
Efectos de diferentes suministros dietéticos de treonina y glicina en pollos de engorde alimentados con dietas bajas en proteínas

Efectos de diferentes suministros dietéticos de treonina y glicina en pollos de engorde alimentados con dietas bajas en proteínas
 Patrik Strifler Boglárka Horváth
Patrik Strifler Boglárka Horváth  Nikoletta Such
Nikoletta Such  Károly Dublecz
Károly Dublecz  László Pál*
László Pál*- Departamento de Nutrición y Fisiología Nutricional, Instituto de Fisiología y Nutrición, Universidad Húngara y Agricultura y Ciencias de la Vida, Keszthely, Hungría
La reducción del contenido de proteína bruta (PC) de las dietas de pollos de engorde con un suministro equilibrado de aminoácidos puede aumentar la eficiencia de la utilización del nitrógeno (N) y reducir la emisión de amoníaco, el riesgo de muchos problemas de salud en las aves. La alimentación con dietas bajas en proteínas (LP) sin el deterioro de los rasgos de rendimiento necesita los niveles dietéticos optimizados de treonina (Thr) y los aminoácidos no esenciales (AA), glicina (Gly) y serina (Ser). Sin embargo, las concentraciones e interacciones requeridas de Thr y Gly + Ser, expresadas como equivalentes de Gly (Glyequi), en las dietas LP no se comprenden completamente. Por lo tanto, el objetivo de este estudio fue investigar los efectos de tres dietas LP (LP1-3) de crecimiento (11-24 días) y de engorde (25-35 días) con una reducción del 2% de la PC en comparación con el control (C), difiriendo en la relación Thr a lisina (Lys) digestible ileal estandarizada (SID) (C, LP1, LP3: 63%, LP2: 72%) y Glyequi (C: 15,65 g/kg, LP1: 13,74 g/kg, LP2: 13,70 g/kg, LP3: 15,77). Los tratamientos con LP no afectaron las características de rendimiento de los pollos de engorde. El tratamiento LP2 con aumento de la relación SID Thr-to-Lys (+9,0%) resultó en una ganancia de peso corporal significativamente mayor y una tasa de conversión alimenticia más ventajosa en todo el engorde en comparación con el tratamiento control con un nivel normal de PB (p < 0,05). El tratamiento LP3 que contiene harina de carne de cerdo con Gly similarequi En comparación con el tratamiento normal de PC, se obtuvo la tasa de conversión alimenticia más ventajosa en la fase de engorde y la mayor eficiencia de retención de nitrógeno (p < 0,05). Sin embargo, el tratamiento con LP3 con una alta relación almidón/PB influyó negativamente en el peso relativo de la canal y en la relación de grasa abdominal de los pollos de engorde (p < 0,05).
1 Introducción
La alimentación de los pollos de engorde con dietas bajas en proteínas (LP) ha sido el método más eficaz para reducir la excreción de nitrógeno (N) y aumentar la eficiencia de la utilización del N, reduciendo el riesgo de problemas en la cama húmeda y la incidencia de disbiosis (1-3). Nuestro conocimiento actual permite la reducción de hasta un 2% de proteína bruta (PC) en dietas de pollos de engorde a base de maíz/harina de soja en cada fase de alimentación sin pérdida de rendimiento ni deterioro de la calidad del producto (4). Este nivel de reducción de la PB puede lograrse con éxito mediante un ajuste preciso del suministro de aminoácidos esenciales (AA) de las aves utilizando el «concepto de proteína ideal» sobre una base de aminoácidos digeribles ileales (SID) normalizados (5). Existe una amplia gama de AA cristalinos libres como suplementos alimenticios que pueden utilizarse para este fin.
La treonina (Thr) ha sido generalmente el tercer AA limitante en las dietas de los pollos de engorde, y el grado alimenticio L-Thr ha estado disponible comercialmente desde la década de 1980 (6). El Thr sirve principalmente como sustrato para la síntesis de proteínas, mucina e inmunoglobulinas y desempeña un papel crucial en la respuesta al estrés y el mantenimiento de la integridad del epitelio intestinal (7). Las proporciones ideales de AA esenciales deben considerarse aún más cuidadosamente en las dietas LP. La proporción ideal de Thr a lisina digestible (Lys) se ha incrementado hasta más del 65% en las dietas para pollos de engorde modernos, que se puede ajustar aún más de acuerdo con el rasgo de rendimiento objetivo, el nivel de estrés y los desafíos del sistema inmunológico (8, 9). El Thr puede ser metabolizado por la Thr aldolasa o la Thr deshidrogenasa a glicina (Gly) en las aves de corral, que es el precursor AA de la síntesis de ácido úrico (10). Además, Gly puede metabolizarse a partir de la serina (Ser), y esta reacción puede revertirse (11, 12). La interconversión metabólica de Gly y Ser es continua e ilimitada, y Ser tiene los mismos efectos que Gly sobre una base equimolar (11). Por lo tanto, el cálculo de un equivalente de Gly en la dieta (Glyequi) se ha recomendado utilizar el equivalente molar de Ser de 0,7143 (13). Los niveles de Gly + Ser en la dieta disminuyen en gran medida cuando se reduce la PC en las dietas vegetarianas, y estos AA se convierten en los primeros AA no esenciales que limitan el crecimiento cuando el contenido de PC de las dietas está por debajo del 19% durante los 7 a 21 días de edad y por debajo del 17% durante los 21 a 35 días de edad (14, 15). La interrelación de la dieta Thr y Glyequi es bastante compleja y ha sido reportada por muchos estudios (16-21). Los niveles de Thr en la dieta más altos que los recomendados pueden reducir el requerimiento de Gly en la dietaequi para alcanzar ciertos niveles de respuesta de los rasgos de rendimiento (22). Este llamado efecto de ahorro o reemplazo puede atribuirse en parte a la conversión de Thr en Gly (23). Además, el aumento de los niveles dietéticos de Thr puede reducir el catabolismo de otros AA, reduciendo así la necesidad de Glyequi para la formación de ácido úrico (23). Por otro lado, la suplementación con Gly disminuye la actividad de las enzimas Thr aldolasa y Thr deshidrogenasa, lo que puede conducir a una disminución de la degradación de Thr y una mayor disponibilidad de Thr para necesidades fisiológicas (19).
La interacción entre Thr y Gly con respecto al rendimiento de los pollos de engorde se ha estudiado principalmente durante las primeras 3 semanas de vida de los pollos de engorde. Las interacciones entre Thr y Glyequi de 7 a 21 días fueron cuantificados por Siegert et al. (22). Descubrieron que el aumento de la Thr en la dieta reducía la Glyequi necesarias para lograr ciertas respuestas BWG y FCR. Los efectos de un 0,2% o 0,4% adicional de Gly dependieron del nivel de PB de la dieta administrada desde el día 0 hasta el día 21 en el estudio de Waldroup et al. (16). Cuando se añadió Gly a dietas con 16% o 18% de PC (1,30 o 1,62% Gly + Ser), el peso corporal de las aves a los 21 días de edad mejoró significativamente. Sin embargo, la adición de Gly a dietas con 20, 22% o 24% de PC (1,86, 2,08 y 2,28% de Gly + Ser) mostró poco o ningún beneficio. Los autores no observaron ninguna mejora en el rendimiento de un 0,2% o 0,4% de Thr adicional (0,80-0,98% de Thr en dietas basales) ni interacción entre Gly y Thr. Ospina-Rojas et al. (18) investigaron una dieta LP con 19% de PB alimentado desde el día 0 hasta el día 21 con dos niveles de Thr (0,93 y 1,07% Thr) y cuatro concentraciones de Gly + Ser (1,84 a 2,26%). El aumento suplementario de Gly + Ser mejoró el FCR de las aves de manera cuadrática al nivel de 0.93% de Thr, pero no tuvo un efecto significativo en el FCR a una concentración de 1.07% de Thr. La investigación centrada en la interacción entre Thr y Gly en la fase de engorde de engorde es más limitada y necesita una cuantificación más precisa. El FCR de los pollos de engorde de 21 a 35 días mostró una respuesta más fuerte a Gly cuando la proporción de Thr a Lys digerible de las dietas fue del 65% en comparación con la proporción de Thr a Lys digerible del 72% (19). Los pollos de engorde alimentados con una dieta con un nivel más bajo de digestibilidad (Gly + Ser)-to-Lys mostraron una respuesta más fuerte de FCR al nivel de Thr en la dieta (135% vs. 149%) (19). Corzo et al. (17) también observaron interacciones similares para BWG de aves de 21 a 42 días de edad. En el estudio de Star et al. (21), el BWG de pollos de engorde de 7 a 28 días de edad solo respondió cuando los niveles de Gly y Thr en la dieta eran muy bajos. Además de sus respectivas concentraciones en las dietas, los efectos de Thr y Gly en la dieta también están relacionados con el nivel dietético de Met+Cys, colina, Arg, ácido guanidino acético y creatina, según lo revisado por Siegert y Rodehutscord (23).
La mayoría de los estudios centrados en las interacciones Thr-Gly aplicaron solo una dieta LP y suplementos cristalinos de Gly y Thr (17-19). El objetivo de este estudio fue utilizar dietas formuladas de forma práctica con contenido de PB normal y bajo sin glis cristalino, que solo está permitido como agente aromatizante en la UE. Para aumentar la concentración de Gly en las dietas LP, en nuestro experimento se utilizó harina de carne de cerdo rica en Gly. Además, el presente estudio se centra no solo en las interacciones Thr y Gly en relación con el rendimiento de los pollos de engorde, sino también en las características de la canal, la calidad de la carne, la eficiencia de la retención de nitrógeno y las formas nitrogenadas de los excrementos utilizando dietas LP formuladas en la práctica adecuadas en la UE.
2 Materiales y métodos
2.1 Animales de experimentación y tratamientos
Se llevó a cabo una prueba de corral en el suelo en la granja experimental del Instituto de Fisiología y Nutrición, Campus Georgikon, Universidad Húngara de Agricultura y Ciencias de la Vida (Keszthely, Hungría). Se compraron un total de 576 pollos de engorde machos de 1 día de edad (Ross 308) en un criadero local (Gallus Ltd., Devecser, Hungría) y se dividieron aleatoriamente en 24 corrales de piso a una tasa de carga de 24 aves por corral (14 aves/m2). Los animales fueron vacunados contra la bronquitis infecciosa (CEVAC BRON), la enfermedad de Newcastle (CEVAC VITAPEST) y la bursitis infecciosa (CEVAC TRANSMUNE) en la planta de incubación utilizando vacunas producidas por Ceva (Ceva Santé Animale, Francia). Se utilizó paja de trigo picada como material de cama. A los animales se les proporcionó agua y alimento ad libitum durante toda la duración del experimento. Las condiciones climáticas y el programa de luz, basados en las directrices del criador, fueron controlados por ordenador e idénticos para todos los corrales. La temperatura ambiente se fijó en 34 °C el día 0 y se redujo gradualmente a 24 °C a los 18 días de edad. La intensidad de la luz fue de 30 lux en la primera semana y de 10 lux a partir de entonces, con una duración constante de 23 h desde el día 0 hasta el día 7 y 20 h de luz y 4 h de oscuridad a partir de entonces.
Durante el experimento de 35 días de duración se utilizaron tres fases dietéticas: iniciador (de 0 a 10 días), de crecimiento (de 11 a 24 días) y de engorde (de 25 a 35 días). Todas las aves fueron alimentadas con la misma dieta de la fase inicial, y se establecieron 4 tratamientos dietéticos que consistieron en 6 repeticiones con 24 aves en cada una y se alimentaron dietas experimentales en las fases de crecimiento y engorde en la forma peletizada. El diseño del experimento se describe en la Tabla 1. Las dietas del tratamiento control C se formularon de acuerdo con las recomendaciones del criador para Ross 308 (Aviagen, Newbridge, Reino Unido) y niveles adecuados de PB y SID AA esenciales. Las dietas bajas en proteínas (LP), LP1, LP2 y LP3, contenían un 2,0% menos de proteína bruta que la dieta C con un nivel de PB controlado en cada fase de la dieta. Los tratamientos dietéticos fueron diferentes en el nivel de SID Thr y la relación SID Thr-to-Lys, así como en la relación SID (Gly + Ser)-Lys y la relación Glyequi. La composición de las dietas experimentales se muestra en la Tabla 2, mientras que el contenido de nutrientes calculado y medido de las dietas experimentales se puede ver en la Tabla 3. Las dietas experimentales de LP fueron isocalóricas con la dieta C. El aumento de Glyequi en la dieta LP3 se logró mediante la sustitución parcial de harina de soja por harina de carne de cerdo rica en Gly. Las dietas se formularon a base de AA estandarizados digestibles ileales (SID) de acuerdo con el concepto de proteína ideal. Las dietas LP se suplementaron con seis AA esenciales cristalinos de grado alimenticio (Lys, Met, Val, Thr, Arg e Ile) para cumplir con las concentraciones calculadas de AA SID en las dietas C, excepto para el nivel de SID Thr de la dieta LP2. Todas las dietas contenían enzimas fitasa y xilanasa, pero no se consideró el impacto de estas enzimas en la liberación de aminoácidos en las formulaciones de los piensos.
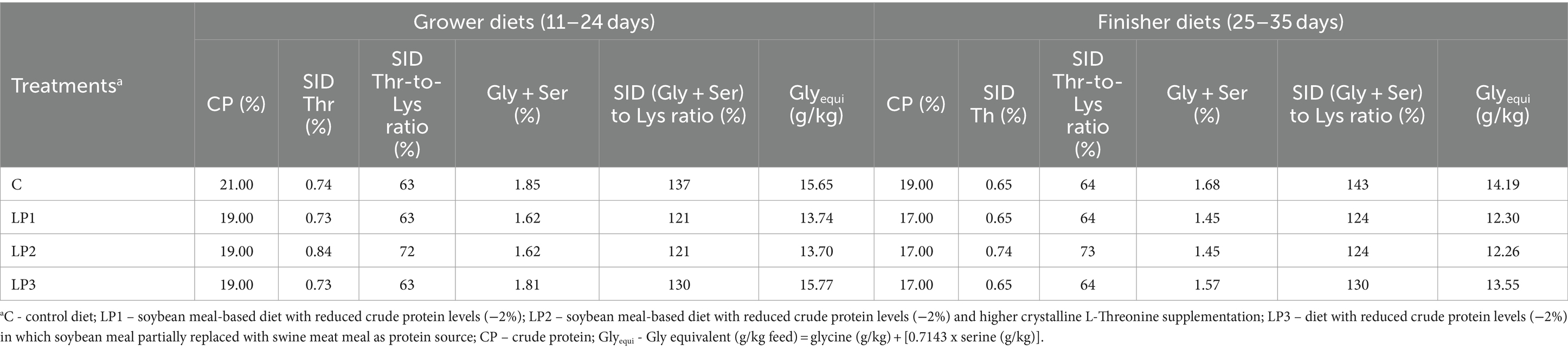 Tabla 1. Diseño experimental.
Tabla 1. Diseño experimental.
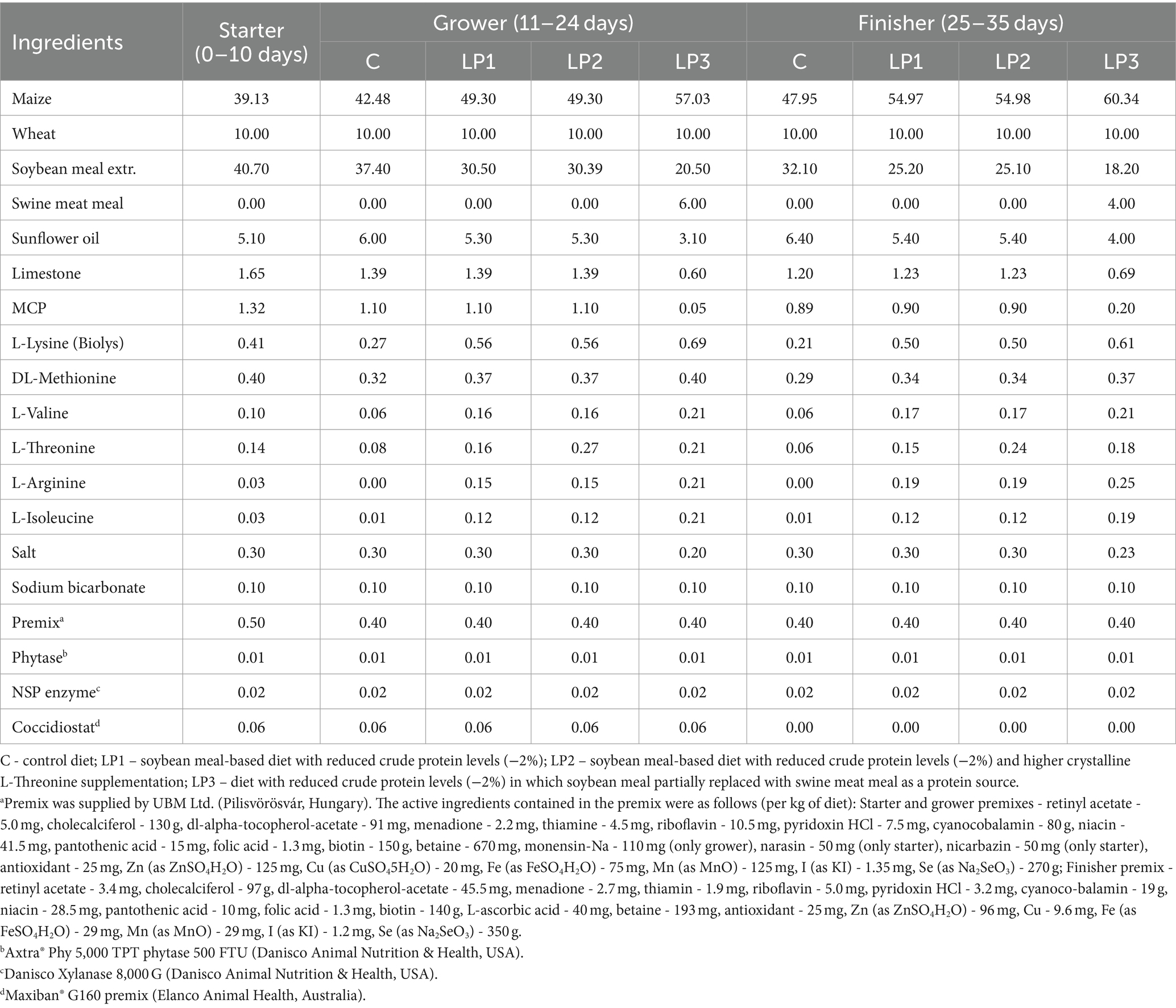 Tabla 2. Composición de las dietas experimentales (%).
Tabla 2. Composición de las dietas experimentales (%).
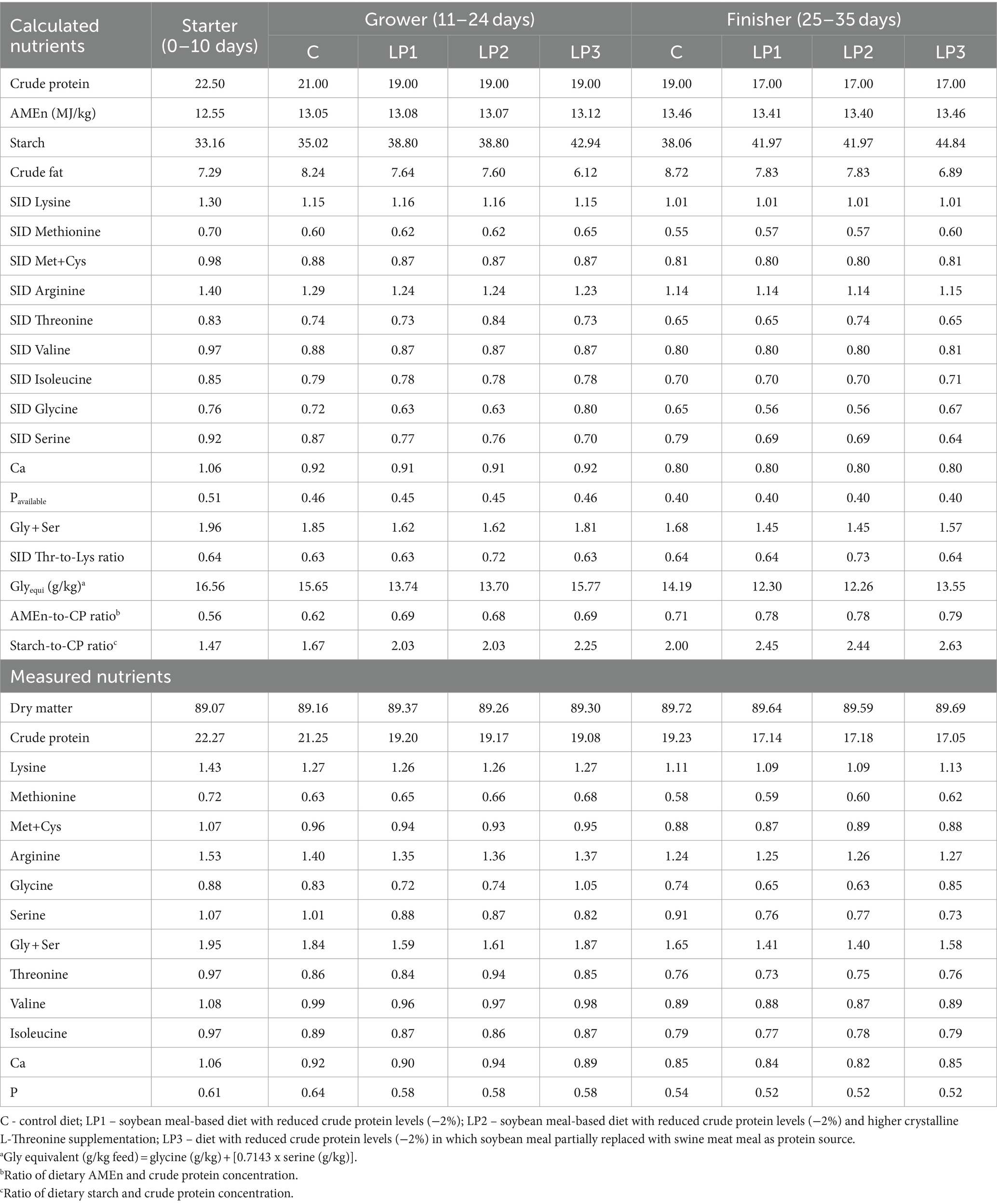 Tabla 3. Se calculó y midió el contenido de nutrientes de las dietas experimentales (%).
Tabla 3. Se calculó y midió el contenido de nutrientes de las dietas experimentales (%).
2.2 Medidas
El peso corporal (PC) de los pollos de engorde se midió individualmente al inicio del ensayo y al final de cada fase dietética, y se calculó el peso corporal medio para cada corral. La ingesta de alimento (IF) de los pollos de engorde se registró por corral al final de cada fase de la dieta. Se calculó la ganancia de peso corporal (BWG) y el índice de conversión alimenticia (FCR) por corral al final de cada fase y durante todo el período de prueba. La mortalidad y el peso de las aves muertas se registraron diariamente durante todo el ensayo. A los 35 días, se seleccionaron aleatoriamente dos pollos con peso corporal promedio de cada corral (12 aves por tratamiento) y se transfirieron a jaulas de equilibrio, donde los pollos consumieron las mismas dietas de engorde pero suplementadas con 0,5% de TiO2 como marcador interno indigerible. Después de un período de adaptación de 5 días, se recolectaron muestras representativas de excretas de cada ave diariamente durante 2\u00B00días consecutivos (días 41 y 42). Las muestras de 12 aves por tratamiento se agruparon, se mezclaron completamente, se congelaron y se almacenaron a -20 °C hasta nuevos análisis. Antes de los análisis, los excrementos se homogeneizaron adecuadamente, y luego el contenido de materia seca, N total, N de amonio (NH4+-N) y se determinó el contenido de ácido úrico-N. El contenido de materia seca de las muestras de excretas se midió en un exicador (100°C durante 24 h). El N total de excretas se determinó según el método de Kjeldahl con el Analizador Foss-Kjeltec 8.400 (Nils Foss Allé 1, DK-3400 Hilleroed, Dinamarca), el N de amonio por el método de Peters (24) y el N de ácido úrico descrito por Marquardt (25). Todos los parámetros de N se ajustaron sobre la base de la materia seca. La suma de NH4+El contenido de N y el nitrógeno úrico se consideró como contenido de N urinario (26). Las muestras de alimento se analizaron para determinar el contenido de materia seca (ISO 6496), proteína cruda (ISO 5983-1:2005), fósforo (ISO 6491), calcio (ISO 6896) y composición de aminoácidos (ISO 13903:2005) utilizando métodos de la Organización Internacional de Normalización (ISO). El TiO2\u00B0La concentración de las dietas experimentales y las muestras de excretas se determin\u00f3 mediante un ensayo de espectroscopía UV (27). La retención de nitrógeno se calculó mediante la siguiente ecuación (28): Retención aparente de nitrógeno = 1 – [([TiO2] dieta/[TiO2] excretas) × (excretas [de nitrógeno]/dieta [de nitrógeno])]. Al final del experimento, se seleccionaron dos aves por corral (12 aves por tratamiento) que representaban el peso corporal promedio del corral para ser sacrificadas por luxación cervical. Después de la evisceración, se determinó la composición de la canal (% del peso de la canal, % de carne de pechuga, % de peso del muslo, % de grasa abdominal) y la calidad de la carne de la pechuga. El pH del músculo pectoral, Pectoralis major (P. major), se midió inmediatamente después del sacrificio (pH0h) y después de 24 h de almacenamiento a 4 °C (pHu) con un medidor de pH portátil (Testo 205; Testo Ltd., Hungría) insertando un electrodo de vidrio directamente en la parte más gruesa del músculo. La capacidad de retención de agua de la carne se estimó midiendo la pérdida por goteo de la carne cruda: el músculo P. major se pesó inmediatamente después del sacrificio y se colocó en una bolsa de plástico, se colgó de un gancho y se almacenó a 4 °C durante 24 h. Después de colgar, la muestra se limpió con un papel absorbente y se pesó nuevamente. La diferencia de peso correspondiente a la pérdida por goteo se expresó como el porcentaje del peso muscular inicial (29).
2.3 Análisis estadístico
Los promedios de los parámetros examinados se analizaron en un diseño completamente aleatorizado utilizando un análisis de varianza de una vía (ANOVA) con los tratamientos dietéticos como efectos principales. Para los resultados de rendimiento (BW, BWG, FI y FCR), el corral fue la unidad experimental, mientras que para otras variables, el ave individual fue la unidad experimental. Cuando la prueba F reveló un efecto significativo del tratamiento, las diferencias significativas entre los grupos se probaron mediante la prueba de Tukey HSD. Todos los análisis estadísticos se realizaron con el paquete SPSS 22.0 para Windows (IBM Corp., Armonk, NY, Estados Unidos). La significación estadística se ha declarado en p < 0,05.
3 Resultados
No hubo diferencias notables entre los valores medidos y calculados de la AA total de la dieta (Tabla 3). En la Tabla 4 se presenta el efecto de los tratamientos dietéticos sobre la ingesta total de SID esencial AA balanceada sin Thr y la ingesta de SID Thr y Gly + Ser de pollos de engorde en las fases de engorde y engorde. La ingesta total de AA esencial de SID balanceado sin Thr no fue significativamente diferente entre los grupos de tratamiento. Además, los resultados muestran que las diferencias calculadas entre los niveles de SID Thr y Gly + Ser en los tratamientos dietéticos dieron lugar a diferencias significativas en la ingesta de SID Thr y Gly + Ser de los pollos de engorde.
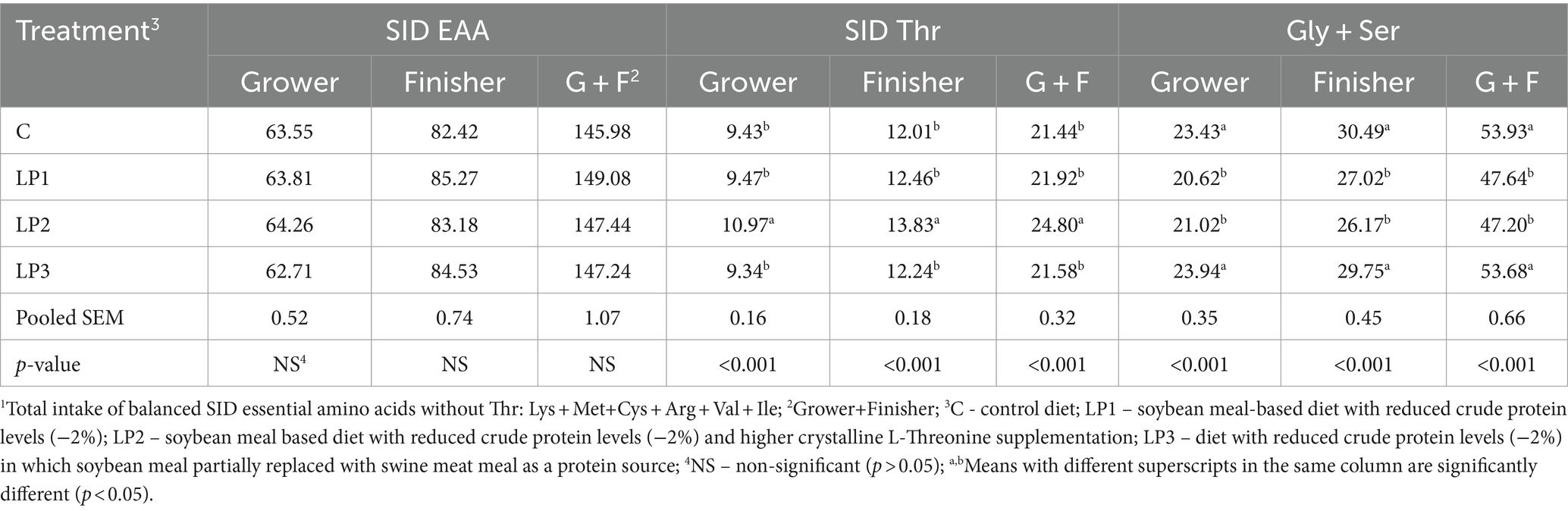 Tabla 4. Ingesta de EAA SID1, SID Thr y Gly + Ser de pollos de engorde durante las fases de alimentación (g/ave, medias de 6 corrales por tratamiento; n = 6).
Tabla 4. Ingesta de EAA SID1, SID Thr y Gly + Ser de pollos de engorde durante las fases de alimentación (g/ave, medias de 6 corrales por tratamiento; n = 6).
Los resultados de los parámetros de producción se muestran en la Tabla 5. Los parámetros de rendimiento no se vieron influenciados significativamente por la misma dieta inicial alimentada en todos los corrales cuando la alimentación de las dietas experimentales comenzó el día 10. No hubo diferencias significativas entre el peso corporal de los grupos de tratamiento al final de la fase de engorde, mientras que en la fase de engorde, los pollos de engorde alimentados con la dieta LP2 mostraron un peso corporal significativamente mayor que los pollos de engorde que consumieron la dieta C (p < 0.05). El BWG de las aves en el período de engorde, engorde y todo el período de prueba se vio afectado significativamente por las dietas experimentales (p < 0.05). En la fase de crecimiento, la alimentación con dietas LP no condujo a una BWG significativamente diferente en comparación con el tratamiento C. Sin embargo, hubo una diferencia significativa entre los grupos LP2 y LP3: el aumento del nivel de SID Thr-to-Lys de la dieta LP2 resultó en un mayor BWG de las aves que la dieta LP3 con mayor Glyequi (p < 0,05). El BWG de los pollos de engorde en los grupos LP1 y LP3 fue significativamente mayor que el del grupo C en la fase de engorde (p < 0.05). Al igual que en todo el experimento, solo los pollos de engorde alimentados con la dieta LP2 lograron un peso corporal significativamente mayor que las aves en el tratamiento C (p < 0.05). A diferencia de los datos de BW y BWG, no se encontraron diferencias significativas en el IF de los animales de experimentación entre los tratamientos en ninguna de las fases del experimento. El FCR de los pollos de engorde se vio significativamente influenciado por los tratamientos dietéticos tanto en la fase de engorde como en la de engorde así como durante todo el engorde (p < 0,05). En la fase de crecimiento, las dietas LP no condujeron a valores de FCR significativamente diferentes a los de la dieta C. En cuanto a la FCR en los tratamientos con LP, se observó la misma diferencia significativa entre LP2 y LP3 que en el caso de la BWG (p < 0,05). Los animales experimentales alimentados con la dieta LP3 mostraron una mejor FCR que las aves control que consumieron la dieta C en la fase de engorde (p < 0.05). El valor de FCR calculado para todo el estudio en el grupo LP2 superó el FCR en el grupo C, pero no difirió significativamente de los otros dos tratamientos con LP.
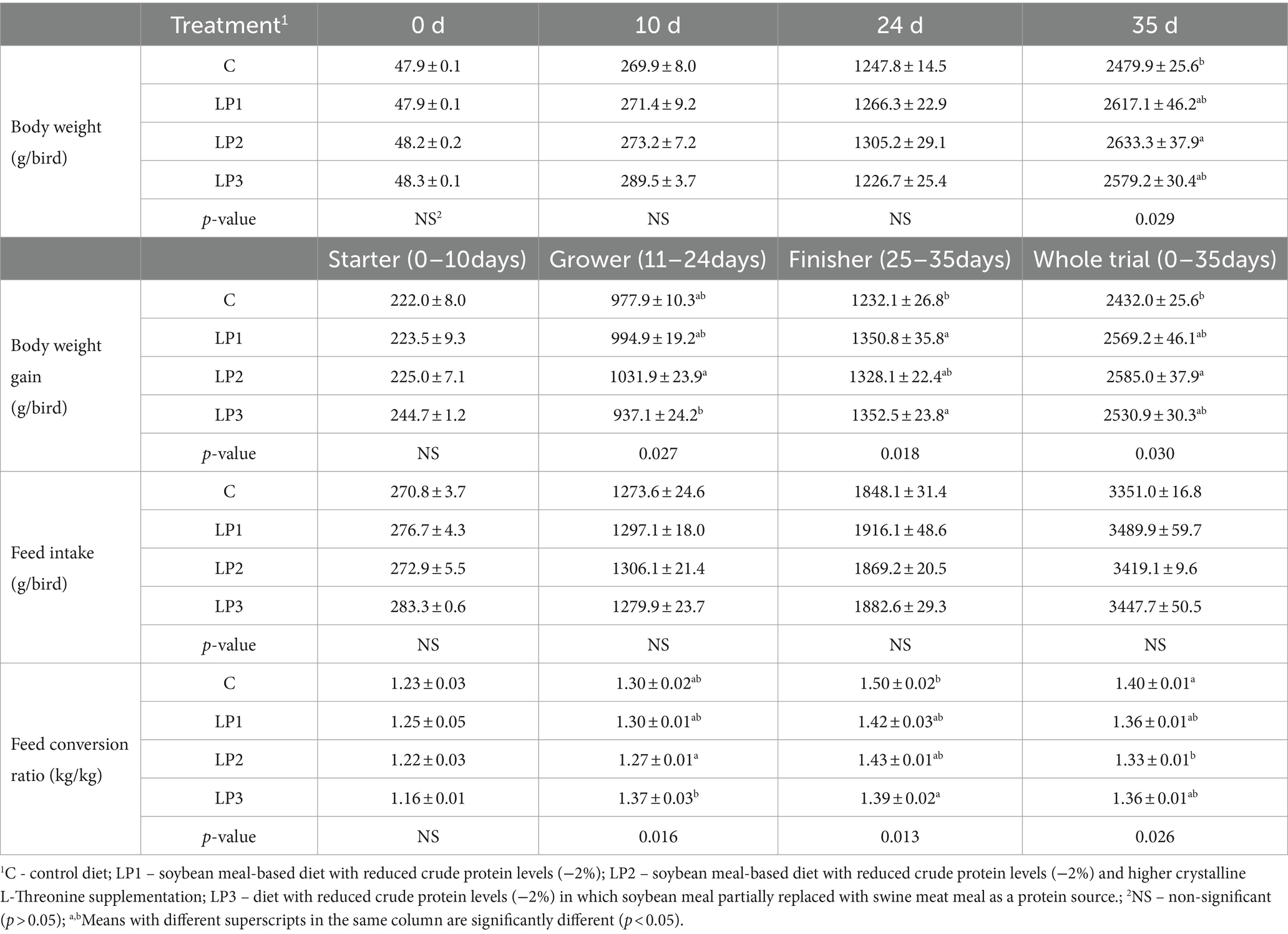 Tabla 5. Parámetros de comportamiento de las aves en las fases de inicio, engorde y engorde y en todo el experimento (media ± SEM; n = 6 plumas por tratamiento).
Tabla 5. Parámetros de comportamiento de las aves en las fases de inicio, engorde y engorde y en todo el experimento (media ± SEM; n = 6 plumas por tratamiento).
Los tratamientos dietéticos afectaron significativamente el peso relativo de la canal y la almohadilla abdominal (p < 0,05), mientras que el rendimiento relativo de carne de pechuga y el peso del muslo no fueron influenciados significativamente por la alimentación experimental (Tabla 6). El peso relativo de la canal de los pollos de engorde en los grupos LP1 y LP2 no fue diferente del grupo C. Sin embargo, este rasgo en el tratamiento con LP3 fue significativamente menor que en los tratamientos con C y LP2. Además, la alimentación de la dieta LP3 resultó en una mayor almohadilla de grasa abdominal relativa en comparación con la dieta C (p < 0,05). Los tratamientos dietéticos no influyeron significativamente en la pérdida por goteo de la carne de pechuga ni en el pH del filete de pechuga medido inmediatamente después del sacrificio o después de 24 h (p > 0,05; Tabla 7).
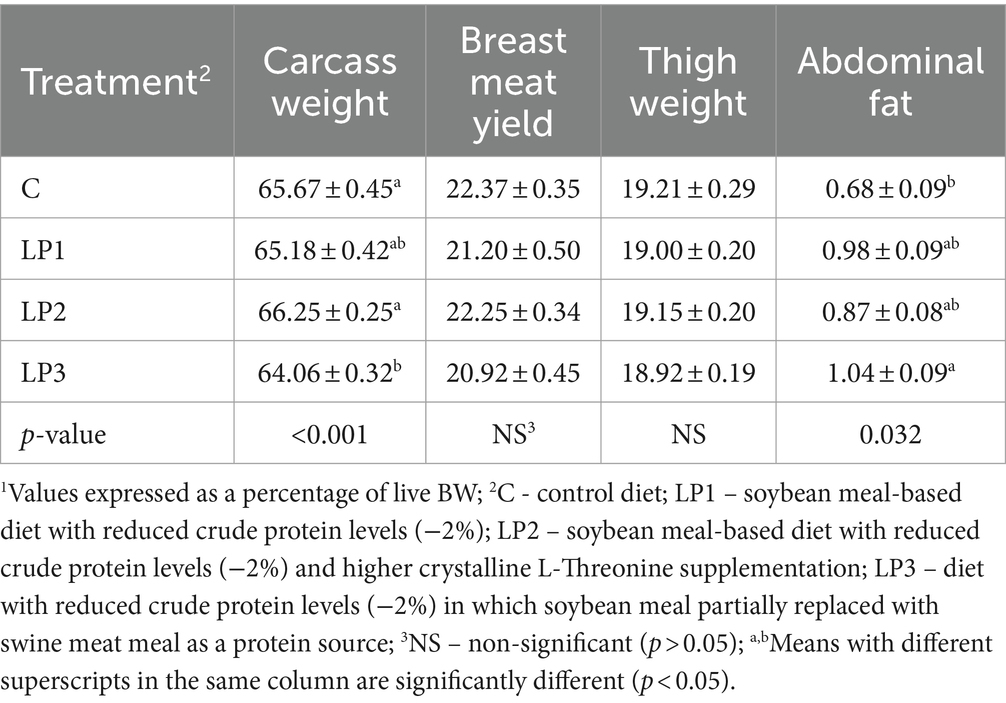 Tabla 6. Peso y composición de la canal1 (%, media ± SEM; n = 12 pollos de engorde por tratamiento).
Tabla 6. Peso y composición de la canal1 (%, media ± SEM; n = 12 pollos de engorde por tratamiento).
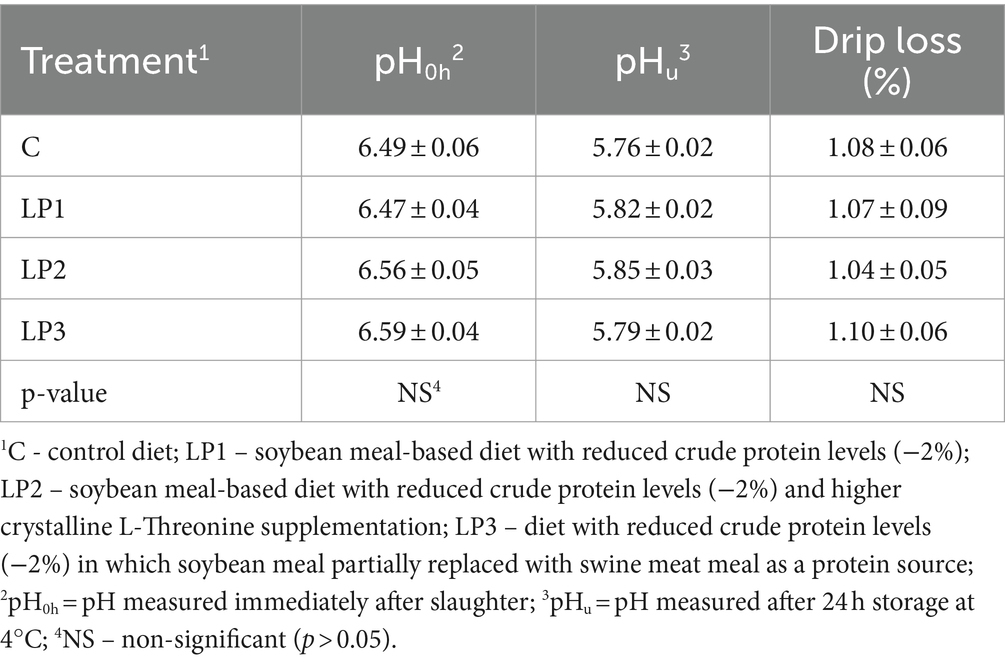 Tabla 7. Parámetros de calidad de la carne de pechuga (media ± SEM; n = 12 pollos de engorde por tratamiento).
Tabla 7. Parámetros de calidad de la carne de pechuga (media ± SEM; n = 12 pollos de engorde por tratamiento).
El efecto de los tratamientos dietéticos sobre la eficiencia de retención de nitrógeno de los pollos de engorde fue significativo (66,4, 72,3, 69,3 y 73,5% en los grupos C, LP1, LP2 y LP3, respectivamente; Figura 1; p < 0,05). Los animales experimentales del tratamiento LP3 lograron una eficiencia de retención de nitrógeno significativamente mayor que las aves del grupo C (p < 0.05). El contenido medio de materia seca de las excretas fue de 21,0, 23,9, 24,4 y 22,4% en los grupos de tratamiento C, LP1, LP2 y LP3, respectivamente, y mostró solo la tendencia al aumento en los grupos de LP en comparación con el tratamiento C (p = 0,109). Los tratamientos dietéticos no influyeron significativamente en la concentración de N fecal, ácido úrico-N, NH4+-N, N urinario y N total en los excrementos de pollos de engorde. La relación de N urinario dentro del N total en excretas fue de 43,1, 42,7, 42,3 y 42,7% en los grupos C, LP1, LP2 y LP3, respectivamente, y estos valores no se vieron afectados por los tratamientos dietéticos (ver Tabla 8).
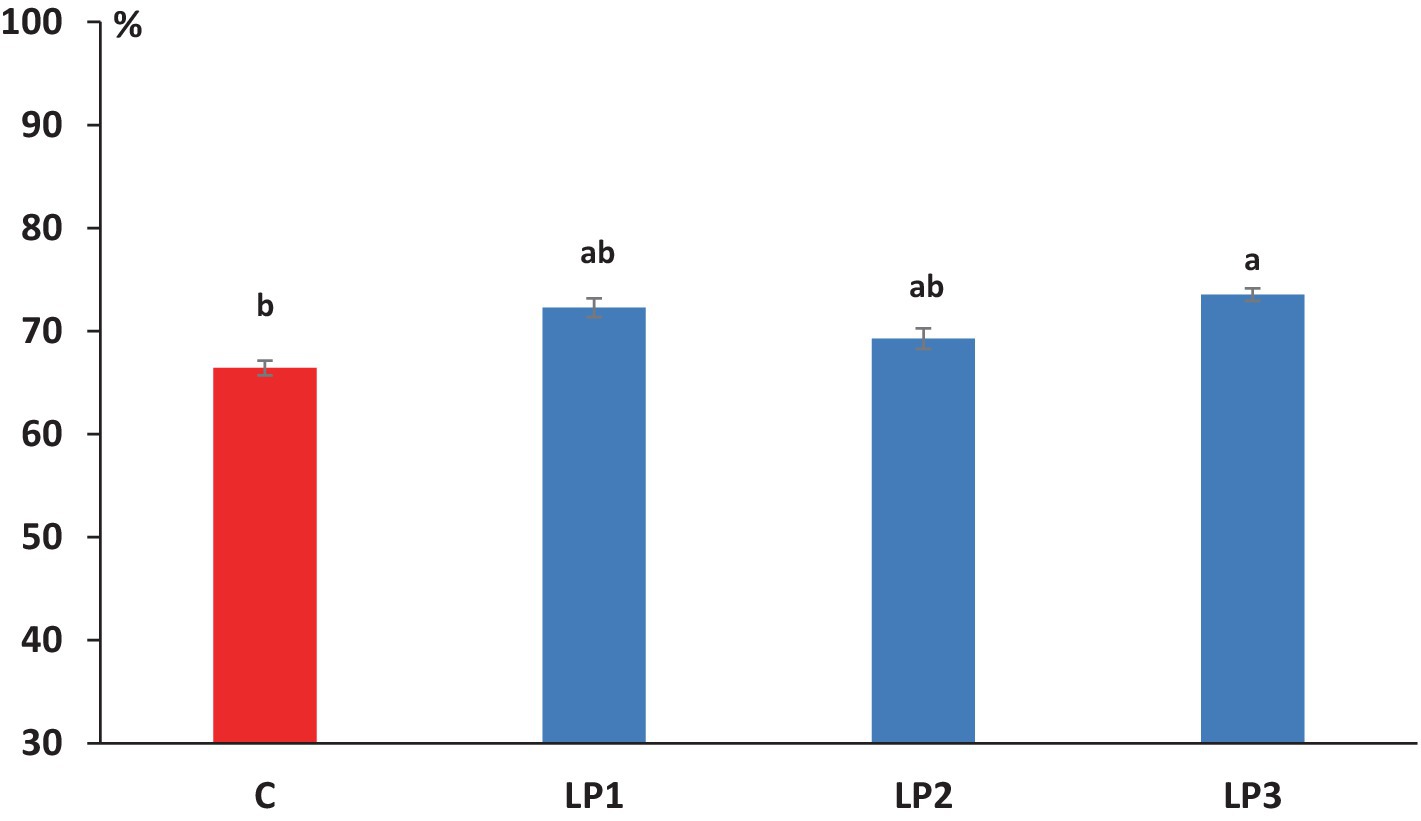 Figura 1. Efecto de los tratamientos dietéticos sobre la eficiencia de la retención de N en la dieta (media + SEM; n = 12 pollos de engorde por tratamiento). A,BLas medias con diferentes superíndices son significativamente diferentes; C – dieta de control; LP1 – dieta a base de harina de soja con niveles reducidos de proteína bruta (−2%); LP2 – dieta a base de harina de soja con niveles reducidos de proteína bruta (-2%) y mayor suplementación con L-treonina cristalina; LP3 – dieta con niveles reducidos de proteína bruta (−2%) en la que la harina de soja se sustituyó parcialmente por harina de carne de cerdo como fuente de proteína.
Figura 1. Efecto de los tratamientos dietéticos sobre la eficiencia de la retención de N en la dieta (media + SEM; n = 12 pollos de engorde por tratamiento). A,BLas medias con diferentes superíndices son significativamente diferentes; C – dieta de control; LP1 – dieta a base de harina de soja con niveles reducidos de proteína bruta (−2%); LP2 – dieta a base de harina de soja con niveles reducidos de proteína bruta (-2%) y mayor suplementación con L-treonina cristalina; LP3 – dieta con niveles reducidos de proteína bruta (−2%) en la que la harina de soja se sustituyó parcialmente por harina de carne de cerdo como fuente de proteína.
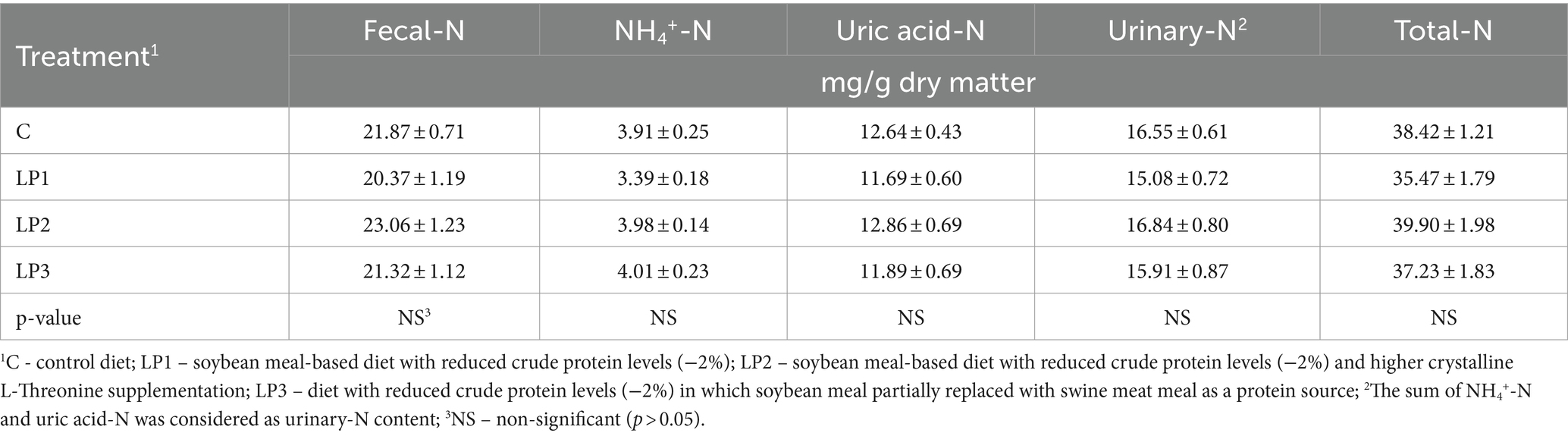 Tabla 8. La concentración de formas N en los excrementos de pollos de engorde (media ± SEM; n = 12 pollos de engorde por tratamiento).
Tabla 8. La concentración de formas N en los excrementos de pollos de engorde (media ± SEM; n = 12 pollos de engorde por tratamiento).
4 Discusión
En nuestro estudio, los tratamientos dietéticos de LP se aplicaron solo a partir de la fase de crecimiento. Se administró una dieta de inicio común porque una reducción temprana de la PC habría tenido un impacto mínimo en la reducción total de la PC y la harina de soja, la emisión de nitrógeno y el rendimiento final. En la mayoría de los ensayos, la reducción de la PC en la dieta en un 2% se pudo lograr sin alterar los rasgos de producción cuando se utilizó al menos la alimentación trifásica, se administraron dietas peletizadas y las dietas se equilibraron en al menos seis AA limitantes (15, 30, 31). En nuestro estudio, todos los rasgos de rendimiento de los pollos de engorde alimentados con dietas LP cumplieron o superaron los parámetros de rendimiento de las aves de control alimentadas con la dieta C. Los contenidos esenciales de AA de las dietas se establecieron de acuerdo con el concepto de proteína ideal, y la formulación se basó en los requerimientos de AA totales y SID. En cuanto a la suplementación con AA esencial cristalino, no solo se utilizaron los cuatro primeros aminoácidos limitantes (Lys, Met, Thr y Val) sino también L-Arg y L-Ile. Los pollos de engorde alimentados con dietas LP tuvieron un rendimiento generalmente bajo en algunos estudios, lo que puede explicarse por el hecho de que no se utilizaron L-Val, L-Arg y L-Ile cristalinos (32, 33). En algunos estudios, cuando las dietas de LP proporcionaron un suministro desequilibrado de AA esencial, las aves regularon su ingesta desequilibrada de AA a través de la hiperfagia, y se observó un FI significativamente más alto (34, 35). La alimentación con dietas LP en nuestro estudio no condujo a un aumento significativo de la IF en comparación con el tratamiento C. Además, se obtuvo la misma ingesta prevista de SID AA en el caso de seis AA esenciales principales y las diferencias específicas de la ingesta de SID Thr y Gly + Ser de pollos de engorde entre los grupos LP y C.
El rendimiento de los pollos de engorde del grupo LP1 fue similar o incluso superior (BWG en la fase de engorde) en comparación con los del grupo C tanto en la fase de engorde como en la de engorde. Significa que el grado de disminución del suministro de Gly (Glyequi de 15.65 vs. 13.74 y 14.19 vs. 12.30 en las fases de engorde y engorde, respectivamente) mientras que el cumplimiento de los requerimientos de Thr de las aves no afectó el rendimiento de los pollos de engorde de manera significativa. La explicación de la mayor BWG de las aves en la fase de engorde no está clara, pero puede asociarse con el aumento de la digestibilidad de AA en la dieta LP1 en comparación con la dieta C. Liu et al. (4) demostraron que la alimentación reducida con PC generalmente aumenta la digestibilidad del AA en el yeyuno distal, probablemente debido a la mayor proporción de AA cristalino esencial que se encuentra en las dietas reducidas en proteínas. En la fase de engorde, la mejora del FCR de los pollos de engorde en el grupo LP3 en comparación con el grupo C fue incluso mejor que la del grupo LP1. Este resultado sugiere que un aumento en el Glyequi de las dietas de LP de engorde de 12,3 a 13,55 g/kg es aconsejable en relación con el FCR de los pollos de engorde. De manera similar a nuestros resultados, estudios previos han demostrado que el aumento del suministro de Gly tuvo el mayor impacto en la mejora de la FCR (36, 37), posiblemente al mejorar la utilización de nutrientes y la síntesis de proteínas mediante el desarrollo de enterocitos y la secreción de mucina (38). De acuerdo con la bibliografía existente, el requisito de Glyequi Se estima que los pollos de engorde de 0 a 21 días de edad varían entre 11 y 20 g/kg dependiendo de la formación de ácido úrico, el suministro de Thr, la relación Met-a-(Met+Cys) y el nivel de colina (22, 23). Otros estudios han encontrado un óptimo Gly+Ser que oscila entre el 1,8 y el 2,3% de 0 a 35 días de edad (16, 18, 19). El nivel dietético de Gly + Ser necesario para optimizar la FCR de 21 a 35 días en dietas LP se estimó en 1,54% al nivel de Thr digestible de 0,77% (19). La concentración calculada del 1,57% de Gly + Ser en nuestra dieta LP3 de engorde, junto con el nivel de Thr de SID del 0,65%, parece ser baja, y podría ser posible un aumento efectivo adicional. Las proporciones mínimas de SID (Gly + Ser)/Lys en las fases de engorde 2 (21-31 días) y de engorde (31-41 días) son de 1,42 y 1,40, respectivamente, según lo sugerido por Rostagno et al. (39) y confirmado por Mansilla et al. (40). Los efectos positivos del aumento de la relación SID (Gly + Ser) a Lys en nuestra dieta LP3 de engorde en comparación con la dieta LP1 (1,30 vs. 1,24) en el FCR de los pollos de engorde respaldan un aumento adicional hacia las proporciones SID (Gly + Ser) a Lys sugeridas.
La relación entre la dieta Glyequi y Thr en cuanto a los rasgos de rendimiento se ha demostrado (17, 18) y cuantificado para la fase de 7 a 21 días (22, 23). La disminución de Glyequi deteriora el BWG y el FCR de los pollos de engorde de manera lineal y no lineal, respectivamente, y el efecto negativo es más pronunciado a un nivel dietético más bajo de Thr. En el caso de la FCR, la respuesta positiva de las aves al mismo aumento del nivel de Thr en la dieta es mayor en el nivel más bajo de Glyequi concentración. Además, el efecto positivo de un aumento de 1 g/kg en el nivel de Thr en la dieta sobre la FCR es mayor que el efecto del mismo aumento de Glyequi concentración en el alimento. De acuerdo con la relación, es posible mejorar el FCR aumentando el suministro de Thr mientras que el Glyequi está disminuyendo en la dieta. El tratamiento LP2 significó cambios similares en la dieta Thr y Glyequi en comparación con el tratamiento C en nuestro estudio, y se observaron efectos positivos en el rendimiento de las aves. Este efecto positivo seguía siendo una tendencia en la fase de crecimiento, pero se hizo significativo al final del engorde. La mejora del rendimiento del tratamiento con LP2 significa que el aumento de la concentración de SID Thr en un 0,10% (0,74% frente a 0,84% en el engorde y 0,65% frente al 0,74% en la fase de engorde) y el aumento de la relación SID Thr-to-Lys en un 9,0% (63 frente a 72% en el engorde y 64% frente a 73% en la fase de engorde) en comparación con un tratamiento control puede ser beneficioso cuando la PC dietética se reduce en un 2% y Glyequi en 2 g/kg. El aumento del suministro de aves con Thr en el grupo de tratamiento con LP2 podría apoyar las funciones primarias de Thr para la síntesis de proteínas. La conversión enzimática de Thr a Gly, el llamado efecto de reemplazo, también puede explicar estos resultados (23). Además, la reducción del catabolismo de AA que no sea Thr durante el aumento del suministro de Thr puede reducir la necesidad de Glyequi en la formación de ácido úrico, lo que podría servir como otro efecto ahorrador de Gly (23). Desafortunadamente, la relación entre la dieta Glyequi y el Thr en cuanto a las características de rendimiento para todo el engorde desde la eclosión hasta los 35 o 42 días de edad no se cuantifica con tanta precisión como se hizo para la fase de 7 a 21 días (22). Sin embargo, interacciones similares entre Thr y Glyequi puede existir hasta el final de la fase de acabado en función de nuestros resultados. Del mismo modo, estudios previos centrados en la segunda mitad del engorde confirmaron la interacción Thr-Gly mencionada anteriormente, en la que la respuesta positiva del rendimiento de las aves al mismo aumento del nivel de Thr en la dieta es mayor al nivel más bajo de Glyequi concentración. La respuesta de FCR al nivel de Thr en la dieta fue más fuerte en el nivel más bajo de digestibilidad (Gly + Ser) a Lys (135 vs. 149%) (19). El BWG de pollos de engorde de 21 a 42 días respondió más para Thr cuando se alimentaron al 143% en comparación con el nivel de relación digestible (Gly + Ser) a Lys del 153% (17). Se necesitan más estudios para cuantificar los efectos de la interacción Thr-Gly en el rendimiento de los pollos de engorde durante todo el período de engorde.
En el presente estudio, los resultados de las aves en el tratamiento con LP1 están en línea con estudios previos similares que no muestran ningún efecto de las dietas esenciales de LP suplementadas con AA hasta una reducción del 2% de la PC en el rendimiento de la canal y las partes valiosas de la canal (30, 40-42). En el estudio de Mansilla et al. (40), las reducciones de la relación SID (Gly + Ser)-Lys paralelas a la reducción de 2% de PC en comparación con la dieta control fueron de 5.0, 9.0 y 12.0% en las dietas de engorde1, engorde2 y engorde, respectivamente. Las reducciones de la relación SID (Gly + Ser)-Lys fueron mayores (15,0% en el engorde y 19% en el engorde) en el presente experimento, pero los resultados del peso en canal fueron similares en ambos estudios. Solo se han publicado unos pocos resultados sobre los efectos de la interacción Thr-Gly en la calidad del producto de pollos de engorde alimentados con dietas LP. De manera similar a nuestros resultados observados en el grupo LP2, el aumento de la concentración de Thr digestible de 0.57 a 0.65% mientras que la disminución del nivel de Gly + Ser de 1.65 a 1.55% no influyó en el peso relativo de la canal y el peso del filete de pechuga de las aves alimentadas con una dieta LP con 18.2% de PC de 21 a 42 días de edad (17). Un aumento de la Thr en la dieta de 9,3 a 10,7 g/kg puede incluso disminuir el peso relativo de la mama, lo que puede deberse al efecto tóxico del aumento de las concentraciones plasmáticas de ácido úrico y amoníaco (18). En el mismo estudio, hubo un efecto lineal significativo positivo del aumento de las concentraciones de Gly + Ser de 18,4 a 22,6 g/kg en el peso relativo de la pechuga de los pollos de engorde alimentados con una dieta LP que contenía 19% de PC de 0 a 21 días (18).
Nuestros resultados sugieren que los efectos negativos significativos del tratamiento con LP3 sobre el peso y la composición de la canal podrían estar asociados con la relación almidón/PB de las dietas experimentales. El contenido de almidón como principal nutriente proveedor de energía suele aumentar cuando se reduce la PC dietética en las dietas isocalóricas de LP (4). Por el contrario, el nivel de lípidos en la dieta suele disminuir con el nivel de proteínas, que también fue el caso en este experimento. La relación almidón/PB aumentó en los tratamientos con LP en comparación con el tratamiento con C, y fue la más alta en la dieta LP3 que contenía harina de carne de cerdo. La mayor relación almidón/PC deterioró el valor de FCR en la fase inicial de nuestro experimento anterior de forma cuadrática (43). Se observó una relación cuadrática similar entre estos dos factores de 7 a 35 días en dos estudios (44, 45). En el presente estudio, la mayor relación almidón/PC de las dietas LP no afectó la FCR de las aves en comparación con la dieta C. Sin embargo, en nuestra opinión, la mayor relación almidón/PC en las dietas LP3 (2,25 y 2,63 en las fases de engorde y engorde, respectivamente) dio lugar a un peso relativo en canal significativamente menor en comparación con las dietas C (1,67 y 2,0 en las fases de engorde y engorde, respectivamente). Este efecto podría verse como una fuerte tendencia en el caso del rendimiento de carne de pechuga. Con base en la dinámica digestiva del almidón dietético, se ha demostrado que la absorción de su contenido de glucosa compite con la absorción de AA, lo que puede afectar la disponibilidad de AA para la acreción de proteínas tisulares (46, 47). Sin embargo, Hilliar et al. (48, 49) encontraron que la adición de Gly cristalino en una dieta LP redujo el rendimiento de carne de pechuga en comparación con el control sin Gly adicional. En este caso, se puede excluir el efecto teórico del almidón como explicación, pero la causa no está clara. A diferencia de los resultados de muchos ensayos previos (47, 50, 51), la alimentación con la dieta isoenergética LP1 y LP2 no aumentó significativamente la almohadilla de grasa abdominal en comparación con la dieta C. Sin embargo, también se observó una ligera tendencia negativa en estos dos tratamientos, lo que tuvo un efecto negativo significativo en el grupo de tratamiento con LP3. La relación AMEn-PC/PC en la dieta aumenta mientras se mantiene constante la AMEn dietética de las dietas LP, y el exceso de energía puede aumentar la grasa abdominal (50, 51). En otros estudios, la reducción de la energía dietética y la PC mientras se mantenía la misma proporción de AMEn-PC previno con éxito la acumulación de grasa abdominal, pero se suprimió el rendimiento de crecimiento de los pollos de engorde (52, 53). La proporción de AMEn a PC de las dietas LP fue casi la misma, pero solo el tratamiento con LP3 resultó en un aumento significativo en la proporción de almohadillas de grasa abdominal. El uso de suplementos sintéticos de Gly puede reducir la grasa de los pollos de engorde alimentados con dietas LP (37, 54, 55). De acuerdo con estudios con aves de corral, ratas y cerdos, se ha demostrado que el aumento de la glico-glucosa o betaína (trimetilglicina) estimula la oxidación de lípidos y reduce las concentraciones plasmáticas de triglicéridos y la deposición de grasa (56-58). Por el contrario, el uso de harinas de carne de cerdo para aumentar la concentración de Gly + Ser de las dietas LP3 condujo a resultados opuestos en el presente estudio. Suponemos que la mayor relación almidón/PB en la dieta LP3 en comparación con las dietas LP1 y LP2 se asociócon el aumento significativo de la proporción de grasa abdominal. Si la acumulación de proteínas tisulares disminuyó en el grupo LP3 debido a la alta relación almidón/PC, como se supone en función del resultado del peso de la canal, el exceso de energía formada podría conducir a una acumulación de almohadilla de grasa abdominal de manera significativa. Otra posible explicación es la elevada concentración hepática de acetil-CoA derivada de los niveles relativamente altos de almidón en la dieta de las aves a las que se les ofrecieron dietas LP (4). El acetil-CoA puede servir como precursor de la síntesis de ácidos grasos e influir en la actividad de numerosas enzimas (4). Estas consecuencias negativas de las dietas de LP suplementadas con 6 y 4% de harina de carne de cerdo sobre el peso de la canal y la almohadilla de grasa abdominal necesitan investigaciones más enfocadas. Además del equilibrio dietético de AA, la dinámica digestiva de los principales nutrientes, especialmente el almidón, los lípidos y las proteínas, debe tenerse en cuenta en el desarrollo posterior de las dietas LP.
Como sabemos, no se han investigado los efectos de la interacción Thr-Gly en la dieta sobre la calidad de la carne de pollos de engorde alimentados con dietas LP. El pH y la pérdida por goteo de la carne de pechuga en el presente experimento no mostraron cambios significativos debido a los tratamientos dietéticos de LP con diferentes Thr y Glyequi Suministros. La pérdida por goteo es uno de los parámetros que se asocia a la capacidad de retención de agua de la carne e influye en su calidad sensorial y tecnológica. La relación negativa entre la pérdida por goteo y el pH final en la carne de ave es bien conocida (52, 59). La alimentación con dietas LP puede resultar en un pH final más alto y una disminución de la pérdida por goteo (43, 60). La mayor pérdida por goteo de la carne resultó ser más ácida con un mayor nivel de glucosa, glucógeno y potenciales glucolíticos (52, 53, 59). La descomposición post-mortem del glucógeno acumulado en los tejidos musculares es responsable de la acidez adecuada de la carne después del sacrificio. Si no se dispone de una cantidad adecuada de glucógeno, el pH de la carne se vuelve menos ácido y la capacidad de retención de agua de la carne es mayor. La pérdida por goteo de carne podría estar asociada con un exceso de AA (60). Después de la desaminación de los AA no utilizados, la cadena de carbono es utilizada por el tejido muscular para la síntesis de diversos carbohidratos, como el glucógeno. Esto sugiere que nuestras dietas experimentales proporcionaron un perfil de AA equilibrado similar sin o con una cantidad similar de exceso de AA.
La eficiencia de retención de N en la dieta de los pollos de engorde medida en nuestro experimento mejoró en un 3-7% con las dietas LP en comparación con la dieta C, y esta mejora está en línea con los resultados publicados anteriormente (2-13%) de las dietas LP que proporcionan un suministro adecuado de AA esencial (40, 61, 62). Sin embargo, la eficiencia de retención de N de los pollos de engorde no ha sido reportada por los estudios dosis-respuesta con dietas LP suplementadas con Thr-Gly (17-19, 21). El aumento medio de la eficiencia de la retención de N fue de +2,63%/punto porcentual de PC, un poco inferior al valor correspondiente de +3,2% publicado por Belloir et al. (15). Las mejoras fueron fuertes tendencias en los grupos LP1 y LP2 en comparación con el grupo C, y fue una diferencia significativa entre los tratamientos LP3 y C. De acuerdo con nuestro resultado, el aumento del nivel de Glyequi en dietas de PL con una reducción de PC del 2% se puede recomendar para aumentar la eficiencia de retención de N, ya que también fue recomendable aumentar el FCR de las aves de engorde. Se midieron valores de eficiencia extremadamente ventajosos superiores al 70% en los tratamientos LP1 y LP3, que también han sido reportados por otros grupos de investigación (14, 15). El aumento de la eficiencia de utilización de N disminuye la Gly dietéticaequi necesario para la síntesis de ácido úrico basado en cálculos de modelos (63, 64). La estimación muestra que una eficiencia de utilización de N superior al 70% requiere solo menos de 10 g/kg Glyequi para la producción de ácido úrico. La mayor parte del amoníaco liberado por el estiércol de aves de corral se origina en la descomposición del ácido úrico (65). La alimentación con dietas LP puede disminuir el nivel de ácido úrico del plasma sanguíneo (21) y las concentraciones de N total y urinario y ácido úrico en los excrementos de pollos de engorde (15, 31, 43). Sin embargo, los resultados del presente experimento no confirmaron los resultados ventajosos de estudios previos asociados con la emisión de N. La concentración de ácido úrico de los excrementos en los pollos de engorde alimentados con la dieta Control C ya era bastante baja (12,54 mg/g de materia seca), y tal vez no fuera posible una disminución adicional alimentados con dietas LP. En nuestros dos experimentos anteriores, las dietas LP redujeron los niveles de ácido úrico en los excrementos cuando se midieron niveles de ácido úrico más altos que en el presente ensayo en los excrementos de pollos de engorde alimentados con dietas normales de PC (17,65 y 15,30 mg/g de materia seca) (31, 43).
Además de las características de rendimiento, la eficacia económica de las dietas LP depende de los precios reales de las materias primas, especialmente la harina de soja y los suplementos de AA cristalinos. De acuerdo con los precios húngaros actuales, el costo de las dietas experimentales basadas únicamente en los ingredientes fue de 364, 362, 363 y 352 euros por tonelada en el caso del productor, y de 346, 343, 344 y 339 euros por tonelada en el caso de las dietas de engorde de los tratamientos C, LP1, LP2 y LP3, respectivamente. Todas las dietas LP tuvieron precios más bajos que la dieta C, y asumiendo un rendimiento similar de las aves alimentadas con dietas C y LP, pueden contribuir a una mayor rentabilidad de la producción de pollos de engorde.
5 Conclusión
De acuerdo con los resultados de este experimento, el aumento de la relación SID Thr-to-Lys y Glyequi de las dietas de PL con un contenido de PB un 2% inferior al adecuado pueden tener efectos positivos en el rendimiento de los pollos de engorde y en la eficiencia de retención de nitrógeno. La relación SID Thr-to-Lys de las dietas de LP más alta que la recomendada puede mejorar el peso corporal, el peso corporal y el FCR finales de las aves. La harina de carne de cerdo como fuente de Gly se puede utilizar para aumentar Glyequi en dietas prácticas de LP en la UE, lo que puede conducir a una FCR más ventajosa en la fase de engorde y a una mayor eficiencia de retención de N. Sin embargo, las dietas LP que contienen harina de carne de cerdo pueden tener una alta proporción de almidón a PB, lo que puede contribuir a una disminución del peso relativo de la canal y a un aumento de la proporción de almohadillas de grasa abdominal de los pollos de engorde a la edad comercial.
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán puestos a disposición por los autores, sin reservas indebidas.
Declaración ética
El estudio en animales fue aprobado por el Comité de Bienestar Animal de la Universidad Húngara de Agricultura y Ciencias de la Vida, Campus Georgikon, con el número de licencia MÁB-3/2022. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
PD: Curación de datos, Análisis formal, Metodología, Administración de proyectos, Redacción – borrador original. BH: Curación de datos, Análisis formal, Metodología, Administración de proyectos, Redacción – borrador original. NS: Curación de datos, Análisis formal, Metodología, Administración de proyectos, Redacción – borrador original. KD: Recursos, Supervisión, Validación, Redacción – borrador original, Redacción – revisión y edición. LP: Conceptualización, Investigación, Metodología, Administración de Proyectos, Recursos, Supervisión, Validación, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue apoyada por la beca de doctorado de PS.
Reconocimientos
Los autores desean expresar su gratitud y aprecio a quienes ayudaron con esta investigación en el Instituto de Fisiología y Nutrición de la Universidad Húngara de Agricultura y Ciencias de la Vida.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
References
1. Nahm, KH . Evaluation of the nitrogen content in poultry manure. Worlds Poult Sci J. (2003) 59:77–88. doi: 10.1079/WPS20030004
2. European CommissionJoint Research CentreGeorgitzikis, K, Giner Santonja, G, Roudier, S, Montobbio, P, et al. Best Available Techniques (BAT) reference document for the intensive rearing of poultry or pigs – Industrial Emissions Directive 2010/75/EU (Integrated Pollution Prevention and Control). Publications Office (2017). Available at: https://data.europa.eu/doi/10.2760/020485
3. Wu, S-B, Stanley, D, Rodgers, N, Swick, RA, and Moore, RJ. Two necrotic enteritis predisposing factors, dietary fishmeal and Eimeria infection, induce large changes in the caecal microbiota of broiler chickens. Vet Microbiol. (2014) 169:188–97. doi: 10.1016/j.vetmic.2014.01.007
4. Liu, SY, Macelline, SP, Chrystal, PV, and Selle, PH. Progress towards reduced-crude protein diets for broiler chickens and sustainable chicken-meat production. J Anim Sci Biotechnol. (2021) 12:20. doi: 10.1186/s40104-021-00550-w
5. Kidd, MT, Maynard, CW, and Mullenix, GJ. Progress of amino acid nutrition for diet protein reduction in poultry. J Anim Sci Biotechnol. (2021) 12:45. doi: 10.1186/s40104-021-00568-0
6. Fernandez, SR, Aoyagi, S, Han, Y, Parsons, CM, and Baker, DH. Limiting order of amino acids in corn and soybean meal for growth of the Chick. Poult Sci. (1994) 73:1887–96. doi: 10.3382/ps.0731887
7. Kidd, MT . Nutritional considerations concerning threonine in broilers. Worlds Poult Sci J. (2000) 56:139–51. doi: 10.1079/WPS20000011
8. Dozier, WA, Meloche, KJ, Tillman, PB, and Jiang, Z. Growth performance of male broilers fed diets varying in digestible threonine to lysine ratio from 1 to 14 days of age1. J Appl Poult Res. (2015) 24:457–62. doi: 10.3382/japr/pfv047
9. Star, L, Rovers, M, Corrent, E, and van der Klis, JD. Threonine requirement of broiler chickens during subclinical intestinal Clostridium infection. Poult Sci. (2012) 91:643–52. doi: 10.3382/ps.2011-01923
10. Tang, Q, Tan, P, Ma, N, and Ma, X. Physiological functions of threonine in animals: beyond nutrition metabolism. Nutrients. (2021) 13:2592. doi: 10.3390/nu13082592
11. Sugahara, M, and Kandatsu, M. Glycine serine interconversion in the rooster. Agric Biol Chem. (1976) 40:833–7. doi: 10.1080/00021369.1976.10862153
12. Meléndez-Hevia, E, De Paz-Lugo, P, Cornish-Bowden, A, and Ml, C. A weak link in metabolism: the metabolic capacity for glycine biosynthesis does not satisfy the need for collagen synthesis. J Biosci. (2009) 34:853–72. doi: 10.1007/s12038-009-0100-9
13. Dean, DW, Bidner, TD, and Southern, LL. Glycine supplementation to low protein, amino acid-supplemented diets supports optimal performance of broiler Chicks1. Poult Sci. (2006) 85:288–96. doi: 10.1093/ps/85.2.288
14. Siegert, W, Wild, KJ, Schollenberger, M, Helmbrecht, A, and Rodehutscord, M. Effect of glycine supplementation in low protein diets with amino acids from soy protein isolate or free amino acids on broiler growth and nitrogen utilisation. Br Poult Sci. (2016) 57:424–34. doi: 10.1080/00071668.2016.1163523
15. Belloir, P, Méda, B, Lambert, W, Corrent, E, Juin, H, Lessire, M, et al. Reducing the CP content in broiler feeds: impact on animal performance, meat quality and nitrogen utilization. Animal. (2017) 11:1881–9. doi: 10.1017/S1751731117000660
16. Waldroup, PW, Jiang, Q, and Fritts, CA. Effects of Glycine and threonine supplementation on performance of broiler chicks fed diets low in crude protein. Int J Poult Sci. (2005) 4:250–7. doi: 10.3923/ijps.2005.250.257
17. Corzo, A, Kidd, MT, Dozier, WA, y Kerr, BJ. Efectos interactivos de la glicina y la treonina en la dieta en pollos de engorde. J Appl Poult Res. (2009) 18:79–84. doi: 10.3382/japr.2008-00078
18. Ospina-Rojas, IC, Murakami, AE, Moreira, I, Picoli, KP, Rodrigueiro, RJB y Furlan, AC. Respuestas dietéticas de glicina + serina de pollos de engorde machos que recibieron dietas bajas en proteínas con diferentes concentraciones de treonina. Hno. Poult Sci. (2013) 54:486–93. doi: 10.1080/00071668.2013.794257
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Ospina-Rojas, IC, Murakami, AE, Oliveira, C, y Guerra, AFQG. Efectos suplementarios de glicina y treonina sobre el rendimiento, el desarrollo de la mucosa intestinal y la utilización de nutrientes de pollos de engorde en crecimiento. Poult Sci. (2013) 92:2724–31. doi: 10.3382/ps.2013-03171
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Siegert, W, Ahmadi, H y Rodehutscord, M. Metaanálisis de la influencia de la glicina y la serina en la dieta, teniendo en cuenta la metionina y la cisteína, en el crecimiento y la conversión alimenticia de los pollos de engorde. Poult Sci. (2015) 94:1853–63. doi: 10.3382/ps/pev129
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Star, L, Tesseraud, S, van Tol, M, Minussi, I, Corrent, E y Lambert, W. Rendimiento de la producción y concentraciones plasmáticas de metabolitos de pollos de engorde alimentados con dietas bajas en proteína bruta que difieren en Thr y Gly. Anim Nutr. (2021) 7:472–80. doi: 10.1016/j.aninu.2020.09.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Siegert, W, Ahmadi, H, Helmbrecht, A, y Rodehutscord, M. Un estudio cuantitativo de los efectos interactivos de la glicina y la serina con treonina y colina sobre el rendimiento del crecimiento en pollos de engorde. Poult Sci. (2015) 94:1557–68. doi: 10.3382/ps/pev109
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Siegert, W, y Rodehutscord, M. La relevancia de la glicina y la serina en la nutrición de las aves: una revisión. Hno. Poult Sci. (2019) 60:579–88. doi: 10.1080/00071668.2019.1622081
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Peters, J, Combs, S, Hoskins, B, Jarman, J, Kovar, J, Watson, M, et al. «Métodos recomendados para el análisis del estiércol», Actas de los resúmenes de la reunión anual de ASA-CSSA-SSSA. Madison, WI, ESTADOS UNIDOS: ASA-CSSASSSA (2003). págs. 25-29.
25. Marquardt, RR, Ward, AT, y Campbell, LD. Método cromatográfico líquido rápido y de alta resolución para la cuantificación de ácido úrico en muestras de excretas y tejidos. Poult Sci. (1983) 62:2099–105. doi: 10.3382/ps.0622099
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. O’dell, BL, Woods, WD, Laerdal, OA, Jeffay, AM, y Savage, JE. Distribución de los principales compuestos nitrogenados y aminoácidos en la orina de pollo1. Poult Sci. (1960) 39:426–32. doi: 10.3382/ps.0390426
27. Short, FJ, Gorton, P, Wiseman, J y Boorman, KN. Determinación de dióxido de titanio añadido como marcador inerte en estudios de digestibilidad de pollos. Anim Feed Sci Technol. (1996) 59:215–21. doi: 10.1016/0377-8401(95)00916-7
28. Scott, ML, Nesheim, MC y Young, RJ. Nutrición del pollo. (1976) Disponible en: https://www.cabdirect.org/cabdirect/abstract/19771453582 (consultado el 14 de febrero de 2023).
29. Northcutt, JK, Foegeding, EA, y Edens, FW. Propiedades de retención de agua de la carne de pechuga y pierna de pollo preacondicionada térmicamente1. Poult Sci. (1994) 73:308–16. doi: 10.3382/ps.0730308
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. van Harn, J, Dijkslag, MA, y van Krimpen, MM. Efecto de las dietas bajas en proteínas suplementadas con aminoácidos libres sobre el rendimiento del crecimiento, el rendimiento del sacrificio, la calidad de la camada y las lesiones de las almohadillas de los pollos de engorde machos. Poult Sci. (2019) 98:4868–77. doi: 10.3382/ps/pez229
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Tales, N, Pál, L, Strifler, P, Horváth, B, Koltay, IA, Rawash, MA, et al. Efecto de la alimentación con dietas bajas en proteínas sobre las características de producción y la composición de nitrógeno de los excrementos de pollos de engorde. Agricultura. (2021) 11:781. doi: 10.3390/agriculture11080781
32. Khajali, F, y Moghaddan, HN. Suplementación con metionina en dietas bajas en proteínas para pollos de engorde: influencia sobre el rendimiento del crecimiento y la eficiencia de la utilización de proteínas. Int J Poult Sci. (2006) 5:569–73. doi: 10.3923/ijps.2006.569.573
33. Guaiume, EA . Efectos de las dietas reducidas en proteínas y suplementos de aminoácidos sobre la producción y el rendimiento económico de pollos de engorde comerciales alimentados desde la eclosión hasta la edad comercial. [Ph. D. Columbia: Universidad de Missouri (2007).
34. Smith, ER, y Pesti, GM. Influencia del cruce de cepas de pollos de engorde y la proteína de la dieta en el rendimiento de los pollos de engorde. Poult Sci. (1998) 77:276–81. doi: 10.1093/ps/77.2.276
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Swennen, Q, Decuypere, E y Buyse, J. Implicaciones de los macronutrientes dietéticos para el crecimiento y el metabolismo en pollos de engorde. Worlds Poult Sci J. (2007) 63:541–56. doi: 10.1017/S0043933907001602
36. Kriseldi, R, Tillman, PB, Jiang, Z, y Dozier, WA. Efectos de la suplementación con glicina y glutamina en dietas reducidas de proteína cruda sobre el rendimiento del crecimiento y las características de la canal de pollos de engorde machos durante un período de producción de 41 días1. J Appl Poult Res. (2017) 26:558–72. doi: 10.3382/japr/pfx030
37. Lee, DT, Lee, JT, Ruan, C, y Rochell, SJ. Influencia del aumento de las concentraciones de glicina en dietas reducidas de proteína bruta alimentadas a pollos de engorde de 0 a 48 días. Poult Sci. (2022) 101:102038. doi: 10.1016/j.psj.2022.102038
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Wang, W-W, Wang, J, Wu, S-G, Zhang, H-J y Qi, G-H. Respuesta de los pollos de engorde a la reducción gradual de proteínas en la dieta con o sin un nivel adecuado de glicina más serina. Ital J Anim Sci. (2020) 19:127–36. doi: 10.1080/1828051X.2019.1704634
39. Rostagno, HS, Albino, LFT, Donzele, JL, Gomes, PC, Oliveira, RT, Lopes, DC, et al. Mesas brasileñas para aves de corral y cerdos. Composición de los piensos y necesidades nutricionales. 4ª edición Viçosa, MG, Brasil: Universidade Federal de Viçosa (2017).
40. Mansilla, WD . Reducción de proteínas en la dieta con adición escalonada de aminoácidos cristalinos y el efecto de considerar un contenido mínimo de glicina-serina en las dietas de pollos de engorde. Poult Sci. (2023) 102:102684. doi: 10.1016/j.psj.2023.102684
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Lemme, A, Hiller, P, Klahsen, M, Taube, V, Stegemann, J y Simon, I. La reducción de la proteína dietética en las dietas de los pollos de engorde no solo reduce las emisiones de n, sino que también va acompañada de varios beneficios adicionales. J Appl Poult Res. (2019) 28:867–80. doi: 10.3382/japr/pfz045
42. Ospina-Rojas, IC, Murakami, AE, Duarte, CRA, Eyng, C, Oliveira, C, y Janeiro, V. Suplementación con valina, isoleucina, arginina y glicina de dietas bajas en proteínas para pollos de engorde durante las fases de inicio y crecimiento. Hno. Poult Sci. (2014) 55:766–73. doi: 10.1080/00071668.2014.970125
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Strifler, P, Horváth, B, Such, N, Farkas, V, Wágner, L, Dublecz, K, et al. Efectos de la alimentación con dietas bajas en proteínas con diferentes proporciones de energía a proteína sobre el rendimiento, las características de la canal y la excreción de nitrógeno de los pollos de engorde. Animales. (2023) 13:1476. doi: 10.3390/ani13091476
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Chrystal, PV, Moss, AF, Khoddami, A, Naranjo, VD, Selle, PH y Liu, SY. Impactos de las dietas reducidas en proteína cruda en parámetros clave en pollos de engorde machos ofrecidos a dietas a base de maíz. Poult Sci. (2020) 99:505–16. doi: 10.3382/ps/pez573
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Chrystal, PV, Moss, AF, Khoddami, A, Naranjo, VD, Selle, PH y Liu, SY. Los efectos de la reducción de los niveles de proteína cruda, el equilibrio electrolítico de la dieta y la densidad energética en el rendimiento de los pollos de engorde ofrecieron dietas a base de maíz con evaluaciones del metabolismo del almidón, las proteínas y los aminoácidos. Poult Sci. (2020) 99:1421–31. doi: 10.1016/j.psj.2019.10.060
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. van der Meulen, J, Bakker, JG, Smits, B, y de Visser, H. Efectos de la fuente de almidón en el flujo portal neto de glucosa, lactato, ácidos grasos volátiles y aminoácidos en el cerdo. Hno. J Nutr. (1997) 78:533–44. doi: 10.1079/bjn19970173
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Li, T-J, Dai, Q-Z, Yin, Y-L, Zhang, J, Huang, R-L, Ruan, Z, et al. Las fuentes de almidón de la dieta afectan la apariencia portal neta de aminoácidos y glucosa en cerdos en crecimiento. Animal. (2008) 2:723–9. doi: 10.1017/S1751731108001614
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Hilliar, M, Hargreave, G, Girish, CK, Barekatain, R, Wu, S-B y Swick, RA. Uso de aminoácidos cristalinos para complementar las necesidades de pollos de engorde en dietas reducidas en proteínas. Poult Sci. (2020) 99:1551–63. doi: 10.1016/j.psj.2019.12.005
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Hilliar, M, Huyen, N, Girish, CK, Barekatain, R, Wu, S, y Swick, RA. Suplementación con glicina, serina y treonina en dietas bajas en proteínas para pollos de tipo carne. Poult Sci. (2019) 98:6857–65. doi: 10.3382/ps/pez435
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Rosebrough, RW, y Steele, Carolina del Norte. Relaciones energéticas y proteicas en pollos de engorde: 1. Efecto de los niveles de proteínas y los regímenes de alimentación sobre el crecimiento, la composición corporal y la lipogénesis in vitro de pollos de engorde. Poult Sci. (1985) 64:119–26. doi: 10.3382/ps.0640119
51. Swennen, Q, Janssens, GPJ, Collin, A, Le Bihan-Duval, E, Verbeke, K, Decuypere, E, et al. Termogénesis inducida por la dieta y oxidación de la glucosa en pollos de engorde: influencia del genotipo y la composición de la dieta. Poult Sci. (2006) 85:731–42. doi: 10.1093/ps/85.4.731
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Bihan-Duval, EL, Alnahhas, N, Pampouille, E, Berri, C y Abasht, B. Genética y genómica de los rasgos de calidad de la carne en especies de aves de corral En: Avances en genética y genómica avícola. Londres, Reino Unido: Burleigh Dodds Science Publishing (2020).
53. Przybylski, W, Sałek, P, Kozłowska, L, Jaworska, D, y Stańczuk, J. El análisis metabolómico indica que una mayor pérdida por goteo puede estar relacionada con la producción de metilglioxal como subproducto de la glucólisis. Poult Sci. (2022) 101:101608. doi: 10.1016/j.psj.2021.101608
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Hejdysz, M, Bogucka, J, Ziółkowska, E, Perz, K, Jarosz, Ł, Ciszewski, A, et al. Efectos del bajo contenido de proteína cruda y la suplementación con glicina en el rendimiento de los pollos de engorde, las características de la canal y la calidad de la camada. Livest Sci. (2022) 261:104930. doi: 10.1016/j.livsci.2022.104930
55. Elahi, U, Wang, J, Ma, Y, Wu, S, Qi, G y Zhang, H. La respuesta de los pollos de engorde a la reducción de la harina de soja en la dieta con inclusión de glicina y cisteína en la deficiencia marginal de aminoácidos azufrados (SAA). Animales. (2020) 10:1686. doi: 10.3390/ani10091686
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Fouad, AM, y El-Senousey, HK. Factores nutricionales que afectan la deposición de grasa abdominal en aves de corral: una revisión. Australas Asiáticas J Anim Sci. (2014) 27:1057–68. doi: 10.5713/ajas.2013.13702
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. El Hafidi, M, Pérez, I, Zamora, J, Soto, V, Carvajal-Sandoval, G, y Baños, G. La ingesta de glicina disminuye los ácidos grasos libres plasmáticos, el tamaño de las células adiposas y la presión arterial en ratas alimentadas con sacarosa. Am J Physiol Regul Integr Comp Physiol. (2004) 287:R1387-93. doi: 10.1152/ajpregu.00159.2004
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Zhong, Y, Yan, Z, Song, B, Zheng, C, Duan, Y, Kong, X, et al. La suplementación dietética con betaína o glicina mejora las características de la canal, la calidad de la carne y el metabolismo lipídico de los minicerdos de engorde. Anim Nutr. (2021) 7:376–83. doi: 10.1016/j.aninu.2020.08.010
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Beauclercq, S, Hennequet-Antier, C, Praud, C, Godet, E, Collin, A, Tesseraud, S, et al. El análisis del transcriptoma muscular revela vías moleculares y biomarcadores implicados en la aparición de pH final extremo y defectos cárnicos en el pollo. Sci Rep. (2017) 7:6447. doi: 10.1038/s41598-017-06511-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Belloir, P, Lessire, M, Lambert, W, Corrent, E, Berri, C y Tesseraud, S. Cambios en la composición corporal y la calidad de la carne en respuesta al suministro de aminoácidos en la dieta en pollos de engorde de engorde. Animal. (2019) 13:1094–102. doi: 10.1017/S1751731118002306
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Aletor, VA, Hamid, II, Nieß, E y Pfeffer, E. Dietas suplementadas con aminoácidos bajas en proteínas en pollos de engorde: efectos sobre el rendimiento, las características de la canal, la composición de todo el cuerpo y la eficiencia de la utilización de nutrientes. J Sci Food Agric. (2000) 80:547–54. doi: 10.1002/(SICI)1097-0010(200004)80:5<547::AID-JSFA531>3.0.CO; 2-C
62. Jackson, S, Summers, JD y Leeson, S. Efecto de la proteína y la energía de la dieta en la composición de la canal de pollos de engorde y la eficiencia de la utilización de nutrientes. Poult Sci. (1982) 61:2224–31. doi: 10.3382/ps.0612224
Resumen de PubMed | Texto completo de Crossref | Google Académico
63. Goldstein, DL y Skadhauge, E. CAPÍTULO 11 – Regulación renal y extrarrenal de la composición de los fluidos corporales En: GC Whittow editor. Fisiología aviar de Sturkie. 5ª ed. San Diego: Academic Press (2000). 265–97.
64. Hofmann, P, Siegert, W, Kenéz, Á, Naranjo, VD y Rodehutscord, M. La proteína bruta muy baja y las concentraciones variables de glicina en la dieta afectan el rendimiento del crecimiento, las características de la excreción de nitrógeno y el metaboloma sanguíneo de los pollos de engorde. J Nutr. (2019) 149:1122–32. doi: 10.1093/jn/nxz022
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: pollos de engorde, dieta baja en proteínas, treonina, glicina, nitrógeno
Cita: Strifler P, Horváth B, Such N, Dublecz K y Pál L (2024) Efectos de diferentes suministros dietéticos de treonina y glicina en pollos de engorde alimentados con dietas bajas en proteínas. Frente. Vet. Sci. 11:1373348. doi: 10.3389/fvets.2024.1373348
Editado por:
Tugay Ayasan, Universidad Osmaniye Korkut Ata, Türkiye
Revisado por:
Majid Shakeri, Departamento de Agricultura de los Estados Unidos, Estados
Unidos Ilias Giannenas, Universidad Aristóteles de Tesalónica, Grecia
Anna Stępniowska, Universidad de Ciencias de la Vida de Lublin, Polonia
Derechos de autor © 2024 Strifler, Horváth, Such, Dublecz y Pál. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: László Pál, pal.laszlo@uni-mate.hu
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía