
Efectos de la meteorología y el ciclo lunar en la calidad post-descongelación del esperma aviar
 Efectos de la meteorología y el ciclo lunar en la calidad post-descongelación del esperma aviar
Efectos de la meteorología y el ciclo lunar en la calidad post-descongelación del esperma aviar
 Esther Díaz Ruizç1
Esther Díaz Ruizç1  Juan Vicente Delgado Bermejo1
Juan Vicente Delgado Bermejo1  Antonio González Ariza2*
Antonio González Ariza2*  José Manuel León Jurado2 Ander Arando Arbulu1
José Manuel León Jurado2 Ander Arando Arbulu1  Francisco Javier Navas González1
Francisco Javier Navas González1- 1Departamento de Genética, Facultad de Ciencias Veterinarias, Universidad de Córdoba, Córdoba, España
- 2Centro Provincial Agropecuary, Diputación Provincial de Córdoba, Córdoba, España
Introducción: Diversos parámetros climatológicos y del ciclo lunar tienen un impacto directo en la reproducción animal, y en el caso de las especies aviares, los espermatozoides son extremadamente sensibles al estrés térmico. Estos parámetros podrían influir en la congelabilidad de los espermatozoides, lo que en última instancia afectará a la calidad del semen tras la descongelación, siendo la motilidad de los espermatozoides en gallos un indicador relevante de esta calidad ya que está muy relacionada con la fertilidad. Por lo tanto, el objetivo del presente estudio es determinar cuáles son los parámetros climatológicos y del ciclo lunar que tienen un mayor efecto sobre la congelabilidad de espermatozoides en gallos.
Métodos: Se obtuvieron espermatozoides de 16 gallos de raza Utrerana y se realizaron un total de 27 repeticiones. Se realizó un pool con aquellos eyaculados que cumplían con los criterios mínimos de calidad para cada réplica, y se analizaron cuatro muestras de congelación-descongelación por réplica. Se descongelaron las pajuelas y se evaluó la motilidad de los espermatozoides, clasificando los resultados obtenidos en cuatro grupos de calidad seminal según las directrices de la Organización de las Naciones Unidas para la Alimentación y la Agricultura (Grupo 1: Bueno, Grupo 2: Satisfactorio, Grupo 3: Aceptable pero indeseable y Grupo 4: Insatisfactorio). Se registraron los siguientes rasgos para cada día de recolección de semen: temperatura máxima, temperatura mínima, presión barométrica máxima, presión barométrica mínima, ráfaga máxima, dirección del viento, velocidad media del viento, horas de sol, precipitación, fase lunar y porcentaje de superficie lunar iluminada sobre el área total.
Resultados: Se realizó un análisis canónico discriminante para determinar cuál de estos parámetros ofrecía más información a la hora de clasificar un eyaculado en cada grupo de calidad, siendo la temperatura mínima, la luna nueva como fase lunar, la presión barométrica mínima y la precipitación las variables más significativas.
Discusión: De acuerdo con los resultados obtenidos, la calidad del semen disminuye cuando la temperatura y la precipitación son más bajas, la presión es mayor y cuando hay una fase de luna nueva. Por lo tanto, estas condiciones ambientales deben evitarse para la recolección y el procesamiento de espermatozoides.
1 Introducción
Las condiciones climáticas tienen un impacto en la reproducción animal, lo que se refleja en los diferentes parámetros reproductivos. Se ha reportado que la temperatura, la humedad y la presión atmosférica son factores importantes que determinan la calidad seminal de los gallos (1, 2). El estrés térmico genera numerosas especies reactivas de oxígeno (ROS) que provocan un desequilibrio oxidativo que se acentúa en las especies aviares debido a la elevada cantidad de ácidos grasos poliinsaturados (AGPI) presentes en los espermatozoides aviares, que son más propensos a la peroxidación lipídica en presencia de ROS (3, 4). Además, una temperatura alta se asocia con una menor concentración y motilidad de los espermatozoides (5). El ciclo lunar es otro factor que también influye en la reproducción animal, provocando fluctuaciones en los niveles de corticosterona y melatonina en las aves (6).
Dado que múltiples factores pueden influir en los parámetros reproductivos, en los machos, la evaluación de la calidad seminal de los eyaculados es importante y se puede realizar por diferentes métodos. Existen herramientas para evaluar tanto parámetros macroscópicos, como el volumen y el color del eyaculado, como parámetros microscópicos como la concentración, la morfología o la motilidad de los espermatozoides (7). En el caso de las aves, la motilidad de los espermatozoides es un factor muy relevante ya que está muy relacionado con la fertilidad, ya que solo aquellos espermatozoides con un buen nivel de motilidad podrán ascender por el oviducto hasta llegar a la zona de fecundación (8).
Dentro de las técnicas de reproducción asistida, la criopreservación de espermatozoides es una herramienta muy valiosa para la conservación de los recursos genéticos en especies aviares (9). Sin embargo, la viabilidad de los espermatozoides generalmente se reduce entre un 30 y un 60% después de un ciclo de congelación y descongelación (8, 10). Un factor que puede influir en la congelabilidad de los espermatozoides es la época del año, ya que provoca cambios en la composición bioquímica del eyaculado, lo que se traduce en una motilidad más favorable en el semen descongelado cuando la recogida seminal se realiza en primavera, tal y como se ha descrito en razas avícolas locales españolas mantenidas en condiciones ambientales naturales (11).
Deben mejorarse las técnicas de criopreservación seminal en las especies avícolas. Los bancos de germoplasma aviar son muy incompletos, en comparación con los de otras especies ganaderas (12), ya que el semen de las aves de corral es más vulnerable debido a las características específicas de los espermatozoides aviares (13). Esto se debe principalmente a las peculiaridades fisiológicas y anatómicas de las aves, como la colocación intraabdominal de los testículos y la ausencia de glándulas sexuales accesorias (13). Por lo tanto, se debe realizar un esfuerzo técnico y de investigación por parte de las instituciones públicas para el desarrollo de técnicas y herramientas que mejoren la conservación in vitro de los recursos genéticos locales.
Un banco de un genotipo local debe considerarse completo cuando el material genético almacenado permite la reconstrucción de la raza, si es necesario, así como el aumento del tamaño efectivo de una población mediante la reducción de la deriva genética, entre otros (14). Un ejemplo de raza autóctona en peligro de extinción es la raza aviar utrerana, cuyo censo a 31/12/2022 no superaba los 2000 individuos (12) Esta raza pertenece al tronco mediterráneo y presenta una alta rusticidad, lo que permite su adaptabilidad a sistemas de producción extensivos más sostenibles y con un mínimo impacto en el medio ambiente. Así, la conservación de este tipo de razas tiene un impacto positivo indirecto en la salud humana y sus sistemas de cría también son más respetuosos con el bienestar animal (15, 16). La gallina Utrerana se orientó inicialmente hacia la producción de huevos, sin embargo, la introducción de genotipos híbridos comerciales bastante productivos en Europa produjo el desplazamiento de esta raza a una posición en peligro de extinción (17). Dada su complicada situación actual en cuanto al censo existente de la raza, el desarrollo de un banco de germoplasma para la raza debe ser una herramienta importante para la implementación exitosa de su programa de conservación.
En cualquier caso, el control de los parámetros climatológicos podría aportar beneficios al proceso de criopreservación, por lo que el objetivo principal de la presente investigación es determinar las relaciones existentes entre los diferentes parámetros climáticos y el ciclo lunar sobre la calidad seminal post-descongelación de los eyaculados de los gallos en relación con la motilidad de los espermatozoides.
2 Materiales y métodos
2.1 Aprobación ética
El presente estudio queda excluido del ámbito de evaluación del Comité de Ética de la Universidad de Córdoba ya que los datos obtenidos forman parte de las actividades rutinarias que se realizan en el Centro Provincial Agrícola de la Diputación de Córdoba (Andalucía, España) como centro de reproducción y conservación aviar de razas autóctonas y por tanto no se consideran animales utilizados con fines científicos. Sin embargo, todos los animales utilizados en el estudio fueron tratados siguiendo la legislación europea (Directiva 2010/63/UE «relativa a la protección de los animales utilizados para fines científicos»), que ha sido transpuesta al ordenamiento jurídico español a través del RD 53/2013.
2.2 Muestra animal
Para este estudio, 16 gallos utreranas de raza entre 1 y 3 años de edad fueron alojados en jaulas individuales (95 × 95 × 95 cm) en el Centro Provincial Agropecuary de la Diputación de Córdoba (37°54′50.9″N-4°42′40.4″W, Andalucía, sur de España) bajo un fotoperiodo natural. Todos los animales fueron alimentados con una dieta comercial (15,20% de proteína bruta, 4,60% de grasa y aceites crudos, 3,20% de fibra bruta, 14,00% de ceniza bruta, 4,10% de calcio, 0,66% de fósforo, 0,19% de sodio, 0,31% de metionina, 0,72% de lisina) y se les suministró agua ad libitum.
2.3 Recogida y procesamiento de esperma
La recolección de semen se realizó mediante la técnica de masaje dorsal-abdominal descrita por Burrows y Quinn (18). La recolección de semen se realizó entre septiembre de 2021 y mayo de 2022. Para cada jornada de trabajo se realizó un pool con aquellas muestras que cumplían con los criterios mínimos de calidad previamente establecidos: volumen (>0,2 mL), concentración (>3×109 spz/mL), motilidad (≥80%) y morfología (≤10 a 15%). Se realizaron un total de 27 réplicas, analizando cuatro muestras por réplica. La Tabla 1 muestra el número de muestras utilizadas para cada estación y fase del ciclo lunar.
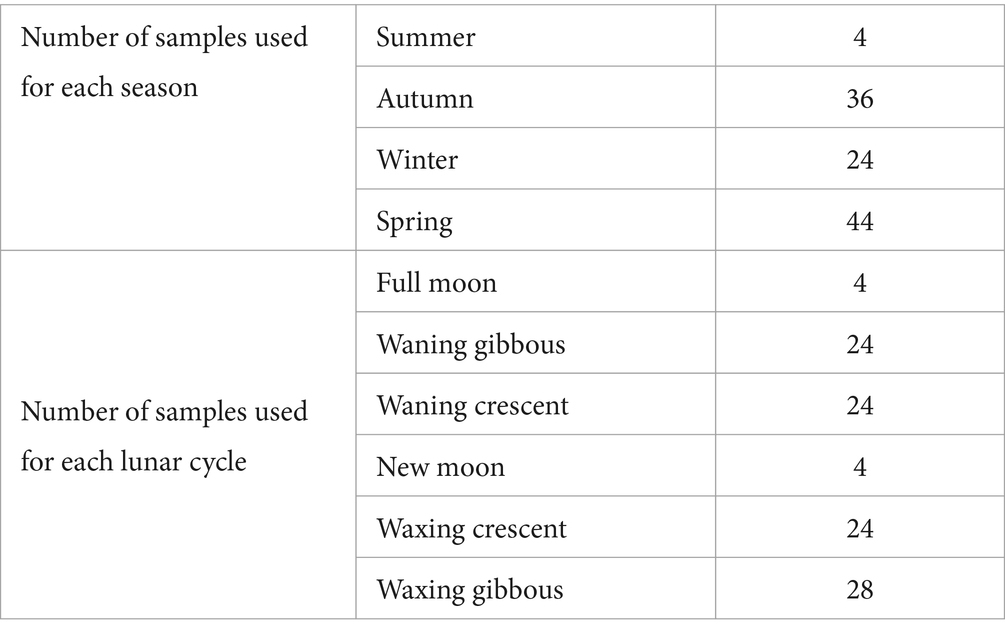 Tabla 1. Número de muestras utilizadas para cada estación y fase del ciclo lunar.
Tabla 1. Número de muestras utilizadas para cada estación y fase del ciclo lunar.
Cada muestra de semen se refrigeró en un refrigerador programable (incubadora celular SH-020S, Welson, Corea) durante 1 h hasta que alcanzó los 5 °C con una tasa de disminución de temperatura de 0,3 °C/min. Después de esa hora, el semen se diluyó con un diluyente compuesto por 0,2 g de D (+) de glucosa, 3,8 g de D (+) de trehalosa dihidratada, 1,2 g de sal monosódica de ácido L-glutámico, 0,3 g de acetato de potasio, 0. 08 g de acetato de magnesio tetrahidratado, 0,05 g de citrato de sodio tribásico dihidratado, 0,4 g de BES, 0,4 g de Bis-Tris y 0,001 g de sulfato de gentamicina (pH = 6,8, osmolaridad = 360 mOsm) (19). Después de 30 min, se realizó una segunda dilución con el diluyente descrito anteriormente, a la que se le añadió N-metilacetamida (NMA) como crioprotector a una concentración del 18% (concentración final del 9%). Después de la segunda dilución, los espermatozoides se envasaron en pajuelas de 0,25 a una concentración final de 250 × 106 spz/paja y 30 min posteriormente se colocan en vapores de nitrógeno a una altura de 4 cm durante 30 min y finalmente se sumergen en nitrógeno líquido (−196°C).
2.4 Evaluación de la motilidad de los espermatozoides
Para la evaluación de la motilidad de los espermatozoides, se descongelaron cuatro pajuelas por réplica (un total de 108 observaciones) sumergiendo cada pajuela en un baño de agua a 5 °C durante 1 min 40 s (19). La motilidad total (TM, %) se analizó utilizando un analizador de espermatozoides asistido por ordenador IVOS 12.3 (Hamilton-Thorne Bioscience, MA, Estados Unidos). Para ello, se depositaron 5 μL de la muestra en una cámara Life Optic. Esta muestra había sido previamente diluida a 50 × 106 spz/mL con el diluyente descrito anteriormente, considerando como espermatozoides aquellas células con un área entre 2 y 60 μm2.
Para clasificar las muestras analizadas, se establecieron cuatro grupos en función de la calidad de los espermatozoides post-descongelados en relación con la MT en base a los criterios publicados por la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) (20), que son los siguientes: Grupo 1: Bueno (>50%), Grupo 2: Satisfactorio (40-50%), Grupo 3: Aceptable pero indeseable (30-40%) y Grupo 4: Insatisfactorio (<30%).
2.5 Recogida de datos: parámetros climáticos y ciclo lunar
Para cada día de extracción seminal, se analizaron las condiciones ambientales en el Centro Provincial Agropecuary de la Diputación de Córdoba para diferentes parámetros climatológicos como temperatura máxima, temperatura mínima, presión barométrica máxima, presión barométrica mínima, ráfaga máxima, dirección del viento, velocidad media del viento, horas de sol y precipitaciones, y el ciclo lunar como fase lunar y porcentaje de superficie lunar iluminada sobre el área total. Los valores mínimo y máximo de cada parámetro se presentan en la Tabla 2. Los datos de los parámetros climatológicos se obtuvieron de la Agencia Estatal de Meteorología1 y los del ciclo lunar de la página web del Departamento de Aplicaciones Astronómicas del Observatorio Naval de los Estados Unidos.2
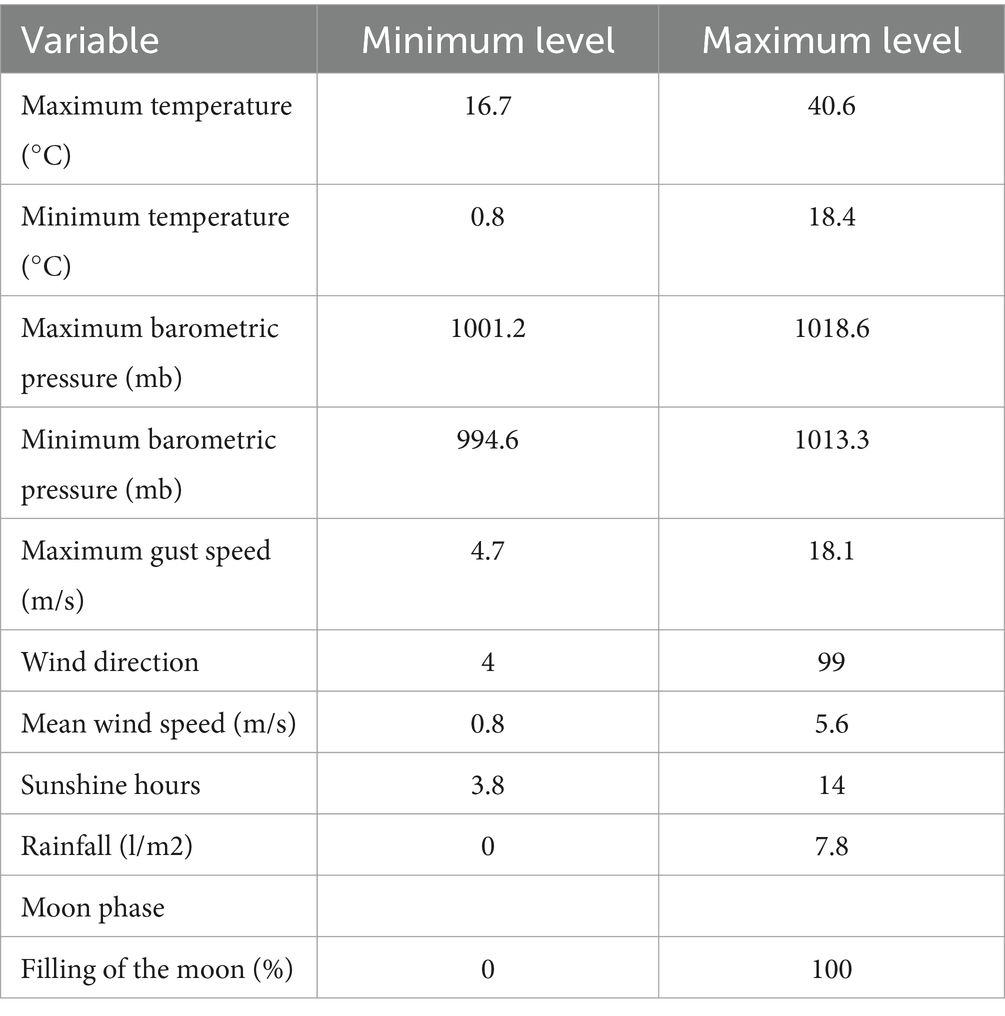 Tabla 2. Valores mínimos y máximos para los parámetros evaluados.
Tabla 2. Valores mínimos y máximos para los parámetros evaluados.
2.6 Estadística descriptiva general
Se estableció la media de cada grupo de calidad seminal para los rasgos meteorológicos y del ciclo lunar registrados: temperatura máxima, temperatura mínima, presión barométrica máxima, presión barométrica mínima, velocidad máxima de la ráfaga, dirección del viento, velocidad media del viento, horas de sol, precipitación, fase lunar y porcentaje de llenado de la luna. Para realizar este análisis, se utilizó la rutina de estadística descriptiva del paquete de descripción de datos del software XLSTAT (Addinsoft Pearson Edition 2014, Addinsoft, París, Francia).
2.7 Análisis estadístico: análisis canónico discriminante
Para el DCA se incluyeron las 11 variables explicativas sobre los rasgos meteorológicos y del ciclo lunar mencionados anteriormente. Los criterios de clasificación establecidos corresponden a los cuatro grupos de calidad seminal mencionados anteriormente (1: buena, 2: satisfactoria, 3: aceptable pero indeseable y 4: insatisfactoria) y se midieron las diferencias entre los grupos.
Según varios autores, se requiere un tamaño mínimo de muestra de al menos 20 observaciones por cada uno de los 4 o 5 predictores, y el número máximo de variables independientes para mitigar los posibles efectos de distorsión debe ser n-2, donde n es el tamaño de la muestra. Teniendo esto en cuenta, el tamaño muestral utilizado en este estudio sería correcto (21, 22).
El análisis de multicolinealidad asegura la existencia de relaciones fuertes e independientes entre los predictores. Entre los métodos de selección hacia adelante, se llevó a cabo el método hacia adelante porque requiere menos tiempo de ejecución (17). Para realizar los análisis de multicolinealidad y DCA, se utilizó la rutina de Análisis Discriminante del paquete de Análisis de Datos del software XLSTAT (Addinsoft Pearson Edition 2014, Addinsoft, París, Francia).
2.7.1 Ensayos preliminares de multicolinealidad
Antes de realizar un DCA, se comprobó el supuesto de multicolinealidad para asegurar que la varianza del potencial explicativo no está sobreinflada por problemas de redundancia entre variables. El factor de inflación de la varianza (VIF) es el indicador más utilizado para detectar la multicolinealidad, y autores previos han recomendado valores inferiores a 5 para descartar los problemas de redundancia (23). El VIF se calculó utilizando la siguiente fórmula como subrutina de la rutina de Análisis Discriminante del paquete de Análisis de Datos del software XLSTAT:
donde R2 fue el coeficiente de determinación de la ecuación de regresión.
2.7.2 Fiabilidad del modelo DCA
Cuando se utiliza DCA y hay tamaños de muestra desiguales, la única prueba aceptable que se puede aplicar es el criterio de traza de Pillai. La asunción de matrices de covarianza iguales se probó a través de este método en el análisis de funciones discriminantes (24). Para el cálculo de este parámetro se utilizó una subrutina de la rutina de Análisis Discriminante del paquete de Análisis de Datos del software XLSTAT, considerando que el conjunto de predictores del DCA fue estadísticamente significativo cuando p ≤ 0,05. Se ha argumentado que el criterio de traza de Pillai es el estadístico más robusto para la protección general contra las desviaciones de la normalidad y la homogeneidad de la varianza de los residuos multivariados. La evidencia de que el conjunto de predictores tiene un efecto estadísticamente significativo sobre los valores de la variable de respuesta aumenta a medida que el valor de la traza de Pillai es mayor.
2.7.3 Eficiencia del DCA
Las contribuciones de las variables a la función discriminante se analizaron mediante el uso de la prueba Lambda de Wilks. La contribución es mayor a medida que el valor lambda de Wilks se aproxima a 0. Las funciones se pueden utilizar para explicar la adscripción de grupo si p ≤ 0,05 (25).
2.7.4 Evaluación del potencial discriminante factorial independiente, coeficientes canónicos e interpretación de la carga
Tras analizar las variables cuyo potencial discriminante se basaba en las diferencias de medias entre los distintos tratamientos, se realizó un análisis de la función discriminante para identificar aquellas cuyo potencial discriminante podía basarse en su capacidad para determinar mayores porcentajes de asignación de observaciones dentro de su grupo. Valores discriminantes de ≥ |0,40| indican que la carga discriminante de las diferentes variables es significativamente discriminante. Para evitar la inclusión de variables redundantes en la función, se utiliza una técnica de procedimiento por pasos. Se obtuvo una mayor capacidad de discriminación y un mayor porcentaje de clasificación correcta cuando los valores absolutos de las cargas de los coeficientes estandarizados de cada variable fueron altos.
2.7.5 Representación espacial
Se calcularon las distancias de Mahalanobis al cuadrado y el análisis de componentes principales, utilizando la siguiente fórmula:
Dónde D2Ij𝐷Yo𝑗2 : distancia entre la población i y j; COV−1: inversa de la matriz de covarianza de la variable medida x; Υ¯¯¯Yoϒ¯Yo y Υ¯¯¯jϒ¯𝑗 : medias de la variable x en las poblaciones i-ésima y j-ésima, respectivamente (25).
Se construyó un dendrograma convirtiendo la matriz de distancias de Mahalanobis al cuadrado en una matriz de distancias euclidianas. Para ello, se utilizaron promedios aritméticos del método pare-grupo de infraponderación (UPGMA; Universidad Rovira i Virgili, Tarragona, España) y el procedimiento de Filogenia de MEGA X 10.0.5 (Institute of Molecular Evolutionary Genetics, The Pennsylvania State University, State College, PA, Estados Unidos).
2.7.6 Validación cruzada de funciones discriminantes
La probabilidad de que una observación de un fondo desconocido se clasifique correctamente en un grupo determinado puede determinarse calculando la proporción de aciertos (26). Para determinar si las diferentes funciones discriminantes pueden validarse, se utilizó la opción de validación cruzada de dejar una salida (27). Cuando la tasa de clasificación es al menos un 25% más alta que la obtenida por azar, se puede considerar que el DCA logra la precisión de la clasificación.
Estos resultados obtenidos deben estar respaldados por el estadístico Q de Press, que es un parámetro que puede comparar el poder discriminativo de la función validada de forma cruzada mediante el uso de la fórmula:
donde n es el número de observaciones de la muestra; n’ es el número de observaciones correctamente clasificadas, y K es el número de grupos.
El valor del estadístico Q de Press debe compararse con el valor crítico de 6,63 para χ2 con un grado de libertad en un nivel de significación de 0,01. Para considerar que la clasificación validada cruzada es significativamente mejor que el azar, el valor de Q de Press debe exceder el valor crítico de χ2 = 6,63.
3 Resultados
3.1 Estadística descriptiva general
La media para cada rasgo relacionado con la meteorología o el ciclo lunar en cada grupo seminal relacionado con la calidad se muestra en la Tabla 3. Los valores más altos de temperatura mínima (13.205) se reportaron para el grupo 1. Por otro lado, los valores más bajos para este rasgo se reportaron en el grupo 4 (6.400). Todas las muestras tomadas en el ciclo lunar de la luna nueva pertenecían al grupo 4. Por último, las muestras obtenidas en el grupo 4 obtuvieron los valores más altos para la presión barométrica mínima (1007.433).
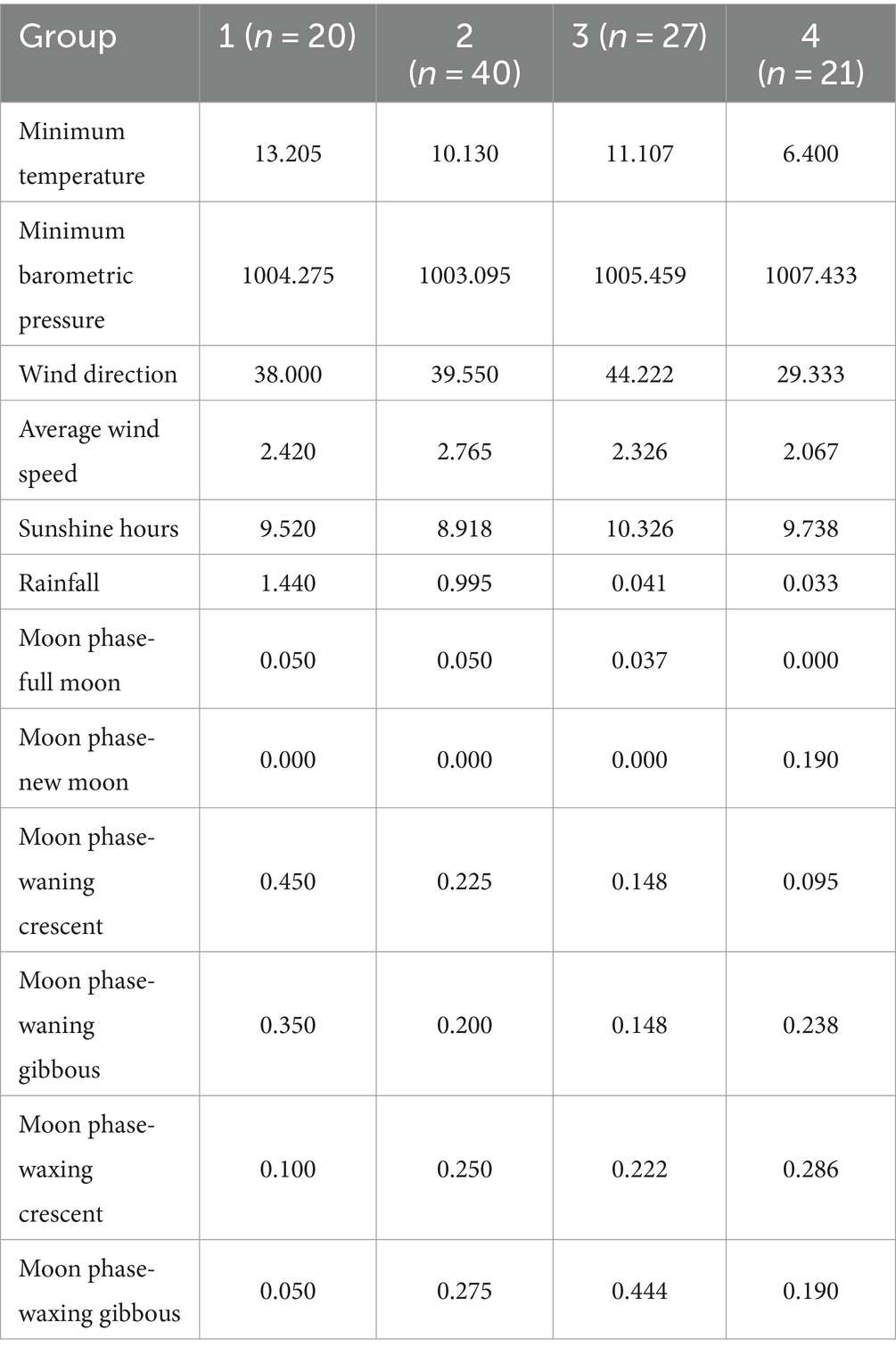 Tabla 3. Medios por clase; n: tamaño de la muestra para cada grupo; Los valores de MT [expresados como media (DE)] para cada grupo fueron los siguientes: 1 = 57,75 (7,38), 2 = 44,48 (3,21), 3 = 34,59 (3,00) y 4 = 20,62 (6,14).
Tabla 3. Medios por clase; n: tamaño de la muestra para cada grupo; Los valores de MT [expresados como media (DE)] para cada grupo fueron los siguientes: 1 = 57,75 (7,38), 2 = 44,48 (3,21), 3 = 34,59 (3,00) y 4 = 20,62 (6,14).
3.2 Fiabilidad y eficiencia del modelo DCA
La presión barométrica máxima, la temperatura máxima, el porcentaje de llenado de la luna y la velocidad máxima de la ráfaga se descartaron para los siguientes análisis, ya que estas variables mostraron un valor de VIF mayor a 5. Las variables que quedaron después del análisis preliminar de multicolinealidad (VIF < 5) se muestran en la Tabla 4.
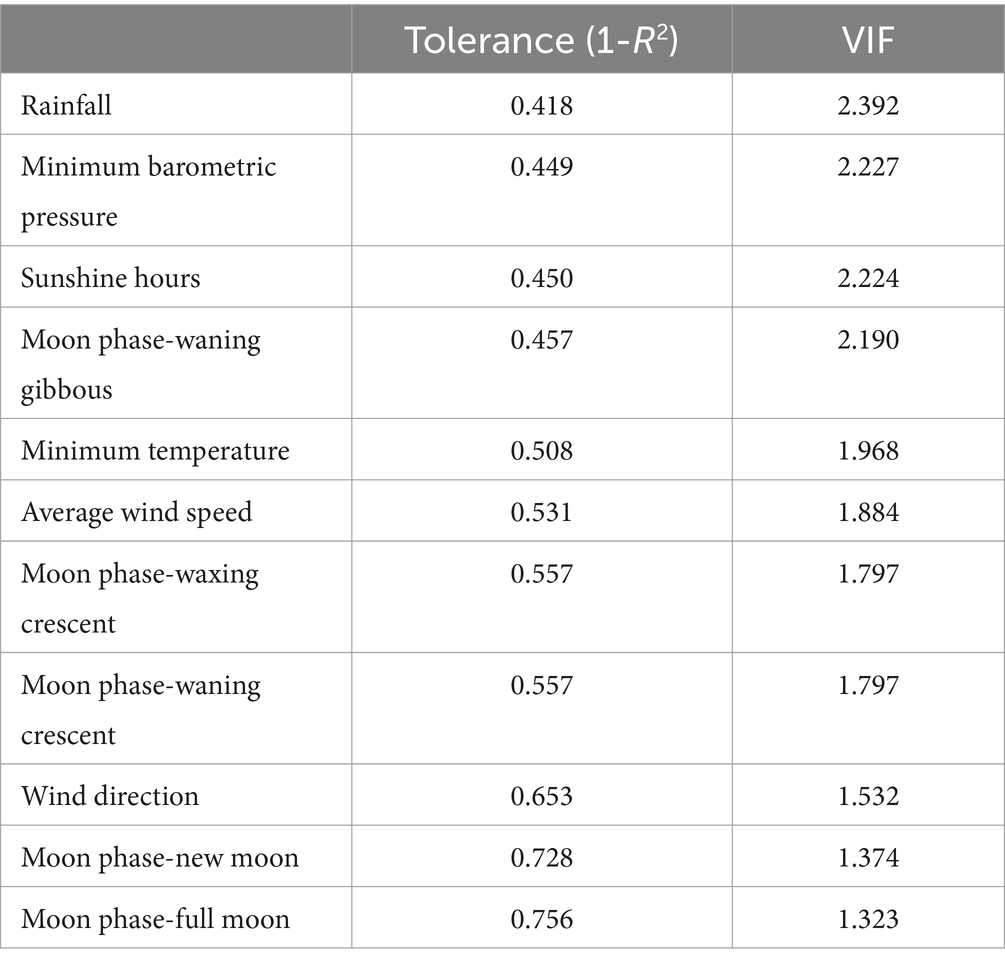 Tabla 4. Variables que se mantuvieron en el análisis preliminar de multicolinealidad utilizando el factor de inflación de la varianza (VIF) de las variables explicativas.
Tabla 4. Variables que se mantuvieron en el análisis preliminar de multicolinealidad utilizando el factor de inflación de la varianza (VIF) de las variables explicativas.
El criterio de traza significativa de Pillai determinó la validez del DCA (p < 0,0001; Tabla 5). De las tres funciones discriminantes reveladas después de los análisis discriminantes, dos mostraron una capacidad discriminante significativa (Significación de 0,010 y 0,041 para F1 y F2, respectivamente; Tabla 6). El poder discriminatorio de la función F1 fue alto (valor propio de 0,457) con un 85,28% de la varianza explicada por F1 y F2 (Figura 1).
 Tabla 5. Resumen de los resultados de la traza de pillai de matrices de igualdad de covarianza de funciones discriminantes canónicas.
Tabla 5. Resumen de los resultados de la traza de pillai de matrices de igualdad de covarianza de funciones discriminantes canónicas.
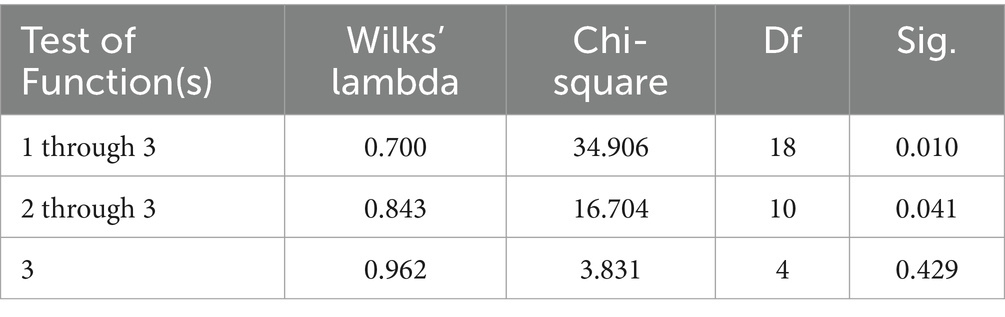 Tabla 6. Parámetros de eficiencia del análisis canónico discriminante para determinar la importancia de cada función discriminante canónica.
Tabla 6. Parámetros de eficiencia del análisis canónico discriminante para determinar la importancia de cada función discriminante canónica.
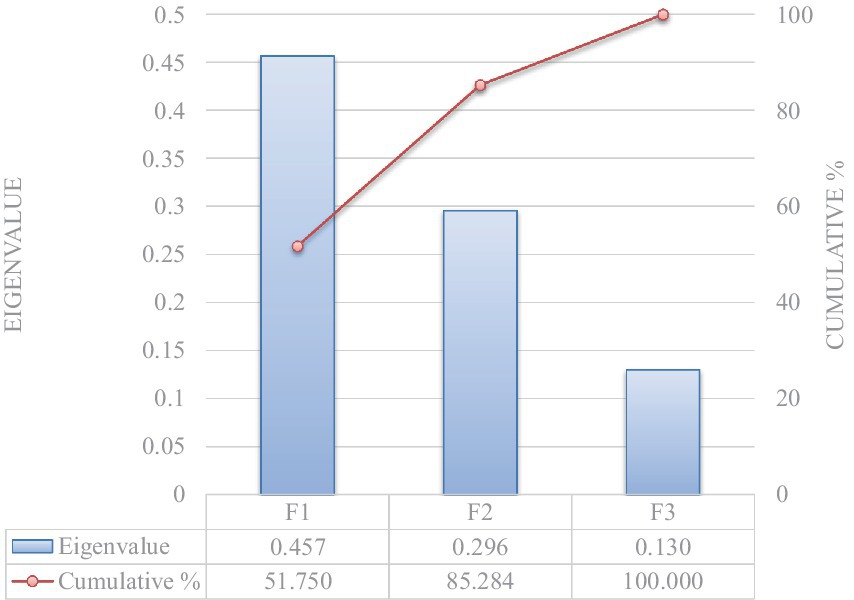 Figura 1. Valor propio y potencial explicativo de variabilidad acumulada de variables explicativas independientes.
Figura 1. Valor propio y potencial explicativo de variabilidad acumulada de variables explicativas independientes.
3.3 Evaluación del potencial discriminante factorial independiente, coeficientes canónicos e interpretación de la carga
La capacidad discriminatoria de las diferentes variables estudiadas se muestra en la Tabla 7. El mayor poder discriminatorio de una variable en cuestión se relaciona con un valor alto de F y, en consecuencia, valores más bajos de Lambda de Wilks, lo que se traduce en una mejor posición en el rango. El presente análisis reveló que la temperatura mínima (Lambda de Wilks = 0,803; F = 8.504), luna nueva como fase lunar (Lambda de Wilks = 0.841; F = 6,571), presión barométrica mínima (Lambda de Wilks = 0,886; F = 4,476), precipitaciones (Lambda de Wilks = 0,897; F = 3,988), gibosas crecientes (Lambda de Wilks = 0,908; F = 3,518) y media luna menguante (Lambda de Wilks = 0,918; F = 3.084) ya que las fases lunares contribuyeron significativamente (p < 0.05) a la capacidad discriminante de las funciones discriminantes significativas.
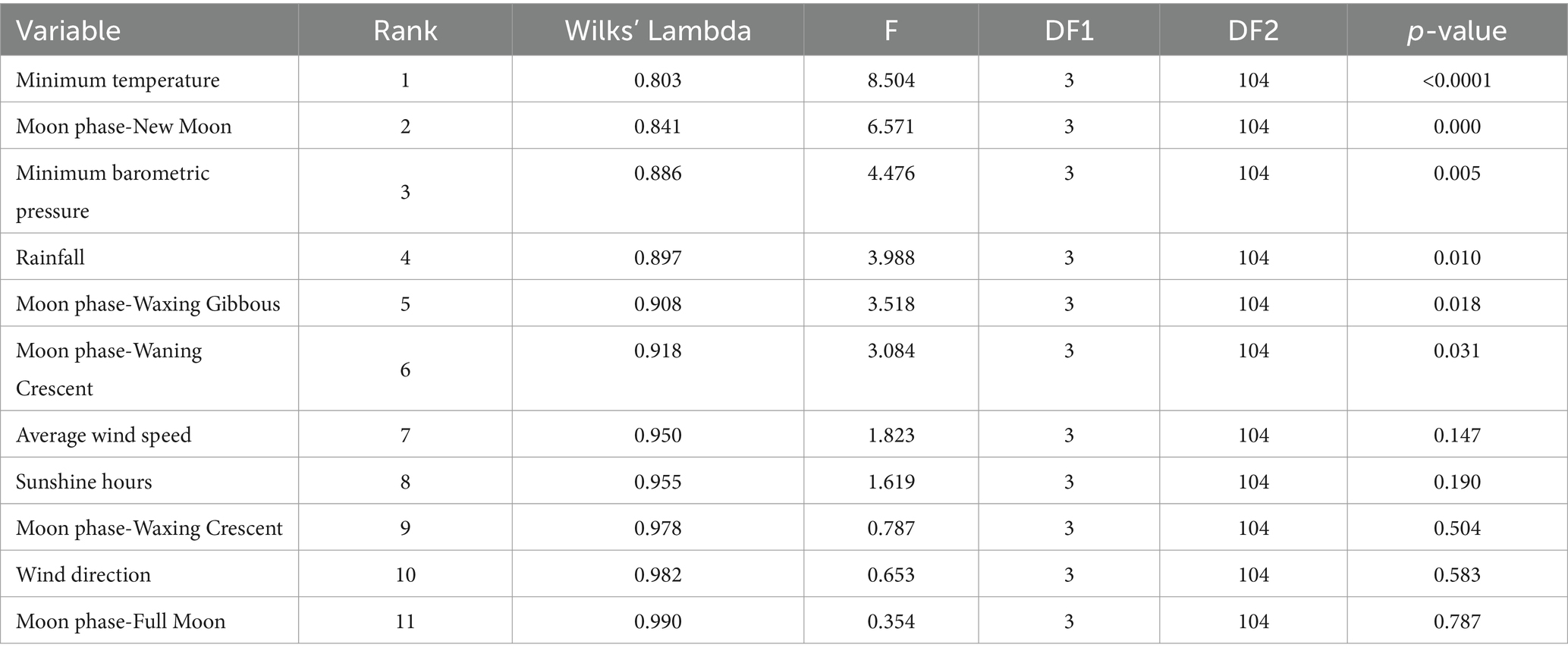 Tabla 7. Resultados de las pruebas de igualdad de medias grupales para probar la diferencia en las medias entre los grupos de muestra una vez que se han eliminado las variables redundantes.
Tabla 7. Resultados de las pruebas de igualdad de medias grupales para probar la diferencia en las medias entre los grupos de muestra una vez que se han eliminado las variables redundantes.
En la Figura 2 se muestran las cargas de coeficientes canónicos discriminantes para variables representativas en funciones discriminantes. La temperatura mínima (valor del coeficiente = │0,741│), la luna nueva (valor del coeficiente = │0,693│) y la presión mínima (valor del coeficiente = │0,499│) fueron los rasgos con los coeficientes de función discriminante canónicos estandarizados más altos para F1. El giboso creciente (valor del coeficiente = │0,496│), la media luna menguante (valor del coeficiente = │0,379│) y el giboso menguante (valor del coeficiente = │0,306│) fueron los rasgos que mostraron los valores de coeficiente más altos para F2.
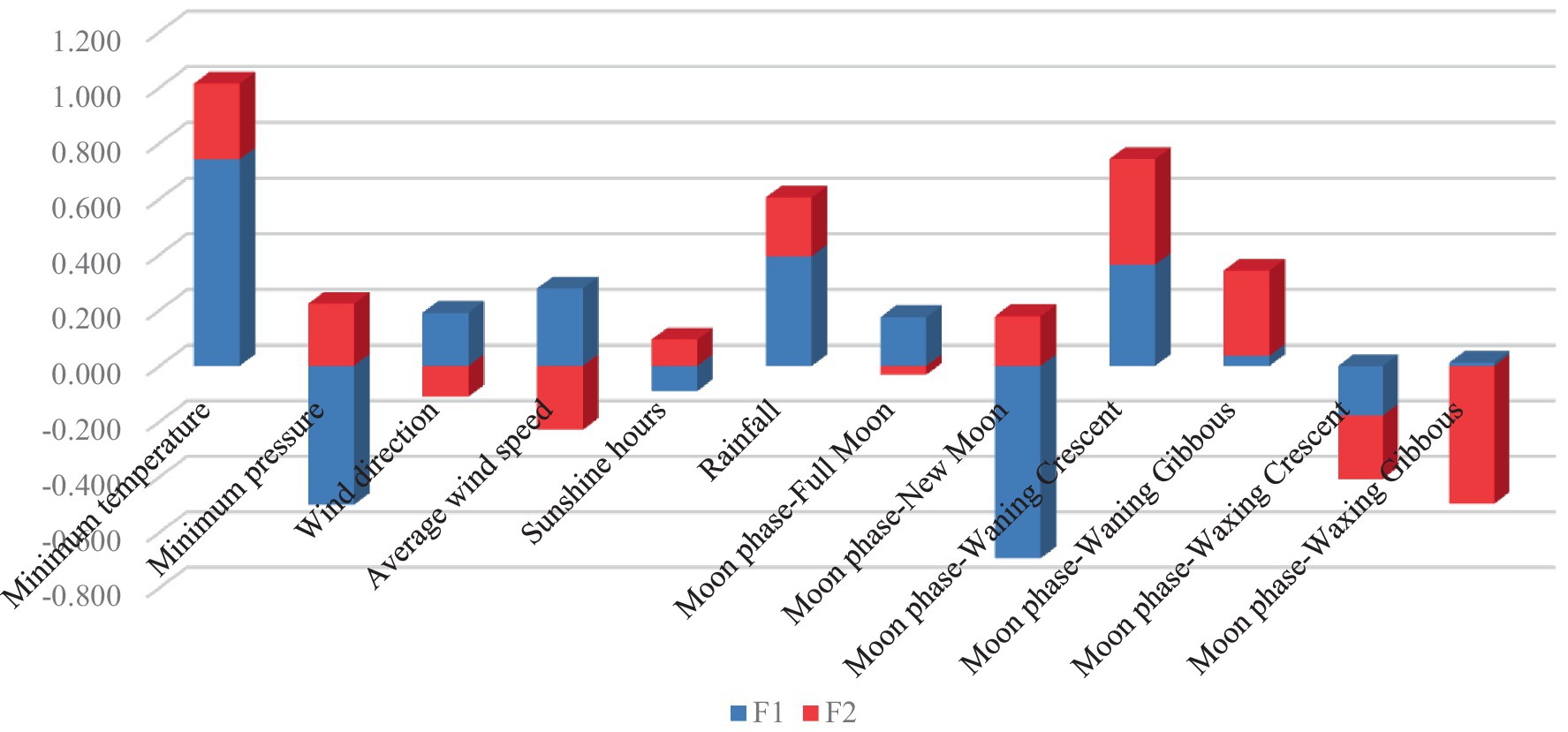 Figura 2. Coeficientes de función discriminante canónicos estandarizados.
Figura 2. Coeficientes de función discriminante canónicos estandarizados.
3.4 Representación espacial
Se observa una clara diferenciación entre tratamientos (Figura 3). Para obtener las coordenadas de los ejes x e y, se sustituye el valor medio de las observaciones en cada término de las dos primeras funciones discriminantes (F1 y F2) para obtener la posición relativa de los centroides. El poder predictivo de la función discriminante canónica para clasificar las observaciones será mayor a medida que aumente la distancia entre los centroides.
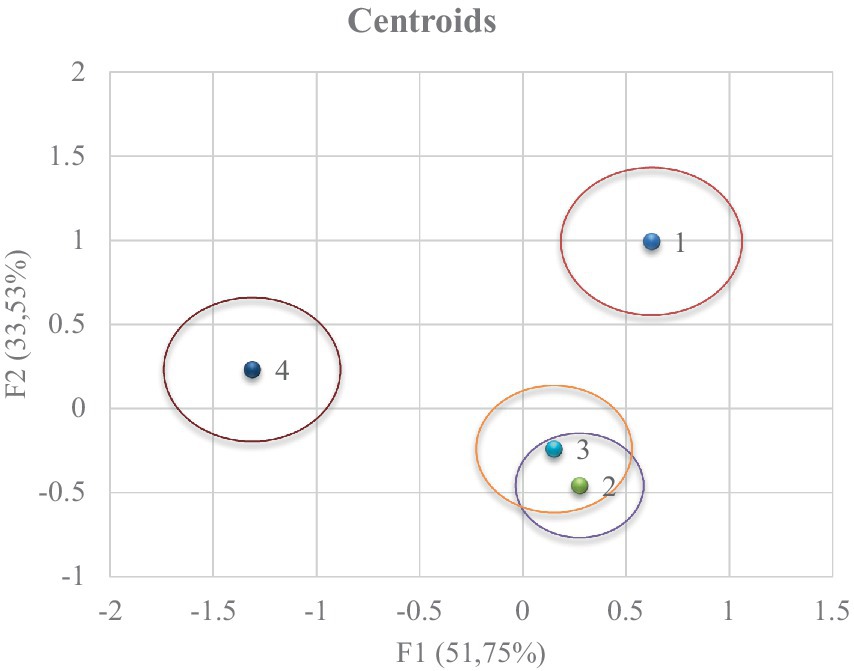 Figura 3. Mapa territorial que muestra los resultados del análisis discriminante canónico sobre los diferentes grupos.
Figura 3. Mapa territorial que muestra los resultados del análisis discriminante canónico sobre los diferentes grupos.
La distancia de Mahalanobis se calcula por la distancia relativa de la observación del problema al centroide de su grupo más cercano, lo que representa la probabilidad de que una observación que muestra un fondo desconocido pertenezca a un grupo dado, y es necesario calcular la proporción de aciertos que se muestra en la Figura 4. En ambas figuras (Figuras 3, 4), se reportó que el grupo 4 se obtuvo en condiciones meteorológicas y de fase lunar que diferían en gran medida del resto de los grupos.
 Figura 4. Cladograma construido a partir de las distancias de Mahalanobis a través de diferentes grupos.
Figura 4. Cladograma construido a partir de las distancias de Mahalanobis a través de diferentes grupos.
3.5 Validación cruzada de funciones discriminantes
Un valor Q de Press de 215,11 (n = 108; n‘ = 93; K = 4). Por lo tanto, se puede considerar que las predicciones son mejores que el azar en un 95%. En la Tabla 8 se muestra la validación cruzada de los resultados de la clasificación discriminante. La herramienta discriminante desarrollada en el presente estudio permitió clasificar correctamente el 86,11% de las muestras estudiadas.
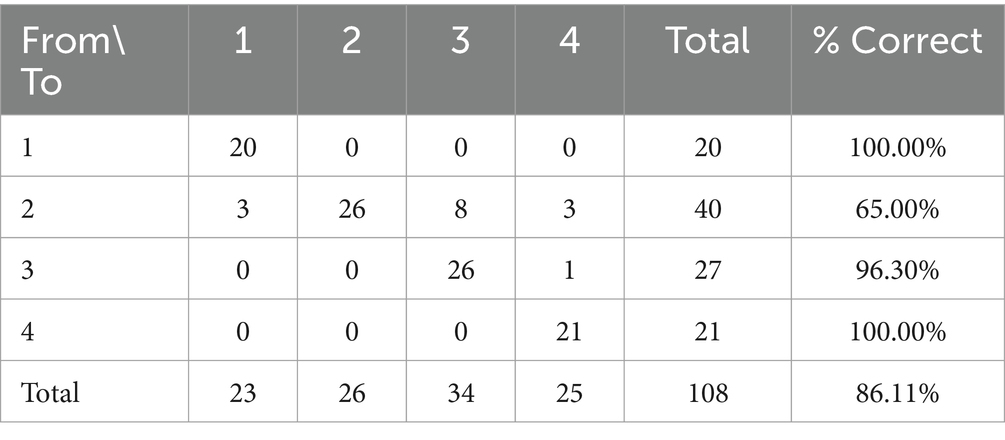 Tabla 8. Validación cruzada de los resultados de la clasificación.
Tabla 8. Validación cruzada de los resultados de la clasificación.
4 Discusión
Esta investigación analiza una serie de características climáticas y del ciclo lunar. La verificación de las relaciones entre las variables explicativas es útil para decidir qué combinación de ellas conforma un modelo predictivo eficaz. Para ello, fue necesaria la selección de aquellas variables independientes que no se superponen en cuanto a su capacidad para explicar la variabilidad de los datos. Por lo tanto, las variables con problemas de multicolinealidad (VIF > 5) no se tuvieron en cuenta en los siguientes análisis.
La presión barométrica máxima y la temperatura máxima se descartaron debido a problemas de multicolinealidad con presión barométrica mínima y temperatura mínima, respectivamente. En la atmósfera, elementos como la temperatura y la presión barométrica muestran variaciones diurnas, que pueden explicarse por la combinación de múltiples factores, siendo el componente solar el factor determinante, y las temperaturas máximas y mínimas que ocurren aproximadamente a la misma hora local cada día (28, 29). La curva de temperatura diaria puede ser aproximada por una función sinusoidal, pero puede estar influenciada por la latitud, la época del año y otros factores climáticos. Sin embargo, factores como las nubes o el viento pueden modificar la curva temporalmente (30).
Por otro lado, los problemas de multicolinealidad con el porcentaje de llenado lunar podrían estar relacionados con la redundancia con la variable fase lunar ya que ambos parámetros están directamente relacionados. Las fases de la luna se determinan en función de la porción iluminada que se puede observar desde la Tierra a medida que la luna se mueve a su alrededor (31). La fase lunar podría tener un impacto en la fisiología animal, ya que influye en los cambios cíclicos en el agua, que constituye el 65-70% de las aves (32, 33). Además, autores anteriores han planteado la hipótesis de que los animales pueden ser influenciados por la luna debido a los cambios en los campos geomagnéticos al aumentar la sensibilidad a la magnetorrecepción con una luna llena (34). La influencia del ciclo lunar en la reproducción animal ha sido estudiada en varias especies como el ganado vacuno (35), porcino (36) o equino (37), sin embargo, es evidente el desconocimiento de las especies aviares en este campo.
Finalmente, se descartó la variable velocidad máxima de ráfaga, que podría solaparse con la velocidad media del viento. El viento es un parámetro importante ya que influye en la fisiología animal debido a sus efectos sobre el factor de enfriamiento, lo que repercute en la homeotermia, la locomoción y el forrajeo (38). En el caso de las aves marinas, se ha observado que la velocidad del viento tiene un impacto en el coste energético derivado del vuelo (39) o en las estrategias utilizadas para el forrajeo (40). Teniendo todo esto en cuenta, el hecho de que la variable velocidad máxima de la ráfaga no contribuyera significativamente al análisis podría explicarse por el hecho de que los animales utilizados en este estudio se encontraban en condiciones controladas en las que se suministraba alimento ad libitum. En otras especies animales como el bovino, la velocidad del viento es un factor clave en el proceso de termorregulación bovina, con un marcado efecto sobre el enfriamiento testicular (41).
Después de realizar el análisis de multicolinealidad, entre las variables que quedan, la temperatura mínima, la luna nueva como fase lunar, la presión barométrica mínima, la precipitación, la gibosa creciente y la media luna menguante como fases lunares contribuyen significativamente a determinar si un eyaculado de congelación-descongelación pertenece a un grupo de calidad u otro.
La temperatura mínima mostró el mayor poder discriminante. Para el correcto desarrollo de la capacidad reproductiva, los gallos deben alojarse a una temperatura ambiente adecuada. Las desviaciones de 5 °C respecto a su óptimo térmico dan lugar a grandes reducciones en el éxito reproductivo (42). Por lo tanto, la espermatogénesis puede verse alterada por los cambios fisiológicos resultantes del estrés térmico. Por ejemplo, en los mamíferos, se ha demostrado una reducción en la producción de espermatozoides porque el nivel de testosterona disminuye bajo estrés térmico (43). Del mismo modo, las temperaturas demasiado bajas también afectan a la calidad del semen. En la presente investigación, dentro de las temperaturas mínimas, la de mayor valor es la que da lugar a la mayor motilidad espermática. Esto podría deberse a que las bajas temperaturas suprimen el crecimiento testicular e influyen negativamente en la fertilidad de las aves, lo que en última instancia está directamente relacionado con la calidad del semen (44). Además, varios estudios están demostrando cómo el frío disminuye el desarrollo testicular en varias especies de aves (45, 46).
La fase lunar, especialmente la luna nueva, también muestra un alto poder discriminatorio. Las aves, durante las fases en las que la luminosidad es mayor, muestran un comportamiento más activo reflejado en un canto más potente (47, 48). En las aves reproductoras, las concentraciones de melatonina fluctúan según el ritmo lunar, lo que se debe a la intensidad de la luz asociada a las diferentes fases lunares (49). La melatonina juega un papel protector en el desarrollo testicular al ser un potente antioxidante que elimina los radicales libres de una manera muy efectiva. Por el contrario, en los hombres humanos, los niveles bajos de melatonina dan como resultado una menor calidad del esperma debido a la reducción de la motilidad de los espermatozoides (50). En carneros y cabras, el tratamiento con melatonina mejora el desarrollo testicular y la calidad seminal (51, 52). En cualquier caso, de acuerdo con los resultados obtenidos en la presente investigación, cuando hay una fase de luna nueva, la calidad seminal disminuye. Resultados similares se obtuvieron en un estudio realizado en toros, en el que la actividad de los espermatozoides era mayor cuando había una fase de luna llena en comparación con los datos obtenidos durante una fase de luna nueva (53). Por el contrario, en humanos no se observaron diferencias significativas para el parámetro de motilidad espermática durante las fases del ciclo lunar evaluadas (54).
Otra variable con alto poder discriminante es la presión barométrica mínima. En el presente estudio, cuando la presión barométrica mínima es mayor, el eyaculado tiene peor motilidad post-descongelación. Esto coincide con lo reportado por Díaz-Usi, Venturina (55) en búfalos de agua, en los que un aumento de la presión barométrica tiene efectos negativos sobre las características de motilidad de los espermatozoides, tanto en semen fresco como después de la criopreservación, especialmente en lo que respecta a los parámetros cinemáticos. Este mismo efecto se ha observado en toros (56). Por el contrario, en las ovejas se observa una mejor calidad seminal cuando la presión es alta, ya que las ovejas inseminadas con espermatozoides obtenidos en períodos en los que la presión era baja dieron a luz corderos con bajo peso al nacer y al destete (57). Sin embargo, el hecho de que las ovejas sean una especie poliestral estacional de día corto puede explicar este hecho.
Finalmente, la variable precipitación también fue significativa para determinar el grupo de calidad seminal al que pertenece un eyaculado post-descongelación. La precipitación, entre otros parámetros climatológicos, es un factor de estrés ambiental que podría tener un impacto en los diferentes procesos fisiológicos de los animales (58). Sin embargo, en cabras, durante un período seco, se detectó un menor porcentaje de motilidad y un mayor número de espermatozoides anormales por Van Tilburg, Salles (59). Los períodos secos corresponden a temperaturas más altas, lo que podría explicar la peor calidad del semen, ya que la temperatura elevada es una fuente importante de estrés para las aves, que responden modificando las funciones de los órganos y los niveles circulantes de hormonas, glucosa, leucocitos y electrolitos (60, 61). Además, la temperatura alta se asocia con una menor concentración y motilidad de los espermatozoides (5). Además, el estrés térmico provoca infertilidad aviar, que se debe a que las altas temperaturas afectan negativamente a la formación de gametos y al proceso de fecundación (62). En relación con esto, los espermatozoides expuestos al calor han reducido la longevidad dentro de la unión uterovaginal en las gallinas (63, 64). Además, en pollo, cuando la temperatura es superior a 32°C y la humedad es del 55-65%, se pueden encontrar morfoanomalías en las células espermatogénicas a nivel testicular y una disminución en la producción de testosterona (65, 66).
En resumen, los parámetros meteorológicos pueden influir en la reproducción animal a diferentes niveles, como la modulación del momento de la reproducción, la inversión o la supervivencia de las crías, así como originalmente en la espermatogénesis, lo que hace que el eyaculado tenga una determinada calidad (67-69). La elección de la motilidad de los espermatozoides como parámetro de referencia para determinar la calidad seminal de un eyaculado se debió a su importancia, ya que este rasgo relacionado con la calidad es un predictor fiable de la fertilidad tras un ciclo de congelación-descongelación (70, 71), y refleja varios aspectos de la fisiología de los espermatozoides, como la glucólisis, la fosforilación oxidativa y la integridad de la membrana, entre otros (72). La permeabilidad de la membrana, la composición lipídica y la fluidez de la membrana de los espermatozoides son factores determinantes para evitar daños durante el proceso de criopreservación, y el daño final de la membrana asociado a estos factores tiene consecuencias sobre la motilidad de los espermatozoides (8, 73, 74). El hecho de que la motilidad de los espermatozoides en el semen de congelación-descongelación se reduzca se debe a una disminución gradual de la energía que reduce la capacidad de los espermatozoides para realizar un movimiento adecuado para permitir la fertilización. Por lo tanto, la producción de ATP disminuye cuando la integridad mitocondrial se ve afectada, dada la alta sensibilidad de las mitocondrias de los espermatozoides de gallo a la congelación-descongelación (11).
Los espermatozoides aviares tienen una alta sensibilidad a las altas temperaturas ambientales y a los cambios de temperatura durante la criopreservación. Por lo tanto, es necesario eyacular de alta calidad antes de congelar. La evaluación de las variables morfométricas tanto en el semen fresco como en el descongelado puede ayudar a predecir la capacidad de sobrevivir a un ciclo de congelación/descongelación, ya que el tamaño de la cabeza del espermatozoide determina la permeabilidad de la membrana celular al agua y al crioprotector, lo que estaría directamente relacionado con la formación de cristales de hielo intracelulares (75, 76). En este sentido, el estudio de las variables morfométricas podría ser un criterio de calidad a añadir además de la motilidad para determinar la congelabilidad de un eyaculado.
En cualquier caso, como sugiere el presente trabajo, la elección del momento de la colección seminal es importante, ya que los rasgos climáticos y relacionados con las fases lunares pueden afectar la congelabilidad de los espermatozoides. Esto podría deberse a cambios en la composición de los espermatozoides tanto a nivel enzimático como bioquímico, así como a alteraciones a nivel hormonal, que afectarían a la permeabilidad de la membrana, dando lugar a una mayor susceptibilidad a la toxicidad generada por los crioprotectores y al resto de lesiones que se producen durante el proceso de criopreservación (77-79).
5 Conclusión
En conclusión, la climatología y el ciclo lunar afectan a la congelabilidad de los espermatozoides. Después de realizar un DCA, las variables de temperatura mínima, luna nueva como fase lunar, presión barométrica mínima, precipitación, gibosa creciente y creciente menguante como fase lunar proporcionan una gran cantidad de información y muestran un alto poder de discriminación a la hora de diferenciar entre grupos de calidad espermática. En las especies de gallos, cuando la temperatura es baja, el tamaño testicular disminuye, lo que lleva a una disminución de la calidad seminal antes y después de la descongelación. Se informó que las variaciones en la producción de melatonina debidas a la fase lunar afectaban la calidad del semen. Por lo tanto, se encontraron resultados menos deseables en las fases de luna nueva. Este estudio ha desarrollado una herramienta que nos permitirá optimizar el trabajo de criopreservación de semen de aves de corral en centros de reproducción animal. En la actualidad, no se han obtenido buenos resultados en la criopreservación de semen de gallo. Sin embargo, la selección de los momentos óptimos para la congelación del semen puede representar un avance en la mejora de la calidad del semen post-descongelación en esta especie. A través de la conservación de los recursos genéticos de las razas avícolas en peligro de extinción, se puede garantizar la diversidad genética. Por lo tanto, invertir en investigación para la mejora de los bancos de germoplasma será fundamental para la mejora de la biotecnología reproductiva en especies aviares.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio se incluyen en el artículo/material complementario, las consultas adicionales pueden dirigirse al autor correspondiente.
Declaración ética
No se requirió aprobación ética para el estudio con animales de acuerdo con la legislación local y los requisitos institucionales debido a que el presente estudio está excluido del ámbito de evaluación del Comité de Ética de la Universidad de Córdoba ya que los datos obtenidos forman parte de las actividades rutinarias que se llevan a cabo en el Centro Provincial Agrícola de la Diputación de Córdoba (Andalucía, T España) como centro de reproducción aviar y conservación de razas autóctonas y por tanto no se consideran animales utilizados con fines científicos. Sin embargo, todos los animales utilizados en el estudio fueron tratados siguiendo la legislación europea (Directiva 2010/63/UE «relativa a la protección de los animales utilizados para fines científicos»), que ha sido transpuesta al ordenamiento jurídico español a través del RD 53/2013.
Contribuciones de los autores
ED: Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. JD: Conceptualización, Obtención de Fondos, Administración de Proyectos, Recursos, Supervisión, Visualización, Redacción, Revisión y Edición. AGA: Conceptualización, Curación de datos, Análisis formal, Obtención de fondos, Investigación, Metodología, Administración de proyectos, Recursos, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. JL: Curación de datos, análisis formal, adquisición de fondos, metodología, recursos, software, supervisión, redacción, revisión y edición. AAA: Curación de datos, Investigación, Metodología, Redacción – revisión y edición. FN: Conceptualización, Curación de datos, Análisis formal, Metodología, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declaran/n que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. La presente investigación se llevó a cabo durante una beca postdoctoral Ramón y Cajal financiada por MCIN/AEI/10.13039/501100011033 y la Unión Europea «NextGenerationEU»/PRTR (Plan de Recuperación, Transformación y Resiliencia – Financiado por la Unión Europea-NextGenerationEU).
Reconocimientos
Este trabajo no hubiera sido posible sin la colaboración del Centro Provincial Agropecuary de la Diputación de Córdoba y del grupo de investigación PAIDI AGR 218.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Notas
Referencias
1. Shanmugam, M, Rajkumar, U, Reddy, M y Rao, SR. Efecto de la edad sobre la calidad del semen en pollo de cuello desnudo y enano en condiciones climáticas tropicales. Anim Prod Sci. (2012) 52:964–8. doi: 10.1071/AN12033
2. Verratti, V, Di Giulio, C, D’angeli, A, Tafuri, A, Francavilla, S, y Pelliccione, F. La motilidad hacia adelante de los espermatozoides se ve afectada negativamente por la exposición a corto plazo a la hipoxia de altitud. Andrología. (2016) 48:800–6. doi: 10.1111/and.12515
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Rao, M, Zhao, X-L, Yang, J, Hu, S-F, Lei, H, Xia, W, et al. Efecto de la hipertermia escrotal transitoria sobre los parámetros espermáticos, los marcadores bioquímicos del plasma seminal y el estrés oxidativo en hombres. Asiático J Androl. (2015) 17:668–75. doi: 10.4103/1008-682X.146967
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Longobardi, V, Zullo, G, Salzano, A, De Canditiis, C, Cammarano, A, De Luise, L, et al. El resveratrol previene los cambios similares a la capacitación y mejora la capacidad de fertilización in vitro de los espermatozoides congelados y descongelados de búfalo. Teriogenología. (2017) 88:1–8. doi: 10.1016/j.theriogenology.2016.09.046
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Vandana, G, Sejian, V, Lees, A, Pragna, P, Silpa, M, y Maloney, SK. Estrés por calor y producción avícola: impacto y mejora. Int J Biometeorol. (2021) 65:163–79. doi: 10.1007/s00484-020-02023-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Zimecki, M . El ciclo lunar: efectos sobre el comportamiento y la fisiología humana y animal. Postepy Hig Med Dosw. (2006) 60:1–7.
7. Mocé, E, y Graham, JK. Evaluación in vitro de la calidad espermática. Anim Reprod Sci. (2008) 105:104–18. doi: 10.1016/j.anireprosci.2007.11.016
8. Largo, J . Criopreservación de semen aviar: ¿cuáles son los retos biológicos? Poult Sci. (2006) 85:232–6. doi: 10.1093/ps/85.2.232
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Santiago-Moreno, J, Castaño, C, Toledano-Díaz, A, Coloma, M, López-Sebastián, A, Prieto, M, et al. Criopreservación de semen para la creación de un criobanco de razas avícolas españolas: optimización de la velocidad de congelación y del tiempo de equilibrio. Poult Sci. (2011) 90:2047–53. doi: 10.3382/ps.2011-01355
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Blesbois, E . Congelación de semen aviar. Avian Biol Res. (2011) 4:52–8. doi: 10.3184/175815511X13069413108523
11. Santiago-Moreno, J, Castaño, C, Toledano-Díaz, A, Coloma, M, López-Sebastián, A, Prieto, M, et al. Influencia de la temporada en la congelabilidad del semen de aves de corral. Reprod Domest Anim. (2012) 47:578–83. doi: 10.1111/j.1439-0531.2011.01921.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. MAPA . Bancos de germoplasma animal. (2024). Disponible en: https://www.mapa.gob.es/es/ganaderia/temas/zootecnia/razas-ganaderas/bancos-germoplasma/.
13. Churchil, RR, Praveena, PE y Sharma, D. Parámetros de calidad del semen, su interrelación y atributos de espermatozoides posteriores al lavado de gallos rojos de Rhode Island. Mundo Veterinario. (2014) 7:1117–22. doi: 10.14202/vetworld.2014.1117-1122
14. Gandini, G, y Oldenbroek, K. Estrategias para pasar de la conservación a la utilización En: K Oldenbroek editor. Utilización y conservación de los recursos genéticos de los animales de granja. Wageningen, Países Bajos: Wageningen Academic Publishers (2007). 29–54.
15. Campo, J . Razas de gallinas autóctonas andaluzas: Andaluza Azul, Andaluza Utrerana y Combatiente Español. Patrimonio ganadero andaluz: las razas ganaderas de Andalucía (Volumen II); Consejería de Agricultura y Pesca. (2007).
16. Barba, C, Fernández-Tomillo, L, Jiménez, R, Guzmán, J, y García, A. Valor ecológico ambiental y conservación de razas ovinas autóctonas en peligro de extinción en Andalucía. Arq. Zootec. (2016) 65:445–8.
17. González Ariza, A, Arando Arbulu, A, León Jurado, JM, Navas González, FJ, Delgado Bermejo, JV, y Camacho Vallejo, ME. Herramienta canónica discriminante para la caracterización biométrica diferencial de razas multivarietales de gallinas en peligro de extinción. Animales. (2021) 11:2211. doi: 10.3390/ani11082211
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Madrigueras, W, y Quinn, J. La colección de espermatozoides de las aves domésticas y de Turquía. Poult Sci. (1937) 16:19–24. doi: 10.3382/ps.0160019
19. Sasaki, K, Tatsumi, T, Tsutsui, M, Niinomi, T, Imai, T, Naito, M, et al. Un método para criopreservar el semen de los gallos de Yakido utilizando N-metilacetamida como agente crioprotector. J Poult Sci. (2010) 47:297–301. doi: 10.2141/jpsa.009111
20. Boes, J, Boettcher, P, y Honkatukia, M. Innovaciones en crioconservación de recursos zoogenéticos: guía práctica. Roma, Italia: Organización de las Naciones Unidas para la Alimentación y la Agricultura (2023).
21. Poulsen, J, y French, A. Análisis de funciones discriminantes. San Francisco, CA: Universidad Estatal de San Francisco (2008).
22. Navas, CM, Bermejo, JVD, McLean, AK, Jurado, JML, y González, FJN. Análisis canónico discriminante de la contribución de los caballos de pura raza española y árabe a la diversidad genética y estructura poblacional de los caballos hispano-árabes. Animales. (2021) 11:269. doi: 10.3390/ani11020269
23. Rogerson, Pensilvania . Reducción de datos: análisis factorial y análisis de conglomerados. Métodos estadísticos Geogr. (2001) 2001:192–7. doi: 10.4135/9781849209953.n10
24. Zhang, Q, Hu, J y Bai, Z. Estadísticos de traza de Pillai modificados para dos matrices de covarianza muestral de alta dimensión. J Inferencia del plan de estadísticas. (2020) 207:255–75. doi: 10.1016/j.jspi.2020.01.002
25. Anuthama, K, Shankar, S, Ilayaraja, V, Kumar, GS y Rajmohan, M. Determinación del dimorfismo sexual dental en el sur de la India mediante el análisis de la función discriminante. Ciencias Forenses Int. (2011) 212:86–9. doi: 10.1016/j.forsciint.2011.05.018
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. González Ariza, A, Arando Arbulu, A, Navas González, FJ, Delgado Bermejo, JV, y Camacho Vallejo, ME. Análisis canónico discriminante como herramienta de validación para la clasificación de la calidad comercial del huevo multivariedad de razas autóctonas. Seguridad de alimentos. (2021) 10:632. doi: 10.3390/foods10030632
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Toalombo Vargas, PA, Navas González, FJ, Landi, V, León Jurado, JM, y Delgado Bermejo, JV. Dimorfismo sexual y caracterización de razas de gallinas criollas mediante análisis discriminante canónico biométrico en áreas agroecológicas ecuatorianas. Animales. (2019) 10:32. doi: 10.3390/ani10010032
29. Le Blancq, F . Variación de la presión diurna: la marea atmosférica. Clima. (2011) 66:306–7. DOI: 10.1002/wea.857
30. Neild, R . Temperaturas máximas-mínimas como base para evaluar la respuesta termoperiódica. Lun Tiempo Rev. (1967) 95:583–4. doi: 10.1175/1520-0493(1967)095<0583:MMTAAB>2.3.CO; 2
31. Subramaniam, K, y Padalkar, S. Visualización y razonamiento en la explicación de las fases de la luna. Int J Sci Educ. (2009) 31:395–417. DOI: 10.1080/09500690802595805
32. Das, S, Dodd, S, Lewis-Jones, DI, Patel, FM, Drakeley, AJ, Kingsland, CR, et al. ¿Afectan las fases lunares a las tasas de concepción en reproducción asistida? J Assist Reprod Genet. (2005) 22:15–8. doi: 10.1007/s10815-005-0815-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Ellis, HI, y Jehl, JR Jr. Agua corporal total y composición corporal en falaropos y otras aves. Physiol Zool. (1991) 64:973–84. doi: 10.1086/physzool.64.4.30157952
34. Nishimura, T, y Fukushima, M. Por qué los animales responden a la luna llena: hipótesis magnética. Hipótesis de Biosci. (2009) 2:399–401. doi: 10.1016/j.bihy.2009.06.006
35. Yonezawa, T, Uchida, M, Tomioka, M y Matsuki, N. El ciclo lunar influye en el parto espontáneo en las vacas. PLoS Uno. (2016) 11:e0161735. doi: 10.1371/journal.pone.0161735
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Chinchilla-Vargas, J, Kerns, K, y Rothschild, MF. Efectos lunares y climáticos sobre los rasgos de eyaculación de verracos. Anim Reprod Sci. (2018) 193:117–25. doi: 10.1016/j.anireprosci.2018.04.006
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Kollerstrom, N . Efecto lunar en la fertilidad de yeguas pura sangre: un análisis de 14 años de datos, 1986-1999. Biol Rhythm Res. (2004) 35:317–27. doi: 10.1080/0929-1010400000624
38. Møller, A . Tendencias a largo plazo en la velocidad del viento, abundancia de insectos y ecología de un ave insectívora. Ecosfera. (2013) 4:1–11. doi: 10.1890/ES12-00310.1
39. Amélineau, F, Péron, C, Lescroël, A, Authier, M, Provost, P, y Grémillet, D. El paisaje eólico y la tortuosidad dan forma a los costos de vuelo de los alcatraces del norte. J Exp Biol. (2014) 217:876–85. doi: 10.1242/jeb.097915
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Dehnhard, N, Ludynia, K, Poisbleau, M, Demongin, L, y Quillfeldt, P. Días buenos, días malos: el viento como impulsor del éxito de la búsqueda de alimento en un ave marina no voladora, el pingüino penacho amarillo del sur. PLoS Uno. (2013) 8:e79487. doi: 10.1371/journal.pone.0079487
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Overton, M, Sischo, W, Temple, G y Moore, D. Uso de la fotografía de video de lapso de tiempo para evaluar el comportamiento de la vaca lechera en un establo de establo libre. J Dairy Sci. (2002) 85:2407–13. doi: 10.3168/jds. S0022-0302(02)74323-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Schou, MF, Bonato, M, Engelbrecht, A, Brand, Z, Svensson, EI, Melgar, J, et al. Las temperaturas extremas comprometen la fertilidad de machos y hembras en un ave grande del desierto. Nat Commun. (2021) 12:666. doi: 10.1038/s41467-021-20937-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Moreno, RD, Lagos-Cabré, R, Buñay, J, Urzúa, N, y Bustamante-Marín, X. Bases moleculares del daño por estrés térmico en testículos de mamíferos. Testículos. (2012):127–55.
44. Huston, TM . Los efectos de la temperatura ambiental sobre la fertilidad de las aves domésticas. Poult Sci. (1975) 54:1180–4. doi: 10.3382/ps.0541180
45. Perfito, N, Tramontin, AD, Meddle, S, Sharp, P, Afik, D, Gee, J, et al. Desarrollo reproductivo de acuerdo con la elevación en un ave cantora macho que se reproduce estacionalmente. Ecología. (2004) 140:201–10. doi: 10.1007/s00442-004-1576-5
46. Silverin, B, Wingfield, J, Stokkan, K-A, Massa, R, Järvinen, A, Andersson, N-Å, et al. Efectos de la temperatura ambiente sobre los ciclos gonadales fotoinducidos y los patrones de secreción hormonal en carboneros comunes de tres latitudes de cría diferentes. Horm Compórtate. (2008) 54:60–8. doi: 10.1016/j.yhbeh.2008.01.015
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. York, JE, Young, AJ y Radford, AN. Cantando a la luz de la luna: la interpretación del canto del amanecer de un ave diurna varía con la fase lunar. Biol Lett. (2014) 10:20130970. doi: 10.1098/rsbl.2013.0970
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Andrade, RM, Rojas, JA, Espinoza, MM, y Viamonte, KR. Influencia lunar en cultivos, animales y ser humano. Uniandes Episteme Revista de Ciencia, Tecnología e Innovación, no. 4, pp. 37-47. (2017).
49. Tarlow, EM, Hau, M, Anderson, DJ y Wikelski, M. Diel cambios en las concentraciones plasmáticas de melatonina y corticosterona en piqueros tropicales de Nazca (Sula granti) en relación con la fase lunar y la edad. Gen Comp Endocrinol. (2003) 133:297–304. doi: 10.1016/S0016-6480(03)00192-8
50. Awad, H, Halawa, F, Mostafa, T y Atta, H. Perfil hormonal de melatonina en hombres infértiles. Int J Androl. (2006) 29:409–13. doi: 10.1111/j.1365-2605.2005.00624.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Ramadan, T, Taha, T, Samak, M, y Hassan, A. Efectividad de la exposición a un día largo seguido de un tratamiento con melatonina en las características del semen de cabras macho de Damasco durante las temporadas de reproducción y no reproducción. Teriogenología. (2009) 71:458–68. doi: 10.1016/j.theriogenology.2008.07.029
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Casao, A, Vega, S, Palacín, I, Pérez-Pe, R, Laviña, A, Quintín, F, et al. Efectos de los implantes de melatonina durante la época no reproductiva sobre la motilidad espermática y los parámetros reproductivos en carneros rasa aragonesa. Reprod Domest Anim. (2010) 45:425–32. doi: 10.1111/j.1439-0531.2008.01215.x
53. Shestakov, V, Ermoshina, E, Korolev, V, Pimkina, T, y Koroleva, E. Variabilidad de la función generativa de toros de diferentes razas y su tolerancia en relación con el cambio en las fases lunares En: V Shestakov editor. Serie de conferencias del IOP: Ciencias de la Tierra y del medio ambiente. Bristol, Reino Unido: IOP Publishing (2021)
54. Moretti, E, Tallis, V, Trovarelli, S, Gnech, M, Capitani, S, Ponchietti, R, et al. ¿Influyen las fases lunares en los parámetros del semen? J Androl Sci. (2008) 15:158–63.
55. Díaz-Usi, J, Venturina, E, Peralta, M, Durán, P, Mingala, C, Medina, N, et al. Efecto de los cambios semanales en los parámetros ambientales sobre las características de motilidad de los espermatozoides de los toros de búfalo de agua (Bubalus bubalis Linn.). AIP Conf Proc, (2023), 2628: 070010. doi: 10.1063/5.0145393
56. Abilov, A, Amerhanov, CA, Korneyenko-Zhilyaev, YA, Pyzhova, E, Kombarova, N, Vinogradova, I, et al. Efecto de la presión atmosférica sobre los parámetros seminales en toros de selección moderna el día de la recogida. Agric Biol. (2017) 52:314–22. doi: 10.15389/agrobiology.2017.2.314eng
58. Abecia, J, Arrébola, F, Macías, A, Laviña, A, González-Casquet, O, Benítez, F, et al. La temperatura y la pluviometría están relacionadas con la tasa de fertilidad después de la inseminación artificial de primavera en pequeños rumiantes. Int J Biometeorol. (2016) 60:1603–9. doi: 10.1007/s00484-016-1150-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Van Tilburg, M, Salles, M, Silva, M, Moreira, R, Moreno, F, Monteiro-Moreira, A, et al. Variables seminales y perfil proteico de la membrana espermática de machos Saanen (Capra hircus) en épocas secas y lluviosas del nordeste de Brasil (3 S). Int J Biometeorol. (2015) 59:561–73. doi: 10.1007/s00484-014-0869-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Smith, EM, y Blalock, JE. Un bucle regulador completo entre los sistemas inmunitario y neuroendocrino opera a través de moléculas de señalización comunes (hormonas) y receptores. Proc. (1985) 44:108–11. doi: 10.1007/978-1-4899-0557-4_11
61. Mitchell, M, y Kettlewell, P. Estrés fisiológico y bienestar de los pollos de engorde en tránsito: ¡soluciones, no problemas! Poult Sci. (1998) 77:1803–14. doi: 10.1093/ps/77.12.1803
Resumen de PubMed | Texto completo de Crossref | Google Académico
62. Mcdaniel, CD, Bramwell, RK, Wilson, JL y Howarth, B Jr. Fertilidad de reproductoras de pollos de engorde machos y hembras después de la exposición a temperaturas ambientales elevadas. Poult Sci. (1995) 74:1029–38. doi: 10.3382/ps.0741029
Resumen de PubMed | Texto completo de Crossref | Google Académico
63. Mcdaniel, CD, Bramwell, RK, y Howarth, JRB. La contribución de los machos a la infertilidad inducida por el calor de las reproductoras de pollos de engorde, determinada por la penetración de los espermatozoides y los óvulos y el almacenamiento de espermatozoides dentro del oviducto de la gallina. Poult Sci. (1996) 75:1546–54. doi: 10.3382/ps.0751546
Resumen de PubMed | Texto completo de Crossref | Google Académico
64. King, L, Brillard, J, Garrett, W, Bakst, M y Donoghue, A. Segregación de espermatozoides dentro de los túbulos de almacenamiento de esperma de aves y gallinas de pavo. Reproducción. (2002) 123:79–86. doi: 10.1530/rep.0.1230079
Resumen de PubMed | Texto completo de Crossref | Google Académico
65. Chen, Z, Zhang, J, Zhou, Y, Liang, C y Jiang, Y. Efecto del estrés por calor en el desarrollo pituitario y testicular de los pollitos Wenchang. Raza Arch Anim. (2015) 58:373–8. doi: 10.5194/aab-58-373-2015
66. Xiong, Y, Yin, Q, Li, J y He, S. El estrés oxidativo y el estrés del retículo endoplásmico están involucrados en el efecto protector del ácido alfa lipoico contra el daño por calor en los testículos de pollo. Animales. (2020) 10:384. doi: 10.3390/ani10030384
Resumen de PubMed | Texto completo de Crossref | Google Académico
67. Møller, AP, Fiedler, W, y Berthold, P. Efectos del cambio climático en las aves. Oxford, Reino Unido: OUP Oxford (2010).
68. Visser, ME, Both, C, y Lambrechts, MM. El cambio climático global conduce a la reproducción aviar a destiempo. Adv Ecol Res. (2004) 35:89–110. doi: 10.1016/S0065-2504(04)35005-1
69. Karaca, A, Parker, H y McDaniel, C. La temperatura corporal elevada contribuye directamente a la infertilidad por estrés térmico de los machos reproductores de pollos de engorde. Poult Sci. (2002) 81:1892–7. doi: 10.1093/ps/81.12.1892
Resumen de PubMed | Texto completo de Crossref | Google Académico
70. Nguyen, TMD . Principales vías de señalización implicadas en el control de la motilidad de los espermatozoides de las aves. Poult Sci. (2019) 98:1528–38. doi: 10.3382/ps/pey465
Resumen de PubMed | Texto completo de Crossref | Google Académico
71. Blesbois, E, Grasseau, I, Seigneurin, F, Mignon-Grasteau, S, Saint Jalme, M, y Mialon-Richard, M. Predictores del éxito de la criopreservación de semen en pollos. Teriogenología. (2008) 69:252–61. doi: 10.1016/j.theriogenology.2007.09.019
Resumen de PubMed | Texto completo de Crossref | Google Académico
72. Van der Horst, G . Estado de la evaluación de la funcionalidad espermática en especies silvestres: de peces a primates. Animales. (2021) 11:1491. doi: 10.3390/ani11061491
Resumen de PubMed | Texto completo de Crossref | Google Académico
74. Blesbois, E . Fluidez de la membrana y capacidad de sobrevivir a la criopreservación en espermatozoides de aves domésticas. Reproducción. (2005) 129:371–8. doi: 10.1530/rep.1.00454
Resumen de PubMed | Texto completo de Crossref | Google Académico
75. Esteso, MC, Soler, AJ, Fernández-Santos, MR, Quintero-Moreno, AA, y Garde, JJ. Significado funcional del tamaño y la forma morfométrica de la cabeza del espermatozoide para la determinación de la congelabilidad en muestras de semen de epidídimo de ciervo ibérico (Cervus elaphus hispanicus). J Androl. (2006) 27:662–70. doi: 10.2164/jandrol.106.000489
76. Peña, FJ, Saravia, F, García-Herreros, M, Núñezmartínez, I, Tapia, JA, Johannisson, A, et al. Identificación de subpoblaciones morfométricas de espermatozoides en dos porciones diferentes del eyaculado de verraco y su relación con la calidad post-descongelación. J Androl. (2005) 26:716–23. doi: 10.2164/jandrol.05030
Resumen de PubMed | Texto completo de Crossref | Google Académico
77. Bentley, L, Ansah, G, y Buckland, R. Proteínas plasmáticas seminales de una línea de pollos seleccionados para la fertilidad del semen congelado-descongelado y la línea de control. Poult Sci. (1984) 63:1444–5. doi: 10.3382/ps.0631444
Resumen de PubMed | Texto completo de Crossref | Google Académico
78. Blesbois, E, y De Reviers, M. Efecto de diferentes fracciones de plasma seminal sobre la capacidad fertilizante de espermatozoides de aves almacenados in vitro. Reproducción. (1992) 95:263–8. doi: 10.1530/jrf.0.0950263
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: reproducción aviar, condición climática, ciclo lunar, raza en peligro de extinción, motilidad, criopreservación de espermatozoides, congelabilidad de espermatozoides
Cita: Díaz Ruiz E, Delgado Bermejo JV, González Ariza A, León Jurado JM, Arando Arbulu A y Navas González FJ (2024) Efectos de la meteorología y el ciclo lunar en la calidad post-descongelación de espermatozoides aviares. Frente. Vet. Sci. 11:1394004. doi: 10.3389/fvets.2024.1394004
Editado por:
Abdulmojeed Yakubu, Universidad Estatal de Nasarawa, Nigeria
Revisado por:
Nucharin Songsasen, Instituto Smithsonian de Biología de la Conservación (SI), Estados
Unidos Cristina Soriano-Úbeda, Universidad de León, España
Copyright © 2024 Díaz Ruiz, Delgado Bermejo, González Ariza, León Jurado, Arando Arbulu y Navas González. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Antonio González Ariza, angoarvet@outlook.es
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía