
Cambios en los factores de virulencia mediados por la genética y la dieta de fibra en el contenido de colon y las heces de cerdo

Cambios en los factores de virulencia mediados por la genética y la dieta de fibra en el contenido de colon y las heces de cerdo y sus factores impulsores
 Tao Wang1,2†
Tao Wang1,2†  Yuheng Luo1,2†
Yuheng Luo1,2†  Xiangfeng Kong3
Xiangfeng Kong3  Bing Yu1,2
Bing Yu1,2  Ping Zheng1,2
Ping Zheng1,2  Zhiqing Huang1,2
Zhiqing Huang1,2  Xiangbing Mao1,2
Xiangbing Mao1,2  Jie Yu1,2
Jie Yu1,2  JuHui Yan1,2
JuHui Yan1,2  Jun He1,2*
Jun He1,2*- 1Instituto de Nutrición Animal, Universidad Agrícola de Sichuan, Chengdu, China
- 2Laboratorio Clave de Nutrición Resistente a Enfermedades Animales, Chengdu, China
- 3Instituto de Agricultura Subtropical, Academia China de Ciencias, Changsha, China
Los factores de virulencia (FV) son factores clave para que los microorganismos establezcan mecanismos de defensa en el huésped y potencien su potencial patogénico. Sin embargo, el espectro de factores de virulencia en el colon y las heces de cerdo, así como la influencia de los factores dietéticos y genéticos en ellos, sigue sin informarse. En este estudio, en primer lugar, revelamos la diversidad, abundancia y características de distribución de las FV en el contenido colónico de diferentes razas de cerdos (cerdos Taoyuan, Xiangcun y Duroc) alimentados con diferentes niveles de fibra mediante un análisis metagenómico. El análisis dio como resultado la identificación de 1.236 factores de virulencia, que podrían agruparse en 16 características de virulencia. Entre estos, los cerdos Taoyuan exhibieron niveles significativamente más altos de factores de virulencia en comparación con los cerdos Duroc. La dieta alta en fibra redujo significativamente la abundancia de ciertas categorías de factores de virulencia, incluidos los sistemas de absorción de hierro (FbpABC, HitABC) y las categorías de proteasa Ig en el colon, junto con una disminución notable en la abundancia relativa de categorías de plásmidos en elementos genéticos móviles (MGE). Además, examinamos las FV en heces utilizando cuantificación absoluta. Los resultados mostraron que las dietas altas en fibra reducen la excreción fecal de FV y que este efecto está fuertemente influenciado por los MGE y los ácidos grasos de cadena corta (AGCC). Los experimentos de fermentación in vitro confirmaron que el ácido acético (AA) condujo a una disminución en la abundancia relativa de FV (p < 0,1). En conclusión, nuestros hallazgos revelan por primera vez cómo la dieta de fibra y los factores genéticos afectan la distribución de FV en el contenido de colon y las heces de cerdo y sus factores impulsores. Esta información proporciona valiosos datos de referencia para mejorar aún más la inocuidad de los alimentos y la sanidad animal.
Introducción
El tracto gastrointestinal y la materia fecal contienen diversas comunidades microbianas, con microorganismos patógenos que representan riesgos significativos para la salud de los seres humanos y los animales. Aproximadamente el 61% de los patógenos humanos pueden infectar a los animales, lo que pone de relieve la importancia mundial de las enfermedades zoonóticas (1). Los patógenos mejoran sus defensas y patogenicidad dentro de los huéspedes utilizando factores de virulencia (VF) (2, 3), que desempeñan un papel fundamental durante las infecciones y dictan la capacidad de los patógenos para causar enfermedades. Estas FV pueden interactuar directamente con los tejidos del huésped o ayudar a las bacterias a evadir las respuestas inmunitarias (4). Investigaciones anteriores han dilucidado los mecanismos por los cuales las FV impulsadas por bacterias dañan las células huésped, incluida la interrupción de la señalización celular, el desencadenamiento de respuestas inmunitarias inespecíficas y la activación de funciones proteasas específicas. Por ejemplo, las adhesinas codificadas por el gen FimA de Klebsiella pneumoniae promueven la adhesión microbiana (5), mientras que las toxinas producidas por Escherichia coli, como las hemolisinas, pueden inhibir las funciones de la célula huésped o causar lisis celular (6). Sin embargo, a pesar de la amplia investigación clínica sobre los factores de virulencia transmitidos por microorganismos patógenos, los estudios sobre la distribución de las FV en el tracto gastrointestinal y las heces de los animales destinados a la producción de alimentos siguen siendo limitados.
La materia fecal del ganado es un producto directo del contenido gastrointestinal. Sus propiedades fisicoquímicas y su composición microbiana están influenciadas por factores dietéticos e intestinales (7). La alteración de los hábitos alimenticios puede tener un impacto significativo en las comunidades microbianas dentro del tracto gastrointestinal y en las FV que poseen, afectando posteriormente a la excreción de FV en las heces (8). Por ejemplo, después de la infección con Citrobacter, un patógeno de la mucosa murina, en un modelo experimental de ratón, se encontró que la adición de hierro en la dieta induce un mecanismo de defensa cooperativo que debilita la virulencia del patógeno y promueve infecciones asintomáticas prolongadas al desencadenar cambios fisiológicos en el huésped (9, 10). Teniendo en cuenta esto, planteamos la hipótesis de que los cambios en la fibra dietética pueden alterar las actividades metabólicas en el contenido gastrointestinal, afectando a las FV presentes en los microorganismos intestinales e influyendo en los patrones de excreción de la FV.
Históricamente, la investigación de FV en bacterias patógenas ha dependido en gran medida de los métodos de PCR de un solo gen o PCR cuantitativa (qPCR) (11, 12). Sin embargo, estas técnicas se enfrentan a limitaciones técnicas, lo que dificulta una evaluación holística y rápida de la diversidad de la FV en las muestras. Con los avances en la genómica microbiana, especialmente en la secuenciación completa del genoma bacteriano, la metagenómica se ha convertido en una herramienta crucial para identificar las FV. Este enfoque ayuda a estudiar ampliamente las FV y a comprender la dinámica evolutiva de las FV, teniendo en cuenta diversas presiones de selección ambiental y la ganancia o pérdida de genes en escenarios patogénicos específicos (13). En nuestro estudio, seleccionamos las razas de cerdos Taoyuan, Duroc y Xiangcun como modelos y utilizamos una combinación de metagenómica de alto rendimiento y qPCR para caracterizar la extensión de las FV en el colon de diferentes razas de cerdos a diferentes niveles de fibra y para validar la excreción de FV en las heces. Y utilizamos la cromatografía líquida y el modelado de fermentación in vitro para explorar los posibles impulsores de los cambios en las FV en el contenido del colon y las heces. El estudio tuvo como objetivo caracterizar la distribución dietética y genéticamente mediada de las FV en el contenido colónico porcino y, además, investigar los impulsores de la fibra que influyen en la excreción de FV en las heces porcinas. Nuestro estudio proporciona datos de referencia importantes para la cría de animales y los estudios de salud alimentaria.
Materiales y métodos
Declaración ética
La gestión de los experimentos con animales que participen en la investigación se remitirá al «Reglamento sobre la administración de animales de laboratorio» (Ministerio de Ciencia y Tecnología de China, revisado en junio de 2004). La recolección de muestras fue aprobada por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Agrícola de Sichuan, Sichuan, China (No. 20181105), y se llevó a cabo en estricta conformidad con las pautas éticas.
Experimentación con animales y recogida de muestras
El experimento seleccionó a los 60 días de edad cerdos Taoyuan (Peso promedio: 13,87 ± 0,58 kg; Comprado a Xiangcun Hi-Tech Agriculture Co., Ltd.), cerdos Duroc (Peso promedio 18,50 ± 1,09 kg; Comprado a Linli Tianxin Seed Industry Co., Ltd) y sus variedades cruzadas de cerdos Xiangcun (Peso promedio 14,47 ± 0,15 kg; Comprado a Xiangcun Hi-Tech Agriculture Co., Ltd.), y adoptó un diseño factorial de 3 × 2, es decir, 3 variedades de cerdos (20 cerdos para cada variedad, es decir, cada 10 cerdos en cada grupo de tratamiento, 60 cerdos en total) fueron alimentados con una dieta alta en fibra (fibra bruta: 6-7%; energía digestible: 3,5%; proteína bruta: 19,16%) y una dieta baja en fibra (fibra cruda: 2–3%; Energía digestible: 3,49%; Proteína bruta: 19,15%). La fibra de salvado de trigo se compró a Chengdu Tubaite Technology Co., Ltd. (fabricante: JRS, modelo: WF200, pureza >95%). Todos los cerdos se alojaron en corrales individuales, cada corral estaba equipado con comederos y bebederos de tetina, la temperatura ambiente era de 28 °C, el agua y la comida eran gratuitos y el período experimental fue de 28 días. Los cerdos fueron sacrificados el último día del experimento y se recolectó el contenido colónico y las heces.
Secuenciación del metagenoma y análisis de datos
Las bibliotecas de extremos emparejados fueron construidas por NEXTFLEX Rapid DNA Seq (Bioo Scientific, Austin, TX, Estados Unidos) utilizando 1 μg de construcciones de ADN de alta calidad, y luego secuenciadas utilizando la plataforma Novaseq6000 de Shanghai Majorbio BioPharmaceuticals Biotechnology Co., Ltd. (Shanghai, China). Los datos sin procesar se filtraron utilizando Trimmomatic v0.38 para eliminar las lecturas que contenían más de tres nucleótidos ambiguos con una puntuación de calidad media de <20 y para eliminar las lecturas duplicadas artificialmente. A continuación, las lecturas limpias se ensamblaron en contigs utilizando MEGAHIT (14) (https://github.com/voutcn/megahit, versión 1.1.2). Solo se conservaron los conglomerados superpuestos de ≥300 pb para su posterior análisis. Los marcos de lectura abiertos (ORF) de todos los grupos de superposición ensamblados se predijeron utilizando MetaGene y se tradujeron en secuencias de aminoácidos. Se construyó un catálogo de genes no redundantes utilizando CD-HIT (15) (http://www.bioinformatics.org/cd-hit/, versión 4.6.1) con un 90% de identidad de secuencia y una cobertura del 90%. Las lecturas de alta calidad se alinearon con el catálogo de genes no redundantes para calcular la abundancia de genes con una identidad del 95% utilizando SOAPaligner (http://soap.genomics.org.cn/, versión 2.21). La anotación del factor de virulencia se realizó utilizando Diamond (16) (http://www.diamondsearch.org/index.php, versión 0.8.35 http://www.mgc.ac.cn/VFs/) e el valor de corte fue de 1 × 10−5. Los datos de secuencia asociados con este proyecto se han depositado en la base de datos NCBI Short Read Archive (Número de acceso: PRJNA849732).
Determinación de los AGCC
Se pesó aproximadamente 1 g de muestras (contenido de colon y muestras fecales) en un tubo de centrífuga de 2 mL, se agregó agua ultrapura en la proporción de 1:2.5, se mezcló bien y se dejó reposar durante 30 min. Se tomaron 2 mL de sobrenadante en un tubo de centrífuga nuevo y se incubaron durante 30 min a 4°C y 12.000 r/min durante 10 min. Se tomó 1 mL de sobrenadante y se añadieron 0,2 mL de solución de ácido metafosfórico al 25% (p/v), se mezcló bien y se incubó durante 30 min a 4 °C durante 10 min a 12.000 r/min. Se añadieron 0,2 mL de solución de ácido metafosfórico al 25% (p/v) a 1 mL del sobrenadante, se mezcló bien y se incubó a 4 °C durante 30 min. Incubado durante 10 min a 12.000 r/min durante 10 min. Se tomaron 0,6 mL del sobrenadante y se añadieron 11,67 μL de solución de ácido crotónico (210 mmol/L) al sobrenadante, se mezclaron bien y luego se introdujeron 0,3 mL de sobrenadante en un nuevo tubo centrífugo, y luego se añadieron 0,9 mL de solución de metanol. La concentración de AGCC en las muestras se determinó mediante un sistema de cromatografía de gases (CP-3800 GC, Varian, Inc., Walnut Creek, CA, Estados Unidos) y siguiendo el método de Franklin et al. (17).
qPCR
El ADN se extrajo de heces y muestras de fermentación in vitro de acuerdo con las instrucciones del Kit de Extracción de ADN Genómico Fecal (D2700, Solarbio, China). La concentración y la integridad de las muestras de ADN se determinaron mediante el uso de un microespectrofotómetro NanoDrop (NanoDrop 2000, NanoDrop, Estados Unidos) y Gel Imaging (GelDocXR, Bio-Rad, Estados Unidos) para detectar la concentración, pureza e integridad de las muestras de ADN. El análisis cuantitativo del gen de la FV se realizó mediante qPCR utilizando ADN fecal como molde (instrumento de cuantificación de fluorescencia en tiempo real CFX96, ABI7900, Estados Unidos). Se establecieron tres réplicas para cada muestra y se promediaron los resultados. El sistema de reacción consistió en 5 μL de SYBR Premix Ex Taq™ II, 0,5 μL de cada uno de los cebadores aguas arriba y aguas abajo, 2 μL de la plantilla de ADN y 2 μL de ddH2O. La abundancia de 46 genes en las muestras fecales se examinó mediante qPCR (QS6FX, ABI, Estados Unidos) (Tabla suplementaria S5) (11). La abundancia absoluta de FV se calculó según el método de Xie et al. (11).
Ensayo de fermentación in vitro
Se llevaron a cabo experimentos de fermentación in vitro de acuerdo con los métodos existentes (18). Los cerdos Duroc estuvieron en condiciones corporales normales durante 1 mes antes de la recolección fecal. Las heces colectadas se mezclaron fuertemente en condiciones asépticas con inóculo al 32% (p/v) (lodo fecal) en tampón fosfato, pH ajustado a 7.0 con HCl 0.1 M y luego filtrado a través de 4 capas de gasa. El filtrado se burbujeó con N2. Luego, se agregaron 8 mL de medio de fermentación, 2 mL de inóculo fecal y agente de tratamiento a cada tubo de fermentación de 50 mL bajo anaerobio (N2) y condiciones asépticas. A continuación, los tubos de fermentación se incubaron durante 20 h a 37 °C a 120 rpm. Después de 20 h de fermentación, la fermentación se detuvo por inmersión en agua helada durante 15 min y se recolectó el caldo de fermentación para el análisis de qPCR de FV y MGE. El medio de fermentación y la solución reductora se prepararon de acuerdo con el método establecido (18). Con base en el contenido de AGCC en las heces del grupo de dieta alta en fibra, se determinaron las concentraciones finales de AA, PA y BA agregadas en la fermentación in vitro, que fueron de 70 μmol/mL, 30 μmol/mL y 20 μmol/mL, respectivamente.
Estadística
El preprocesamiento de los datos se realizó con Excel 2019 (Microsoft, Estados Unidos) y las estadísticas de datos se realizaron con el programa SPSS 22.0 (IBM Corp., Estados Unidos). La visualización gráfica de los resultados se realizó utilizando GraphPad Prism 10. El valor RPKM se utilizó para los datos del mapa de calor, la puntuación Z se utilizó para la estandarización de los datos y el agrupamiento promedio se utilizó para el análisis de conglomerados. El análisis de correlación utilizó Spearman. Se utilizó la función de «protesta» en el envase vegano para analizar la correlación de Procusto entre el bacterioma y los FV. El análisis de coordenadas principales (PCoA) y el escalamiento multidimensional no métrico (NMDS) se realizaron utilizando el paquete de vegetación pura en el software R, utilizando FV y valores de abundancia normalizados de comunidades bacterianas con una distancia de Bray-Curtis. La distribución de los FV en bacterias de diferentes niveles taxonómicos se graficó como un diagrama de Sankey utilizando el paquete networkD31 en R (v3.6.2). Los mapas de calor de correlación de la prueba de Mantel se trazaron utilizando el paquete ggplot2 (3.3.3) en R (v4.1.3) y las dos correlaciones matriciales se probaron utilizando el paquete linkET (0.0.7.4). El gráfico de morera se dibuja utilizando el paquete plotly en el software R. El análisis de Circos se produjo y analizó utilizando las herramientas de la plataforma en la nube Megi Bio.
Lecturas por kilobase millones (RPKM):
Ri�Yo representa el valor de abundancia de Genei en una muestra, es decir, el número de lecturas alineadas con Genei en la muestra; Li�Yo representa la longitud de nucleótidos de Genei; ∑n1(Rj)∑1��� representa la suma de las lecturas correspondientes a todos los genes de la muestra.
Se utilizó un diseño experimental utilizando un diseño 3 × 2, ANOVA de dos vías para analizar dos efectos principales y efectos de interacción. Los resultados se expresaron como media ± error estándar, y p < 0,05 indicaron una diferencia significativa.
Resultado
Dieta de fibra y cambios mediados por especies en el espectro de FV
Para investigar los efectos de la dieta y la genética de la fibra en la diversidad, abundancia y características de distribución de las FV en el contenido colónico de diferentes razas de cerdos, realizamos análisis metagenómicos del contenido colónico de cerdos Duroc, Taoyuan y Xiangcun en una dieta alta y baja en fibra (Figura 1A). En el contenido colónico se identificaron 1.236 FV (Tabla Suplementaria S1). La clasificación mecanicista reveló que predominaron los factores de virulencia tanto ofensivos como defensivos, constituyendo más del 70% de las FV. No hubo una disparidad significativa en la abundancia general de FV en los diferentes niveles de fibra. Sin embargo, se evidenciaron claras diferencias en la abundancia de FV entre las razas, con cerdos Taoyuan y Xiangcun mostrando niveles elevados de FV (Figura 1B). El análisis de PCoA reveló un agrupamiento distintivo para los cerdos Duroc y Taoyuan a través de los niveles de fibra, mientras que los cerdos Xiangcun se parecían a ambas razas hasta cierto punto (Figuras 1C, D). A través de α análisis de diversidad, no se observaron diferencias notables en la abundancia o diversidad de FV con respecto a los niveles de fibra o razas (Figuras 1E, F). En el nivel 2, las FV se segregaron en 16 categorías en función de las características biológicas. Los factores más abundantes fueron la adherencia, la antifagocitosis, el sistema de captación de hierro, entre otros (Figura 2A; Figura complementaria S1). La dieta alta en fibra disminuyó notablemente la abundancia del sistema de absorción de hierro y la proteasa Ig (Figura suplementaria S1; Cuadro complementario S2). Entre las razas, los cerdos Taoyuan exhibieron niveles de abundancia significativamente más altos de las seis categorías de factores de virulencia en comparación con los cerdos Duroc (Figura Suplementaria S1; Cuadro complementario S2). Para superar las deficiencias de los modelos lineales y reflejar mejor la estructura no lineal, evaluamos la precisión del modelo con valores de tensión NMDS. Confirmamos que los valores de tensión eran inferiores a 0,2, lo que garantizaba la fiabilidad del modelo. El análisis NMDS mostró que en el grupo bajo en fibra, los cerdos Duroc y Taoyuan formaron grupos claramente diferenciados, mientras que los cerdos Xiangcun mostraron cierto grado de similitud con los cerdos Duroc y Taoyuan. En el grupo con alto contenido de fibra, los grupos de diferentes razas estaban más agrupados (Figuras 2B, C).
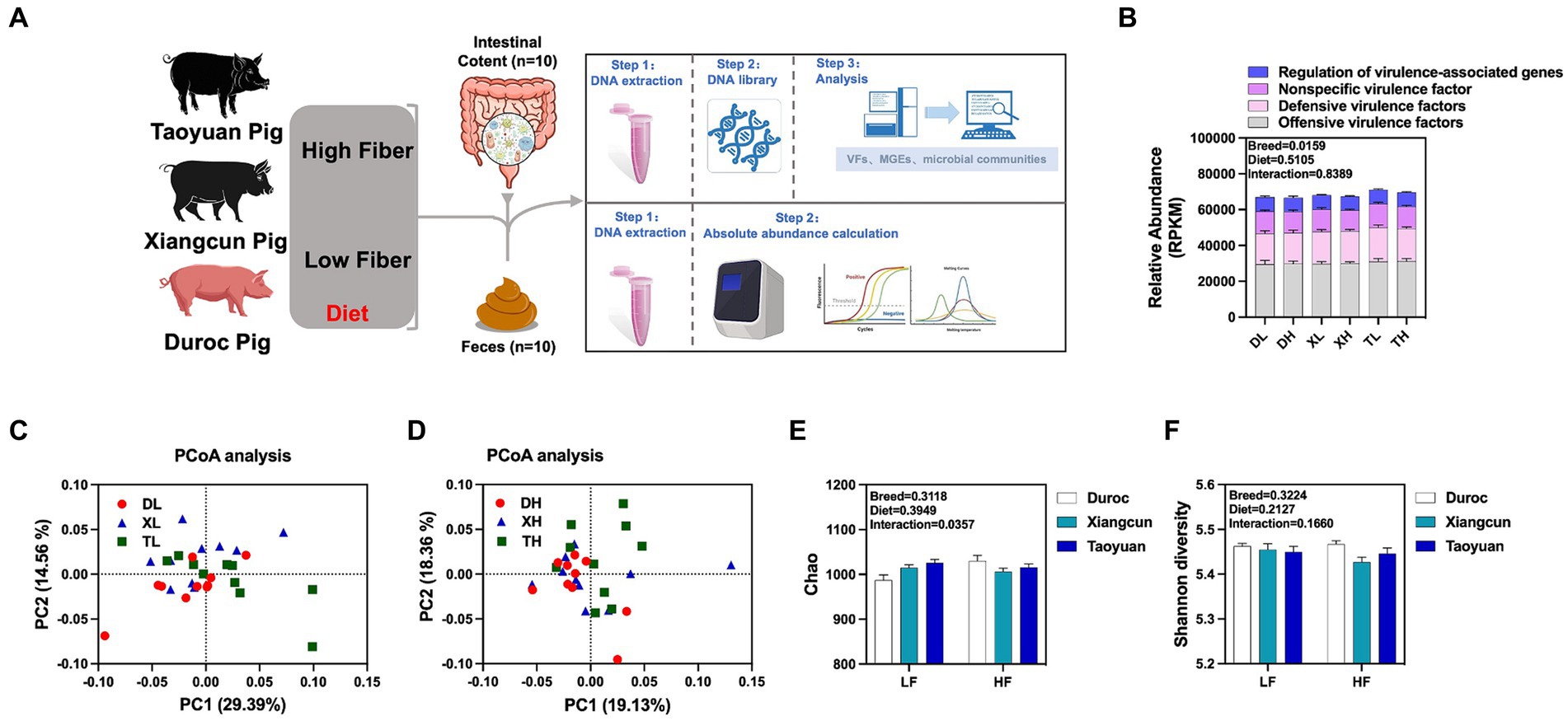 Figura 1. Efectos de diferentes niveles de fibra sobre la abundancia y diversidad de FV en el contenido colónico de diferentes razas de cerdos. Diagrama de flujo experimental (A). Abundancia total de FV en el contenido colónico (B). Análisis de PCoA (C,D) y análisis de diversidad alfa (E,F) de FV en el contenido colónico de diferentes razas porcinas a niveles altos y bajos de fibra. p < 0,05.*
Figura 1. Efectos de diferentes niveles de fibra sobre la abundancia y diversidad de FV en el contenido colónico de diferentes razas de cerdos. Diagrama de flujo experimental (A). Abundancia total de FV en el contenido colónico (B). Análisis de PCoA (C,D) y análisis de diversidad alfa (E,F) de FV en el contenido colónico de diferentes razas porcinas a niveles altos y bajos de fibra. p < 0,05.*
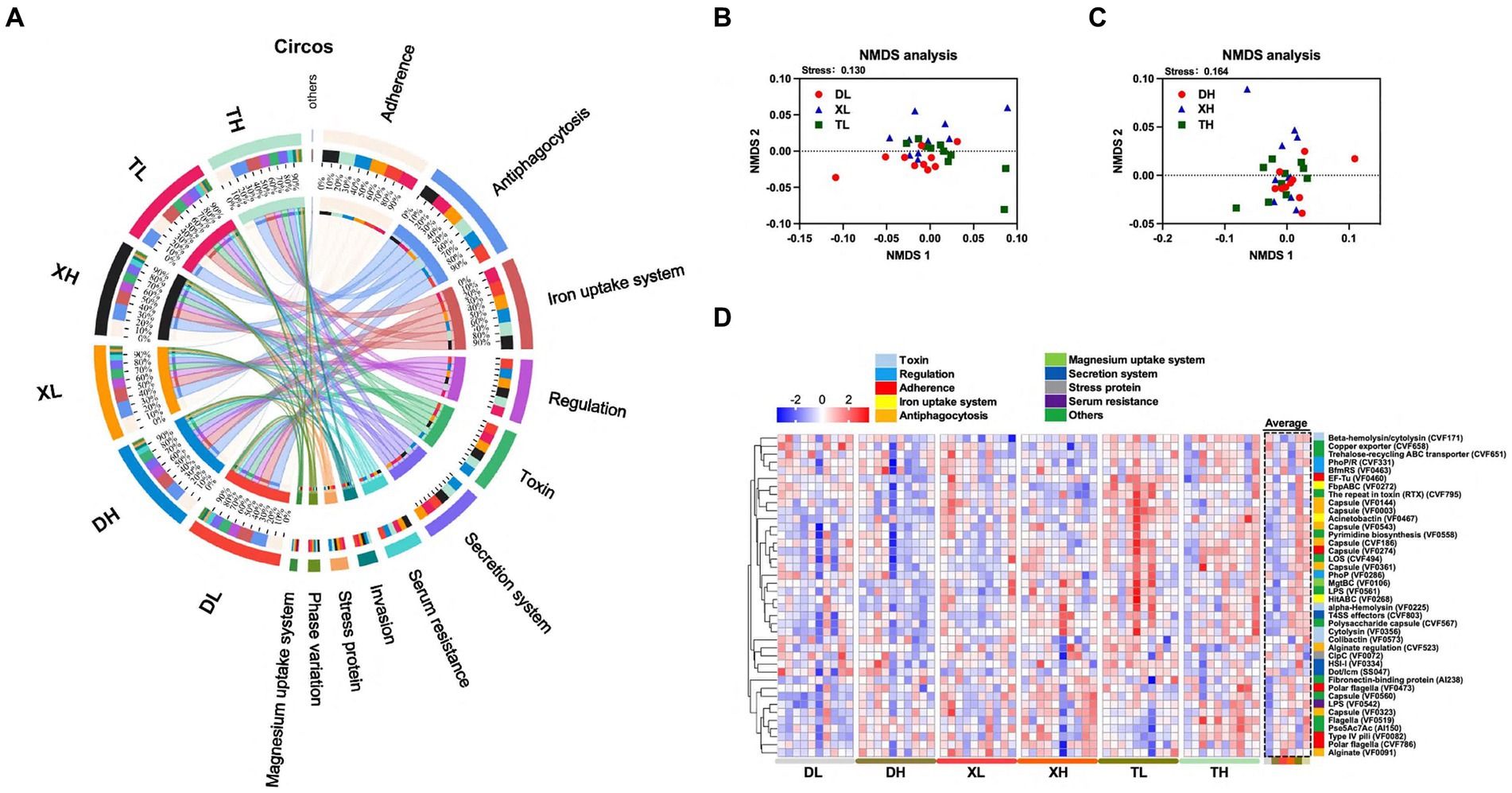 Figura 2. Efecto de diferentes niveles de fibra sobre la composición de FV en el contenido colónico de diferentes razas de cerdos. Análisis circos de FV en el contenido colónico (A). Análisis NMDS de FV en el contenido colónico de diferentes razas de cerdos a niveles altos y bajos de fibra (B,C). Análisis de mapa de calor del enriquecimiento de FV de los 40 genes principales con abundancia media relativa en diferentes muestras y grupos (D). Los diferentes colores en el lado izquierdo del mapa de calor indican diferentes categorías de VF.
Figura 2. Efecto de diferentes niveles de fibra sobre la composición de FV en el contenido colónico de diferentes razas de cerdos. Análisis circos de FV en el contenido colónico (A). Análisis NMDS de FV en el contenido colónico de diferentes razas de cerdos a niveles altos y bajos de fibra (B,C). Análisis de mapa de calor del enriquecimiento de FV de los 40 genes principales con abundancia media relativa en diferentes muestras y grupos (D). Los diferentes colores en el lado izquierdo del mapa de calor indican diferentes categorías de VF.
Ampliando nuestro estudio a las influencias de la raza y la fibra en las FV intestinales de cerdo, examinamos la abundancia de las 40 FV principales. El análisis reveló diferencias entre razas en 18 de las 40 principales FV, con cerdos Taoyuan exhibiendo mayores abundancias en comparación con los cerdos Duroc (Figura 2D; Cuadro complementario S3). Estas diferencias incluyen LOS (CVF494), cápsula (VF0543), beta-hemolisina/citolisina (CVF171), LPS (VF0542), HitABC (VF0268), FbpABC (VF0272) cápsula (VF0274), cápsula de polisacárido (CVF567), proteína de unión a fibronectina (AI238), citolisina (VF0356), biosíntesis de pirimidina (VF0558), alfa-hemolisina (VF0225), efectores T4SS (CVF803), cápsula (VF0003), la repetición en toxina (RTX) (CVF795), EF-Tu (VF0460), acinetobactina (VF0467) y cápsula (VF0560). En un análisis de diferentes niveles de fibra, una dieta alta en fibra redujo significativamente 7 de los 40 principales FV enriquecidos (HitABC (VF0268), FbpABC (VF0272), acondicionamiento de alginato (CVF523), cápsula (VF0144), encapsulante (VF0003), RTX (CVF795) y exportador de cobre (CVF658)) (Figura 2D; Cuadro complementario S3). Los resultados mostraron que la abundancia de FV fue mayor en el contenido colónico de los cerdos locales bajo el mismo patrón de alimentación a escala, mientras que la dieta alta en fibra redujo la abundancia de algunas FV.
Relación entre la cinética de clasificación bacteriana y los factores de virulencia
Las FV intestinales se originan principalmente en la microbiota intestinal. Estos se analizaron a través de una evaluación comparativa de los datos microbianos metagenómicos con la base de datos VFDB. Los 10 principales microorganismos a nivel de género presentes en el colon porcino, que constituyen más del 40% de la población, incluyeron Lactobacillus, Clostridium, Prevotella, Treponema, Oscillibacter, Ruminococcus, unclassified_f_Lachnospiraceae, unclassified_o_Clostridiales, unclassified_p_Firmicutes y unclassified_f_Ruminococcaceae (Figura 3A). El análisis de la diversidad alfa indicó que una dieta alta en fibra mejoró sustancialmente la diversidad microbiana, sin disparidades marcadas entre especies. Un análisis de Procrustes reveló una correlación significativa entre las FV y los taxones bacterianos, mostrando que la composición de las FV se alineaba estrechamente con la composición bacteriana (m2 = 0,1132, p = 0,001) (Figura 3B). Las FV se distribuyeron ampliamente entre una variedad de bacterias, principalmente del filo Firmicutes (Clostridium y Bacillus), el filo Bacteroidetes (principalmente Bacteroidia y Flavobacteriia), el filo Proteobacteria (principalmente Gammaproteobacteria y Betaproteobacteria) y el filo Actinobacteria (principalmente Actinobacteria y Coriobacteriia) (Figura 3C). El análisis de correlación entre la abundancia de especies y la abundancia funcional se realizó con base en las especies relativas y la abundancia funcional de las muestras. Identificamos microorganismos a nivel de especie que fueron los principales contribuyentes en la abundancia de FV, que fueron principalmente s_Lactobacillus_johnsonii, s_Clostridiales_bacterium, s_Oscillibacter_sp., s_Lachnospiraceae_bacterium, s_Lactobacillus_reuteri, s_uncultured_Clostridium_sp., s_Phascolarctobacterium_succinatutens, s_Lactobacillus_amylovorus, s_Faecalibacterium_prausnitzii y s_Alistipes_sp._CAG:435 (Figura 3D).
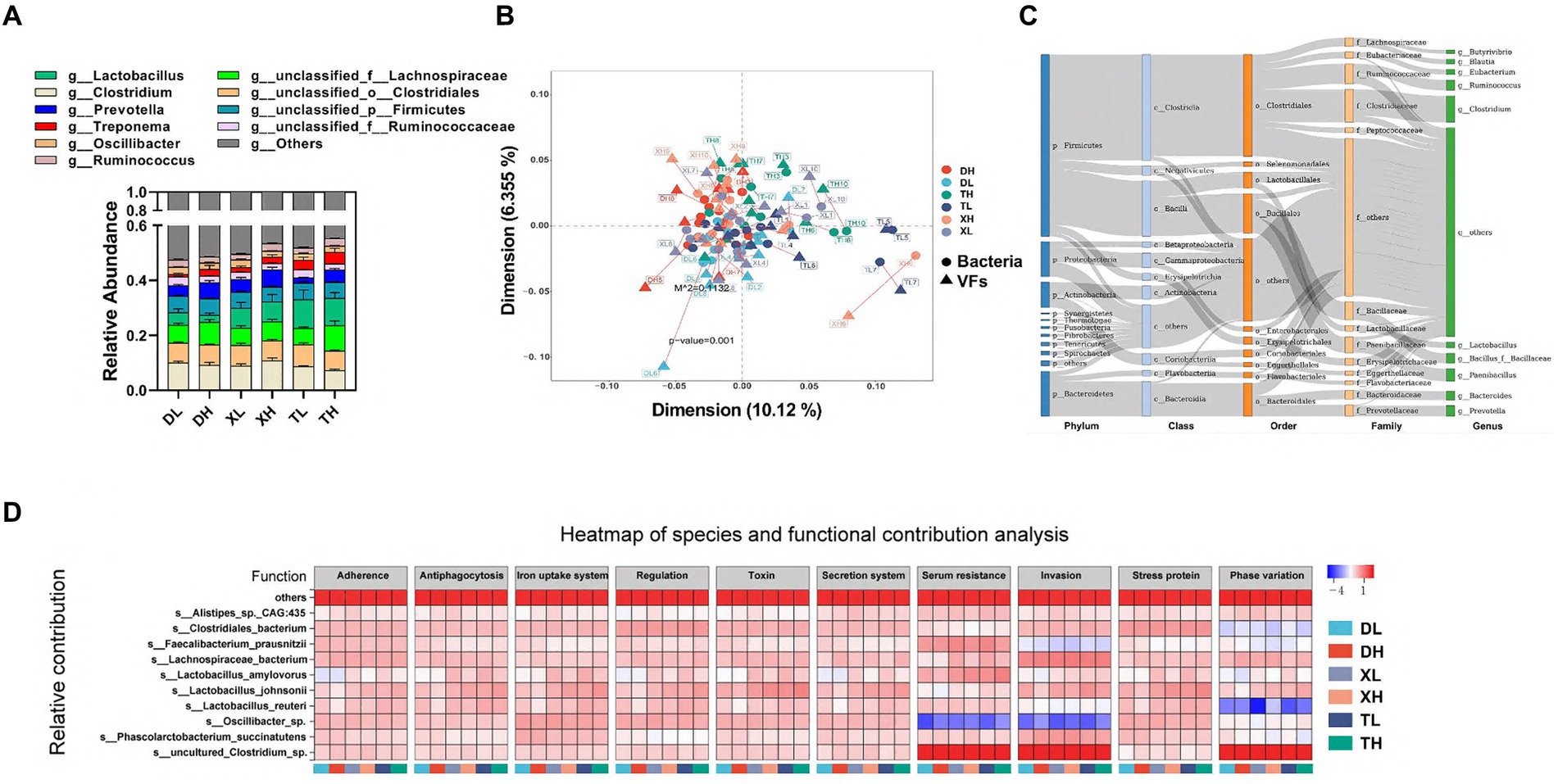 Figura 3. Correlación de las FV con los microbios intestinales. Composición microbiana a nivel de género del contenido de colon de cerdo de diferentes grupos (A). Procrustes analizó la correlación entre las especies bacterianas intestinales y la abundancia de FV (B). Los puntos indican la posición clasificada de la abundancia de especies bacterianas en cada muestra, y los triángulos indican la posición clasificada de la abundancia de FV. La longitud de la línea entre los puntos y los triángulos muestra los residuos de Procusto. (C) Distribución de FV en diferentes clases de bacterias (se eliminaron del análisis microorganismos no clasificados). Los colores de los rectángulos representan los diferentes niveles de clasificación. La longitud del rectángulo representa el número de FV. Los 10 principales microorganismos a nivel de especie que contribuyen a la abundancia de FV (D).
Figura 3. Correlación de las FV con los microbios intestinales. Composición microbiana a nivel de género del contenido de colon de cerdo de diferentes grupos (A). Procrustes analizó la correlación entre las especies bacterianas intestinales y la abundancia de FV (B). Los puntos indican la posición clasificada de la abundancia de especies bacterianas en cada muestra, y los triángulos indican la posición clasificada de la abundancia de FV. La longitud de la línea entre los puntos y los triángulos muestra los residuos de Procusto. (C) Distribución de FV en diferentes clases de bacterias (se eliminaron del análisis microorganismos no clasificados). Los colores de los rectángulos representan los diferentes niveles de clasificación. La longitud del rectángulo representa el número de FV. Los 10 principales microorganismos a nivel de especie que contribuyen a la abundancia de FV (D).
Para dilucidar la relación entre la flora intestinal y los FV, realizamos un análisis de redes de interacción utilizando el software Network. Este análisis tuvo como objetivo evaluar las interacciones y correlaciones entre los 100 principales géneros de microorganismos y FV dentro de la misma muestra. En la gráfica de centralidad de nodos, los números del eje x representan diferentes nodos. Cada nudo corresponde a la interacción de un solo género con otros géneros. Cuando un pico aparece simultáneamente en las tres curvas entre Degree_Centrality, Closeness_Centrality y Betweenness_Centrality, consideramos que el género correspondiente a ese nodo puede ser importante para toda la red. Sobre la base de este principio, identificamos 6 géneros, incluidos g_Lactobacillus, g_unclassified_f_Lachnospiraceae, g_Roseburia, g_unclassified_c_Clostridia, g_ Mediterraneibacter y g_Acetivibrio, así como tres VF, incluidos LPS (VF0542), transportador ABC de reciclaje de rehalosa T (CVF651) y BfmRS (VF0463). Este resultado confirmó aún más la importante influencia de la flora intestinal en la distribución de las FV (Figuras 4A, B; Cuadro complementario S4).
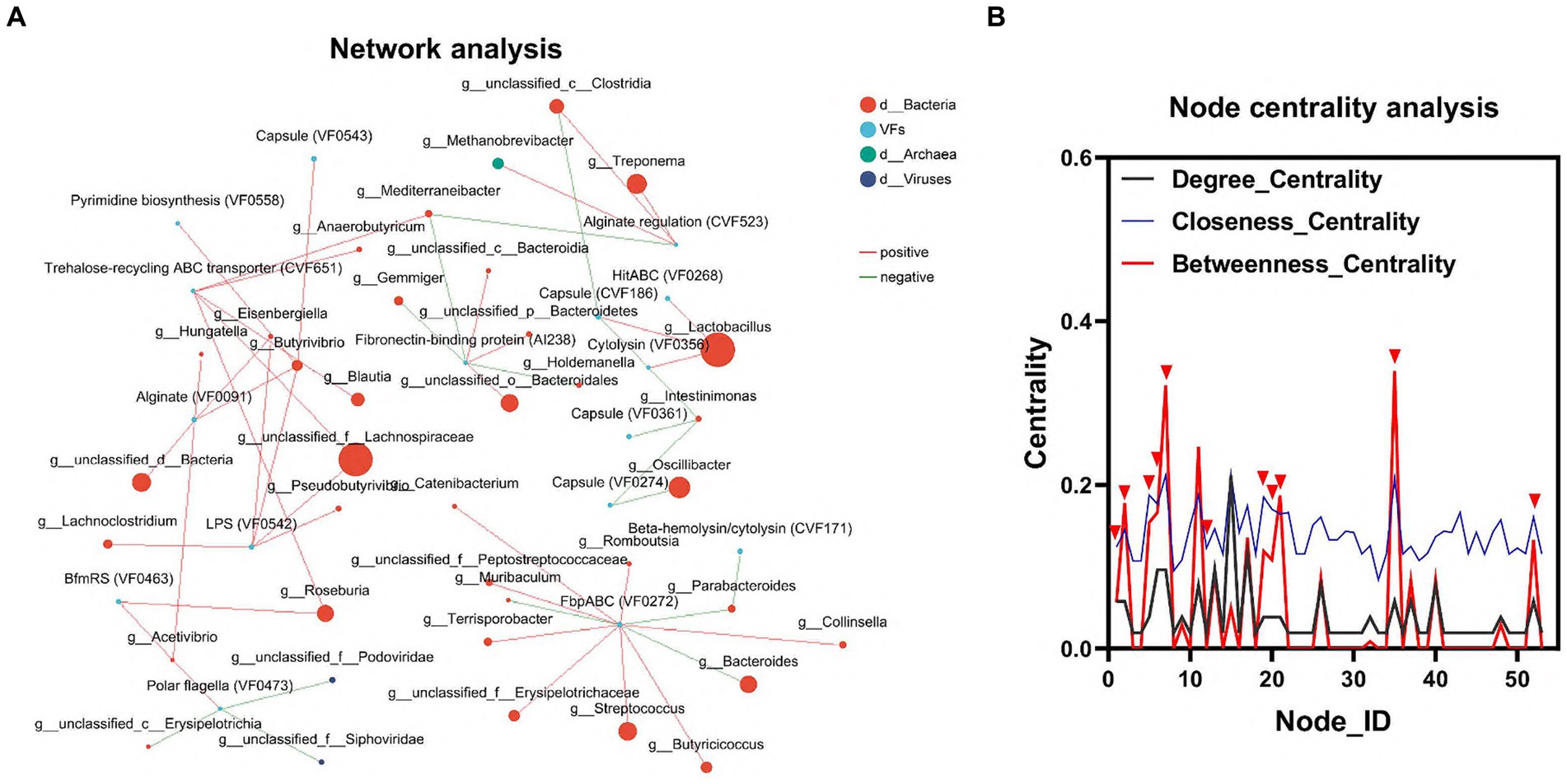 Figura 4. Interacción de la microbiota colónica y las FV. (A) Diagrama de red de la interacción entre la microbiota dominante y las FV. (B) Se analizaron los ganglios de deformación críticos del género principal. Las curvas negras indican centralidad de grados. Las curvas azules indican centralidad de proximidad y las curvas rojas indican centralidad mediada. Los nodos en los que las tres curvas alcanzan su punto máximo simultáneamente pueden ser géneros que desempeñan un papel importante en la red de interacción.
Figura 4. Interacción de la microbiota colónica y las FV. (A) Diagrama de red de la interacción entre la microbiota dominante y las FV. (B) Se analizaron los ganglios de deformación críticos del género principal. Las curvas negras indican centralidad de grados. Las curvas azules indican centralidad de proximidad y las curvas rojas indican centralidad mediada. Los nodos en los que las tres curvas alcanzan su punto máximo simultáneamente pueden ser géneros que desempeñan un papel importante en la red de interacción.
Análisis integrado de microorganismos, AGCC y MGEs con factores de virulencia
Examinamos las disparidades en la abundancia de clases funcionales de MGE en el contenido colónico de diferentes razas porcinas sometidas a diferentes niveles de ingesta de fibra. Nuestros hallazgos revelaron que una dieta alta en fibra disminuyó notablemente la abundancia relativa de plásmidos (p < 0.05) sin afectar el isfinder, el integrall o el iceberg. Curiosamente, las categorías de MGEs dentro de los contenidos colónicos porcinos no mostraron variaciones significativas entre razas (Figura 5A). El análisis de correlación lineal indicó una notable asociación positiva (p < 0,05) entre la abundancia de MGE y FV (Figura 5B). Para determinar la correlación de la diversidad microbiana y los MGE con los FV, evaluamos la relación entre la diversidad microbiana y la abundancia de EG y las diferentes clasificaciones de FV. Los resultados del ensayo de Mantel mostraron que los MGE se asociaron con la antifagocitosis, el sistema de absorción de hierro, la regulación, la toxina, la resistencia sérica, la exoenzima y la antiproteólisis, la riqueza de bacterias se correlacionó significativamente positivamente con la adherencia, el sistema de secreción y la proteína de estrés, la diversidad bacteriana se correlacionó significativamente positivamente con la antifagocitosis, el sistema de absorción de hierro, la resistencia sérica, la variación de fase, el sistema de absorción de magnesio y la motilidad basada en actina ( Figura 5C).
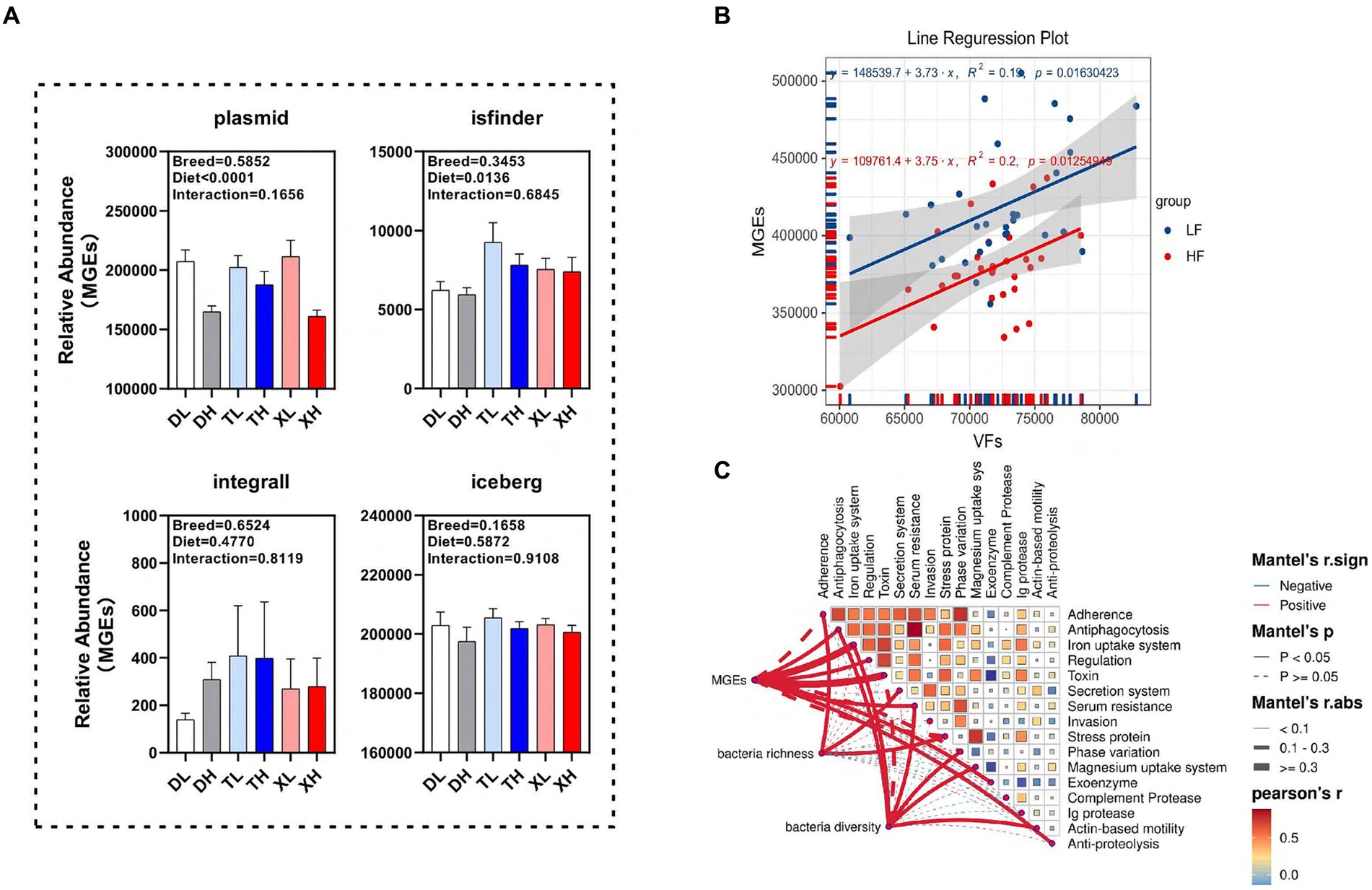 Figura 5. Posibles impulsores que median los cambios en las FV. (A) Diferencias en diferentes clases de MGE (plásmido, isfinder, integral e iceberg) en el contenido colónico de diferentes grupos de cerdos. Análisis de regresión lineal de la abundancia relativa de FV y MGE en el contenido colónico a diferentes niveles de fibra (B). Análisis del manto de MGES, abundancia microbiana y diversidad microbiana y diferentes categorías de FV (C).
Figura 5. Posibles impulsores que median los cambios en las FV. (A) Diferencias en diferentes clases de MGE (plásmido, isfinder, integral e iceberg) en el contenido colónico de diferentes grupos de cerdos. Análisis de regresión lineal de la abundancia relativa de FV y MGE en el contenido colónico a diferentes niveles de fibra (B). Análisis del manto de MGES, abundancia microbiana y diversidad microbiana y diferentes categorías de FV (C).
Efectos de las dietas de razas y fibras en la distribución de FV en el estiércol porcino y sus posibles impulsores
Se detectaron 46 subtipos de FV en las heces de diferentes razas de cerdos por cuantificación absoluta. Estas FV se clasificaron principalmente en 22 categorías por función, que incluyen adherencia, antivirulencia, autotransportador, formación de biopelículas, cápsula, determinantes de adherencia fimbrial, evasión inmune, invasión, absorción de hierro, efectores TTSS codificados por LEE, genes inducibles por macrófagos, absorción de magnesio, determinantes de adherencia no fimbrial, efectores TTSS no codificados por LEE, factor nutricional, proteasa, regulación, sistema de secreción, resistencia sérica, locus Spv, adaptación al estrés y toxina (Figura 6A). Los resultados mostraron que las dietas altas en fibra redujeron significativamente la abundancia normalizada de FV totales en las heces de cerdo, y no hubo diferencias significativas en la abundancia de FV entre las razas (Figuras 6A, C). Estudios de laboratorio previos han demostrado que las dietas altas en fibra resultaron en una reducción significativa en la abundancia normalizada del gen de la integrasa intI1 en las heces (19). Después de examinar la cantidad de AGCC en las heces, encontramos que la dieta alta en fibra elevó significativamente la cantidad de AGCC totales en las heces de cerdo, pero no hubo diferencias entre las razas (Figura 6B). Utilizamos el análisis de correlación para investigar más a fondo estas relaciones. Se encontraron correlaciones positivas significativas entre la abundancia normalizada de intI1 y las categorías de determinantes de adherencia a Fimbrial, evasión inmune, resistencia sérica y abundancia total de FV, mientras que la concentración fecal de AGCC totales se correlacionó significativamente con las concentraciones de evasión inmunitaria, efectores TTSS codificados por LEE, genes inducibles por macrófagos y las categorías de abundancia de FV totales mostraron correlaciones negativas significativas entre sí (Figura 6D). Por el contrario, las intervenciones dietéticas con alto contenido de fibra pueden modular eficazmente la excreción fecal de FV a través de los efectos intermedios de sus subproductos metabólicos, AGCC y MGE. Para confirmar la asociación entre los AGCC y los FV, evaluamos los efectos de BA, AA y PA sobre los FV transportados por microorganismos después de la fermentación de la flora fecal utilizando un modelo de fermentación in vitro. Los resultados mostraron que el AA tendió a reducir la abundancia normalizada de FV en el caldo de fermentación fecal porcino (p = 0,051) (Figura 6E). En conclusión, nuestros resultados sugieren que las dietas de fibra pueden influir en la excreción de FV en diferentes razas porcinas a través de la acción intermedia de su metabolito AA. Además, este efecto puede inhibir la propagación de FV al reducir los MGE en las heces.
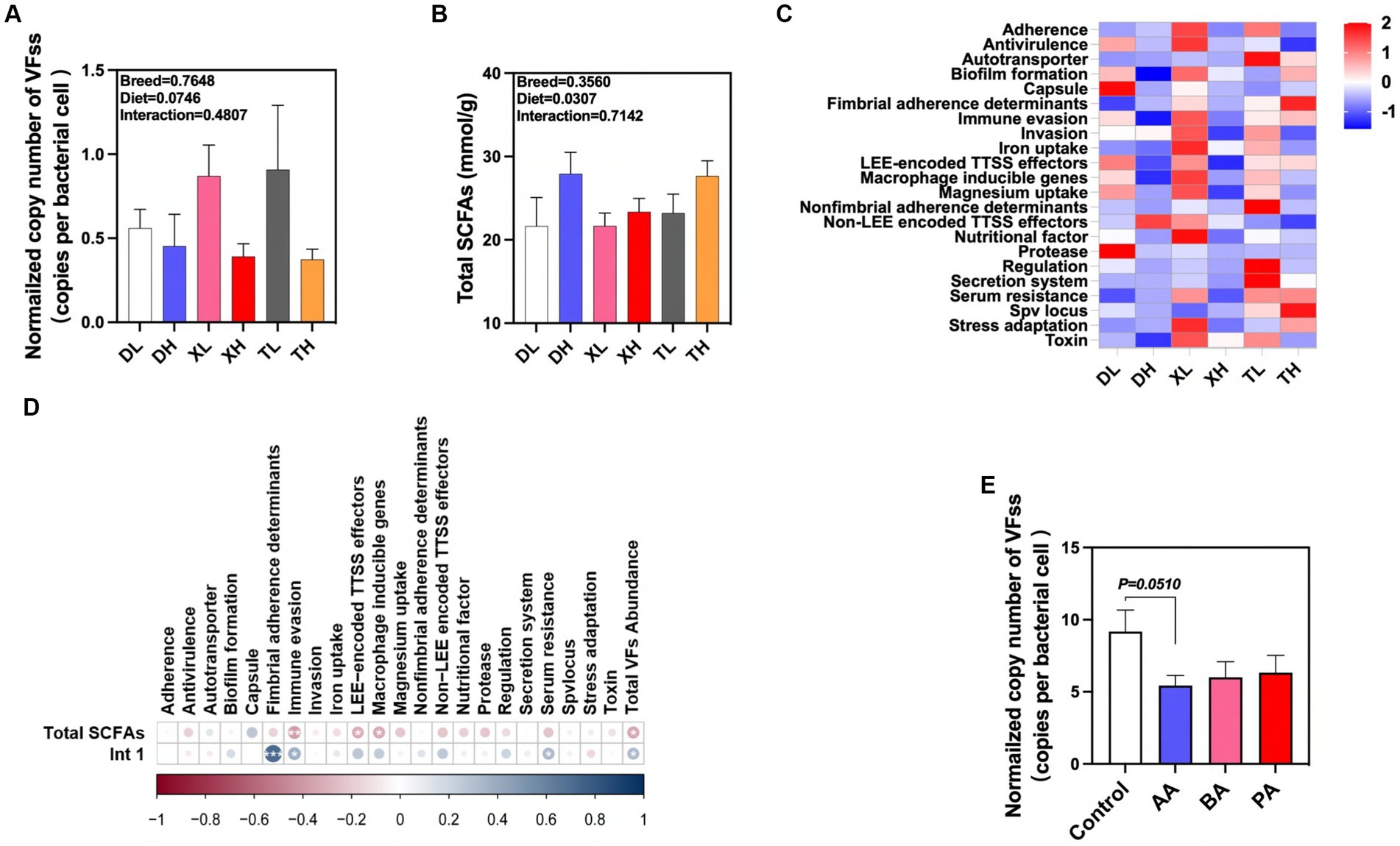 Figura 6. Efectos de diferentes niveles y especies de fibra en la excreción de FV en heces y sus impulsores. (A) Abundancia normalizada de FV en muestras fecales. AGCC totales en muestras fecales (B). (C) Mapa de calor del análisis de enriquecimiento de diferentes clases de FV en heces de diferentes grupos de cerdos. (D) Correlación de las FV con el gen intI1 y los AGCC totales en muestras fecales. (E) Efecto de AA, BA y PA sobre la abundancia estandarizada de FV en modo de fermentación in vitro. aA: ácido acético; PA: ácido propiónico; BA, ácido butírico; VA: ácido valérico; IPA: ácido isopropiónico; IBA: ácido isobutírico; IVA: ácido isovalérico. p < 0,05, p < 0,01 y p < 0,001. El cambio de color indica el valor R.******
Figura 6. Efectos de diferentes niveles y especies de fibra en la excreción de FV en heces y sus impulsores. (A) Abundancia normalizada de FV en muestras fecales. AGCC totales en muestras fecales (B). (C) Mapa de calor del análisis de enriquecimiento de diferentes clases de FV en heces de diferentes grupos de cerdos. (D) Correlación de las FV con el gen intI1 y los AGCC totales en muestras fecales. (E) Efecto de AA, BA y PA sobre la abundancia estandarizada de FV en modo de fermentación in vitro. aA: ácido acético; PA: ácido propiónico; BA, ácido butírico; VA: ácido valérico; IPA: ácido isopropiónico; IBA: ácido isobutírico; IVA: ácido isovalérico. p < 0,05, p < 0,01 y p < 0,001. El cambio de color indica el valor R.******
Discusión
Las FV albergadas por bacterias patógenas plantean importantes problemas de salud, lo que complica el tratamiento de la enfermedad. Las FV presentes en la flora intestinal y fecal son fuentes ambientales importantes (20). Comprender la distribución de estos FV ayuda a controlar los patógenos humanos potenciales, ya que su abundancia y propagación están influenciadas por presiones genéticas, ambientales y dietéticas (21).
En condiciones de alimentación intensiva, los cerdos Taoyuan exhiben una mayor abundancia de factores de virulencia (FV) en su contenido intestinal, y se ha observado que una dieta alta en fibra reduce la abundancia relativa de ciertos FV altamente abundantes. Este fenómeno puede atribuirse a la presencia de diversos microorganismos patógenos en el tracto intestinal de las razas porcinas locales, influenciados por sus características naturales y antecedentes genéticos. Además, la disminución del rendimiento inmunitario durante la cría a gran escala podría conducir a una mayor expresión de factores de virulencia (22). Algunos estudios han demostrado que los pandas gigantes salvajes tienen más bacterias patógenas portadoras de factores de virulencia en comparación con los pandas gigantes en cautiverio (23). Curiosamente, nuestros hallazgos revelan que la función predominante de los FV altamente expresados en los intestinos de los cerdos Taoyuan, en contraste con los cerdos Duroc, es facilitar la colonización de bacterias patógenas en el tracto intestinal del huésped, evadir la inmunidad del huésped y mejorar la absorción de hierro. Esto subraya la mayor resistencia ambiental y adaptabilidad de los microorganismos patógenos en el tracto intestinal de cerdos endémicos como los cerdos Taoyuan. Los estudios han demostrado que los cambios en el metabolismo afectan significativamente la distribución y abundancia de bacterias patógenas en los animales (24). La fibra dietética, conocida por desempeñar un papel crucial en el metabolismo energético, el metabolismo de los lípidos y el metabolismo de la glucosa en animales, se ha asociado con la alteración de los resultados de la enfermedad (25). Se ha demostrado que algunos extractos de plantas, como la populina y la quercetina, pueden mejorar los resultados de la enfermedad en pacientes con S. aureus al reducir la virulencia de S. aureus (26). Los resultados del presente estudio mostraron que un régimen dietético alto en fibra podría reducir significativamente la abundancia relativa de algunas FV. Y las principales funciones de las FV reducidas fueron la colonización de bacterias patógenas en el intestino del huésped, la evasión de la inmunidad del huésped y las FV relacionadas con la absorción de hierro.
La microbiota del colon es portadora de muchas FV. Debemos tener en cuenta que los microbios intestinales son los principales huéspedes de las FV, y una mayor diversidad de microbios intestinales implica múltiples mecanismos genéticos y más tipos de FV que son la fuente de muchos factores de virulencia desconocidos (27). Esto puede explicar los altos niveles de FV en el contenido colónico de los cerdos Taoyuan y Xiangcun. Se identificó un número limitado de especies microbianas que contribuían a muchas FV. s__Phascolarctobacterium_succinatutens se encontró que contribuía con una gran cantidad de FV. s__Phascolarctobacterium_succinatutens resultó ser el microorganismo cultivable a nivel de especie portador de la mayor cantidad de FV en el colon de cerdo. Varios estudios previos han reportado que s__ Phascolarctobacterium_succinatutens enriqueció en los intestinos de pacientes con cirrosis hepática y cáncer de colon, entre otros (28, 29). Esto sugiere que s__Phascolarctobacterium_succinatutens puede ser un agente causal potencial de ciertas enfermedades. Además, cabe destacar que varios microorganismos pertenecientes al género Lactobacillus también albergan cantidades significativas de FV. Algunos estudios incluso han confirmado la presencia de numerosos genes de resistencia en el genoma del género Lactobacillus (30). Esto subraya la importancia de considerar los posibles riesgos de seguridad asociados con el uso de Lactobacillus en los procesos de fermentación de alimentos.
Los microorganismos, los MGE y los AGCC son importantes impulsores de los cambios en las FV en el intestino. Las FV transportadas por las MGE experimentan constantes desplazamientos horizontales y cambios en la abundancia a través de la inducción humana y los cambios ambientales (31). Se observó una correlación lineal y positiva entre la abundancia relativa de MGE y FV. Como razón clave para su supervivencia y propagación, se ha determinado que los MGE se transmiten horizontalmente a una amplia gama de bacterias (tanto patógenas como humanas) (32). En el tracto intestinal de los cerdos se albergan numerosas FV, y los MGE sirven como vectores potenciales para la transferencia horizontal de genes. En consecuencia, es probable que estas FV se transmitan a los seres humanos a través de la cadena alimentaria o de rutas ambientales como ríos y suelos, lo que plantea riesgos potenciales para la salud. Por el contrario, nuestro estudio destaca el potencial de los ácidos grasos de cadena corta, metabolitos esenciales derivados de dietas ricas en fibra, como medio para mitigar el transporte de FV en el ganado y las aves de corral.
La fibra dietética reduce la abundancia de FV transportados por microorganismos en las heces a través de su metabolito AGCC. Las heces de los animales se producen debido a los procesos digestivos y metabólicos de los alimentos a través del tracto gastrointestinal y, por lo tanto, las alteraciones en los procesos metabólicos en el intestino pueden influir en las propiedades fecales (33), que pueden incluir la abundancia de FV. Por lo tanto, el efecto observado de una dieta rica en fibra sobre la abundancia de FV en las heces puede estar influenciado por el papel de los metabolitos derivados de la fibra en el intestino. De acuerdo con estudios previos, nuestros hallazgos mostraron que una dieta de fibra aumentó significativamente el contenido fecal de los AGCC totales (34). Además, se encontraron correlaciones negativas significativas entre los AGCC totales y la abundancia de clases funcionales de factores de virulencia, lo que sugiere que una dieta alta en fibra puede influir en la excreción de FV en las heces a través de sus metabolitos AGCC. Por lo tanto, demostramos además que el AA tendía a reducir la abundancia de FV mediante la validación de un modelo de fermentación in vitro. Estos resultados nos sugieren que el AA puede tener el efecto de reducir las FV al inhibir la proliferación de microorganismos huéspedes de la FV (por ejemplo, microorganismos patógenos en las heces). Además, a través de la aplicación exitosa en el modelado de fermentación, también reconocimos que el AA podría aprovechar su potencial para promover la seguridad ambiental al agregarse a los procesos de producción agrícola como el compostaje.
Conclusión
Esta investigación ofrece un análisis exhaustivo de la influencia de la genética y la fibra dietética en la distribución, composición e impulsores de los factores de virulencia (FV) en el contenido colónico y las heces de cerdo. Se observó que la raza autóctona china, el cerdo Taoyuan, exhibía una mayor abundancia relativa de FV que el cerdo comercial Duroc en la cría a gran escala. En particular, los perfiles de FV difirieron significativamente entre las dos razas. Los FV intracolónicos del híbrido «cerdo Xiangcun» fusionaron características de las cepas Taoyuan y Duroc. Si bien estos conocimientos son invaluables para medir los cambios en las FV durante la reproducción, una muestra más amplia y diversa es esencial para comprender a fondo la transmisión y las alteraciones de las FV. Las dietas altas en fibra redujeron marginalmente la abundancia de FV en el contenido colónico y, en menor medida, en las heces. Los principales contribuyentes a estos cambios fueron los microbios, los MGE (predominantemente plásmidos) y los AGCC (principalmente AA). Dado el uso frecuente de dietas de fibra y el metabolito AA en la producción, esto establece una base sólida para la mitigación del riesgo de FV en entornos de cría porcina. Los resultados de este estudio pueden utilizarse como referencia para optimizar las estrategias de alimentación y reducir el riesgo de que los alimentos sean portadores de FV.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio están a disposición del público. Estos datos se pueden encontrar en: https://www.ncbi.nlm.nih.gov, BioProject: PRJNA849732.
Declaración ética
El estudio en animales fue aprobado por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Agrícola de Sichuan, Sichuan, China (No. 20181105). El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
TW: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Validación, Visualización, Escritura – borrador original, Escritura – revisión y edición. YL: Conceptualización, Curación de datos, Metodología, Visualización, Escritura – borrador original, Escritura – revisión y edición. XK: Conceptualización, Curación de datos, Análisis formal, Metodología, Redacción – revisión y edición. BY: Conceptualización, Investigación, Metodología, Administración de Proyectos, Redacción – revisión y edición. PZ: Conceptualización, Investigación, Metodología, Administración de Proyectos, Redacción – revisión y edición. ZH: Conceptualización, Investigación, Metodología, Administración de Proyectos, Redacción – revisión y edición. XM: Conceptualización, Investigación, Metodología, Administración de Proyectos, Redacción – revisión y edición. JY: Conceptualización, Investigación, Metodología, Administración de Proyectos, Redacción – revisión y edición. JL: Conceptualización, Investigación, Metodología, Administración de Proyectos, Redacción – revisión y edición. HY: Conceptualización, Investigación, Metodología, Administración de Proyectos, Redacción – revisión y edición. JH: Conceptualización, Metodología, Recursos, Supervisión, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declaran/n que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio contó con el apoyo de la Fundación Nacional de Ciencias Naturales de China (n.º U20A2056) y el Programa Clave de Investigación y Desarrollo de la Provincia de Sichuan (2020YFN0147).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
El/los autor/es declararon, en el momento de la presentación, ser miembro del consejo editorial de Frontiers. Esto no tuvo ningún impacto en el proceso de revisión por pares ni en la decisión final.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1351962/full#supplementary-material
Notas
1. ^ https://cran.r-project.org/web/packages/networkD3
Referencias
1. Klous, G, Huss, A, Heederik, DJJ, y Coutinho, RA. Contactos humano-ganadero y su relación con la transmisión de patógenos zoonóticos, una revisión sistemática de la literatura. Una sola salud. (2016) 2:65–76. doi: 10.1016/j.onehlt.2016.03.001
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Liu, B, Zheng, D, Jin, Q, Chen, L y Yang, J. VFDB 2019: una plataforma patogénica comparativa con una interfaz web interactiva. Ácidos nucleicos Res. (2019) 47:D687–D692. doi: 10.1093/nar/gky1080
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Leitão, JH . Factores de virulencia microbiana. Int J Mol Sci. (2020) 21:5320. doi: 10.3390/ijms21155320
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Kadioglu, A, Weiser, JN, Paton, JC y Andrew, PW. El papel de los factores de virulencia de Streptococcus pneumoniae en la colonización respiratoria y la enfermedad del huésped. Nat Rev Microbiol. (2008) 6:288–301. doi: 10.1038/nrmicro1871
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Stella, NA, Brothers, KM, Callaghan, JD, Passerini, AM, Sigindere, C, Hill, PJ, et al. Una proteína de la familia IgaA/UmoB de Serratia marcescens regula la motilidad, la biosíntesis de polisacáridos capsulares y la producción de metabolitos secundarios. Appl Environ Microbiol. (2018) 84:e02575. doi: 10.1128/aem.02575-17
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Schwidder, M, Heinisch, L, y Schmidt, H. Genética, toxicidad y distribución de la hemolisina enterohemorrágica de Escherichia coli. Toxinas. (2019) 11:502. DOI: 10.3390/toxinas11090502
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Mitsou, EK, Kakali, A, Antonopoulou, S, Mountzouris, KC, Yannakoulia, M, Panagiotakos, DB, et al. La adherencia a la dieta mediterránea se asocia con el patrón de la microbiota intestinal y las características gastrointestinales en una población adulta. Hno. J Nutr. (2017) 117:1645–55. doi: 10.1017/s0007114517001593
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Richardson, AR . Virulencia y metabolismo. Microbiol Spectr. (2019) 7. DOI: 10.1128/microbiolspec. GPP3-0011-2018
9. Mayneris-Perxachs, J, Moreno-Navarrete, JM, y Fernández-Real, JM. El papel del hierro en la diafonía entre el huésped y la microbiota y sus efectos sobre el metabolismo sistémico de la glucosa. Nat Rev Endocrinol. (2022) 18:683–98. doi: 10.1038/s41574-022-00721-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Sánchez, KK, Chen, GY, Schieber, AMP, Redford, SE, Shokhirev, MN, Leblanc, M, et al. Las adaptaciones metabólicas cooperativas en el huésped pueden favorecer la infección asintomática y seleccionar la virulencia atenuada en un patógeno entérico. Célula. (2018) 175:146–158.e15. doi: 10.1016/j.cell.2018.07.016
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Xie, ST, Ding, LJ, Huang, FY, Zhao, Y, An, XL, Su, JQ, et al. VFG-Chip: una micromatriz de qPCR de alto rendimiento para perfilar genes de factor de virulencia del medio ambiente. Environ Int. (2023) 172:107761. doi: 10.1016/j.envint.2023.107761
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Rudin, L, Bornstein, MM y Shyp, V. Inhibición de la formación de biopelículas y factores de virulencia del patógeno oral cariogénico Streptococcus mutans por el flavonoide natural floretina. J Microbiol oral. (2023) 15:2230711. doi: 10.1080/20002297.2023.2230711
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Wyres, KL, Lam, MMC y Holt, KE. Genómica poblacional de Klebsiella pneumoniae. Nat Rev Microbiol. (2020) 18:344–59. doi: 10.1038/s41579-019-0315-1
14. Li, D, Liu, CM, Luo, R, Sadakane, K y Lam, TW. MEGAHIT: una solución ultrarrápida de un solo nodo para el ensamblaje de metagenómica grande y compleja a través de un sucinto gráfico de Bruijn. Bioinformática. (2015) 31:1674–6. doi: 10.1093/bioinformatics/btv033
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Fu, L, Niu, B, Zhu, Z, Wu, S y Li, W. CD-HIT: acelerado para agrupar los datos de secuenciación de próxima generación. Bioinformática. (2012) 28:3150–2. doi: 10.1093/bioinformatics/bts565
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Buchfink, B, Xie, C, y Huson, DH. Alineación rápida y sensible de proteínas con DIAMOND. Métodos Nat. (2015) 12:59–60. doi: 10.1038/nmeth.3176
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Franklin, MA, Mathew, AG, Vickers, JR y Clift, RA. Caracterización de poblaciones microbianas y concentraciones de ácidos grasos volátiles en yeyuno, íleon y ciego de cerdos destetados a los 17 vs. 24 días de edad. J Anim Sci. (2002) 80:2904–10. doi: 10.2527/2002.80112904x
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Pérez-Burillo, S, Molino, S, Navajas-Porras, B, Valverde-Moya, ÁJ, Hinojosa-Nogueira, D, López-Maldonado, A, et al. Un protocolo de fermentación discontinua in vitro para estudiar la contribución de los alimentos a la composición y funcionalidad de la microbiota intestinal. Nat Protoc. (2021) 16:3186–209. doi: 10.1038/s41596-021-00537-x
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Wang, T, Luo, Y, Kong, X, Yu, B, Zheng, P, Huang, Z, et al. Cambios en los genes de resistencia a los antibióticos en el contenido del colon y las heces de cerdo y sus factores impulsores mediados por la dieta genética y la fibra. Microorganismos. (2023) 11:2370. doi: 10.3390/microorganismos11102370
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Chuan, J, Belov, A, Cloutier, M, Li, X, Khan, IUH y Chen, W. Análisis genómico comparativo y factores relacionados con la virulencia en nuevas especies de Aliarcobacter faecis y Aliarcobacter lanthieri identificadas como posibles patógenos oportunistas. BMC Genómica. (2022) 23:471. DOI: 10.1186/s12864-022-08663-w
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Siscar-Lewin, S, Hube, B y Brunke, S. Aparición y evolución de la virulencia en hongos patógenos humanos. Tendencias Microbiol. (2022) 30:693–704. doi: 10.1016/j.tim.2021.12.013
22. Morgan, XC, Tickle, TL, Sokol, H, Gevers, D, Devaney, KL, Ward, DV, et al. Disfunción del microbioma intestinal en la enfermedad inflamatoria intestinal y su tratamiento. Genoma Biol. (2012) 13:R79. doi: 10.1186/gb-2012-13-9-r79
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Zhao, M, Li, Y, Wei, W, Zhang, Z y Zhou, H. La variación de la distribución de patógenos y factores de virulencia en diferentes poblaciones geográficas de pandas gigantes. Microbiol frontal. (2023) 14:1264786. doi: 10.3389/fmicb.2023.1264786
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Rosenberg, G, Riquelme, S, Prince, A y Avraham, R. Diafonía inmunometabólica durante la infección bacteriana. Nat Microbiol. (2022) 7:497–507. doi: 10.1038/s41564-022-01080-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Pathirannehelage, NPV, y Joye, IJ. La fibra dietética procedente de cereales integrales y sus beneficios para la salud metabólica. Nutrientes. (2020) 12:3045. doi: 10.3390/nu12103045
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Wu, SC, Liu, F, Zhu, K y Shen, JZ. Productos naturales que se dirigen a los factores de virulencia en el Staphylococcus aureus resistente a los antibióticos. J Agric Food Chem. (2019) 67:13195–211. doi: 10.1021/acs.jafc.9b05595
27. Escudeiro, P, Pothier, J, Dionisio, F y Nogueira, T. La diversidad génica de resistencia a antibióticos y la diversidad génica de virulencia están correlacionadas en los microbiomas intestinales y ambientales humanos. mSphere. (2019) 4:E00135. doi: 10.1128/mSphere.00135-19
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Bucher-Johannessen, C, Birkeland, EE, Vinberg, E, Bemanian, V, Hoff, G, Berstad, P, et al. El seguimiento a largo plazo de los asistentes a las pruebas de detección de cáncer colorrectal identifica diferencias en Phascolarctobacterium spp. utilizando el ARNr 16S y la secuenciación del metagenoma. Frente Oncol. (2023) 13:1183039. doi: 10.3389/fonc.2023.1183039
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Zhang, Y, Yan, S, Sheng, S, Qin, Q, Chen, J, Li, W, et al. Comparación de la microbiota intestinal en pacientes varones con MAFLD con rigidez hepática variable. Las células frontales infectan el microbiol. (2022) 12:873048. doi: 10.3389/fcimb.2022.873048
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Colautti, A, Arnoldi, M, Comi, G e Iacumin, L. Resistencia a los antibióticos y factores de virulencia en lactobacilos: algo a considerar cuidadosamente. Microbiol alimentario. (2022) 103:103934. doi: 10.1016/j.fm.2021.103934
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Lu, J, Zhang, X, Wang, C, Li, M, Chen, J y Xiong, J. Respuestas del resistoma de sedimentos, factores de virulencia y patógenos potenciales a décadas de contaminación por antibióticos en una granja acuícola de camarones. Sci Total Environ. (2021) 794:148760. doi: 10.1016/j.scitotenv.2021.148760
32. Weisberg, AJ, y Chang, JH. La flexibilidad de los elementos genéticos móviles como principio subyacente a la evolución bacteriana. Ann Rev Microbiol. (2023) 77:603–24. doi: 10.1146/annurev-micro-032521-022006
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. de Groot, P, Scheithauer, T, Bakker, GJ, Prodan, A, Levin, E, Khan, MT, et al. Las características metabólicas del donante influyen en los efectos del trasplante de microbiota fecal sobre la sensibilidad a la insulina del receptor, el gasto energético y el tiempo de tránsito intestinal. Tripa. (2020) 69:502–12. doi: 10.1136/gutjnl-2019-318320
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Trompette, A, Gollwitzer, ES, Yadava, K, Sichelstiel, AK, Sprenger, N, Ngom-Bru, C, et al. El metabolismo de la fibra dietética en la microbiota intestinal influye en las enfermedades alérgicas de las vías respiratorias y la hematopoyesis. Nat Med. (2014) 20:159–66. doi: 10.1038/nm.3444
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: factores de virulencia (FV), cerdo, dieta alta en fibra, colon, heces
Cita: Wang T, Luo Y, Kong X, Yu B, Zheng P, Huang Z, Mao X, Yu J, Luo J, Yan H y He J (2024) Cambios mediados por la dieta genética y de fibra en los factores de virulencia en el contenido del colon y las heces de cerdo y sus factores impulsores. Frente. Vet. Sci. 11:1351962. doi: 10.3389/fvets.2024.1351962
Editado por:
Huansheng Yang, Universidad Normal de Hunan, China
Revisado por:
Yuqing Feng, Universidad Agrícola de China, China
Adriana Belas, Universidad Lusofona, Portugal
Derechos de autor © 2024 Wang, Luo, Kong, Yu, Zheng, Huang, Mao, Yu, Luo, Yan y He. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Jun He, hejun8067@163.com
†Estos autores han contribuido igualmente a este trabajo
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía