
Circulación libre versus expresión de microARN asociado a vesículas extracelulares en el linfoma canino de células T

Circulación libre versus expresión de microARN asociado a vesículas extracelulares en el linfoma canino de células T
 Cecilia Capuano1†
Cecilia Capuano1† Antonella Molinari2
Antonella Molinari2 Livia Ferro1Serena Ferraresso2
Livia Ferro1Serena Ferraresso2


- 1Anicura Istituto Veterinario di Novara, Granozzo Monticello, Italia
- número arábigoDepartamento de Biomedicina Comparada y Ciencia de los Alimentos, Universidad de Padua, Legnaro, Italia
- 3Departamento de Medicina, Producción y Sanidad Animal, Universidad de Padua, Legnaro, Italia
Introducción: El linfoma canino (lL) es uno de los cánceres más frecuentes en perros. El linfoma de células T (TcL) no es el fenotipo más común pero presenta un comportamiento agresivo. Los microARN (miARN) son pequeñas moléculas de ARN monocatenario no codificantes que pueden circular libremente en la sangre o estar asociadas a vesículas extracelulares (VE). La desregulación de ciertos miRNAs ha sido identificada en numerosos tipos de cánceres humanos y han sido ampliamente investigados como posibles biomarcadores tumorales en medicina humana, mientras que la investigación en oncología veterinaria es todavía escasa. El objetivo de este estudio fue comparar los patrones de expresión de miRNAs circulantes libres y asociados a EV en perros con linfoma de células T (TcL) y perros sanos.
Métodos: Se seleccionaron ocho perros con TcL como grupo de linfoma (LG) y ocho perros como controles (Ctrl). Se recogieron muestras de plasma en el momento del diagnóstico y se aislaron las VE con ultracentrifugación. Los miRNAs se extrajeron tanto de los EVs circulantes como del sobrenadante plasmático, obteniendo miRNAs asociados a EV y libres. Se realizó una PCR cuantitativa en tiempo real para analizar la expresión de 88 miRNAs diana.
Resultados: Se detectaron diez y siete miRNAs expresados diferencialmente entre LG y Ctrl en miRNAs asociados a EV y libres, respectivamente. Entre los miRNAs asociados a EV y libres, solo has-miR-222-3p se sobreexpresó en ambas condiciones.
Discusión: Todos los miRNAs expresados diferencialmente detectados en este estudio, ya han sido descritos como desregulados en otros cánceres humanos o caninos. Los miARN asociados a EV, que parecen ser más estables y mejor conservados que los miARN libres, podrían investigarse en estudios más amplios para evaluar mejor su uso como posibles biomarcadores de TcL.
1 Introducción
El linfoma canino (lL) es uno de los cánceres más frecuentemente diagnosticados en perros y representa la neoplasia más manejada en oncología veterinaria (1). La mayoría de los casos son linfoma de células B (70%), mientras que los otros pueden ser linfoma de células T (TcL) o linfoma no B/no de células T (1, 2). En general, la lL se diagnostica de forma rutinaria mediante el examen citológico de los tejidos afectados, pero se pueden realizar otras investigaciones diagnósticas para el inmunofenotipado, la clasificación y la estadificación clínica, principalmente para predecir el comportamiento biológico (2). La quimioterapia multifarmacológica representa un tratamiento de referencia para la cL y a menudo se logra la remisión, a pesar de que la TcL puede desarrollar más comúnmente resistencia a los medicamentos (2).
Los microARN (miARN) son pequeñas moléculas de ARN no codificantes de aproximadamente 22 nucleótidos de longitud que participan en la regulación génica postranscripcional. Al tener este papel, están implicados en la regulación de diferentes procesos biológicos, entre ellos la carcinogénesis, ya sea oncogenes o supresores de tumores (3). Muchos estudios han demostrado que la expresión de muchos miRNAs oncogénicos está desregulada en los tumores, lo que mejora la señalización proliferativa, la evasión de los supresores del crecimiento, la resistencia a la muerte celular, la invasividad y la angiogénesis (3). Muchos mecanismos están implicados en esta desregulación e incluyen la amplificación o deleción de genes de miRNA, un control transcripcional anormal de los miRNAs, cambios epigenéticos desregulados y defectos en la maquinaria de biogénesis del miRNA (3).
Dado que los miRNAs existen no solo en las células y tejidos, sino también en diversos fluidos corporales, junto con su importante estabilidad, ofrecen un vasto recurso de biomarcadores mínimamente invasivos (2). Además, los miRNAs pueden circular libremente en la sangre o asociados a vesículas extracelulares (VE).
Las vesículas extracelulares (VE) son vesículas heterogéneas unidas a la membrana liberadas por las células (4). Los vehículos eléctricos, que transportan moléculas como proteínas, azúcares, lípidos y ácidos nucleicos, desempeñan un papel crucial en la comunicación de célula a célula (5).
Los VE se dividen principalmente en exosomas y microvesículas, según su biogénesis (5, 6). Sin embargo, otras partículas derivadas de células pueden clasificarse como VE, incluidos, por ejemplo, los cuerpos apoptóticos, los prostasomas y los oncosomas. Teniendo en cuenta la heterogeneidad de los VE y las dificultades para reconocer su origen, generalmente también se clasifican según su tamaño en vehículos eléctricos pequeños (<200 nm) o vehículos eléctricos grandes (>200 nm) (5, 6).
Entre muchas otras funciones, las VE desempeñan un papel en la promoción del proceso cancerígeno, induciendo la angiogénesis, la diseminación tumoral, el escape inmunológico, la metástasis y la resistencia a los medicamentos (5, 7, 8). Las VE están presentes en todos los fluidos corporales (por ejemplo, sangre, saliva, orina, líquido broncoalveolar, leche materna, semen) y son estables durante mucho tiempo y son fáciles de muestrear, siendo adecuadas para la recolección secuencial (8). En muchos estudios de medicina humana, se ha informado de que los ARN asociados a EV son posibles biomarcadores tumorales para el diagnóstico del cáncer o para monitorizar la progresión del cáncer (8). Sin embargo, a pesar de que se han realizado algunos estudios en medicina veterinaria, el conocimiento sigue siendo limitado con respecto a la expresión de miRNAs de circulación libre y asociados a EV en perros con linfoma (2, 9, 10).
Teniendo en cuenta la necesidad de biomarcadores no invasivos en oncología veterinaria y la escasez de información sobre la expresión de miRNAs en el linfoma canino de células T (TcL), el objetivo de este estudio fue identificar patrones de expresión de miRNAs circulantes específicos en perros con TcL, investigando tanto los miRNAs de circulación libre como los miRNAs asociados a EV como posibles biomarcadores diagnósticos.
2 Materiales y métodos
2.1 Reclutamiento de casos
Se trata de un estudio prospectivo de doble brazo. Se incluyeron ocho perros como controles (Ctrl) y ocho perros diagnosticados con TcL se seleccionaron de acuerdo con los criterios de elegibilidad para el grupo de linfoma (LG). Todas las muestras de LG y algunas de Ctrl fueron proporcionadas por Anicura Istituto Veterinario Novara (Novara, NO, Italia). El resto de las muestras de Ctrl fueron proporcionadas por el Hospital Universitario de Veterinaria de la Universidad de Padua (Legnaro, PD, Italia). Todos los pacientes con linfoma fueron tratados en el AniCura Istituto Veterinario Novara con protocolos de quimioterapia que requirieron controles hematológicos sistemáticos. Las muestras de sangre se obtuvieron en el momento del diagnóstico (T0), durante los procedimientos de rutina de diagnóstico clínico y los restos se utilizaron para el estudio. Las muestras de sangre de Ctrl se obtuvieron durante la cirugía electiva para la esterilización y las sobras se guardaron para el estudio. Por lo tanto, no se requirió la aprobación de un comité de ética para este estudio.
Los criterios de inclusión para el Ctrl fueron: ausencia de alteraciones clínicas, hematológicas o bioquímicas. Los criterios de inclusión para la LG incluyeron un diagnóstico citológico o histológico de linfoma, ausencia de tratamientos previos con quimioterapia o esteroides y ausencia de otras comorbilidades. La presencia o sospecha de linfoma leucémico se consideró criterio de exclusión. Se obtuvo el consentimiento informado de todos los propietarios.
El diagnóstico, la clasificación y la estadificación del linfoma se realizaron de acuerdo con la clasificación actualizada de Kiel (Lennert K, Feller CA: Histologie des Lymphomes Malins Non Hodgkiniens Selon la Classification de Kiel Actualise’. Doin, París, Francia, 1991). Para cada caso, el examen clínico, el hemograma completo (CBC) y el perfil bioquímico, las radiografías de tórax, la ecografía abdominal (US) y la citología de los ganglios linfáticos agrandados en la presentación clínica, el hígado (con una excepción) y el bazo se realizaron de acuerdo con los procedimientos diagnósticos estándar. Para confirmar el origen de los linfomas en células T, el inmunofenotipado se evaluó mediante citometría de flujo y la graduación se determinó por el tamaño de las células (mayoría de células de tamaño pequeño o mayoría de células de tamaño mediano y grande) y el índice mitótico (11). Los fenotipos de cL se determinaron mediante citometría de flujo (CyFlow Space, Sysmex Europe GmBH, Norderstedt, Alemania) y los datos se analizaron con el software FlowMax (Sysmex Europe GmBH, Norderstedt, Alemania). Para la citometría de flujo, se recogieron muestras de ganglios linfáticos en Eppendorff que contenían 500 μL de suero autólogo y se analizaron dentro de las 24 horas posteriores a la recolección. La lisis eritrocitaria se realizó utilizando tampón de lisis eritrocitaria que contenía cloruro de amonio al 8% si se consideró necesario debido a la hemodilución. Los porcentajes de células vivas y muertas se evaluaron utilizando yoduro de propidio (PI) para evaluar la conservación de las muestras antes de la tinción con anticuerpos. Se utilizó un panel de anticuerpos que incluía CD45 y CD44 (panleucocitos), CD3 y CD5 (linfocitos T), CD4 (linfocitos T colaboradores), CD8 (linfocitos T citotóxicos), CD21 (linfocitos B), CD34 (células blásticas). Para cada muestra, se añadieron 50 μL de suspensión celular a los tubos que contenían diferentes combinaciones de anticuerpos (Tabla suplementaria S1).
2.2 Aislamiento y caracterización de vesículas extracelulares a partir de extracción con plasma y microARN
Cada perro recogió un total de 4 mL de sangre venosa y los transfirió a un tubo de EDTA. Tras la recolección, cada muestra se centrifuga a 4 °C a 2000 × g durante 15 min para obtener la fracción plasmática. El plasma se almacenó a -80 °C hasta el aislamiento de los vehículos eléctricos.
Los VE se aislaron del plasma de perros Ctrl y LG con ultracentrifugación (UC). Cada muestra se centrifugó primero a 4 °C (2000 × g durante 10 min) para eliminar los residuos grandes. A continuación, el sobrenadante se transfirió a un nuevo tubo y se centrifugó dos veces a 3850 × g. El sobrenadante final (aproximadamente 1 mL) se diluyó en solución salina tamponada con fosfato (PBS) de 0,2 μm con doble filtro (dfPBS) para alcanzar un volumen final de 4 mL, se transfirió a un tubo de ultracentrífuga (Ultra-Clear Open top, Beckman Coulter, Brea, CA, EE. UU.) y se ultracentrifugó en un rotor de cangilón oscilante (SW55ti, Beckman Coulter, Brea, CA, EE. UU.) a 100.000 × g durante 90 min a 4 °C (Beckam Coulter Optima L-90K, Beckman Coulter, Brea, CA, EE.UU.).
Después de la CU, los gránulos obtenidos que contenían EV (EV-pellet) se lisaron y los ácidos nucleicos se extrajeron utilizando miRNAeasy® Micro Kit (Qiagen, Hilden, Alemania) de acuerdo con las instrucciones del fabricante para obtener finalmente miRNAs asociados a EV. Para extraer los miRNAs de circulación libre (free-miRNAs), se utilizó miRNaeasy Serum/Plasma Advanced Kit (Qiagen, Hilden, Alemania) de acuerdo con las instrucciones del fabricante sobre el sobrenadante UC.
Debido al bajo volumen de muestras de plasma, se utilizaron dos muestras más de Ctrl, no incluidas en el siguiente análisis de miARN, como controles técnicos para evaluar la presencia de EV después de la CU. Después de realizar el aislamiento de EV como se describió anteriormente, una de estas dos muestras se utilizó para realizar el análisis de seguimiento de nanopartículas (NTA) y otra para el Western Blotting (WB).
2.3 Análisis de seguimiento de nanopartículas
Se realizó NTA para evaluar la distribución de tamaños y la concentración de VE. Después de la CU, el pellet de EV se resuspendió en 600 μL de dfPBS y luego se diluyó 1:100 en el mismo tampón. Se realizó NTA con NanoSight NS300 (Malvern Panalytical, Malvern, Reino Unido) y se grabaron y analizaron 3 videos de 60 s cada uno utilizando el software 3.4, con el nivel de la cámara establecido en 14 y el umbral de detección en 5. Los valores considerados fiables fueron los comprendidos en los siguientes rangos: partículas por fotograma de 20 a 120; concentración de partículas: 106–109/mL; La relación entre las partículas totales y las partículas válidas ≥ 1/5.
2.4 Western blot
Se realizó WB para verificar la presencia de marcadores EV en el pellet enriquecido con EV.
Después de la CU, las proteínas de los gránulos enriquecidos con EV se extrajeron utilizando 20 μL de tampón RIPA (Thermo Fisher Scientific, Waltham, MA, EE. UU.) suplementado con inhibidor de la proteinasa. En primer lugar, se cuantificó la concentración de proteínas mediante el tampón Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, EE. UU.), de acuerdo con las instrucciones del fabricante. Se desnaturalizaron 20 μg de proteínas durante 10 min a 70 °C, se resolvieron mediante gel de electroforesis utilizando el gel NuPAGE 4-12% Bis Tris (Thermo Fisher Scientific, Waltham, MA, USA) y se transfirieron a una membrana de nitrocelulosa. Los sitios de unión inespecíficos se bloquearon mediante una incubación de 90 min a temperatura ambiente con una solución de TBS-T (solución salina tamponada con Tris con 0,05% de Tween-20) suplementada con 5% de leche desnatada en polvo.
Las transferencias se incubaron durante la noche en TBS-T suplementado con un 1% de leche desnatada con anticuerpos primarios antihumanos contra TSG101 (proteína citosólica, dilución 1:1000, GTX70255, GeneTex, Irvine, CA, EE. UU.) e integrina-beta (una proteína de membrana, dilución 1:5000, GTX128839, GeneTex, Irvine, CA, EE. UU.) a 4 °C, un marcador EV citosólico y otro de membrana, respectivamente (12). Después de la incubación durante la noche, las membranas se incubaron a temperatura ambiente durante 1 h con un anticuerpo secundario conjugado con peroxidasa (dilución 1:3000, anti-Rabbit #32260 o anti-Mouse #32230, Thermo Fisher Scientific, Waltham, MA, EE. UU.).
Por último, las bandas resultantes de la unión antígeno-anticuerpo se visualizaron utilizando un kit de detección quimioluminiscente (SuperSignal West Pico PLUS Chemluminescent Substrate, Thermo Fisher Scientific, Waltham, MA, EE. UU.) con el instrumento iBright (Thermo Fisher Scientific, Waltham, MA, EE. UU.).
2.5 Transcripción inversa y PCR cuantitativa en tiempo real
El ARN extraído de cada muestra (8 sujetos LG y 8 sujetos ctrl) se transcribió inversamente a ADN complementario (ADNc), utilizando el kit miRCURY LNA RT (Qiagen, Hilden, Alemania) de acuerdo con las instrucciones del fabricante. A cada ARN, se añadieron 0,5 μL de ARN UniSp6 con espiga como control para monitorizar el éxito de la transcripción inversa y para ser utilizado como calibrador entre placas en la qPCR. La solución obtenida se incubó a 42°C durante 60 min y luego a 95°C durante 5 min.
Por último, se realizó inmediatamente la qPCR o se almacenaron muestras de ADNc a -20 °C durante un máximo de 5 semanas.
Se realizó una RT-PCR cuantitativa para cada muestra utilizando un panel de PCR miRNA Focus de miRNA LNA de 96 pocillos en placa de 96 pocillos (Qiagen, YAFD-201Z, Qiagen, Hilden, Alemania), diseñado específicamente por la empresa fabricante para el diagnóstico de linfoma canino, y miRCURY LNA SYBR® Green PCR Kit (Qiagen, Hilden, Alemania), de acuerdo con las instrucciones del fabricante. Estas placas están diseñadas para dirigirse a 84 miRNAs diferentes y 4 genes de referencia (U6 snRNA, 5S rRNA, RNU5G, RNU1A1) mientras que los pocillos restantes son para controles positivos y calibradores interplaca (es decir, IPC UniSp3) (Tabla suplementaria S2).
La mezcla de reacción se preparó añadiendo 495 μL de agua libre de nucleasas, 510 μL de 2x miRCURY SYBR® Green PCR Master Mix, 5,1 μL de ROX y 10 μL de ADNc. Para cada diana/ensayo, la reacción de qPCR se llevó a cabo en un volumen final de 10 μL. El protocolo de amplificación consistió en una etapa inicial de 2 min a 95°C, seguida de 40 ciclos de 10 s a 95°C y 60 s a 56°C. Todos los experimentos se llevaron a cabo en un Thermo ABI 7500 (Applied Biosystems, Waltham, MA, USA).
2.6 Análisis estadístico
Los valores umbral de ciclo (Ct) obtenidos se analizaron utilizando el análisis de datos de matriz de PCR de miRNA de GeneGlobe. 1 Para verificar la eficiencia de la transcripción inversa y las reacciones de qPCR, se utilizaron la espiga inscel-miR-39-3p y Unisp6 como controles de amplificación interna. Se aplicó un primer control basado en el valor de Ct tanto en los miRNAs diana como en los de referencia antes de la normalización. A los valores de Ct por encima de 36 se les asignó el estado indeterminado, ya que la transcripción se considera «indetectable». Los miRNAs con Ct entre 33 y 36 se consideran «detectables pero no cuantificables», y los miRNAs con Ct por debajo de 33 se consideran «cuantificables» y, por lo tanto, utilizables para el análisis estadístico posterior. Se realizó una primera normalización entre placas basada en el IPC UniSp3. El software calculó el factor calibrador (CF), obtenido a partir de la diferencia entre el Ct de la IPC de cada muestra (placa IPC) y la media de los valores de la placa IPC de todas las muestras (IPC en general). Para cada muestra, se corrigió el valor de Ct de todos los genes (CtC) en función del valor de CF. A continuación, los valores de CtC de cada miARN se normalizaron según el método NormFinder. La cuantificación relativa se determinó mediante el método Delta\Delta Ct. El cambio de pliegue (FC) se calculó utilizando 2^-(Delta\DeltaCT) para evaluar las diferencias en la expresión de miRNA entre el grupo de control y el grupo de linfoma: FC mayores que 1 fueron indicativos de sobreexpresión, mientras que FC menores de 1 representaron bajo expresión.
Para comparar la diferencia entre la expresión media de cada miRNA entre los dos grupos, se realizó la prueba t de Student y se consideraron estadísticamente significativos los valores de FC mayores de 1,5 y p (p) menores de 0,05.
3 Resultados
3.1 Características del paciente
Dieciséis perros se inscribieron en el estudio, ocho perros en el LG y 8 perros en el Ctrl. El LG incluyó cuatro hembras y cuatro machos con una edad media de 7,75 años (4-12) y pertenecientes a varias razas (mestizo, bóxer, labrador retriever, Weimaraner, boyero de Berna). La forma más común de linfoma fue el multicéntrico (cinco perros), con un paciente con afectación nasal también. Las otras presentaciones de linfoma fueron mediastínico (un perro), hepatoesplénico (un perro), cardíaco (un perro) y en el ganglio linfático yeyunal (un perro) (Tabla 1).
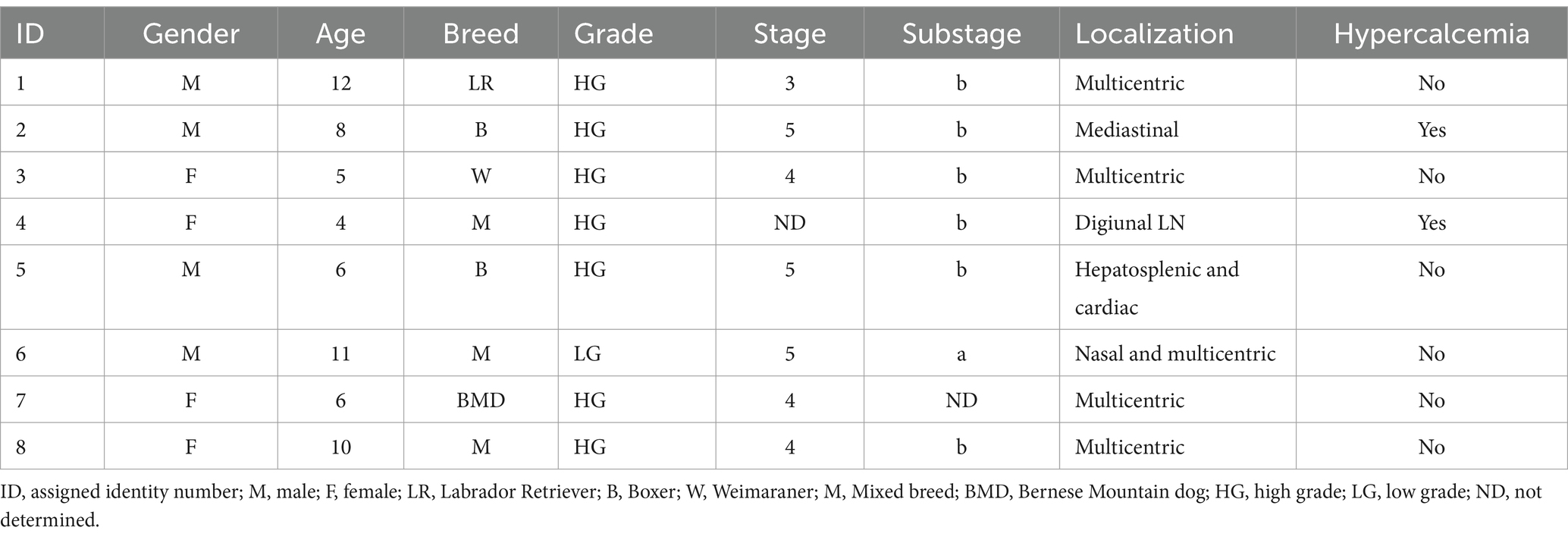
Tabla 1. Características de los perros del grupo de los linfomas.
La evaluación citológica y citométrica de flujo confirmó que siete perros (87,5%) presentaban linfoma de alto grado y un perro con linfoma de bajo grado (ID 6). Un perro tenía linfoma en etapa 3, tres perros tenían etapa 4 y tres perros presentaban un linfoma en etapa 5. Seis perros (75%) fueron clasificados con el subestadio b (Tabla 1).
3.2 Caracterización de vesículas extracelulares
Para confirmar el tamaño y la concentración de EV, se realizó NTA en un pellet enriquecido con EV después de la CU (Figura 1A).
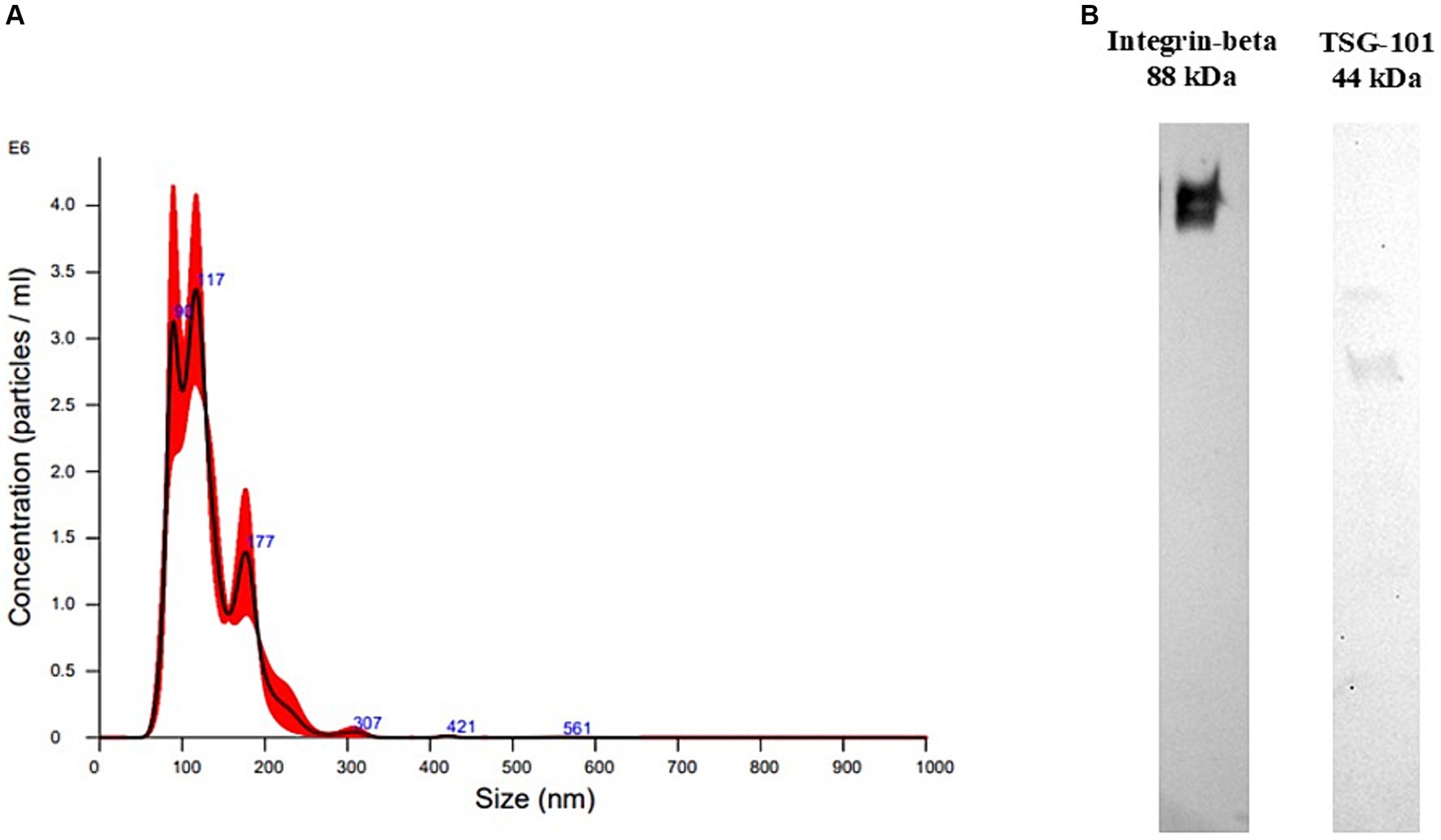
La distribución del tamaño de partícula mostró rangos dentro de los de los VE, principalmente entre 80 y 600 nm, con valores de media y moda de 130,4 +/− 3,1 y 111,3 +/− 12,3 nm, respectivamente, lo que significa que se aislaron principalmente los VE pequeños. La concentración de partículas fue de 2,53*1012 +/− 2,54*1011 partículas/mL.
Se realizó WB en una muestra para confirmar la presencia de marcadores EV en el pellet enriquecido con EV. El gránulo enriquecido con EV dio positivo para dos marcadores típicos de EV, TSG101, una proteína citosólica, e integrina-beta, una proteína de membrana (Figura 1B).
3.3 Cuantificación de miRNAs asociados a vesículas extracelulares y de circulación libre
Para cada perro incluido en el estudio, se llevaron a cabo análisis de miRNA tanto en muestras asociadas a EV (EVs-LG y EVs-Ctrl) como en muestras de miRNA libre (Free-miRNA-LG y Free-miRNA-Ctrl).
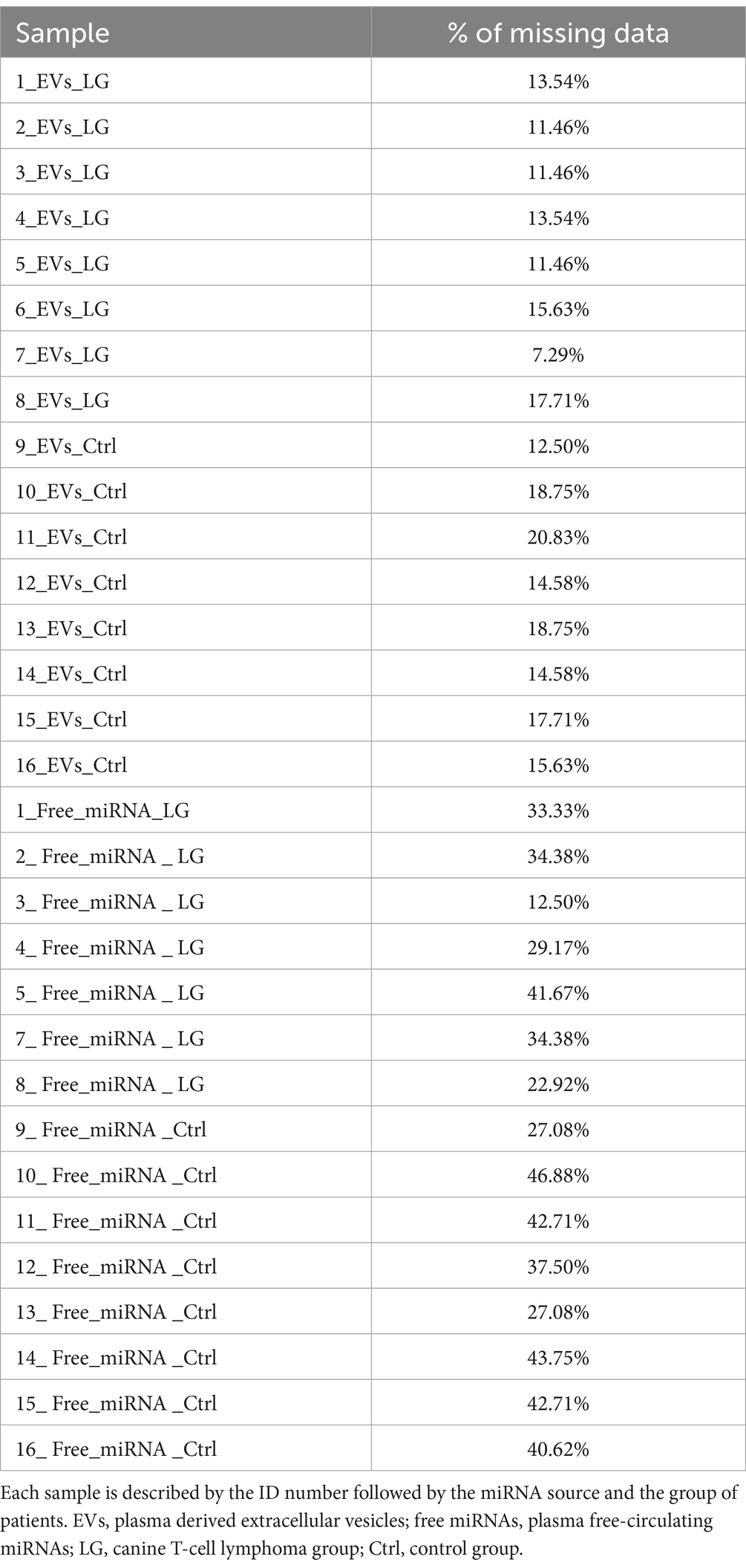
La muestra de miRNA libre ID 6 (6-Free-miRNA-LG, Tabla 1) tenía valores de CtC para todos los miRNAs diana mucho más altos que todas las demás muestras, con el consiguiente bajo porcentaje de miRNAs cuantificables (<30%). Teniendo en cuenta este porcentaje insuficiente de miRNAs cuantificables, esta muestra no se incluyó en los análisis estadísticos del estudio. Para todas las demás muestras, el análisis de los perfiles de amplificación identificó un porcentaje variable de miRNAs indetectables, generalmente mayor para los miRNAs libres (datos faltantes que oscilaban entre el 7,29 y el 20,83% para los miRNAs asociados a EV y entre el 12,50 y el 46,88% para los miRNAs libres) (Tabla 2). Entre los miRNAs asociados a EV y los miRNAs libres, el software seleccionó 5 miRNAs con la mejor puntuación de estabilidad como miRNAs de referencia para cada una de las dos fuentes de miRNAs (Tabla Suplementaria S3). El coeficiente de variación CV%, de los 5 miRNAs de referencia se calculó como el cociente entre la desviación estándar de cada miRNA y su CtC medio entre todas las muestras, multiplicado por 100. Se considera que un valor CV óptimo tiene un valor inferior al 15%. En este estudio, el CV% calculado para los miRNAs de referencia mostró un valor entre 3 y 7% (Tabla Suplementaria S3).
3.4 Expresión diferencial de miRNA entre los dos grupos
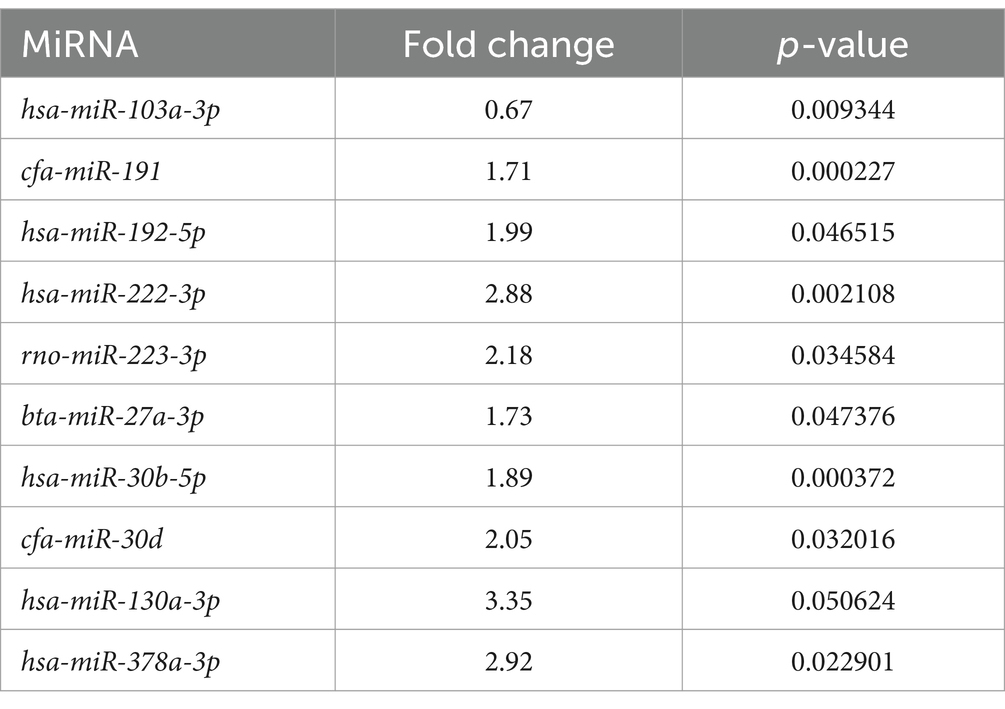
Se encontró que un total de 10 miRNAs asociados a EV se expresaban diferencialmente en LG en comparación con Ctrl. Un miRNA adicional (miR-103a-3p) tuvo un valor de p cercano a la significación (valor de p = 0,0506) (Tabla 3 y Figura Suplementaria S1).
Se encontró que un total de siete miRNAs libres se expresan diferencialmente en LG en comparación con Ctrl (Tabla 4 y Figura Suplementaria S1).
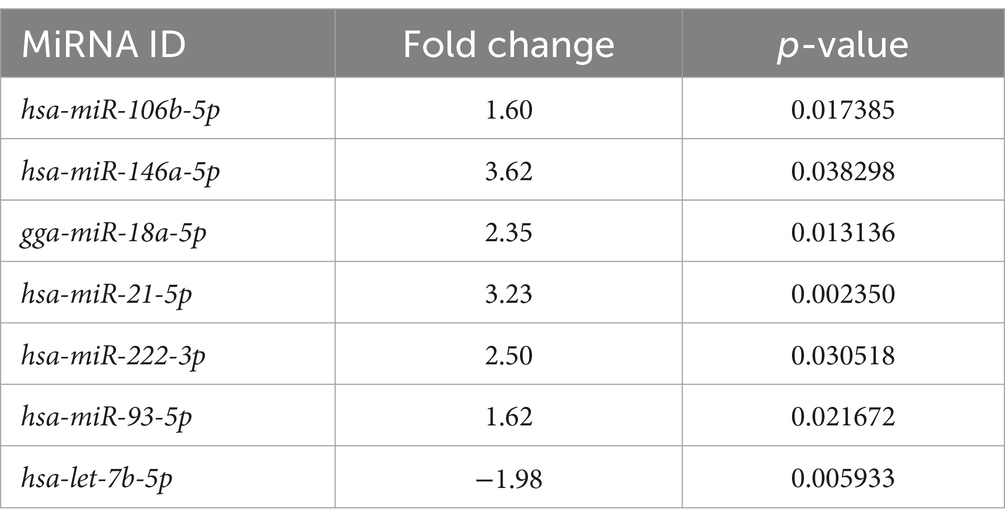
En LG, solo se encontró un miRNA (miR-222-3p) sobreexpresado tanto en los miRNAs asociados a EV como en los miRNAs libres (Tablas 3, 4). Como se muestra en el gráfico del volcán, entre los miRNAs asociados a EV expresados diferencialmente en LG, 8 estaban sobreexpresados estadísticamente significativos, uno estaba significativamente subexpresado y uno estaba sobreexpresado pero marginalmente significativo (Figura 2A).
Figura 2. Diagrama volcánico de genes expresados diferencialmente entre el linfoma y el grupo control. (A) MiRNAs asociados a vesículas extracelulares. (B) MiRNAs de libre circulación. Grupo 1 = grupo de linfoma.
Entre los miRNAs libres expresados diferencialmente, 6 miRNAs se sobreexpresaron de manera estadísticamente significativa, y uno se subexpresó significativamente en el LG (Figura 2B).
Finalmente, un análisis de componentes principales (PCA) sobre genes expresados diferencialmente en muestras de miRNA asociadas a EV, mostró una separación entre LG y Ctrl (Figura 3A). La PCA realizada en genes expresados diferencialmente de las muestras de miARN libre no mostró en cambio ninguna separación entre LG y Ctrl (Figura 3B).
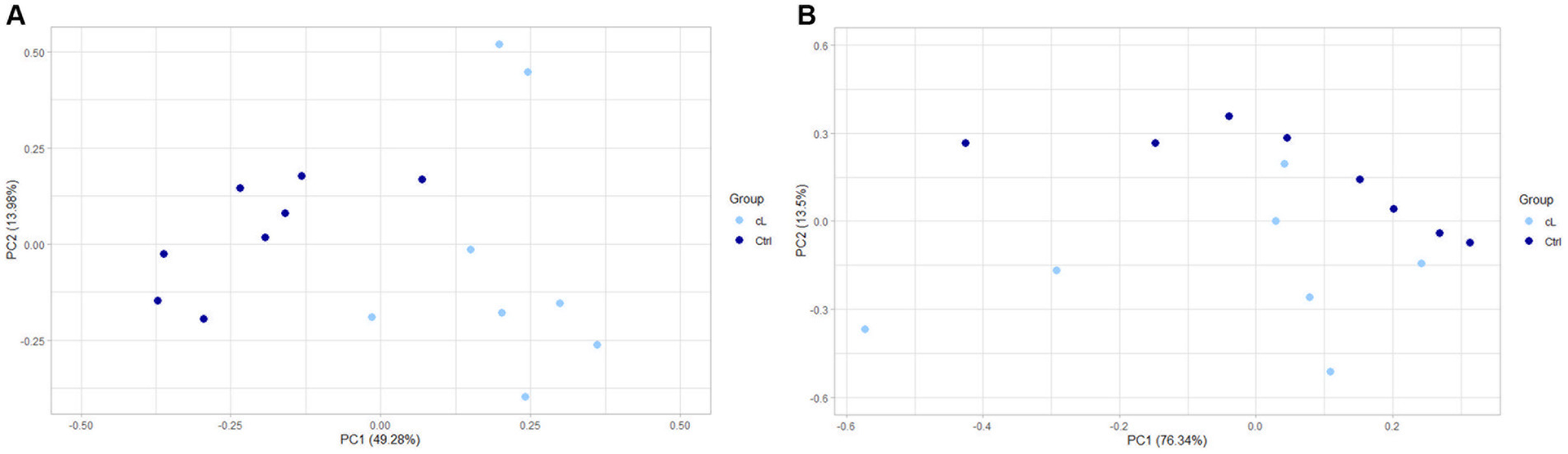
Figura 3. PCA de genes expresados diferencialmente entre el linfoma y el grupo control. (A) MiRNAs asociados a vesículas extracelulares. (B) MiRNAs de libre circulación. cL: grupo de linfomas; Ctrl, grupo de control.
4 Discusión
La TcL canina sigue planteando importantes desafíos para los oncólogos veterinarios, con un comportamiento particularmente rápido y agresivo y sin enfoques curativos desarrollados en los últimos 25 años a pesar de numerosos intentos (1, 13). Al igual que en otros tumores de mascotas, la detección temprana sigue siendo difícil de alcanzar, y el enfoque en la investigación de biomarcadores se considera un faro de esperanza para identificar linfomas en una etapa temprana, o para proporcionar valor pronóstico. En este contexto, los estudios in vivo son cruciales, ya que la investigación in vitro con células cancerosas no logra identificar las primeras etapas de la enfermedad y no replica con precisión la complejidad de las condiciones in vivo (14, 15).
Los miRNAs han ganado popularidad en la medicina veterinaria y humana como biomarcadores tumorales mínimamente invasivos, dada su presencia estable no solo en células y tejidos, sino también en diversos fluidos corporales (2, 3). Este estudio preliminar tuvo como objetivo identificar los patrones de expresión de los miRNAs circulantes en un pequeño grupo de perros caninos con linfoma de células T, y compararlo con perros sanos, investigando la expresión de miRNAs de circulación libre y miRNAs asociados a EV.
La caracterización de EV realizada en dos controles técnicos, mostró que el aislamiento de EV con CU permitió la purificación de partículas que fueron similares en términos de concentración y distribución de tamaño a las de otros estudios en los que se aislaron principalmente VE pequeños utilizando CU de plasma canino (16, 17). Además, las partículas aisladas fueron positivas para dos marcadores EV típicos en WB, un marcador citosólico (TSG101) y un marcador de membrana (integrina-beta) (6).
Después de la caracterización de EV, realizamos qPCR y análisis de datos tanto en miRNAs asociados a EV como en miRNAs de circulación libre. Como consideraciones generales, se detectaron más valores de Ct indeterminados en las muestras de miARN de circulación libre en comparación con las asociadas a EV, lo que significa que se detectaron menos miARN en general en las muestras de miARN libres. Además, hubo un número marginalmente mayor de miRNAs expresados diferencialmente en los asociados a EV en comparación con los miRNAs libres al comparar LG y Ctr (9 vs. 7). Entre los miRNAs expresados diferencialmente, solo uno era común entre las muestras asociadas a EV y las muestras de miRNA libres (miR-222-3p). Esta falta de correspondencia entre los miRNAs expresados diferencialmente que estaban asociados a EV o libres en el plasma podría deberse al hecho de que los miRNAs asociados a EV constituyen solo una pequeña porción del total de miRNAs plasmáticos y su expresión puede diferir de los que circulan libremente (18, 19). Además, los miRNAs asociados a EV son más estables y están mejor protegidos de la actividad de la RNAsa, lo que podría explicar la mayor cantidad de valores determinados de Ct observados en los miRNAs asociados a EV en nuestro estudio (20, 21). De acuerdo con nuestros resultados, una revisión reciente subrayó que, entre los numerosos estudios que examinaron la expresión de miARN en pacientes con cáncer de próstata humano, ya sea como miARN circulantes totales (incluidos los miARN asociados a EV y libres) o miARN asociados a EV, solo se identificó que miR-21 estaba sobreexpresado en ambas fuentes de miARN en diferentes grupos de pacientes (22). Dado que habitualmente los estudios analizan solo los miRNAs asociados a EV o el total de miRNAs circulantes, hasta donde sabemos, nuestro trabajo es el único que compara los miRNAs asociados a EV y los miRNAs de circulación libre extraídos del plasma empobrecido por EV, en el mismo grupo de pacientes (22).
Si bien la literatura sobre el cáncer, tanto los miARN circulantes como los asociados a EV, es abundante en medicina humana, se han realizado menos estudios en medicina veterinaria (2, 9, 10, 23-25).
Garnica y sus coautores investigaron la relación entre los miRNAs séricos asociados a EV y la respuesta a la terapia en perros con linfoma multicéntrico (LCM). Entre los miRNAs investigados, los autores encontraron que mir-222 se expresó más en el grupo que logró una respuesta completa, y mir-93 se sobreexpresó en el grupo que mostró enfermedad progresiva (9). En nuestro estudio, se encontró que miR-222 estaba sobreexpresado tanto en los miRNAs asociados a EV como en los miRNAs libres en el LG. Por otro lado, miR-93 mostró sobreexpresión en los miRNAs libres, pero no en los asociados a EV. La discrepancia en los perfiles de miRNA asociados a EV entre nuestro estudio y el de Garnica y sus colegas podría deberse a varios factores: (i) su enfoque específico en el MCL que involucra tanto a las células B como a las T, (ii) su examen exclusivo de los EV pequeños y (iii) su análisis de los EV derivados del suero en contraposición a los EV plasmáticos, matrices que han demostrado poseer diferentes perfiles de EV (9).
Los miRNAs circulantes en perros portadores de linfoma también se examinaron en otro estudio, que se centró en el análisis de los miRNAs séricos circulantes totales, abarcando pero no diferenciando entre miRNAs libres y asociados a EV (10). En este estudio, el único miRNA que se expresó diferencialmente y fue consistente con nuestros hallazgos fue miR-let-7b, regulado a la baja en casos de cL en ambos estudios y expresado diferencialmente como miRNA libre en nuestro análisis. Una vez más, los diferentes resultados podrían atribuirse de nuevo a los diferentes criterios de selección de casos (exclusivamente TcL frente a varios tipos de linfomas) y a la distinción entre el análisis de los miRNAs asociados a EV y los miRNAs séricos circulantes totales (10).
Por último, también se ha reportado que miR-18a, que se sobreexpresó entre los miRNAs libres de nuestro estudio, se ha reportado como sobreexpresado en perros con cáncer, específicamente en el suero de perros con carcinoma mamario (26).
De acuerdo con la literatura, todos los miRNAs identificados como expresados diferencialmente en muestras libres y asociadas a EV en este trabajo, se han observado previamente por su sobreexpresión o regulación negativa en tumores humanos y parecen desempeñar un papel importante tanto en la oncogénesis tumoral como en la oncosupresión (27-38). De hecho, algunos de los miRNAs que encontramos expresados diferencialmente en TcL han sido reportados como marcadores en el linfoma humano. Se ha encontrado que miR-21 y miR191 están sobreexpresados en el suero de pacientes con estadios avanzados de linfoma difuso de células B grandes (DLBCL), y también se ha informado que miR21 está sobreexpresado en EVs plasmáticos de pacientes con linfoma de Hodgkin (39, 40). El miR-30d, que se encontró sobreexpresado en los miRNAs asociados a EV, estaba, por el contrario, regulado a la baja entre el total de miRNAs séricos circulantes de pacientes humanos con leucemia linfocítica crónica (41). Curiosamente, miR-let-7b y miR-18a, previamente identificados como desregulados en el linfoma canino y el carcinoma mamario canino respectivamente, también se han observado como desregulados en los miRNAs séricos circulantes totales de pacientes humanos que padecen DLBCL. En particular, miR-let-7b se ha relacionado con etapas más avanzadas de esta enfermedad (10, 25, 42).
Los otros miRNAs encontrados como diferencialmente expresados en nuestro estudio, que hasta ahora no se han reportado en neoplasias malignas caninas o linfoma humano, se han investigado en otros tipos de cánceres humanos, y algunos de ellos se han detectado como asociados a VE circulantes. miR-146 y miR-223, que encontramos sobreexpresados en miRNAs libres y asociados a EV respectivamente, estaban subexpresados en EVs séricos de pacientes humanos con carcinoma de células escamosas de laringe (43). Se ha encontrado que miR-223 está sobreexpresado y regulado a la baja en pacientes con cáncer de ovario colorrectal y epitelial, respectivamente, cuando se asocian a EV (44, 45). En pacientes con cáncer colorrectal, también se ha encontrado que miR-192 y miR-27a se expresan diferencialmente cuando se encontró que se asociaba a EV y miR-30b, sobreexpresado entre los miRNAs asociados a EV en nuestro estudio, en las EV plasmáticas de mujeres con recurrencia de cáncer de mama (44, 46, 47).
Por último, miR-103a y miR-130a, que estaban, respectivamente, regulados a la baja y sobreexpresados cuando se asociaron a EV en nuestro estudio, se han encontrado sobreexpresados en los miRNAs séricos circulantes totales de pacientes humanos con cáncer de pulmón y cáncer de vejiga urinaria, respectivamente (48, 49). El miR-106, que hemos encontrado sobreexpresado entre los miRNAs libres, ha sido considerado un potencial biomarcador diagnóstico circulante para el carcinoma hepatocelular y un biomarcador pronóstico para el cáncer gástrico (50).
Si bien los hallazgos de este estudio se alinean hasta cierto punto con investigaciones previas en oncología humana y canina, lo que sugiere un subconjunto de miRNAs como posibles biomarcadores de diagnóstico temprano para TcL, debemos reconocer algunas limitaciones de este estudio. Se incluyó un número limitado de pacientes debido a las dificultades para obtener muestras de sangre de un subconjunto homogéneo de sujetos. Además, para excluir a los pacientes que habían sido sometidos a diferentes protocolos de tratamiento, las muestras solo se recogieron en el momento del diagnóstico. En consecuencia, para lograr resultados más sustanciales, las investigaciones adicionales deben involucrar a un grupo más grande y uniforme de individuos, incluidas muestras adicionales recolectadas en diferentes momentos después del diagnóstico. Estas investigaciones podrían centrarse en el análisis de los miRNAs aquí encontrados expresados diferencialmente, evaluando su eficacia no solo como biomarcadores diagnósticos, sino también pronósticos y predictivos. Esto implicaría agregar datos de seguimiento de grupos de muestra más grandes. Por otra parte, aunque la CU siempre se ha considerado el método de referencia para el aislamiento de VE y es una técnica muy bien establecida que permite un excelente margen de certeza para el aislamiento de VE, solo pudimos analizar la concentración, el tamaño y la caracterización de las VE en dos muestras utilizadas como controles de procedimiento. Además, existen otras técnicas que, según la literatura, permiten la recogida de muestras más puras siendo menos dependientes del operador (6, 51, 52). Por lo tanto, los estudios futuros también podrían considerar el uso y la comparación de diferentes técnicas de aislamiento de EV, posiblemente aplicándolas a todo el subconjunto de muestras analizadas.
5 Conclusión
Este estudio prospectivo preliminar de doble brazo compara por primera vez la expresión de miRNAs asociados a EV y de circulación libre en el plasma de perros con TcL y perros de control sanos. Se encontraron diez miRNAs asociados a EV y siete de circulación libre expresados diferencialmente entre LG y Ctrl, con has-miR-222-3p sobreexpresado tanto en miRNAs asociados a EV como en miRNAs de circulación libre. Varios miRNAs identificados como expresados diferencialmente en este estudio han sido reconocidos previamente como marcadores potenciales en TcL, lo que los convierte en candidatos prometedores para un análisis más profundo. La elección de utilizar miARN asociados a EV como biomarcadores en lugar de miARN de circulación libre tendría varias ventajas, ya que el contenido de EV es estable y está protegido de la degradación por la ARNasa circulante. Los miRNAs encontrados en este estudio podrían ser utilizados en estudios posteriores con un mayor número de pacientes e información terapéutica y de seguimiento adicional para investigar su posible uso también como biomarcadores pronósticos y predictivos. Estos resultados preliminares añaden nuevos conocimientos sobre los miRNAs asociados a EV en el campo de la medicina veterinaria, mejorando nuestra comprensión de su expresión en TcL. Nuestras expectativas son contribuir al desarrollo de herramientas diagnósticas y terapéuticas más rápidas, eficaces e innovadoras.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio están incluidas en el artículo/Material complementario, las consultas posteriores pueden dirigirse al autor correspondiente.
Declaración ética
No se requirió aprobación ética para los estudios con animales de acuerdo con la legislación local y los requisitos institucionales porque todas las muestras de sangre eran restos de los controles clínicos planificados necesarios. Se obtuvo el consentimiento escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
CC: Redacción – borrador original. VM: Análisis formal, Metodología, Validación, Redacción – borrador original. AM: Metodología, Redacción – revisión y edición. FT: Metodología, Redacción – revisión y edición. LF: Metodología, Redacción – revisión y edición. SF: Análisis formal, Metodología, Validación, Redacción – revisión y edición. FB: Metodología, Redacción – revisión y edición. CL: Conceptualización, Obtención de fondos, Administración de proyectos, Supervisión, Redacción, revisión y edición. VZ: Conceptualización, Obtención de fondos, Administración de proyectos, Supervisión, Redacción, revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio ha sido financiado por la beca científica AniCura (2020-2021: 10.000 €), que se ha utilizado para cubrir los costes del laboratorio. Ninguno de los autores ha sido remunerado con el uso de este fondo. Financiación de acceso abierto proporcionada por la Università degli Studi di Padova, Universidad de Padua, Comité de Ciencia Abierta.
Reconocimientos
Nos gustaría agradecer a la Dra. Michele Berlanda por proporcionar algunas de las muestras de control.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1461506/full#supplementary-material
Notas
1. ^www.geneglobe.qiagen.com/analyze
Referencias
1. Zandvliet, M . Linfoma canino: una revisión. Veterinario Q. (2016) 36:76–104. doi: 10.1080/01652176.2016.1152633
2. Craig, KKL, Wood, GA, Keller, SM, Mutsaers, AJ y Wood, RD. Perfil de microARN en el linfoma multicéntrico canino. PLoS Uno. (2019) 14:e0226357. doi: 10.1371/journal.pone.0226357
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Peng, Y, y Croce, CM. El papel de los microARN en el cáncer humano. Objetivo de transducción de señal. (2016) 1:15004. doi: 10.1038/sigtrans.2015.4
4. Abels, ER, y Breakefield, XO. Introducción a las vesículas extracelulares: biogénesis, selección de la carga de ARN, contenido, liberación y absorción. Célula Mol Neurobiol. (2016) 36:301–12. DOI: 10.1007/S10571-016-0366-Z
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Maacha, S, Bhat, AA, Jiménez, L, Raza, A, Haris, M, Uddin, S, et al. Comunicación intercelular mediada por vesículas extracelulares: roles en el microambiente tumoral y resistencia a fármacos anticancerosos. Cáncer de topo. (2019) 18:55. doi: 10.1186/s12943-019-0965-7
6. Welsh, JA, Goberdhan, DCI, O’Driscoll, L, Buzas, EI, Blenkiron, C, Bussolati, B, et al. Información mínima para estudios de vesículas extracelulares (MISEV2023): desde abordajes básicos hasta avanzados. J Vesículas extracelulares. (2024) 13:E12404. doi: 10.1002/jev2.12404
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Van Niel, G, D’Angelo, G, y Raposo, G. Arrojando luz sobre la biología celular de las vesículas extracelulares. Nat Rev Mol Cell Biol. (2018) 19:213–28. doi: 10.1038/nrm.2017.125
8. Urabe, F, Kosaka, N, Ito, K, Kimura, T, Egawa, S y Ochiya, T. Vesículas extracelulares como biomarcadores y dianas terapéuticas para el cáncer. Am J Physiol Célula Physiol. (2020) 318:C29-39. doi: 10.1152/ajpcell.00280.2019
9. Garnica, TK, Lesbon, JCC, Ávila, ACFCM, Rochetti, AL, Matiz, ORS, Ribeiro, RCS, et al. La biopsia líquida basada en pequeñas vesículas extracelulares predice la respuesta quimioteriana de los linfomas multicéntricos caninos. Sci Rep. (2020) 10:20371. doi: 10.1038/s41598-020-77366-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Fujiwara-Igarashi, A, Igarashi, H, Mizutani, N, Goto-Koshino, Y, Takahashi, M, Ohno, K, et al. Perfil de expresión de microRNAs séricos circulantes en perros con linfoma. Veterinario J. (2015) 205:317–21. doi: 10.1016/j.tvjl.2015.04.029
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Ponce, F, Marchal, T, Magnol, JP, Turinelli, V, Ledieu, D, Bonnefont, C, et al. Estudio morfológico de 608 casos de linfoma maligno canino en Francia, centrado en las similitudes comparativas entre la morfología del linfoma canino y humano. Veterinario Pathol. (2010) 47:414–33. doi: 10.1177/0300985810363902
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Théry, C, Witwer, KW, Aikawa, E, Alcaraz, MJ, Anderson, JD, Andriantsitohaina, R, et al. Información mínima para estudios de vesículas extracelulares 2018 (MISEV2018): una declaración de posición de la Sociedad Internacional de Vesículas Extracelulares y actualización de las guías MISEV2014. J Vesículas extracelulares. (2018) 7:1535750. doi: 10.1080/20013078.2018.1535750
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Moore, AS . Tratamiento del linfoma de células T en perros. Rec. Veterinario. (2016) 179:277–81. doi: 10.1136/vr.103456
14. Valli, VE, Kass, PH, Myint, MS y Scott, F. Linfomas caninos: asociación del tipo de clasificación, estadio de la enfermedad, subtipo tumoral, tasa mitótica y tratamiento con la supervivencia. Veterinario Pathol. (2013) 50:738–48. doi: 10.1177/0300985813478210
15. Paterson, E, Blenkiron, C, Danielson, K y Henry, C. Recomendaciones para la investigación de biomarcadores de miARN de vesículas extracelulares en el contexto del cáncer de endometrio. Trans Oncol. (2022) 23:101478. doi: 10.1016/j.tranon.2022.101478
16. Narita, M, Nishida, H, Asahina, R, Nakata, K, Yano, H, Ueda, T, et al. Identificación de genes de referencia para microRNAs de vesículas extracelulares aisladas de muestras de plasma de perros sanos mediante métodos de ultracentrifugación, precipitación y cromatografía de afinidad de membrana. AJVR. (2019) 80:449–54. doi: 10.2460/ajvr.80.5.449
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Kulka, M, Brennan, K y Mc Gee, M. Investigación de vesículas extracelulares caninas en linfomas difusos de células B grandes. PLoS Uno. (2022) 17:e0274261. doi: 10.1371/journal.pone.0274261
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Endzelinš, E, Berger, A, Melne, V, Bajo-Santos, C, Sobolevska, K, Abols, A, et al. Detección de miRNAs circulantes: análisis comparativo de miRNAs extracelulares incorporados a vesículas y miRNAs libres de células en plasma entero de pacientes con cáncer de próstata. BMC Cáncer. (2017) 17:730. doi: 10.1186/s12885-017-3737-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Chevillet, JR, Kang, Q, Ruf, IK, Briggs, HA, Vojtech, LN, Hughes, SM, et al. Análisis cuantitativo y estequiométrico del contenido de microRNA de los exosomas. Proc Natl Acad Sci U S A. (2014) 111:14888–93. doi: 10.1073/pnas.1408301111
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Dhondt, B, Rousseau, Q, De Wever, O y Hendrix, A. Función de los miRNAs asociados a vesículas extracelulares en la metástasis. Res. de Tejido Celular. (2016) 365:621–41. doi: 10.1007/s00441-016-2430-x
21. Groot, M y Lee, H. Mecanismos de clasificación de microARN en vesículas extracelulares y sus enfermedades asociadas. células. (2020) 9:1044. doi: 10.3390/cells9041044
22. de Nóbrega, M, dos Reis, MB, Pereira, ÉR, de Souza, MF, y de Syllos Cólus, IM. El potencial de los microRNAs libres de células y exosomales como biomarcadores en la biopsia líquida en pacientes con cáncer de próstata. J Cáncer Res Clin Oncol. (2022) 148:2893–910. doi: 10.1007/s00432-022-04213-9
23. Varvil, MS, y dos Santos, AP. Una revisión sobre los estudios de detección y expresión de microARN en perros. Frente Vet Sci. 10:1261085. doi: 10.3389/fvets.2023.1261085
24. Petroušková, P, Hudáková, N, Maloveská, M, Humeník, F y Cizkova, D. MiRNAs no exosomales y derivados de exosomas como biomarcadores prometedores en el cáncer de mama canino. Vida. (2022) 12:524. doi: 10.3390/vida12040524
25. Fish, EJ, Martínez-Romero, EG, DeInnocentes, P, Koehler, JW, Prasad, N, Smith, AN, et al. MicroARN circulantes como biomarcadores del carcinoma mamario canino en perros. J Veterinario Pasante Med. (2020) 34:1282–90. doi: 10.1111/jvim.15764
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Fish, EJ, Irizarry, KJ, DeInnocentes, P, Ellis, CJ, Prasad, N, Moss, AG, et al. Las células epiteliales mamarias caninas malignas desprenden exosomas que contienen microARN expresados diferencialmente que regulan las redes oncogénicas. BMC Cáncer. (2018) 18:832. doi: 10.1186/s12885-018-4750-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Liang, J, Tang, J, Shi, H, Li, H, Zhen, T, Duan, J, et al. miR-27a-3p dirigido a RXRα promueve la progresión del cáncer colorrectal mediante la activación de la vía Wnt/β-catenina. Oncotarget. (2017) 8:82991–3008. doi: 10.18632/oncotarget.19635
28. Liu, Z, Wei, X, Zhang, A, Li, C, Bai, J y Dong, J. El ARN largo no codificante HNF1A-AS1 funcionó como oncogén y promotor de autofagia en el carcinoma hepatocelular a través de la esponja has-miR-30b-5p. Biochem Biophys Res Commun. (2016) 473:1268–75. doi: 10.1016/j.bbrc.2016.04.054
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Rao, X, Wan, L, Jie, Z, Zhu, X, Yin, J y Cao, H. El miR-27a-3p regulado al alza indica un mal pronóstico en pacientes con carcinoma de páncreas y promueve la angiogénesis y la migración mediante el silenciamiento epigenético de GATA6 y la activación de la vía de señalización VEGFA/VEGFR2. Onco se dirige a Ther. (2019) 12:11241–54. doi: 10.2147/OTT. S220621
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Yang, Y, Yang, Z, Zhang, R, Jia, C, Mao, R, Mahati, S, et al. MiR-27a-3p mejora la sensibilidad al cisplatino en células de carcinoma hepatocelular mediante la inhibición de la vía PI3K/Akt. Biosci Rep. (2021) 41:BSR20192007. DOI: 10.1042/BSR20192007
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Eldosoky, MA, Hammad, R, Elmadbouly, AA, Aglan, RB, Abdel-Hamid, SG, Alboraie, M, et al. Importancia diagnóstica del panel de has-miR-21-5p, has-miR-192-5p, has-miR-155-5p, has-miR-199a-5p y proporciones en el carcinoma hepatocelular además de la cirrosis hepática en pacientes infectados por el VHC. Int J Mol Sci. (2023) 24:3157. doi: 10.3390/ijms24043157
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Furuke, H, Konishi, H, Arita, T, Kataoka, S, Shibamoto, J, Takabatake, K, et al. El microRNA-192-5p plasmático puede predecir la respuesta a la quimioterapia neoadyuvante y el pronóstico en el cáncer de esófago. Cáncer Sci. (2023) 114:1686–96. doi: 10.1111/cas.15703
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Gao, X, Xie, Z, Wang, Z, Cheng, K, Liang, K y Song, Z. La sobreexpresión de miR-191 predice un mal pronóstico y promueve la proliferación e invasión en el carcinoma de células escamosas de esófago. Yonsei Med J. (2017) 58:1101–10. doi: 10.3349/ymj.2017.58.6.1101
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Él, Y, Cui, Y, Wang, W, Gu, J, Guo, S, Ma, K, et al. La hipometilación del locus has-miR-191 causa una alta expresión de has-miR-191 y promueve la transición epitelial a mesenquimal en el carcinoma hepatocelular. Neoplasia. (2011) 13:841–IN23. doi: 10.1593/neo.11698
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Polioudakis, D, Abell, NS, e Iyer, VR. MiR-191 regula la proliferación primaria de fibroblastos humanos y se dirige directamente a múltiples oncogenes. PLoS Uno. (2015) 10:e0126535. doi: 10.1371/journal.pone.0126535
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Peng, WZ, Ma, R, Wang, F, Yu, J y Bin, LZ. Papel del grupo miR-191/425 en la tumorigénesis y el diagnóstico del cáncer gástrico. Int J Mol Sci. (2014) 15:4031–48. doi: 10.3390/ijms15034031
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Tian, F, Yu, C, Wu, M, Wu, X, Wan, L y Zhu, X. MicroRNA-191 promueve la proliferación de células de carcinoma hepatocelular por eje has_circ_0000204 / miR-191 / KLF6. Célula Prolif. (2019) 52:E12635. doi: 10.1111/cpr.12635
38. Di Leva, G, Garofalo, M, y Croce, CM. MicroARN en el cáncer. Ann Rev Pathol. (2014) 9:287–314. doi: 10.1146/annurev-pathol-012513-104715
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. van Eijndhoven, MAJ, Zijlstra, JM, Groenewegen, NJ, Drees, EEE, van Niele, S, Baglio, SR, et al. MiRNAs de vesículas plasmáticas para la monitorización de la respuesta terapéutica en pacientes con linfoma de Hodgkin. Perspectiva de la JCI. (2016) 1:e89631. doi: 10.1172/jci.insight.89631
40. Ren, JL, Cui, LZ, Hao, XX y Li, XY. Correlación entre miR-21, miR-191 y el estadio clínico de los pacientes con linfoma difuso de células B grandes. Zhongguo Shi Yan Xue Ye Xue Za Zhi. (2023) 31:115–9. doi: 10.19746/j.cnki.issn.1009-2137.2023.01.018
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Farzadfard, E, Kalantari, T y Tamaddon, G. Expresión sérica de siete microARN en pacientes con leucemia linfocítica crónica. J Sangre Med. (2020) 11:97–102. doi: 10.2147/JBM. S230842
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Beheshti, A, Stevenson, K, Vanderburg, C, Ravi, D, McDonald, JT, Christie, AL, et al. Identificación de firmas séricas circulantes de múltiples micro-microARN en modelos humanos de DLBCL. Sci Rep. (2019) 9:17161. DOI: 10.1038/s41598-019-52985-X
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Genc, S, Yagci, T, Vageli, DP, Dundar, R, Doukas, PG, Doukas, SG, et al. Perfiles exosomales de MicroRNA-223, MicroRNA-146 y MicroRNA-21 y cambios bioquímicos en el cáncer de laringe. ACS Pharmacol Trans Sci. (2023) 6:820–8. doi: 10.1021/acsptsci.3c00038
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. de Miguel, PD, Rodriguez Martínez, A, Ortigosa Palomo, A, Delgado Ureña, M, Garcia Puche, JL, Robles Remacho, A, et al. MiRNAs de vesículas extracelulares como biomarcadores de biopsia líquida para la identificación y el pronóstico de enfermedades en pacientes con cáncer colorrectal metastásico. Sci Rep. (2020) 10:1–13. doi: 10.1038/s41598-020-60212-1
45. Pan, C, Stevic, I, Müller, V, Ni, Q, Oliveira-Ferrer, L, Pantel, K, et al. MicroRNAs exosomales como marcadores tumorales en el cáncer epitelial de ovario. Mol Oncol. (2018) 12:1935–48. doi: 10.1002/1878-0261.12371
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Liu, X, Pan, B, Sun, L, Chen, X, Zeng, K, Hu, X, et al. Los miR-27a y miR-130a exosomales circulantes actúan como nuevos biomarcadores diagnósticos y pronósticos del cáncer colorrectal. Cáncer Epidemiol Biomark Preven. (2018) 27:746–54. doi: 10.1158/1055-9965.EPI-18-0067
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Ni, Q, Stevic, I, Pan, C, Müller, V, Oliviera-Ferrer, L, Pantel, K, et al. Diferentes firmas de miR-16, miR-30b y miR-93 en exosomas de pacientes con cáncer de mama y CDIS. Sci Rep. (2018) 8:12974. DOI: 10.1038/s41598-018-31108-Y
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Canatan, D, Sönmez, Y, Yılmaz, Ö, Coşkun, HŞ, Göksu, SS, Uçar, S, et al. MicroRNAs como biomarcador en cáncer de pulmón. Acta Biomedica. (2023) 94:e2023045. doi: 10.23750/abm.v94i1.13334
49. Wang, J, Zhao, L, Peng, X, Liu, K, Zhang, C, Chen, X, et al. Evaluación de los miembros de la familia miR-130 como biomarcadores circulantes para el diagnóstico de cáncer de vejiga. J Clin Lab Anal. (2020) 34:E23517. doi: 10.1002/jcla.23517
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Jiang, L, Li, X, Cheng, Q y Zhang, BH. El microARN plasmático podría ser un biomarcador potencial para el carcinoma hepatocelular y el cribado de enfermedades hepáticas crónicas. Tumor Biol. (2015) 36:7167–74. doi: 10.1007/s13277-015-3446-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Coumans, FAW, Brisson, AR, Buzas, EI, Dignat-george, F, Drees, EEE, El-andaloussi, S, et al. Revisar las guías metodológicas de vesículas extracelulares para el estudio de las vesículas extracelulares. Circ Res. (2017) 120:1632–48. doi: 10.1161/CIRCRESAHA.117.309417
52. Witwer, KW, Buzás, EI, Bemis, LT, Bora, A, Lässer, C, Lötvall, J, et al. Estandarización de los métodos de recogida, aislamiento y análisis de muestras en la investigación de vesículas extracelulares. J Vesículas extracelulares. (2013) 2:27. doi: 10.3402/jev.v2i0.20360
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: vesículas extracelulares, miRNA, biopsia líquida, oncología veterinaria, linfoma canino
Cita: Capuano C, Moccia V, Molinari A, Torrigiani F, Ferro L, Ferraresso S, Bonsembiante F, Leo C y Zappulli V (2024) Expresión de microARN asociada a vesículas extracelulares versus circulación libre en el linfoma de células T caninas. Frente. Vet. Sci. 11:1461506. doi: 10.3389/fvets.2024.1461506
Editado por:
Attilio Corradi, Universidad de Parma, Italia
Revisado por:
Fabrizio Ceciliani, Universidad de Milán, Italia
Naoki Miura, Universidad de Kagoshima, Japón
Derechos de autor © 2024 Capuano, Moccia, Molinari, Torrigiani, Ferro, Ferraresso, Bonsembiante, Leo y Zappulli. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Atribución (CC BY).
*Correspondencia: Valentina Moccia, valentina.moccia@unipd.it
†Estos autores han contribuido por igual a este trabajo y comparten la primera autoría
Página webEstos autores han contribuido por igual a este trabajo y comparten la última autoría
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía