada metabolito. Los parámetros principales se establecieron de la siguiente manera: tolerancia de tiempo de retención = 0,2 min, tolerancia de masa = 5 ppm; tolerancia de intensidad de señal = 30%; relación señal/ruido = 3; Intensidad mínima = 100.000. Las intensidades máximas se normalizaron a la intensidad espectral total. Los datos normalizados se utilizaron para predecir la fórmula molecular basada en iones aditivos, picos de iones moleculares e iones fragmentados. Los picos se compararon con la base de datos mzCloud2 y ChemSpider3 para la anotación y cuantificación de picos.
2.9. Análisis estadístico
El análisis de los datos se realizó con el programa SPSS 25.0 (SPSS Inc., Chicago, IL, Estados Unidos). El ANOVA de un factor se llevó a cabo utilizando las mediciones de los valores medios seguidos de la prueba de diferencia mínima significativa protegida de Fisher, con cerdo individual (n = 8) como unidad experimental. Los datos de la evaluación sensorial se analizaron utilizando un procedimiento de modelo mixto en el que los tratamientos dietéticos, las sesiones y sus interacciones se trataron como efectos fijos y los panelistas como efectos aleatorios. Los niveles de diferencias significativas se establecieron en p < 0,05. Se informaron los valores medios y los errores estándar de las medias. Se realizó un análisis de componentes principales (ACP) para explorar las relaciones entre los rasgos de calidad sensorial y el contenido total de lípidos entre los tratamientos dietéticos. Los datos se normalizaron estableciendo los valores medios en cero y escalando sobre la base de una desviación estándar. Los gráficos se trazaron utilizando el software OriginPro 8.
3. Resultados y discusión
3.1. La dieta baja en proteínas suplementada con glicina aumentó la IMF pero disminuyó el glucógeno en el músculo
Muchos estudios han informado que las dietas reducidas en proteínas aumentan la acumulación de grasa corporal, el contenido de IMF y el marmoleado de los cerdos en crecimiento y engorde (1), lo que demuestra que la restricción de proteínas podría utilizarse para mejorar la calidad de la carne. Sin embargo, esto suele ir acompañado de una reducción de la tasa de crecimiento de los animales y de la producción de carne magra (7, 8). Por esta razón, se estudió el efecto de LPD suplementado con glicina sobre el crecimiento de los cerdos, el metabolismo muscular y la calidad de la carne. El rendimiento del crecimiento de los cerdos se muestra en la Tabla Suplementaria S2. La LPD disminuyó la ganancia de peso diaria promedio (GMD) de 855 ± 17 g a 777 ± 15 g (p < 0.05) en comparación con la ND, pero la suplementación de glicina a LPD aumentó la GMD a 839 ± 31 g, que no fue diferente (p > 0.05) a la del tratamiento con ND, lo que demuestra que la suplementación dietética de glicina a LPD eliminó el efecto negativo de la restricción proteica en el rendimiento del crecimiento animal.
Como se muestra en la Tabla 1, LPD y LPDG no tuvieron ningún efecto sobre el contenido de humedad, proteínas y cenizas en el músculo porcino, pero LPDG aumentó significativamente (p < 0.05) la deposición de grasa dentro del músculo, que es aproximadamente un 60% más alta que la de los cerdos control ND. Aunque no es estadísticamente significativo, la dieta baja en proteínas sin suplementación con glicina aumentó la IMF de 1,83 ± 0,23 a 2,64 ± 0,27% en comparación con la dieta normal control. Esto concuerda con la literatura de que la LPD combinada con o sin suplementación con aminoácidos aumenta el contenido de grasa porcina (1). Dado que el IMF es un factor clave que determina la calidad de la carne (3), el aumento del contenido de IMF en el músculo de los cerdos LPDG debería mejorar la calidad de la carne. Al mismo tiempo, tanto el contenido de glucógeno como el potencial glucolítico fueron significativamente más bajos en las dos dietas bajas en proteínas con o sin suplementación con glicina en comparación con la dieta normal de control. Dado que el glucógeno es el sustrato de la glucólisis postmortem, que desempeña un papel clave en la determinación de los valores de pH final del músculo y en la formación postmortem de la calidad de la carne, el contenido reducido de glucógeno en el músculo del cerdo indica que una dieta baja en proteínas influye en la calidad de la carne de cerdo al alterar el metabolismo de los carbohidratos musculares. Tomados en conjunto, estos datos muestran que LPD y LPDG alteraron el metabolismo de nutrientes en el músculo de los cerdos en crecimiento y engorde, con una disminución de la acumulación de glucógeno pero un aumento de la deposición de grasa. La LPD perjudicó el rendimiento del crecimiento de los animales, mientras que la suplementación con glicina al LPD recuperó la tasa de crecimiento de los cerdos.
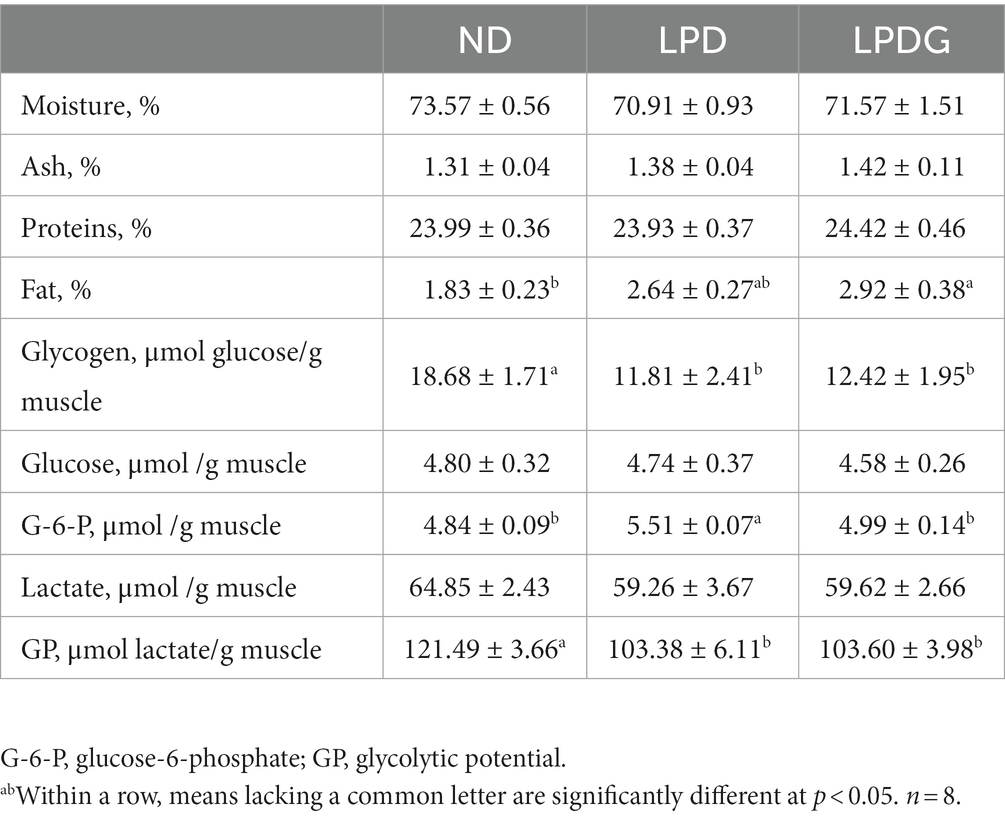
3.2. La dieta baja en proteínas suplementada con glicina mejoró la calidad de la carne
Los datos sensoriales se presentan en la Tabla 2. De acuerdo con el contenido de IMF, LPDG aumentó significativamente la terneza, la jugosidad y el puntaje general de gusto del lomo de cerdo en comparación con ND. Esto es razonable, ya que el IMF afecta en gran medida la ternura, la jugosidad y la aceptabilidad general de la carne, que se ven afectadas negativamente cuando el FMI < 2-2,5% (3). Tanto el LPD como el LPDG aumentaron el IMF de la carne de cerdo de 1,83 a más del 2,5%, lo que demuestra que el LPD con o sin suplementación con glicina tuvo un efecto positivo en la calidad de la carne. De hecho, la LPD sin suplementación con glicina también aumentó la ternura de la carne y la puntuación general de agrado, aunque la jugosidad de la carne de cerdo no se vio afectada (Tabla 2). Dado que los lípidos son la principal fuente de compuestos aromatizantes en la carne de cerdo (18), especialmente los compuestos saborizantes volátiles, como aldehídos, cetonas y alcoholes que se forman comúnmente por oxidación de grasas o degradación térmica (17, 19), es razonable observar que el LPD y el LPDG mejoraron el olor carnoso del lomo de cerdo (Tabla 2).
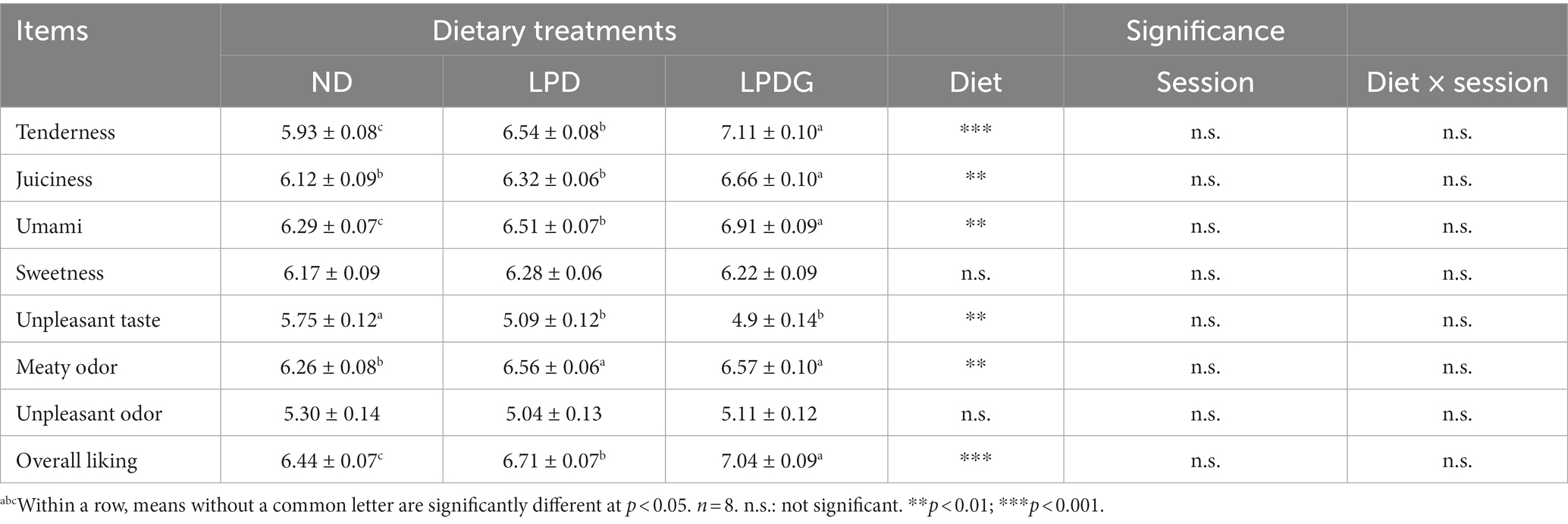
Tabla 2. Evaluación de panel sensorial de lomo de cerdo de cerdos en crecimiento y engorde que reciben una dieta normal, baja en proteínas o baja en proteínas suplementada con glicina.
Para mostrar mejor el impacto de las dietas en la calidad de la carne, se realizó un análisis de componentes principales para explorar las relaciones entre los rasgos de calidad sensorial, el contenido total de lípidos y los tratamientos dietéticos. Como se muestra en la Figura Suplementaria S1, el PC1 explicó el 65,30 de la varianza asociada con los rasgos de calidad de la carne y podría representar la calidad general de la carne. El PC1 separó LPDG y LPD de los tratamientos con ND, mostrando que la carne de los dos tratamientos con dieta baja en proteínas tenía mayor calidad que la carne del tratamiento con ND. Además, el contenido de lípidos, la jugosidad de la carne, la ternura y el gusto general se encuentran en el primer cuadrante, lo que muestra la estrecha relación entre el contenido de lípidos de la carne y estos rasgos de calidad de la carne. En resumen, estos datos muestran que el FMI tuvo un impacto importante en la calidad de la carne. El LPDG mejoró la calidad de la carne al aumentar el contenido del FMI.
3.3. Dieta baja en proteínas suplementada con glicina, alteró las actividades de las enzimas metabólicas y la tipificación de fibras musculares
Para verificar y comprender mejor el metabolismo alterado dentro del músculo, se midieron las actividades de algunas enzimas glucogeníticas/glucolíticas y oxidativas en el músculo LT. Como se muestra en la Figura 1, las actividades de PK a las 0 h PM fueron significativamente menores, pero las actividades de CcO fueron significativamente mayores (p < 0.05) para los tratamientos con LPD y LPDG en comparación con el tratamiento con ND, que representan las actividades enzimáticas in vivo. Esto demuestra que la reducción de proteínas en las dietas aumentó la capacidad del metabolismo oxidativo del músculo animal, pero disminuyó la capacidad glucolítica. Además, se determinaron mayores actividades de GPa y PK tanto a las 0,75 como a las 4 h PM, y menores actividades de CS a las 0,75 h PM, así como menores actividades de CcO a las 0,75 y 4 h PM para los cerdos ND en comparación con los cerdos LPD y LPDG. Estos datos muestran además que la reducción de proteínas en las dietas aumentó la capacidad oxidativa del músculo animal, pero disminuyó la capacidad glucolítica. La actividad de la LDH fue similar entre los tratamientos hasta las 4 h PM. Esto es lógico porque la LDH no es una enzima limitante de la velocidad y su actividad puede no correlacionarse con la capacidad glucolítica/oxidativa muscular. La diferencia en la actividad de la LDH entre los tratamientos dietéticos a las 24 h PM puede reflejar una diferencia en la estabilidad de esta proteína.
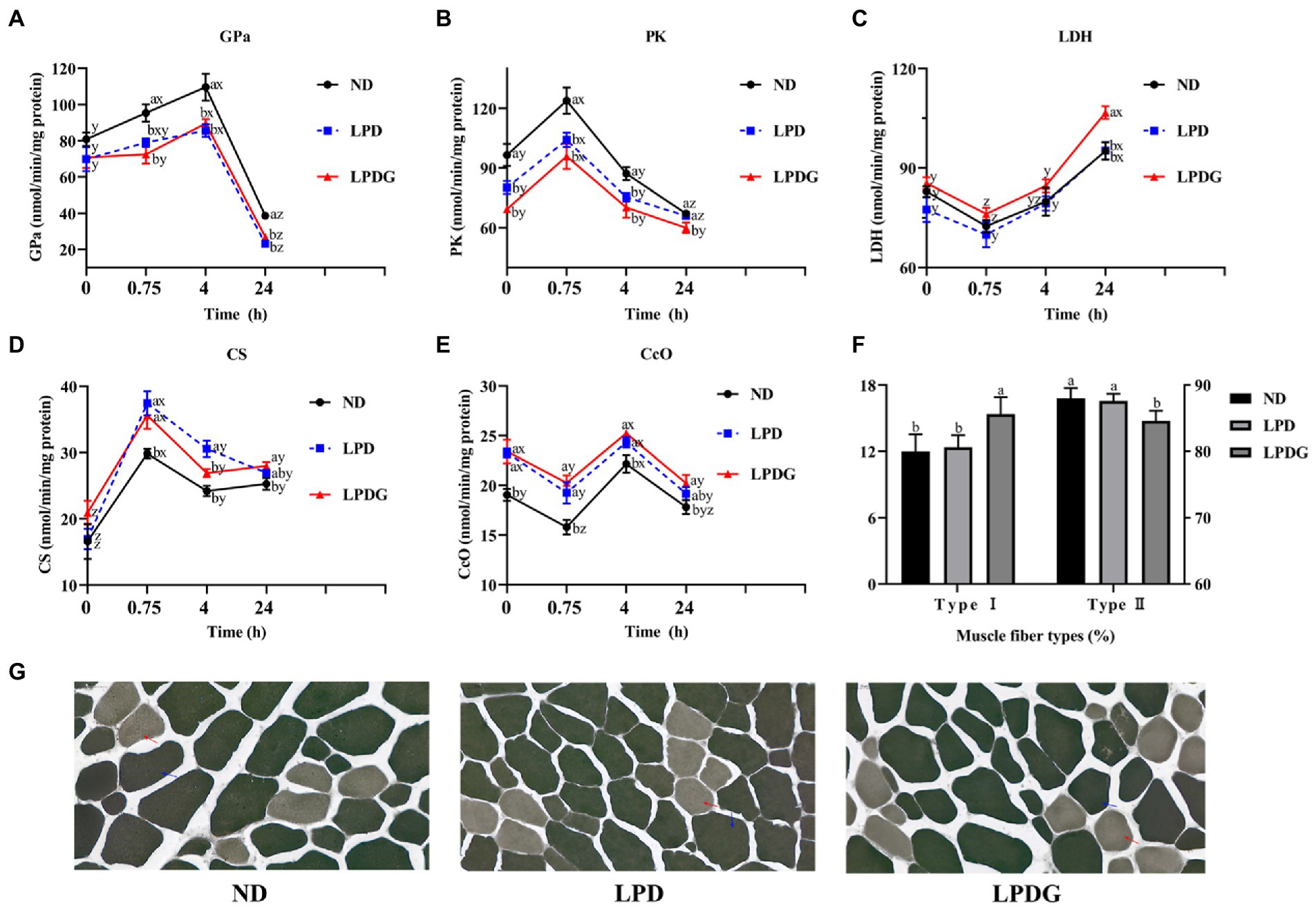
De acuerdo con la actividad m-ATPasa, las fibras musculares se pueden clasificar en fibras musculares tipo I y tipo II (20, 21). La fibra muscular tipo I también se conoce como fibra muscular de contracción lenta, fibra muscular oxidativa o fibra muscular roja. Las fibras de tipo I son ricas en mitocondrias y lípidos, tienen una alta actividad de enzimas metabólicas aeróbicas (como la citocromo oxidasa, la succinato deshidrogenasa, la citrato sintasa, etc.) y una baja actividad de la ATPasa. Además, los músculos lentos son ricos en capilares y mioglobina, tienen una gran capacidad de transporte de oxígeno, se contraen lenta y duraderamente. Las fibras musculares de tipo II son fibras musculares de contracción rápida, fibras musculares glucolíticas o fibras musculares blancas. Las fibras de tipo II son ricas en glucógeno, tienen altas actividades enzimáticas glucolíticas y actividad m-ATPasa, se contraen rápidamente pero no de forma duradera. El análisis histoquímico de la actividad de la m-ATPasa reveló que la LPDG aumentó el número relativo de fibras musculares de tipo I y simultáneamente disminuyó el número de fibras musculares de tipo II dentro del músculo LT (Figuras 1F, G), lo que demuestra que la LPDG promovió la transición de la fibra muscular porcina del tipo II al tipo I. Las miofibras de tipo I suelen tener un diámetro más pequeño y un mayor contenido de lípidos. es razonable que la carne del grupo LPDG fuera más tierna y de mejor sabor (Tabla 2). Varios estudios han reportado que los músculos porcinos con más fibras musculares tipo I tienen enzimas del metabolismo oxidativo reguladas al alza, menor potencial glucolítico, menor glucólisis postmortem y mayor calidad de la carne (22-24). Con base en la literatura y los datos obtenidos en el presente estudio, se puede concluir que el LPDG promovió la transición de las fibras musculares del tipo II al tipo I, alteró el metabolismo energético muscular, aumentó el contenido de IMF y, por lo tanto, mejoró la calidad de la carne.
3.4. Dieta baja en proteínas suplementada con o sin metaboloma alterado por la glicina en el músculo porcino
Para una mejor exploración de los cambios inducidos por las dietas, se realizó un perfil metabólico del músculo LT utilizando UHPLC-MS/MS, que identificó 859 metabolitos en modo positivo y 805 metabolitos en modo negativo. Dado que se identificaron 156 metabolitos tanto en modo positivo como negativo, se identificaron un total de 1.508 metabolitos y se aplicaron para el análisis de MetaboAnalyst en el presente estudio.
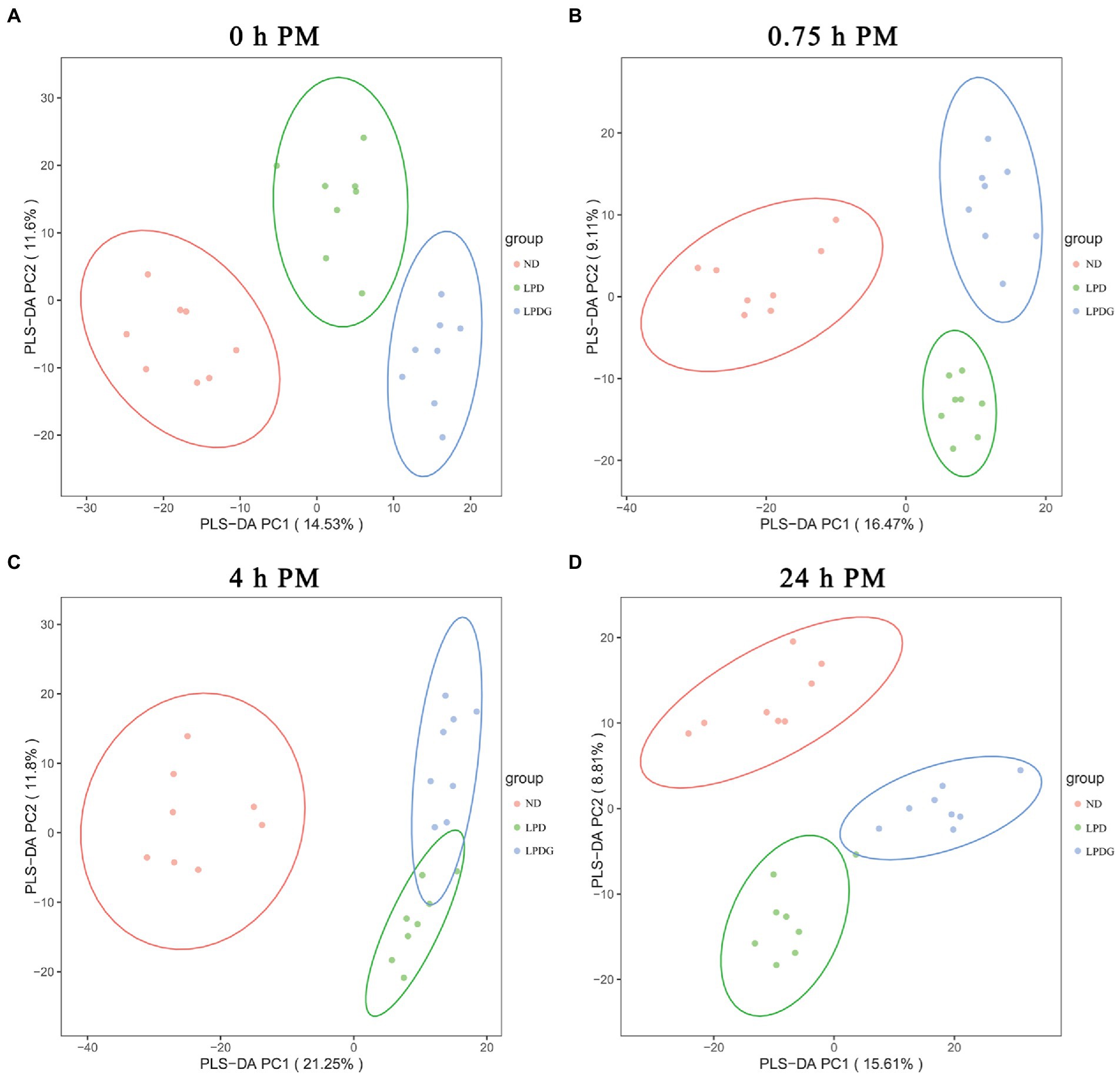
Para proporcionar una visión general de la diferencia en los perfiles metabólicos, se realizó un análisis de discriminación de mínimos cuadrados parciales (PLS-DA). Como se muestra en la Figura 2, PLS-DA separó de manera efectiva y distinta las muestras de músculo LT de cada cohorte en función de las dietas de los animales, lo que demuestra que LPD y LPDG causaron un cambio significativo en el perfil general de metabolitos del músculo LT dentro de las 24 h PM. En otras palabras, los cerdos que recibieron una dieta normal, baja en proteínas o una dieta baja en proteínas suplementada con glicina tenían patrones metabólicos distintos en el músculo esquelético postmortem. Dado que el perfil de metabolitos determinado en el músculo recogido a las 0 h PM representa el metabolismo in vivo, la buena segregación de las muestras de músculo ND y LPDG con muestras de LPD colocadas entre estas dos cohortes (Figura 2A) sugiere que la suplementación con glicina tuvo efectos aditivos en la dieta baja en proteínas y alteró aún más el metabolismo en el músculo esquelético de los cerdos. Además, las muestras de músculo ND se ubicaron relativamente más lejos de los dos grupos de dieta baja en proteínas (Figura 2), lo que sugiere que la dieta baja en proteínas causó mayores cambios en el metabolismo que la suplementación con glicina. Esto concuerda con el contenido de glucógeno (Tabla 1) y las actividades de PK y CcO a las 0 h PM (Figura 1), todas las cuales fueron significativamente mayores o menores (p < 0,05) en el grupo ND, pero no diferentes entre las dos dietas bajas en proteínas, lo que sugiere que el contenido de glucógeno, las actividades PK y CcO podrían ser buenos indicadores del metabolismo muscular. Para proporcionar una visión general de los metabolitos diferencialmente abundantes en los tres grupos dietéticos, se realizó un análisis de agrupamiento jerárquico (HCA) utilizando todos los metabolitos anotados significativamente expresados diferencialmente y los resultados se muestran en un mapa de calor (Figura suplementaria S2). El HCA también mostró que las muestras de músculo porcino a las 0 h PM se separaron de acuerdo con los tratamientos dietéticos, con todas las muestras de LPDG agrupadas en el lado izquierdo, la mayoría de las muestras de ND (6/8) agrupadas en el lado derecho, y las muestras de LPD entremezcladas con una muestra de ND en el centro del panel (Figura suplementaria S2A). Una vez más, este patrón de perfil reveló que la dieta baja en proteínas obviamente cambió el metabolismo en animales vivos y la suplementación de glicina con LPD amplió la brecha. Además, el perfil de metabolitos agrupó bien las ocho muestras de ND, pero las muestras de LPD se mezclaron con muestras de LPDG recogidas a las 0,75 y 4 h PM (Figura suplementaria S2B,C), lo que revela que la dieta baja en proteínas causó mayores cambios metabólicos postmortem en comparación con la dieta normal que la suplementación con glicina en comparación con la dieta baja en proteínas.
Los metabolitos diferenciales entre los grupos de dieta se filtraron de acuerdo con los siguientes criterios de significación estadística: umbrales con VIP > 1, p < 0.05 y FC > 1.5 o FC < 0.667 y se resumieron visualmente en gráficos de volcanes (Figura Suplementaria S3). Los metabolitos que contribuyeron a la diferencia metabolómica en las comparaciones por pares se marcaron con el tamaño de los círculos y se codificaron por colores en función de los valores de VIP y p del análisis PLS-DA. La información detallada de estos metabolitos diferenciales del músculo LT a las 0, 0,75, 4 y 24 h PM se enumera en la Tabla 3 y la Tabla Suplementaria S1. La mayoría de los metabolitos diferenciales son hidratos de carbono, aminoácidos y sus derivados, lípidos, purinas y cofactores.
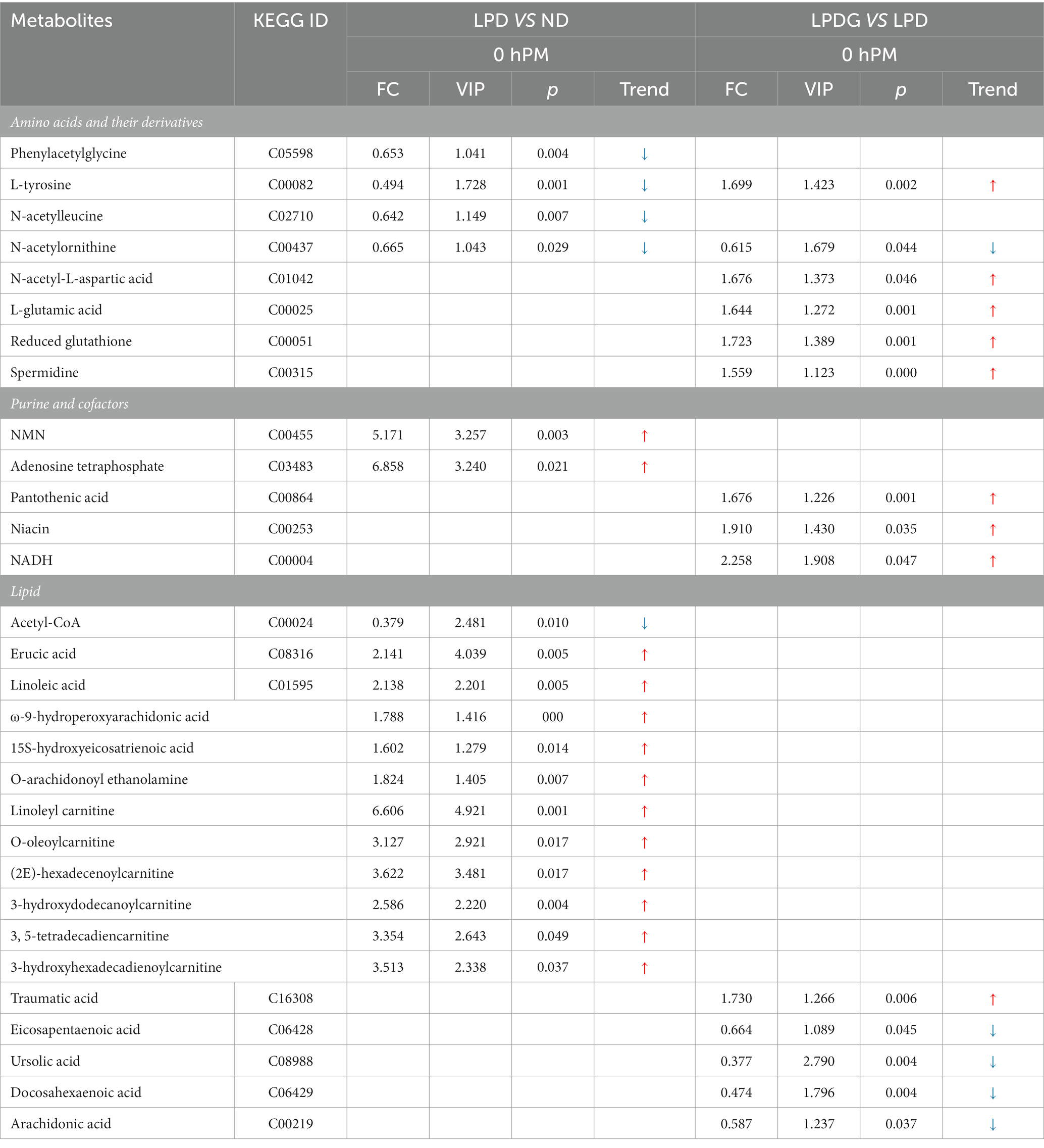
Tabla 3. Metabolitos significativamente alterados en el músculo LT (recogidos a las 0 h PM) de cerdos alimentados con una dieta baja en proteínas en comparación con los cerdos alimentados con la dieta normal, y cerdos alimentados con una dieta baja en proteínas suplementada con glicina en comparación con los cerdos alimentados con la dieta baja en proteínas.
De acuerdo con lo informado previamente por investigadores que las dietas influyen en el metabolismo de proteínas/aminoácidos (25-27), la dieta baja en proteínas redujo el contenido de múltiples aminoácidos en el músculo LT porcino en comparación con la dieta normal (Tabla 3), incluyendo L-tirosina, fenilacetilglicina, N-acetilecina y N-acetilornitina. Sin embargo, la suplementación de glicina con LPD revirtió el contenido de L-tirosina. Además, la suplementación con glicina aumentó las concentraciones de ácido L-glutámico y ácido N-acetil-L-aspártico, así como redujo el glutatión y la espermidina. Con base en estos datos y en el rendimiento del crecimiento de los cerdos, se puede especular que la insuficiencia de aminoácidos inducida por LPD, especialmente la tirosina, podría ser responsable de la disminución de la deposición de proteínas musculares y, por lo tanto, de la disminución de la GMD en los animales (28). La suplementación con glicina puede haber promovido la síntesis y reservado aminoácidos esenciales (EAA) para aminoácidos no esenciales (NEAA), promoviendo la síntesis de proteínas musculares y el metabolismo energético (29). La N-acetilornitina disminuyó en ambos grupos de dieta baja en proteínas en comparación con la dieta normal.
De acuerdo con el aumento de la concentración de glutatión reducido, la glicina aumentó las concentraciones de NADH y su precursor niacina (Tabla 3), mostrando un elevado potencial antioxidante y reflejando un metabolismo alterado del músculo de los cerdos LPDG. El ácido pantoténico es un precursor de la coenzima A (CoA) y un elemento estructural de la proteína transportadora de acilo, que participa en numerosas reacciones catabólicas y sintéticas celulares. La deficiencia severa de ácido pantoténico en ratas causa pérdida de peso, alteraciones en el pelaje y finalmente la muerte (30). Estudios previos informan que el ácido pantoténico en la dieta aumenta la eficiencia de retención de nitrógeno, la tasa de crecimiento y el peso corporal final de los peces (31). En el presente estudio, la suplementación de glicina con LPD aumentó el contenido de ácido pantoténico en el músculo porcino, lo que debería haber contribuido a la recuperación del rendimiento del crecimiento de los cerdos LPDG.
El metabolismo de los lípidos se ve muy alterado por la restricción de proteínas. Como se muestra en la Tabla 3, múltiples ácidos grasos y derivados, incluyendo ácido erúcico, ácido linoleico, ácido ω-9-hidroperoxiaraquidónico, ácido 15-s-hidroxieicosatrienoico y O-araquidonoiletanolamina aumentaron significativamente en ambos grupos de dieta baja en proteínas en comparación con el grupo de ND. Además, se determinó que múltiples acilcarnitinas de diferentes especificidades de longitud de cadena aumentaron significativamente con dietas bajas en proteínas, que incluyen linoleil carnitina, O-oleoilcarnitina, (2E)-hexadecenoilcarnitina, 3-hidroxidodecanoilcarnitina, 3, 5-tetradecadiencarnitina y 3-hidroxihexadecadienoilcarnitina. Estos datos revelaron que las dietas bajas en proteínas promovieron no solo la deposición de IMF como se informó anteriormente (1), sino también el transporte de ácidos grasos a través de la membrana mitocondrial y la oxidación. Dado que las fibras musculares lentas dependen más del catabolismo de los ácidos grasos y de la fosforilación oxidativa para la producción de ATP, el aumento de la movilización de grasa en el músculo de los cerdos alimentados con dietas bajas en proteínas sugiere una mejora de la capacidad oxidativa, como lo revela el aumento de las actividades enzimáticas de CS y CcO (Figuras 1D, E). Esto es especialmente cierto para los cerdos LPDG, ya que la suplementación con glicina aumentó significativamente el porcentaje de fibra muscular de tipo lento (tipo I) en el músculo LT porcino (Figuras 1F, G). Cuando se compararon los dos grupos de dietas bajas en proteínas, la suplementación con glicina disminuyó el contenido muscular de algunos ácidos grasos poliinsaturados, incluidos el ácido eicosapentaenoico, el ácido docosahexaenoico y el ácido araquidónico, lo que probablemente esté relacionado con el aumento del contenido de ácido pantoténico del grupo LPDG, ya que el ácido pantoténico altera la acumulación de ácidos grasos saturados e insaturados en el hígado y el músculo de los peces (31). El acetil-CoA es un metabolito intermedio importante del metabolismo energético. Los carbohidratos, los ácidos grasos y los aminoácidos convergieron en el acetil-CoA antes de entrar en el ciclo tricarboxílico para la oxidación completa y la generación de ATP a través de la fosforilación. El acetil-CoA es también el sustrato para la gluconeogénesis, la cetogénesis y la síntesis de ácidos grasos. La reducción de acetil-CoA en los dos grupos de dieta baja en proteínas reveló una alteración completa en el metabolismo muscular inducida por la restricción de proteínas en la dieta.
Las dietas también indujeron alteración del metabolismo en el músculo post mortem. Algunos metabolitos expresados diferencialmente, que pueden estar relacionados con la conversión de músculo en carne, se enumeran en la Tabla Suplementaria S3. La glucólisis determinó la tasa de disminución del pH muscular y los valores finales de pH muscular. En el presente estudio, se detectaron tres intermediarios de glucólisis, glucosa 6-fosfato, gliceraldehído 3-fosfato y ácido 3-fosfoglicérico que diferencian en abundancia entre el ND y los dos grupos de dieta baja en proteínas a las 0,75 PM. Otra característica del músculo postmortem es que muchos aminoácidos y péptidos aumentaron en abundancia en el músculo postmortem por la restricción de proteínas, de los cuales se detectó que algunos eran más bajos en el grupo LPDG a las 24 h PM cuando se compararon los dos grupos de dieta baja en proteínas, lo que indica que las dietas bajas en proteínas promovieron la proteólisis, lo que influye en el envejecimiento de la carne y la calidad final de la carne. como la ternura y el sabor de la carne. También se determinaron concentraciones más altas de AMP, ADP y UMP en las dos dietas bajas en proteínas. En resumen, todos estos datos mostraron que la restricción de proteínas en la dieta y la suplementación con glicina influyeron en la glucólisis, la proteólisis y el metabolismo de nucleótidos en el músculo postmortem y, probablemente, en la conversión del músculo en carne y en la calidad de la carne.
Para evaluar aún más las perturbaciones bioquímicas en la etapa de crecimiento de los cerdos inducidas por LPD y LPDG, se realizó un análisis de enriquecimiento de la vía basado en los metabolitos identificados que se expresaron significativamente de manera diferencial. El análisis de enriquecimiento de KEGG (Tablas suplementarias S4, S5) mostró que 27 y 20 vías metabólicas, incluidas las vías metabólicas de aminoácidos, vías metabólicas de lípidos, vías metabólicas de carbohidratos y vías de metabolismo de nucleótidos, se predijeron de acuerdo con los metabolitos identificados en los grupos LPD y LPDG, respectivamente. De acuerdo con los resultados del análisis de la ruta metabólica del programa MetaboAnalyst 4.0 (Figura 3, datos brutos con una p < 0,05), la biosíntesis de fenilalanina, tirosina y triptófano, la síntesis y degradación de cuerpos cetónicos y el metabolismo del ácido linoleico se vieron significativamente afectados por la LPD (Figura 3A). Por su parte, el metabolismo de la D-glutamina y D-glutamato, el metabolismo del glutatión, la biosíntesis de ácidos grasos insaturados, la biosíntesis de arginina, el metabolismo de la alanina, el aspartato y el glutamato, la biosíntesis de fenilalanina, tirosina y triptófano fueron alterados significativamente por el LPDG (Figura 3B).
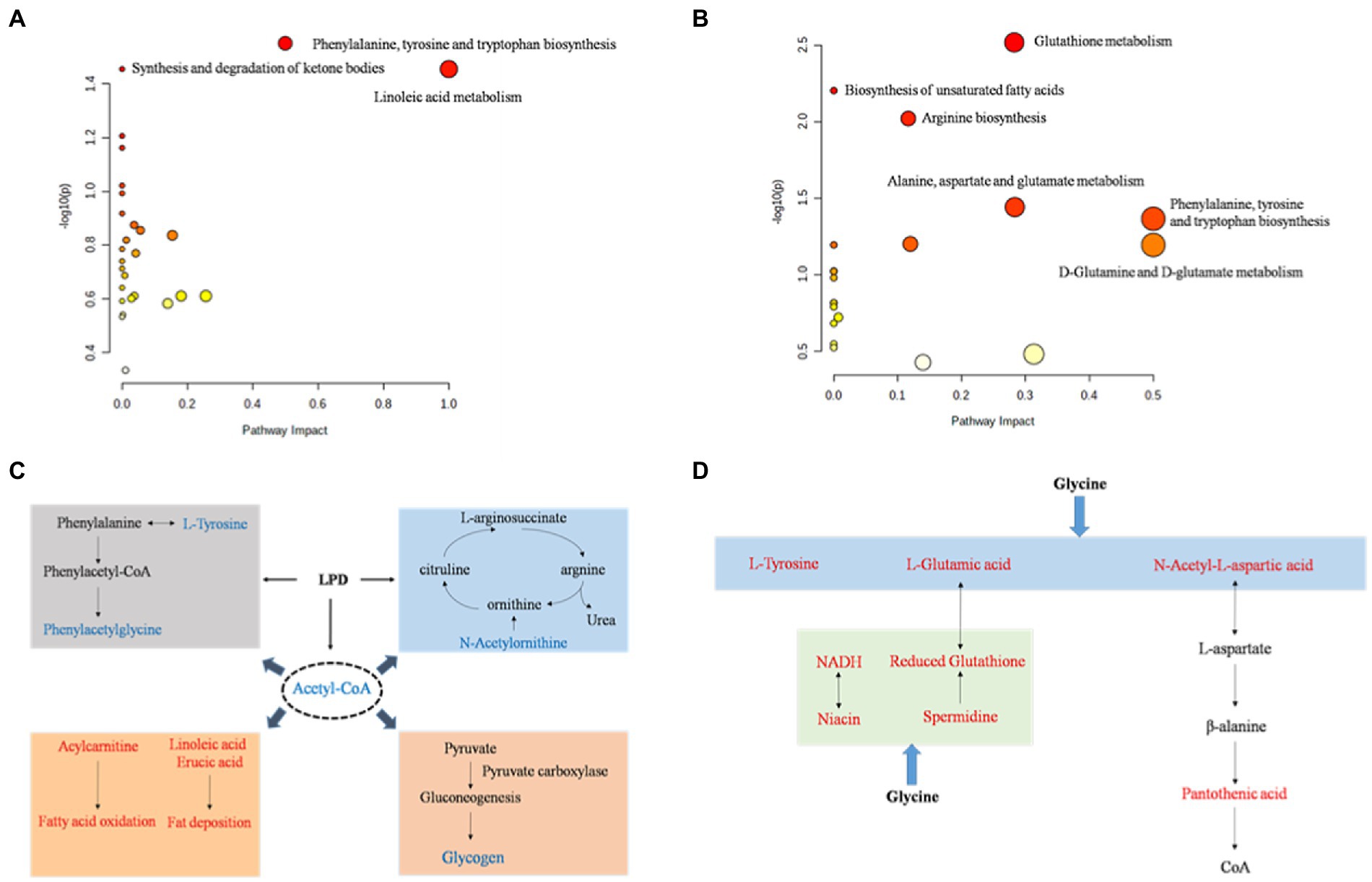
Sobre la base de la base de datos KEGG, se resumieron y esbozaron las principales vías que contribuyen a las diferencias de metabolitos del músculo LT porcino por LPD y LPDG en las Figuras 3C, D. Tradicionalmente, los aminoácidos se clasifican como EAA y NEAA. Sin embargo, cada vez más literatura informa que algunos de los NEAA tradicionalmente clasificados desempeñan importantes funciones reguladoras en la señalización celular, la expresión génica, el recambio de proteínas, el metabolismo de nutrientes y la defensa oxidativa (32). En el presente estudio, la LPD perjudicó el crecimiento de los animales y redujo significativamente la GMD de los cerdos en crecimiento y engorde (28). Al mismo tiempo, disminuyeron en abundancia múltiples aminoácidos en el músculo LT de los cerdos LPD, incluyendo L-tirosina, fenilacetilglicina, N-acetilleucina y N-acetilornitina (Tabla 3). Aunque la función reguladora fisiológica de estos aminoácidos no se conoce bien, es probable que la restricción proteica del grupo LPD condujera a una insuficiencia de tirosina y fenilacetilglicina y, por lo tanto, redujera la síntesis de proteínas y el crecimiento animal (Figura 3C). La N-acetilornitina disminuyó en ambos grupos de dieta baja en proteínas en comparación con la dieta normal. Dado que la N-acetilornitina se desacetila para producir ornitina, la disminución de la N-acetilornitina podría ser un mecanismo para reducir la emisión de nitrógeno a través del ciclo de la urea inducido por dietas bajas en proteínas (Figura 3C). El acetil-CoA se redujo significativamente en ambos grupos de dietas bajas en proteínas, lo que podría ser, al menos parcialmente, un efecto acumulado de la síntesis de ácidos grasos y la oxidación inducida por la restricción de proteínas (Figura 3C). Por otro lado, la reducción de acetil-CoA mediante dietas bajas en proteínas puede regular el metabolismo intracelular. Estudios previos revelan que el acetil-CoA modula la actividad de la piruvato carboxilasa (PC) y la disminución del acetil-CoA en el hígado suprime la producción de glucosa hepática (33). Es probable que la dieta baja en proteínas indujera la movilización de ácidos grasos redujera el acetil-CoA en el músculo porcino y, por lo tanto, probablemente en el hígado, lo que a su vez suprimió la actividad del PC, la producción hepática de glucosa y la acumulación de glucógeno en el músculo de los animales (Figura 3C).
La suplementación con glicina a una dieta baja en proteínas revirtió la L-tirosina en el músculo porcino al mismo nivel que en el grupo ND (Tabla 3). Además, el suplemento de glicina aumentó significativamente el contenido muscular de ácido L-glutámico y ácido N-acetil-L-aspártico, que se pueden utilizar para la síntesis de ácido L-aspártico y ácido pantoténico. Estos datos revelaron que la suplementación con glicina a LPD podría promover la síntesis de NEAA como la tirosina, el ácido glutámico y el ácido aspártico, y el ácido pantoténico, el precursor de la coenzima A y la proteína transportadora de acilo, para promover la acumulación de proteínas y el crecimiento animal (Figura 3D).
4. Conclusión
La restricción proteica a los cerdos de engorde mejoró la deposición de grasa intramuscular, las actividades de las enzimas glucolíticas GPa y PK y la calidad de la carne, pero disminuyó el contenido de glucógeno y las actividades de las enzimas metabólicas oxidativas CS y CcO en el músculo esquelético porcino en comparación con las dietas normales. La suplementación con glicina en una dieta baja en proteínas promovió la transición de la fibra muscular del tipo II al tipo I, y mejoró aún más la ternura de la carne de cerdo y la calidad sensorial general. El análisis metabólico reveló que las dietas afectaban no solo a los nutrientes y al metabolismo energético in vivo, sino también al metabolismo energético y a la proteólisis en el músculo postmortem. El análisis de la vía sugirió que las dietas bajas en proteínas probablemente causaron insuficiencia de tirosina y sus metabolitos, lo que podría conducir a una disminución de la deposición de proteínas musculares y un deterioro del rendimiento del crecimiento animal, y una reducción de la emisión de nitrógeno a través del ciclo de la urea. Las dietas bajas en proteínas también disminuyeron la abundancia de acetil-CoA en el músculo, lo que puede estar involucrado en el aumento de la movilización de lípidos y regular a la baja la acumulación de glucógeno en el músculo. La suplementación de glicina con dietas bajas en proteínas aumentó el contenido muscular de tirosina, ácido glutámico y ácido N-acetil-aspártico, lo que podría promover la síntesis de NEAA y ácido pantoténico, guardar EAA para la síntesis de NEAA y, por lo tanto, promover la acumulación de proteínas y el crecimiento animal. Se necesitan más estudios para comprender el mecanismo detallado de la transición de miofibras inducida por la glicina y la mejora del rendimiento del crecimiento animal, que están directamente relacionados con la eficiencia de la producción de carne.
Declaración de disponibilidad de datos
Los datos presentados en el estudio se encuentran depositados en el repositorio Metabolights, número de acceso MTBLS7345. https://www.ebi.ac.uk/metabolights/MTBLS7345.
Declaración ética
El estudio en animales fue revisado y aprobado por el Comité de Gestión y Revisión de Protocolo del Instituto de Agricultura Subtropical de la Academia China de Ciencias.
Contribuciones de los autores
SJ: curación de datos, análisis formal e investigación. WQ: software y metodología. JL: análisis formal y análisis formal. AL: metodología. XZ: recursos. FL: supervisión y validación. QS: adquisición de fondos, conceptualización y redacción: borrador original. No se declaran conflictos de interés para ninguno de los autores. Todos los autores contribuyeron al artículo y aprobaron la versión presentada.
Financiación
Este estudio fue apoyado por el Programa Nacional de Investigación y Desarrollo Clave de China [Subvención n.º 2018YFD0500405] y el Proyecto Nacional de Desarrollo de Disciplinas Superiores para Equipos de Innovación [Subvención n.º kxk201801004].
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2023.1170573/full#supplementary-material
Notas
1. ^http://www.stats.gov.cn/xxgk/sjfb/tjgb2020/201902/t20190228_1768642.html
2. ^https://www.mzcloud.org/
3. ^http://www.chemspider.com/
Referencias
1. Alfaia, C. M., Lopes, P. A., Madeira, M. S., Pestana, J. M., Coelho, D., Toldrá, F., et al. (2019), «Estrategias de alimentación actuales para mejorar el contenido de grasa intramuscular del cerdo y su calidad nutricional». 89, 53–94, doi: 10.1016/bs.afnr.2019.03.006
Resumen de PubMed | Texto completo de CrossRef | Google Académico
2. Hocquette, J. F., Gondret, F., Baéza, E., Médale, F., Jurie, C., y Pethick, D. W. (2010), «Contenido de grasa intramuscular en animales productores de carne: desarrollo, control genético y nutricional, e identificación de marcadores putativos». Animal, 4:303–319, doi: 10.1017/S1751731109991091
Resumen de PubMed | Texto completo de CrossRef | Google Académico
3. DeVol, D. L., McKeith, F. K., Bechtel, P. J., Novakofski, J., Shanks, R. D. y Carr, T. R. (1988), «Variación en los rasgos de composición y palatabilidad y relaciones entre las características musculares y la palatabilidad en una muestra aleatoria de canales de cerdo». J. Anim. Sci., 66, 385–395. doi: 10.2527/jas1988.662385x
4. Daza, A., Mateos, A., Rey, A. I., Ovejero, I., y López-Bote, C. J. (2007), «Efecto de la duración de la alimentación en condiciones de pastoreo sobre los resultados de producción y la calidad de la canal y la grasa en cerdos ibéricos». Meat Sci., 76, 411–416, doi: 10.1016/j.meatsci.2006.10.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
5. Madeira, M. S., Pires, V. M. R., Alfaia, C. M., Costa, A. S. H., Luxton, R., Doran, O., et al. (2013), «Efectos diferenciales de las dietas reducidas en proteínas sobre la composición de ácidos grasos y la expresión génica en el músculo y el tejido adiposo subcutáneo de cerdos cruzados de raza pura Alentejana y blancos grandes x raza autóctona x Pietrain. H. J. Nutr., 110, 216–229, doi: 10.1017/S0007114512004916
Resumen de PubMed | Texto completo de CrossRef | Google Académico
6. Castell, A. G., Cliplef, R. L., Poste-Flynn, L. M. y Butler, G. (1994), «Rendimiento, características de la canal y de la carne de cerdo de las dietas autoalimentadas de castrados y primerizas que difieren en el contenido de proteínas y la relación lisina:energía». Can. J. Anim. Sci., 74, 519–528, doi: 10.4141/cjas94-073
7. Goerl, K. F., Eilert, S. J., Mandigo, R. W., Chen, H. Y. y Miller, P. S. (1995), «Características de la carne de cerdo afectadas por dos poblaciones de cerdos y seis niveles de proteína bruta». J. Anim. Sci., 73, 3621–3626. doi: 10.2527/1995.73123621x
8. Kerr, B. J., McKeith, F. K., y Easter, R. A. (1995), ‘Efecto sobre el rendimiento y las características de la canal de los cerdos de matriz a engorde alimentados con proteínas brutas reducidas, dietas suplementadas con aminoácidos’, J. Anim. Sci., 73, 433–440, doi: 10.2527/1995.732433x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
9. Ma, X., Zheng, C., Hu, Y., Wang, L., Yang, X. y Jiang, Z. (2015), «La suplementación dietética con L-arginina afecta al proteoma del músculo Longissimus esquelético en cerdos de engorde». PLoS Uno, 10:e0117294. doi: 10.1371/journal.pone.0117294
10. Tan, B., Yin, Y., Liu, Z., Li, X., Xu, H., Kong, X., et al. (2009), ‘La suplementación dietética con l-arginina aumenta la ganancia muscular y reduce la masa grasa corporal en cerdos en crecimiento y engorde’, Aminoácidos, 37, 169–175, doi: 10.1007/s00726-008-0148-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Madeira, M. S., Alfaia, C. M., Costa, P., Lopes, P. A., Lemos, J. P. C., Bessa, R. J. B., et al. (2014), «La combinación de suplementos de arginina y leucina de dietas reducidas de proteínas brutas para verracos aumenta la calidad alimentaria de la carne de cerdo», J. Anim. Sci., 92, 2030–2040, doi: 10.2527/jas.2013-6876
12. Tous, N., Lizardo, R., Vilà, B., Gispert, M., Font-i-Furnols, M., y Esteve-Garcia, E. (2016), «Adición de arginina y leucina a dietas proteicas bajas o normales: rendimiento, características de la canal y grasa intramuscular de cerdos de engorde», Span. J. Agric. Res., 14, doi: 10.5424/sjar/2016144-9351
13. Dean, D., Bidner, T. D. y Southern, L. L. (2006), «La suplementación con glicina a las dietas bajas en proteínas y suplementadas con aminoácidos apoya el rendimiento óptimo de los pollos de engorde», Poult. Sci., 85, 288–296, doi: 10.1093/ps/85.2.288.
14. Ospina-Rojas, I. C., Murakami, A. E., Oliveira, C. A. L. y Guerra, A. F. Q. G. (2013), «Efectos suplementarios de glicina y treonina en el rendimiento, el desarrollo de la mucosa intestinal y la utilización de nutrientes de los pollos de engorde en crecimiento», Poult. Sci., 92, 2724–2731, doi: 10.3382/ps.2013-03171
Resumen de PubMed | Texto completo de CrossRef | Google Académico
15. Monin, G., y Sellier, P. (1985), «Carne de cerdo de baja calidad tecnológica con una tasa normal de caída del pH muscular en el período post-mortem inmediato: el caso de la raza Hampshire», Meat Sci., 13, 49–63, doi: 10.1016/S0309-1740(85)80004-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
16. Hwang, Y. H., Kim, G. D., Jeong, J. Y., Hur, S. J., and Joo, S. T. (2010), ‘The relationship between muscle fiber characteristics and meat quality traits of highly marbled Hanwoo (Korean native cattle) steers’, Meat Sci., 86, 456–461, doi: 10.1016/j.meatsci.2010.05.034
17. Guo, Q., Kong, X., Hu, C., Zhou, B., Wang, C., and Shen, Q. W. (2019), ‘Fatty acid content, flavor compounds, and sensory quality of pork loin as affected by dietary supplementation with l-arginine and glutamic acid’, J. Food Sci., 84, 3445–3453, doi: 10.1111/1750-3841.14959
18. Scollan, N. D., Dannenberger, D., Nuernberg, K., Richardson, I., MacKintosh, S., Hocquette, J. F., et al. (2014), ‘Enhancing the nutritional and health value of beef lipids and their relationship with meat quality’, Meat Sci., 97:384–394, doi: 10.1016/j.meatsci.2014.02.015
19. Calkins, C. R., and Hodgen, J. M. (2007), ‘A fresh look at meat flavor’, Meat Sci., 77, 63–80, doi: 10.1016/j.meatsci.2007.04.016
20. Brooke, M. H., and Kaiser, K. K. (1970), ‘Muscle fiber types: how many and what kind?’, Arch. Neurol., 23, 369–379, doi: 10.1001/archneur.1970.00480280083010
21. Lind, A., and Kernell, D. (1991), ‘Myofibrillar ATPase histochemistry of rat skeletal muscles: a «two-dimensional» quantitative approach’, J. Histochem. Cytochem., 39, 589–597, doi: 10.1177/39.5.1826695
22. Kim, G. D., Yang, H. S., and Jeong, J. Y. (2018), ‘Intramuscular variations of proteome and muscle fiber type distribution in semimembranosus and semitendinosus muscles associated with pork quality’, Food Chem., 244, 143–152, doi: 10.1016/j.foodchem.2017.10.046
23. Li, Y., Li, J., Zhang, L., Yu, C., Lin, M., Gao, F., et al. (2015), «Efectos de las fuentes de energía dietética en la glucólisis post mortem, la calidad de la carne y la transformación del tipo de fibra muscular de los cerdos de engorde», PLoS One, 10,:e0131958, doi: 10.1371/journal.pone.0131958
Resumen de PubMed | Texto completo de CrossRef | Google Académico
24. Zhang, S. H., Zhu, L., Wu, Z. H., Zhang, Y., Tang, G. Q., Jiang, Y. Z., et al. (2013), «Efecto del tipo de fibra muscular en la expresión génica de la glucogenina-1 y su relación con el potencial glucolítico y el pH de la carne de cerdo», Genet. Mol. Res., 12, 3383–3390, doi: 10.4238/2013.septiembre.4.4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
25. Giannenas, I., Grigoriadou, K., Sidiropoulou, E., Bonos, E., Cheilari, A., Vontzalidou, A., et al. (2021), Perfil metabólico UHPLC-MS no dirigido como herramienta valiosa para la evaluación de los parámetros de calidad de los huevos después de la suplementación dietética con orégano, tomillo, té de sideritis y manzanilla en gallinas ponedoras marrones. Metabolómica, 17:51, doi: 10.1007/s11306-021-01801-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
26. Rocchetti, G., Becchi, P. P., Salis, L., Lucini, L. y Cabiddu, A. (2022), Impacto de las dietas basadas en pastos en el perfil metabolómico no dirigido de la leche de oveja Sarda. Alimentos, 12:143, doi: 10.3390/foods12010143
Resumen de PubMed | Texto completo de CrossRef | Google Académico
27. Wang, H., Xia, P., Lu, Z., Su, Y. y Zhu, W. (2023), La alimentación restringida en el tiempo afecta el perfil transcriptómico del hipotálamo en cerdos a través de la regulación del metabolismo de los aminoácidos aromáticos. J. Sci. Food Agric., 103:1578–1587, doi: 10.1002/jsfa.12256
Resumen de PubMed | Texto completo de CrossRef | Google Académico
28. Zhou, X., Liu, Y., Zhang, L., Kong, X. y Li, F. (2021), Las proporciones de serina a glicina en dietas bajas en proteínas regulan la grasa intramuscular al afectar el metabolismo de los lípidos y la transición del tipo de miofibra en el músculo esquelético de cerdos en crecimiento y engorde. Anim. Nutr. 7:384–392. doi: 10.1016/j.aninu.2020.08.011
29. Wang, X., Wei, H., Cao, J., Li, Z. y He, P. (2015), «Análisis metabolómico del músculo de lechones alimentados con dietas bajas en proteínas suplementadas con aminoácidos de cadena ramificada mediante HPLC-MS de alta resolución», Electroforesis, 36, 2250–2258, doi: 10.1002/elps.201500007
Resumen de PubMed | Texto completo de CrossRef | Google Académico
30. Barboriak, J. J., Krehl, W. A., y Cowgill, G. R. (1957), «Necesidad de ácido pantoténico de la rata adulta y en crecimiento», J. Nutr., 61, 13-21, doi: 10.1093/jn/61.1.13
31. Qian, Y., Li, X. F., Zhang, D. D., Cai, D. S., Tian, H. Y. y Liu, W. B. (2015), «Efectos del ácido pantoténico dietético sobre el crecimiento, la función intestinal, el estado antioxidante y la síntesis de ácidos grasos de la dorada juvenil Megalobrama amblycephala», PLoS One, 10:e0119518, doi: 10.1371/journal.pone.0119518
Resumen de PubMed | Texto completo de CrossRef | Google Académico
32. Wu, G. Y. (2010), ‘Aminoácidos funcionales en el crecimiento, la reproducción y la salud’, Adv. Nutr., 1, 31–37, doi: 10.3945/an.110.1008
Resumen de PubMed | Texto completo de CrossRef | Google Académico
33. Perry, R. J., Camporez, J. P. G., Kursawe, R., Titchenell, P. M., Zhang, D., Perry, C. J., et al. (2015), «El acetil CoA hepático vincula la inflamación del tejido adiposo con la resistencia a la insulina hepática y la diabetes tipo 2», Cells, 160, 745–758, doi: 10.1016/j.cell.2015.01.012
Keywords: low protein diet, glycine, metabolomics, growing-finishing pigs, meat quality
Citation: Jiang S, Quan W, Luo J, Lou A, Zhou X, Li F and Shen QW (2023) Low-protein diets supplemented with glycine improves pig growth performance and meat quality: An untargeted metabolomic analysis. Front. Vet. Sci. 10:1170573. doi: 10.3389/fvets.2023.1170573
Edited by:
Panagiotis E. Simitzis, Agricultural University of Athens, Greece
Reviewed by:
Ilias Giannenas, Aristotle University of Thessaloniki, Greece
George P. Laliotis, Agriculture University of Athens, Greece
Biao Li, Southwest Minzu University, China
Derechos de autor © 2023 Jiang, Quan, Luo, Lou, Zhou, Li y Shen. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Fengna Li, lifengna@isa.ac.cn; Qingwu W. Shen, yaoyao3153@aliyun.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía