
Efectos de la suplementación con ácido guanidina acético desde la gestación hasta la lactancia sobre el rendimiento reproductivo

Efectos de la suplementación con ácido guanidina acético desde la gestación hasta la lactancia sobre el rendimiento reproductivo, la calidad del calostro, la bioquímica sanguínea y la diversidad de la microflora intestinal de las cerdas
 Guanglei Cong1
Guanglei Cong1 Shuangshuang Xia1Chunxue Liu1,2*Junbo Li1Ifen Hung1
Shuangshuang Xia1Chunxue Liu1,2*Junbo Li1Ifen Hung1- 1Anyou Biotechnology Group Co., Ltd., Taicang, Jiangsu, China
- número arábigoFacultad de Ciencia y Tecnología Animal, Universidad de Agricultura de Nanjing, Nanjing, Jiangsu, China
Este experimento tuvo como objetivo estudiar los efectos del ácido guanidina acético (GAA) sobre el rendimiento reproductivo, el rendimiento de la lactancia y los índices bioquímicos sanguíneos de las cerdas, así como el rendimiento de los lechones de descendencia. Se utilizaron un total de 20 cerdas (Landrace × Yorkshire, paridad 4). La mitad de las cerdas de cada parto fueron alimentadas con una dieta control (GC; dieta básica, n = 10) o dieta GAA (dieta básica +1 g/kg GAA, n = 10) desde el día 85 de gestación hasta el destete. Los resultados del estudio se presentan de la siguiente manera: La suplementación con GAA desde el final de la gestación hasta la lactancia no afectó negativamente la ingesta de alimento de las cerdas, el grosor de la grasa dorsal o los índices de rutina sanguínea (p > 0,05). La suplementación con GAA mostró una tendencia a aumentar el número de lechones sanos y su actividad al nacer (p = 0,06; p = 0,08), mientras que aumentó significativamente la puntuación del RCIU de los lechones (p < 0,05). La suplementación con GAA aumentó significativamente el contenido de proteína del calostro (p < 0,05) y tendió a aumentar la producción diaria de leche en las cerdas (p = 0,07). La suplementación con GAA aumentó el nivel de inmunoglobulina A en el calostro de las cerdas (p < 0,05) y mostró una tendencia a aumentar el contenido de prolina (p = 0,10). La suplementación con GAA disminuyó significativamente el contenido de triglicéridos en la sangre del cordón umbilical de las cerdas (p < 0,05), sin que se observaran efectos significativos sobre HDL-C, LDL-C, TC y GLU (p > 0,05). La suplementación con GAA aumentó significativamente los niveles de eNOS en la sangre del cordón umbilical de las cerdas (p < 0,05), mientras que no mostró efectos significativos sobre la IL-6 y la IL-10 (p > 0,05). La suplementación con GAA no afectó significativamente la diversidad α de la flora intestinal de las cerdas (ACE, Shannon, Chao1, Simpson, observed_otus, pielou_e y good_cover), pero el análisis de PCoA reveló diferencias en la estructura de la flora intestinal entre los grupos. Además, GAA disminuyó la abundancia relativa de Sarciha y unidentified_ruminococcaceae y aumentó la abundancia relativa de Lactobacillus, Parabacteroides y Pedobacter en el intestino. El GAA aumenta la óxido nítrico sintasa en la sangre del cordón umbilical de las cerdas, lo que mejora el desarrollo de los vasos sanguíneos de la placenta. Esto mejora la salud y la vitalidad de los lechones, aumenta las bacterias intestinales beneficiosas (Lactobacillus, Parabacteroides, Pedobacter) y eleva los niveles de proteína del calostro y el volumen de la lactancia, lo que conduce a un mejor crecimiento y rendimiento de los lechones.
1 Introducción
El ácido guanidina acético (GAA) es indispensable para la síntesis de creatina en vertebrados y desempeña un papel fundamental en el metabolismo energético de las células somáticas en animales (1, 2). La L-arginina glicina amidinotransferasa cataliza la conversión de arginina y glicina en GAA en los riñones, que posteriormente se transporta a través del torrente sanguíneo hasta el hígado. Aquí, reacciona con la n-dimetiltransferasa y la S-adenosilmetionina para producir creatina (3, 4). La creatina sirve como un depósito de energía vital en los tejidos animales, convirtiendo rápidamente el fosfato de creatina en ATP.
En los últimos años, la reducción del uso de proteína animal en los piensos para el ganado chino ha aumentado la preocupación por la pérdida de creatina durante el procesamiento de los piensos y el tratamiento térmico. Esto ha llevado a un mayor interés en la suplementación con creatina exógena, a pesar de su costo, inestabilidad y posible inhibición de la síntesis endógena de creatina (5, 6). Por el contrario, el GAA exhibe estabilidad y eficacia superior en comparación con la creatina para mejorar los niveles de creatina en animales (7), lo que lo convierte en la opción preferida para la suplementación con creatina.
Los estudios han explorado la suplementación con GAA en lechones destetados y cerdos de engorde, revelando que el 0,12% de GAA mejora la ganancia diaria promedio (ADG) y la eficiencia alimenticia (Gratio) a lo largo de la fase de crecimiento, aumenta el rendimiento de carne magra y reduce el grosor de la grasa dorsal (8). He et al. (9) informaron mejoras en el rendimiento del crecimiento con 300 mg/kg de GAA, atribuidas a niveles elevados de creatina y ATP en los tejidos. Además, la suplementación con 2,4 g/kg de GAA durante la etapa de destete mejora la eficiencia de la conversión alimenticia y regula al alza las expresiones de ARNm de mTOR y AMPK en el músculo esquelético (10). La suplementación con 0,06% de GAA durante las etapas tardías de engorde aumenta la GMD y el porcentaje de carne magra al influir en el desarrollo muscular a través de la modulación de la expresión génica de mioblastos y las características de las fibras musculares (11). Además, la suplementación alrededor de 86 kg de peso corporal no mejora el rendimiento del crecimiento, pero aumenta los niveles de aminoácidos libres, sus metabolitos en plasma y tejidos, y enzimas antioxidantes (12, 13).
Sin embargo, la investigación sobre el impacto de GAA en la producción de cerdas sigue siendo limitada y arroja resultados variados (14, 15). Por lo tanto, este estudio investiga los efectos de la suplementación con GAA durante la gestación tardía y la lactancia en el rendimiento reproductivo de las cerdas, la composición del calostro (incluido el contenido de inmunoglobulinas y aminoácidos), los parámetros rutinarios de la sangre del cordón umbilical, los niveles de óxido nítrico en la sangre del cordón umbilical, los marcadores inflamatorios, la flora intestinal de las cerdas y el rendimiento del crecimiento de los lechones lactantes.
2 Materiales y métodos
El experimento se llevó a cabo en la unidad experimental porcina de Anyou Biotechnology Group Co., Ltd. (Nanjing, Chian). Todos los procedimientos experimentales fueron aprobados por el Comité de Ética en el Uso de Animales (CEUA) de Anyou Biological Technology Group Co., Ltd (Nº: ANS-CEUA-PJT/ PL/202309/084).
2.1 Diseño experimental, dietas y manejo
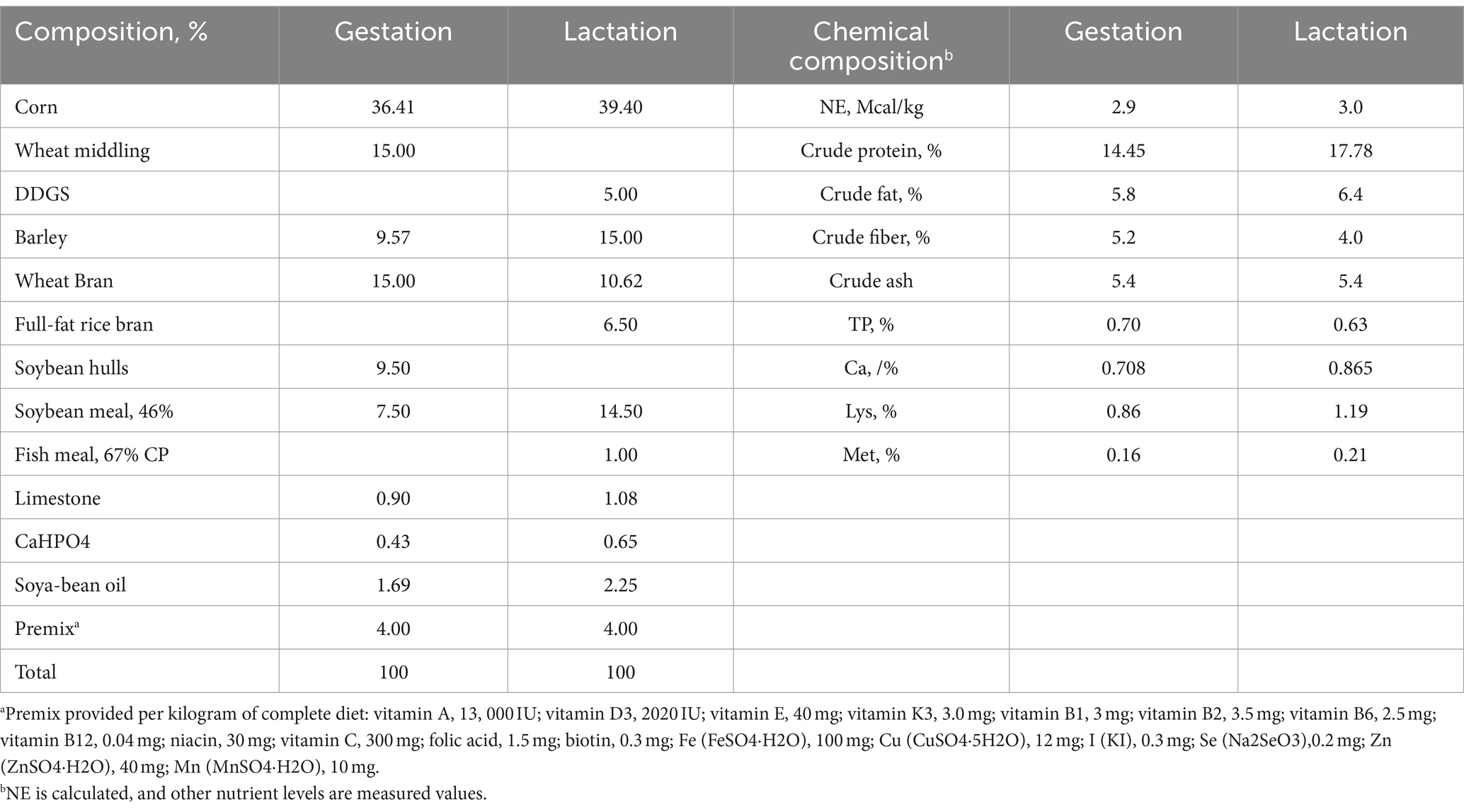
Se utilizaron un total de 20 cerdas (Landrace × Yorkshire, paridad 4). La mitad de las cerdas de cada parto fueron alimentadas con una dieta control (GC; dieta básica, n = 10) o dieta GAA (dieta básica +1 g/kg GAA, polvo blanco con 98% de pureza, n = 10) desde el día 85 de gestación hasta el destete. La composición detallada y los niveles de nutrientes de las dietas basales se describen en la Tabla 1.
En el momento del parto (el día de parto se tomó como el día 0 de lactancia), se registró el número de lechones vivos, lechones nacidos muertos (los lechones deformes se consideraron nacidos muertos) y fetos momificados, y se midió individualmente el peso al nacer de los lechones vivos. Teniendo en cuenta el mismo número de lechones vivos totales que se produjeron por tratamiento, no se realizó la acogida cruzada. Se programó que los lechones recibieran marcas auriculares de plástico y hierro suplementario, así como procedimientos de rutina para el corte de la cola, el corte de dientes y la castración al tercer día de edad. Los lechones se mantuvieron en incubadoras configuradas a 22 ~ 32 ° C, con la temperatura controlada por lámparas de calefacción suplementarias. El alimento (proporcionado a las 07:30, 13:00 y 18:00 h) se administró a partir de 1 kg en el día 1 de lactancia y se incrementó gradualmente en 1,0 kg por día hasta el día 6. Después de eso, la cerda pudo acceder libremente para alimentarse hasta el día 21 de lactación. Durante la lactancia, se registró diariamente el consumo de alimento de cada cerda.
2.2 Medición
Se midió el grosor de la LM y el peso corporal de las cerdas no alimentadas en el 85º día prenatal y en el 1º y 21º días postnatales. El grosor del BF se midió en la línea media dorsal del lado izquierdo (distancia 65 mm) de la 10ª costilla con ultrasonido (Shu Shuang® Lean-Meater, China).
Para calcular el peso medio de los lechones por camada y el peso medio de los lechones nacidos vivos, se registró el peso de cada lechón durante el parto (antes de comer el calostro). Se registró el número de lechones por camada (incluyendo el total de lechones nacidos, nacidos vivos, sanos, lechones débiles y fetos momificados y nacidos muertos). Los lechones sanos se refieren a aquellos con un peso al nacer de ≥0,8 kg, mientras que los lechones débiles son aquellos con un peso al nacer de <0,8 kg (según las normas de gestión de la granja). Registre la duración del parto. La duración del parto significa el tiempo desde el nacimiento del primer lechón hasta el nacimiento del último lechón de la camada. Se registró la ingesta diaria de alimento de cada cerda para calcular la ingesta total de alimento y la ingesta diaria promedio de alimento.
La evaluación visual de la escala de vitalidad de los lechones (VS) se realizó inmediatamente después del nacimiento según Baxter et al. (16). También se midió el retraso del crecimiento intrauterino (RCIU) después del nacimiento, con una puntuación de 1 que indica un desarrollo normal. Una puntuación de 2 indica un RCIU leve, con al menos un RCIU. Una puntuación de 3 indica RCIU grave en lechones con al menos una presentación de RCIU (17).
A los 5 y 21 días de lactancia se pesó el peso corporal de los lechones individuales. Durante el período postnatal, se registró diariamente la mortalidad y diarrea de los lechones. Al mismo tiempo, se registró la ingesta de alimento de las cerdas desde el día 85 de gestación hasta el día 21 de lactación.
2.3 Recogida de muestras
Al parto del parto, se seleccionaron aleatoriamente seis cerdas para recolectar sangre de las venas umbilicales (se seleccionaron aleatoriamente seis venas umbilicales en cada cerda). Las muestras de suero se obtuvieron centrifugando la sangre a 3000 r/min y 4 °C durante 15 min y luego se almacenaron a -80 °C hasta el análisis. Se recolectaron alrededor de 20 mL de calostro del tercer y cuarto par de cabezas de agujero en un lado de la cerda a las 8 h después del parto, se mezclaron suavemente y se almacenaron a -20 °C hasta el análisis.
2.3.1 Determinación del nivel sanguíneo rutinario de sangre de cordón umbilical
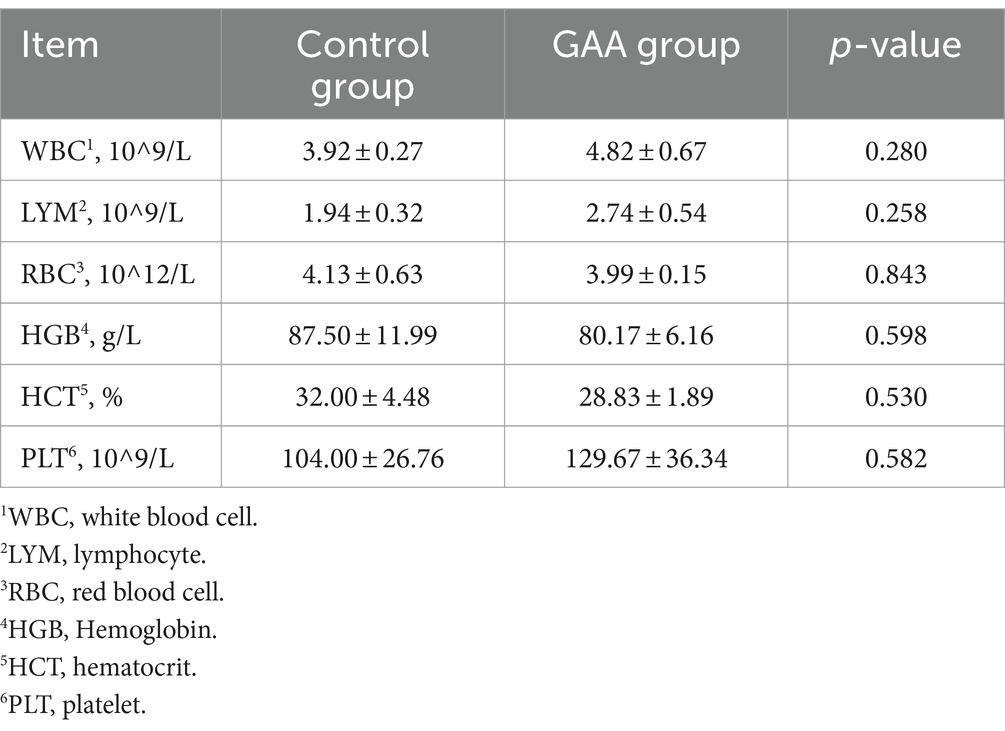
Los niveles de glóbulos blancos (WBC), linfocitos (LYM), glóbulos rojos (RBC), hemoglobina (HGB), hematocrito (HCT) y plaquetas (PLT) en la sangre del cordón umbilical de las cerdas se determinaron mediante el analizador automático de hematología de cinco clasificaciones de animales veterinarios BH-5160 (URIT, China).
2.3.2 Análisis del contenido de hormonas y metabolitos en la sangre del cordón umbilical
Los contenidos de óxido nítrico sintasa endotelial (eNOS) se analizaron utilizando los respectivos kits de ensayo de inmunoabsorción enzimática (ELISA) (Jiangsu Meimian Industrial Co., Ltd., China) siguiendo las instrucciones del fabricante y se determinaron las concentraciones séricas de glucosa (GLU), triglicéridos (TG), colesterol total (TC), colesterol de lipoproteínas de alta densidad (HDL-C) y colesterol de lipoproteínas de baja densidad (LDL-C) utilizando el analizador bioquímico automático Hitachi 3.100 (Hitachi Diagnostic Productos Co., Ltd., China). Los límites mínimos de detección para LRP y eNOS fueron de 8 ngL/L y 0,1 μmol/L, respectivamente, y el coeficiente de variación (CV) intraensayo de todos los kits fue del 10%, y el CV entre ensayos fue del 12%.
El contenido de óxido nítrico (NO) se analizó utilizando los respectivos kits de ensayo (Nanjing Jiancheng Bioengineering Institute Co., Ltd., China). Todas las mediciones se realizan de acuerdo con los procedimientos del fabricante.
2.3.3 Análisis del contenido de componentes en el calostro y determinación de la producción de leche
Las muestras de calostro descongelado se analizaron utilizando el analizador de leche MilkoScan™ FT3 (FOSS, Dinamarca) para evaluar el contenido de grasa, proteína y lactosa. Los resultados se calcularon como porcentajes de calostro y leche. La evaluación de la producción de leche durante la lactancia dependió de la ganancia diaria promedio (GMD) de lechones individuales y del número de camadas según la siguiente ecuación (18): producción de leche = GMD de lechones individuales × número de camadas × días de lactancia × 4. A partir de esto, concluyó la producción media diaria de leche de las cerdas.
2.3.4 Análisis del contenido de inmunoglobulinas en el calostro
Los contenidos de inmunoglobulina A (IgA), IgG e IgM se analizaron utilizando los respectivos kits de ensayo de inmunoabsorción enzimática (ELISA) (Jiangsu Meimian Industrial Co., Ltd., China) siguiendo las instrucciones del fabricante. Los niveles más bajos detectables de IgA, IgG e IgM fueron de 1 μg/mL, 1,2 μg/mL y 12 μg/mL respectivamente, y el coeficiente de variación (CV) intraensayo de todos los kits fue del 10%, y el CV entre ensayos fue del 12%.
2.3.5 Análisis del contenido de aminoácidos en el calostro
De acuerdo con el método propuesto por Nascimento et al. (19), las muestras apropiadas se transfirieron a un tubo de hidrólisis de 50 mL, se añadieron 20 mL de HCL de 6 moL/L y luego se hidrolizaron a 110 °C durante 24 h en un horno de secado rápido eléctrico. Retirar y enfriar, transferir a un tubo colorimétrico de 25 mL de volumen constante.
Tome con precisión una muestra de 100 μL en un tubo de centrífuga de 15 ml, póngala en un horno de secado al vacío, séquela durante 2 h a 60 °C (seque todos los solventes), llene el tubo de centrífuga con nitrógeno y agregue con precisión 50 μL de reactivos derivados: Etanol: isotiocianato de fenilo: Agua: trietilamina = 7:1:1:1 (lista para usar, llena de nitrógeno durante la preparación), derivada a temperatura ambiente durante 30 min, fase móvil A añadida (31,815 g de acetato de sodio +3.880 mL de agua +120 mL de acetonitrilo), volumen fijo a 0,5 mL, bien mezclado, sobre 0,45 μm de recubrimiento de membrana orgánica.
Las concentraciones de AA del calostro se analizaron utilizando el método de análisis de oxidación en un sistema LC 1.260 Infinity II Prime (Agilent, Estados Unidos) equipado con un RP-C18 SHISEIDO (250 mm de longitud, 4,6 mm de diámetro, 5 mm de tamaño de partícula).
2.3.6 Análisis del nivel inmune en sangre de cordón umbilical
Los contenidos de interleucina-6 (IL6) e IL-10 se analizaron utilizando los respectivos kits de ensayo de inmunoabsorción enzimática (ELISA) (Jiangsu Meimian Industrial Co., Ltd., China) siguiendo las instrucciones del fabricante. Los niveles más bajos detectables de IL-6 e IL-10 fueron de 50 ng/L y 8 ng/L, respectivamente, y el coeficiente de variación (CV) intraensayo de todos los kits fue del 10%, y el CV entre ensayos fue del 12%.
2.3.7 Análisis de la diversidad de la flora intestinal
Utilizamos DNA Kit (DP328, Tiangen Biotechnology Co., Ltd.) para extraer el ADN genómico total. La integridad y concentración de ARN fueron detectadas por NanoDrop ND 2000 (Thermo, Estados Unidos). De acuerdo con el fragmento objetivo, amplificación de PCR en la región V3-V4 de 16 S rDNA, 341F-(5′-CCTAYGGGRBGCASCAG-3′) y 806R-(5′-GGACTACNNGGGTATCTAAT-3′). A continuación, utilizamos electroforesis en gel de agarosa al 1,5% para extraer productos de PCR de fragmentos de 400-450 pb, purificados con el kit de extracción de gel GeneJET (Thermo, Estados Unidos). La biblioteca fue establecida por Ion Plus Fragment Library Kit (Thermo, Estados Unidos). Después de la cuantificación de qubits y las pruebas de biblioteca, cada ADNr 16 S replicado se agrupó y se secuenció en el extremo emparejado en las plataformas de secuenciación IonS5TMXL (Novogene Biotechnology Co., Ltd., China).
Sobre la base de las plataformas de secuenciación IonS5TMXL, la calidad de las etiquetas sin procesar se filtró mediante FLASH (V1.2.7) y se extrajeron las etiquetas efectivas. Todas las etiquetas efectivas se agruparon en unidades taxonómicas operativas (OTU) con una similitud de homología del 97% mediante Uparse (V7.0.1001), utilizando la base de datos SSUrRNA para anotar estas secuencias. La diversidad alfa y beta se analizaron en función de los niveles de OTUs.
2.4 Análisis estadístico
Todos los datos se recopilaron inicialmente con Excel 2021 y se analizaron con SAS 9.4 (SAS Institute Inc.). Antes del análisis, se utilizó el procedimiento UNIVARIADO para comprobar los valores atípicos y evaluar la normalidad con un intervalo de confianza del 95%. Los datos que se ajustan o se aproximan a una distribución normal se sometieron a una prueba T de muestras independientes; cuando no se satisfizo la homogeneidad de la varianza, se aplicó el ajuste de Satterthwaite. Para los datos que no cumplían con los supuestos de normalidad, se empleó la prueba U de Mann-Whitney. Se analizó el rendimiento reproductivo de las cerdas utilizando como unidad experimental a las cerdas y sus tamaños de camada. Para el resto de datos, se utilizaron como unidad experimental muestras relevantes de cerdas seleccionadas aleatoriamente dentro de cada grupo. Los resultados se presentan como medias y errores estándar de la media (SEM). La significación estadística se determinó con p < 0,05 y 0,05 ≤ p ≤ 0,10 se consideró una tendencia.
3 Resultados
3.1 Espesor de la grasa dorsal e ingesta de alimento
Como se muestra en la Tabla 2, la suplementación dietética con GAA no mostró un impacto significativo sobre la grasa dorsal inicial, la grasa dorsal del parto, la grasa dorsal al destete, la pérdida de grasa dorsal, la ingesta de alimento durante el embarazo y la ingesta de alimento durante la lactancia (p > 0,05).
3.2 Rutina de sangre de cordón umbilical
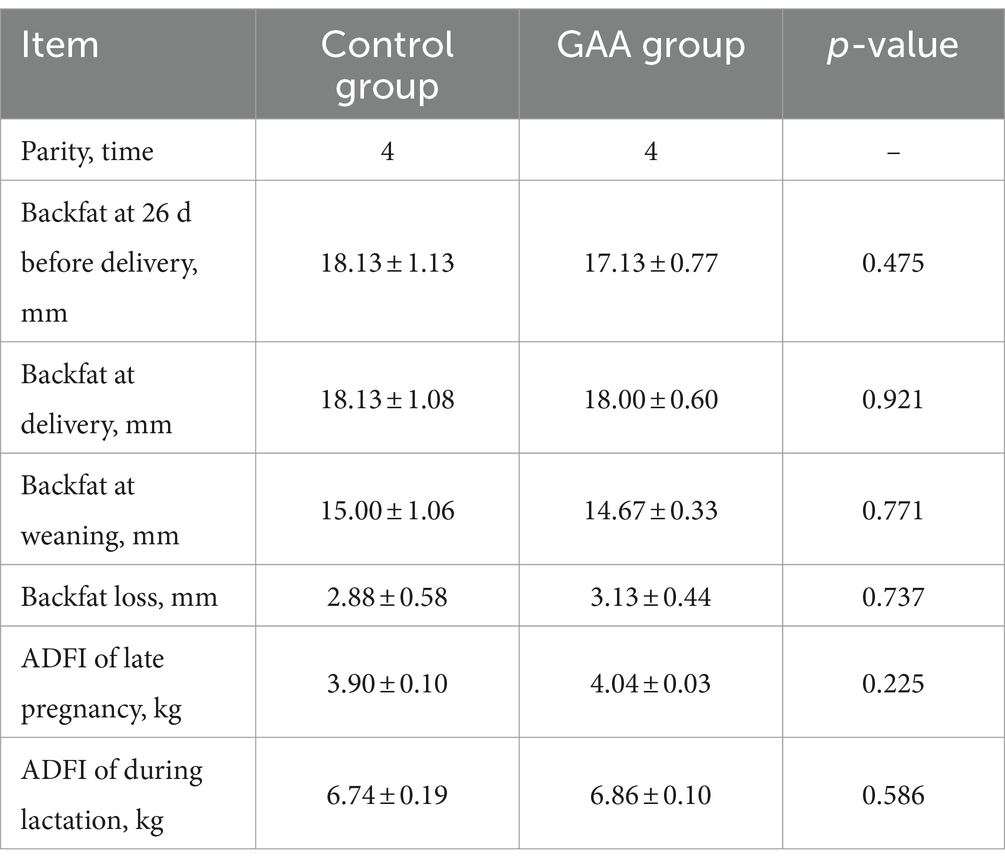
Como se muestra en la Tabla 3, la suplementación dietética con GAA no mostró efectos significativos sobre el recuento de glóbulos blancos (WBC), el recuento de linfocitos (LYM), el recuento de glóbulos rojos (RBC), la concentración de hemoglobina (HGB), el hematocrito (HCT) y el recuento de plaquetas (PLT) en la sangre del cordón umbilical de las cerdas (p > 0,05).
3.3 Rendimiento reproductivo
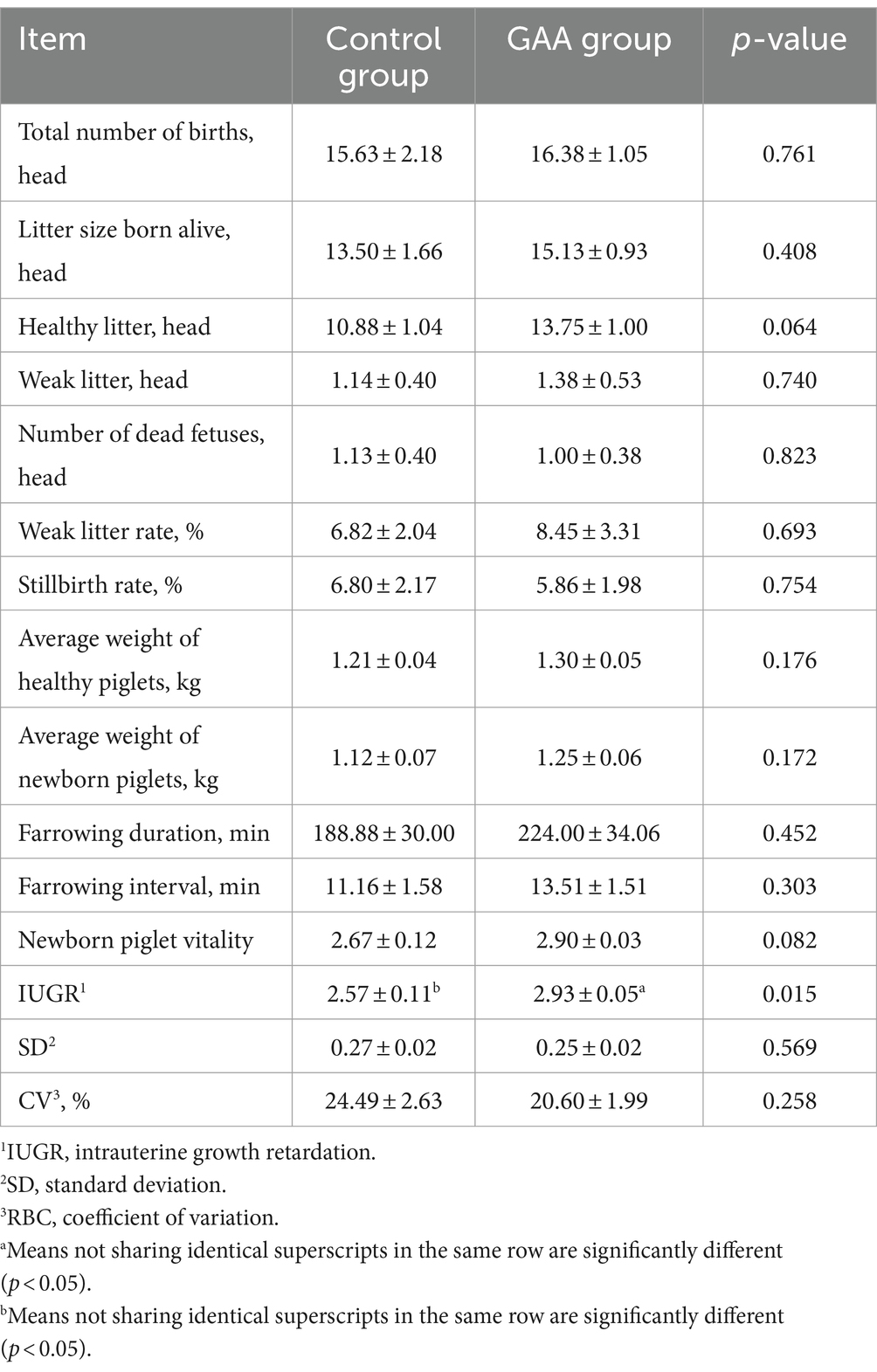
Como se muestra en la Tabla 4, la suplementación dietética con GAA aumentó significativamente la puntuación de restricción del crecimiento intrauterino (RCIU) (p < 0,05). Si bien hubo una tendencia hacia un mayor número de camadas sanas y un mayor vigor de los lechones con la suplementación con GAA (p = 0,06; p = 0,08), estos efectos no alcanzaron significación estadística. Sin embargo, la suplementación con GAA no afectó significativamente parámetros como el número total de camadas, el número de camadas vivas, el número de camadas débiles, el número de mortinatos, el peso saludable de la camada, el peso promedio de la camada viva, la tasa de mortinatos, la tasa de camada débil, la duración del parto, el intervalo entre partos y la uniformidad de los lechones en las cerdas (p > 0,05).
3.4 Composición y contenido del calostro
Como se muestra en la Tabla 5, la suplementación dietética de GAA aumentó significativamente el porcentaje de proteína de calostro en las cerdas (p < 0,05). Además, la suplementación con GAA aumentó la producción de leche (p < 0.05) sin afectar significativamente los porcentajes de grasa y lactosa del calostro (p > 0.05).
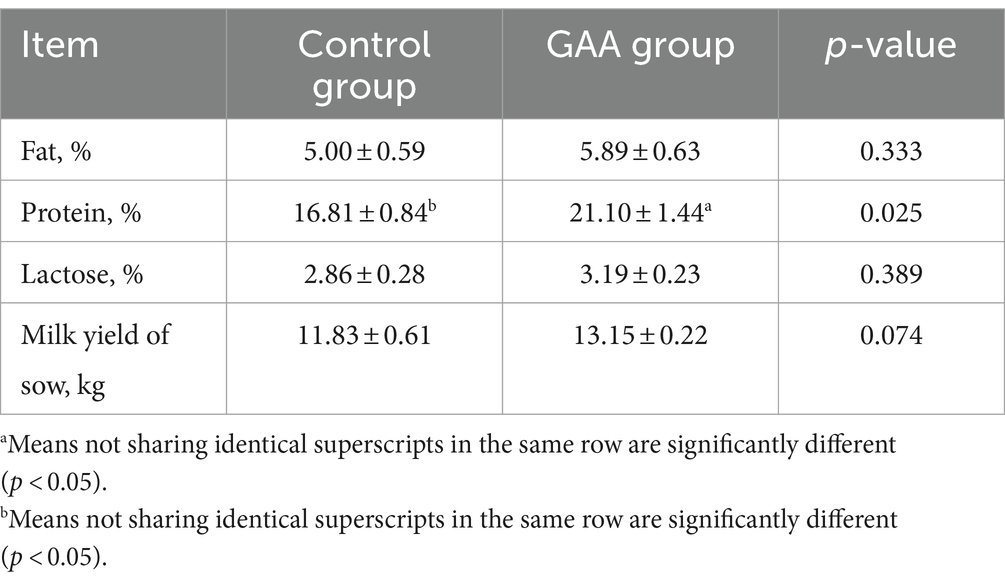
La suplementación dietética con GAA aumentó la inmunoglobulina A (IgA) en el calostro de las cerdas (p < 0,05; Tabla 6), sin efecto significativo sobre los niveles de IgG e IgM en calostro (p > 0,05).
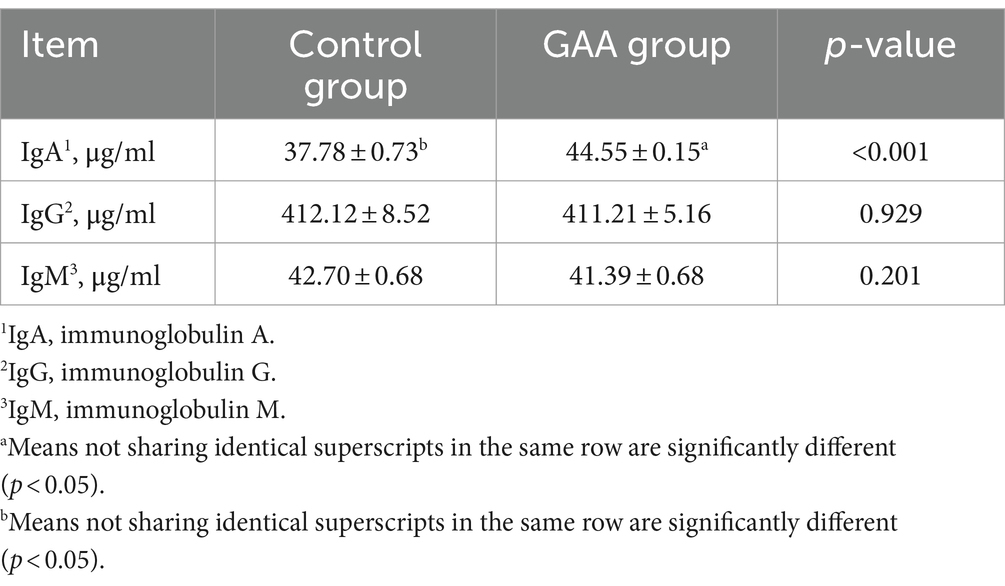
La suplementación dietética con GAA tendió a aumentar el contenido de prolina (Pro) en el calostro de las cerdas (p = 0,10; Tabla 7). La suplementación dietética con GAA no tuvo efectos significativos sobre los contenidos de ácido aspártico (Asp), ácido glutámico (Glu), serina (Ser), glicina (Gly), histidina (His), arginina (Arg), treonina (Thr), alanina (Ala), tirosina (Tyr), valina (Val), metionina (Met), isoleucina (Iso), leucina (Leu), fenilalanina (Phe) y lisina (Lys) en el calostro de las cerdas (p > 0,05).
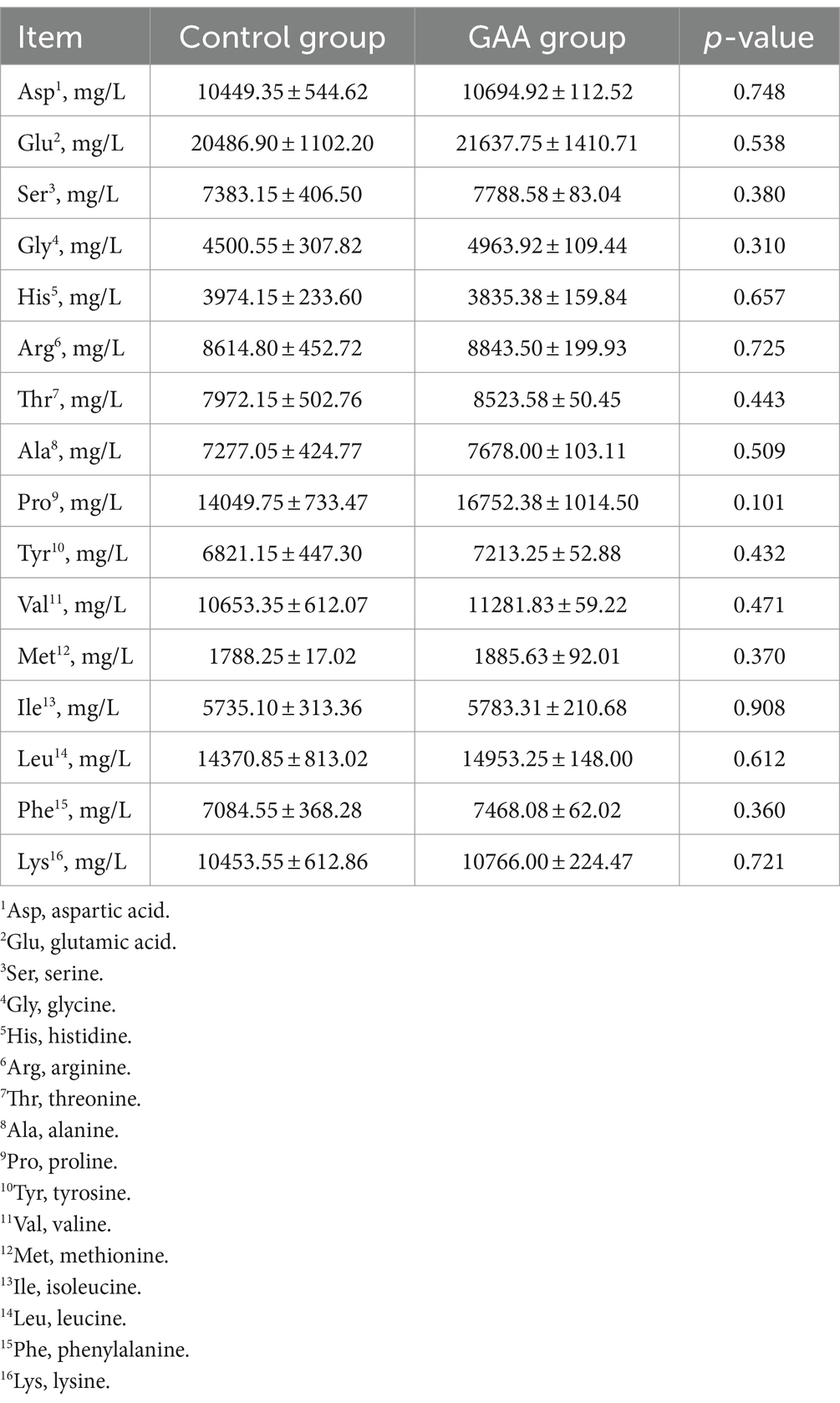
3.5 Metabolitos séricos, NO y rendimiento inmunitario
Como se muestra en la Tabla 8, la suplementación dietética con GAA tendió a disminuir los niveles séricos de TG en la sangre del cordón umbilical de las cerdas (p = 0,072). Sin embargo, la suplementación dietética con GAA no afectó significativamente los niveles de HDL-C, LDL-C, GLU y TC en la sangre del cordón umbilical de las cerdas (p > 0,05).
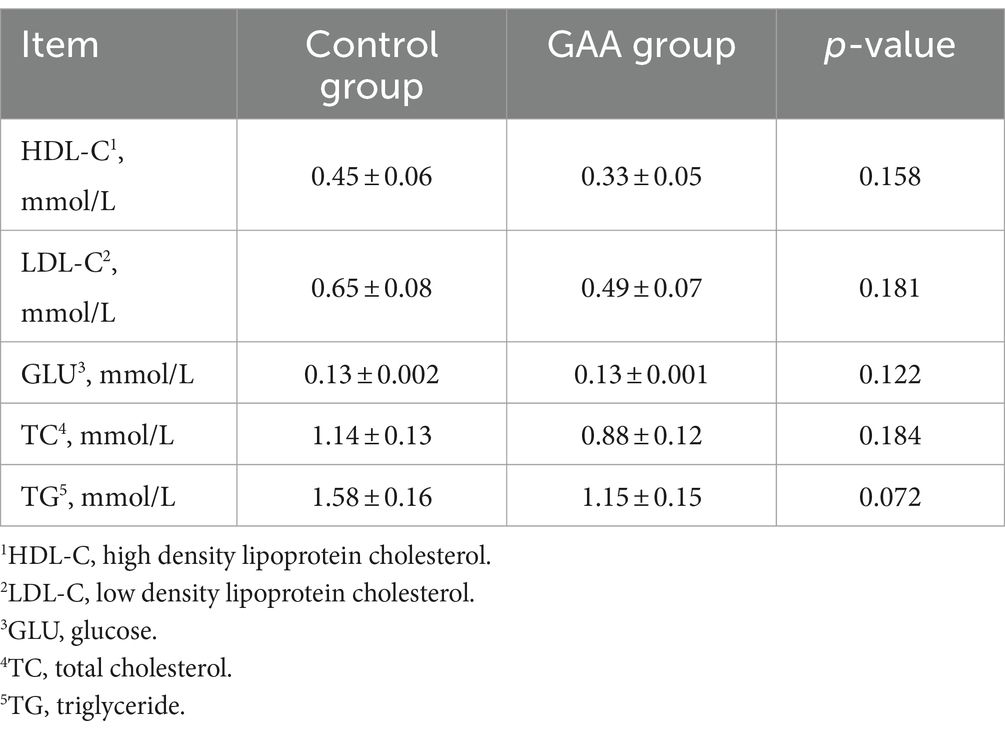
La suplementación dietética con GAA aumentó significativamente el nivel de eNOS en la sangre del cordón umbilical de las cerdas (p < 0,05; Tabla 9). Sin embargo, los niveles de NO, IL-6 e IL-10 en la sangre del cordón umbilical de las cerdas no se alteraron significativamente con la suplementación dietética con GAA (p > 0,05).
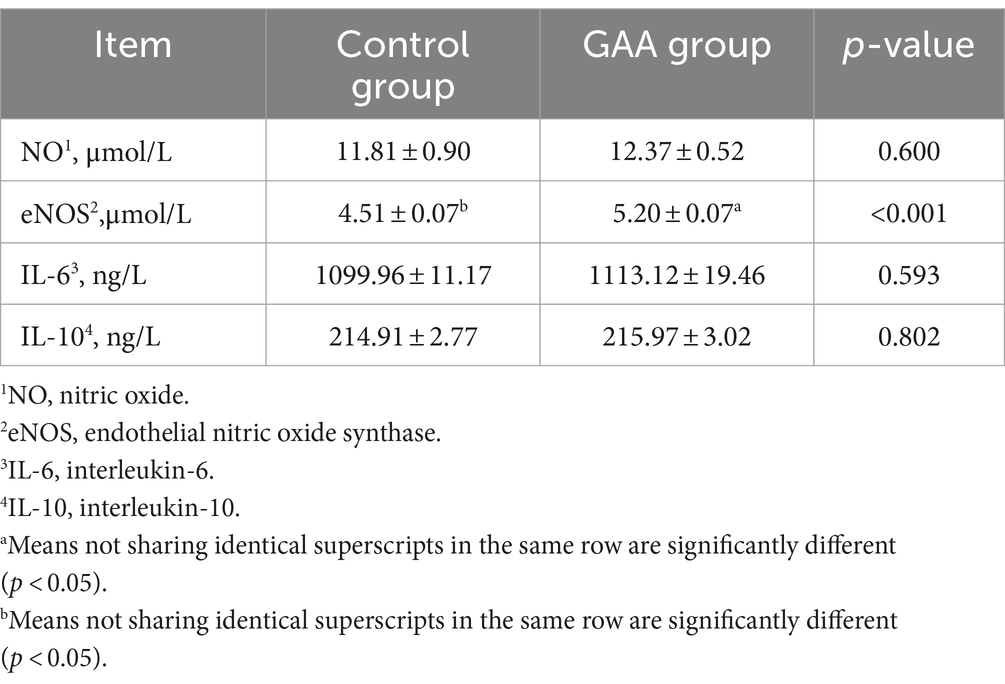
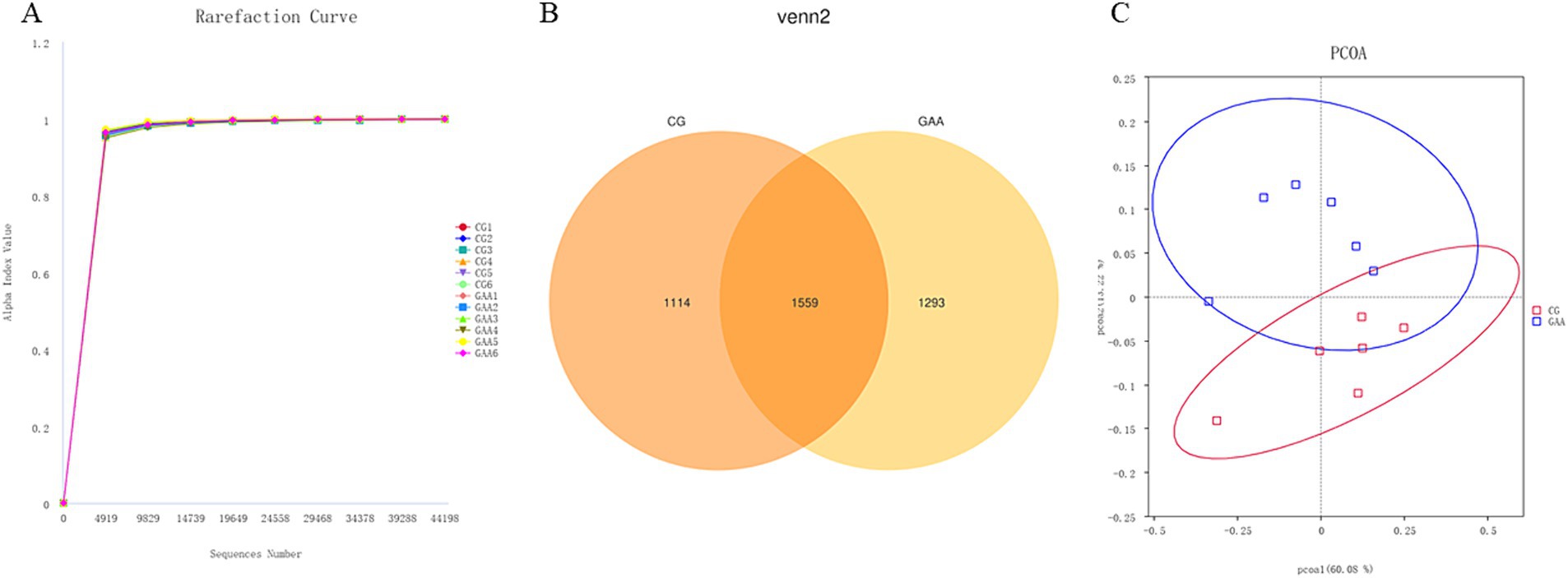
3.6 Diversidad de la flora intestinal
Con base en la Figura 1, las curvas de dilución de las muestras fecales de ambos grupos de cerdas mostraron una tendencia ascendente constante, lo que indica una extracción completa de la muestra. Según el diagrama de Venn, el grupo de control (CG) exhibió 1.114 secuencias únicas, mientras que el grupo de ácido acético guanidina (GAA) tuvo 1.293 secuencias únicas, con 1.559 secuencias compartidas entre los dos grupos. El análisis del PCoA reveló diferencias significativas en la composición y la estructura de la microbiota intestinal entre los dos grupos.
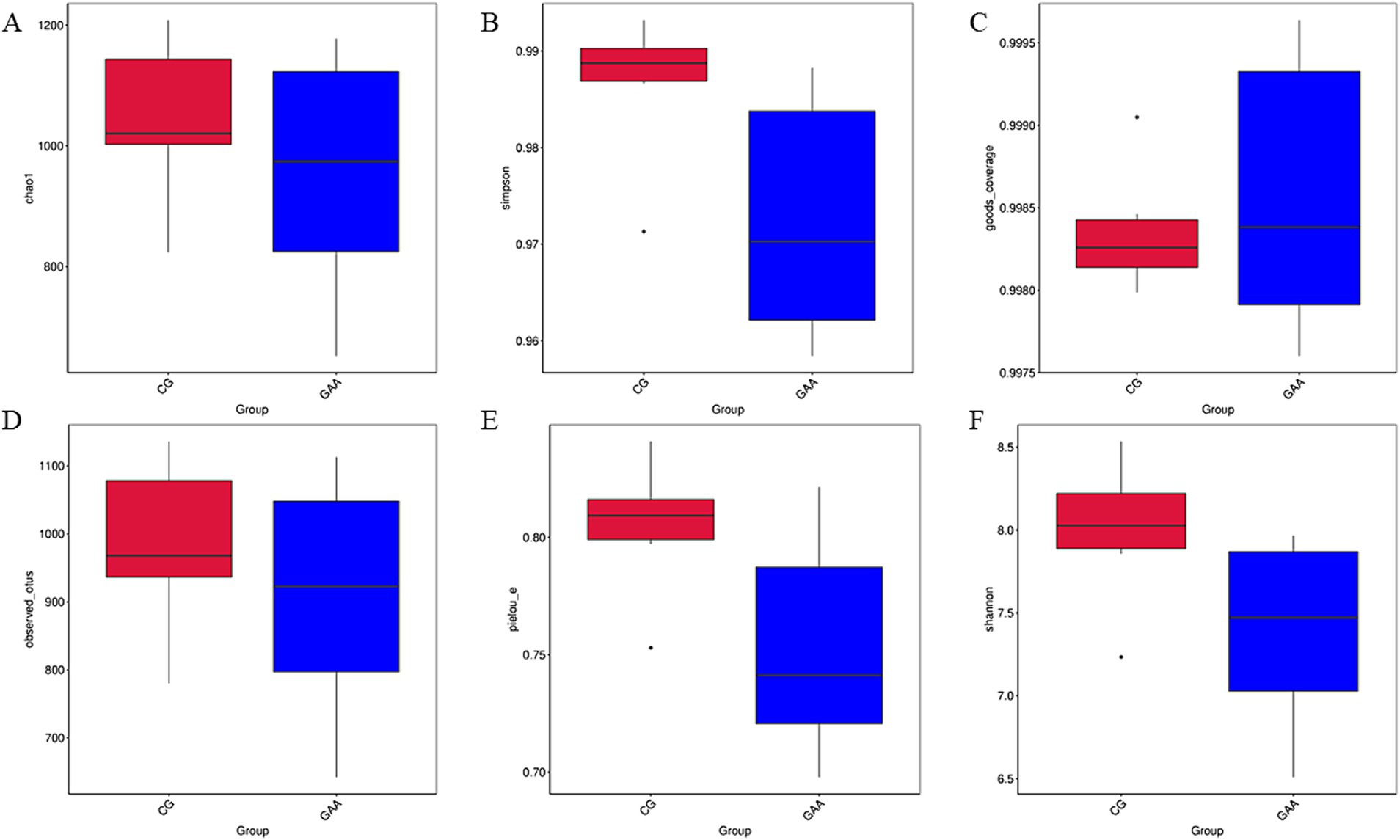
Como se muestra en la Figura 2, no hubo diferencias significativas (p > 0,05) en los índices de α diversidad (chao1, Simpson, good_coverage, observed_otus, pielou_e y Shannon) de la microbiota intestinal entre los dos grupos.
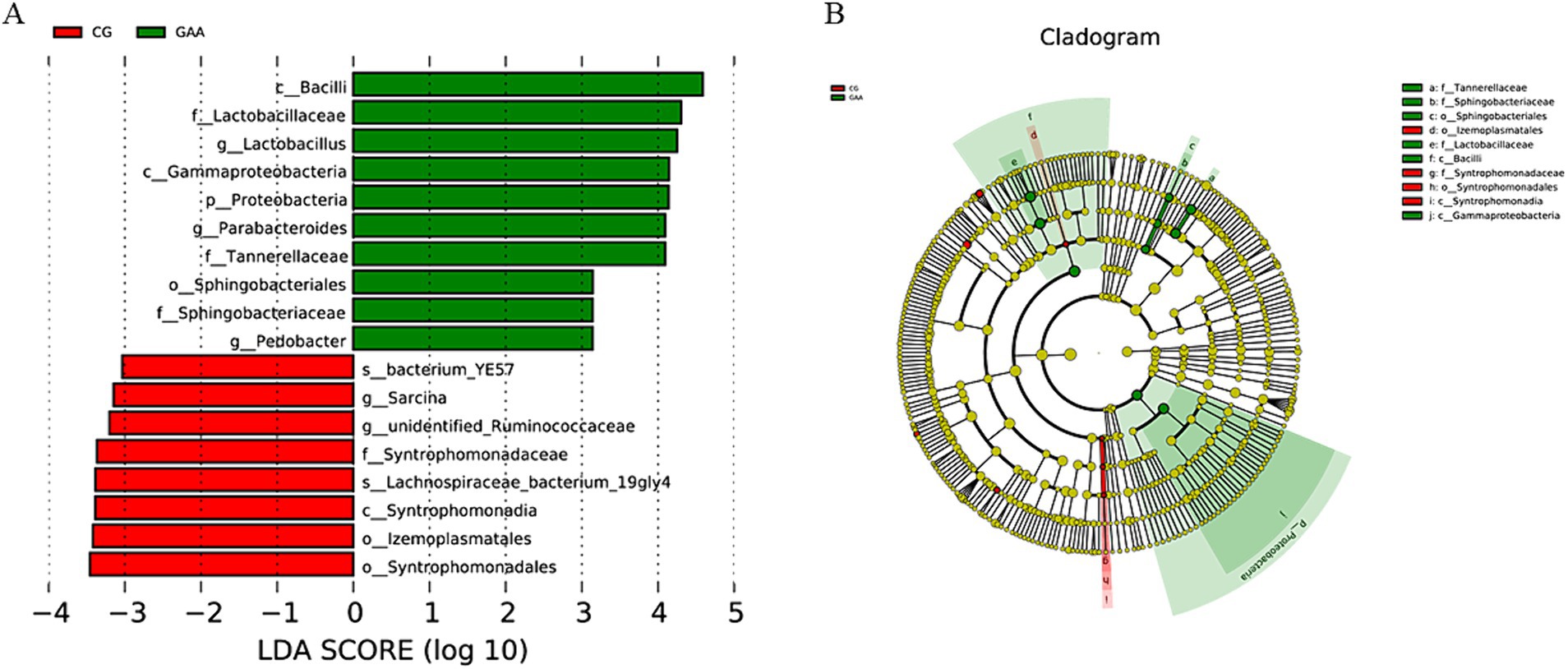
El análisis LEfSe de la Figura 3, LDA = 3, identificó a lactobacillus, Pedobacter y Parabacteroides como taxones significativamente enriquecidos en la microbiota intestinal de cerdas suplementadas con ácido guanidina acético.
Además, el análisis de Simper de la Figura 4 identificó además a lactobacillus y Terrisporobacter como taxones dominantes en la microbiota intestinal del grupo del ácido guanidina acético.
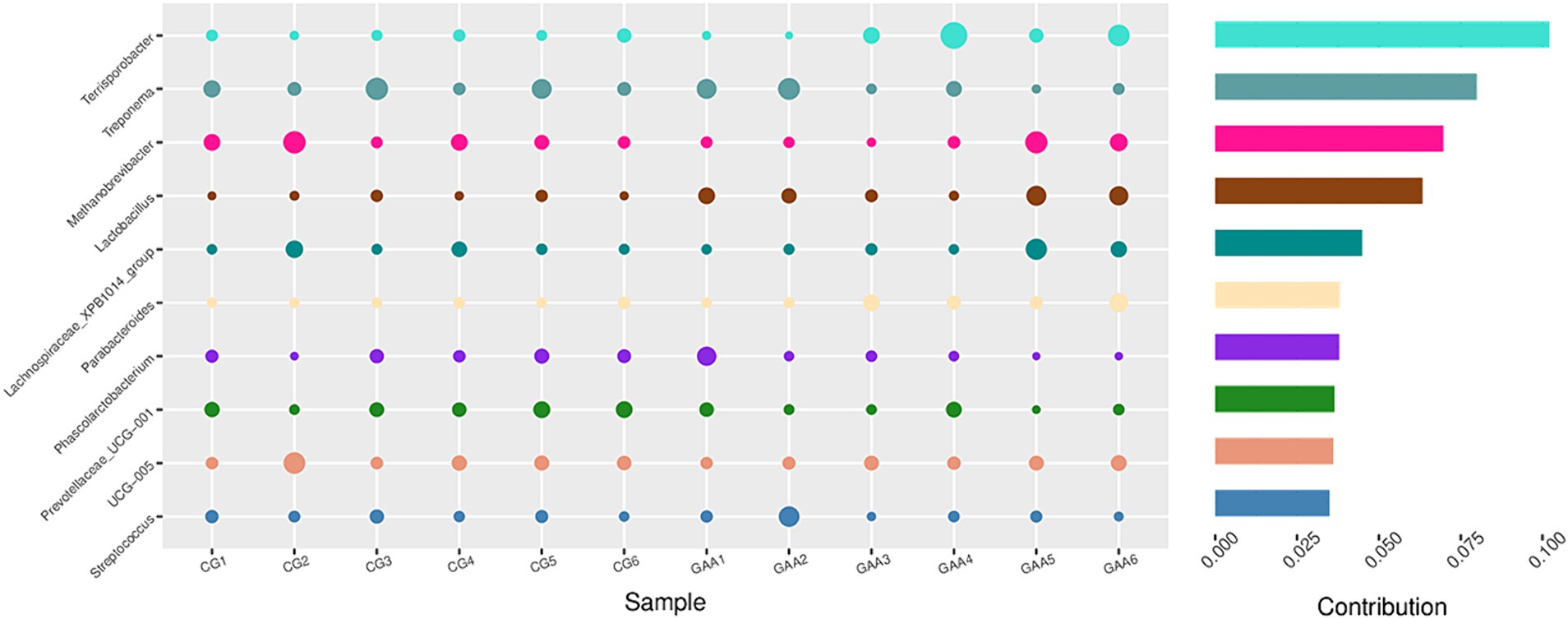
3.7 Rendimiento de crecimiento y salud de los lechones lactantes
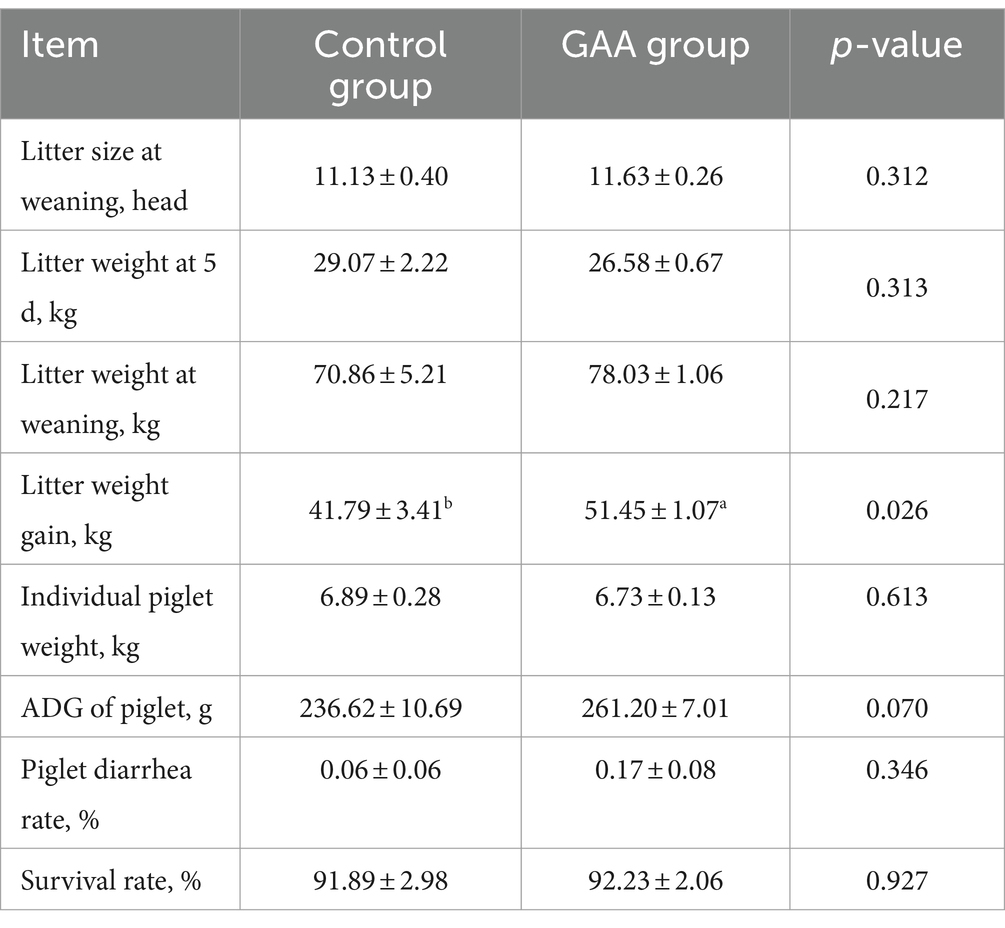
Como se muestra en la Tabla 10, la suplementación dietética con GAA resultó en un aumento significativo en la ganancia de peso de la camada de los lechones descendientes (p < 0,05). Además, la adición de GAA a la dieta mostró una tendencia a aumentar la ganancia media diaria (GMD) de los lechones (p < 0 0,05). Sin embargo, la suplementación con GAA no afectó significativamente el tamaño de la camada al destete, el peso de la camada a los 5 días, el peso de la camada al destete, la tasa de diarrea de los lechones o la supervivencia de los lechones lactantes.
4 Discusión
Actualmente, la investigación sobre la aplicación del ácido guanidínico acético (GAA) en cerdas es limitada. Las cerdas gestantes y lactantes tienen necesidades energéticas máximas, y la suplementación con GAA durante este período puede mejorar la eficiencia energética, aumentando así la productividad de las cerdas y las crías.
Las investigaciones indican que la suplementación con GAA durante la gestación por sí sola puede aumentar el número de lechones vivos por cerda. El inicio de la suplementación con GAA al final de la gestación aumenta el peso medio al nacer de los lechones, mientras que la suplementación durante la lactancia aumenta las concentraciones de proteínas y aminoácidos en el calostro de las cerdas (20). La suplementación con creatina en la última semana de gestación mejora la vaina de mielina de los lechones de bajo peso al nacer y aumenta la tasa de supervivencia de los lechones más débiles, aunque no tiene ningún impacto en la duración del trabajo de parto, los intervalos entre partos o las tasas de mortinatos (21). La adición de un 0,1% de GAA a las dietas de primerizas aumenta el número de lechones vivos producidos, con una variación mínima en el peso al nacer por camada. Además, el GAA mejora la ganancia media diaria (GMD), el peso al destete y la producción diaria de leche de los lechones. Durante la lactancia, el GAA aumenta el contenido de aminoácidos en la leche a los 7 días, en particular lisina, metionina, arginina, valina y glutamina (15).
Nuestros resultados indican que la suplementación con GAA aumenta significativamente el número de lechones vivos y sanos, pero no afecta a la duración de los partos, los intervalos entre las camadas, las tasas de mortinatos, la ingesta de alimento o la pérdida de grasa dorsal. Este hallazgo se alinea con estudios previos, que sugieren que los efectos promotores del crecimiento de GAA en animales monogástricos están relacionados principalmente con su papel en el anabolismo de proteínas tisulares y la mejora del metabolismo energético. La síntesis de GAA implica su producción en el hígado y su posterior metilación en los riñones. La síntesis de GAA requiere la participación de dos aminoácidos, la arginina y la glicina. La arginina transfiere su grupo amidino al grupo amino de la glicina para producir ornitina y GAA, catalizada por L-arginina: glicina amidinotransferasa (AGAT). A continuación, el acetato de guanidinocetato se metila utilizando un grupo metilo de la S-adenosilmetionina (SAM), que se sintetiza a partir de la metionina. Esta reacción, catalizada por la N-metiltransferasa (GAMT) del guanidinoacetato produce creatina y S-adenosilhomocisteína (SAH) (2). En consecuencia, la suplementación dietética con GAA reduce la descomposición de la arginina y la glicina en el cuerpo, mejorando así su eficacia en la síntesis de proteínas y promoviendo el crecimiento (22, 23). Además, el GAA puede estar mediado por proteínas transportadoras como el transportador de creatina (CRT/SLC6A8), el transportador de taurina (TauT/SLC6A6) y el transportador de ácido γ-aminobutírico (SLC6A13), y se transporta a las células diana a través de la difusión pasiva a través de la membrana plasmática (24), mejorando así el rendimiento reproductivo de las cerdas. Además, las funciones reguladoras fisiológicas del GAA pueden incluir la estimulación de la secreción del factor de crecimiento similar a la insulina 1 (IGF-1), mejorando así el rendimiento del crecimiento (25). Además, la suplementación dietética con GAA podría aumentar la secreción de ácido gamma-aminobutírico (GABA), que promueve la secreción de la hormona liberadora de la hormona del crecimiento por el hipotálamo y, posteriormente, aumenta la secreción de la hormona del crecimiento por la adenohipófisis, fomentando el crecimiento animal (26). Sin embargo, nuestro estudio actual sobre GAA en cerdas tiene limitaciones en cuanto a los mecanismos involucrados, lo que justifica una mayor investigación y validación.
Junto con los hallazgos de óxido nítrico de nuestro experimento, los niveles de óxido nítrico sintasa y óxido nítrico en la sangre del cordón umbilical de las cerdas aumentaron en 0,7 μmol/L y 0,5 μmol/L en el grupo GAA, respectivamente. Los niveles óptimos de óxido nítrico estimulan la proliferación de vasos sanguíneos en la placenta y el cordón umbilical, mejorando el flujo de nutrientes de la madre al lechón y aumentando el peso de los lechones al nacer. Esto subraya por qué el ácido guanidina acético puede aumentar el número de lechones sanos en las cerdas, lo que resulta en un aumento de peso promedio de 130 g para lechones vivos y de 90 g para lechones sanos.
Durante la gestación, la deficiencia de arginina puede mitigarse eficazmente mediante la suplementación con ácido guanidina acético (GAA). Mateo et al. (27) observaron que la suplementación diaria de arginina al 1% en cerdas lactantes en el 7º día de lactancia aumentaba las concentraciones plasmáticas de arginina y los niveles de insulina, mejorando así el metabolismo de la sustancia. Esta suplementación también elevó el contenido de nutrientes en la leche, con concentraciones más altas de la mayoría de los aminoácidos en comparación con el grupo de control. Además, la arginina promueve el desarrollo de la glándula mamaria, mejora la circulación sanguínea, afecta el transporte de nutrientes y potencialmente mejora el rendimiento de la lactancia al aumentar la producción y la calidad de la leche (28).
Además, nuestro estudio demostró que la suplementación con GAA aumentó la proteína de la leche, el nitrógeno ureico y los sólidos totales en el calostro de las cerdas. Esto sugiere que el GAA puede influir en la composición del calostro a través de su impacto en los niveles de arginina. En este experimento no se llevaron a cabo más investigaciones sobre la composición normal de la leche en las cerdas, lo que pone de manifiesto la necesidad de futuras investigaciones.
La microbiota intestinal desempeña un papel crucial en la regulación del desarrollo fisiológico y el estado de salud de los cerdos, así como en la prevención de la colonización de patógenos (29). Si bien la investigación sobre el efecto del GAA en la flora intestinal es escasa, los estudios sobre la arginina han demostrado que la utilización de bacterias intestinales puede conducir a la liberación de poliaminas en el intestino (30). La arginina también regula la utilización de aminoácidos en cultivos de bacterias intestinales obtenidos de intestinos de cerdo (31, 32).
En nuestro experimento, el grupo GAA no afectó la diversidad α de la flora intestinal de las cerdas, pero sí mostró diferencias en la diversidad de β. Los análisis de LEfSe y Simper identificaron a Lactobacillus y Terrisporobacter como los principales géneros afectados. Lactobacillus, un probiótico, inhibe los microorganismos dañinos (33), altera el metabolismo microbiano (actividad enzimática), mejora la absorción de nutrientes (por ejemplo, proteínas, monosacáridos) (34) y estimula las respuestas inmunes (35). El género Terrisporobacter incluye bacterias beneficiosas como Bacillus subtilis, Bacillus fungoides y Bacillus polymyxis, que mejoran el rendimiento de la producción animal. Estos hallazgos sugieren que el GAA altera las bacterias intestinales dominantes, mejorando potencialmente el metabolismo y la absorción de nutrientes, aumentando así la proteína de la leche, los sólidos totales y el nitrógeno ureico en las cerdas.
El GAA entra en la leche de cerda a través del torrente sanguíneo y aumenta eficazmente el contenido de creatina y fosfato de creatina en el tejido muscular. Esto reduce los suministros de energía de carbohidratos, grasas y proteínas, al tiempo que mejora la hidratación para inhibir la descomposición de proteínas y promover la síntesis de proteínas y glucógeno, acelerando el crecimiento animal (36). Además, el GAA influye en el hipotálamo para que secrete la hormona liberadora de la hormona del crecimiento, lo que afecta la secreción de ácido gamma-aminobutírico y, posteriormente, estimula la secreción de la hormona del crecimiento hipofisaria (37). En combinación con el aumento de los niveles de proteína láctea en la leche de cerda, esto contribuye a aumentar en cierta medida la ganancia media diaria (GMD) en los lechones de descendencia.
5 Conclusión
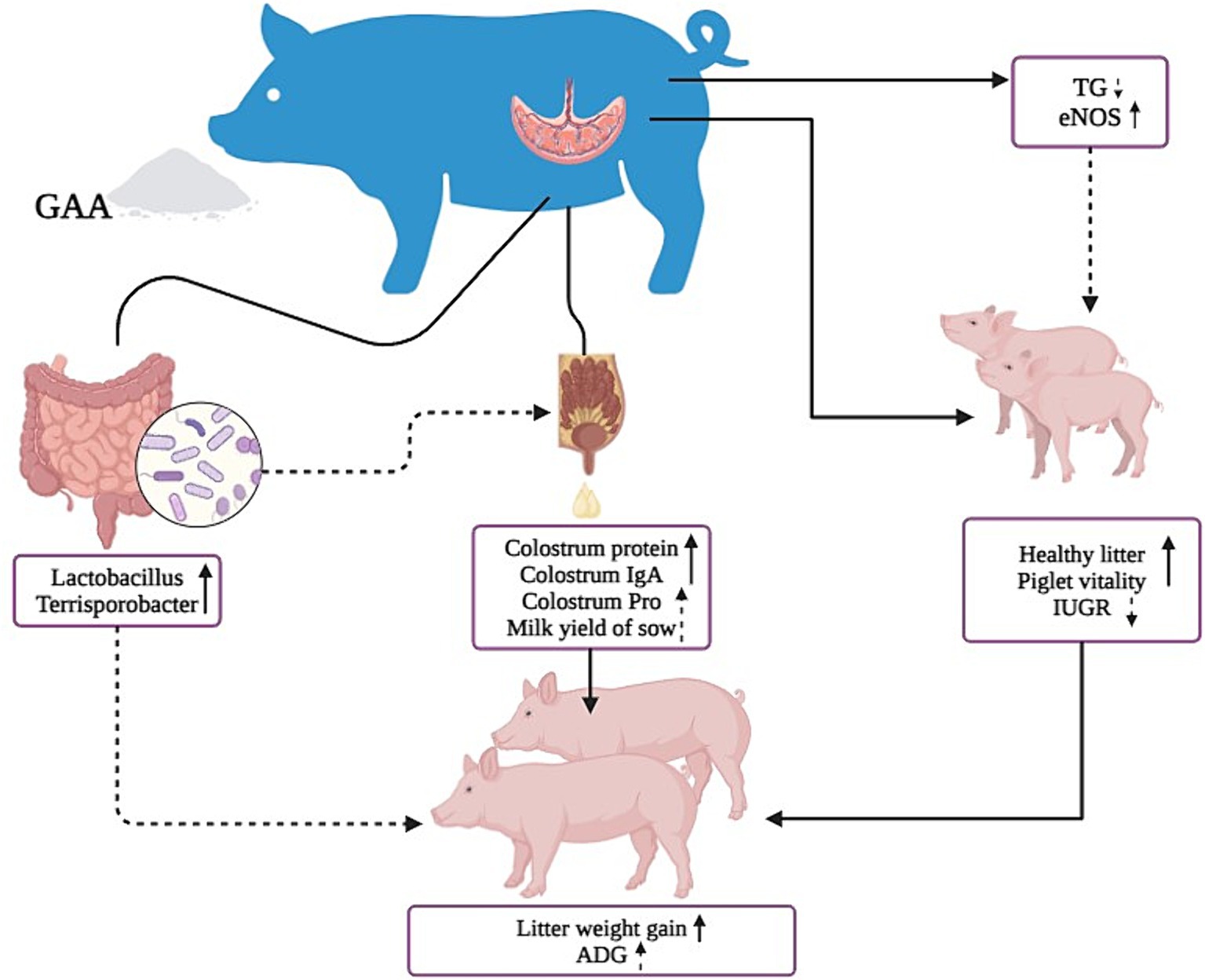
El GAA puede aumentar los niveles de óxido nítrico sintasa en la sangre del cordón umbilical de las cerdas, promoviendo el desarrollo de vasos sanguíneos placentarios y el crecimiento de lechones. Aumenta el número de lechones sanos y su vitalidad, al tiempo que aumenta la abundancia de bacterias beneficiosas como Lactobacillus, Parabacteroides y Pedobacter en el intestino. Estos efectos conducen a niveles más altos de proteína en el calostro y a un aumento del volumen total de la lactancia, mejorando así el rendimiento del crecimiento de los lechones descendientes (Figura 5).
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán puestos a disposición por los autores, sin reservas indebidas.
Declaración ética
El estudio en animales fue aprobado por Ética en el Uso de Animales (CEUA) de Anyou Biological Technology Group Co., LTD. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
GC: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Software, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. SX: Conceptualización, Administración de proyectos, Recursos, Validación, Redacción – revisión y edición. CL: Conceptualización, Metodología, Recursos, Redacción – revisión y edición. JL: Administración de proyectos, Recursos, Redacción – revisión y edición. IH: Redacción – revisión y edición, análisis formal.
Financiación
El (los) autor(es) declara(n) que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. El Proyecto (Innovación) del Plan de Talentos Científicos y Tecnológicos de la Ciudad de Taicang de China (TCRC2022067) proporcionó apoyo financiero para el presente estudio.
Reconocimientos
Los autores desean agradecer a Shuaipeng Guo, Bo Zhao, Fang Zhang y Lili Zhang del Instituto de Investigación de Anyou Biotechnology Group Co., Ltd. por su asistencia y apoyo en la realización de experimentos con animales.
Conflicto de intereses
GC, SX, CL, JL e IH, fueron empleados de la empresa Anyou Biotechnology Group Co., Ltd.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Bloch, K, y Schoenheimer, R. Los precursores biológicos de la creatina. J Biol Chem. (1941) 138:167–94. doi: 10.1016/S0021-9258(18)51422-8
2. Wyss, M, y Kaddurah-Daouk, R. Metabolismo de la creatina y la creatinina. Physiol Rev. (2000) 80:1107–213. doi: 10.1152/physrev.2000.80.3.1107
3. Fisher, H, Salander, RC y Taylor, MW. El crecimiento y la biosíntesis de creatina en el pollito se ven afectados por las deficiencias de aminoácidos de la caseína. J Nutr. (1956) 58:459–70. doi: 10.1093/jn/58.4.459
4. Fisher, H, Salander, RC y Taylor, MW. Influencia de la biosíntesis de creatina en el requerimiento de arginina del pollito. J Nutr. (1956) 59:491–9. doi: 10.1093/jn/59.4.491
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Lemme, A, Ringel, J, Sterk, A, y Young, JF. (2007). El ácido guanidino acético suplementario afecta el metabolismo energético de los pollos de engorde. 16º Simposio Europeo sobre Nutrición Avícola; del 26 al 30 de agosto; Estrasburgo, Francia.
6. Tossenberger, J, Rademacher, M, Németh, K, Halas, V y Lemme, A. Digestibilidad y metabolismo del ácido guanidino acético dietético alimentado con pollos de engorde. Poult Sci. (2016) 95:2058–67. doi: 10.3382/ps/pew083
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. McBreairty, LE, Robinson, JL, Furlong, KR, Brunton, JA y Bertolo, RF. El acetato de guanidinocetato es más efectivo que la creatina para mejorar las reservas de creatina en los tejidos y, en consecuencia, limitar la disponibilidad de metionina en cerdos miniatura de Yucatán. PLoS Uno. (2015) 10:e0131563. doi: 10.1371/journal.pone.0131563
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Jayaraman, B, La, KV, La, H, Doan, V, Carpena, EM, Rademacher, M, et al. Suplementación con ácido guanidinoacético en dietas de cerdos: efectos sobre el rendimiento, las características de la canal y la calidad de la carne. J Anim Sci. (2018) 96:2332–41. doi: 10.1093/jas/sky137
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Él, DT, Gai, XR, Yang, LB, Li, JT, Lai, WQ, Sun, XL, et al. Efectos del ácido guanidinoacético sobre el rendimiento del crecimiento, la creatina y el metabolismo energético, y las características de la canal en cerdos de engorde y engorde. J Anim Sci. (2018) 96:3264–73. doi: 10.1093/jas/sky186
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Valini, GAC, Duarte, MS, Rodrigues, GA, Veroneze, R, Saraiva, A, Hausman, G, et al. Suplementación con ácido guanidinoacético sobre el rendimiento del crecimiento y los mecanismos moleculares de la ganancia de masa magra en cerdos de destete. Cienc Rural. (2020) 50:e20190948. doi: 10.1590/0103-8478cr20190948
11. Lu, Y, Zou, T, Wang, Z, Yang, J, Li, L, Guo, X, et al. El ácido guanidinoacético dietético mejora el rendimiento del crecimiento y el desarrollo del músculo esquelético de los cerdos de engorde mediante el cambio en la expresión génica miogénica y las características de la miofibra. J Anim Physiol Anim Nutr. (2020) 104:1875–83. doi: 10.1111/jpn.13351
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Cui, Y, Tian, Z, Yu, M, Deng, D, Lu, H, Song, M, et al. La suplementación con ácido guanidina acético alteró los perfiles de aminoácidos libres plasmáticos y tisulares en cerdos de engorde. Gestión de la Salud Porcina. (2022) 8:24. doi: 10.1186/s40813-022-00269-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Cui, Y, Yu, M, Li, Z, Song, M, Tian, Z, Deng, D, et al. El ácido guanidina acético altera los perfiles de aminoácidos unidos a los tejidos y el estado oxidativo en los cerdos de engorde. Animales. (2023) 13:1626. doi: 10.3390/ani13101626
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Mendonça, IW, Watanabe, PH, Silva, BAN, Boiago, MM, Panisson, JC, Andrade, TS, et al. Suplementación dietética de ácido guanidinoacético para cerdas y sus progenies: rendimiento, parámetros sanguíneos y viabilidad económica en la fase de destete. Livest Sci. (2019) 227:105–10. doi: 10.1016/j.livsci.2019.07.011
15. Panisson, J, Silva, BAN, Oliveira, SG, Maiorka, A, y Azevedo, AM. (2019). Ácido guanidinoacético: ¿Cómo pueden beneficiarse las cerdas de este novedoso aditivo para piensos? Editorial Académica de Wageningen. 8–22. doi: 10.3920/978-90-8686-891-9_31
16. Baxter, EM, Jarvis, S, Palarea-Albaladejo, J, y Edwards, SA. ¿El sexo débil? La propensión a la mortalidad de lechones sesgados por machos. PLoS Uno. (2012) 7:E30318. doi: 10.1371/journal.pone.0030318
17. Magallón, E, García, A, Bautista, R, Sánchez, BA, Almenara, SD, Prieto, P, et al. Prácticas de cría y manejo en parideras I: Parideras. Zaragoza: Grupo Asís Biomedia (2014).
18. Bolduan, G, Schnabel, E, y Voss, S. Postpartale N-Bilanzen laktierender Sauen. Arch Anim Nutr. (1990) 40:923–32. doi: 10.1080/17450399009428443
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Nascimento, TMT, Mansano, CFM, Peres, H, Rodrigues, FHF, Khan, KU, Romaneli, RS, et al. Determinación del perfil óptimo de aminoácidos esenciales en la dieta para la fase de crecimiento de la tilapia del Nilo mediante el método de deleción. Acuicultura. (2020) 523:735204. doi: 10.1016/j.aquaculture.2020.735204
20. Mendoza, SM, Richter, JF y Htoo, JK. Efecto del ácido guanidinoacético, precursor de la creatina, en cerdas gestantes y lactantes. Arch Latinoam prod anim. (2022) 30:95–101. doi: 10.53588/alpa.300509
21. Vallet, JL, Miles, JR y Rempel, LA. Efecto de la suplementación con creatina durante la última semana de gestación sobre los intervalos entre partos, muerte fetal y mortalidad antes del destete en cerdos. J Anim Sci. (2013) 91:2122–32. doi: 10.2527/jas.2012-5610
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Panadero, DH. Avances en la nutrición proteico-aminoacídica de las aves. Aminoácidos. (2009) 37:29–41. doi: 10.1007/s00726-008-0198-3
23. Kodambashi Emami, N, Golian, A, Rhoads, DD y Danesh, MM. Efectos interactivos de la temperatura y la suplementación dietética de arginina o ácido guanidinoacético en las respuestas nutricionales y fisiológicas en pollos de engorde machos. Hno. Poult Sci. (2017) 58:87–94. doi: 10.1080/00071668.2016.1257779
24. Tachikawa, M, Kasai, Y, Yokoyama, R, Fujinawa, J, Ganapathy, V, Terasaki, T, et al. Transporte de la barrera hematoencefálica y distribución cerebral del acetato de guanidinocetato en ratas: implicación de los transportadores de creatina y taurina. J Neuroquímica. (2009) 111:499–509. doi: 10.1111/j.1471-4159.2009.06332.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Adriano, E, Garbati, P, Salis, A, Damonte, G, Millo, E y Balestrino, M. Las sales de creatina proporcionan neuroprotección incluso después de un deterioro parcial del transportador de creatina. Neurociencia. (2017) 340:299–307. doi: 10.1016/j.neuroscience.2016.02.038
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Ostojic, SM. Ácido guanidinoacético como agente que mejora el rendimiento. Aminoácidos. (2016) 48:1867–75. doi: 10.1007/s00726-015-2106-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Mateo, RD, Wu, G, Moon, HK, Carroll, JA y Kim, SW. Efectos de la suplementación dietética con arginina durante la gestación y la lactancia en el rendimiento de cerdas primíparas lactantes y lechones lactantes. J Anim Sci. (2008) 86:827–35. doi: 10.2527/jas.2007-0371
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Gao, AG, Wen, XL, Guo, CY, Wang, L, Ban, WJ, Yang, XF, et al. Efecto de la relación arginina/lisina en la dieta en la lactancia sobre los índices bioquímicos y el rendimiento de las cerdas lactantes. J Anim Sci. (2020) 98:skaa261. doi: 10.1093/jas/skaa261
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Schokker, D, Zhang, J, Zhang, LL, Vastenhouw, SA, Heilig, HG, Smidt, H, et al. Las variaciones ambientales en los primeros años de vida afectan a la microbiota intestinal y al desarrollo inmunitario de los lechones recién nacidos. PLoS Uno. (2014) 9:E100040. doi: 10.1371/journal.pone.0100040
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Matsumoto, M, Kitada, Y y Naito, Y. La función endotelial mejora al inducir la producción de poliaminas microbianas en el intestino: un ensayo aleatorizado controlado con placebo. Nutrientes. (2019) 11:1188. doi: 10.3390/nu11051188
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Dai, ZL, Li, XL, Peng, BX, Zhang, J, Wu, GY y Zhu, WY. Papel regulador de la l-arginina en la utilización de aminoácidos por las bacterias del intestino delgado de cerdos. Aminoácidos. (2012) 43:233–44. doi: 10.1007/s00726-011-1067-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Ren, WK, Duan, JL, Yin, J, Liu, G, Cao, Z, Xiong, X, et al. La suplementación dietética con L-glutamina modula la comunidad microbiana y activa la inmunidad innata en el intestino del ratón. Aminoácidos. (2014) 46:2403–13. doi: 10.1007/s00726-014-1793-0
33. Muralidhara, KS, Sheggeby, GG, Elliker, PR, Inglaterra, DC, y Sandine, WE. Efecto de la alimentación con lactobacilos sobre la flora coliforme y Lactobacillus del tejido intestinal y las heces de lechones. J Food Prot. (1977) 40:288–95. doi: 10.4315/0362-028X-40.5.288
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Li, RL, Liu, JW, Liu, YH, Cao, LT, Qiu, WF y Qin, M. Efectos probióticos de Bacillus subtilis en el rendimiento del crecimiento y el equilibrio microecológico intestinal de cerdos de crecimiento a engorde. J Bioquímica de los Alimentos. (2023) 2023:1–16. doi: 10.1155/2023/7150917
35. Lan, Q. (2008). El estudio de la distribución de la flora principal en el tracto intestinal de los cerdos bebés y la influencia de los probióticos en el crecimiento y desarrollo y la función inmune de los cerdos bebés [tesis]. Chengdu: Universidad Agrícola de Sichuan.
36. Wang, YQ, Liu, Q, Jiang, FB, Yuan, QQ, Rui, Y y Zhuang, S. Efectos del ácido guanidinoacético sobre el rendimiento y la capacidad antioxidante en patos de Cherry Valley. J Nanjing Agric Univ. (2016) 39:269–74. doi: 10.7685/jnau.201505024
37. Michiels, J, Maertens, L, Buyse, J, Lemme, A, Rademacher, M, Dierick, NA, et al. Suplementación con ácido guanidinoacético en dietas de pollos de engorde: efectos sobre el rendimiento, las características de la canal, la calidad de la carne y el metabolismo energético. Poult Sci. (2012) 91:402–12. doi: 10.3382/ps.2011-01585
Palabras clave: cerdo, ácido guanidina acético, rendimiento inmunológico, eficiencia reproductiva, aminoácido calostro, óxido nítrico
Cita: Cong G, Xia S, Liu C, Li J y Hung I (2024) Efectos de la suplementación con ácido acético con guanidina desde la gestación hasta la lactancia en el rendimiento reproductivo, la calidad del calostro, la bioquímica sanguínea y la diversidad de la microflora intestinal de las cerdas. Frente. Vet. Sci. 11:1476328. doi: 10.3389/fvets.2024.1476328
Editado por:
Jie Yu, Universidad Agrícola de Sichuan, China
Revisado por:
Cesar Augusto Pospissil Garbossa, Universidad de São Paulo, Brasil
Zhiyong Fan, Universidad Agrícola de Hunan, China
Derechos de autor © 2024 Cong, Xia, Liu, Li y Hung. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Chunxue Liu, cx_liu@anschina.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía