
Efecto de la sincronicidad del aporte de aminoácidos en la síntesis de proteínas en miotubos C2C12 cultivados in vitro

Efecto de la sincronicidad del aporte de aminoácidos en la síntesis de proteínas en miotubos C2C12 cultivados in vitro
 Qiyu Zhang
Qiyu Zhang Mengmeng Mi
Mengmeng Mi Tianjio EXin FuNan Bao
Tianjio EXin FuNan Bao Li Pan
Li Pan Yuan Zhao*Guixin Qin*
Yuan Zhao*Guixin Qin*- Laboratorio Clave de Producción Animal, Calidad y Seguridad de Productos, Ministerio de Educación, Provincia de Jilin Laboratorio Clave de Nutrición Animal y Ciencia de Piensos, Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Jilin, Changchun, China
Estudios previos dedujeron que la tasa de síntesis/eficiencia de las proteínas en el tejido corporal probablemente se vea afectada por la sincronicidad del suministro de diferentes aminoácidos (AA) en su reserva metabólica. Con el fin de observar más a fondo la influencia de la sincronicidad del suministro de AA en la síntesis de proteínas a nivel celular, se realizó un experimento de cultivo celular in vitro con miotubos C2C12. Los miotubos C2C12 se cultivaron durante 24 h, mientras que el medio de cultivo se reemplazó cada 8 h. Esos miotubos se sometieron a 3 tratamientos (1 para control y 2 para ensayos), los miotubos control se cultivaron con el mismo medio completo normal durante las 24 h completas, y los 2 miotubos analizados se cultivaron con un medio de suministro de aminoácidos asíncrono en el que los niveles de diferentes AA (lisina, treonina, metionina, leucina, valina y ácido glutámico) aumentaron y luego disminuyeron o disminuyeron y luego aumentaron, en diferentes puntos de tiempo medios reemplazados (a las 0, 8 y 16 h). Sin embargo, durante todo el período del experimento, los 3 miotubos tratados recibieron la misma cantidad de cada AA. La muestra de los miotubos se utilizó para el análisis de la morfología de los miotubos, proteínas, AA y proteómica. Los resultados mostraron que la nutrición asincrónica de AA afecta la síntesis y degradación de las proteínas miotubas, y los AAAS en el medio aumentan, disminuyendo así la tasa de síntesis de proteínas miotubas (p < 0,05) y disminuyendo el diámetro de los miotubos (p < 0,05). El proceso de reducción de la síntesis de proteínas afecta a la vía de señalización PI3K-AKT y FoxO al regular a la baja los niveles de IRS1 y EGFR, y la amplitud de degradación es mayor que la amplitud de síntesis. Por lo tanto, este estudio reveló aún más el efecto del suministro asincrónico de aminoácidos en la síntesis de proteínas miotubulares y el mecanismo subyacente y proporcionó una referencia teórica para la precisión de la nutrición de los animales.
1 Antecedentes
De acuerdo con la teoría anterior de la nutrición animal, generalmente se cree que el hecho de que los nutrientes de la dieta puedan satisfacer las necesidades nutricionales de las actividades y la producción de la vida animal depende principalmente del tipo, la cantidad y la proporción de nutrientes efectivos proporcionados por la dieta (1). En cuanto a la nutrición proteica y aminoacídica de los cerdos, tanto la expresión de valor nutricional en el alimento como la expresión de las necesidades nutricionales de los cerdos se miden mediante la cantidad digerible al final del íleon (2, 3). Sin embargo, muchos estudios nacionales y extranjeros han demostrado que las dietas equilibradas formuladas con diferentes fuentes de proteínas y los mismos aminoácidos digeribles en el extremo del íleon no pueden obtener resultados estables, y el rendimiento del crecimiento y el efecto de la deposición de nitrógeno de los cerdos son muy diferentes (4, 5). Estudios posteriores han demostrado que es probable que la eficiencia de la deposición de nitrógeno esté relacionada con el equilibrio del tiempo de digestión de aminoácidos y la sincronización de diferentes tipos de aminoácidos en la dieta (6, 7).
El principal sitio de síntesis de aminoácidos para la deposición de proteínas es el músculo esquelético (8), que representa aproximadamente el 40% de la masa corporal total (9). La regulación de la masa muscular esquelética depende del equilibrio dinámico entre la síntesis y la degradación de proteínas de aminoácidos (10, 11), por lo que mejorar la eficiencia de la deposición de proteínas del músculo esquelético es crucial para la producción de carne de ganado (8). Algunos estudios han demostrado que la tasa de síntesis de proteínas de varios tejidos y órganos de los lechones se detectó cuando se alimentaban con dietas de aminoácidos desequilibradas, y los resultados mostraron que el crecimiento de los lechones estaba limitado por la disminución de la tasa de síntesis de proteínas del músculo esquelético (12, 13). Por lo tanto, observar la síntesis de proteínas de las células musculares es el objetivo principal para reflejar con precisión el estado de síntesis de proteínas del cuerpo. Si la síntesis de proteínas en el cuerpo está influenciada por el patrón de suministro de aminoácidos, entonces la sincronía y el equilibrio temporal del suministro de aminoácidos pueden afectar la síntesis de proteínas en las células musculares.
Muchos estudios que exploran la nutrición proteica en el músculo esquelético han utilizado la línea celular C2C12 como modelo in vitro (14, 15). Por lo tanto, este estudio verificó y reveló el mecanismo regulador de la sincronización de la liberación de aminoácidos que afecta la síntesis de proteínas a través de in vitro. En el grupo de control, se utilizó el medio normal como modo de suministro de aminoácidos sincrónico, y el miotubo C2C12 se cultivó mediante un medio personalizado en el grupo de suministro de aminoácidos asíncrono.
2 Materiales y métodos
2.1 Cultivo celular y diferenciación
Las células C2C12 del mioblasto de ratón se compraron a Shanghai EK-Bioscience Biotechnology Co., Ltd., se inocularon en frascos de cultivo celular y se cultivaron en medio DMEM/alto contenido de glucosa (icell, Shanghai, China). El medio se suplementó con 1% de penicilina-estreptomicina (cytiva, SV30010, Utah, América) y 10% de suero fetal bovino (Clark, FB25015, origen australiano) a 37 °C en una incubadora humidificada (HERA CELL 240i; Thermo, EE.UU.) con 5% de CO2. Después de que las células habían proliferado hasta el 80%, el medio se cambió a un medio de diferenciación [DMEM que contenía un 2% de suero de caballo (Cytiva, SH30074, EE. UU.) y una solución de penicilina-estreptomicina al 1%] durante 5 días. El medio se cambió cada dos días después de dos lavados con PBS (cytiva, SH30256.01, EE. UU.) para promover la maduración de los miotubos multinucleados maduros.
2.2 Esquema de tratamiento
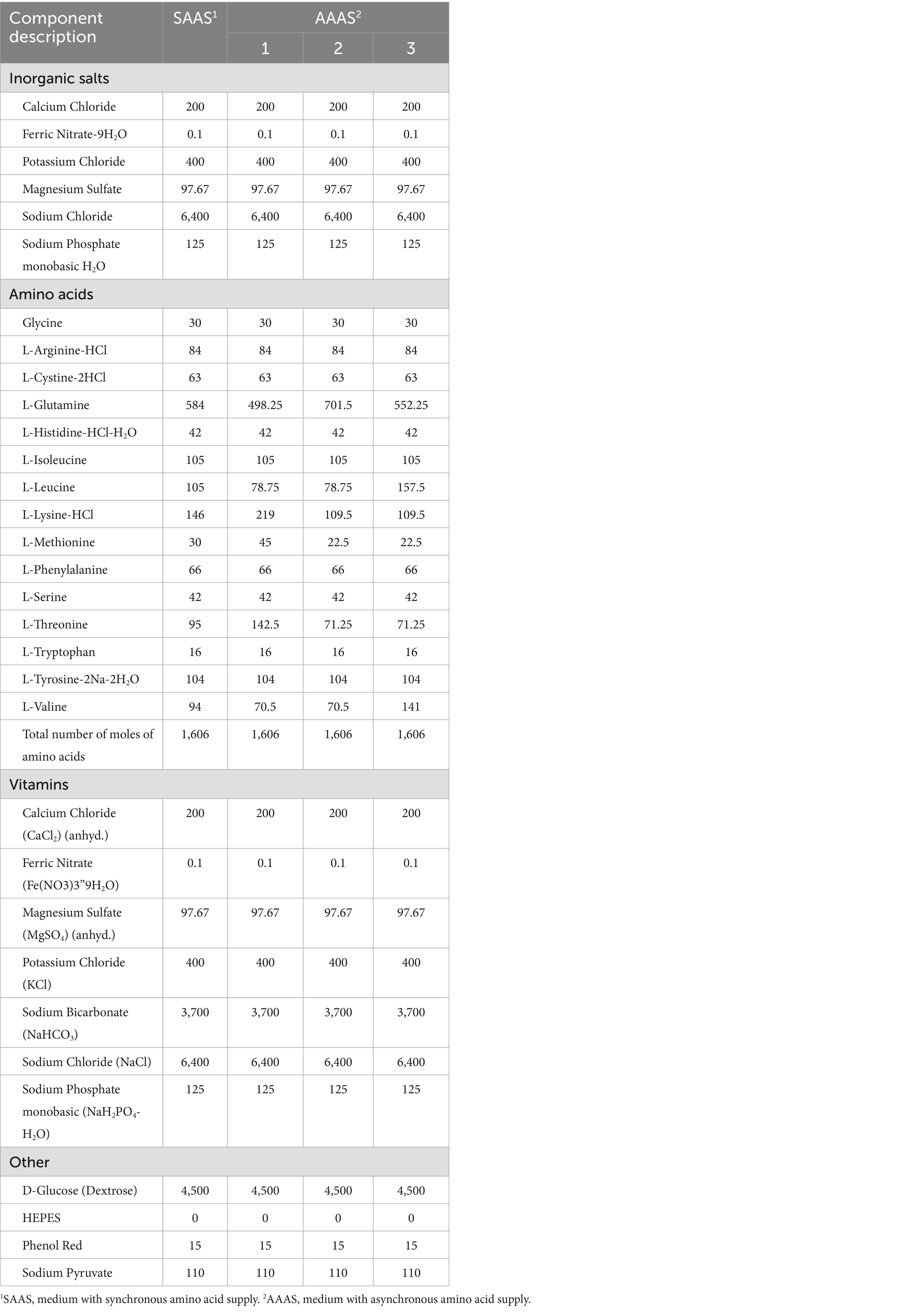
Los miotubos totalmente diferenciados se trataron con dos medios con diferentes concentraciones de AA durante 24 h, y el medio se cambió cada 8 h. El suministro sincronizado de aminoácidos (SAAS) se cultivó con medio normal, y el suministro asíncrono de aminoácidos (AAAS) se cultivó con medio personalizado. La cantidad de cada adición fue de 2 mL. El orden de adición media fue 1-2-3 en el primer grupo y 2-1-3 en el segundo grupo para el patrón AAAS. De acuerdo con la característica de que la concentración de aminoácidos en la sangre de la vena porta de los cerdos aumentó primero y luego disminuyó después de comer diferentes dietas de fuentes de proteínas con diferentes concentraciones de aminoácidos libres in vitro (16), se diseñó la fórmula de suministro de medio celular con suministro de aminoácidos no sincronizado. Los AA seleccionados para el ajuste se consideraron principalmente de la siguiente manera: uno es el AA limitante relacionado con la síntesis de proteínas musculares (17), y el otro es el AA de cadena ramificada, que es la principal fuente de energía del metabolismo del nitrógeno en el músculo esquelético (18). Para preparar el medio con el suministro de aminoácidos asíncronos, se ajustó la secuencia en la que la concentración de lisina, treonina y metionina aumentó primero y luego disminuyó, y también se ajustó la secuencia en la que la leucina y la valina cayeron primero y luego aumentaron. La concentración de glutamato se utilizó para equilibrar el número molar total de AA en cada período, que fue consistente con el del grupo control. La fórmula específica se muestra en la Tabla 1.
2.3 Determinación de la proteína miotuba
Después de 5 días de diferenciación de miocitos y la formación de miotubos maduros, los miotubos no tratados se colocaron como blancos. Se recogieron miotubos del grupo SAAS y del grupo AAAS después del tratamiento durante 24 h. Las células se lisaron con un kit de extracción de proteínas totales (Bestbio, BB-3101, Shanghái, China) durante 30 min en hielo. Los lisados de células enteras se centrifugaron a 12.000 × g durante 15 min a 4 °C, y el sobrenadante se transfirió a un nuevo tubo. La concentración de proteínas de cada muestra se cuantificó utilizando el kit de ensayo de proteínas BCA (Shanghai Epyzime Biotechnology Co., Ltd., ZJ102L, Shanghai, China). El contenido de proteínas de los grupos SAAS y AAAS se obtuvo restando el espacio en blanco.
2.4 Determinación del contenido de urea en el sobrenadante de los miotubos
Los miotubos maduros se trataron durante 0, 8, 16 y 24 h, y se colectó el sobrenadante del medio de cultivo. El contenido de urea en las muestras en cada punto de tiempo se determinó mediante un kit de ensayo de contenido de nitrógeno ureico (Boxbio, AKM002M, Beijing, China), y la suma se utilizó como el contenido total de urea para el análisis estadístico. De acuerdo con los resultados de la determinación de proteínas, se seleccionó el grupo AAAS1 con menor proteína de los miotubos y mayor nitrógeno ureico del medio para detectar los siguientes ítems.
2.5 Determinación de AA en el sobrenadante miotubado
Durante el período de tratamiento, las muestras se recolectaron cada ocho horas y se analizó el sobrenadante del medio de miotubos inicial de la siguiente manera.
Paso 1: Se realizó el pretratamiento de la muestra según lo descrito por Dai et al. (19). El contenido de AA libre en suero se determinó mediante cromatografía líquida de alta resolución (HPLC). Pretratamiento de la muestra: Las muestras se extrajeron a -80 °C y una alícuota de 200 μL de las muestras se colocó en un tubo de centrífuga de 1,5 mL después de la fusión. A continuación, se añadieron 200 μ L de 1,5 mol/L de HCIO4, se agitaron, se mezclaron y se dejaron reposar a 4°C durante 30 min. Para ajustar el pH de la solución, se añadieron 200 μ L de 2 mol/L de K2CO3, se agitaron y se mezclaron bien. Las muestras se centrifugaron a 10000 r/min durante 2 min, se colocaron 100 μL de sobrenadante en un tubo centrífugo de 1,5 mL y se añadieron 100 μL de ácido benzoico al 1,2% y 1.400 μ L de agua ultrapura. Las soluciones se mezclaron y se transfirieron a una botella de muestreo automático UPLC para su análisis.
Paso 2: Determinación de la muestra Se utilizó un sistema ACQUITY UPLC FLR de Waters (Waters, Milford, MA, EE. UU.) y una columna AcQUITY UPLC (2,1 mm x 100 mm, P/N:186002352) con los siguientes parámetros. Se añadió fase A móvil (acetato de sodio 0,1 M, pH 7,2), 27,3 g de acetato de sodio trihidrato a 1,6 L H2O en un frasco de vidrio, seguido de la adición secuencial de 96 μL de HCl 6 M, 180 mL de metanol y 10 mL de tetrahidrofurano. Después de que la mezcla se mezcló uniformemente, se agregó H2O y el volumen final de la solución fue de 2 L. Fase B móvil: 100% metanol, botella de vidrio marrón. Caudal, 0,3 mL/min; volumen de inyección, 1 μL; temperatura de la columna, 35°C; temperatura de la muestra, 5°C; longitud de onda de detección, 450 nm; tasa de recaudación, 20 puntos/s; constante de tiempo, 0,1 s; y duración, 19 min.
Calculando el porcentaje del contenido de aminoácidos remanentes de cada aminoácido en el medio inicial cada 8 h, se obtuvo la sincronización de la absorción de aminoácidos en cada punto de tiempo. Las curvas cinéticas de absorción de un solo AA se trazaron utilizando el software Origin 9.1 (EE. UU.). El grado de sincronización de AA se cuantificó según el método de Wang et al. (6). Se calculó la desviación estándar de la liberación de AA (%) del monómero en cada punto de tiempo, que se utilizó como índice de liberación de AA de forma sincrónica. El índice de sincronización (SI) de cada medio es la suma del índice de sincronización de cada punto de tiempo. Cuanto menor sea el SI, mejor será la sincronización.
2.6 Mediciones no radiactivas de la síntesis de proteínas con SUnSET
La síntesis de proteínas se midió utilizando la tecnología de detección de superficie traslacional (SUnSET) (20). En el nivel de dosis inferior, la puromicina se une a la proteína recién sintetizada, reflejando directamente la tasa de traducción del ARNm in vitro (21, 22). Las células C2C12 se cultivaron como se describió anteriormente y se incubaron con 1 μg/mL de puromicina (23) durante 30 min. Después de que los miotubos se recolectaron en lisado de células heladas, los cambios en la incorporación de puromicina se detectaron mediante Western blot (24).
2.7 Tinción con inmunofluorescencia
El método de ensayo se basó en el método de Caldow (25). Los cultivos se retiraron por succión y las células se lavaron (2 × 5 min) con solución salina tamponada con fosfato (PBS) y se fijaron en paraformaldehído al 4% (Beijing Dingguo Changsheng Biotechnology Co. LTD, AR-0211, Beijing, China) durante 15 min. A continuación, las células se lavaron con PBS (3 × 5 min), se infiltraron con PBS que contenía 0,1% de Triton X-100 (Coolaber, DZSL1466-100, Pekín, China) durante 20 min, se lavaron con PBS (3 × 5 min) y se incubaron durante 2 h en PBS que contenía un 3% de albúmina V bovina (Biotopped, A6020, Pekín, China). A continuación, las células se lavaron con PBS (3 × 5 min), se incubaron con el anticuerpo primario MYH (1:300, sc-376157, Santa Cruz, Dallas TX, EE.UU.) durante la noche a 4 °C, se lavaron con PBS (3 × 5 min) y se incubaron con el anticuerpo secundario anti-conejo de cabra YF488 (1:500, Y6105L, UElandy, Suzhou, China) durante 2 h a temperatura ambiente en la oscuridad. A continuación, las células se lavaron con PBS (3 × 5 min) y se incubaron con una solución de tinción 1× DAPI (tipo readyuse) (SL7100, Coolaber, Pekín, China) durante 5 min a temperatura ambiente en la oscuridad para teñir los núcleos. Finalmente, las células se lavaron nuevamente con PBS (3 × 5 min). Los miotubos se fotografiaron mediante escaneo con un microscopio confocal láser (Leica SP8, Alemania).
2.8 Medición del diámetro del miotubo
Los miotubos se fotografiaron con un microscopio confocal láser y sus diámetros se midieron con el software Leica. El diámetro medio de los miotubos se calculó midiendo el diámetro máximo de cada uno. Se tomaron diez bloques de cultivo aleatorios para cada muestra y se contaron al menos 100 miotubos.
2.9 Análisis proteómico de las vías de síntesis y degradación de proteínas en miotubos
2.9.1 Extracción de proteínas e hidrólisis enzimática peptídica
Se añadió una cantidad adecuada de lisado de SDT (4% (p/v) de SDS, 100 mM de Tris/HCl, pH 7,6) a todas las muestras para la extracción de proteínas, seguido de la cuantificación de proteínas mediante ensayo de BCA. Se añadieron veinte microgramos de proteína de cada muestra a una cantidad adecuada de tampón de carga 5X, se sometieron a SDS-PAGE (gel de gradiente preformado al 4-20%, presión constante 180 V, 45 min) en un baño de agua hirviendo durante 5 min, y se tiñeron con azul Coomassie Brilliant R-250. El nivel de expresión de la proteína en las muestras fue cuantificado por Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China). Una cantidad apropiada de proteína de cada muestra se tripsinió mediante el método de preparación del proteoma asistido por filtro (FASP) (26), y el péptido se desalinizó mediante un cartucho de C18. El péptido se liofilizó y se volvió a disolver añadiendo 40 μL de solución de ácido fórmico al 0,1%. Se realizó la cuantificación peptídica (OD280).
2.9.2 Adquisición de datos LC–MS/MS
Cada muestra se separó utilizando un sistema de fase líquida de HPLC NanoElute con un caudal de nanolitros. El tampón A fue una solución acuosa de ácido fórmico al 0,1%, y el tampón B fue una solución acuosa de acetonitrilo de ácido fórmico al 0,1% (acetonitrilo 99,9%). La columna se equilibró con un 95 % de A líquido y las muestras se separaron en una columna analítica de fase reversa C18 (columna Thermo Scientific EASY, 25 cm, ID 75 μm, 1,9 μm) a un caudal de 300 nL/min.
Las muestras se separaron por cromatografía y se analizaron por espectrometría de masas utilizando un espectrómetro de masas timsTOF Pro. El voltaje de la fuente de iones se fijó en 1,5 kV, la MS y la MSMS se detectaron y analizaron mediante TOF, y el rango de exploración del espectro de la MS se fijó en 100-1700 m/z. El modo de adquisición de datos fue el modo de fragmentación en serie acumulativa paralela (PASEF). Los parámetros específicos de adquisición de datos fueron los siguientes: la movilidad iónica (1/K0) fue de 0,6-1,6 Vs/cm2, y un espectro de masas primario correspondió a 10 espectros secundarios del modo PASEF. El tiempo de exclusión dinámica para la exploración MS en tándem se fijó en 24 s para evitar exploraciones repetidas de iones padres.
2.9.3 Identificación de proteínas y análisis cuantitativo
Los datos originales del análisis de espectrometría de masas se muestran en file.d. Se utilizó el software MaxQuant (versión 1.6.14) (27) para la identificación de bibliotecas y el análisis cuantitativo. Análisis estadístico de los resultados proteómicos. En diferentes proteínas, se tomó como estándar el cociente de expresión (cambio de pliegue, FC) >2.0 (aumento mayor de 2.0 veces o disminución menor de 0.5 veces) y p < 0.05 (prueba T u otra).
2.9.4 Bioinformática y análisis estadístico
En primer lugar, se normalizó la información cuantitativa del conjunto de proteínas diana (normalizada al intervalo (−1,1)). A continuación, se utilizó el paquete ComplexheatmapR (RVersion3.4) para clasificar simultáneamente las dimensiones de expresión de la muestra y de la proteína (algoritmo de distancia: Euclidean, método de conexión: Averagelinkage) y generar mapas de calor de agrupamiento jerárquico.
En segundo lugar, se utilizó Blast2GO (número de versión: BLASTP 2.8.0+) para realizar la anotación de GO con el conjunto de proteínas objetivo. El proceso se puede resumir a grandes rasgos en los siguientes pasos: alineación de secuencia (Blast), extracción de elementos GO (Mapping), anotación GO y anotación complementaria InterProScan (aumento de anotaciones). Se utilizó el software KOBAS (versión: KOBAS3.0) para anotar la vía KEGG del conjunto de proteínas objetivo. El conjunto de proteínas objetivo se comparó con la distribución de cada clasificación de GO (o vía KEGG, o dominio) en el conjunto de proteínas objetivo y el conjunto general de proteínas mediante la prueba exacta de Fisher (prueba exacta de Fisher), y el análisis de enriquecimiento de las anotaciones de GO o (o vía KEGG, o dominio) se realizó en el conjunto de proteínas objetivo. Sobre la base de IntAct1 o STRING,2 la información de la base de datos se utilizó para encontrar las interacciones directas e indirectas entre las proteínas diana. Se generó y analizó la red de interacción.
2.10 Western blot
Los miotubos tratados se escindieron en hielo con un kit de extracción de proteínas totales (Bestbio, BB-3101, Shanghái, China) durante 30 min. El lisado de toda la célula se centrifugó a 12000 × g a 4 °C durante 15 min, y el sobrenadante se transfirió a los nuevos tubos. La concentración de proteína de cada muestra se cuantificó mediante un kit de ensayo de proteínas BCA (Shanghai Epyzime Biotechnology Co., Ltd., ZJ102L, Shanghai, China).
Se cargaron cantidades iguales de proteínas de diferentes muestras en geles de poliacrilamida al 7,5, 10% o 12,5% SDS (Shanghai Epyzime Biotechnology Co., Ltd., PG113, PG114, PG112, Shanghai, China) y se separaron por electroforesis. Las proteínas se transfirieron a membranas de PVDF y posteriormente se bloquearon con V albúmina bovina al 5% durante 1 h a temperatura ambiente. Las membranas se incubaron con diferentes anticuerpos primarios contra p-4E-BP1 (1:100, Santa Cruz, sc-293124, Dallas, EE. UU.), p-mTOR (1:1000, Abcam, AP0115, Cambridge, Reino Unido), IRS1 (1:100, Santa Cruz, sc-8038, Dallas, EE. UU.), EGFR (1:100, Santa Cruz, sc-373746, Dallas, EE. UU.), PDGFR (1:100, Santa Cruz, sc-80991, Dallas, EE. UU.), p-ERK (1:100, Santa Cruz, sc-7383, Dallas, EE. UU.), p-Akt (1:500, Cell Signaling, 4.060 T, Boston, EE. UU.), FOXO1 (1:1500, GeneTex, GTX135251, Irvine, California, EE. UU.), ubiquitina (1:100, Santa Cruz, sc-8017, Dallas, EE. UU.), puromicina (1:5000, Sigma, MABE343, Darmstadt, Alemania) y β-actina (1:100000, ABclonal, AC026, Cambridge, Reino Unido). A continuación, las membranas se lavaron tres veces con solución salina Tween 20 (TBST) tamponada con Tris (iScience, CP17207M, Jiangsu, China) y se incubaron con el anticuerpo secundario anti-ratón o anti-conejo correspondiente (1:5000, SA00001-2, SA00001-1, Proteintech Group, Shanghai, China) durante 1 h a 37 °C. Finalmente, las membranas se lavaron tres veces con TBST y se desarrollaron con un kit de quimioluminiscencia ECL hipersensible (NCM Biotech, P10200, Suzhou, China) y un sistema de detección de Western blot (Kodak, EE. UU.). La intensidad de las bandas se cuantificó mediante el software ImageJ (National Institutes of Health, EE.UU.) y la expresión relativa de la proteína diana se normalizó a la de la β-actina.
2.11 Análisis estadístico
La secuenciación del proteoma se realizó en 4 réplicas y todas las demás pruebas se realizaron en 3 réplicas. Los datos se clasificaron a través de Excel. Se utilizó el software SPSS 23.0 para analizar la significancia de un solo factor (ANOVA) de los datos, p < 0,05. 0,05 fue significativo. Trazar con GraphPad Prism 10.0.2. Además de los resultados estadísticos indicados de otra manera, se proporcionan otros resultados para facilitar la visualización y normalizar los datos al contenido relativo del grupo de control SAAS.
3 Resultados
3.1 Efecto del patrón de suministro asíncrono de AA sobre la deposición de proteínas en miotubos
El contenido total de proteínas de los miotubos en el grupo SAAS fue significativamente mayor que el del grupo AAAS cuando los miotubos se cultivaron durante 24 h en el patrón de suministro de aminoácidos asíncrono (p < 0,05, Figura 1A). Sin embargo, la producción de nitrógeno ureico en el grupo SAAS fue significativamente menor que en el grupo AAAS (p < 0.05, Figura 1B). De acuerdo con los resultados de la deposición de proteínas, se seleccionó AAAS1 como representante del patrón de suministro de aminoácidos asíncrono (p < 0.05, Figura 1C). El contenido de proteínas recién generadas en los miotubos en el grupo SAAS fue significativamente mayor que en el grupo AAAS (p < 0,05, Figura 1D).
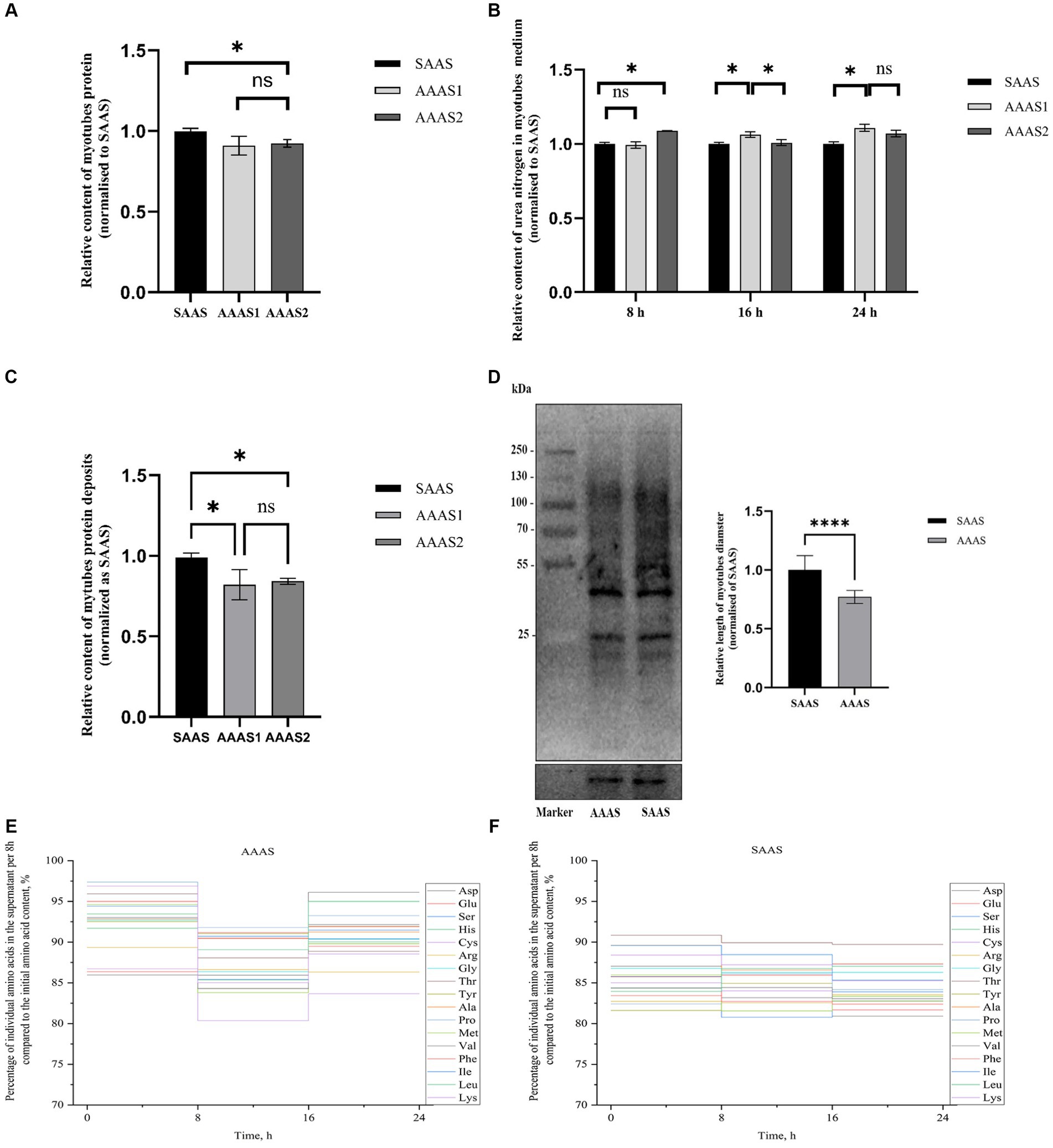
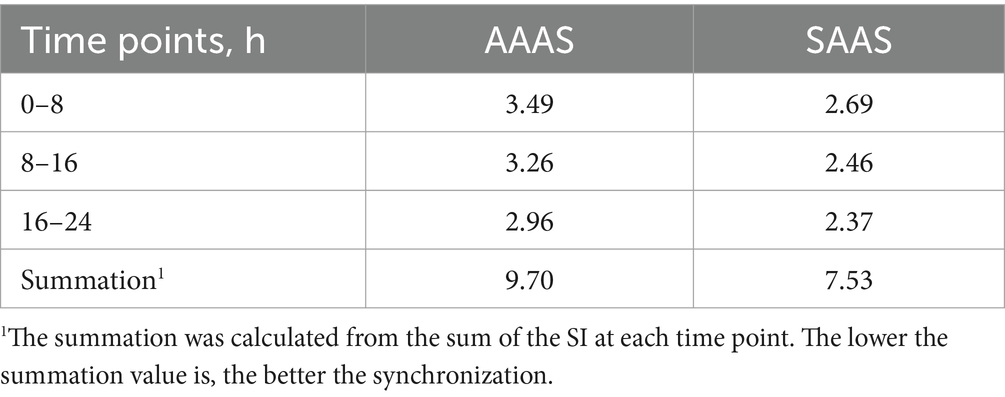
Para 17 AA individuales, se trazó una curva escalonada vertical de liberación de AA cada 8 h, que se expresó como porcentaje de la concentración final, para indicar la sincronización de la absorción de AA. En comparación con el medio con SAAS, la sincronicidad AA del medio con AAAS fue pobre en cada tiempo (Figura 1E). Además, en la Tabla 2 se muestra el índice de sincronización de AA cada 8 h. Las proporciones de medios AAAS y SAAS fueron de 9,70 y 7,53%, respectivamente. El SI del medio AAAS es mayor que el del medio SAAS.
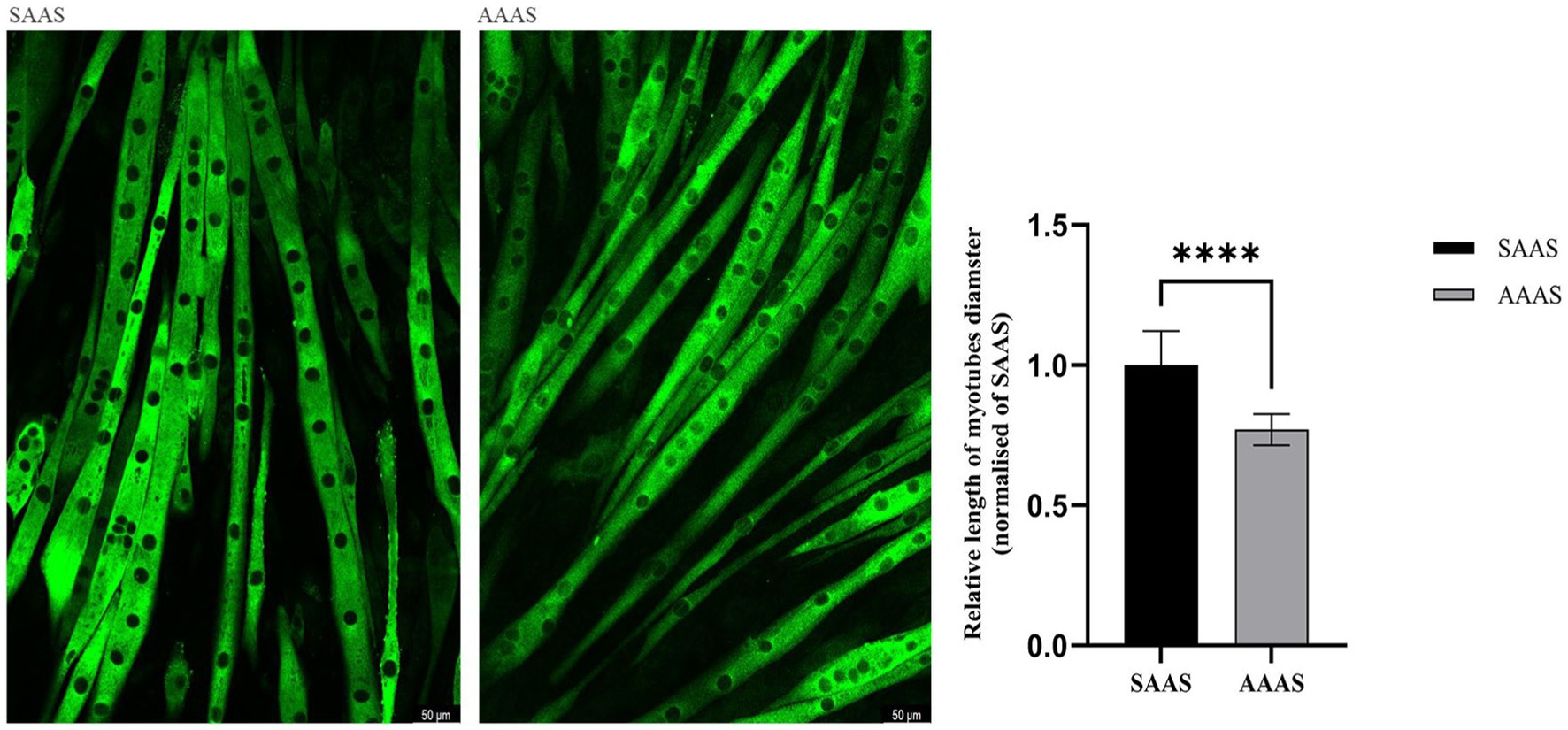
3.2 Efectos del patrón de suministro de AA en el tamaño de los miotubos C2C12
El suministro de aminoácidos no convencionales (24 h) en DMEM resultó en un crecimiento lento de los miotubos C2C12. En comparación con el SAAS, el medio de diferenciación con AAAS presentó una disminución del diámetro del miotubo (p < 0,05; Figura 2).
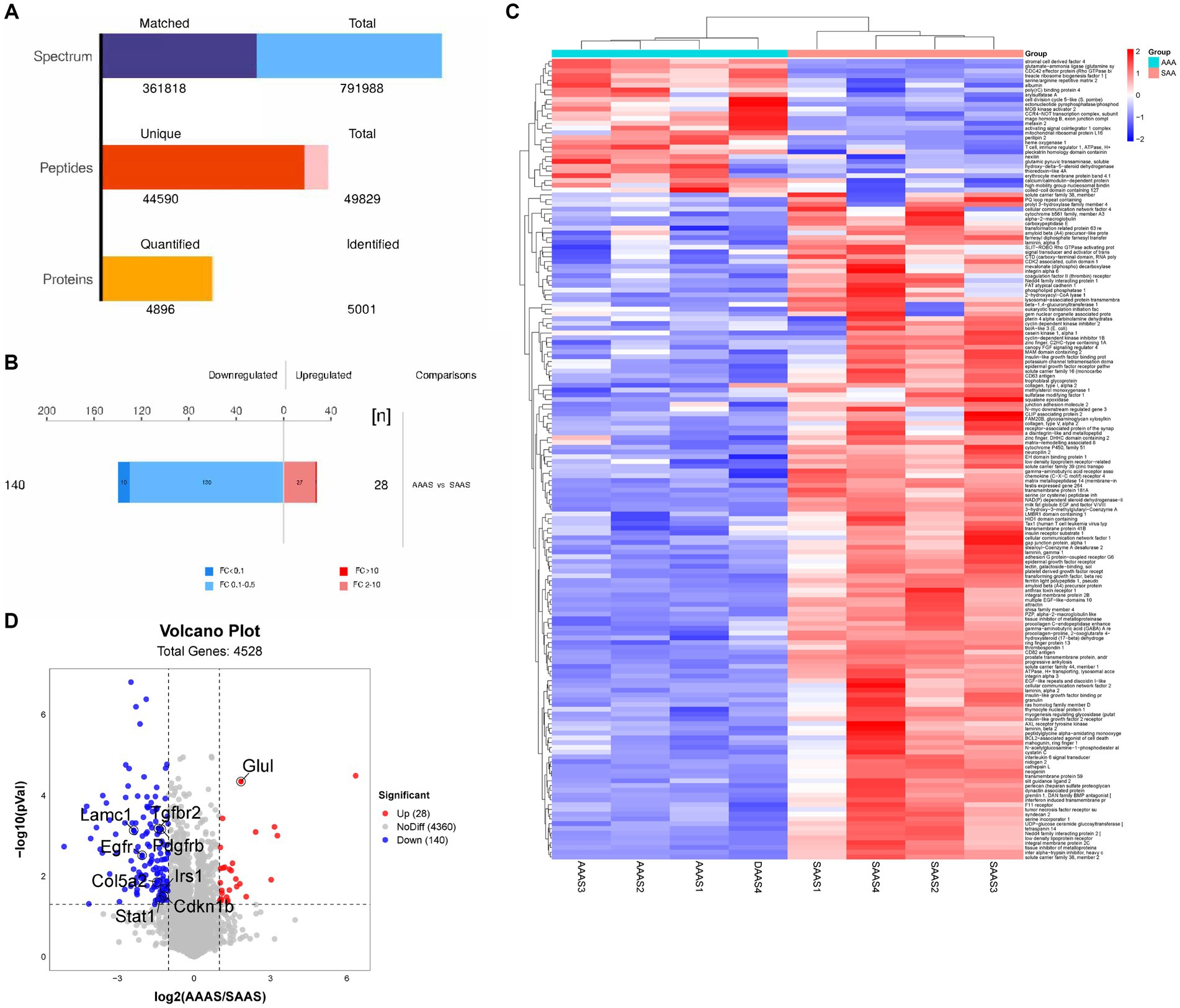
3.3 Identificación y cuantificación de proteínas miotubas
Se analizaron dos muestras de miotubos de los grupos AAAS y SAAS mediante LC-MS/MS, y se identificaron 50.193 péptidos únicos y 5.004 proteínas (Figura 3A). Se identificaron 168 proteínas expresadas diferencialmente, de las cuales 28 proteínas estaban significativamente reguladas al alza y 140 proteínas estaban significativamente reguladas a la baja (Figura 3B). La visualización del mapa de calor muestra las 100 proteínas principales afectadas por el patrón de suministro de aminoácidos asíncrono. En comparación con el grupo SAAS, el patrón de suministro asincrónico afectó a tres grupos de proteínas asociados con la síntesis y degradación de proteínas (Figura 3C). En comparación con SAAS, el patrón de suministro asíncrono de AA afecta a las vías de síntesis y degradación de proteínas al regular a la baja los niveles de proteínas de EGFR, PDGFRB, IRS1 y Col5a2 (Figura 3D).
3.4 Anotación funcional de proteínas expresadas diferencialmente
Para investigar mejor la importancia del enriquecimiento de vías para las proteínas diferenciales, se realizó un análisis de enriquecimiento de las vías KEGG en proteínas expresadas diferencialmente reguladas al alza y a la baja, que se muestran en forma de diagrama de mariposa. Los resultados se describen a continuación (Figura 4A). Las proteínas diferencialmente expresadas al alza se enriquecieron significativamente en la biosíntesis de arginina y en las vías metabólicas de alanina, aspartato y glutamato. Las proteínas expresadas diferencialmente reguladas a la baja se enriquecieron significativamente en las interacciones entre el receptor de ECM, la biosíntesis de esteroides, la vía de señalización PI3K-Akt, la vía de señalización FoxO y otras vías.
Por lo tanto, analizamos más a fondo las proteínas diferenciales EGFR, PDGFRB e IRS1 en la vía de señalización PI3K-Akt/FoxO para la síntesis y degradación de proteínas (Figuras 4B, C). Para verificar la expresión de las proteínas anteriores, se realizó un análisis de Western blot y los resultados mostraron que el patrón AAAS disminuyó significativamente la expresión de las proteínas IRS1, EGFR y PDGFRB (p < 0.05) (Figura 4D). Además, para estudiar los efectos del patrón AAAS en la síntesis de proteínas y la degradación de otras proteínas clave, se realizó un análisis cuantitativo de Akt y mTOR aguas abajo y sus proteínas fosforiladas en la vía PI3K-Akt. Los resultados mostraron que, aunque el patrón de suministro de AA no tuvo ningún efecto sobre el nivel de expresión de la proteína mTOR, el SAAS aumentó significativamente los niveles de expresión de las proteínas p-Akt y p-mTOR (p < 0,05). Además, analizamos cuantitativamente la proteína clave ERK y su proteína fosforilada en la vía de señalización FoxO. Los resultados también mostraron que el patrón de suministro de AA no tuvo ningún efecto sobre el nivel de expresión de la proteína ERK, pero el AAAS aumentó significativamente el nivel de expresión de la proteína p-ERK (p < 0,05). Los estudios han demostrado que el patrón AAAS regula la síntesis y degradación de proteínas a través de proteínas mediadas por IRS-1 relacionadas con la vía de señalización PI3K-AKT/FoxO.
4 Discusión
Los estudios de cinética digestiva han demostrado que los AA en la dieta son retenidos de forma asincrónica por los tejidos intestinales y hepáticos (28), lo que conduce a un desequilibrio en la entrada de aminoácidos a las células, lo que afecta la coordinación del suministro de AA en los sitios de síntesis de proteínas en el cuerpo (29). Los resultados se caracterizaron por cambios significativos en la síntesis de proteínas del músculo esquelético (30). Por lo tanto, este estudio revela la producción y pérdida de proteínas durante la síntesis y el metabolismo de los miotubos C2C12 mediante el establecimiento de un patrón AAAS in vitro.
La homeostasis del músculo esquelético se mantiene a través de una relación equilibrada entre la síntesis y la degradación de proteínas. Encontramos que las dietas de liberación de aminoácidos desincronizadas afectan la deposición de nitrógeno en el cuerpo (6, 7), similar a los resultados obtenidos en este estudio; por lo tanto, el patrón AAAS podría reducir la eficiencia de la deposición de proteínas en los miotubos, lo que conduce a la atrofia de los miotubos. Esto puede deberse a la respuesta sensible de los mioblastos a la regulación de diferentes concentraciones de nutrientes (31) y a que los aminoácidos son la principal fuente de sustrato para la síntesis de proteínas en el músculo esquelético; Además, la concentración equilibrada de aminoácidos desempeña un papel crucial en la síntesis de proteínas musculares y el metabolismo del cuerpo (32). Por lo tanto, cuando la tasa de degradación de la proteína muscular supera su tasa de síntesis, se produce la atrofia muscular.
Los aminoácidos son el principal reservorio de nitrógeno celular (33, 34). En este estudio, revelamos los cambios en AA en el medio bajo el patrón AAAS, y el contenido residual de glutamato en el medio del grupo SAAS fue significativamente menor que el del grupo AAAS en un 6%. Esto puede deberse a que el glutamato se utiliza como fuente de nitrógeno durante la síntesis de aminoácidos para mantener el proceso de biosíntesis; Sin embargo, cuando no se utiliza para la biosíntesis, el glutamato se convierte en amonio libre a través del ciclo de la glutamina sintasa, y el amoníaco restante se convierte en urea (35). Además, el índice asincrónico del grupo AAAS fue el más alto, lo que indica que la síntesis y el metabolismo de los conductos musculares estaban en un estado desequilibrado. Se especula que esta es la razón por la que el modo AAAS reduce la síntesis de proteínas y aumenta los niveles de urea.
Los resultados de este estudio demostraron que con el suministro asincrónico de aminoácidos a lo largo del tiempo, el contenido de proteínas recién formadas disminuyó y el contenido de nitrógeno ureico aumentó, lo que indica que la tasa de síntesis de proteínas fue menor que la tasa de descomposición. A través del análisis de enriquecimiento de la vía KEGG, la transducción de señales de proteínas diferenciales en el nivel 2 pertenece principalmente a la vía de señalización PI3K-Akt, la vía de señalización FoxO, la vía de señalización del calcio y la vía de señalización Rap1. Estudios previos han demostrado que la vía PI3K-Akt es una vía clave de transducción de señales involucrada en la regulación de la proliferación, adhesión y diferenciación celular (36, 37). IRS1 es el representante más importante de la familia de proteínas IRS y un factor clave en la vía de señalización PI3K-Akt. Durante el crecimiento muscular, IRS1 es un medio clave de transducción de señales PI3K-Akt que regula el crecimiento muscular y el metabolismo (38-40). Los resultados de este experimento mostraron que los niveles de expresión de IRS1, PDGFR y EGFR se redujeron significativamente por el modo de suministro de aminoácidos asíncrono durante el proceso de síntesis de proteínas. IRS1 es un intermediario importante en la vía de señalización de la insulina, así como un nodo clave en la vía PI3K-AKT, y promueve la generación de músculo en el músculo esquelético (35). EGFR es un receptor transmembrana aguas abajo de IRS1, y su mecanismo de acción es a través de la cascada de señalización de iniciación de dímero inducida por ligando (41). Puede activar múltiples efectores aguas abajo (42), y el factor de crecimiento transformante aguas abajo (TGF) puede activar EGFR para promover la fosforilación de cascadas de señales, y también desempeña un papel clave en la comunicación de PI3K-AKT y otras vías entre células animales (43). Por lo tanto, el patrón de suministro no sincrónico de aminoácidos afecta la reducción de la síntesis de proteínas en los conductos musculares, que pueden estar regulados por la vía de señalización PI3K-Akt mediada por IRS1, incluida la síntesis de proteínas, la absorción de glucosa, la progresión del ciclo celular y otros procesos. Por lo tanto, después de confirmar que el patrón asíncrono de suministro de aminoácidos en el sitio de síntesis de proteínas (miotubo C2C12) reduce la síntesis de proteínas, se encuentra que esto se logra afectando significativamente la vía de señalización PI3K-Akt y regulando significativamente a la baja los niveles de expresión de las proteínas EGFR y PDGFRB durante la vía de señalización.
Además, la proteína IRS1 no solo aumenta la síntesis de proteínas al mediar la vía PI3K, sino que también bloquea el proceso de degradación de proteínas (44, 45), afectando así a la hipertrofia de los miotubos (46, 47). Los estudios han demostrado que la inhibición de la degradación de proteínas se logra a través de la inhibición mediada por IRS1 de la familia de factores de transcripción Foxo (44, 45). El FoxO se activa por señales ascendentes en condiciones de bajo suministro de nutrientes o inanición. Por ejemplo, en el hígado, un estado nutricional bajo y niveles bajos de señalización de insulina activan FoxOs, restaurando los niveles de glucosa a través de la glucogenólisis y el gluconógeno (47). Nuestros resultados también muestran que el patrón de suministro de aminoácidos asíncrono afecta significativamente la expresión de la proteína IRS1 y la actividad de la vía de señalización FoxO aguas abajo de RTK.
Con el fin de explorar más a fondo los efectos de la síntesis y degradación de proteínas en las vías de señalización PI3K-Akt y FoxO, verificamos cuantitativamente los niveles y los niveles de fosforilación de proteínas clave en las vías. En las células del músculo esquelético de los mamíferos, la expresión de la proteína Akt puede conducir a la hipertrofia muscular (48). Akt es una señal descendente activada por PI3K, y Akt y PI3K juntas juegan un papel en la hipertrofia muscular (49). Por lo tanto, el Western blot se utiliza para detectar una determinada proteína en una muestra de sarcotubulina de acuerdo con la unión específica del antígeno al anticuerpo. Los resultados de la inmunotransferencia mostraron cambios significativos en la fosforilación de Akt y ningún cambio en Akt. Esto sugiere que el modo de suministro de aminoácidos asíncrono induce la fosforilación de Akt a través de la vía PI3K. La transducción de señales PI3K-Akt: la transducción de señales de rapamicina (mTOR) en mamíferos es una vía central que controla el crecimiento, la proliferación y el metabolismo celular (50, 51). En este estudio, se validaron aún más mTOR y mTOR fosforilado y se encontró que el modo de suministro de aminoácidos asíncrono disminuyó el nivel de proteína mTOR fosforilada pero no cambió el nivel de proteína mTOR. Se plantea la hipótesis de que la razón se debe a la activación de Akt, que fosforila una variedad de sustratos, incluidas las proteínas que regulan la síntesis de proteínas y la transcripción génica de la proliferación celular. Se ha informado que la vía MAPK desempeña un papel importante en el crecimiento de los miotubos y, en particular, la fosforilación de ERK es esencial para la proliferación de los miotubos (52). En este estudio, el análisis mostró que después del suministro asincrónico de aminoácidos, el agua fosforilada de ERK se incrementó significativamente a través de la reducción de la proteína EGFR, lo que promovió la actividad de la vía de señalización FoxO para regular la descomposición de proteínas. Mientras tanto, se verificó que los resultados de otras proteínas diferenciales clave eran los mismos que los del proteoma. Por lo tanto, el experimento de células cultivadas con miotubos demostró que el efecto asincrónico de la liberación de aminoácidos en la dieta sobre la deposición de nitrógeno puede estar relacionado con la sincronización del suministro de aminoácidos en el sitio de síntesis de proteínas (células musculares) en el cuerpo. El mecanismo molecular del efecto del patrón de suministro de aminoácidos asíncrono en la disminución de la síntesis de proteínas se realiza principalmente a través del efecto de que la amplitud de síntesis de la vía de señalización PI3K-AKT es menor que la amplitud de degradación de la vía de señalización FoxO.
5 Conclusión
En resumen, los experimentos in vitro confirmaron que el suministro asincrónico de aminoácidos redujo la cantidad de proteína depositada en las células. El análisis proteómico identificó cuatro proteínas distintas y dos vías de señalización relacionadas con la síntesis de proteínas, incluida la vía de señalización PI3K-AKT y la vía de señalización FoxO. Al detectar estos marcadores proteicos, es posible evaluar con precisión el grado en que se ve afectada la síntesis de proteínas. Los resultados de este estudio tienen una importancia teórica importante para comprender mejor los requisitos dinámicos de la nutrición de aminoácidos y proteínas animales y promover el proceso de precisión de la nutrición animal.
Declaración de disponibilidad de datos
Extraemos las cifras anteriores de acuerdo con los datos de prueba. Los conjuntos de datos analizados durante el presente estudio están disponibles a través del autor correspondiente previa solicitud razonable. Los resultados proteómicos presentados en el estudio están a disposición del público. Estos datos se pueden encontrar en: ProteomeXchange Consortium a través del repositorio de socios PRIDE con un identificador de conjunto de datos PXD051755.
Declaración ética
No se requirió aprobación ética para los estudios en animales de acuerdo con la legislación local y los requisitos institucionales porque solo se utilizaron líneas celulares establecidas disponibles comercialmente.
Contribuciones de los autores
QZ: Redacción – borrador original. MM: Análisis formal, Escritura – borrador original. TE: Análisis formal, Redacción – borrador original. XF: Análisis formal, Escritura – borrador original. NB: Metodología, Redacción – borrador original. LP: Metodología, Redacción – borrador original. YZ: Conceptualización, Obtención de fondos, Redacción – revisión y edición. GQ: Conceptualización, Redacción – revisión y edición.
Financiación
El (los) autor(es) declara(n) que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio contó con el apoyo del Programa Provincial de Desarrollo Científico y Tecnológico de Jilin (No. 20220202042NC).
Reconocimientos
Agradecemos a todos los miembros que trabajaron en la recolección de muestras y estadísticas de datos. Nos gustaría agradecer a los revisores y editores por sus valiosos comentarios y sugerencias y por su cuidadosa revisión de nuestro manuscrito.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Abreviaturas
AA, Aminoácidos; PI3K-AKT, fosfatidilinositol-3-quinasa-proteína quinasa B; FoxO, Forkhead Box O; IRS1: Sustrato del receptor de insulina; EGFR: Receptor del Factor de Crecimiento Epidérmico; DEME, el medio de águila modificado de Dulbecco; HCL, Cloruro de hidrógeno; PDGFRB: receptor beta del factor de crecimiento derivado de plaquetas; ECM: Matriz extracelular; mTOR, diana de la rapamicina en mamíferos; ERK: Proteínas quinasas reguladas extracelulares; AKT: Proteína quinasa B; RTK: Receptor de tirosina quinasas.
Notas
Referencias
1. NRC . Requerimientos de nutrientes de los cerdos. 11ª ed. Washington DC: National Academies Press (2012).
2. Stein, HH, Pedersen, C, Wirt, AR, y Bohlke, RA. Aditividad de los valores de digestibilidad ileal aparente y estandarizada de los aminoácidos en dietas mixtas alimentadas con cerdos en crecimiento. J Anim Sci. (2005) 83:2387–95. doi: 10.2527/2005.83102387x
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. van Milgen, J, y Dourmad, JY. Concepto y aplicación de proteína ideal para cerdos. J Anim Sci Biotechnol. (2015) 6:15. doi: 10.1186/s40104-015-0016-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Zhong, RZ, Xia, JQ, Sun, H y Qin, GX. Efectos de diferentes fuentes de proteínas sobre el rendimiento del crecimiento, la química sanguínea y los perfiles de polipéptidos en la digesta del tracto gastrointestinal de lechones recién destetados. J Anim Physiol Anim Nutr. (2017) 101:e312e22. doi: 10.1111/jpn.12607
5. Hu, NZ, Shen, ZW, Pan, L, Qin, GX, Zhao, Y y Bao, N. Efectos del contenido de proteínas y la inclusión de fuentes de proteínas con diferentes dinámicas de liberación de aminoácidos en la utilización de nitrógeno de lechones destetados. Anim Biosci. (2022) 35:260–71. doi: 10.5713/ab.21.0142
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Wang, B, Mi, MM, Zhang, QY, Bao, N, Pan, L y Zhao, Y. Relación entre la cinética de liberación de aminoácidos de las proteínas alimenticias y el equilibrio de nitrógeno en cerdos de engorde. Animal. (2021) 10:100359. doi: 10.1016/j.animal.2021.100359
7. Zhang, QY, Wang, B, Hu, NZ, Bao, N, Pan, L, Zhao, Y, et al. Relación entre la cinética de liberación de aminoácidos en la dieta y la deposición de nitrógeno en cerdos en crecimiento. Anim Nutr. (2022) 9:233–9. doi: 10.1016/j.aninu.2022.01.001
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Shavlakadze, T, y Grounds, M. De osos, ranas, carne, ratones y hombres: complejidad de los factores que afectan la masa muscular esquelética y la grasa. BioEnsayos. (2006) 28:994–1009. doi: 10.1002/bies.20479
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Roths, M, Abeyta, MA, Wilson, B, Rudolph, TE, Hudson, MB, Rhoads, RP, et al. Efectos del estrés térmico sobre marcadores de proteólisis del músculo esquelético en ganado lechero. J Lácteos Sci. (2023) 106:5825–34. doi: 10.3168/jds.2022-22678
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Atherton, PJ, Etheridge, T, Watt, PW, Wilkinson, D, Selby, A, Rankin, D, et al. Efecto muscular completo después de la proteína oral: concordancia y discordancia dependiente del tiempo entre la síntesis de proteínas musculares humanas y la señalización de mTORC1. Am J Clin Nutr. (2010) 92:1080–8. doi: 10.3945/ajcn.2010.29819
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Sandri, M . Señalización en atrofia e hipertrofia muscular. Fisiología. (2008) 23:160–70. doi: 10.1152/physiol.00041.2007
12. Deng, D, Yao, K, Chu, WY, Li, TJ, Huang, RL, Yin, YL, et al. Alteración de la activación del inicio de la traducción y reducción de la síntesis de proteínas en lechones destetados alimentados con una dieta baja en proteínas. J Nutr Biochem. (2009) 20:544–52. doi: 10.1016/j.jnutbio.2008.05.014
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Suryawan, A, Torrazza, RM, Gazzaneo, MC, Orellana, RA, Fiorotto, ML, el-Kadi, SW, et al. La suplementación enteral con leucina aumenta la síntesis de proteínas en los músculos esqueléticos y cardíacos y en los tejidos viscerales de los cerdos neonatos a través de las vías dependientes de mTORC1. Pediatr Res. (2012) 71:324–31. doi: 10.1038/pr.2011.79
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Tannu, NS, Rao, VK, Chaudhary, RM, Giorgianni, F, Saeed, AE, Gao, Y, et al. Los proteomas comparativos de los mioblastos C2C12 en proliferación y los miotubos completamente diferenciados revelan la complejidad del programa de diferenciación del músculo esquelético. Proteómica de células molares. (2004) 3:1065–82. DOI: 10.1074/MCP. M400020-MCP200
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Shi, YF, Wang, Y, Shi, XY, Zhang, XY y Zhang, S. La naringenina promueve la expresión de miofibras oxidadas a través de la vía de señalización PKA en ratones C57BL/6J y células C2C12. J Alimentos Fungt. (2023) 111:105902. doi: 10.1016/j.jff.2023.105902
16. Wang, J . Liberación de aminoácidos de diferentes dietas de fuentes de proteínas y su efecto sobre la dinámica de absorción de aminoácidos de la vena porta de cerdos en crecimiento. Changchun: Universidad Agrícola de Jilin (2019).
17. Yan, GK, Li, XZ, Cheng, XF, Peng, Y, Long, BS, Fan, QW, et al. El perfil proteómico revela que la vía de fosforilación oxidativa está suprimida en el músculo longissimus dorsi de lechones destetados alimentados con una dieta baja en proteínas suplementada con aminoácidos limitantes. Int J Biochem Cell Biol. (2016) 79:288–97. doi: 10.1016/j.biocel.2016.08.024
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Harper, AE, Miller, RH y Block, KP. Metabolismo de aminoácidos de cadena ramificada. Annu Rev Nutr. (1984) 4:409–54. doi: 10.1146/annurev.nu.04.070184.002205
19. Dai, ZL, Wu, ZL, Jia, SC y Wu, GY. Análisis de la composición de aminoácidos en proteínas de tejidos animales y alimentos como derivados de o-ftaldialdehído pre-columna por HPLC con detección de fluorescencia. J Cromatógrafo B. (2014) 964:116–27. doi: 10.1016/j.jchromb.2014.03.025
20. Goodman, CA, Mabrey, DM, Frey, JW, Miu, MH, Schmidt, EK, Pierre, P, et al. Nuevos conocimientos sobre la regulación de la síntesis de proteínas del músculo esquelético revelados por una nueva técnica in vivo no radiactiva. FASEB J. (2011) 25:1028–39. doi: 10.1096/fj.10-168799
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Lelouard, H, Ferrand, V, Marguet, D, Bania, J, Camosseto, V, David, A, et al. Las estructuras inducidas por agresomas similares a las células dendríticas son áreas dedicadas para la ubiquitinación y el almacenamiento de proteínas defectuosas recién sintetizadas. J Cell Biol. (2004) 164:667–75. doi: 10.1083/jcb.200312073
22. Schmidt, EK, Clavarino, G, Ceppi, M y Pierre, P. SUnSET, un método no radiactivo para monitorear la síntesis de proteínas. Métodos Nat. (2009) 6:275–7. doi: 10.1038/nmeth.1314
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Deng, Z, Luo, P, Lai, W, Song, TX, Peng, J y Wei, HK. La miostatina inhibe eEF2K-eEF2 mediante la regulación de la AMPK para suprimir la síntesis de proteínas. Biochem Biophys Res Commun. (2017) 494:278–84. doi: 10.1016/j.bbrc.2017.10.040
24. Ham, DJ, Gleeson, BG, Chee, A, Baum, DM, Caldow, MK, Lynch, GS, et al. La L-citrulina protege las células del músculo esquelético de los estímulos caquécticos a través de un mecanismo dependiente de iNOS. PLoS Uno. (2015) 10:e0141572. doi: 10.1371/journal.pone.0141572
25. Caldow, MK, Ham, DJ, Trieu, J, Chung, JD, Lynch, GS y Koopman, R. La glicina protege las células musculares del desgaste in vitro a través de la señalización mTORC1. Nuerca delantera. (2019) 6:172. doi: 10.3389/fnut.2019.00172
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Wisniewski, JR, Zougman, A, Nagaraj, N y Mann, M. Método universal de preparación de muestras para el análisis del proteoma. Métodos Nat. (2009) 6:359–62. doi: 10.1038/nmeth.1322
27. Cox, J y Mann, M. MaxQuant permite altas tasas de identificación de péptidos, precisiones de masa individualizadas en el rango de ppb y cuantificación de proteínas en todo el proteoma. Nat Biotechnol. (2008) 26:1367–72. doi: 10.1038/nbt.1511
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Wu, GY, Bazer, FW, Dai, ZL, Li, DF, Wang, JJ y Wu, ZL. Nutrición de aminoácidos en animales: síntesis de proteínas y más allá. Annu Rev Anim Biosci. (2014) 2:387–417. doi: 10.1146/annurev-animal-022513-114113
29. Nørgaard, JV, Florescu, IC, Krogh, U, y Nielsen, TS. Perfiles de absorción de aminoácidos en cerdos en crecimiento alimentados con diferentes fuentes de proteínas. Animales. (2021) 11:1740. doi: 10.3390/ani11061740
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Zheng, LF, Zuo, FR, Zhao, SJ, He, PL, Wei, HK, Xiang, QH, et al. La suplementación dietética de aminoácidos de cadena ramificada aumenta los flujos netos de aminoácidos musculares a través de la elevación de su disponibilidad de sustrato y el catabolismo intramuscular en cerdos jóvenes. Hno. J Nutr. (2017) 117:911–22. doi: 10.1017/S0007114517000757
31. Shah, OJ, Anthony, JC, Kimball, SR y Jefferson, LS. 4E-BP1 y S6K1: sitios de integración traslacional para información nutricional y hormonal en músculo. Am J Physiol Endocrinol Metab. (2000) 279:E715-29. doi: 10.1152/ajpendo.2000.279.4.E715
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Volpi, E, Kobayashi, H, Sheffield-Moore, M, Mittendorfer, B, y Wolfe, RR. Los aminoácidos esenciales son los principales responsables de la estimulación de aminoácidos del anabolismo de proteínas musculares en adultos mayores sanos. J Clin Nutr. (2003) 78:250–8. doi: 10.1093/ajcn/78.2.250
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Hosios, AM, Hecht, VC, Danai, LV, Johnson, MO, Rathmell, JC, Steinhauser, ML, et al. Los aminoácidos, en lugar de la glucosa, representan la mayor parte de la masa celular en las células de mamíferos en proliferación. Célula de desarrollo. (2016) 36:540–9. doi: 10.1016/j.devcel.2016.02.012
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Palm, W, y Thompson, CB. Estrategias de adquisición de nutrientes de células de mamíferos. Naturaleza. (2017) 546:234–42. doi: 10.1038/nature22379
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Kiran, K, y Haigis, MC. Metabolismo del nitrógeno en el cáncer y la inmunidad. Tendencias Cell Biol. (2020) 17:1584. doi: 10.1016/j.t.cb.2020.02.005
36. Dietz, C, Infanger, M, Romswinkel, A, Strube, F y Kraus, A. Inducción de apoptosis y alteración de la adherencia celular en células de cáncer de pulmón humano bajo microgravedad simulada. Int J Mol Sci. (2019) 20:3601. doi: 10.3390/ijms20143601
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Madhala-Levy, D, Williams, VC, Hughes, SM, Reshef, R y Halevy, O. La cooperación entre Shh e IGF-I para promover la proliferación y diferenciación miogénica a través de las vías MAPK/ERK y PI3K/Akt requiere actividad Smo. J Célula Physiol. (2012) 227:1455–64. doi: 10.1002/jcp.22861
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Wang, G, Pan, J, Zhang, L, Wei, Y y Wang, C. Esponjas CRNDE de ARN largo no codificante miR-384 para promover la proliferación y metástasis de las células de cáncer de páncreas a través de la regulación positiva de IRS1. Célula Prolif. (2017) 50:E12389. doi: 10.1111/cpr.12389
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Motohashi, N, Alexander, MS, Shimizu-Motohashi, Y, Myers, JA, Kawahara, G, y Kunkel, LM. Regulación de la señalización de insulina IRS1/Akt por microRNA-128a durante la miogénesis. Célula J Sci. (2013) 126:2678–91. doi: 10.1242/jcs.119966
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Li, Z, Cai, B, Abdalla, BA, Zhu, X, Zheng, M, Han, P, et al. LncIRS1 controla la atrofia muscular mediante la esponja de la familia miR-15 para activar la vía IGF1-PI3K/AKT. J Caquexia Músculo Sarcopenia. (2019) 10:391–410. doi: 10.1002/jcsm.12374
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Yarden, Y, y Sliwkowski, MX. Desenredando la red de señalización ErbB. Nat Rev Mol Cell Biol. (2001) 2:127–37. doi: 10.1038/35052073
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Prensa, MF, y Lenz, HJ. Vías EGFR, HER2 y VEGF. Drogas. (2007) 67:2045–75. doi: 10.2165/00003495-200767140-00006
43. Yarden, Y, y Shilo, BZ. SnapShot: Vía de señalización EGFR. Célula. (2007) 131:1018. doi: 10.1016/j.carolcarrollell.2007.11.013
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Sandri, M, Sandri, C, Gilbert, A, Skurk, C, Calabria, E, Picard, A, et al. Los factores de transcripción de Foxo inducen la ubiquitina ligasa atrogina-1 relacionada con la atrofia y causan atrofia del músculo esquelético. Célula. (2014) 117:399–412. doi: 10.1016/s0092-8674(04)00400-3
45. Stitt, TN, Drujan, D, Clarke, BA, Panaro, F, Timofeyva, Y, Kline, WO, et al. La vía IGF-1/PI3K/Akt previene la expresión de ubiquitina ligasa inducida por atrofia muscular mediante la inhibición de los factores de transcripción FOXO. Célula molar. (2004) 14:395–403. doi: 10.1016/S1097-2765(04)00211-4
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Lai, KM, González, M, Poueymirou, WT, Kline, WO, Na, E, Zlotchenko, E, et al. La activación condicional de akt en el músculo esquelético adulto induce una hipertrofia rápida. Mol Cell Biol. (2004) 24:9295–304. doi: 10.1128/MCB.24.21.9295-9304.2004
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Brown, AK, y Webb, AE. Regulación de factores FOXO en células de mamíferos. Curr Top Dev Biol. (2018) 127:165–92. doi: 10.1016/bs.ctdb.2017.10.006
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Vivanco, I, y Sawyers, CL. La vía AKT de la fosfatidilinositol 3-quinasa en el cáncer humano. Nat Rev Cáncer. (2002) 2:489–501. doi: 10.1038/nrc839
49. Pallafacchina, G, Calabria, E, Serrano, AL, Kalhovde, JM, y Schiaffino, S. Una vía dependiente de la proteína quinasa B y sensible a la rapamicina controla el crecimiento del músculo esquelético, pero no la especificación del tipo de fibra. PNAS. (2002) 99:9213–8. doi: 10.1073/pnas.142166599
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Latres, E, Amini, AR, Amini, AA, Griffiths, J, Martin, FJ, Wei, Y, et al. El factor de crecimiento similar a la insulina-1 (IGF-1) regula inversamente los genes inducidos por la atrofia a través de la vía de la fosfatidilinositol 3-quinasa/Akt/diana de la rapamicina en mamíferos (PI3K/Akt/mTOR). J Biol Chem. (2005) 280:2737–44. doi: 10.1074/jbc. M407517200
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Bonaldo, P, y Sandri, M. Mecanismos celulares y moleculares de la atrofia muscular. Dis Model Mech. (2013) 6:25–39. doi: 10.1242/dmm.010389
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Zanou, N, y Gailly, P. Hipertrofia y regeneración del músculo esquelético: interacción entre las vías de los factores reguladores miogénicos (MRF) y los factores de crecimiento similares a la insulina (IGF). Célula Mol Life Sci. (2013) 70:4117–30. doi: 10.1007/s00018-013-1330-4
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: nutrición, miotubos C2C12, aporte de aminoácidos, sincronicidad, síntesis proteica
Cita: Zhang Q, Mi M, E T, Fu X, Bao N, Pan L, Zhao Y y Qin G (2024) Efecto de la sincronicidad del suministro de aminoácidos en la síntesis de proteínas en miotubos C2C12 cultivados in vitro. Frente. Vet. Sci. 11:1423604. doi: 10.3389/fvets.2024.1423604
Editado por:
Kai Wang, Academia China de Ciencias Agrícolas (CAAS), China
Revisado por:
Xiang Bing Mao, Universidad Agrícola de Sichuan, China
Chaoyue Wen, Universidad Agrícola de Hunan, China
Derechos de autor © 2024 Zhang, Mi, E, Fu, Bao, Pan, Zhao y Qin. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Yuan Zhao, zhaoyuan4CL52@126.com; Guixin Qin, qgx@jlau.edu.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía