
Diferentes fuentes de heno de alfalfa alteran la composición de la microbiota ruminal en vacas

Diferentes fuentes de heno de alfalfa alteran la composición de la microbiota ruminal en vacas Holstein a mitad de lactancia sin afectar el rendimiento productivo
 Shaokai LaUn †
Shaokai LaUn † Hao LiUn †
Hao LiUn † Yan Zhang1
Yan Zhang1 Muhammad Abaidullah1
Muhammad Abaidullah1 Jiakuan Niu1
Jiakuan Niu1 Zimin Gao1
Zimin Gao1 Boshuai Liu1
Boshuai Liu1 Sen Ma1,2,3
Sen Ma1,2,3 Yalei Cui1,2,3
Yalei Cui1,2,3 Defeng Li1,2,3
Defeng Li1,2,3 Yinghua Shi1,2,3*
Yinghua Shi1,2,3*- 1Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Henan, Zhengzhou, China
- número arábigoLaboratorio Clave de Innovación y Utilización de Recursos de Pastizales de Henan, Zhengzhou, China
- 3Centro de Investigación de Tecnología de Ingeniería de Forraje de Henan, Zhengzhou, China
El heno de alfalfa es un ingrediente importante y de uso común en la producción lechera. Para expandir mejor el mercado de suministro de alfalfa, es de gran importancia explorar el impacto del heno de alfalfa de diferentes fuentes en el rendimiento de la producción de vacas lecheras. Este estudio comparó los efectos del heno de alfalfa importado de América (AAH) y España (SAH) sobre el rendimiento de la lactancia y la microbiota ruminal de las vacas. Trescientas sesenta vacas Holstein sanas a mitad de lactancia con peso corporal, producción de leche y paridad similares se dividieron aleatoriamente en dos grupos alimentados con dietas basadas en AAH o SAH durante un período experimental de 70 días. Cada grupo estaba compuesto por cuatro corrales, con 45 vacas en cada corral. Se llevaron registros diarios de la IM por vaca y de la ingesta de materia seca por corral. Se tomaron muestras de doce vacas seleccionadas al azar por grupo para recolectar leche, heces, líquido ruminal y sangre. Los resultados no revelaron diferencias significativas entre los dos grupos en términos de rendimiento de producción, digestibilidad aparente de nutrientes, índices bioquímicos séricos o parámetros de fermentación ruminal. Sin embargo, la composición microbiana del rumen difirió significativamente entre los dos grupos de vacas en función de la diversidad de β. A nivel de género, la abundancia relativa de Prevotella, Succinivibrionaceae_UCG-002 aumentó, mientras que la de NK4A214_group, Ruminococcus, norank_f_F082 y Lachnospiraceae_NK3A20_group disminuyó en el grupo SAH en comparación con el grupo AAH. No hubo correlación significativa entre estas bacterias diferenciales centrales y las proporciones molares de acetato y propionato, la concentración de ácidos grasos volátiles totales y la producción de leche. En conclusión, los efectos alimentarios de la HSA fueron similares a los de la AAH. Estos hallazgos proporcionaron una referencia para la aplicación de heno de alfalfa de diferentes fuentes y para la mejora del beneficio económico de las granjas lecheras.
1 Introducción
La alfalfa es ampliamente utilizada en la ganadería debido a su buena palatabilidad, bajo contenido de fibra y alta concentración de proteínas (que oscila entre 17 y 22%) (1). La alimentación con alfalfa puede reducir eficazmente la proporción de concentrados, especialmente la de ingredientes ricos en proteínas en las dietas (2). Por lo tanto, la alfalfa se ha convertido en un forraje insustituible en la ganadería lechera, y el impacto beneficioso del heno de alfalfa en el rendimiento lácteo de las vacas lecheras ha sido ampliamente reconocido.
La demanda de heno de alfalfa de primera calidad ha aumentado significativamente en los últimos años debido a un aumento en el número de vacas lecheras y una mejora en la producción de leche (MY) de las vacas lecheras (3). Por lo general, las granjas lecheras dependen de múltiples proveedores para su alfalfa, lo que contribuye al funcionamiento estable de las granjas. Los productores de leche deben seleccionar heno de alfalfa de diferentes fuentes en función de factores como el precio para maximizar sus beneficios. Especialmente desde el estallido de la pandemia de COVID-19, el precio y la estabilidad del suministro de heno de alfalfa se vieron desafiados debido a problemas de transporte y otros factores. Como resultado, la sustitución de heno de alfalfa de diferentes fuentes se había vuelto más frecuente. Es bien sabido que los impactos de la alfalfa en el rendimiento productivo de las vacas lecheras no son fijos, ya que su composición de nutrientes es variable según la raza (4), los métodos de almacenamiento (5) y el período de cosecha. Por ejemplo, Vagnoni y Broderick (6) encontraron que se degradaba más proteína bruta en el rumen en las vacas que recibieron ensilado de alfalfa que en las que recibieron heno de alfalfa, mientras que las vacas que recibieron heno de alfalfa tuvieron una mayor ingesta de materia seca (DMI). Además, estudios previos han realizado ensayos comparando el heno de alfalfa con otros forrajes como la paja de trigo y las plántulas de maní (7-9). Sin embargo, ha habido pocos estudios que comparen cómo el heno de alfalfa de diversas fuentes afecta a las vacas lecheras. A pesar de los perfiles nutricionales similares, los cambios frecuentes de heno de alfalfa de diversas fuentes, especialmente los de diferentes países, han suscitado preocupaciones entre los productores de leche debido a la amenaza potencial para la estabilidad del rendimiento de la producción de vacas lecheras.
Los rumiantes poseen una estructura fisiológica única que interactúa sinérgicamente con las bacterias del rumen para facilitar la fermentación y la degradación del forraje en ácidos grasos volátiles (AGV). Estos AGV sirven como una fuente vital de nutrición para los rumiantes (10). De hecho, la fermentación de la fibra en el rumen produce dos AGV lipogénicos diferentes: acetato y butirato (11). Una porción significativa de butirato se convierte en β-hidroxibutirato dentro del tejido de la pared del rumen. Este acetato y β-hidroxibutirato sirven como sustrato para aproximadamente la mitad de la grasa que se encuentra en la leche. El heno de alfalfa, como forraje rico en proteínas, desempeña un papel crucial en los microorganismos del rumen y, por lo tanto, afecta el rendimiento de la producción de las vacas lecheras (12).
Por lo tanto, este experimento comparó los efectos de diferentes fuentes de heno de alfalfa importado de América (AAH) o España (SAH) con una composición de nutrientes similar sobre el rendimiento de la lactancia, la digestibilidad aparente de los nutrientes, los índices bioquímicos séricos, la fermentación ruminal y la microbiota de las vacas lecheras. Nuestra hipótesis es que el heno de alfalfa de diferentes fuentes con perfiles nutricionales similares no afectará el rendimiento de la producción de vacas lecheras, proporcionando a los productores lecheros más opciones para mejorar la eficiencia económica de la granja lechera.
2 Materiales y métodos
2.1 Heno de alfalfa
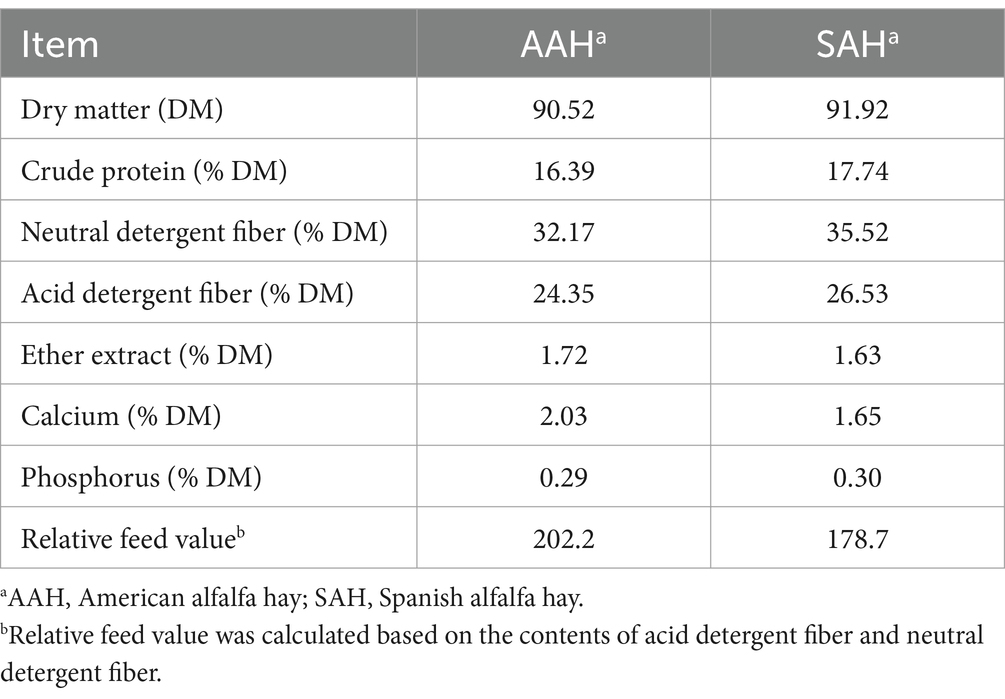
El heno de alfalfa utilizado en este estudio, obtenido en España y Estados Unidos, respectivamente, se compró a Literana, LLC y Stone Wings II, LLC. Compramos 60 toneladas de heno de alfalfa americana y otra española, con 60 fardos de cada uno. Se seleccionaron aleatoriamente 10 pacas y se recolectaron 500 g de muestras de las capas superior, media e inferior de cada paquete, y se mezclaron para la determinación de la composición nutricional. La Tabla 1 muestra los nutrientes presentes en el heno de alfalfa.
2.2 Vacas, diseño experimental y dietas
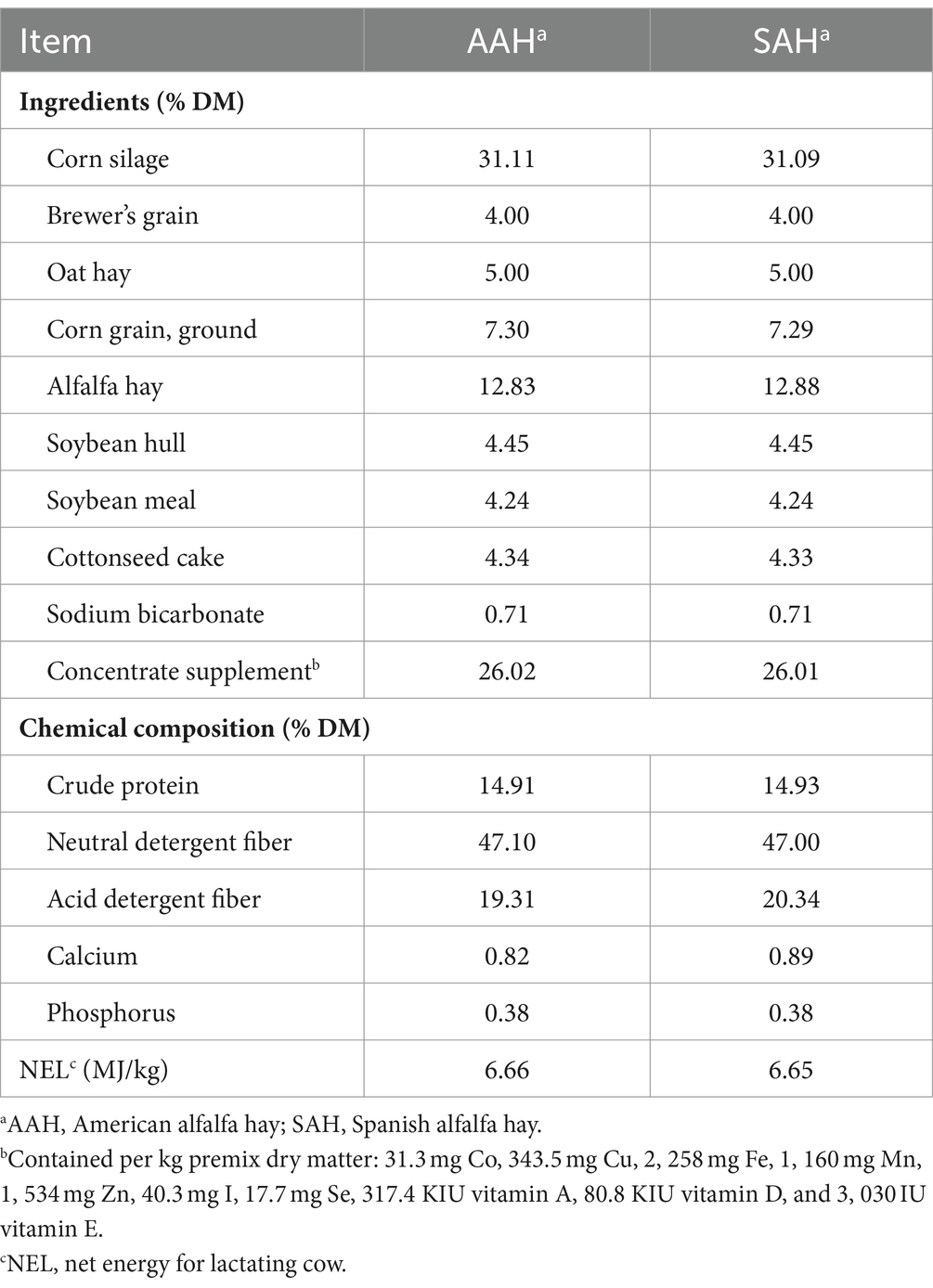
Este estudio fue aprobado por el Comité Institucional de Cuidado y Uso de Animales de la Universidad de Agricultura de Henan (Zhengzhou, China) (Número de aprobación: HNND2021062812). El ensayo se llevó a cabo en la granja de cría de animales Xincai Ruiya, provincia de Henan. Trescientas sesenta vacas lecheras Holstein sanas a mitad de lactancia (MY = 21,98 ± 5,02 kg; días en leche = 208 ± 19,42 d; paridad = 2,34 ± 0,47, media ± DE) fueron asignadas aleatoriamente a dos grupos (cuatro réplicas en cada grupo y 45 vacas en cada réplica) alimentadas con dietas basadas en AAH o SAH. El ensayo duró 70 días, con 10 días para la adaptación y 60 días para la recolección de datos y muestras. La composición dietética y los niveles de nutrientes se muestran en la Tabla 2. Todas las vacas fueron alimentadas dos veces al día (07:00 y 19:00 h) y ordeñadas 3 veces (06:30, 14:30 y 22:30 h) al día, y tuvieron acceso a agua en todo momento. El establo se limpiaba y desinfectaba una vez a la semana.
2.3 Recogida de datos y muestras
Durante el período de recolección, se anotó diariamente el alimento ofrecido y rechazado para cada replicación para determinar el DMI. Diariamente se registró el IM de cada una de las 360 vacas experimentales. Además, se recogieron diariamente muestras de ración mezclada total (TMR). Una vez finalizado el experimento, las muestras de TMR se mezclaron en proporción para determinar la composición nutricional de la TMR. Para determinar si había diferencias en la composición de la leche, la digestibilidad aparente de los nutrientes y la microbiota ruminal entre los dos grupos de vacas, se seleccionaron aleatoriamente 12 vacas de cada grupo (había tres vacas en cada repetición) para tomar muestras de leche, heces, sangre y líquido ruminal. Las muestras de leche se recolectaron los días 0, 14, 28, 42, 56 y los últimos 4 días del experimento. Para cada vaca, se recolectaron 40 mL de muestras de leche por día de recolección (en una proporción de 4:3:3 por la mañana, al mediodía y por la noche) y se colocaron en un tubo de centrífuga que contenía conservante de dicromato de potasio. Las muestras de leche se refrigeraron a 4°C para el análisis de la composición de la leche. Las muestras fecales se recogieron 4 veces al día (06:00, 12:00, 18:00 y 24:00 h) durante los últimos 4 días del experimento. Se recolectaron aproximadamente 250 g de heces por vaca cada vez utilizando un método de muestreo rectal, mezclado, y 120 g de las muestras fecales se pesaron y mezclaron con 30 mL de ácido tartárico al 10%. Las muestras de heces recogidas durante los 4 días se mezclaron por separado al final del experimento para su análisis químico. La sangre se recolectó a través de la vena caudal con tubos de vacío de 5 mL antes de la alimentación matutina el último día del ensayo. La sangre se envió inmediatamente al laboratorio y se centrifugó a 3.000 × g durante 15 min a 4 °C. Y luego el suero se juntó en tubos de centrífuga de 2 mL y se mantuvo a -20 °C hasta que se analizó. El líquido ruminal se recolectó el último día del ensayo utilizando una sonda gástrica con un diámetro exterior de 1 cm, un diámetro interno de 0,8 cm y una longitud de 300 cm, 2 h después de la alimentación matutina. Para evitar la contaminación de la saliva, se extrajeron y desecharon alrededor de 200 mL de líquido ruminal de vacas individuales. A continuación, se recogieron 200 mL de líquido ruminal en un vaso de precipitados de 500 mL y se enviaron inmediatamente al laboratorio para su análisis de pH utilizando un medidor PHS-10 (Sartorius, Göttingen, Alemania) y registro. Después de la filtración a través de 4 capas de gasa, el líquido ruminal se dividió en 3 porciones, cada una con 10 mL. A continuación, estas muestras se congelaron y almacenaron a -80 °C para la posterior determinación de AGV y amoníaco (NH3-N), así como para la secuenciación del ARNr 16S.
2.4 Métodos analíticos
Las concentraciones de materia seca (MS), extracto de éter y fibra detergente ácida en heno de alfalfa, heces y muestras de TMR, así como las cenizas insolubles en ácido en heces y muestras de TMR, se determinaron utilizando los métodos descritos en AOAC (13). La concentración de fibra detergente neutra en muestras de heno de alfalfa, heces y TMR se determinó utilizando el método de Van Soest et al. (14). Las concentraciones de proteína bruta y nitrógeno se determinaron utilizando un analizador automático de N Kjeldahl (SKD-2000, Haineng Experimental Instrument Technology Co., Ltd., Shanghai, China). Las concentraciones de calcio y fósforo en heno de alfalfa y TMR se analizaron con base en el método de Mattioli et al. (15). Se utilizó como indicador endógeno cenizas insolubles en ácido, según la fórmula: la digestibilidad aparente de un nutriente (%) = 100 − 100 × la concentración de nutrientes en las heces (%) × la concentración de cenizas insolubles en ácido en TMR (%)/la concentración de nutrientes en TMR (%)/concentración de cenizas insolubles en ácido en las heces (%), calculando la digestibilidad aparente de la proteína bruta, el extracto de éter, la fibra detergente ácida, y fibra detergente neutra.
Un analizador de leche automatizado de infrarrojo cercano (Seris300 CombiFOSS; Foss Electric, Hillerd, Dinamarca) para medir las concentraciones de grasa láctea, proteína, lactosa, sólidos lácteos, nitrógeno ureico (MUN) y recuento de células somáticas en muestras de leche entregadas al Centro de Pruebas de Mejora del Rebaño Lechero en la provincia de Henan. Las concentraciones séricas de glucosa, proteínas totales, albúmina, globulina, triglicéridos, colesterol, ácidos grasos no esterificados, β-hidroxibutirato, urea, alanina aminotransferasa y aspartato aminotransferasa se evaluaron utilizando kits de ensayo disponibles comercialmente obtenidos de Nanjing Jian Cheng Biological Technology Co. (Nanjing, Jiangsu, China).
Para las muestras de fluido ruminal utilizadas para la determinación de AGV, después de descongelar a 4 °C, se trataron con ácido metafosfórico al 25%. A continuación, se determinaron las concentraciones de AGV (acetato, propionato, butirato, isobutirato, valerato e isovalerato) utilizando un cromatógrafo iónico alemán Sykam y una columna Dionex AS11-HC (4 × 250 mm, Thermo Fisher Science, Waltham, MA, Estados Unidos). Se aplicaron las siguientes circunstancias cromatográficas: una temperatura de columna de 30 °C, un caudal de 1 mL/min y un volumen de inyección de 10 μL; la fase móvil fue de 0,1 mmol/L y 50 mmol/L de soluciones de NaOH. El primero se mantuvo durante 28 min y el segundo durante 5 min. Finalmente, se mantuvieron 0,1 mmol/L de solución de NaOH durante 10 min. Para las muestras de fluido ruminal utilizadas para la determinación de NH3-N, después de la descongelación a 4 °C, la concentración de NH3-N se determinó utilizando un espectrofotómetro UV-2100 (UV2100, Shanghai Younike Instrument Co., Ltd., Shanghai, China) según AOAC (13).
2.5 Extracción de ADN y secuenciación del gen 16S rRNA
Se seleccionaron aleatoriamente seis muestras de líquido ruminal de cada grupo para la extracción de ADN y la secuenciación del gen 16S rRNA. De acuerdo con las instrucciones del fabricante, el ADN bacteriano total del líquido ruminal se obtuvo utilizando el kit de ADN del suelo E.Z.N.A. (® Qiagen, Estados Unidos). Se utilizó el espectrofotómetro UV-vis NanoDrop-2000 (Thermo Scientific, Wilmington, Estados Unidos) para detectar las concentraciones de ADN. Además, se utilizó electroforesis en gel de agarosa al 1% para evaluar la calidad del ADN. La región bacteriana V3-V4 del gen 16S rRNA se amplificó utilizando cebadores para 338F (5′-ACTCCTACGGGAGGCAGCAGCAG-3′) y 806R (5′-GGACTACHVGGGTWTCTAAT-3′) a una temperatura de 55 °C, como se describió anteriormente (16). Tras la purificación y cuantificación, los productos de PCR fueron utilizados por Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China) para construir bibliotecas y secuenciarlas en la plataforma Illumina MiSeq PE300/NovaSeq PE250 (Illumina, San Diego, Estados Unidos) de acuerdo con los protocolos establecidos. Todos los resultados se basaron en lecturas secuenciadas y unidades taxonómicas operativas. El número de acceso de BioProject para los datos de secuenciación del ARNr 16S, que se cargaron en la base de datos del NCBI, es PRJNA898964.
2.6 Tratamiento de los datos de secuenciación
Las secuencias optimizadas se obtuvieron mediante el empalme de los resultados de la secuenciación con control de calidad de secuencia de doble extremo (17) y utilizando FLASH versión 1.2.7 (18). Las secuencias optimizadas se basaron en la base de datos de clasificación de especies silva138/16s_bacteria y se sometieron a un análisis de agrupamiento de unidades taxonómicas operativas (OTU) (nivel de confianza 0,7) con un 97% de similitud utilizando la versión 7.1 de UPARSE (19, 20), y se generó una tabla de estadísticas de clasificación de especies de OTU mediante el cálculo de la paridad con el número mínimo de secuencias de muestra. El análisis del número de especies, el análisis de la diversidad alfa (índices Chao1 y Simpson), el análisis de la diversidad beta (análisis de coordenadas principales), el análisis de la composición de la comunidad y el mapeo se llevaron a cabo en la herramienta online de Majorbio Cloud Platform. 1 Los diagramas de Venn, que muestran visualmente el número de OTUs comunes y únicas entre grupos, y la curva de rarefacción (índice de Shannon) fueron dibujados por el software R (versión 3.3.1). Con el fin de ilustrar la variedad y abundancia de las comunidades microbianas, los índices de Chao1 y Simpson fueron calculados por Mothur (versión 1.30.2) (21). Posteriormente, se calculó la distancia de Bray-Curtis utilizando el software Qiime (versión 1.9.1), y el gráfico de análisis de coordenadas principales se generó utilizando el software R (versión 3.3.1). Se utilizó el paquete Vegan basado en el software R (versión 3.6.0) para probar las diferencias en la estructura de la comunidad microbiana entre diferentes tratamientos a través de ANOSIM. La abundancia relativa de filos y géneros se mostró mediante un gráfico de barras y un mapa de calor, que se dibujaron con el software R (versión 3.3.1). Se utilizó el método de la prueba de suma de rangos de Wilcoxon para analizar e identificar las diferencias entre los dos grupos. Para calcular el impacto de la abundancia de especies en el efecto de diferencia a nivel de género, se utilizó el análisis discriminante lineal del tamaño del efecto (LEfSe) y el análisis discriminante lineal. Se calcularon los coeficientes de correlación de Spearman entre las 20 especies principales a nivel de género y varios parámetros de fermentación ruminal, así como la producción de leche y la composición de la leche utilizando el paquete vegano en lenguaje R (versión 3.3.1) y se mostraron en el mapa de calor. Para predecir las funciones de la microbiota ruminal, se obtuvieron predicciones de función de PICRUSt2 basadas en la base de datos KEGG. 2 Las diferencias en las vías de KEGG entre dos grupos se evaluaron mediante la prueba t de Welch bilateral utilizando el software STAMP (versión 2.1.3). El método de tasa de falsos descubrimientos de Storey se utilizó para múltiples correcciones de prueba según lo recomendado por los desarrolladores de STAMP. Las 20 información biológica más importante (funcional) se seleccionaron mediante una clasificación basada en el tamaño del efecto (22).
2.7 Análisis estadístico
Para el análisis estadístico se utilizó la MY diaria de 360 vacas y el DMI por repetición a lo largo del experimento. Los datos de 24 vacas utilizados para la recolección de muestras no mostraron valores atípicos (puntos de datos fuera de ±3 desviaciones estándar de la media), y para el análisis estadístico se utilizaron su IM, composición de la leche, parámetros bioquímicos sanguíneos y parámetros de fermentación ruminal. Las mediciones de la composición de la leche, los parámetros bioquímicos de la sangre y los parámetros de fermentación ruminal se repitieron al menos 3 veces. La prueba de normalidad y la prueba de homogeneidad de varianza se llevaron a cabo utilizando el modelo PROC UNIVARIATE y el modelo PROC DISCRIM, respectivamente. De acuerdo con las características de los datos, se realizó un ANOVA de una vía o prueba no paramétrica utilizando el modelo PROC MIXED, y los resultados se expresaron como medias. Los valores de p >0,05 y <0,10 se consideraron una tendencia significativa, mientras que los valores de p <0,05 se consideraron significativos.
3 Resultados
3.1 Influencia en el rendimiento de la producción y la digestibilidad de los nutrientes
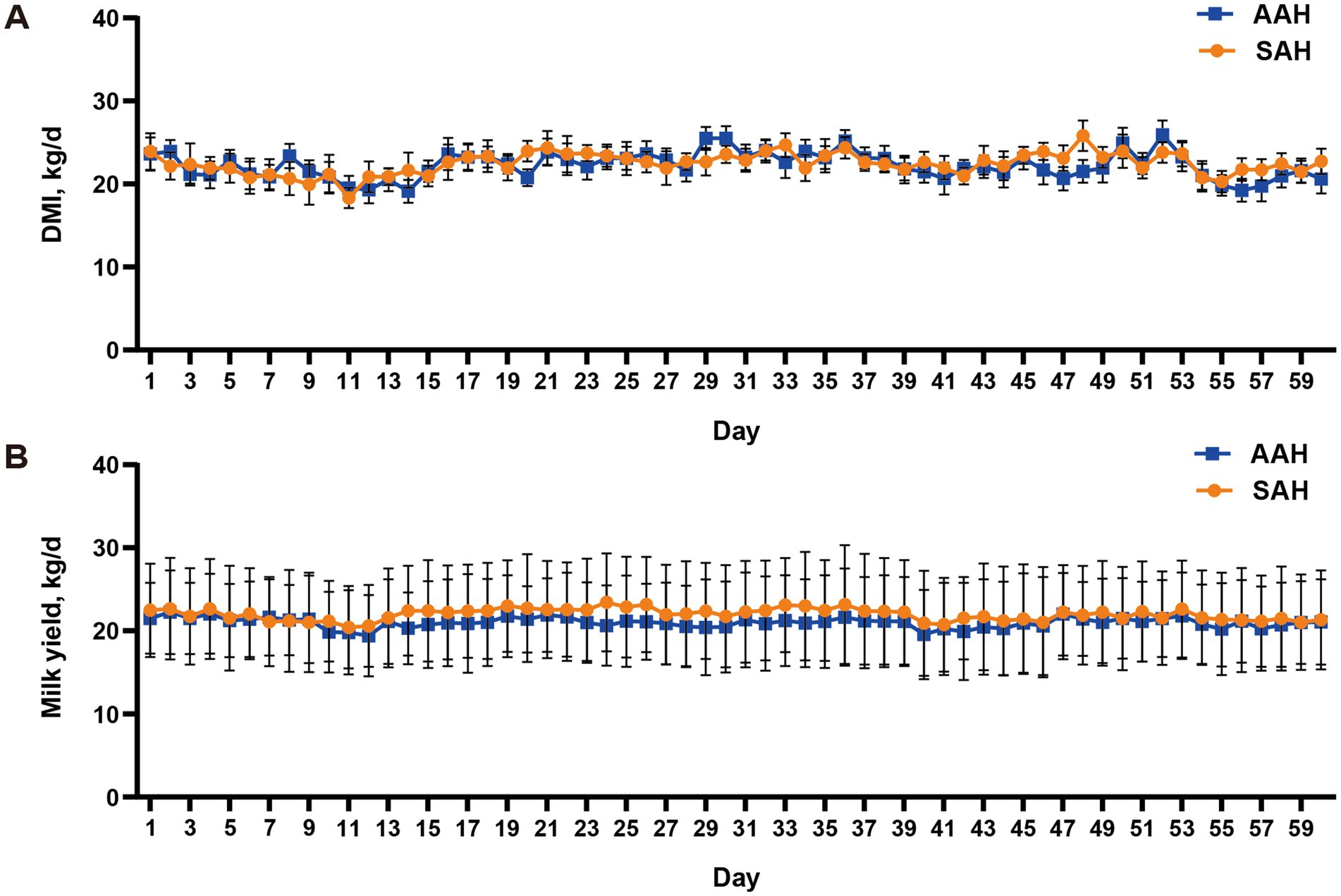
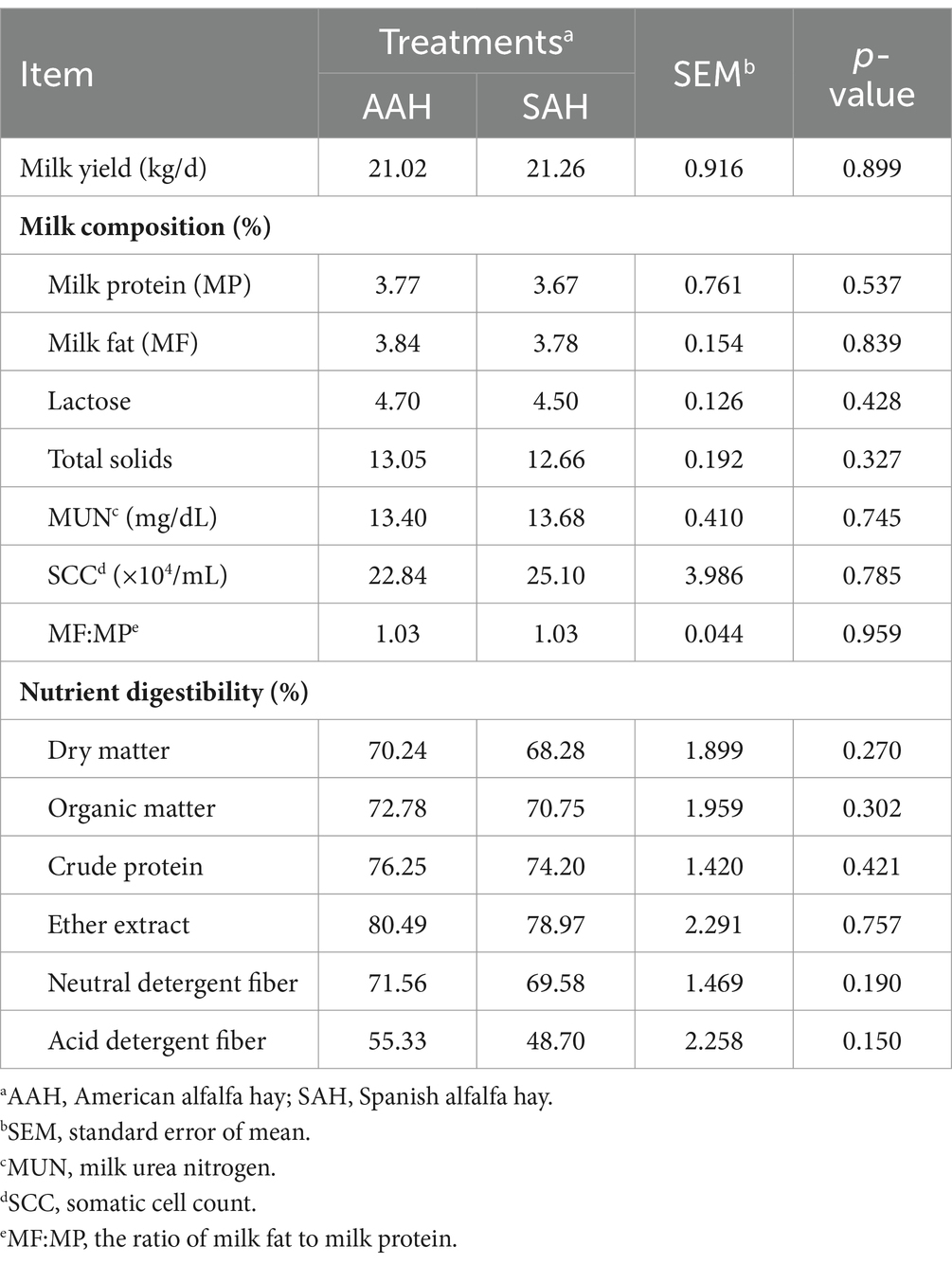
Durante el experimento, se observó que el heno de alfalfa de diferentes fuentes no tuvo efecto significativo sobre el IMD (Figura 1A) y el IM diario (Figura 1B) de los dos grupos de vacas lecheras (p > 0.05). Esta consistencia se mantuvo a lo largo de todo el período de recolección. De manera similar, en los últimos 4 días del experimento, no hubo diferencias en la IM entre las vacas utilizadas para la recolección de muestras (p > 0.05, Tabla 3). Además, el análisis no reveló diferencias en los componentes de la leche entre los dos grupos de vacas lecheras en días específicos del período de recolección, incluidos los días 14, 28, 42, 56 y los últimos 4 días (p > 0.05, Figura 2 y Tabla 3). Este hallazgo también se extendió a la digestibilidad aparente de los nutrientes, lo que indica que la fuente de heno de alfalfa tampoco tuvo un impacto significativo en este aspecto (p > 0.05, Tabla 3).
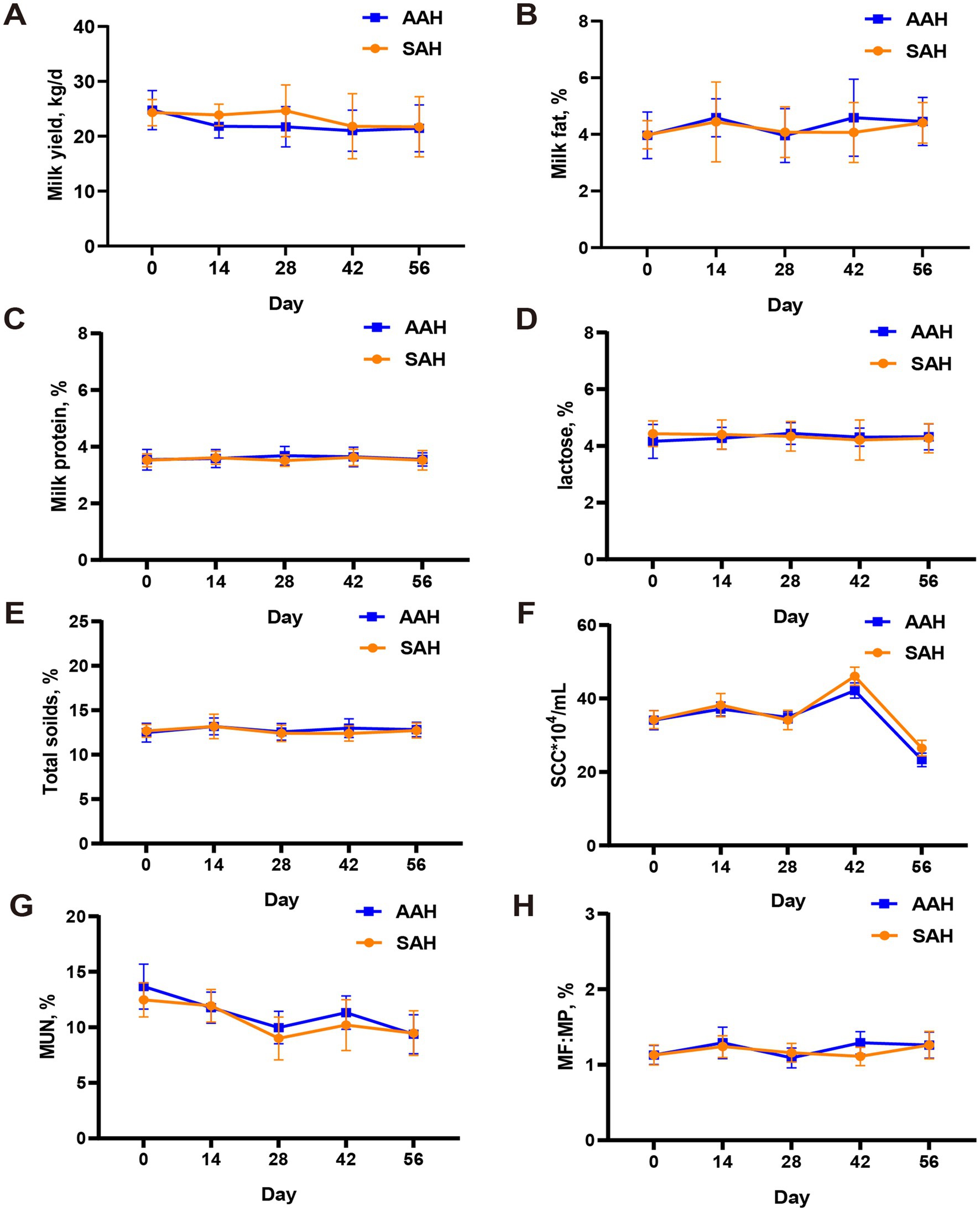
3.2 Influencia en los indicadores bioquímicos sanguíneos y en los parámetros de fermentación ruminal
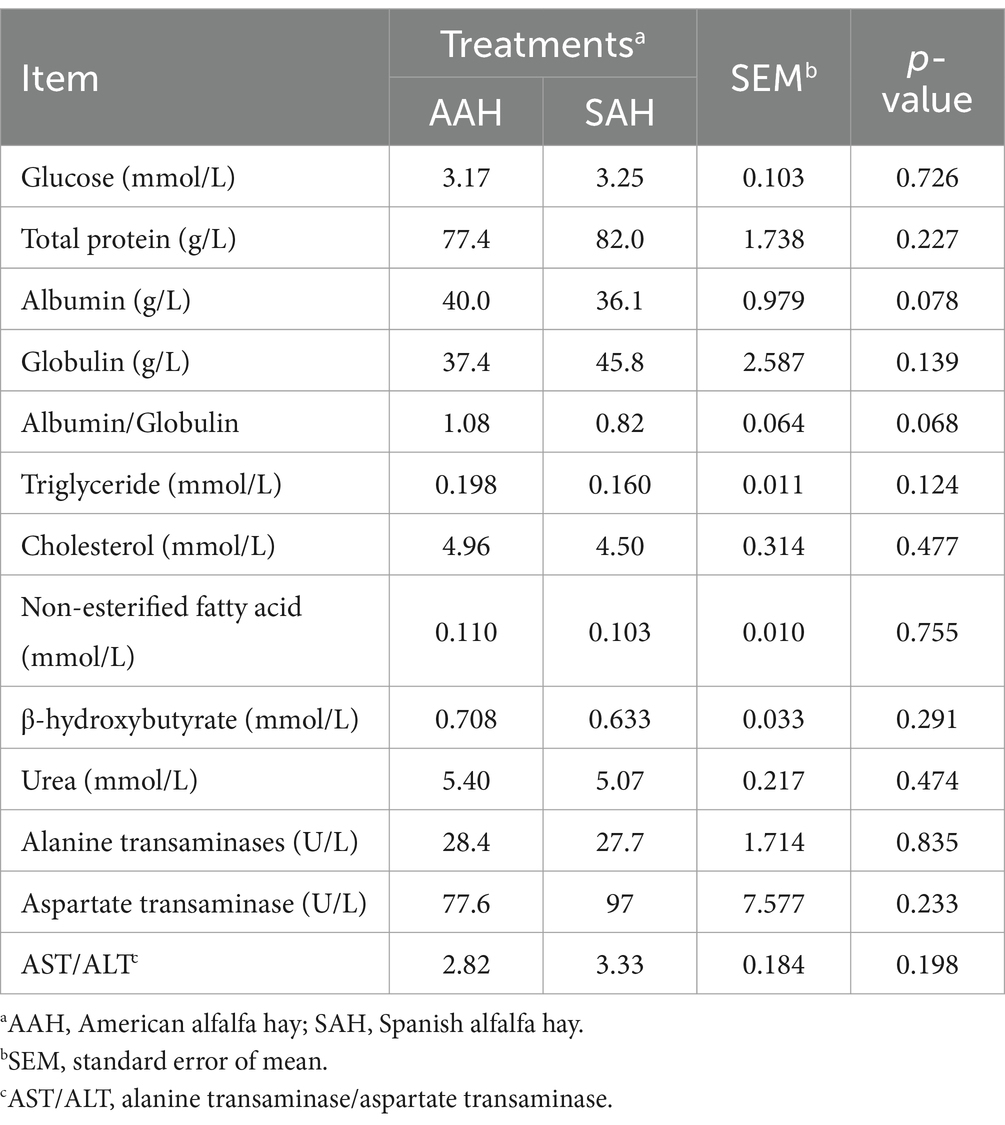
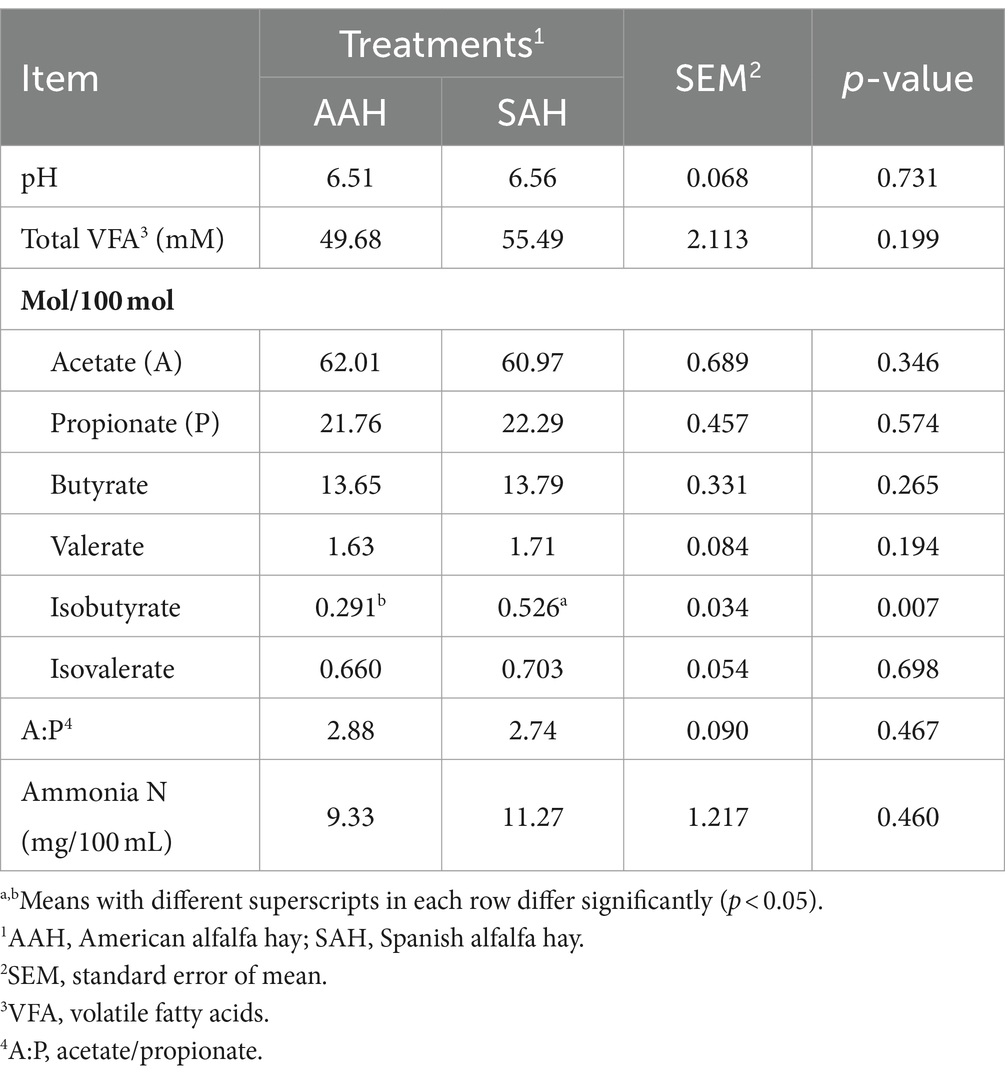
Las concentraciones séricas de glucosa, proteína total, albúmina, globulina, triglicéridos, colesterol, ácidos grasos no esterificados, β-hidroxibutirato y urea, así como las actividades de la alanina aminotransferasa y la aspartato aminotransferasa, no mostraron diferencias significativas entre los dos grupos de vacas lecheras (p > 0,05, Tabla 4). Del mismo modo, el pH ruminal se mantuvo dentro del rango normal y no varió significativamente entre los grupos (p > 0,05, Tabla 5). Los parámetros de fermentación ruminal fueron generalmente similares entre los dos grupos, con la excepción de la proporción molar de isovalerato, que fue significativamente mayor en el grupo SAH en comparación con el grupo AAH (p = 0,007). Otros parámetros de fermentación no mostraron diferencias estadísticamente significativas (p > 0,05).
3.3 Influencia en la diversidad y composición a nivel filo de la microbiota ruminal
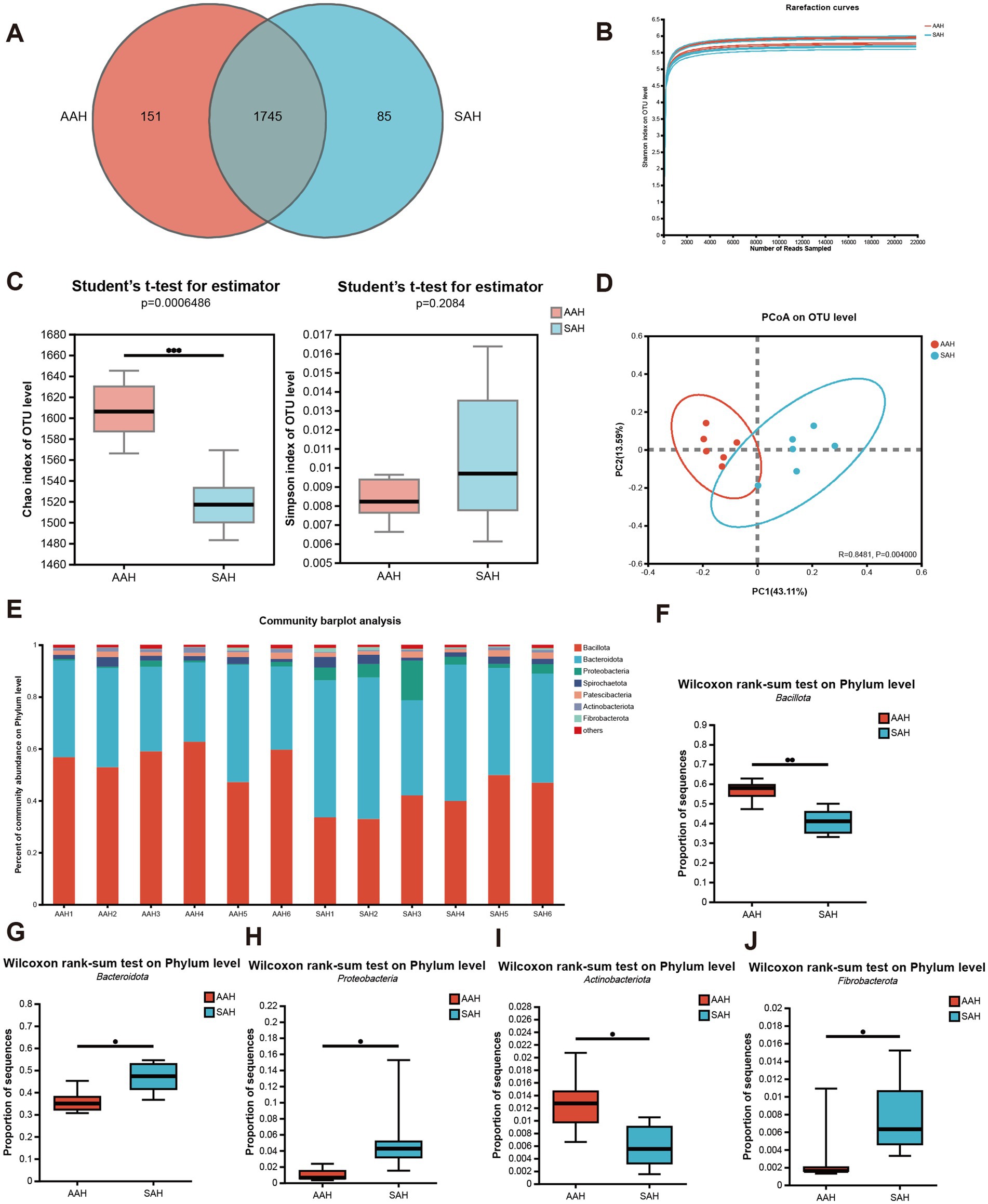
El análisis de 12 muestras de fluido ruminal dio como resultado 602.999 secuencias de alta calidad después del control de calidad, con una longitud de secuencia promedio de 417 pb y 251.558.244 bases. El análisis taxonómico con un umbral de similitud del 97% identificó 1.981 OTU en varios niveles taxonómicos: Dominio (1), Reino (1), Filo (17), Clase (34), Orden (86), Familia (149), Género (304) y Especie (589). Entre estos, los dos grupos juntos tenían 1.745 OTU, con 151 exclusivas del grupo AAH y 85 exclusivas del grupo SAH (Figura 3A). La diversidad alfa, evaluada utilizando el índice de Shannon, se acercó a una meseta, lo que indica una profundidad de secuenciación suficiente (Figura 3B). El índice Chao1 fue significativamente mayor en el grupo de AAH en comparación con el grupo de HSA (p < 0,05, Figura 3C), mientras que el índice de Simpson no mostró diferencias significativas entre los grupos (p > 0,05, Figura 3C). El gráfico de análisis de coordenadas principales reveló un agrupamiento distinto de los grupos AAH y SAH (Figura 3D). En cuanto a la composición de los filos, ambos grupos estaban dominados por Bacillota y Bacteroidota, que en conjunto constituían más del 86% de la microbiota de cada grupo (Figura 3E). Se observaron diferencias significativas entre los grupos: el grupo de SAH tuvo mayores abundancias relativas de Bacteroidota (Figura 3G), Proteobacteria (Figura 3H) y Fibrobacterota (Figura 3J) (p < 0.05), pero menores abundancias relativas de Bacillota (Figura 3F) y Actinobacteria (Figura 3I) (p > 0.05) en comparación con el grupo AAH.
3.4 Influencia en la composición y diferencia a nivel de género de la microbiota ruminal
Los cinco géneros más abundantes en el rumen del grupo AAH y del grupo SAH fueron Prevotella (12,65 y 25,76%), Succiniclasticum (6,31 y 6,18%), Rikenellaceae_RC9_gut_group (6,27 y 4,26%), NK4A214_group (6,46 y 3,30%) y Ruminococcus (5,77 y 3,29%), respectivamente (Figura 4A). Los perfiles de microbiota en el grupo AAH y en el grupo SAH mostraron distintos patrones de agrupamiento. En particular, la mayoría de los géneros dominantes en el grupo AAH pertenecían al filo Bacillota, mientras que los del grupo SAH pertenecían predominantemente al filo Bacteroidota (Figura 4B). Un análisis posterior utilizando LEfSe y un análisis discriminante lineal reveló varios géneros con abundancias significativamente diferentes entre los dos grupos (Figuras 4C,D). Específicamente, las vacas en el grupo SAH mostraron un aumento significativo en la abundancia relativa de Prevotella (Figura 4E) y Succinivibrionaceae_UCG-002 (Figura 4J) (p < 0.05), mientras que la abundancia de NK4A214_group (Figura 4F), Ruminococcus (Figura 4G), norank_f_F082 (Figura 4H) y Lachnospiraceae_NK3A20_group (Figura 4I) (p < 0,05) se redujo significativamente en comparación con el grupo AAH.
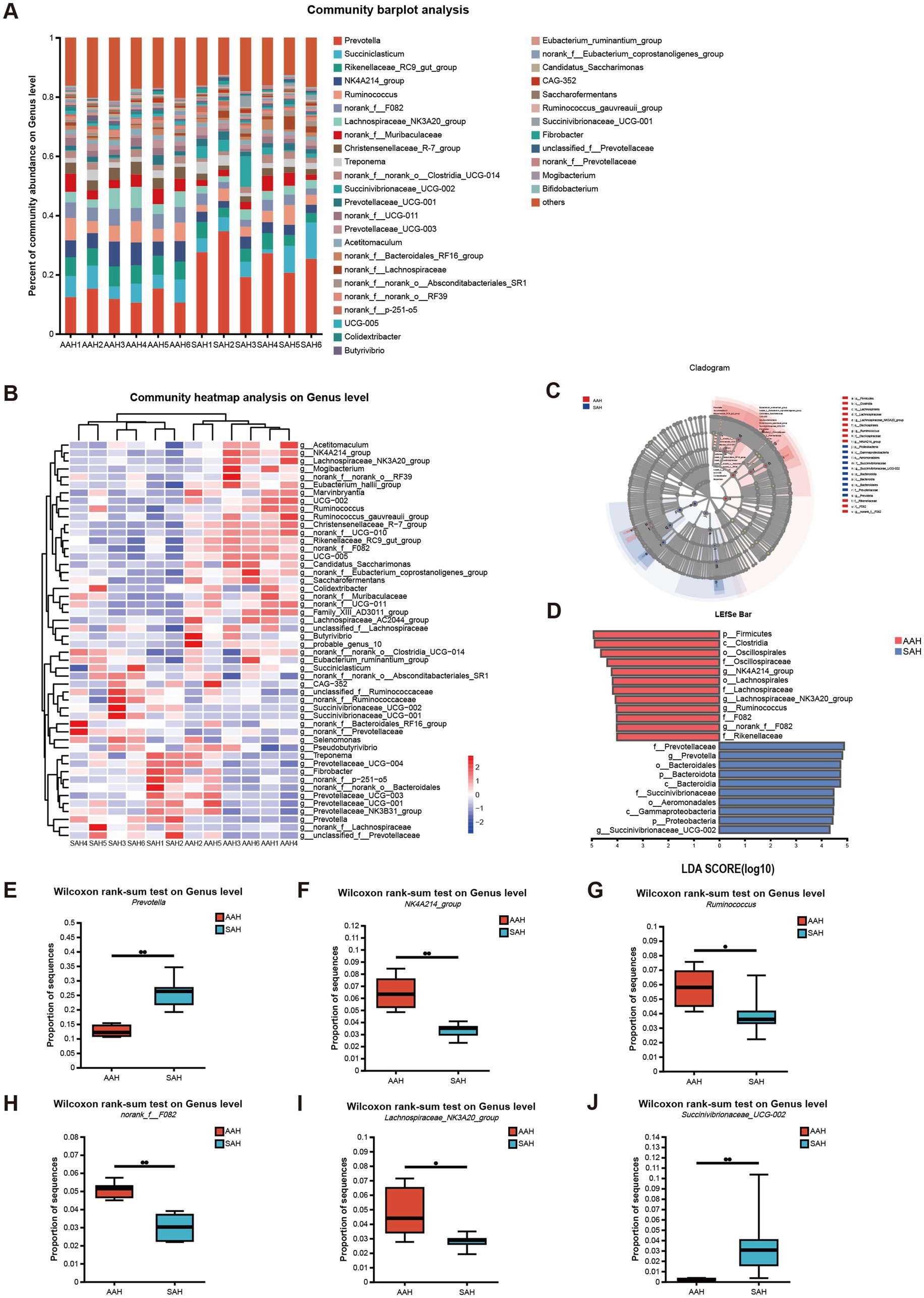
3.5 Influencia en la correlación entre los microbios ruminales y el DMI, la producción de leche y la composición de la leche
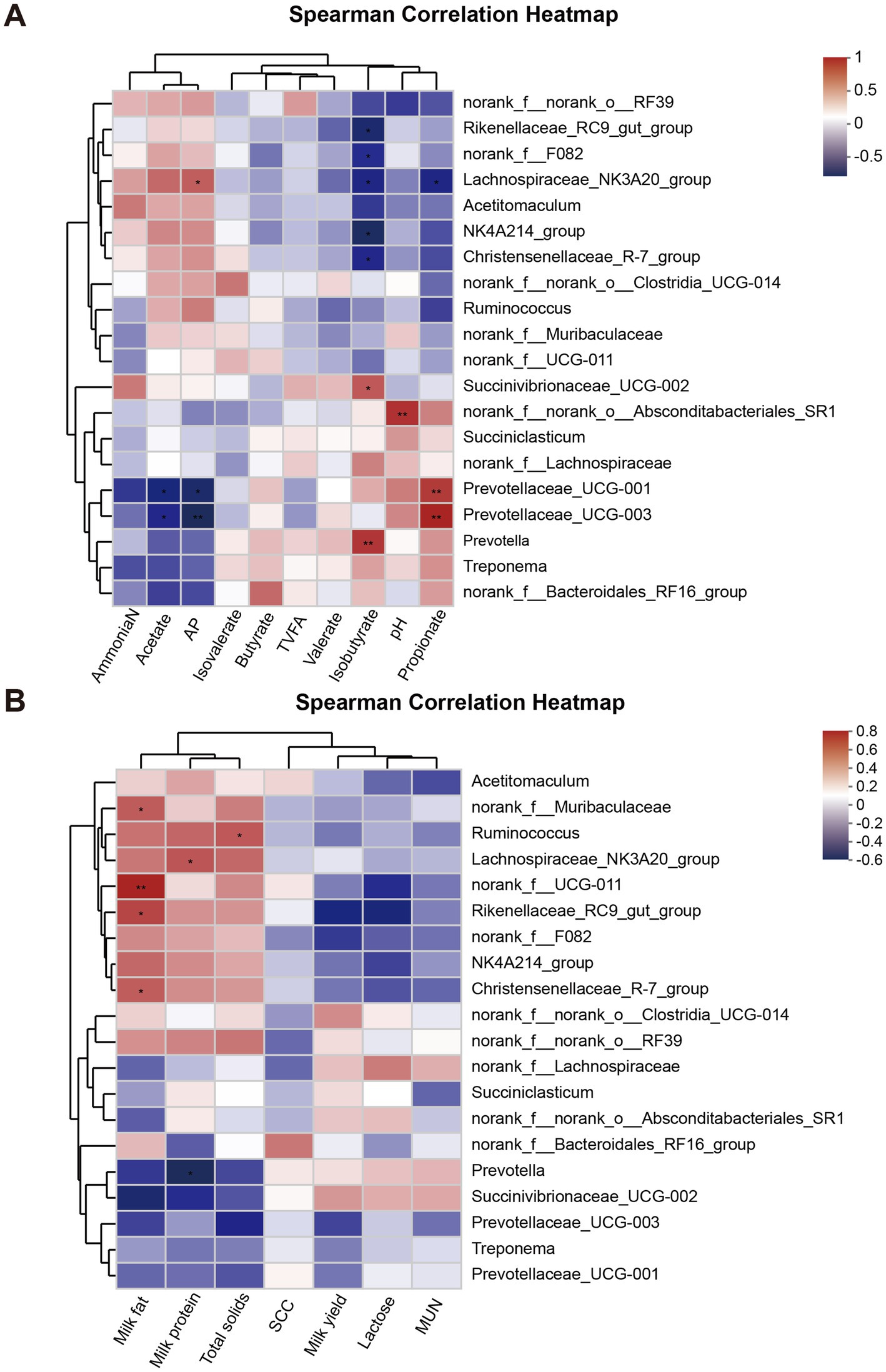
El análisis de correlación de Spearman reveló que Prevotella y Succinivibrionaceae_UCG-002 se correlacionaron significativamente positivamente con el isobutirato, mientras que Christensenellaceae_R-7_group, Lachnospiraceae_NK3A20_group, norank_f_F082, NK4A214_group y Rikenellaceae_RC9_gut_group se correlacionaron negativamente con el isobutirato (p < 0,05). Además, Lachnospiraceae_NK3A20_group mostraron una correlación positiva significativa con A/P (p < 0,05), mientras que Prevotellaceae_UCG-003 y Prevotellaceae_UCG-001 se correlacionaron positivamente con el propionato (p < 0,05) pero negativamente con A/P (p < 0,05, Figura 5A). Además, el análisis indicó que la grasa láctea se asoció significativamente positivamente con norank_f_UCG-011, Christensenellaceae_R-7_group, norank_f_Muribaculaceae y Rikenellaceae_RC9_gut_group (p < 0,05). En contraste, Prevotella mostró una correlación negativa con la proteína de la leche (p < 0.05), mientras que Lachnospiraceae_NK3A20_group mostró una correlación positiva con la proteína de la leche (p < 0.05). Se encontró que Ruminococcus tiene una correlación positiva significativa con los sólidos totales (p < 0.05). Sin embargo, no se observó correlación significativa entre la IM y los microorganismos ruminales (p > 0,05, Figura 5B).
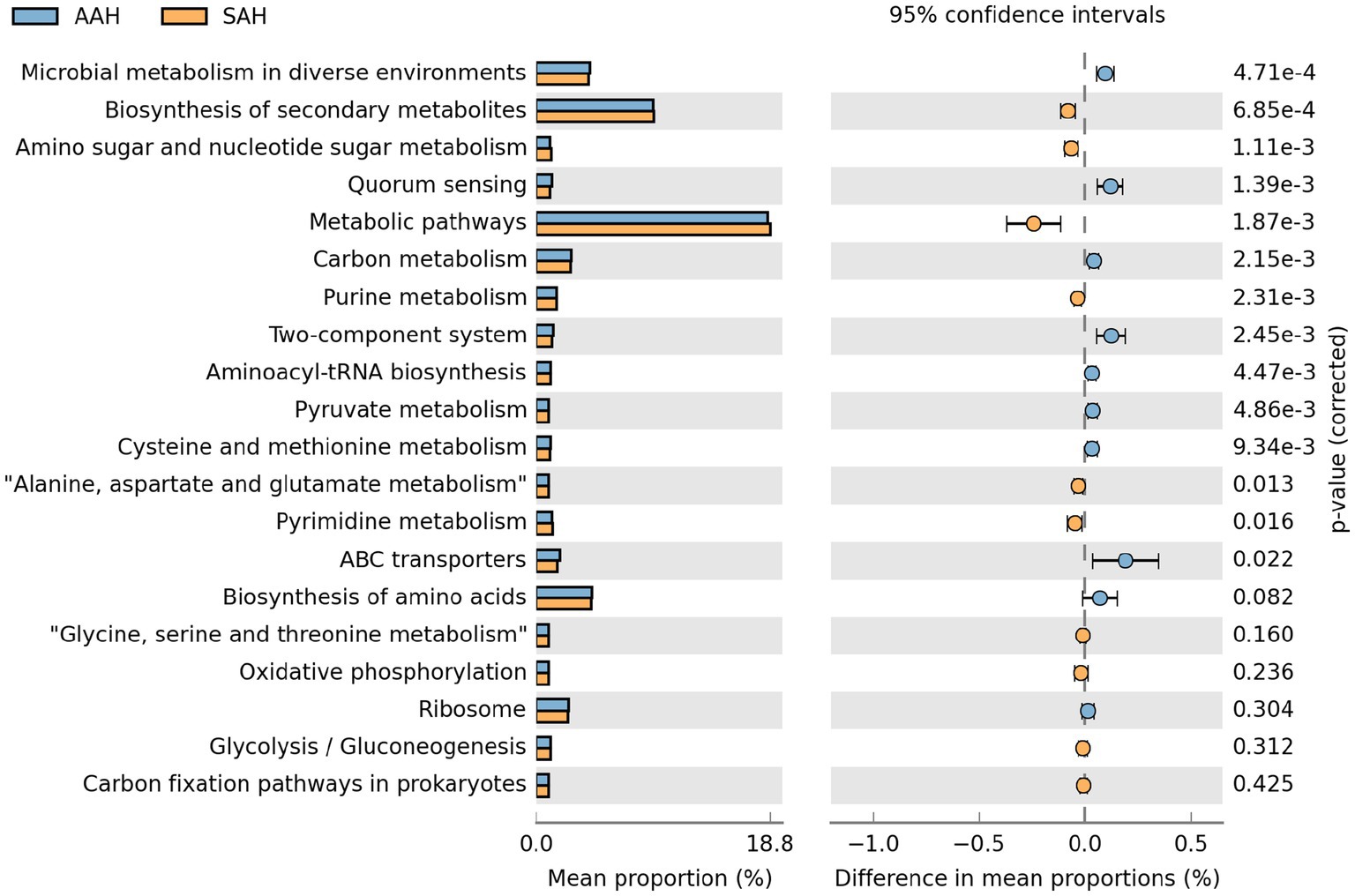
3.6 Predicciones de las funciones microbianas ruminales
A partir de la base de datos KEGG, se obtuvieron predicciones funcionales de los microorganismos ruminales utilizando PICRUSt y se analizaron con el software STAMP. Se identificaron las 20 principales vías metabólicas con mayor abundancia funcional para la comparación entre los dos grupos (Figura 6). El análisis reveló que 7 vías metabólicas fueron significativamente más abundantes en el grupo de HSA en comparación con el grupo de EHA. Estas vías incluían la biosíntesis de metabolitos secundarios, el metabolismo de aminoácidos y nucleótidos, las vías metabólicas generales, el metabolismo de las purinas, el metabolismo de la alanina, el aspartato y el glutamato, y el metabolismo del piruvato. Por el contrario, ocho vías metabólicas se redujeron notablemente en el grupo de HSA en relación con el grupo de EHA. Estas vías incluían el metabolismo microbiano en diversos entornos, la detección de quórum, el metabolismo del carbono, los sistemas de dos componentes, la biosíntesis de aminoacil-ARNt, el metabolismo de la pirimidina, el metabolismo de la cisteína y la metionina, y los transportadores ABC.
4 Discusión
El heno de alfalfa importado de Estados Unidos es ampliamente utilizado en la industria láctea china. Recientemente, la HSA se ha convertido en una importante fuente alternativa para estabilizar el mercado de suministro de heno de alfalfa, especialmente a raíz de la pandemia de COVID-19. A pesar de su utilidad, existen preocupaciones sobre los posibles impactos de la HSA en la comercialización y la calidad de la leche, que son factores cruciales que influyen en los ingresos de los productores de leche. Por lo tanto, se necesita más investigación para evaluar sus efectos dietéticos. Este estudio tuvo como objetivo comparar los efectos alimentarios de dos tipos de heno de alfalfa importado: SAH y AAH, al proporcionar ambos henos a vacas en mitad de lactancia con una inclusión de materia seca en la dieta de 12.88% durante un período de 60 días. Cada heno fue diseñado para proporcionar el mismo nivel nutricional para garantizar una comparación justa.
Después de 60 días de monitoreo continuo en 360 vacas lecheras, el MY de las vacas AAH y SAH cambió de 21,52 kg/d y 22,48 kg/d a 21,00 kg/d y 21,98 kg/d, respectivamente, lo que estuvo en línea con las fluctuaciones esperadas de la MY en vacas de mitad de lactancia (23). Del mismo modo, no hubo fluctuaciones significativas en el DMI entre los dos grupos, ya que las vacas AAH consumieron aproximadamente 22,24 kg/día y las vacas SAH consumieron 22,47 kg/día. Además, se recogieron muestras de leche de 12 vacas representativas de cada grupo para evaluar la composición de la leche. El análisis no mostró diferencias significativas en el porcentaje de grasa láctea, el porcentaje de proteína láctea o la composición general de la leche entre los grupos de SAH y AAH. Estos resultados indican que los efectos de la HSA sobre la IM y la composición de la leche son comparables a los de la HA. Esta conclusión está respaldada por la falta de diferencias significativas en la digestibilidad de los nutrientes entre los dos grupos.
Para explorar más a fondo los efectos alimentarios de AAH y SAH, en este estudio se detectaron los indicadores representativos que reflejan el metabolismo de energía, proteínas y lípidos en el suero, que no se vieron afectados por las diferentes fuentes de heno de alfalfa. Además, no hubo diferencias significativas en la actividad de la alanina aminotransferasa y la aspartato aminotransferasa, que son indicadores sensibles del metabolismo hepático y la salud cardíaca en los animales. Investigaciones previas han reportado concentraciones séricas de glucosa y urea en vacas a mitad de lactancia de alrededor de 3.2 y 4.6 mmol/L, respectivamente (23). Los resultados de este estudio se alinean con estos hallazgos, a pesar de las variaciones que pueden ser influenciadas por la dieta animal (24). Estos resultados sugieren que las diferentes fuentes de heno de alfalfa no afectan la condición fisiológica normal de las vacas lecheras durante la mitad de la lactancia, lo que respalda aún más la similitud en los efectos de la alimentación entre la AAH y la SAH.
Para comparar aún más los efectos de SAH y AAH en vacas lecheras, se determinaron los parámetros de fermentación ruminal en este experimento. El rumen, un órgano digestivo especializado en los rumiantes, desempeña un papel fundamental en el proceso de digestión debido a su gran población de microorganismos (25). En el rumen, los carbohidratos se degradan para producir AGV, como el ácido propiónico, el acetato y el ácido butírico, que son las principales sustancias productivas y constituyen el 70-80% de la energía total (26). En este estudio, los valores de pH ruminal para ambos grupos de vacas oscilaron entre 6,51 y 6,56, lo que favorece el crecimiento de bacterias fibrolíticas y la digestión efectiva de la fibra (27). Los resultados mostraron que no hubo diferencias significativas en el pH o en las concentraciones totales de AGV entre los dos grupos. La estabilidad de la microecología ruminal en rumiantes adultos puede ser la razón de este hallazgo (28, 29). Además, se encontró que la proporción de ácido acético a ácido propiónico, un indicador importante de la eficiencia de la fermentación ruminal (30), fue similar entre los dos grupos de vacas lecheras en el estudio actual, lo que indica que la fermentación ruminal no se vio afectada por ninguno de los tipos de heno de alfalfa. Además, el NH3-N ruminal, un marcador de la degradación de proteínas y la síntesis de proteínas microbianas (31), no mostró diferencias significativas entre los grupos. Este resultado se alinea con la digestibilidad de la proteína bruta. En resumen, la HSA no causó cambios en los parámetros de fermentación ruminal.
Para demostrar la abundancia de especies microbianas en las muestras de fluido ruminal, se construyó una curva de rarefacción entre la cantidad de secuencias obtenidas a través de muestreo aleatorio y los índices de diversidad de Shannon. Una curva plana indicaba que la profundidad de secuenciación era suficiente y que había capturado la mayoría de las OTU. Además, evaluamos la diversidad alfa para determinar si las diferentes fuentes de heno de alfalfa influían en la diversidad microbiana en el rumen. El índice Chao1, que refleja el número total de especies (32), mostró diferencias significativas entre los dos grupos. Las comunidades microbianas del rumen en los grupos de AAH y SAH fueron significativamente diferentes según el análisis de β-diversidad. Estos hallazgos indican que la fuente de heno de alfalfa afectó la abundancia, composición y estructura de la comunidad microbiana del rumen.
En este estudio, la secuenciación del ARNr 16S reveló que Bacteroidota y Bacillota tenían el mayor porcentaje de bacterias en la flora ruminal a nivel taxonómico de filos (33, 34). Los bacteroidota participan principalmente en la descomposición de materiales orgánicos macromoleculares intrincados, como la conversión de carbohidratos en monosacáridos. Mientras tanto, Bacillota produce enzimas extracelulares, incluidas proteasas, lipasas y celulasas, que ayudan a hidrolizar proteínas, lípidos, aminoácidos y hemicelulosa (35). A nivel de filo, Bacillota y Bacteroidota fueron los grupos bacterianos dominantes en el rumen en este estudio. En concreto, la abundancia relativa de Bacillota fue del 56,35 y 40,88% en los grupos AAH y SAH, respectivamente, mientras que Bacteroidota constituyó el 35,99 y el 46,64% en estos grupos. Estos hallazgos están en línea con los hallazgos de estudios previos (16).
Los estudios microecológicos ruminales han propuesto la existencia de un microbioma ruminal central y han informado de cambios significativos en la abundancia de géneros bacterianos centrales entre los animales (36). Debido a las diferencias en la dieta, los días en la leche, la paridad y el tamaño de la muestra, los microbios centrales del rumen pueden diferir entre los estudios en diversos grados (37). Prevotella y Succinivibri-onaceae_UCG-002, que son más abundantes en el grupo SAH a nivel de género en el rumen, pueden ser las bacterias diferenciales centrales. La Prevotella, perteneciente a la familia Bacteroidota, puede descomponer las proteínas vegetales, los péptidos, la hemicelulosa y la pectina en ácido acético, ácido succínico y una pequeña cantidad de isobutirato, que son utilizados directamente por las vacas lecheras (16). Según Calabrò et al. (38), el mayor contenido de Provetella en el grupo SAH podría deberse al mayor contenido de hemicelulosa en este estudio. Succinivibrionaceae_UCG-002, perteneciente a la familia Bacillota, es una bacteria típica que degrada la fibra y la celobiosa y la celobiosa en ácido succínico, ácido acético y dióxido de carbono (39). El análisis de correlación reveló una relación positiva significativa entre la abundancia de Prevotella y Succinivibrionaceae_UCG-002 y la proporción molar de isobutirato en el rumen. El grupo SAH exhibió una mayor proporción molar de isobutirato en comparación con el grupo AAH. Además, NK4A214_group, Ruminococcus, norank_f_F082 y Lachnospiraceae_NK3A20_group fueron más prevalentes en el grupo de AAH y podrían representar las bacterias diferenciales centrales en este estudio. Ruminococcus y NK4A214_group, pertenecientes a la familia Bacillota, son bacterias beneficiosas en el rumen, que degradan eficientemente el almidón y la fibra, respectivamente, y producen AGV para proporcionar energía al animal (40). Norank_f_F082 y Lachnospiraceae_NK3A20_group son de manera similar los principales componentes de la microbiota ruminal en los rumiantes y están estrechamente relacionados con la producción de AGV (41, 42). En este estudio, NK4A214_group, Ruminococcus, norank_f_F082 y Lachnospiraceae_NK3A20_group se correlacionaron significativamente negativamente con la proporción molar de isobutirato. Sin embargo, estos microorganismos diferenciales centrales no se correlacionaron significativamente con los AGV totales, las proporciones molares de acetato y ácido propiónico, y la MY. De hecho, a excepción de la proporción molar de isobutirato, no hubo otras diferencias significativas entre los dos grupos en los parámetros de fermentación ruminal. Esto indicó que los cambios en estos microorganismos centrales no alteraron los patrones de fermentación ruminal, que es una de las razones por las que las diferentes fuentes de heno de alfalfa no causaron cambios en el rendimiento de las vacas. Los estudios futuros podrían explorar la combinación de heno de alfalfa de diversas fuentes con otros ingredientes de los piensos para mejorar la eficiencia alterando la composición microbiana del rumen y mejorando la eficiencia de la alimentación de las vacas lecheras, que es un enfoque de la alimentación refinada.
Investigamos el impacto de diferentes fuentes de heno de alfalfa en las vías metabólicas microbianas ruminales en vacas lecheras. Utilizando el software STAMP para analizar las funciones microbianas, encontramos diferencias notables entre los dos grupos. La alimentación con SAH aumentó significativamente las funciones relacionadas con la biosíntesis de metabolitos secundarios, el metabolismo de aminoácidos y azúcares nucleótidos, las vías metabólicas, el metabolismo de las purinas, el metabolismo de la alanina, el aspartato y el glutamato, así como el metabolismo del piruvato. Por el contrario, la alimentación con AAH condujo a una regulación positiva significativa del metabolismo microbiano en diversos entornos, detección de quórum, metabolismo del carbono, sistemas de dos componentes, biosíntesis de aminoacil-ARNt, metabolismo de pirimidina, metabolismo de cisteína y metionina y transportadores ABC. A pesar de estas diferencias en la composición microbiana y las funciones metabólicas, no hubo cambios en la MY, el DMI, los parámetros bioquímicos de la sangre o los patrones de fermentación ruminal entre los dos grupos. Se necesita más investigación para comprender y verificar las razones detrás de estos hallazgos.
5 Conclusión
Encontramos que la HSA y la AAH con niveles nutricionales similares tuvieron los mismos efectos alimenticios, como lo demuestra que no hubo diferencias significativas en la MY, la composición de la leche, los parámetros bioquímicos de la sangre o los parámetros de fermentación ruminal entre los dos grupos de vacas lecheras. Sin embargo, las abundancias relativas de Prevotella, Succinivibrionaceae_UCG-002, NK4A214_group, Ruminococcus, norank_f_F082 y Lachnospiraceae_NK3A20_group se modificaron significativamente entre los dos grupos. A pesar de estos cambios, no se encontró una correlación significativa entre estas variaciones microbianas y la producción de leche. Estos hallazgos proporcionaron referencia para la aplicación de heno de alfalfa de diferentes fuentes, la mejora del beneficio económico de la granja lechera y la expansión del mercado de suministro de alfalfa.
Declaración de disponibilidad de datos
Los conjuntos de datos presentados en este estudio se pueden encontrar en repositorios en línea. Los nombres de los repositorios y los números de acceso se pueden encontrar en el artículo/material complementario.
Declaración ética
El estudio en animales fue aprobado por el Comité Institucional de Cuidado y Uso de Animales (IACUC) de la Universidad de Agricultura de Henan (Zhengzhou, China) (Número de aprobación: HNND2021062812). El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
SL: Investigación, Metodología, Validación, Redacción – borrador original. NS: Investigación, Metodología, Validación, Redacción – borrador original. YZ: Software, Escritura – revisión y edición. MA: Escritura – revisión y edición. JN: Software, Redacción – revisión y edición. ZG: Análisis formal, Redacción – revisión y edición. BL: Análisis formal, Redacción – revisión y edición. SM: Curación de datos, redacción, revisión y edición. YC: Curación de datos, redacción, revisión y edición. DL: Curación de datos, redacción, revisión y edición. YS: Conceptualización, Obtención de fondos, Administración de proyectos, Recursos, Redacción – revisión y edición.
Financiación
Los autores declaran que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio contó con el apoyo del fondo asignado al Sistema de Investigación de Forrajes y Pastos de China (CARS-34).
Reconocimientos
Los autores desean expresar su gratitud a la granja de cría de animales Xincai Ruiya por proporcionar los animales de experimentación y las instalaciones, y al personal por su dedicado cuidado de los animales.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Notas
1. ^ https://cloud.majorbio.com/page/tools/
2. ^ http://www.genome.jp/kegg/
Referencias
1. Wang, B, Tu, Y, Jiang, LS, y Liu, JX. Efecto de la dieta de paja de cereales y heno de alfalfa sobre el perfil de aminoácidos de la digesta gastrointestinal en vacas lecheras lactantes. J Anim Physiol Anim Nutr. (2018) 102:421–8. doi: 10.1111/jpn.12749
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Woll, FW. Alfalfa como único alimento para el ganado lechero. J Dairy Sci. (1918) 1:447–61. DOI: 10.3168/JDS. S0022-0302(18)94391-2
3. Wang, Q, y Zou, Y. El mercado y las importaciones de alfalfa de China: desarrollo, tendencias e impactos potenciales de la disputa comercial y las represalias entre EE. UU. y China. J Integr Agric. (2020) 19:1149–58. doi: 10.1016/s2095-3119(19)62832-7
4. Wang, X, Yan, X, Mi, F y Li, H. Análisis de correlación de variedades de alfalfa basado en el rendimiento de la producción, las tasas de supervivencia invernal y las latencias otoñales. Legum Res. (2021) 44:15–20. DOI: 10.18805/LR-551
5. Xu, W, Shi, S, Qi, J, Squires, VR, Hua, L, Zhang, W, et al. Efecto de diferentes métodos de procesamiento en la calidad del heno de alfalfa. J Anim Vet Adv. (2013) 12:689–93. doi: 10.3923/javaa.2013.689.693
6. Vagnoni, DB, y Broderick, GA. Efectos de la suplementación de energía o proteína no degradada ruminalmente a vacas lactantes alimentadas con heno de alfalfa o ensilaje. J Dairy Sci. (1997) 80:1703–12. DOI: 10.3168/JDS. S0022-0302(97)76102-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Christensen, RG, Yang, SY, Eun, JS, Young, AJ, Hall, JO y MacAdam, JW. Efectos de la alimentación con heno de trébol de pata de pájaro en la digestión de fibra con detergente neutro, la eficiencia de la utilización del nitrógeno y el rendimiento de la lactancia de las vacas lecheras. J Dairy Sci. (2015) 98:7982–92. doi: 10.3168/jds.2015-9348
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Dai, Q, Hou, Z, Gao, S, Li, Z, Wei, Z y Wu, D. La sustitución de ramio forrajero fresco por heno de alfalfa en las dietas afecta el rendimiento de la producción, la composición de la leche y los parámetros séricos de las vacas lecheras. Trop Anim Salud Prod. (2019) 51:469–72. doi: 10.1007/s11250-018-1692-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Obeidat, BS, Subih, HS y Taylor, JB. La alimentación con heno de alfalfa mejora el rendimiento de lactancia de las ovejas Awassi y el rendimiento de los corderos en crecimiento cuando se utiliza como fuente de forraje en comparación con la paja de trigo. J Anim Sci. (2018) 96:32–3. doi: 10.1093/jas/sky027.061
10. Cammack, KM, Austin, KJ, Lamberson, WR, Conant, GC y Cunningham, HC. Simposio de nutrición ruminnat: pequeño pero poderoso: el papel de los microbios ruminales en la producción ganadera. J Anim Sci. (2018) 96:752–70. doi: 10.1093/jas/skx053
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Cavaliere, G, Trinchese, G, Musco, N, Infascelli, F, De Filippo, C, Mastellone, V, et al. La leche de vacas alimentadas con una dieta alta en la proporción de forraje: concentrado mejora el estado inflamatorio, el estrés oxidativo y la función mitocondrial en ratas. J Dairy Sci. (2018) 101:1843–51. doi: 10.3168/jds.2017-13550
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Fustini, M, Palmonari, A, Canestrari, G, Negri, N y Formigoni, A. Efecto de la digestibilidad del heno de alfalfa y la tasa de inclusión en las dietas de vacas lecheras lactantes sobre la ingesta de alimento, la digestibilidad, el pH ruminal y el rendimiento productivo. Ital J Anim Sci. (2015) 18:101066. doi: 10.1016/j.animal.2023.101066
14. Van Soest, PJ, Robertson, JB y Lewis, BA. Métodos para la fibra dietética, la fibra detergente neutro y los polisacáridos sin almidón en relación con la nutrición animal. J Dairy Sci. (1991) 74:3583–97. DOI: 10.3168/JDS. S0022-0302(91)78551-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Mattioli, S, Dal Bosco, A, Duarte, JMM, D’Amato, R, Castellini, C, Beone, GM, et al. Uso de hojas de olivo enriquecidas con selenio en la alimentación de conejos en crecimiento: efecto sobre el estado oxidativo, el perfil mineral y la especiación del selenio de la carne de longissimus dorsi. J Trace Elem Med Biol. (2019) 51:98–105. doi: 10.1016/j.jtemb.2018.10.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Cui, Y, Liu, H, Gao, Z, Xu, J, Liu, B, Guo, M, et al. El ensilado de maíz de planta entera mejora la fermentación ruminal y el rendimiento del crecimiento del ganado vacuno al alterar la microbiota ruminal. Aplicación Microbiol Biotechnol. (2022) 106:4187–98. doi: 10.1007/s00253-022-11956-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Chen, S, Zhou, Y, Chen, Y y Gu, J. fastp: un preprocesador FASTQ todo en uno ultrarrápido. Bioinformática. (2018) 34:i884–90. doi: 10.1093/bioinformática/bty560
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Magoc, T, y Salzberg, SL. FLASH: ajuste rápido de la longitud de lecturas cortas para mejorar los ensamblajes del genoma. Bioinformática. (2011) 27:2957–63. doi: 10.1093/bioinformática/btr507
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Edgar, RC. UPARSE: secuencias OTU de alta precisión a partir de lecturas de amplicones microbianos. Métodos Nat. (2013) 10:996–8. doi: 10.1038/nmeth.2604
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Stackebrandt, E, y Goebel, BM. Nota taxonómica: un lugar para la reasociación ADN-ADN y el análisis de secuencias de ARNr 16S en la definición actual de especie en bacteriología. Int J Syst Evol Microbiol. (1994) 44:846–9. doi: 10.1099/00207713-44-4-846
21. Schloss, PD, Westcott, SL, Ryabin, T, Hall, JR, Hartmann, M, Hollister, EB, et al. Presentamos mothur: software de código abierto, independiente de la plataforma y respaldado por la comunidad para describir y comparar comunidades microbianas. Appl Environ Microbiol. (2009) 75:7537–41. doi: 10.1128/AEM.01541-09
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Langille, MG, Zaneveld, J, Caporaso, JG, McDonald, D, Knights, D, Reyes, JA, et al. Perfil funcional predictivo de comunidades microbianas utilizando secuencias de genes marcadores de ARNr 16S. Nat Biotechnol. (2013) 31:814–21. doi: 10.1038/nbt.2676
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. NRC. Requerimientos de nutrientes del ganado lechero. Washington, DC: National Academy Press (2001).
24. Musco, N, Tudisco, R, Grossi, M, Mastellone, V, Morittu, V, Pero, M, et al. Efecto de un alto forraje: relación de concentrado sobre la producción de leche, los parámetros sanguíneos y el estado oxidativo en vacas lactantes. Anim Prod Sci. (2020) 60:1531–8. DOI: 10.1071/AN18041
25. Anantasook, N, Wanapat, M, Cherdthong, A y Gunun, P. Cambios en la población microbiana en el rumen de los novillos lecheros influenciados por la planta que contiene taninos y saponinas y la proporción de forraje a concentrado. Australas Asiáticas J Anim Sci. (2013) 26:1583–91. doi: 10.5713/ajas.2013.13182
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Sommer, F, y Baeckhed, F. La microbiota intestinal, maestra del desarrollo y la fisiología del huésped. Nat Rev Microbiol. (2013) 11:227–38. doi: 10.1038/nrmicro2974
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Russell, JB y Wilson, DB. ¿Por qué las bacterias celulolíticas ruminales no pueden digerir la celulosa a pH bajo? J Dairy Sci. (1996) 79:1503–9. DOI: 10.3168/JDS. S0022-0302(96)76510-4
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Chen, XL, Wang, JK, Wu, YM y Liu, JX. Efectos de los tratamientos químicos de la paja de arroz sobre las características de fermentación ruminal, las actividades de las enzimas fibrolíticas y las poblaciones de microbios ruminales asociados a líquidos y sólidos in vitro. Anim Feed Sci Technol. (2008) 141:1–14. doi: 10.1016/j.anifeedsci.2007.04.006
29. Zhou, YW, McSweeney, CS, Wang, JK y Liu, JX. Efectos del fumarato disódico sobre la fermentación ruminal y las comunidades microbianas en ovejas alimentadas con dietas ricas en forraje. Animal. (2012) 6:815–23. doi: 10.1017/S1751731111002102
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Wei, X, Yin, Q, Zhao, H, He, J, Cao, Y y Yao, J. La suplementación con nicotinamida durante el posparto y el periparto modula la energía hepática y el metabolismo de los lípidos, el estado oxidativo y el perfil metabolómico, así como los lípidos en el tejido adiposo de las cabras. Anim Feed Sci Technol. (2021) 274:114849. doi: 10.1016/j.anifeedsci.2021.114849
31. Annison, EF, y Bryden, WL. Perspectivas sobre la nutrición y el metabolismo de los rumiantes. II. Metabolismo en tejidos de rumiantes. Nutr Res Rev. (1999) 12:147–77. doi: 10.1079/095442299108728866
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Barger, K, Langsetmo, L, Orwoll, ES, y Lustgarten, MS. Investigación del eje dieta-intestino-músculo en el estudio de fracturas osteoporóticas en hombres. J Nutr Salud Envejecimiento. (2020) 24:445–52. doi: 10.1007/s12603-020-1344-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. de Oliveira, MNV, Jewell, KA, Freitas, FS, Benjamin, LA, Tótola, MR, Borges, AC, et al. Caracterización de la microbiota a través del tracto gastrointestinal de un novillo Nelore brasileño. Microbiol veterinario. (2013) 164:307–14. doi: 10.1016/j.vetmic.2013.02.013
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Zhang, R, Ye, H, Liu, J y Mao, S. Las dietas altas en granos alteraron la fermentación ruminal y la comunidad bacteriana epitelial y resultaron en lesiones epiteliales ruminales de cabras. Aplicación Microbiol Biotechnol. (2017) 101:6981–92. doi: 10.1007/s00253-017-8427-x
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Li, X, y Yu, YH. Biodiversidad y cribado de bacterias halófilas con actividades hidrolíticas y antimicrobianas del lago salado de Yuncheng, China. Biología. (2015) 70:151–6. doi: 10.1515/biolog-2015-0033
36. Jami, E, y Mizrahi, I. Similitud de las bacterias ruminales entre vacas lactantes individuales. Anaerobio. (2012) 18:338–43. doi: 10.1016/j.anaerobe.2012.04.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Xue, M, Sun, H, Wu, X, Guan, LL y Liu, J. La evaluación de la microbiota ruminal de una gran cohorte de ganado lechero revela que los bacteriomas pan y central contribuyen a fenotipos variados. Appl Environ Microbiol. (2018) 84:E00970. doi: 10.1128/aem.00970-18
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Calabrò, S, Infascelli, F, Bovera, F, Moniello, G, y Piccolo, V. Degradabilidad in vitro de tres forrajes: cinética de fermentación y producción de gas de FDN y fracción soluble en detergente neutro de forrajes. J Sci Alimentos Agric. (2002) 82:222–9. DOI: 10.1002/jsfa.1021
39. Wang, Y, Jiang, M, Zhang, Z y Sun, H. Efectos de la sobrecarga de hierro en la digestibilidad de nutrientes, la hematobioquímica, la fermentación ruminal y las comunidades bacterianas en ovejas. J Anim Physiol Anim Nutr. (2020) 104:32–43. doi: 10.1111/jpn.13225
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Liu, C, Wu, H, Liu, S, Chai, S, Meng, Q y Zhou, Z. Alteraciones dinámicas en la comunidad de bacterias del rumen yak y las características del metaboloma en respuesta al tipo de alimento. Microbiol frontal. (2019) 10:1116. doi: 10.3389/fmicb.2019.01116
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Han, H, Zhang, L, Shang, Y, Wang, M, Phillips, CJ, Wang, Y, et al. Sustitución del ensilado de maíz y la harina de soja por ensilado de morera en la dieta de corderos Hu sobre el crecimiento, la morfología del tejido gastrointestinal, los parámetros de fermentación ruminal y la diversidad microbiana. Animales. (2022) 12:1406. doi: 10.3390/ani12111406
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Yi, S, Dai, D, Wu, H, Chai, S, Liu, S, Meng, Q, et al. La relación entre el concentrado y el forraje en la dieta afecta a la composición de la comunidad bacteriana del rumen y al metaboloma de los yaks. Tuerca delantera. (2022) 9:927206. doi: 10.3389/fnut.2022.927206
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: heno de alfalfa, rendimiento de la lactancia, fermentación ruminal, microbiota, vaca lechera
Cita: La S, Li H, Zhang Y, Abaidullah M, Niu J, Gao Z, Liu B, Ma S, Cui Y, Li D y Shi Y (2024) Diferentes fuentes de heno de alfalfa alteran la composición de la microbiota ruminal en vacas Holstein a mitad de lactancia sin afectar el rendimiento de la producción. Frente. Vet. Sci. 11:1433876. doi: 10.3389/fvets.2024.1433876
Editado por:
Shengguo Zhao, Instituto de Ciencias Animales (CAAS), China
Revisado por:
Jun Zhang, Universidad del Noroeste A&F, China
Sijia Liu, Universidad Agrícola de Anhui, China
Derechos de autor © 2024 La, Li, Zhang, Abaidullah, Niu, Gao, Liu, Ma, Cui, Li y Shi. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Atribución (CC BY).
*Correspondencia: Yinghua Shi, annysyh@henau.edu.cn
†Estos autores han contribuido por igual a este trabajo y comparten la primera autoría
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía