
Efecto de diferentes niveles de multienzimas sobre la respuesta inmune, la hematología y bioquímica de la sangre
Efecto de diferentes niveles de multienzimas sobre la respuesta inmune, la hematología y bioquímica de la sangre, el estado de los antioxidantes y la histología de los órganos de los pollos de engorde alimentados con dietas estándar y de baja densidad
 Y. A. Attia1,2*
Y. A. Attia1,2*  H. Al-Khalaifah3*
H. Al-Khalaifah3*  H. S. Abd El-Hamid4
H. S. Abd El-Hamid4  M. A. Al-Harthi1 A. A. El-shafey2
M. A. Al-Harthi1 A. A. El-shafey2- 1Departamento de Agricultura de Tierras Áridas, Facultad de Meteorología, Medio Ambiente y Agricultura de Tierras Áridas, Universidad Rey Abdulaziz, Jeddah, Arabia Saudita
- 2Departamento de Producción Animal y Avícola, Facultad de Agricultura, Universidad de Damanhur, Damanhur, Egipto
- 3Centro de Investigación sobre el Medio Ambiente y las Ciencias de la Vida, Instituto de Investigaciones Científicas de Kuwait, Ciudad de Kuwait, Kuwait
- 4Departamento de Enfermedades de las Aves de Corral y los Peces, Facultad de Medicina Veterinaria, Universidad de Damanhour, Damanhur, Egipto
Este estudio se llevó a cabo para investigar el efecto de la suplementación de tres niveles multienzimáticos (0, 0,1 y 0,2 %) con dos tipos de dieta [dieta estándar (DE) vs. dieta de baja densidad (LDD)] sobre la respuesta inmunitaria, la hematología y bioquímica de la sangre, el estado antioxidante y la histología de los órganos de los pollos de engorde durante 1 a 38 días de edad. Un total de 216 pollos de engorde de Arbor Acres de 1 día de edad sin sexar se distribuyeron aleatoriamente, en un diseño factorial (2 × 3), a seis tratamientos cada uno con seis repeticiones. Hubo seis polluelos por réplica. Los resultados mostraron que la LDD disminuyó significativamente la ganancia de peso corporal (BWG) de los pollos de engorde, pero no afectó el Índice Europeo de Eficiencia de la Producción (EPEI). La adición de multienzimas en ambos niveles (0,1 y 0,2%) aumentó significativamente el peso corporal y mejoró la EPEI, en comparación con la dieta control. La alanina aminotransferasa (ALT), la aspirada aminotransferasa (AST), el malondialdehído (MDA), los linfocitos, la prueba de transformación de linfocitos (LTT) y la actividad de los fagocitos (PA) fueron significativamente más altos para la LDD que para la SD, pero el eosinófilo fue menor. La suplementación con multienzimas disminuyó significativamente la ALT, la AST y la MDA, en comparación con el grupo de control, pero aumentó el volumen de células empaquetadas (PCV), la hemoglobina (Hgb), los linfocitos y los monocitos. Los órganos inmunitarios, como el bazo, el timo y la bolsa de Fabricio, aumentaron significativamente con la suplementación multienzimática. Se pudo concluir que la suplementación multienzimática al 0,1 o al 0,2% de SD o LDD mejoró el EPEI y el estado inmunológico de los pollos de engorde.
Introducción
El costo de los ingredientes de los piensos para aves de corral representa alrededor del 60-70% del costo total de producción y, por lo tanto, la formulación de piensos es un enfoque crítico en la industria avícola. La utilización de los piensos puede satisfacerse con la inclusión de enzimas, antimicrobianos, probióticos o productos prebióticos o naturales (1-9).
El efecto inmunomodulador de la suplementación de los piensos para aves de corral con multienzimas ha sido bien documentado en la literatura. Hosseindoust et al. (10) concluyeron que la β-mananasa tiene el potencial de mejorar la salud intestinal de los pollos de engorde alimentados con una dieta a base de maíz y harina de soja (SBM). En el mismo estudio, también se mejoró el rendimiento del crecimiento y la retención total de nutrientes en el tracto. Liu et al. (11) investigaron el efecto de multienzimas que contienen fitasa, proteasa y xilanasa a 1.000, 2.000 y 2.000 U/kg de alimento para pollos de engorde, respectivamente. Los autores demostraron que las multienzimas mejoraron significativamente la ingesta de alimento, el aumento de peso corporal, el receptor polimérico de Ig (pIgR), la secretaria de IgA (sIgA) y los recuentos ileales de lactobacilos y bifidobacterias, y redujeron significativamente las lesiones en el intestino, los anticuerpos séricos contra la toxina a, la expresión de mucina 2 y el recuento ileal de Clostridium perfringens. Sin embargo, la fuerza del efecto multienzimático depende del contenido de proteínas en la dieta. Los autores señalaron que un alto contenido de proteínas no convencionales en la dieta puede conducir a una mayor aparición de enteritis necrótica subclínica, mientras que la suplementación multienzimática puede reducir este efecto en pollos de engorde al mejorar la inmunidad intestinal (11).
La suplementación de las raciones de alimento para aves de corral con enzimas puede mejorar la utilización del alimento y eliminar el efecto negativo de los polisacáridos sin almidón (NSP) en el rendimiento de los pollos de engorde (12-14). Se afirma que la suplementación multienzimática, como la amilasa, la xilanasa y la proteasa, estimula la desintegración del almidón, las paredes celulares y las proteínas endógenas y, por lo tanto, mejora la utilización de la energía en las dietas de maíz y sorgo (15). Attia et al. (16) revelaron que la suplementación de la dieta de las aves de corral con multienzimas mejoró el costo económico de las dietas. Sin embargo, la composición y el tipo de mezcla enzimática determinan el efecto de la suplementación multienzimática en el rendimiento de la producción avícola (17-19).
Se afirma que el uso de complejos enzimáticos que contienen carbohidrasas y proteasas mejora la utilización de energía, proteínas, P y Ca por pollos de engorde, gallinas ponedoras, patos y codornices japonesas (16, 17, 20, 21). Sin embargo, todavía hay resultados contradictorios sobre el efecto beneficioso de las multienzimas en el rendimiento de las aves, y esto podría atribuirse a la composición de la dieta, la edad y el tipo de pollitos, lo que sugiere la necesidad de más investigación (17, 22).
El uso de la dieta de baja densidad (LDD) con o sin suplementación enzimática en relación con las respuestas fisiológicas e inmunológicas y la bioquímica del estado antioxidante de los pollos de engorde es un nuevo concepto en la nutrición de los pollos de engorde. La mayoría de las investigaciones previas se centraron en el rendimiento productivo y las características de la canal como criterios económicos (18, 22, 23). En la literatura, se encontró que la suplementación con enzimas, en particular las dietas bajas en energía o los alimentos que contienen alto contenido de fibra y/o factores antinutricionales, mejoran el uso de energía para el crecimiento (12-18, 21, 23-26). Por otro lado, la inmunidad y el estado antioxidante y el desarrollo de los órganos internos pueden necesitar más nutrientes que los necesarios para el crecimiento y la calidad de la canal y pueden reflejar perfectamente el estado nutricional de los animales (24, 25). Por lo tanto, este estudio se centra en la intervención nutricional entre la LDD y la suplementación enzimática desde el punto de vista de la bioquímica, las respuestas fisiológicas e inmunológicas, el estado antioxidante y la histología de los órganos.
Materiales y métodos
Pollitos, dieta y diseño experimental
El comité departamental de Producción Animal y Avícola acepta todos los procedimientos realizados en el presente estudio. Estos procedimientos sugieren una tensión mínima para los animales a fin de garantizar los derechos y el bienestar mediante la eliminación del daño o el sufrimiento de los animales de acuerdo con los decretos oficiales del Ministerio de Agricultura de Egipto sobre el bienestar animal [Decreto Nº 27 (1967) que impone el trato humanitario de los animales en general].
Se compraron un total de 216 pollos de engorde Arbor Acres de 1 día de edad a Cairo Poultry Company, anillados con alas y distribuidos de manera aleatoria, independientemente de su sexo, con igual peso corporal inicial (45,8 ± 1,42) en 36 jaulas de 6 aves por jaula. Cada tratamiento consta de seis repeticiones, con seis pollitos por repetición.
El experimento se diseñó con dos dietas [dieta estándar (DE) vs. LDD] y tres niveles multienzimáticos (sin suplementar, 0,1 y 0,2%) en un diseño factorial (2 × 3) durante 1-38 días de edad. Galzym-M son productos de Tex Biosciences, Reino Unido. Galzym es una multienzima que contiene celulasa 100.000.000 U/kg, 4-β-xilanasa 1.500.000 U/kg, lipasa 6.500 U/g, α-amilasa 250.000 U/kg, proteasa 40.000 U/kg, pectinasa 30.000 U/kg y benzoato de sodio (conservante) 50 mg/kg. La composición química de las dietas basales experimentales administradas durante las etapas experimentales se muestra en la Tabla 1 (27). La dieta de inicio se alimentó desde la eclosión hasta los 7 días de edad (1 semana); Se administró una dieta de crecimiento de 8 a 21 días de edad (2-3 semanas); y se alimentó con una dieta de engorde de 22 a 38 días de edad.®
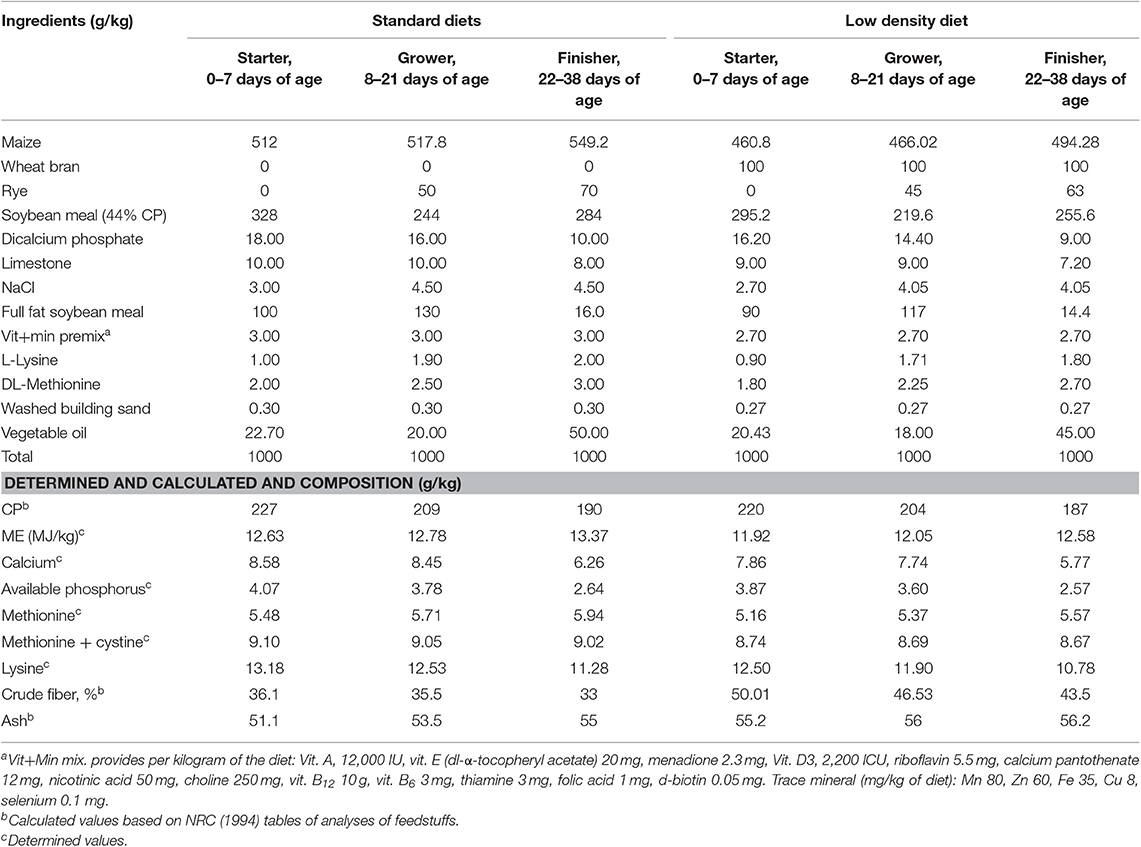 TABLA 1. Ingredientes y composición química de las dietas basales experimentales alimentadas durante las etapas experimentales.
TABLA 1. Ingredientes y composición química de las dietas basales experimentales alimentadas durante las etapas experimentales.
Alojamiento de animales
Los pollitos se distribuyeron en jaulas en batería en una habitación semiabierta. Las raciones de alimento se suministraban ad libitum y se accedía libremente al agua. Se siguió un programa de luz comercial hasta el 7º día de luz de 23 h, seguido de luz de 20 h desde el 8º día a través de la fase de cría hasta el día 38 de edad. Las temperaturas mínimas y máximas exteriores y la humedad relativa durante la fase de cría se promediaron en 22.9 y 28.3°C y 60.1 y 62.2%, respectivamente. La temperatura interior fue de 33,2, 28,3 y 26,7 °C durante los días 1-7, 8-18 y 15-20 de edad, respectivamente. El programa de vacunación consiste en el clon 30 el día 8, la inyección doble de la gripe H5 N2 muerta y el virus de la enfermedad de Newcastle (VEN) debajo de la piel del cuello el día 10, y el clon 30 y Gumboro el día 21.
Recogida de datos
El BWG (g/ave) se calculó con base en el peso de los pollos de engorde (g) a los 1, 21 y 38 días de edad. Se calculó la tasa de conversión alimenticia (FCR) (g de alimento/g de ganancia) en función de la ingesta de alimento (g/ave) y se calculó la tasa de supervivencia (SR, 100 – tasa de mortalidad) durante 1-21, 22-38 y 1-38 días de edad. El Índice Europeo de Eficiencia de la Producción (EPEI) se calculó utilizando la ecuación de la Guía de Manejo de Pollos de Engorde Hubbard (28) de la siguiente manera:
donde PP = Período de producción (días).
Recolección de muestras de sangre
Las muestras de sangre (n = 6) se extrajeron aleatoriamente de cada grupo experimental el día 8 después de la vacunación (29 días de edad), así como el día 0 (justo antes de la vacunación). Las muestras de sangre se extrajeron de la vena branquial mediante el uso de un tubo vacutainer utilizando tubos heparinizados y no heparinizados. El suero y el plasma se separaron por centrifugación a 1.500 × g durante 15 min. Las muestras de sangre utilizadas para los análisis se recogieron antes del inicio de todas las vacunaciones (día 0) y después del final de la última vacunación (día 29 de edad).
Rasgos hematológicos
El volumen de células empaquetadas (% de PCV) se determinó centrifugando las muestras de sangre durante 20 min a 2.000 × g utilizando tubos de hematocrito Wintrobe (Jiangdu Sunflower Glass Instrument Factory, Jiangdu, China). Se utilizó el método de la cianometahemoglobina para determinar la concentración de hemoglobina (Hgb) (29). El recuento de glóbulos rojos se determinó mediante el método de (30) y los índices de glóbulos rojos se calcularon según lo descrito por Jain (31) de acuerdo con las siguientes ecuaciones:
Volumen corpuscular medio (VCM) (μm3) = PCV × 10/RBC’s
Hemoglobina corpuscular media (MCH) (pg) = Hgb × 10/RBC’s
Concentración media de hemoglobina corpuscular (MCHC) (g/dl) = Hgb × 100/PCV.
Índices inmunológicos
La actividad fagocítica y el índice se determinaron de acuerdo con Kawahara et al. (32). Actividad fagocítica (PA) = porcentaje de células fagocíticas que contienen células de levadura. Índice fagocítico (IP) = número de células de levadura fagocitadas/número de células fagocíticas. Esta prueba indica la actividad de los glóbulos blancos que fagocitan las partículas extrañas dañinas, las bacterias y las células muertas o moribundas. Es un índice inmunológico importante, que indica fuertemente y refleja el estado inmunológico. Estos fagocitos son principalmente monocitos y macrófagos, granulocitos y células dendríticas. El índice fagocítico indica la fuerza de la célula fagocítica, representada por el número de bacterias ingeridas. La actividad fagocítica se considera la primera línea de defensa contra antígenos y agentes patógenos (33, 34).
La producción total de anticuerpos específicos para la vacuna contra el VEN se determinó en suero utilizando kits comerciales de ELISA (35). La respuesta de anticuerpos se determinó mediante la prueba de inhibición de la hemaglutinación (IH) según King y Seal (36). El ensayo está diseñado para medir los anticuerpos de la EII unidos a placas recubiertas de antígeno de influenza (37). Se utilizó el método de Takatsy y Hamar (38) para determinar el IH frente al VEN y la influenza aviar. La prueba de transformación de linfocitos se determinó siguiendo el método de Balhaa et al. (39). Esta prueba mide la proliferación de células T a un fármaco in vitro, a partir de la cual se puede simular una reacción previa in vivo contra la sensibilización a un fármaco. La prueba de transformación de linfocitos se ha implementado ampliamente para el diagnóstico médico de inmunodeficiencia, enfermedades patógenas y alergia tipo IV (40, 41). Se utilizó el método de Rainger y Rowley (42) para determinar la actividad bactericida sérica de la cepa Aeromonas hydrophila, y los resultados se expresaron como índice de supervivencia. Esta prueba se utiliza para medir la actividad bactericida del suero durante el tratamiento antimicrobiano contra bacterias aisladas del mismo paciente. Esta prueba se ha utilizado ampliamente en pacientes con endocarditis infecciosa, osteomielitis, bacteriemia u otras infecciones bacterianas graves (43, 44). Se utilizó el método turbidimétrico de Engstad et al. (45) para medir la actividad de la lisozima sérica. Esta prueba representa la actividad de monocitos/macrófagos y se ha utilizado ampliamente para medir la actividad de diversas enfermedades (46, 47). La reducción de la absorbencia de 0,001/min refina el resultado de esta prueba, que se expresa como una unidad de actividad de lisozima. Actividad de la lisozima = (A0 – A)/A.
Rasgos bioquímicos
Se utilizaron kits comerciales producidos por Diamond Diagnostics (23 EL-Montazah St. Heliopolis, El Cairo, Egipto, http://www.diamonddiagnostics.com) para medir los rasgos bioquímicos de la sangre. Se utilizaron los métodos de Armstrong y Carr (48), Doumas et al. (49) y Doumas y Peters (50) para medir las concentraciones séricas totales de proteínas y albúmina. Se utilizó el método de Cole (51) para estimar la concentración de globulina a través de la resta de la concentración de albúmina de la proteína total sérica. Se utilizó el método de Bossuyt et al. (52) para medir los diferentes tipos de globulina (α-β- y γ-globulina).
Se utilizó el método de Reitman y Frankel (53) para determinar las actividades de la alanina aminotransferasa (ALT, U/L) y la aspartato aminotransferasa (AST, U/L). Las actividades de las enzimas fosfatasa alcalina (ALKP) se ensayaron en muestras por el método de McComb y Bowers (54). Se determinó la capacidad antioxidante total sérica (TAC) y el malondialdehído (MDA) según Erel (55).
Estudio Histopatológico
El intestino (es decir, el íleon), la bolsa de Fabricius, el timo y los especímenes de bazo se recolectaron a los 38 días de edad de seis aves de cada tratamiento dietético, n = 6, y se fijaron directamente en solución salina de formalina tamponada al 10% (BFS) durante al menos 24 h. Se utilizó la técnica convencional de inclusión en parafina para procesar las muestras fijas. Esta técnica consiste en deshidratar a través de grados ascendentes de etanol, aclarar en cloroformo e incrustar en cera de parafina a 60 °C. Los bloques de parafina resultantes se cortaron en rodajas de 5 μm de espesor y se tiñeron con hematoxilina y eosina (H&E) siguiendo el método descrito por Culling (56).
Para la medición cuantitativa morfométrica del eje longitudinal del folículo bursal grande, se examinaron cinco secciones por réplica por tratamiento utilizando el analizador de imágenes Optika montado en un microscopio binocular Optika. Además, se midió cualitativamente la relación entre cortical y medular del timo. Se examinó el bazo para determinar la presencia de hiperplasia de células linfoblásticas donde (–) significa pocas; (+) significa moderado; y (++) significa grave.
Análisis estadístico
Se aplicó el procedimiento del Modelo Lineal General del Software de Análisis Estadístico del Instituto SAS (57) utilizando el diseño factorial de dos vías (dos tipos de alimentación por tres niveles de multienzima) de la siguiente manera:
yijk = μ + Ai + βj + (Aβ)ij + eijk
Aquí, μ = media general, Ai = efecto de los tipos de alimento, βj = efecto de los niveles de multienzima, (Aβ)ij = interacción entre los alimentos y las multienzimas, y eijk = error aleatorio.
Antes del análisis, se realizó la transformación de arcoseno para normalizar la distribución de los datos. Para confirmar la homogeneidad (prueba de normalidad) de los datos, se utilizó la prueba de Kolmogorov-Smirnov (K-S) (57). Las medias se consideran diferentes a P ≤ 0,05 mediante la prueba de Student-Newman-Keuls.
Resultados
Rendimiento del crecimiento
La Tabla 2 muestra el efecto de diferentes tipos de dúos y niveles de multienzimas en el crecimiento de pollos de engorde alimentados con SD y LDD durante los días 1 a 38 de edad. Los resultados de la Tabla 2 muestran que la LDD disminuyó significativamente el BWG de los pollos de engorde en comparación con la DE, pero no afectó a la EPEI durante el período experimental. La interacción entre el nivel multienzimático y el tipo de dieta en BWG y EPEI no fue significativa. Los resultados también indicaron que la suplementación multienzimática al 0,1 y 0,2% aumentó de manera significativa y similar el BWG y mejoró la EPEI, en comparación con la dieta control.
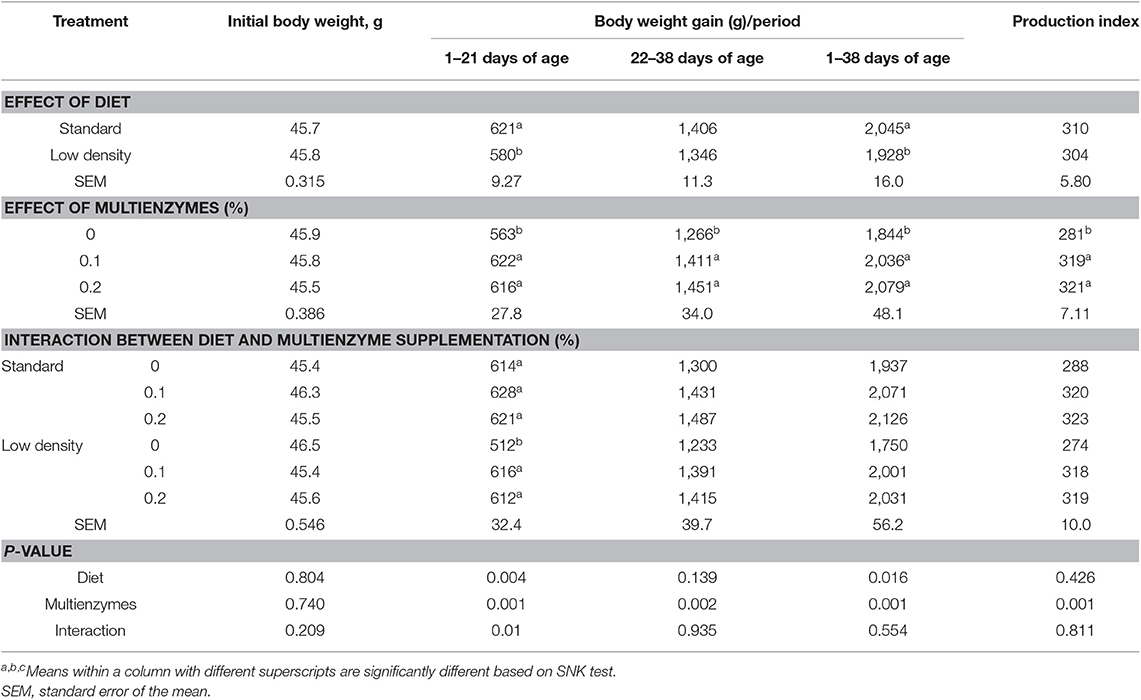 TABLA 2. Efecto de diferentes niveles de multienzimas en el crecimiento de pollos de engorde alimentados con dietas estándar y de baja densidad durante los días 1-38 de edad.
TABLA 2. Efecto de diferentes niveles de multienzimas en el crecimiento de pollos de engorde alimentados con dietas estándar y de baja densidad durante los días 1-38 de edad.
Rasgos hematológicos de la sangre
En las tablas 3 y 4 se muestra el efecto de las dietas experimentales sobre las características hematológicas de la sangre y los glóbulos blancos diferenciales, respectivamente. PCV, Hgb, glóbulos rojos, MCV, MCH, MCHC, GLÓBULOS BLANCOS y diferentes tipos de leucocitos no se vieron afectados por el tipo de dieta. Los linfocitos presentaron una LDD significativamente más elevada, mientras que en el caso de los eosinófilos se observó la tendencia opuesta (Tabla 3). La suplementación con multienzimas aumentó significativamente la Hgb, el PCV (Tabla 3), los linfocitos y los monocitos (Tabla 4) en comparación con la dieta de control. Hubo una interacción significativa entre el tipo de dieta y las concentraciones multienzimáticas sobre Hgb, PCV (Tabla 3) y leucocitos (Tabla 4), mostrando que la multienzima aumentó la Hgb y el PCV de la SD de manera escalonada, pero no afectó la LDD, lo que demuestra que el impacto de la multienzima sobre Hgb y PCV depende del tipo de dieta. La respuesta de los linfocitos a la dosis multienzimática fue escalonada (Tabla 4).
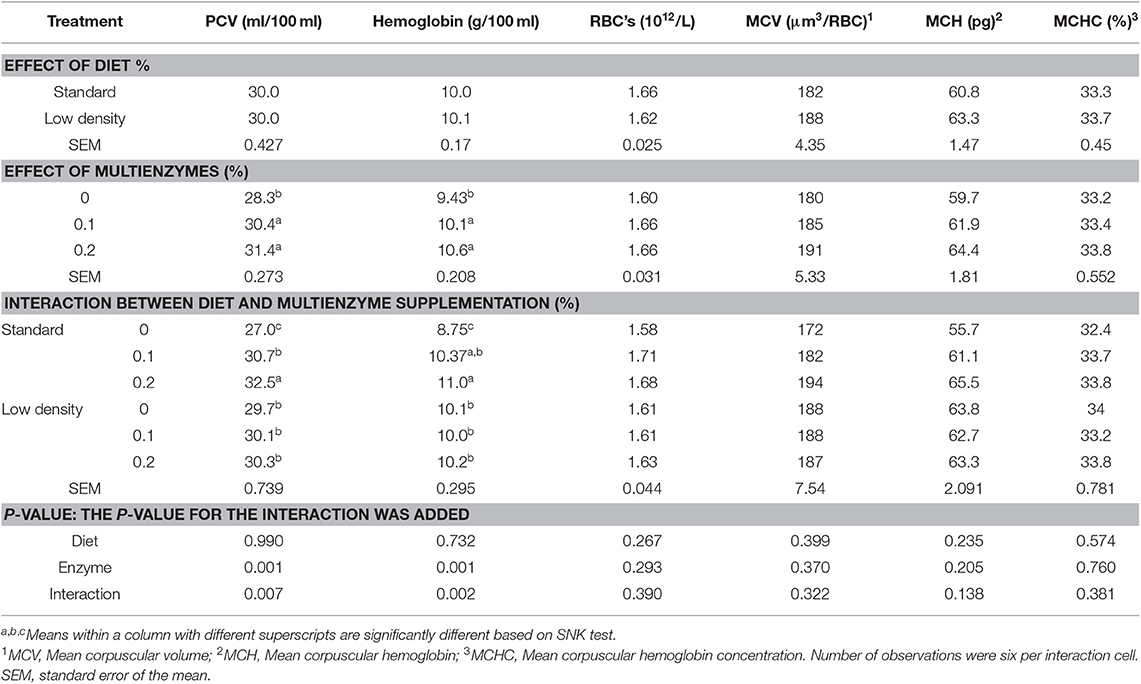 TABLA 3. Efecto de diferentes concentraciones de cóctel enzimático sobre los rasgos hematológicos sanguíneos de pollos de engorde alimentados con dietas estándar y de baja densidad.
TABLA 3. Efecto de diferentes concentraciones de cóctel enzimático sobre los rasgos hematológicos sanguíneos de pollos de engorde alimentados con dietas estándar y de baja densidad.
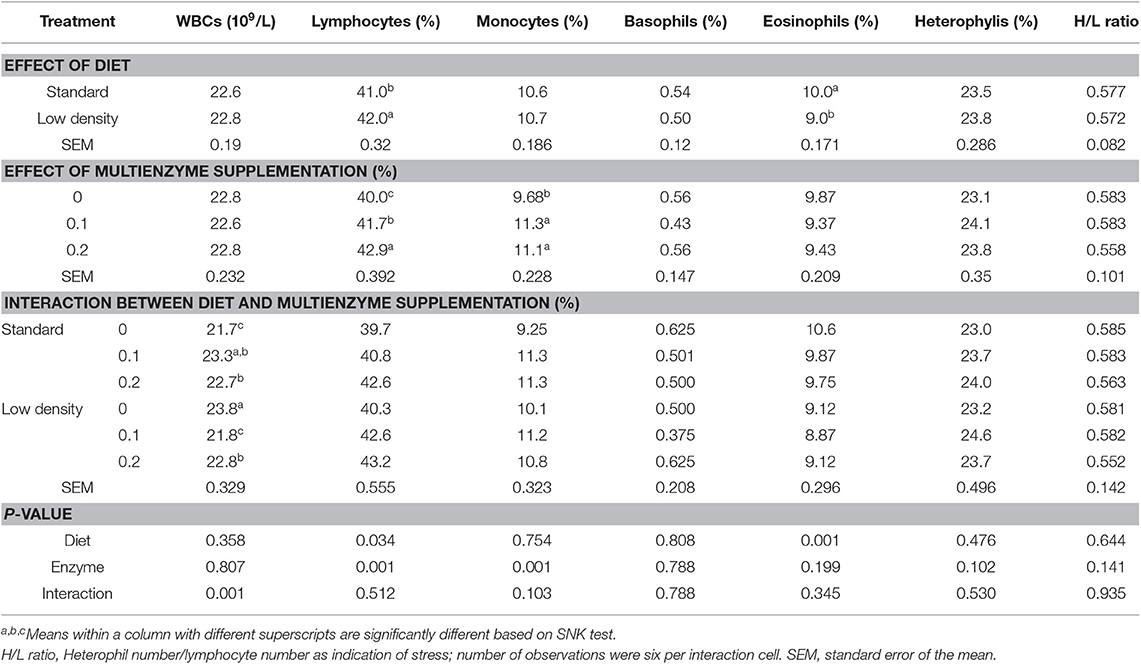 TABLA 4. Efecto de diferentes concentraciones de cóctel enzimático sobre glóbulos blancos y sus fracciones de pollos de engorde alimentados con dieta estándar y de baja densidad.
TABLA 4. Efecto de diferentes concentraciones de cóctel enzimático sobre glóbulos blancos y sus fracciones de pollos de engorde alimentados con dieta estándar y de baja densidad.
Hubo una interacción significativa entre el tipo de dieta y el nivel multienzimático solo en los leucocitos, lo que demuestra que la suplementación con CE a la DE aumentó significativamente los leucocitos pero disminuyó la LDD, lo que demuestra que el impacto de las multienzimas en los leucocitos depende del tipo de dieta.
Componentes bioquímicos de la sangre
Los datos relativos a los componentes bioquímicos de la sangre se muestran en los cuadros 5 y 6. La ALT, la AST y la MAD fueron significativamente mayores en la LDD que en la SD, pero el TAC/MDA (balance de antioxidantes) fue menor (Tabla 5). La proteína total, la albúmina, la α, la β y la γ-globulina, la globulina y la relación globulina/albúmina no se vieron influenciadas significativamente por el tipo de dieta (Tabla 6).
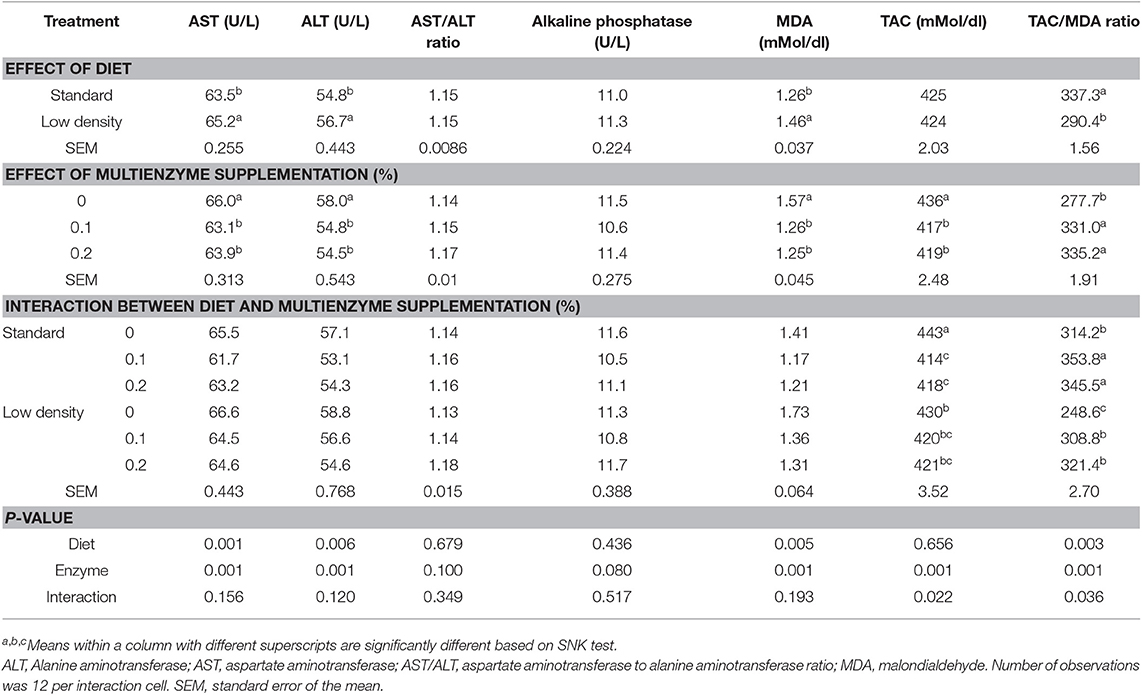 TABLA 5. Efecto de diferentes concentraciones de enzimas sobre las enzimas hepáticas, el malondialdehído sérico en sangre y la capacidad antioxidante total de pollos de engorde alimentados con dieta estándar y de baja densidad.
TABLA 5. Efecto de diferentes concentraciones de enzimas sobre las enzimas hepáticas, el malondialdehído sérico en sangre y la capacidad antioxidante total de pollos de engorde alimentados con dieta estándar y de baja densidad.
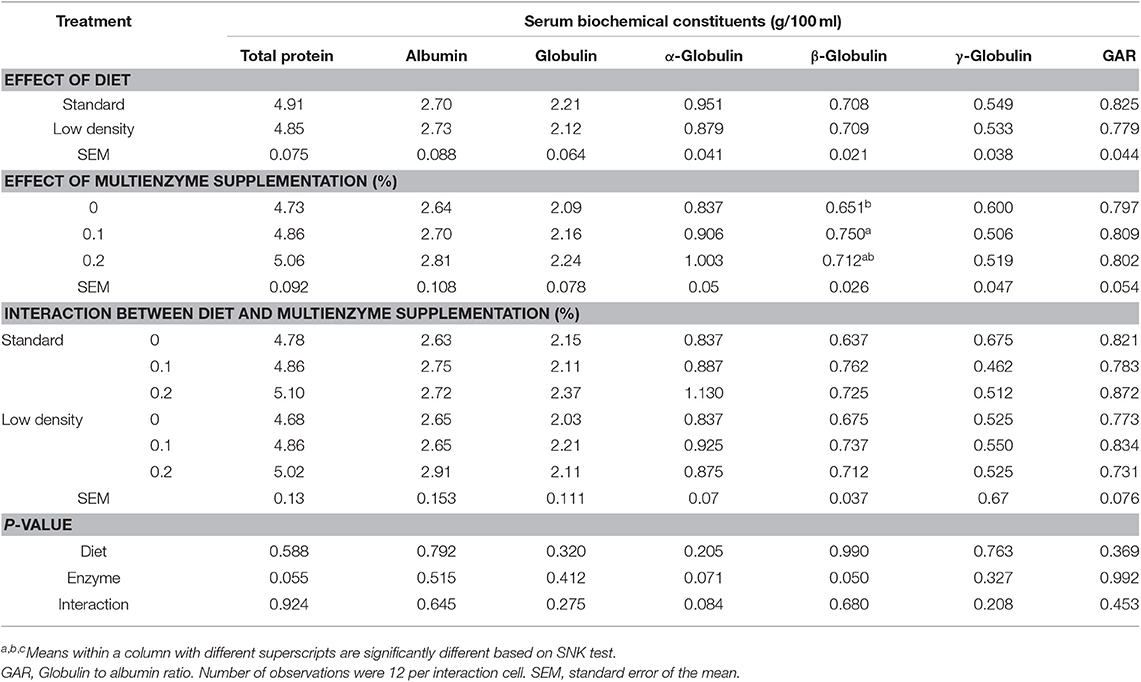 TABLA 6. Efecto de diferentes concentraciones de cóctel enzimático sobre los constituyentes bioquímicos del suero sanguíneo de pollos de engorde alimentados con dieta estándar y de baja densidad.
TABLA 6. Efecto de diferentes concentraciones de cóctel enzimático sobre los constituyentes bioquímicos del suero sanguíneo de pollos de engorde alimentados con dieta estándar y de baja densidad.
La suplementación con multienzimas al 0,1 y 0,2% disminuyó significativamente el ALT, AST, MDA y TAC en comparación con el grupo control (Tabla 5), pero aumentó el equilibrio antioxidante (TAC/MDA). La relación enzimas hepáticas (ALT/AST) y la fosfatasa alcalina no se vieron afectadas significativamente por la suplementación multienzimática. Hubo un efecto significativo de las multienzimas sobre la β-globulina, mostrando mayores valores de los pollitos en la dieta con 0,2 y 0,1% de multienzimas que los de la dieta control sin suplementación multienzimática (Tabla 6). El efecto multienzimático sobre la proteína total fue moderado (P = 0,057).
No hubo una influencia significativa de la interacción entre la suplementación multienzimática y el tipo de dieta sobre el MDA en sangre, las proteínas y las funciones hepáticas reflejadas por las enzimas sanguíneas (ALT, AST, ALT/AST y fosfatasa alcalina). Sin embargo, se observó un efecto significativo de la interacción en el TAC y el balance antioxidante (TAC/MDA). Los resultados indicaron un aumento en la relación TAC/MDA de pollos de engorde suplementados con enzimas de SD y LDD.
Títulos de órganos linfáticos y anticuerpos
La Tabla 7 muestra el efecto de diferentes concentraciones de multienzimas sobre los órganos inmunes y el título de anticuerpos de pollos de engorde alimentados con SD y LDD. El tipo de dieta no tuvo un efecto significativo en los órganos linfáticos, como el bazo, el timo y la bolsa de Fabricio y el HI para el VN. Sin embargo, el IH para la IA fue significativamente mayor en LDD que en SD. Estos órganos también fueron significativamente más altos para los pollos de engorde con dieta suplementada con multienzimas al 0,1 o 0,2% que los que seguían una dieta sin suplementos de EC.
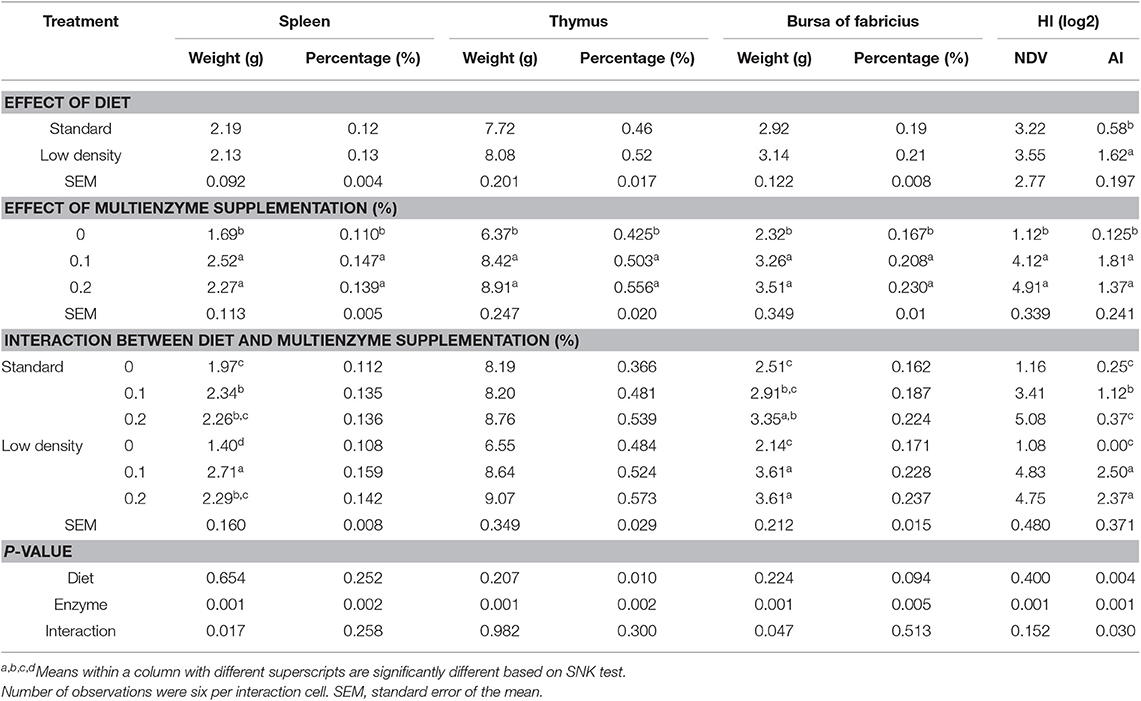 TABLA 7. Efecto de diferentes concentraciones de cóctel enzimático sobre los órganos inmunes y el título de anticuerpos de pollos de engorde alimentados con dieta estándar y de baja densidad.
TABLA 7. Efecto de diferentes concentraciones de cóctel enzimático sobre los órganos inmunes y el título de anticuerpos de pollos de engorde alimentados con dieta estándar y de baja densidad.
Hubo interacciones significativas entre la suplementación multienzimática y el tipo de dieta sobre el peso absoluto del bazo y la bolsa de Fabricius y HI para la IA. Los resultados mostraron que la suplementación multienzimática aumentó significativamente el peso del bazo y la bursa de Fabricius y HI para la IA. Sin embargo, la suplementación multienzimática al 0,1% aumentó significativamente el peso del bazo en comparación con el control no suplementado, mientras que la multienzima al 0,2% aumentó el peso absoluto de la bolsa de Fabricio.
Índices inmunológicos
En la Tabla 8 se muestra el efecto de las diferentes dietas experimentales sobre los índices inmunes. Hubo un efecto significativo del tipo de dieta sobre LTT, BACT y PA, mostrando el efecto potenciador de la LDD sobre la respuesta inmune (Tabla 8).
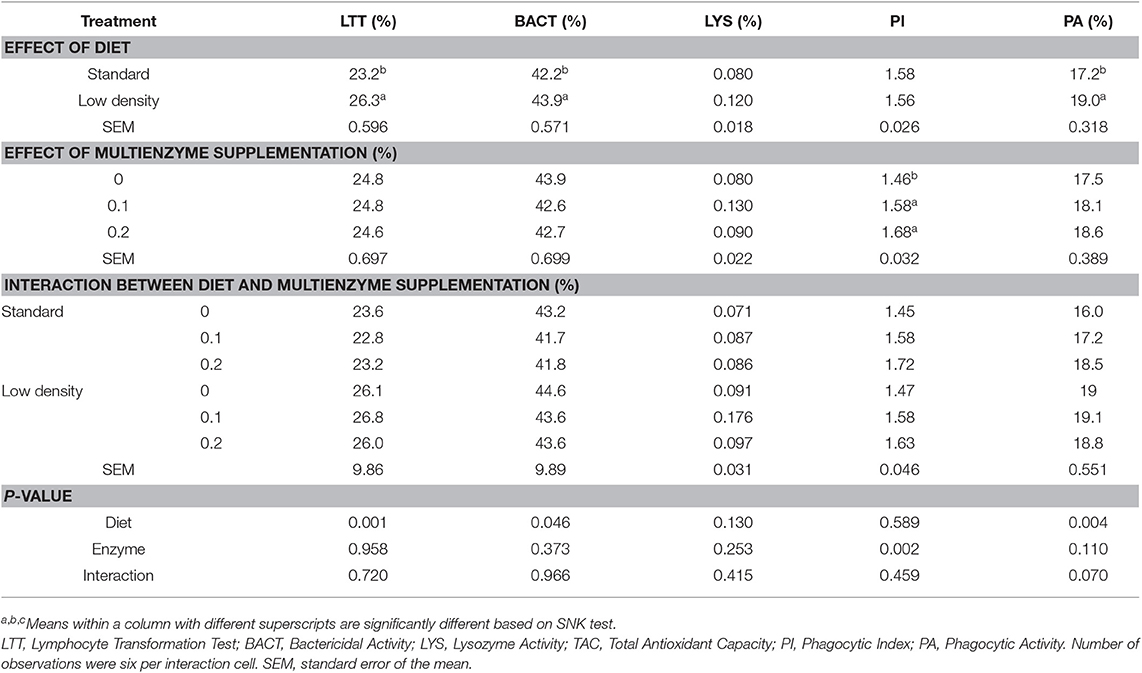 TABLA 8. Efecto de diferentes concentraciones de cóctel enzimático sobre el índice inmunológico de pollos de engorde alimentados con dieta estándar y de baja densidad.
TABLA 8. Efecto de diferentes concentraciones de cóctel enzimático sobre el índice inmunológico de pollos de engorde alimentados con dieta estándar y de baja densidad.
La suplementación con multienzimas aumentó significativamente el IP en comparación con el grupo de control y no tuvo ningún efecto sobre los otros índices inmunes, como LTT, BACT, LYS y PA.
No hubo influencia significativa de la interacción entre la dosis de multienzimas y el tipo de dieta sobre LTT, BACT, LYS, PI y PA.
Estudio histológico
El efecto del tipo de dieta y la suplementación multienzimática sobre la morfología del intestino y la bolsa de Fabricius se muestra en la Tabla 9. El tipo de dieta no tuvo un efecto significativo sobre el diámetro del folículo grande de la bolsa de Fabricio. Hubo un aumento significativo en el diámetro del folículo grande de la bolsa de Fabricio (Figuras 1, 2; Tabla 9) debido a la suplementación de multienzimas al 0,1%, en comparación con las otras concentraciones multienzimáticas. El aumento de la bolsa de Fabricio ascendió al 16,6%, respectivamente. También hubo un aumento insignificante debido a la suplementación con multienzimas al 0,2%.
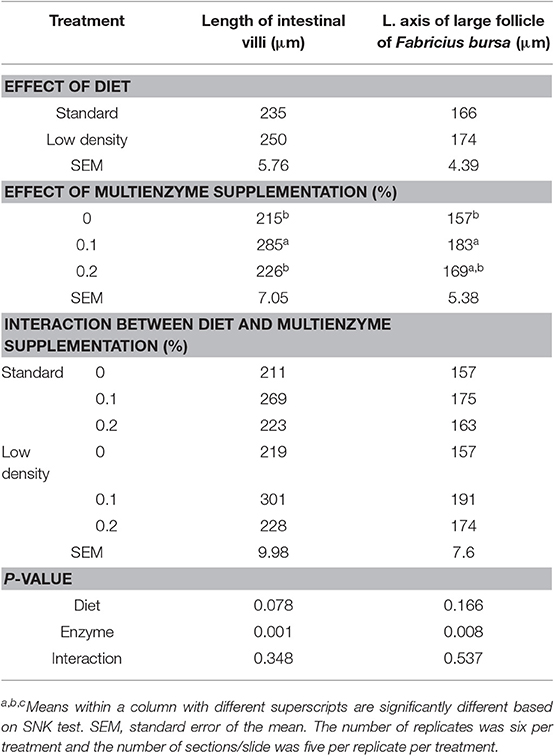 TABLA 9. Efecto del tipo de dieta y la suplementación con cóctel enzimático de la morfología del intestino y de la bursa de Fabricius.
TABLA 9. Efecto del tipo de dieta y la suplementación con cóctel enzimático de la morfología del intestino y de la bursa de Fabricius.
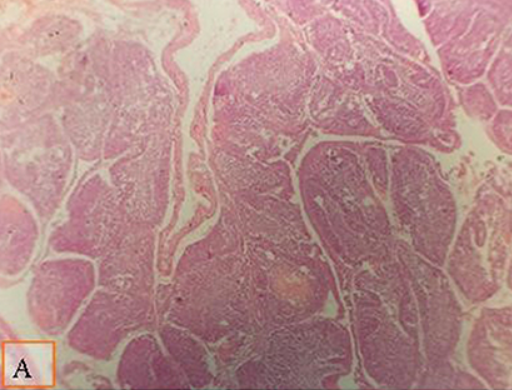
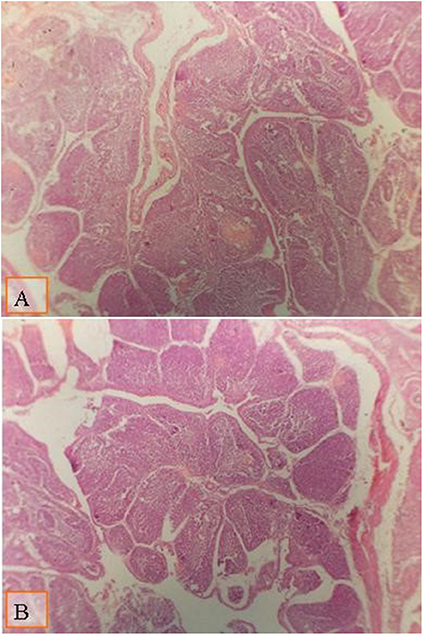 FIGURA 1. Micrografía de la bolsa de Fabricius de pollos de engorde a los 28 días de edad teñida con H&E (×40) para comparar el diámetro del folículo en diferentes grupos; la distancia entre dos polares foliculares como se muestra en todos los grupos por líneas: (A) pollos de engorde alimentados con una dieta estándar suplementada con un cóctel enzimático al 0,1% ml; (B) pollos de engorde alimentados con una dieta suplementada con 0,2% de multienzima. Se observó un aumento moderado en el diámetro folicular en pollos de engorde alimentados con dieta suplementada con 0,1% de multienzima (A).
FIGURA 1. Micrografía de la bolsa de Fabricius de pollos de engorde a los 28 días de edad teñida con H&E (×40) para comparar el diámetro del folículo en diferentes grupos; la distancia entre dos polares foliculares como se muestra en todos los grupos por líneas: (A) pollos de engorde alimentados con una dieta estándar suplementada con un cóctel enzimático al 0,1% ml; (B) pollos de engorde alimentados con una dieta suplementada con 0,2% de multienzima. Se observó un aumento moderado en el diámetro folicular en pollos de engorde alimentados con dieta suplementada con 0,1% de multienzima (A).
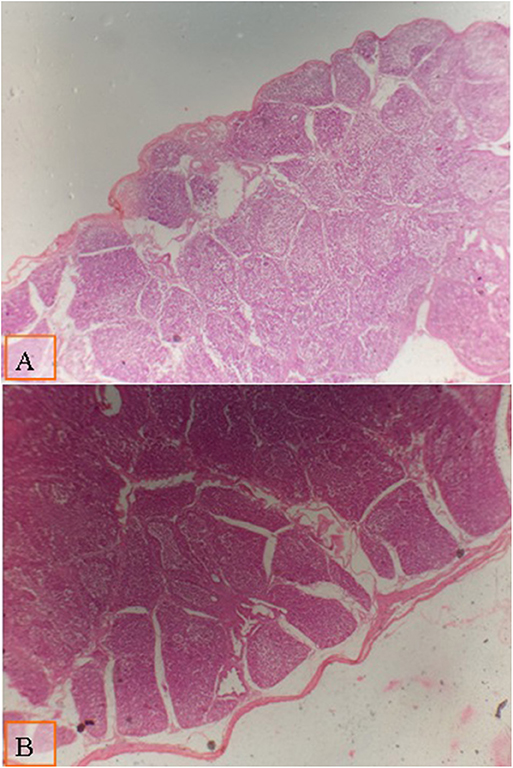 FIGURA 2. Micrografía de la bolsa de Fabricius de pollos de engorde a los 28 días de edad teñida con H&E (×40) para comparar el diámetro del folículo en diferentes grupos; la distancia entre dos polares foliculares como se muestra en todos los grupos por líneas: (A) pollos de engorde alimentados con dieta de baja densidad suplementada con 0,1% de multienzima; (B) pollos de engorde alimentados con una dieta suplementada con 0,2% de multienzima. Se observó un aumento moderado en el diámetro folicular en pollos de engorde alimentados con una dieta suplementada con 0,1% de multienzima (A).
FIGURA 2. Micrografía de la bolsa de Fabricius de pollos de engorde a los 28 días de edad teñida con H&E (×40) para comparar el diámetro del folículo en diferentes grupos; la distancia entre dos polares foliculares como se muestra en todos los grupos por líneas: (A) pollos de engorde alimentados con dieta de baja densidad suplementada con 0,1% de multienzima; (B) pollos de engorde alimentados con una dieta suplementada con 0,2% de multienzima. Se observó un aumento moderado en el diámetro folicular en pollos de engorde alimentados con una dieta suplementada con 0,1% de multienzima (A).
No hubo cambios significativos en el bazo y el timo debido al tipo de dieta y a la suplementación multienzimática y su interacción (Figuras 3 y 4). Sin embargo, la relación cortical/medular en el timo disminuyó con el aumento de la edad de los polluelos. Con el tiempo, el timo comenzó a atrofiarse y la disminución de la relación cortical/medular se volvió fisiológica.
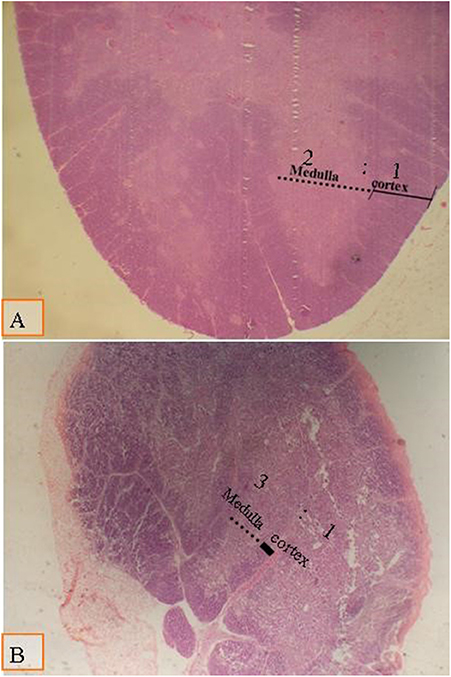 FIGURA 3. Micrografía del timo teñida con H&E (×40) para comparar la relación tímica cortical/medular: (A) la relación cortical/medular es de 2:1 como se muestra en todos los grupos a los 28 días de edad; (B) la ración cortical/medular es de 3:1 como se muestra en todos los grupos a los 38 días de edad, lo que se asoció con atrofia tímica fisiológica envejecida.
FIGURA 3. Micrografía del timo teñida con H&E (×40) para comparar la relación tímica cortical/medular: (A) la relación cortical/medular es de 2:1 como se muestra en todos los grupos a los 28 días de edad; (B) la ración cortical/medular es de 3:1 como se muestra en todos los grupos a los 38 días de edad, lo que se asoció con atrofia tímica fisiológica envejecida.
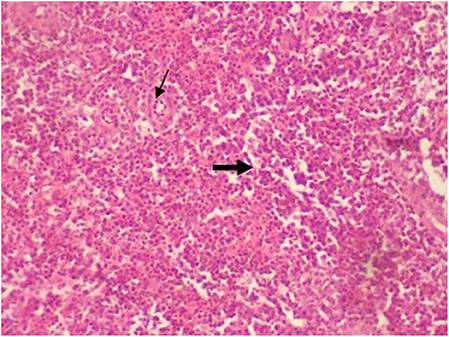 FIGURA 4. Micrografía del bazo teñida con H&E (×40) del grupo control con histología esplénica normal con arteriola esplénica (flecha fina) con pulpa blanca y roja (flecha gruesa). Todos los grupos mostraron la histología esplénica normal como control.
FIGURA 4. Micrografía del bazo teñida con H&E (×40) del grupo control con histología esplénica normal con arteriola esplénica (flecha fina) con pulpa blanca y roja (flecha gruesa). Todos los grupos mostraron la histología esplénica normal como control.
Discusión
Este trabajo se llevó a cabo para estudiar el efecto de la suplementación de tres niveles multienzimáticos (0, 0,1 y 0,2%) con dos tipos de dieta (SD vs. LDD) sobre la respuesta inmunitaria, la hematología y bioquímica de la sangre, el estado antioxidante y la histología de los órganos de los pollos de engorde durante 1 a 38 días de edad. La tendencia en el uso de LDD en la alimentación de pollos de engorde se ha discutido recientemente y puede ser un posible enfoque para disminuir la presión de crecimiento en el sistema esquelético de las aves y reducir el costo de alimentación y la contaminación ambiental (23, 24, 58). Los datos mostraron que la inclusión de salvado de trigo en las dietas de pollos de engorde al 10% indujo una depresión significativa del crecimiento en BWG y FCR solo durante el período de crecimiento temprano, alcanzando 6.61 y 6.74%, respectivamente, que persistió solo en BWG durante todo el período, alcanzando 5.6%. Esto demostró que la LDD no proporcionó los nutrientes adecuados para el crecimiento muscular durante los diferentes períodos experimentales. Sin embargo, la tolerancia de los pollos de engorde a la composición de la dieta aumentó con el aumento de la edad de los pollitos (17, 59). Los resultados actuales concuerdan con los reportados por Al-Harthi (60) y Attia (16, 17). El aumento del peso corporal y el peso corporal como resultado de la suplementación multienzimática puede deberse a la mayor disponibilidad y absorción de nutrientes como resultado del aumento de la digestibilidad de las dietas ingeridas, como sugieren Choct (22), Attia et al. (18) y El-Kelawy (61). Sin embargo, los efectos de la multienzima dependen de la composición de la dieta y del tipo de enzima (17, 62). Anteriormente se informó que la suplementación multienzimática aumentó la BWG y la FCR, al tiempo que redujo la ingesta de alimento. La multienzima aumentó significativamente el crecimiento durante los 7-21 días de edad, y esto se explica por la existencia de enzimas degradadoras de amilasa y NSPs (23, 60, 63, 64). La adición de enzimas exógenas que hidrolizan el NSP de los ingredientes vegetales en las dietas para monogástricos mejora la disponibilidad de energía y el uso de nutrientes y, por lo tanto, mejora la tasa de conversión alimenticia (65). El aumento en la liberación de nutrientes debido a la suplementación enzimática resultó en un mayor nutriente disponible para la absorción, como lo demuestra el aumento en la longitud de las vellosidades intestinales y, por lo tanto, para la reacción bioquímica a favor de la reacción anabólica y la acumulación muscular (18) y también para la función inmune (21, 25).
El uso de LDD mejoró la LTT y la PA, lo que indica una mayor inmunidad. Esta mejora de la inmunidad podría atribuirse a la redistribución de los nutrientes hacia la inmunidad en lugar del crecimiento. El crecimiento disminuyó debido a la alimentación con LDD y, por lo tanto, aumentó la disponibilidad de nutrientes para la respuesta fisiológica, la inmunidad y la utilización de antioxidantes para eliminar los radicales libres resultantes de las reacciones enzimáticas y no enzimáticas debido a la disminución de las demandas de nutrientes para el crecimiento. Por ejemplo, la relación TAC/MDA (equilibrio antioxidante) aumentó significativamente debido a la alimentación con una dieta suplementada con enzimas. Este aumento reflejó la disminución significativa de los antioxidantes (TAC) para su uso en la protección celular de los radicales libres (7, 25).
Se ha informado que la inclusión de salvado de trigo en la dieta y particularmente los polisacáridos son antioxidantes, inmunomoduladores, antiinflamatorios, antitusivos, anticancerígenos y antimutagénicos (1, 66-68). Además, se ha demostrado que el salvado de trigo y los arabinoxilanos mejoran la fagocitosis de los macrófagos en modelos animales. También estimulan la respuesta humoral en los pollos. Mejoró la producción de títulos totales de anticuerpos anti-SRBC de IgS, IgG e IgM en los días 7 y 14 PPI y PSI de SRBC, en relación con el estándar (69, 70). La administración de arabinoxilanos en la dieta aumentó significativamente la respuesta de anticuerpos anti-SRBC en pollos a los que se administraron arabinoxilanos y sugirió un aumento de la respuesta inmune humoral, lo que puede atribuirse a un mayor potencial de un animal para encontrarse con agentes patógenos (71). Además, el salvado de trigo es rico en la enzima fitasa, que puede mejorar la disponibilidad de nutrientes por parte de los pollos, como proteínas, energía y minerales (17, 72).
Curiosamente, los resultados actuales mostraron que la CE mejoró el EPEI de los pollos de engorde. Este resultado está en línea con los mostrados por Al-Harthi (60) y Attia (16, 17). La mejoría de la EPPI debido a la suplementación con AE coincidió con una mayor longitud de las vellosidades en el grupo con suplementación multienzimática al 0,1%; esto está en línea con los hallazgos de Choct (22) y Attia et al. (18). Sin embargo, el efecto de la multienzima depende de la composición de la dieta y del tipo de enzima (17, 62). El efecto de la suplementación enzimática en el rendimiento de los animales podría ser el resultado del efecto de la amilasa y las enzimas que degradan las NSP (23, 60, 63, 64). La adición de enzimas exógenas que hidrolizan el NSP de los ingredientes vegetales en los piensos para monogástricos mejora la disponibilidad de energía y el uso de nutrientes y, por lo tanto, mejora el FCR (65). Además, Gao et al. (73) informaron que la suplementación con xilanasa de las dietas a base de trigo para gallos mejoró significativamente la producción de respuesta de anticuerpos séricos al VEN como medio de respuesta humoral. Por el contrario, Basmacioglu Malayoglu et al. (74) revelaron que la suplementación con enzimas degradadoras de NSP no tuvo un efecto significativo sobre la respuesta inmune representada por títulos de IgG e IgM. Además, Khaksar et al. (75) encontraron que el peso relativo de los órganos inmunes (timo, bazo y la bolsa de Fabricio) no estaba influenciado por la suplementación enzimática.
Además, la intervención de la composición de la dieta con enzimas fue estudiada recientemente por Head et al. (76) quienes sugirieron que si los pollos de engorde se suplementan con linaza, la digestibilidad de los nutrientes y la disponibilidad de AG n-3 se verán limitadas debido a la presencia de polisacáridos no amiláceos (NSP), al afectar el proceso genético durante el metabolismo de los lípidos en el hígado. Los resultados revelaron que la adición de enzimas a las dietas suplementadas con un 10% de linaza disminuyó el ácido araquidónico y el AG n-6 de cadena larga total. Los tratamientos dietéticos con linaza y enzimas aumentaron la regulación de los genes diana de PPARα CPT1A y ACOX1, al tiempo que redujeron la expresión de genes relacionados con la síntesis de AG de novo (76). Además, Seidavi et al. (77) demostraron que la suplementación de la dieta con una mezcla de probióticos y enzimas no tuvo ningún efecto sobre la respuesta humoral contra la IA y el VEN en el día 42. Hubo una diferencia significativa de IgG después del segundo desafío con los SRBC (P = 0,003). Se encontró un menor peso corporal de las aves en la mayoría de los grupos tratados, en relación con el estándar (P = 0,031). El peso absoluto y relativo del bazo fue significativamente diferente en comparación con el grupo control (P = 0,003 y P = 0,001, respectivamente). Los pesos del timo y de la bolsa de Fabricio no eran diferentes (77). Un estudio adicional fue realizado por Abdel-Hafeez et al. (78) quienes encontraron que el uso de cáscaras de papa al 15% y pulpa de remolacha azucarera al 7,5% disminuyó el peso corporal. La inclusión enzimática mejoró significativamente el peso corporal en las cáscaras de patata y la pulpa de remolacha azucarera. Los niveles séricos de colesterol total y colesterol de lipoproteínas de baja densidad se redujeron en todos los grupos experimentales. Además, el contenido de grasa de la canal se redujo utilizando las cáscaras de patata y la pulpa de remolacha azucarera, con o sin enzima (78). Esto indica que la suplementación con enzimas afecta la bioquímica sanguínea de los pollos de engorde (61) y, por lo tanto, la calidad de la carne mejoró debido a la disminución de la grasa de la canal (18).
Estos resultados sugieren que es posible diluir los perfiles nutricionales de las dietas de los pollos de engorde durante las fases de crecimiento y engorde sin efectos negativos sobre el EPEI y la EC, al tiempo que se mejora la respuesta inmune y la eficiencia económica. Estos resultados concuerdan con los reportados por Abudabos (23).
El uso de LDD en el presente estudio no afectó negativamente a la longitud de las vellosidades. La relación entre la morfología intestinal y el tipo de dieta fue reportada en la literatura; el uso de cereales con un alto nivel de NSP puede aumentar el tamaño del tracto gastrointestinal (72, 79). Se ha reportado una correlación positiva significativa entre el nivel de arabinoxilano en trigo y los pesos relativos del duodeno, yeyuno e íleon (80). Se sabe que la composición de la dieta puede inducir cambios microscópicos en la mucosa intestinal, y es posible que los niveles de NSP en la dieta también puedan afectar la morfología del tracto gastrointestinal (81). Iji (82) informó que la profundidad de la cripta tanto del yeyuno como del íleon mejoró debido a la suplementación con goma guar y goma xantana, lo que indica que la NSP puede estimular el recambio celular del tracto gastrointestinal. El aumento de la profundidad de las criptas indica un aumento de la estimulación de las células vellosas y, por lo tanto, un aumento de los nutrientes que se absorben y, por lo tanto, son utilizados por el tracto gastrointestinal. Esto indica que el tipo de cereal afecta el tamaño del tracto gastrointestinal y la morfología del intestino (83).
Es bien sabido que el aumento de la corteza en relación con la relación con la médula aumentó el número de linfoblastos T y aumentó el linfocito T, lo que se refleja en la estimulación de la inmunidad celular (84-86).
Un estudio reciente de Woyengo et al. (87) mostró que el uso de amilasa, NSPasa o una combinación de amilasa y NSPasa en la dieta basal suplementada con fitasa mejoró aún más (P < 0,05) la digestibilidad del íleon a 63,4, 69,9 y 67,3%, respectivamente. Sin embargo, el valor de energía metabolizable aparente corregido por nitrógeno de la dieta no se vio afectado por la adición de fitasa, amilasa o una combinación de amilasa y NSPasa. Woyengo et al. (87) concluyeron que la adición de amilasa y NSPasa a las dietas suplementadas con fitasa de pollos de engorde es beneficiosa para mejorar la energía metabolizable aparente corregida por nitrógeno. En otro estudio realizado por Sateri et al. (88), no mostraron efectos significativos de la harina de aceituna y las enzimas sobre el rendimiento del crecimiento y sobre la microflora del ciego. Los títulos de anticuerpos contra el virus de la bronquitis infecciosa (IBV) y la enfermedad de Gumboro fueron más altos en las aves alimentadas con harina de aceituna al 4%. La suplementación de la dieta con enzimas no afectó a los parámetros antes mencionados (88).
Los presentes resultados indican que la suplementación con CE mejoró la inmunidad de los pollos de engorde medida por los cambios en los órganos. Esto coincidió con un aumento del diámetro del folículo grande de la bolsa de Fabricio para el grupo suplementado con multienzimas al 0,1%. El aumento en el diámetro folicular de la bolsa de Fabricius indica un aumento en el número de linfoblastos B que conduce a la formulación de linfocitos B que internamente forman anticuerpos. Esto refleja la estimulación de la inmunidad humoral debido al aumento de los nutrientes disponibles para la formación de anticuerpos (89, 90). Además, el índice fagocítico aumentó de manera similar con la suplementación de concentraciones multienzimáticas de 0,1 y 0,2%. El mecanismo o mecanismos por los que la suplementación con salvado de trigo y/o enzimas afectó positivamente a la inmunidad de los pollitos puede implicar una mejora de la salud intestinal debido a la disminución de la microbiota dañina en el intestino posterior, el aumento de la longitud de las vellosidades intestinales y/o el aumento de los nutrientes disponibles para la función inmunitaria (21, 24, 25, 73, 89, 90). A este respecto, Gao et al. (73) informaron que la alimentación de gallos con dietas a base de trigo suplementadas con xilanasa aumenta significativamente los anticuerpos séricos contra el VVEN, lo que indica que los suplementos enzimáticos mejoran la respuesta humoral debido a una mayor utilización de nutrientes para la función inmune. Pero en otro estudio, no hubo un efecto significativo en la respuesta humoral (IgG e IgM) con la suplementación con enzimas que degradan NSP (74). Además, Khaksar et al. (75) encontraron que el peso relativo de los órganos inmunes (timo, bazo y la bolsa de Fabricio) no estaba influenciado por la suplementación enzimática. Esta contraindicación en la literatura publicada indica que la inmunidad de las aves es un concepto complejo que se ve afectado por la composición de la dieta, la edad y el estrés de los animales (24, 25, 84).
En conclusión, la suplementación multienzimática al 0,1 o al 0,2% de SD o LDD mejoró el índice de producción al tiempo que mejoró la respuesta inmunitaria de los pollos de engorde durante la edad de 1 a 38 días.
Declaración de disponibilidad de datos
Los conjuntos de datos generados para este estudio están disponibles previa solicitud al autor correspondiente.
Declaración de Ética
El estudio en animales fue revisado y aprobado por el Comité Departamental de Producción Animal y Avícola, los decretos oficiales del Ministerio de Agricultura de Egipto relativos al bienestar animal son el Nº 27 (1967).
Contribuciones de los autores
Todos los autores enumerados han hecho una contribución sustancial, directa e intelectual al trabajo, y lo han aprobado para su publicación.
Financiación
Este artículo fue financiado por el Decanato de Investigación Científica (DSR) de la Universidad Rey Abdulaziz, Jeddah; los autores, por lo tanto, agradecen el apoyo técnico y financiero de DSR.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
El editor encargado declaró una coautoría pasada con uno de los autores YA.
Reconocimientos
Los autores expresaron su agradecimiento a la dirección del Instituto de Investigación Científica de Kuwait (KISR) por su amable ayuda y apoyo.
Referencias
1. Escuela Secundaria Al-Khalaifah. Beneficios de los probióticos y/o prebióticos para las aves de corral reducidas en antibióticos. Poult Sci. (2018) 97:3807–15. doi: 10.3382/ps/pey160
Resumen de PubMed | Texto completo de CrossRef | Google Académico
2. Kutlu HR, Saber SN, Kutay H, Celik L, Uzun Y, Toy N, et al. Efecto de la multienzima producida por un solo hongo sobre el rendimiento del crecimiento y algunos parámetros de la canal de pollos de engorde alimentados con dietas a base de maíz-soja. Kafkas üniv vet fakült dergisi. (2019) 25:221–30. doi: 10.9775/kvfd.2018.20765
3. Sun HY, Kim IH. Efectos de la multienzima en el rendimiento de la producción, la calidad del huevo, la digestibilidad de los nutrientes y la emisión de gases nocivos de excretas de gallinas marrones Hy-line en fase temprana. Poult Sci. (2019) 98:4889–95. doi: 10.3382/ps/pez237
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Attia YA, Addeo NF, Abd Al-Hamid AA-HE, Bovera F. Efectos de la suplementación con fitasa a dietas con o sin adición de zinc sobre el rendimiento del crecimiento y la utilización de zinc de patos pekín blancos. Animales (Basilea). (2019) 9:280. doi: 10.3390/ani9050280
Resumen de PubMed | Texto completo de CrossRef | Google Académico
5. Al-Khalaifa H, Al-Nasser A, Al-Surayee T, Al-Kandari S, Al-Enzi N, Al-Sharrah T, et al. Efecto de los probióticos y prebióticos dietéticos en el rendimiento de pollos de engorde. Poult Sci. (2019) 98:4465–79. doi: 10.3382/ps/pez282
6. Al-Khalifa H. Técnicas inmunológicas en estudios aviares. Worlds Poult Sci J. (2016) 72:573–84. doi: 10.1017/S0043933916000532
7. Al-Khalifa H. Enriquecimiento de las dietas de las aves de corral con ácidos grasos poliinsaturados (AGPI) para el consumo humano. Acércate a Poult Dairy Vet Sci. (2017) 1:APDV.000523. doi: 10.31031/APDV.2017.01.000523
8. Al-Khalifa H, Al-Nasser A, Al-Bahouh M, Ragheb G, Al-Qalaf S, Al-Omani N, et al. Efecto de los ácidos grasos poliinsaturados en las subpoblaciones de células inmunitarias aviares en sangre periférica, bazo y timo. Worlds Poult Sci J. (2016) 72:531–4. doi: 10.1017/S0043933916000428
9. Al-Khalifa H. Producción de carne de aves de corral de valor añadido: enriquecimiento con ácidos grasos poliinsaturados n-3. Worlds Poult Sci J. (2015) 71:319–26. doi: 10.1017/S004393391500032X
10. Hosseindoust A, Lee S, Gook Nho W, Song YH, Shin JS, Laxman Ingale S, et al. Un estudio dosis-respuesta para evaluar los efectos de la β-mananasa estable al pH derivada de Trichoderma citrinoviride sobre el rendimiento del crecimiento, la retención de nutrientes y la morfología intestinal en pollos de engorde. Ital J Anim Sci. (2019) 18:147–54. doi: 10.1080/1828051X.2018.1500872
11. Liu N, Wang JQ, Gu KT, Deng QQ, Wang JP. Efectos de los niveles de proteínas en la dieta y la suplementación multienzimática en el rendimiento del crecimiento y los marcadores de la salud intestinal de los pollos de engorde alimentados con una dieta basada en comidas diversas. Anim Feed Sci Technol. (2017) 234:110–7. doi: 10.1016/j.anifeedsci.2017.09.013
12. Douglas MW, Parsons CM, Bedford MR. Efecto de varias fuentes de harina de soja y avizima en el rendimiento del crecimiento de los pollitos y la energía digestible ileal. J Appl Poult Res. (2000) 9:74–80. doi: 10.1093/japr/9.1.74
13. Suresh G, Santos DU, Rouissi T, Brar SK, Mehdi Y, Godbout S, et al. Producción y evaluación in vitro de una formulación enzimática como alternativa potencial a los antibióticos alimentarios en aves de corral. Bioquímica de procesos. (2019) 80:9–16. doi: 10.1016/j.procbio.2019.01.023
14. Bedford MR. Nuevas tecnologías enzimáticas para la alimentación de aves de corral. Hno. Poult Sci. (2003) 44:14–6. doi: 10.1080/713655277
15. Aftab U, Bedford MR. El uso de enzimas NSP en la nutrición avícola: mitos y realidades. Worlds Poult Sci J. (2018) 74:277–86. doi: 10.1017/S0043933918000272
16. Attia YA, El-Razak A, El-Din ET, Zeweil HS, Hussein AS, Qota ESM, et al. El efecto de la suplementación de la enzima sobre la puesta y el rendimiento reproductivo en gallinas de codorniz japonesas alimentadas con harina de semillas de nigella. J Poult Sci. (2008) 45:110–5. doi: 10.2141/jpsa.45.110
17. Attia YA. Valor del salvado de arroz, su utilización máxima y mejora mediante fitasa y otras enzimas y formulación de la dieta basada en los aminoácidos disponibles para los pollos de engorde. Archiv Geflugelk. (2003) 67:157–66.
18. Attia YA, El-Tahawy WS, Abd El-Hamid AE-HE, Hassan SS, Nizza A, El-Kelaway MI. Efecto de la fitasa con o sin suplementación multienzimática sobre el rendimiento y la digestibilidad de los nutrientes de los pollos de engorde jóvenes alimentados con dietas de puré o desmenuzado. Ital J Anim Sci. (2012) 11:E56. doi: 10.4081/ijas.2012.e56
19. Hussein EOS, Suliman GM, Abudabos AM, Alowaimer AN, Ahmed SH, Abd El-Hack ME, et al. Efecto de una dieta baja en energía y suplementada con enzimas sobre el crecimiento de los pollos de engorde, las características de la canal y la calidad de la carne. Raza Arch Anim. (2019) 62:297–304. doi: 10.5194/aab-62-297-2019
Resumen de PubMed | Texto completo de CrossRef | Google Académico
20. Cowieson AJ, Singh DN, Adeola O. Predicción de la calidad de los ingredientes y el efecto de una combinación de xilanasa, amilasa, proteasa y fitasa en las dietas de pollos de engorde. 1. Rendimiento de crecimiento e ingesta de nutrientes digeribles. Hno. Poult Sci. (2006) 47:477–89. doi: 10.1080/00071660600830603
Resumen de PubMed | Texto completo de CrossRef | Google Académico
21. Yang X, Zhang B, Guo Y, Jiao P, Long F. Efectos de los lípidos dietéticos y Clostridium butyricum en la deposición de grasa y la calidad de la carne de pollos de engorde. Poult Sci. (2010) 89:254–60. doi: 10.3382/ps.2009-00234
Resumen de PubMed | Texto completo de CrossRef | Google Académico
22. Choct M. Enzimas para la industria de piensos: pasado, presente y futuro. Worlds Poult Sci J. (2006) 62:5–16. doi: 10.1079/WPS200480
23. Attia YA. Efectos de la fitasa microbiana sin o con enzimas que rompen la pared celular sobre el rendimiento de pollos de engorde alimentados con niveles marginales de proteína dietética y energía metabolizable. Egipto Poult Sci. (2001) 21:521–47.
24. Abudabos AM. Efecto de la suplementación enzimática en dietas normales y de baja densidad para pollos de engorde a base de harina de maíz-soja. Asiático J Anim Vet Adv. (2012) 7:139–48. doi: 10.3923/ajava.2012.139.148
25. Attia YA, Al-Khalaifah H, Ibrahim MS, Al-Hamid AEA, Al-Harthi MA, El-Naggar A. Constituyentes hematológicos y bioquímicos de la sangre, enzimas antioxidantes, inmunidad y órganos linfoides de pollos de engorde suplementados con propóleos, polen de abeja y manano-oligosacáridos de forma continua o intermitente. Poult Sci. (2017) 96:4182–92. doi: 10.3382/ps/pex173
26. Cooper EL, Ma MJ. Comprender la nutrición y la inmunidad en el manejo de enfermedades. J Tradit Complemento Med. (2017) 7:386–91. doi: 10.1016/j.jtcme.2016.12.002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
27. NRC. Requerimientos de nutrientes de las aves de corral. Washington, DC: National Academy Press (1994).
29. Eilers RJ. Notificación de la adopción final de un método internacional y una solución estándar para las especificaciones de hemoglobinometría para la preparación de la solución estándar. Am J Clin Pathol. (1967) 47:212–4. doi: 10.1093/ajcp/47.2.212
Resumen de PubMed | Texto completo de CrossRef | Google Académico
31. Jain SK. Peroxidación lipídica de membrana en eritrocitos del recién nacido. Clin Chim Acta. (1986) 161:301–6. doi: 10.1016/0009-8981(86)90014-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
32. Kawahara E, Ueda T, Nomura S. Actividad fagocítica in vitro de las células sanguíneas carbonizadas con manchas blancas después de la inyección con productos extracelulares de Aeromonas salmonicida. Pathol de pescado. (1991) 26:213–4. doi: 10.3147/jsfp.26.213
33. Hannum LG, Chen T. La edad avanzada no disminuye la actividad fagocítica ni la producción de NET en los leucocitos renales del pez cebra (Danio rerio). J Immunol. (2018) 200:46.22.
34. Grasse M, Rosenkrands I, Olsen A, Follmann F, Dietrich J. Ensayo basado en citometría de flujo para determinar la actividad fagocítica de muestras de anticuerpos clínicos y no clínicos contra Chlamydia trachomatis. Citometría A. (2018) 93:525–32. doi: 10.1002/cyto.a.23353
Resumen de PubMed | Texto completo de CrossRef | Google Académico
35. Jeffery N, Sanderson P, Sherrington E, Newsholme E, Calder P. La proporción de ácidos grasos poliinsaturados n-6 a n-3 en la dieta de ratas altera los niveles séricos de lípidos y las funciones de los linfocitos. Lípidos. (1996) 31:737–45. doi: 10.1007/BF02522890
Resumen de PubMed | Texto completo de CrossRef | Google Académico
36. King DJ, Seal BS. Caracterización biológica y molecular de aislados de campo del virus de la enfermedad de Newcastle (VN) con comparaciones con cepas de referencia del VN. Avian Dis. (1998) 42:507–16. doi: 10.2307/1592677
Resumen de PubMed | Texto completo de CrossRef | Google Académico
37. Cosgrove A. Una enfermedad aparentemente nueva de los pollos: la nefrosis aviar. Avian Dis. (1962) 6:385–9. doi: 10.2307/1587909
38. Takatsy GY. El uso de bucles en espiral en micrométodos serológicos y virológicos. Acta Microbiol Acad Sci. Hung. (1956) 3:197.
39. Balhaa RL, Hinz HH, Luders H, Siegmann O. Experiencias clínicas con los fármacos para la transformación de linfocitos en parvadas de pollos y pavos. Tierarztliche Umschau. (1985) 43:507–8.
40. Azoury ME, Filì L, Bechara R, Scornet N, de Chaisemartin L, Weaver RJ, et al. Identificación de epítopos de células T de bencilpenicilina conjugados con albúmina sérica humana e implicación en la alergia a la penicilina. Alergia. (2018) 73:1662–72. doi: 10.1111/all.13418
Resumen de PubMed | Texto completo de CrossRef | Google Académico
41. Mori F, Fili L, Barni S, Giovannini M, Capone M, Novembre EM, et al. La sensibilización a la amoxicilina/ácido clavulánico puede ser la base de erupciones cutáneas graves en niños tratados por mononucleosis infecciosa. J Alergia Clin Immunol Pract. (2019) 7:728–31.e1. doi: 10.1016/j.jaip.2018.06.022
Resumen de PubMed | Texto completo de CrossRef | Google Académico
42. Rainger G, Rowley A. Actividad antibacteriana en el suero y el moco de la trucha arco iris, Oncorhynchus mykiss, después de la inmunización con Aeromonas salmonicida. Pescados y mariscos Immunol. (1993) 3:475–82. doi: 10.1006/fsim.1993.1046
43. Loose M, Naber KG, Hu Y, Coates A, Wagenlehner FME. Actividad bactericida sérica de combinaciones de colistina y azidotimidina frente a Escherichia coli resistente a coli mcr-1 positiva. Int J Agentes antimicrobianos. (2018) 52:783–9. doi: 10.1016/j.ijantimicag.2018.08.010
Resumen de PubMed | Texto completo de CrossRef | Google Académico
44. Nahm MH, Yu J, Weerts HP, Wenzel H, Tamilselvi CS, Chandrasekaran L, et al. Desarrollo, evaluaciones interlaboratorios y aplicación de un ensayo bactericida sérico Shigella simple y de alto rendimiento. mSphere. (2018) 3:E00146–18. doi: 10.1128/mSphere.00146-18
Resumen de PubMed | Texto completo de CrossRef | Google Académico
45. Engstad RE, Robertsen B, Frivold E. El glucano de levadura induce un aumento de la actividad hemolítica mediada por lisozima y complemento en la sangre del salmón del Atlántico. Pescados y mariscos Immunol. (1992) 2:287–97. doi: 10.1016/S1050-4648(06)80033-1
46. Ceballos-Francisco D, Guardiola FA, Cordero H, Cuesta A, Esteban MÁ. Parámetros inmunes humorales en suero de dorada (Sparus aurata L.) después de una lesión cutánea inducida. Pescados y mariscos Immunol. (2018) 75:291–4. doi: 10.1016/j.fsi.2018.02.017
Resumen de PubMed | Texto completo de CrossRef | Google Académico
47. Luo C, Gwekwe B, Choto P, Miao W, Chen M, Xue C, et al. Los péptidos amargos de la proteína hidrolizada enzimáticamente aumentan el número de leucocitos y la actividad de la lisozima de la corvina amarilla grande (Larimichthys crocea). Pescados y mariscos Immunol. (2018) 81:130–4. doi: 10.1016/j.fsi.2018.07.013
Resumen de PubMed | Texto completo de CrossRef | Google Académico
48. Armstrong WD, Carr CW. Estimación de la proteína total sérica. En: Instrucciones del Laboratorio de Química Fisiológica, 3ª ed. Minneapolis, MN: Burges Publishing Co. (1964). 153 págs.
49. Doumas BT, Watson WA, Biggs HG. Patrones de albúmina y medición de albúmina sérica con verde de bromcresol. Clin Chim Acta. (1997) 258:21–30. doi: 10.1016/S0009-8981(96)06447-9
50. Doumas BT, Peters T. Suero y albúmina en orina: un informe de progreso sobre su medición e importancia clínica. Clin Chim Acta. (1997) 258:3–20. doi: 10.1016/S0009-8981(96)06446-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Cole DF. Acción de la bradiquinina sobre la presión intraocular y el diámetro pupilar. Res. Oftalmológica. (1974) 6:308–14. doi: 10.1159/000264717
52. Bossuyt X, Lissoir B, Mariën G, Maisin D, Vunckx J, Blanckaert N, et al. Electroforesis automatizada de proteínas séricas por capilares®. Clin Chem Lab Med. (2003) 41:704–10. doi: 10.1515/CCLM.2003.107
53. Reitman S, Frankel S. Método colorimétrico para la determinación de transaminasas glutámicas oxalacéticas y glutámicas pirúvicas séricas. Am J Clin Pathol. (1957) 28:56–63. doi: 10.1093/ajcp/28.1.56
Resumen de PubMed | Texto completo de CrossRef | Google Académico
54. McComb RB, Bowers GN. Estudio de las condiciones óptimas de tampón para medir la actividad de la fosfatasa alcalina en suero humano. Clin Chem. (1972) 18:97–104.
55. Erel O. Un novedoso método automatizado de medición directa de la capacidad antioxidante total utilizando un catión radical ABTS de nueva generación y más estable. Clin Biochem. (2004) 37:277–85. doi: 10.1016/j.clinbiochem.2003.11.015
Resumen de PubMed | Texto completo de CrossRef | Google Académico
56. Culling CF. Manual de Técnicas de Tinción Histopatológica e Histoquímica. 3ª ed. Londres: Butterworth (1983).
57. SAS Intstitute Inc. SAS/STAT Software, versión 9. Cary, Carolina del Norte: SAS Institute, Inc. (2002).
58. Martínez Y, Carrión Y, Rodríguez R, Valdivié M, Olmo C, Betancur C, et al. Rendimiento de crecimiento, peso de órganos y algunos parámetros sanguíneos de pollitas ponedoras de reemplazo alimentadas con niveles crecientes de salvado de trigo. J Braz J Poult Sci. (2015) 17:347–54. doi: 10.1590/1516-635X1703347-354
59. Jeroch H, Dänicke S, Brufau J. Influencia de las preparaciones enzimáticas en el valor nutricional de los cereales para aves de corral. Una revisión. J Anim Feed Sci. (1995) 4:263–85. doi: 10.22358/jafs/69800/1995
60. Al-Harthi MA. Impacto de las enzimas suplementarias para piensos, la mezcla de condimentos o su combinación en el rendimiento de los pollos de engorde, la digestibilidad de los nutrientes y los componentes plasmáticos. Int J Poult Sci. (2006) 5:764–71. doi: 10.3923/ijps.2006.764.771
61. El-Kelawy MI. Efecto de la forma del alimento, el diámetro de los gránulos y la suplementación enzimática en el rendimiento productivo y fisiológico de los pollos de engorde (tesis doctoral), Facultad de Agricultura de la Universidad de Damanhour, Damanhur, Egipto (2012).
62. Zanella I, Sakomura N, Pejerreyes F, Fiqueirdo A, Pack M. Efecto de la suplementación enzimática de las dietas de pollos de engorde a base de maíz y soja. Poult Sci. (1999) 78:561–8. doi: 10.1093/ps/78.4.561
Resumen de PubMed | Texto completo de CrossRef | Google Académico
63. Cowieson AJ, Acamovic T, Bedford MR. Suplementación de dietas que contienen harina de guisantes con enzimas exógenas: efectos sobre el aumento de peso, la conversión alimenticia, la digestibilidad de los nutrientes y la morfología bruta del tracto gastrointestinal de los pollos de engorde en crecimiento. Hno. Poult Sci. (2003) 44:427–37. doi: 10.1080/00071660310001598292
Resumen de PubMed | Texto completo de CrossRef | Google Académico
64. Ghazalah AA, Abd EI-Gawad AH, Soliman MS, Youssef A, W. Efecto de la preparación enzimática en el rendimiento de pollos de engorde alimentados con dietas a base de harina de maíz y soja. Egipto Poult Sci J. (2005) 25:295–316.
65. Shirmohammad F, Mehr M. Efectos de la suplementación dietética del complejo multienzimático sobre la utilización de energía en el gallo y el rendimiento de los pollos de engorde. Afr J Biotechnol. (2011) 10:7541–7. doi: 10.5897/AJB10.2260
66. Zhou X, Wang Y, Gu Q, Li W. Efecto del probiótico dietético, Bacillus coagulans, sobre el rendimiento del crecimiento, la composición química y la calidad de la carne del pollo amarillo de Guangxi. Poult Sci. (2010) 89:588–93. doi: 10.3382/ps.2009-00319
Resumen de PubMed | Texto completo de CrossRef | Google Académico
67. Al-Khalaifah H, Al-Nasser A, Ragheb G, Al-Qalaf S, Al-Omani N, Aneesh N, et al. El efecto de los probióticos y prebióticos dietéticos en el rendimiento de los pollos de engorde en Kuwait. Informe técnico n.º 2 (1 de noviembre – 31 de enero de 2017), Instituto de Investigación Científica de Kuwait (2017).
68. Akhtar M, Tariq AF, Awais MM, Iqbal Z, Muhammad F, Shahid M, et al. Estudios sobre el salvado de trigo Arabinoxylan por sus efectos inmunoestimulantes y protectores frente a la coccidiosis aviar. Carbohydr Polym. (2012) 90:333–9. doi: 10.1016/j.carbpol.2012.05.048
Resumen de PubMed | Texto completo de CrossRef | Google Académico
69. Cao Y, Xu Y, Auchoybur ML, Chen W, He S, Qin W, et al. ¿Cuál es el papel regulador de IKK? en la lesión por isquemia/reperfusión miocárdica mediante la determinación de la polarización M1 frente a M2 de los macrófagos. J Mol Cell Cardiol. (2018) 123:1–12. doi: 10.1016/j.yjmcc.2018.08.021
70. Cao L, Liu X, Qian T, Sun G, Guo Y, Chang F, et al. Actividad antitumoral e inmunomoduladora de los arabinoxilanos: un componente principal del salvado de trigo. Int J Biol Macromol. (2011) 48:160–4. doi: 10.1016/j.ijbiomac.2010.10.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
71. Korte J, Fröhlich T, Kohn M, Kaspers B, Arnold GJ, Härtle S. El análisis DIGE 2D de la bolsa de Fabricius revela perfiles proteómicos característicos para diferentes etapas del desarrollo de células B de pollo. Proteómica. (2013) 13:119–33. doi: 10.1002/pmic.201200177
Resumen de PubMed | Texto completo de CrossRef | Google Académico
72. Attia YA. Valor nutritivo de la harina de girasol sin descascarillar afectada por la suplementación multienzimática en las dietas de pollos de engorde. Archiv Geflugelk. (2003) 67:97–106.
73. Gao F, Jiang Y, Zhou GH, Han ZK. Los efectos de la suplementación con xilanasa sobre el crecimiento, la digestión, los niveles de hormonas y metabolitos circulantes, la inmunidad y la microflora intestinal en gallos alimentados con dietas a base de trigo. Hno. Poult Sci. (2007) 48:480–8. doi: 10.1080/00071660701477320
Resumen de PubMed | Texto completo de CrossRef | Google Académico
74. Basmacioglu Malayoglu H, Baysal S, Misirlioglu Z, Polat M, Yilmaz H, Turan N. Efectos del aceite esencial de orégano con o sin enzimas alimenticias sobre el rendimiento del crecimiento, las enzimas digestivas, la digestibilidad de los nutrientes, el metabolismo de los lípidos y la respuesta inmunitaria de pollos de engorde alimentados con dietas de harina de trigo y soja. Hno. Poult Sci. (2010) 51:67–80. doi: 10.1080/00071660903573702
Resumen de PubMed | Texto completo de CrossRef | Google Académico
75. Khaksar V, Golian A, Kermanshahi H. Respuesta inmunitaria y microflora ileal en pollos de engorde alimentados con una dieta a base de trigo con o sin enzima Endofeed W y suplementación con aceite esencial de tomillo o probiótico PrimaLac®. Afr J Biotechnol. (2012) 11:14716–23. doi: 10.5897/AJB12.1237
Resumen de PubMed | Texto completo de CrossRef | Google Académico
76. Head B, Bionaz M, Cherian G. La suplementación con enzimas de linaza y carbohidrasa altera las especies moleculares de ácidos grasos poliinsaturados n-3 hepáticos y la expresión de genes asociados con el metabolismo de los lípidos en pollos de engorde. Veterinaria Sci. (2019) 6:25. doi: 10.3390/vetsci6010025
Resumen de PubMed | Texto completo de CrossRef | Google Académico
77. Seidavi A, Dadashbeiki M, Alimohammadi-Saraei M, Hoven R, Payan-Carreira R, Laudadio V, et al. Efectos del nivel de inclusión dietética de una mezcla de cultivos probióticos y enzimas en la respuesta inmunitaria de los pollos de engorde. Environ Sci Pollut Res. (2017) 24:4637–44. doi: 10.1007/s11356-016-8206-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
78. Abdel-Hafeez HM, Saleh ESE, Tawfeek SS, Youssef IMI, Abdel-Daim ASA. Utilización de cáscaras de patata y pulpa de remolacha azucarera con y sin suplementación enzimática en dietas de pollos de engorde: efectos sobre el rendimiento, los índices bioquímicos séricos y las características de la canal. J Anim Physiol Anim Nutr. (2018) 102:56–66. doi: 10.1111/jpn.12656
Resumen de PubMed | Texto completo de CrossRef | Google Académico
79. El-Ghamry AA, Al-Harthi MA, Attia YA. Posibilidad de mejorar la utilización del pulido del arroz en dietas de pollos de engorde mediante enzimas o formulación dietética basada en aminoácidos digestibles. Archiv Geflügelk. (2005) 69:49–56.
80. Steenfeldt S. El efecto dietético de diferentes cultivares de trigo para pollos de engorde. Hno. Poult Sci. (2001) 42:595–609. doi: 10.1080/00071660120088416
Resumen de PubMed | Texto completo de CrossRef | Google Académico
81. Yamauchi K-E. Revisión sobre las alteraciones histológicas de las vellosidades intestinales de pollo relacionadas con la función intestinal. J Poult Sci. (2002) 39:229–42. doi: 10.2141/jpsa.39.229
82. Iji PA. El impacto de los polisacáridos sin almidón de los cereales en el desarrollo y la función intestinal en pollos de engorde. World Poult Sci J. (1999) 55:375–87. doi: 10.1079/WPS19990026
83. Thomas DV, Ravindran V. Efecto del tipo de cereal en el rendimiento, el desarrollo del tracto gastrointestinal y la morfología intestinal del pollito de pollo de engorde recién nacido. J Poult Sci. (2008) 45:46–50. doi: 10.2141/jpsa.45.46
84. Luskin MR, DeAngelo DJ. Leucemia linfoblástica aguda de células T: enfoque actual y direcciones futuras. Adv Cell Gene Ther. (2019) 2:E70. DOI: 10.1002/acg2.70
85. Maddu N, Raghavendra PB. Revisión de los efectos del litio sobre las células inmunitarias. Immunopharmacol Inmunotoxicol. (2015) 37:111–25. doi: 10.3109/08923973.2014.998369
Resumen de PubMed | Texto completo de CrossRef | Google Académico
86. Robert J, Sung M, Cohen N. Diferenciación in vitro de timocitos en larvas de Xenopus MHC clase I negativas. Dev Compar Immunol. (2001) 25:323–36. doi: 10.1016/S0145-305X(00)00066-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
87. Woyengo TA, Bogotá KJ, Noll SL, Wilson J. Mejora de la utilización de nutrientes de pollos de engorde a través de enzimas suplementarias. Poult Sci. (2018) 98:1302–9. doi: 10.3382/ps/pey452
Resumen de PubMed | Texto completo de CrossRef | Google Académico
88. Sateri S, Seidavi A, Bouyeh M, Neumann P, Kutzler M, Laudadio V, et al. Efecto de la harina de aceituna y las enzimas suplementarias sobre los rasgos de rendimiento, la bioquímica sanguínea, la respuesta inmunitaria humoral y la microbiota cecal de pollos de engorde. Sur Afr J Anim Sci. (2017) 47:804–12. doi: 10.4314/sajas.v47i6.8
89. Hashem NM, Soltan YA, El-Desoky NI, Morsi AS, Sallam SMA. Efectos de los extractos de Moringa oleifera y monensina sobre el rendimiento de conejos en crecimiento. Livest Sci. (2019) 228:136–43. doi: 10.1016/j.livsci.2019.08.012
Palabras clave: pollos de engorde, densidad de nutrientes, multienzimas, respuesta inmune, suplementación
Cita: Attia YA, Al-Khalaifah H, Abd El-Hamid HS, Al-Harthi MA y El-shafey AA (2020) Efecto de diferentes niveles de multienzimas en la respuesta inmunitaria, la hematología y bioquímica de la sangre, el estado de los antioxidantes y la histología de los órganos de los pollos de engorde alimentados con dietas estándar y de baja densidad. Frente. Vet. Sci. 6:510. doi: 10.3389/fvets.2019.00510
Recibido: 14 de septiembre de 2019; Aceptado: 23 de diciembre de 2019;
Publicado: 04 Febrero 2020.
Editado por:
Kyung-Woo Lee, Universidad de Konkuk, Corea del Sur
Revisado por:
Alireza Seidavi, Universidad Islámica Azad, Sucursal Rasht, Irán
Nesrein M. Hashem, Universidad de Alejandría, Egipto
Derechos de autor © 2020 Attia, Al-Khalaifah, Abd El-Hamid, Al-Harthi y El-shafey. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Y. A. Attia, yaattia@kau.edu.sa; H. Al-Khalaifah, hkhalifa@kisr.edu.kw
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía