
Efectos de la suplementación con extracto de semilla de uva en corderos

Efectos de la suplementación con extracto de semilla de uva sobre el rendimiento del crecimiento, la digestión de nutrientes y la inmunidad de los corderos destetados
 Jian Ma
Jian Ma Tao LiLu LinYuezhang LuXi ChenSibing LiChunmei DuChen WeiFuquan Yin
Tao LiLu LinYuezhang LuXi ChenSibing LiChunmei DuChen WeiFuquan Yin Shangquan Gan*
Shangquan Gan*- Facultad de Ciencias Agrícolas Costeras, Universidad Oceánica de Guangdong, Zhanjiang, China
El extracto de semilla de uva (GSE) tiene una variedad de funciones biológicas. En la actualidad, ha habido poca información sobre la utilización de GSE como aditivo alimentario en corderos destetados. El objetivo de este experimento fue estudiar la influencia potencial de la suplementación dietética con GSE sobre el rendimiento del crecimiento, las características de fermentación ruminal, la digestibilidad aparente, los parámetros sanguíneos y la inmunidad en corderos destetados. En total, 30 corderos machos de oveja Hu con peso corporal (15,43 ± 0,49 kg) y edad (48 ± 2 kg) similares se dividieron aleatoriamente en dos tratamientos: control (CON, alimentados con ración basal) y GSE [alimentados con ración basal y GSE 0,6 g/d (composiciones principales: proantocianidina 50%, catequina 24%, ácido gálico 16% y epicatequina 6%) por cordero]. El experimento de alimentación duró 60 d. Los resultados mostraron que la suplementación con GSE aumentó significativamente (p = 0,008) la ganancia media diaria de corderos. En comparación con el grupo CON, las concentraciones ruminales de propionato y butirato aumentaron significativamente (p < 0,05) en el grupo GSE, mientras que el nitrógeno amoniacal disminuyó (p = 0,007). Además, la proteína cruda, la fibra detergente neutro y la digestibilidad del extracto etéreo del grupo GSE fueron mayores (p < 0,05) que las del grupo CON. Los contenidos séricos de glucosa, triglicéridos, inmunoglobulina G, glutatión peroxidasa y capacidad antioxidante total aumentaron significativamente (p < 0,05) en el grupo GSE en comparación con los del grupo CON. Sin embargo, se observó una tendencia opuesta de nitrógeno ureico, ácido graso no esterificado, interleucina-1β, factor de necrosis itumoral-α y malondialdehído entre los dos grupos. Además, la suplementación con GSE aumentó (p < 0,05) los Lactobacillus y disminuyó (p < 0,05) los recuentos de Escherichia coli y Salmonella en las heces de los corderos. En resumen, la suplementación con GSE puede mejorar el rendimiento del crecimiento, la digestión de nutrientes y la inmunidad de los corderos destetados.
1 Introducción
En los últimos años, la creciente demanda de consumo de carne ha estimulado en gran medida el desarrollo de la industria ganadera, particularmente en la cría de ovejas ovinas. El cordero desempeña un papel esencial en la seguridad alimentaria y nutricional mundial porque es una importante fuente de alimento con proteínas, grasas, vitaminas y elementos minerales de alta calidad para los seres humanos (1, 2). En algunas partes del mundo, debido a los limitados recursos de tierra y a la urgente demanda de conservación y restauración ecológica de las praderas, la producción ovina ha pasado gradualmente del régimen tradicional de pastoreo a la cría intensiva moderna a gran escala (3). Sin embargo, debido a la limitación de espacio y a los cambios en los piensos, el régimen de alimentación intensivo provoca algunos efectos adversos, como la disminución de la resistencia al estrés oxidativo. Además, el bajo rendimiento del crecimiento, así como la alta mortalidad y morbilidad de los corderos, restringen seriamente el desarrollo estable de la industria de cría de ovejas ovinas (4, 5). La etapa de cordero es un período vital que puede afectar la productividad futura de las ovejas adultas. El destete precoz puede disminuir el costo de alimentación y promover el desarrollo del tracto digestivo de los corderos hasta cierto punto, y mientras tanto, el destete temprano disminuye la duración del ciclo reproductivo en la producción ovina (6). Después del destete, se cambia el método de alimentación y el tipo de alimentación de los corderos. Debido al sistema inmunitario inmaduro de los corderos, el estrés del destete precoz tiene una influencia obviamente adversa en la tasa de crecimiento y la salud (6). Además, el tracto gastrointestinal de los corderos se encuentra en la fase de desarrollo y tiene baja resistencia a los cambios del entorno externo (7). Debido a esto, los corderos destetados son vulnerables a las bacterias dañinas que pueden provocar diarrea y mortalidad (5). Por lo tanto, aliviar el estrés del destete y mejorar el crecimiento saludable de los corderos tienen una importancia importante en la alimentación intensiva a gran escala de la producción de ovejas ovinas.
La utilización de extractos de plantas ha atraído una amplia atención en las industrias ganadera y avícola (8-12). Se ha demostrado que los extractos de plantas, como el polifenol, la saponina, la flavona, el esterol y los aceites esenciales, mejoran la digestión de nutrientes, la inmunidad y la capacidad antioxidante, lo que tiene efectos beneficiosos sobre la productividad y la salud de los animales (13-17). Como subproducto de la transformación de la uva, el extracto de semilla de uva (GSE) se extrae de la semilla de uva e incluye principalmente proantocianidina, galato de epicatequina, ácido gálico y epicatequina (18). Recientemente, el uso del GSE está cada vez más extendido, ya que tiene múltiples características que incluyen capacidades antiinflamatorias, antibacterianas, antioxidantes y antidiabéticas (19). Con la prohibición de los antibióticos y algunos aditivos farmacéuticos para piensos, los productos de semilla de uva se han utilizado en la producción animal. Un estudio previo encontró que la suplementación dietética con 1% de semilla de uva y extracto de harina de orujo de uva no tuvo una influencia obvia en la ingesta de materia seca (DMI) de las vacas lecheras, pero aumentó la producción de leche (20). En los corderos de engorde, la dieta alta en concentración suplementada con proantocianidinas de semilla de uva a un nivel de 20 mg/kg de peso corporal (PC) aumentó diariamente el IMD y la ganancia diaria promedio (GMD), además de mejorar la calidad de la carne (21). Debido a estar llenos de polifenoles, se ha demostrado que los productos de semilla de uva (el nivel suplementado fue de 50 mg / kg de peso corporal) mejoran las capacidades antioxidantes y antiinflamatorias de los terneros antes del destete, lo que fue beneficioso para atenuar el estrés oxidativo causado por el estrés térmico (22). Además, los resultados de un estudio in vitro mostraron que el GSE puede aliviar la reacción inflamatoria al reducir los niveles de expresión de interleucina-1β (IL-1β), IL-6 y factor de necrosis tumoral-α (TNF-α) (23). Una investigación reciente en terneros lecheros informó que la adición de GSE en la dieta a una dosis de 50 mg/kg de peso corporal promovió el crecimiento de los terneros lecheros afectados por estrés térmico al mejorar el metabolismo energético, el sistema endocrino y la fermentación intestinal (24).
El desarrollo saludable del tracto gastrointestinal desempeña un papel fundamental en el cambio de alimento de los rumiantes jóvenes de líquido a sólido, lo que tiene un efecto importante en la digestión de nutrientes y el rendimiento del crecimiento de los animales (7). Como órgano digestivo único de los rumiantes para la digestión de nutrientes y la absorción de ácidos grasos volátiles (AGV), la salud y el desarrollo del rumen están fuertemente asociados con la producción de AGV, especialmente el butirato y el propionato (25). Una investigación previa in vitro encontró que el producto de semilla de uva tenía la capacidad de regular la producción de AGV y mejorar la fermentación ruminal (26). Después del destete, los jóvenes rumiantes se someten a un estrés evidente y son vulnerables a las bacterias dañinas, lo que resulta en una reducción de la tasa de crecimiento, la capacidad antioxidante y la inmunidad (27). Como se mencionó anteriormente, estudios previos en corderos, vacas lecheras y terneros de engorde verificaron que la suplementación dietética con productos de semilla de uva alivió el estrés oxidativo y la inflamación, y mejoró la capacidad antioxidante y la inmunidad, que fueron útiles para mejorar el rendimiento del crecimiento (20-22, 24). Sin embargo, en la actualidad, la información sobre el papel del GSE como aditivo alimentario en corderos destetados sigue siendo escasa. Sobre la base de estudios previos, planteamos la hipótesis de que la suplementación dietética con GSE puede mejorar la fermentación ruminal, la digestibilidad de los nutrientes y la inmunidad de los corderos destetados. Por lo tanto, el experimento actual se llevó a cabo para estudiar los efectos de GSE en el rendimiento del crecimiento, la digestión de nutrientes, la capacidad antioxidante y la inmunidad de los corderos destetados.
2 Materiales y métodos
2.1 Diseño experimental y manejo de animales
En este estudio, se seleccionaron 30 corderos machos de oveja Hu con edad similar (48 ± 2 días) y peso corporal (15,43 ± 0,49 kg) después del destete. Los corderos fueron marcados con una marca en la oreja y asignados aleatoriamente a uno de dos grupos con 15 corderos cada uno. Un grupo se utilizó como control sin suplementación con GSE (CON), y el otro grupo fue el grupo GSE que se suplementó con GSE (Beisong Plant Technology Co., Ltd., Xi’an, China; componentes principales: proantocianidina 50%, catequina 24%, ácido gálico 16% y epicatequina 6%). La cantidad de aditivo de GSE fue de 0,6 g/d por cordero, lo que se basó en investigaciones previas en corderos de engorde (21).
Todos los corderos experimentales se alojaron en 30 conejeras con 1 cordero en cada conejera. A los corderos se les proporcionó la misma dieta basal dos veces al día a las 09:00 y a las 17:00 respectivamente, lo que permitió un 5 a 10% de orts, y tuvieron acceso ad libitum para beber agua limpia. El ensayo de alimentación duró 70 d, consistente en 10 d de fase adaptativa, seguido de 60 d de período experimental. En la alimentación matutina, el GSE se mezcló completamente con la dieta experimental para alimentar a los corderos y asegurar una ingesta completa. La dieta experimental se formuló de acuerdo con el NRC (28) y el nivel nutricional de la dieta se ajustó al estándar del NRC. Las composiciones de los alimentos y los niveles nutricionales de la dieta se describen en la Tabla 1.
2.2 Medición del rendimiento del crecimiento
Antes de la alimentación matutina en d 0 y 60, se utilizó la báscula electrónica para determinar el peso corporal de los corderos. El GMD se calculó a partir del peso corporal inicial y final. La ingesta de alimento de cada cordero se registró a partir de la diferencia entre el alimento diario ofrecido y los orts, y posteriormente se transformó en DMI. La eficiencia alimenticia de los corderos se obtuvo dividiendo el DMI por la GMD.
2.3 Toma de muestras de sangre, líquido ruminal y heces
Antes de la alimentación matutina con d 0 y 60, la sangre de todos los corderos se recogía de la vena yugular mediante el uso de tubos de vacío sin ningún aditivo (Kangcai Medical Equipment Manufacturing Co., Ltd., Guangzhou, China). A continuación, se utilizaron muestras de sangre para obtener suero por centrifugación a 3200 rpm durante 12 min. Las muestras de suero se almacenaron en tubos centrífugos a -20 °C para su posterior análisis. Además, se recogieron muestras de líquido ruminal de corderos (30 mL) utilizando un tubo estomacal (Huazhi Kaiwu Technology Co., Ltd., Chengdu, China) el día 60 a las 4 h después de la alimentación matutina. Después de medir el pH del fluido ruminal con un medidor de pH, las muestras se filtraron con un paño de nailon y luego se conservaron en tubos de centrífuga a -20 °C para determinar la fermentación ruminal.
Las muestras fecales de cada cordero se recogieron desde el día 54 hasta el 59 mediante placas de tamiz de nylon bajo el suelo de una conejera individual. Durante la recolección de muestras fecales, se recolectó el alimento ofrecido y rechazado para cada cordero. Las muestras fecales se mezclaron minuciosamente durante 6 días consecutivos y se pesaron. Luego, se recolectó el 10% del total de heces para los siguientes procedimientos. El alimento fresco, las heces y las heces fueron mezcladas por cada cordero, submuestreadas y secadas a 65°C para obtener un peso constante. A continuación, todas las muestras se rompieron a través de una pantalla de 1 mm (Jinzhen Machinery manufacturing Co., Ltd., Xinxiang, China) para el análisis de la digestibilidad aparente. Además, una parte de las muestras fecales de cada cordero se conservaron en tubos estériles a -80 °C para medir los recuentos microbianos.
2.4 Medición de la fermentación ruminal y de los parámetros séricos
Las muestras de fluido ruminal se descongelaron primero y luego se centrifugaron a 15000 rpm durante 10 min para obtener sobrenadante. Las muestras de sobrenadante recolectadas se utilizaron para analizar las características de la fermentación ruminal, incluidos los AGV (29), el nitrógeno amoniacal (NH3-N) (30) y la proteína microbiana (MCP) (31). Antes de la medición de los parámetros séricos, las muestras se descongelaron y se mezclaron completamente. El contenido de indicadores bioquímicos, como alanina transaminasa (ALT), aspartato transaminasa (AST), fosfatasa alcalina (ALP), triglicéridos (TG), ácidos grasos no esterificados (NEFA), glucosa (GLU), nitrógeno ureico (UN), proteínas totales (TP), albúmina (ALB) y globulina (GLB) en suero se analizaron utilizando un analizador bioquímico. Los parámetros antioxidantes séricos, incluyendo glutatión peroxidasa (GSH-Px), malondialdehído (MDA), catalasa (CAT), superóxido dismutasa (SOD) y capacidad antioxidante total (T-AOC) se midieron con kits comerciales (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Además, los contenidos de inmunoglobulinas séricas y citocinas incluyendo IL-1β, IL-6, IL-10, TNF-α, inmunoglobulina M (IgM), IgA e IgG se determinaron mediante kits comerciales (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) siguiendo los procedimientos de las especificaciones.
2.5 Medición de la digestibilidad de los nutrientes y del recuento microbiano
Los contenidos de MS (método 934.01), materia orgánica (MO, método 942.05), EE (método 954.02) y PC (método 984.13) en alimento fresco, orts y heces se analizaron siguiendo los métodos de AOAC (32). Además, los contenidos de FDN y FDAF en estas muestras se midieron utilizando un analizador de fibra Ankom. Con el contenido de nutrientes en las heces y el alimento, la digestibilidad aparente (DA) se calculó mediante una ecuación de la siguiente manera: AD (%) = [(ingesta de nutrientes − contenido de nutrientes en las heces)/ingesta de nutrientes] × 100.
Se utilizó el método en placa para contar los Lactobacillus, Bifidobacterium, Escherichia coli y Salmonella en las heces de corderos de acuerdo con los procedimientos de estudios previos (33, 34). Brevemente, en una consola aséptica, se disolvieron 10 g de muestras fecales de cada cordero en 90 mL de agua estéril y se vibraron para mezclar bien. Luego, el líquido mezclado se diluyó en serie para enumerar la composición microbiana. Las pequeñas protuberancias que se formaron en la placa MRS con bordes ligeramente blancos, húmedos y limpios eran Lactobacillus. Las colonias de anillos concéntricos dobles que formaban una protuberancia central en la placa BBL se consideraron Bifidobacterium. Además, pequeñas colonias con brillo negro violáceo y metálico que se formaban en la placa de azul de metileno de eosina eran Escherichia coli. Las colonias con centro negro que se formaron en el medio SS se consideraron Salmonella.
2.6 Análisis estadístico
Cada cordero fue considerado como una unidad experimental para analizar los datos. Después de verificar la normalidad y homogeneidad de las pruebas de varianza, los datos que incluían el rendimiento del crecimiento, la fermentación ruminal, la digestibilidad aparente, los índices sanguíneos y el recuento microbiano se analizaron a través de la prueba t de muestras independientes del software SPSS (versión 22.0). Los resultados se muestran como medias y error estándar de media. Se declaró diferencia significativa a p < 0,05 y se indicó tendencia a 0,05 < p < 0,01.
3 Resultados
3.1 Evolución del crecimiento
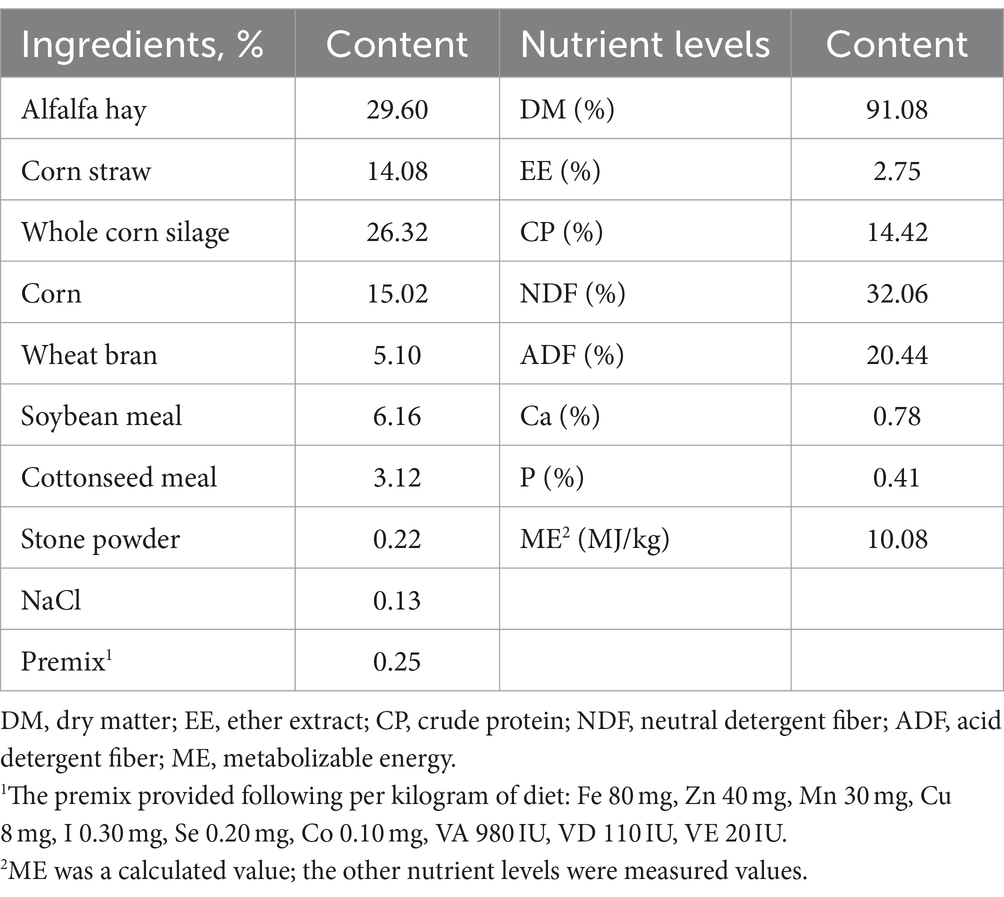
Los efectos de la suplementación con GSE en el rendimiento del crecimiento de los corderos destetados se presentan en la Tabla 2. No hubo diferencia significativa (p > 0,05) de BW y DMI iniciales entre los grupos CON y GSE. Sin embargo, la suplementación dietética con GSE aumentó significativamente (p < 0,05) el peso corporal final y la GMD. Además, en comparación con el grupo CON, la eficiencia alimenticia mejoró en un 7,02% (p = 0,041) en el grupo GSE.
3.2 Fermentación ruminal
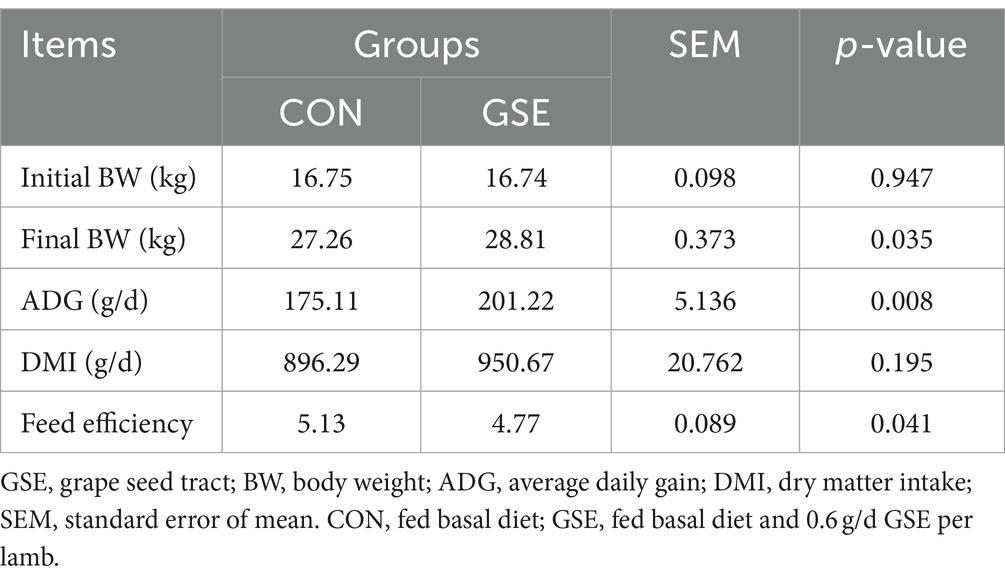
Como se ilustra en la Figura 1A, el pH ruminal promedio de los grupos CON y GSE fue de 6,46 y 6,54 respectivamente, y no mostró diferencias evidentes (p > 0,05). Del mismo modo, no se observaron diferencias evidentes (p > 0,05) de las concentraciones ruminales totales de AGV (Figura 1D) y acetato (Figura 1E) entre los dos grupos. Sin embargo, la suplementación con GSE redujo significativamente (p < 0,05) el contenido de NH3-N (Figura 1B). Por el contrario, las concentraciones de propionato ruminal (Figura 1F) y butirato (Figura 1G) del grupo GSE fueron mayores (p < 0,05) que las del grupo CON. En comparación con el grupo CON, la relación acetato a propionato (Figura 1H) disminuyó en un 12,37% (p < 0,05) en el grupo GSE. Además, la concentración de MCP (Figura 1C) en el grupo GSE tendió a ser mayor (p = 0,088) que en el grupo CON.
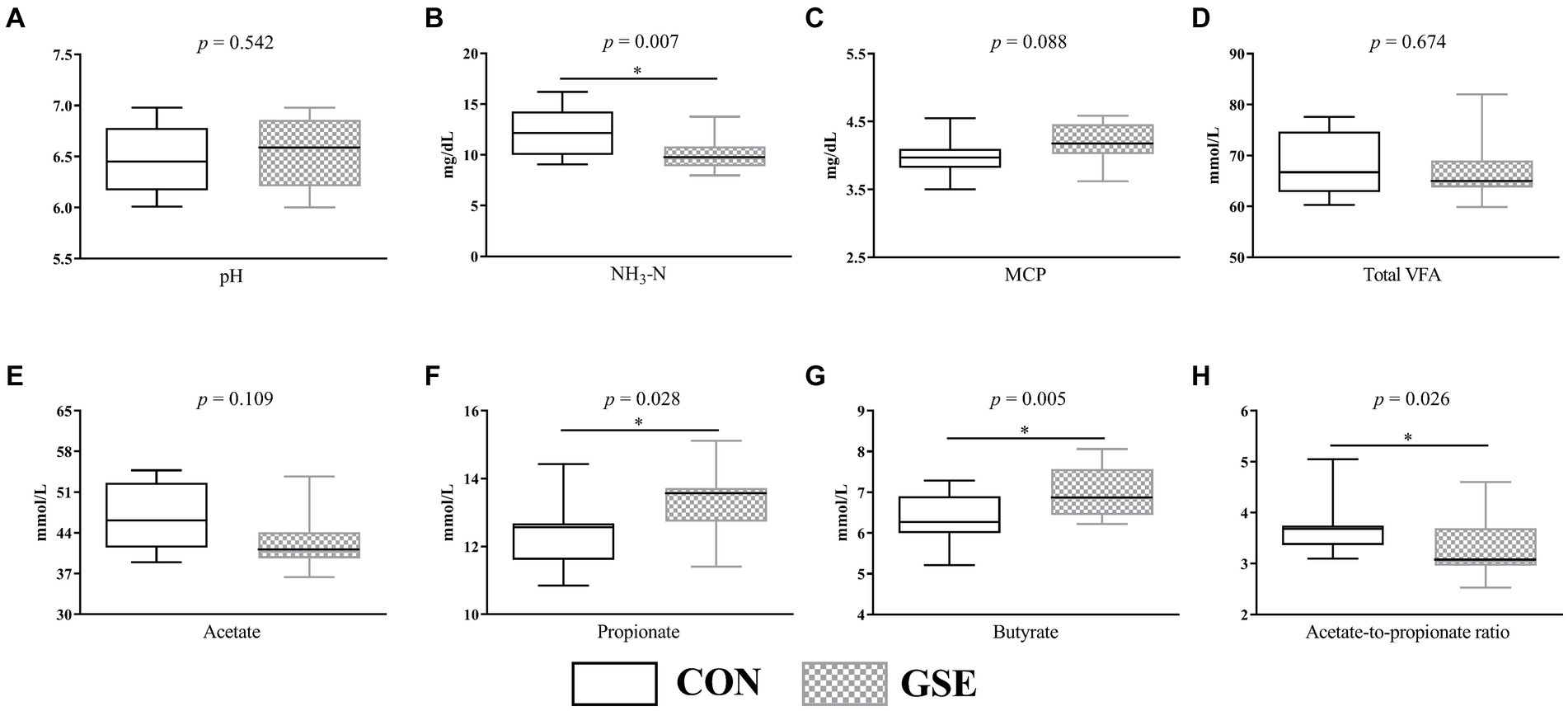
Figura 1. Efectos de la suplementación con GSE sobre las características de fermentación ruminal de los corderos. A) pH; (B) NH3-N; (C) MCP; (D) AGV total; E) acetato; (F) propionato; (G) butirato; (H) Relación acetato a propionato. GSE, tracto de semillas de uva; NH3-N, nitrógeno amonical; MCP: proteína microbiana; AGV: ácidos grasos volátiles. CON, alimentado con dieta basal; GSE, alimentado con dieta basal y 0,6 g/d GSE por cordero. El asterisco representa una diferencia significativa (p < 0,05) entre los tratamientos CON y GSE.
3.3 Digestibilidad aparente de los nutrientes
Obviamente, la digestibilidad aparente de MS (Figura 2A), MO (Figura 2B) y ADF (Figura 2E) fueron similares (p > 0.05) entre los grupos CON y GSE. Sin embargo, los corderos alimentados con GSE aumentaron significativamente (p < 0.05), la digestibilidad de la PCD (Figura 2C), la FDN (Figura 2D) y la EE (Figura 2F).
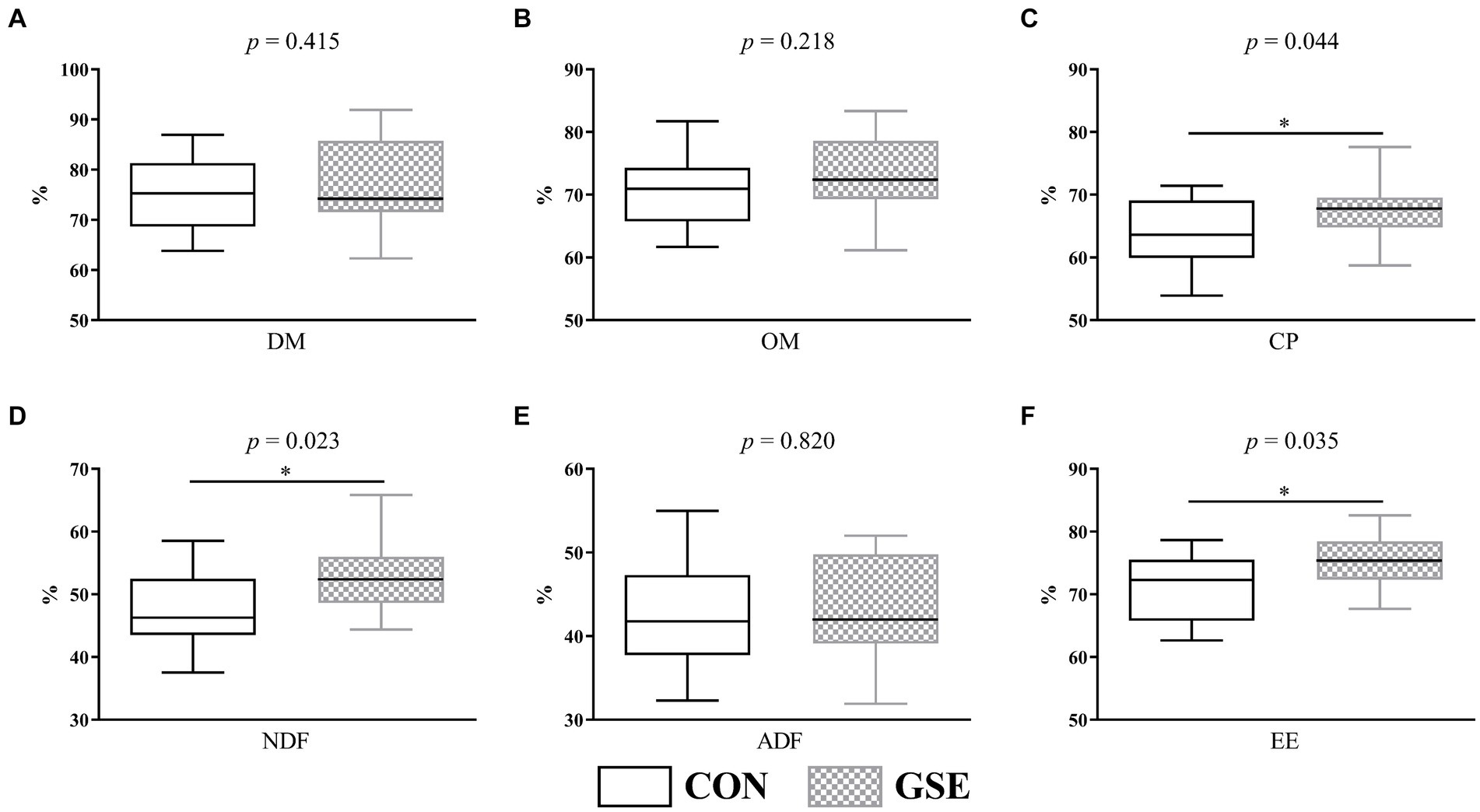
Figura 2. Efectos de la suplementación con GSE sobre la digestibilidad aparente de nutrientes de los corderos. (A) DM; (B) OM; C) CP; d) DENP; e) ADF; f) EE. GSE, tracto de semillas de uva; MS: materia seca; Materia orgánica OM; CP: proteína bruta; NDF, fibra detergente neutra; ADF, fibra detergente ácida; EE, extracto de éter. CON, alimentado con dieta basal; GSE, alimentado con dieta basal y 0,6 g/d GSE por cordero. El asterisco representa una diferencia significativa (p < 0,05) entre los tratamientos CON y GSE.
3.4 Indicador bioquímico del suero
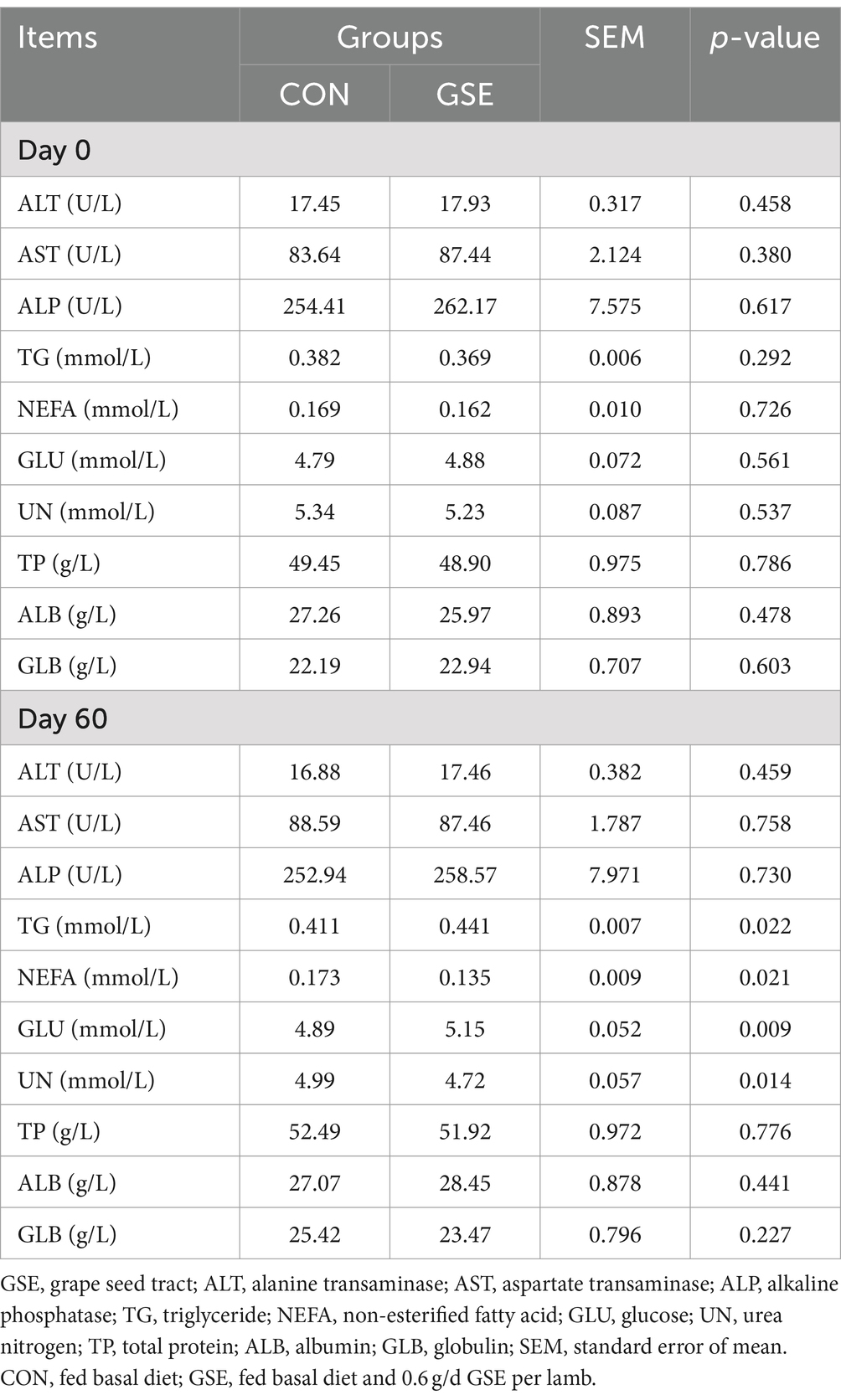
En d 0, las concentraciones de todos los índices bioquímicos en suero de corderos no fueron diferentes (p > 0,05) en los grupos CON y GSE (Tabla 3). Asimismo, los contenidos séricos de ALT, AST, ALP, TP, ALB y GLB fueron similares (p > 0,05) entre los dos grupos en el día 60. Sin embargo, los contenidos de TG y GLU del grupo GSE fueron mayores (p < 0,05) que los del grupo CON, mientras que se encontró una tendencia opuesta en los contenidos de NEFA y UN entre los dos grupos.
3.5 Inmunoglobulina sérica
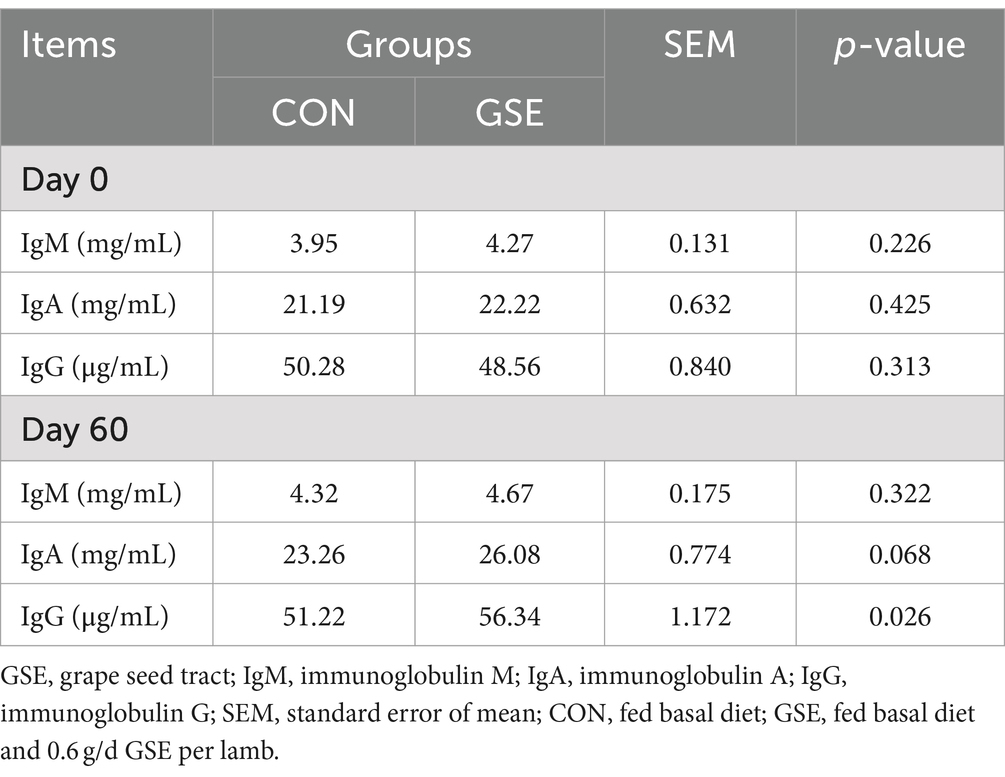
No se encontró diferencia significativa (p > 0,05) de los contenidos de IgM, IgA e IgG entre los tratamientos CON y GSE en d 0 (Tabla 4). Se observó una tendencia similar de IgM entre los dos grupos en el día 60. En comparación con el grupo CON, las concentraciones de IgA e IgG en suero de corderos aumentaron un 12,12% (p = 0,068) y un 10,00% (p = 0,026) en el grupo GSE, respectivamente.
3.6 Citocinas séricas
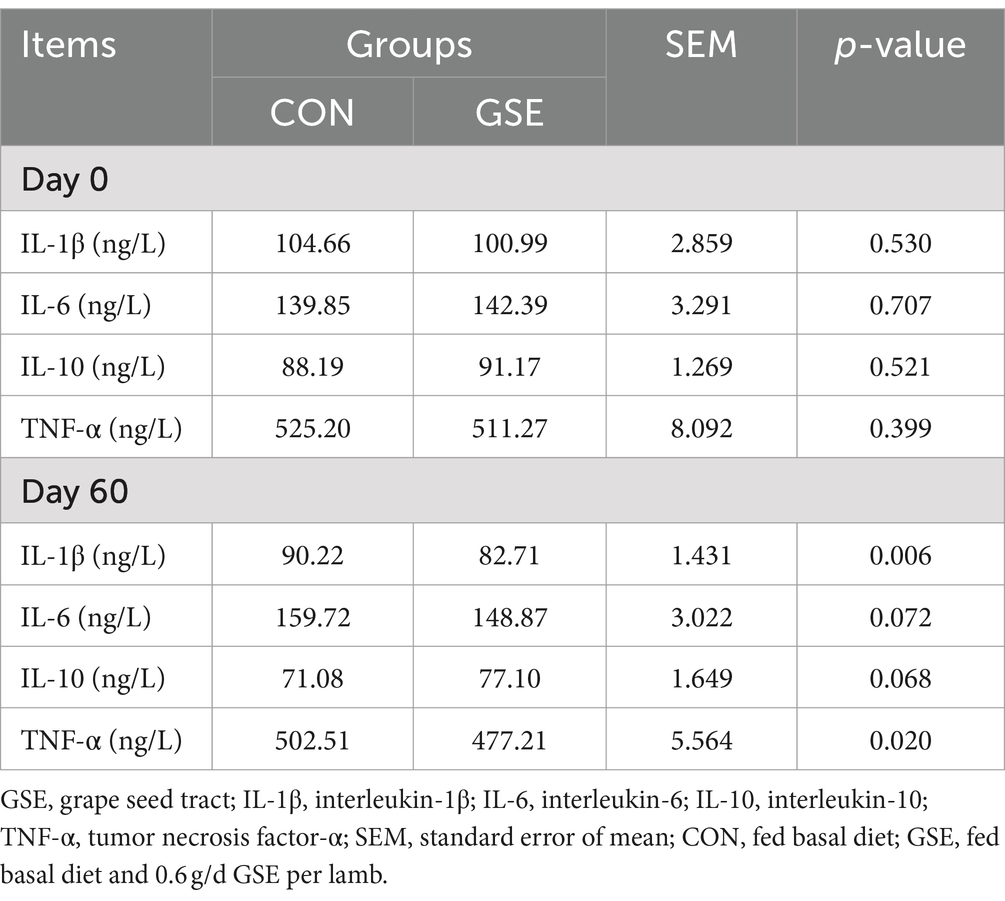
La Tabla 5 muestra la diferencia de las concentraciones de citocinas entre los grupos CON y GSE. En el día 0, no hubo diferencia evidente (p > 0,05) en los contenidos séricos de IL-1β, IL-6, IL-10 y TNF-α entre los dos grupos. En el día 60, la suplementación dietética de GSE redujo significativamente (p < 0,05) las concentraciones séricas de IL-1β y TNF-α en corderos. Además, el contenido de IL-6 en el tratamiento con GSE fue ligeramente inferior (p = 0,072) al del grupo CON, mientras que se encontró una tendencia opuesta a la IL-10 entre los dos grupos.
3.7 Índice antioxidante del suero
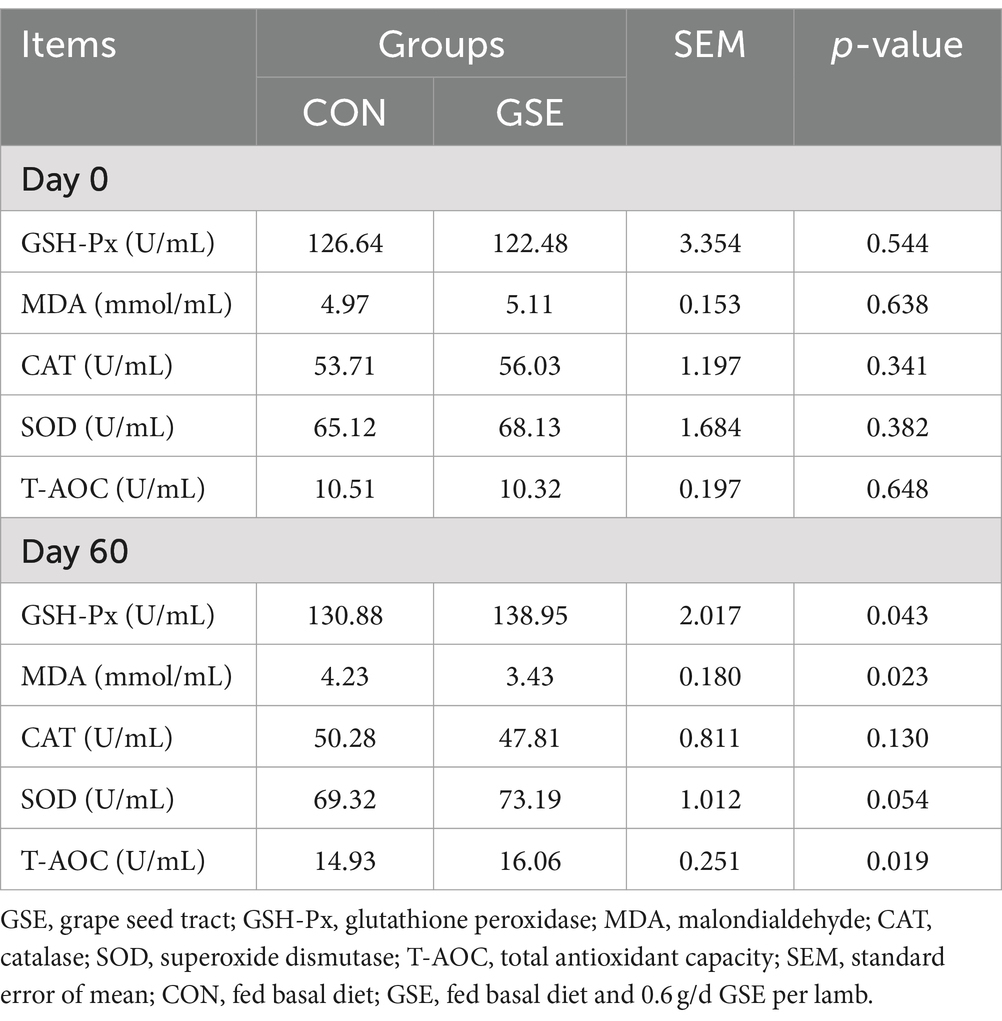
Los parámetros antioxidantes séricos no mostraron diferencia significativa (p > 0,05) entre los tratamientos CON y GSE en el día 0 (Tabla 6). Después de la suplementación con GSE, las actividades séricas de GSH-Px y T-AOC de los corderos aumentaron significativamente (p < 0,05), mientras que la concentración de MDA mostró una tendencia opuesta entre los dos grupos. Además, se encontró una ligera mejora (p = 0,054) de la concentración de SOD en el grupo GSE en comparación con el grupo CON.
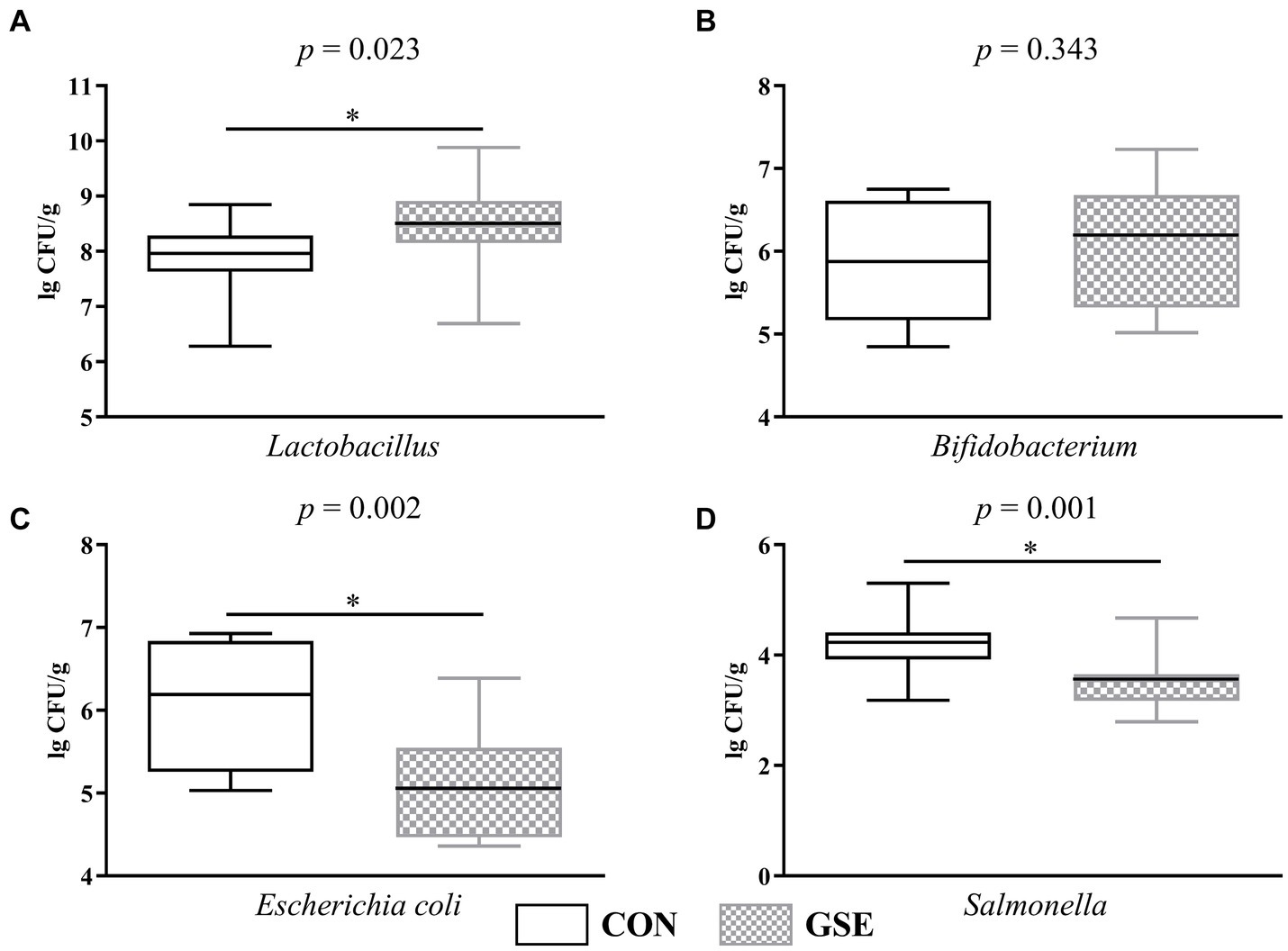
3.8 Recuento microbiano de heces
En particular, la suplementación dietética con GSE aumentó significativamente (p < 0,05) el recuento de Lactobacillus en las heces de los corderos (Figura 3A). El número de Bifidobacterium no mostró diferencia significativa (p > 0,05) entre los grupos CON y GSE (Figura 3B). En comparación con el grupo CON, la suplementación dietética con GSE disminuyó notablemente (p < 0,05) los recuentos de Escherichia coli (Figura 3C) y Salmonella (Figura 3D) en las heces de los corderos.
4 Discusión
La nutrición del cordero ha atraído cada vez más atención en la industria ovina actual. La cría saludable de corderos es de gran importancia para la tasa de crecimiento futura y la productividad de las ovejas de engorde (35, 36). Debido a que el tracto gastrointestinal y el sistema inmunológico aún se encuentran en etapa de desarrollo, los corderos son extremadamente vulnerables a los microbios dañinos. Después del destete, los corderos experimentan ciertos desafíos, como cambios en el alimento, el régimen de alimentación y el metabolismo fisiológico, lo que conduce a una reducción de la tasa de crecimiento y la inmunidad, así como a una mayor tasa de incidencia (5, 37). El alivio del estrés del destete tiene impactos importantes en la promoción del crecimiento rápido de los corderos (36). El GSE, como aditivo verde para piensos, tiene una variedad de funciones fisiológicas, que incluyen efectos antioxidantes, antibacterianos y antiinflamatorios (19). En ovejas de engorde, una investigación encontró que la ración alta en concentrado suplementada con proantocianidina de semilla de uva aumentó la ingesta de alimento y el aumento de peso (21). En el experimento actual, el peso corporal y el GMD de los corderos aumentaron obviamente después de la adición dietética de GSE, pero la ingesta de alimento no mostró cambios evidentes. Las diferencias en los resultados de la investigación pueden deberse a que la etapa de desarrollo (post-destete y engorde respectivamente) de los animales de experimentación fue distinta. Los resultados de la investigación obtenidos de nuestro experimento también mostraron que la suplementación con GSE mejoró significativamente la conversión alimenticia de los corderos. Anteriormente, un experimento en conejos encontró que la conversión alimenticia mejoró después de la suplementación con GSE en condiciones de estrés por calor (38), lo que fue consistente con nuestro hallazgo. La proantocianidina de la semilla de uva puede oxidar las proteínas en la superficie de las bacterias patógenas, lo que puede destruir la estructura y la función de las moléculas de proteínas, lo que resulta en la muerte bacteriana (39). El efecto positivo de la proantocianidina mencionado anteriormente es propicio para inhibir el crecimiento de bacterias patógenas, reduciendo así la morbilidad y atenuando la influencia adversa sobre el crecimiento de los corderos inducida por el estrés del destete.
El rango de fluctuación normal del pH ruminal es de 6 a 7, y es un índice vital para juzgar visualmente la condición saludable del rumen (6). Una investigación reciente informó que una ración baja de forraje suplementada con subproducto de uva redujo el pH ruminal (40). Desafortunadamente, no encontramos diferencias obvias de pH ruminal entre los grupos CON y GSE. Sin embargo, los valores de pH de los dos tratamientos oscilaron entre 6,15 y 6,98, lo que sugiere en parte que el GSE no tuvo un impacto adverso sobre la salud ruminal. Por otro lado, los corderos destetados alimentados con GSE redujeron el contenido ruminal de NH3-N, lo que indica que el GSE podría mejorar la utilización de NH3-N hasta cierto punto. De acuerdo con nuestro estudio, una investigación previa encontró que los polifenoles tenían la capacidad de disminuir el NH3-N del rumen y aumentar las concentraciones de MCP (41). Los microorganismos ruminales pueden sintetizar MCP utilizando amoníaco y otros nutrientes. Curiosamente, el contenido de MCP en el rumen de los corderos aumentó ligeramente después de la suplementación con GSE, lo que correspondió al resultado de NH3-N. En el rumen, la suplementación de polifenoles vegetales es útil para debilitar la degradación de proteínas por parte de los microorganismos ruminales y mejorar las proteínas dietéticas absorbibles para el intestino delgado (42). El papel positivo de la GSE en la mejora de la MCP ruminal puede atribuirse a la ventaja potencial de los polifenoles mencionados anteriormente.
El AGV es la principal fuente de energía para mantener el metabolismo y la productividad de los rumiantes, proporcionando entre el 70 y el 80% de las necesidades energéticas (43). En nuestro experimento, los contenidos de propionato y butirato en el rumen de los corderos fueron mayores en el grupo GSE que en el grupo CON. Como importante fuente de energía, la GLU necesaria para el metabolismo de los rumiantes procede principalmente de la gluconeogénesis en el hígado. El propionato es un precursor primario de la gluconeogénesis hepática (44). Por lo tanto, el aumento de propionato en el rumen podría promover el crecimiento de los corderos. El butirato es una sustancia energética importante para el epitelio ruminal y puede regular al alza las expresiones génicas relacionadas con la proliferación celular, lo que desempeña un papel esencial en la estimulación del desarrollo ruminal (25). Además, una investigación preliminar ha confirmado que el butirato puede acelerar el desarrollo del tejido mucoso a través de la disminución de la apoptosis (45). El aumento de la concentración de butirato en el rumen después de la suplementación con GSE podría ser beneficioso para promover el desarrollo ruminal de los corderos. También encontramos que la suplementación dietética de GSE disminuyó significativamente el acetato ruminal a propionato, lo que sugiere que se produjo una fermentación de tipo propionato en los corderos. La fermentación del propionato ruminal puede mejorar la degradación de la fibra y la utilización del alimento (43), lo que proporciona más energía a los corderos para promover el crecimiento. Anteriormente, una investigación encontró que el subproducto de la semilla de uva aumentó la abundancia relativa de bacterias productoras de butirato utilizando la técnica de simulación ruminal (46), lo que contribuyó a un mayor contenido ruminal de butirato. Sin embargo, la influencia de GSE en el microbioma ruminal de los corderos aún necesita más exploración.
Debido a la secreción inadecuada de enzimas digestivas, la digestibilidad aparente de los nutrientes de los corderos recién destetados es relativamente menor (47). En el presente estudio, la adición de GSE aumentó significativamente la digestibilidad de PC, FDN y EE, lo que indicó que los corderos del grupo GSE pueden obtener más nutrientes para promover el crecimiento. Los microbios ruminales degradan las dietas para producir MCP y péptidos pequeños, y estas sustancias pueden ser fácilmente absorbidas por el intestino delgado para mejorar la digestibilidad de la PC (48). Se especuló que la mejora de la digestibilidad de la PB mediante la suplementación con GSE podría atribuirse al aumento del contenido de MCP. Además, la digestibilidad de la FDN de los corderos GSE fue mayor que la de los corderos CON, lo que concuerda con investigaciones previas de Juráček et al. (49), quienes descubrieron que los animales alimentados con residuos de uva aumentaban la digestibilidad de la FDN. En un estudio previo de terneros lecheros antes del destete, se verificó que la suplementación dietética con ácido gálico, un componente de GSE, aumenta la abundancia relativa de microorganismos asociados con la degradación de la fibra y la producción de AGV en el rumen, que incluían principalmente Ruminococcaceae, Bacteroides y Christensenellaceae (50). La mayor digestibilidad de la FDN del tratamiento con GSE fue probablemente una conexión con la regulación del microbioma gastrointestinal. Además, el aumento de la digestibilidad de la EE de los corderos GSE podría tener efectos beneficiosos sobre el aumento de peso y la deposición de grasa. Por último, la enzima digestiva, existente en el intestino, tiene una función importante en la mejora de la digestibilidad de los nutrientes. Por lo tanto, las investigaciones futuras deberían prestar más atención al estudio de la influencia del GSE en la actividad de las enzimas digestivas en corderos destetados.
Los parámetros bioquímicos séricos están fuertemente relacionados con la condición saludable de los animales, y estos parámetros bioquímicos pueden reflejar el estado metabólico del cuerpo (13, 49). Los cambios en las concentraciones de ALT, AST y ALP en el suero son indicadores vitales relacionados con la función hepática, y los contenidos de GLU, TG y NEFA reflejan el metabolismo lipídico y energético. Además, hasta cierto punto, el metabolismo de las proteínas puede evaluarse mediante cambios en el contenido sérico de TP y UN (24, 49). Un estudio previo en ovejas lecheras lactantes encontró que la suplementación dietética de semillas de uva no tuvo efectos significativos sobre las concentraciones de ALT y AST en la sangre (51). En nuestro experimento, no se observaron diferencias obvias en los contenidos séricos de ALT, AST y ALP, lo que indica que la adición dietética de GSE no tuvo influencia adversa en la función hepática de los corderos. Sin embargo, los resultados obtenidos de nuestro experimento mostraron que las concentraciones séricas de GLU y TG aumentaron significativamente con el tratamiento con GSE, mientras que la suplementación con GSE redujo la concentración sérica de NEFA de los corderos. La posible razón de la mejora en el metabolismo energético de los corderos es que el GSE aumenta el contenido de propionato ruminal y acelera el proceso de gluconeogénesis, lo que resulta en un mayor contenido sérico de GLU. Pero el posible mecanismo molecular requiere más exploración. Después de someterse a estrés como el calor, el frío y el destete de los animales, el TG en el tejido adiposo se descompondrá en NEFA para proporcionar energía para varios metabolismos a través de la circulación sanguínea (52). Los efectos negativos sobre el metabolismo energético de los corderos causados por el destete fueron mejorados por el tratamiento GSE, que favoreció un mejor crecimiento de los corderos. Por otro lado, encontramos que el tratamiento con GSE redujo significativamente el contenido sérico de UN, lo que indica que la conversión de nitrógeno fue mayor en los corderos GSE. La tendencia a la variación en la conversión de nitrógeno fue básicamente consistente con la digestibilidad de la PC, lo que sugiere que la mejor conversión de nitrógeno de los corderos GSE podría atribuirse al aumento de la digestibilidad de la PC. Un estudio previo en vacas lecheras informó que las altas concentraciones de UN, nitrógeno ureico de la leche y NH3-N ruminal se correlacionaron negativamente con la eficiencia del nitrógeno (53). En este experimento, teniendo en cuenta las bajas concentraciones de NH3-N ruminal y UN sérico en corderos GSE, se debe esperar una mayor eficiencia de la conversión de nitrógeno. Una gran cantidad de nitrógeno excretado por los rumiantes reduce la eficiencia de la utilización del alimento y, al mismo tiempo, causa contaminación ambiental (47). La suplementación dietética de GSE aumentó el contenido ruminal de MCP, lo que podría ser beneficioso para mejorar la absorción de nitrógeno en el intestino delgado de los corderos. En el futuro, se deben recopilar más datos, incluida la ingesta de nitrógeno, así como la excreción fecal y urinaria de nitrógeno, para investigar más a fondo los posibles efectos positivos del GSE en la conversión de nitrógeno.
Las inmunoglobulinas séricas son una clase de moléculas proteicas con amplias funciones inmunitarias, que pueden combinarse específicamente con los antígenos, activar el complemento y unirse a los receptores Fc en la superficie celular para obtener inmunidad específica (54). Las concentraciones séricas de inmunoglobulinas se pueden utilizar para reflejar la inmunidad de los animales. En nuestro estudio, la suplementación con GSE aumentó el contenido sérico de IgG e IgA, lo que indica que el GSE mejoró la inmunidad de los corderos destetados. Los polifenoles pueden mejorar la función del sistema inmunológico y la resistencia del cuerpo a las enfermedades al promover la actividad de los glóbulos blancos, aumentar la producción de anticuerpos y mejorar la respuesta inmune celular (55), lo que podría explicar la mejora de GSE en la función inmune. El estrés del destete de los corderos a menudo induce una disfunción del epitelio gastrointestinal y luego resulta en la liberación de factores proinflamatorios, lo que finalmente causa una respuesta inflamatoria (56). Nuestra investigación mostró que la suplementación con GSE disminuyó significativamente la IL-1β sérica y el TNF-α de los corderos, mientras que aumentó la concentración de IL-10. La IL-1β y el TNF-α son las principales citocinas proinflamatorias, y el aumento de las concentraciones se asocia con la respuesta inflamatoria (23). Como citocina antiinflamatoria, la IL-10 puede regular a la baja la expresión de citocinas proinflamatorias como TNF-α, IL-6 e IL-1β mediante la activación de macrófagos e inhibiendo la combinación entre citocinas y receptores (57). Los resultados del estudio actual indicaron que el GSE alivió la reacción inflamatoria de los corderos, lo que contribuyó a un crecimiento saludable. Un estudio previo en terneros sometidos a estrés térmico informó que el GSE disminuyó el contenido plasmático de TNF-α y alivió la reacción inflamatoria (22), lo que estuvo de acuerdo con nuestro resultado. Otra investigación informó que la proantocianidina, un ingrediente principal de GSE, redujo la expresión de factores proinflamatorios a través de la inhibición de la vía de señalización NF-κB (58). En el futuro, se deben llevar a cabo más estudios, incluso in vivo e in vitro, para investigar los efectos potenciales de la vía de señalización NF-κB en la atenuación de la inflamación por GSE.
El destete es una fuente de estrés para los corderos y puede causar estrés oxidativo (5). El MDA, un índice de estrés oxidativo, daña la integridad de las células y puede utilizarse para reflejar el grado de lesión de las células y los tejidos (59). El GSH-Px, el SOD y el T-AOC son índices importantes para evaluar la actividad antioxidante del cuerpo, que puede eliminar los radicales libres y prevenir el daño de las macromoléculas (59). En el presente estudio, las actividades de GSH-Px, SOD y T-AOC en suero de corderos se potenciaron con el tratamiento con GSE, mientras que la actividad de MDA se redujo. Los resultados de nuestro estudio indicaron que la suplementación con GSE aumentó la capacidad antioxidante de los corderos. Experimentos previos en corderos de engorde, terneros lecheros y conejos han demostrado que los productos de semilla de uva pueden aumentar la actividad de la enzima antioxidante en condiciones de estrés (21, 22, 38), lo que está en línea con el estudio actual. Se ha verificado que el polifenol vegetal desempeña el papel de antioxidante por la vía de señalización Nr2 / Keap1 (60), que puede ser responsable de la mejora en la capacidad antioxidante de GSE. En conjunto con los índices séricos, la suplementación con GSE puede mejorar eficazmente el metabolismo energético y la capacidad antioxidante, y finalmente mejorar la inmunidad de los corderos destetados.
El microorganismo intestinal es una barrera eficaz contra los patógenos intestinales, que desempeña un papel importante en la protección de los tejidos intestinales y en el mantenimiento de la función normal del sistema inmunitario (5). El estrés del destete se acompaña de cambios en el entorno intestinal, lo que hace que los corderos sean vulnerables a la invasión de patógenos, lo que provoca diarrea y otras enfermedades (5). La Escherichia coli y la Salmonella son los principales patógenos que inducen diarrea y amenazan la salud de los corderos (61). En este estudio, la suplementación con GSE disminuyó significativamente los recuentos de Escherichia coli y Salmonella, lo que indica que el GSE puede proteger el intestino de los corderos contra las enfermedades. Un estudio reciente informó que GSE tenía bacteriostasis obvia para Escherichia coli (62), lo cual fue consistente con nuestro resultado. La razón puede ser que las proantocianidinas y otros componentes contenidos en la semilla de uva pueden inhibir eficazmente el crecimiento de bacterias patógenas intestinales. Sin embargo, el mecanismo específico aún debe ser estudiado. Los lactobacilos pueden producir ácido láctico en el intestino para mantener un ambiente ácido y luego prevenir la colonización de otros patógenos, protegiendo así la salud intestinal (63). En los terneros lecheros, se ha verificado que el aumento del recuento de Lactobacillus se asocia positivamente con una alta ganancia de peso y eficiencia alimenticia (33). Nuestros resultados microbianos mostraron que la suplementación con GSE tenía la capacidad de proteger la salud intestinal de los corderos. Este efecto positivo también se encontró en la mejora de los parámetros inmunológicos séricos de nuestra investigación. Se requieren futuros experimentos para investigar las funciones de la comunidad microbiana mediante metagenómica y la función del huésped mediante transcriptómica para obtener más resultados sobre el papel del microbioma en el intestino de los corderos y su respuesta al GSE.
5 Conclusión
La adición de GSE puede mejorar la ADG y la eficiencia alimenticia, así como la digestibilidad de PC, FDN y EE de los corderos destetados. También puede mejorar la fermentación ruminal de los corderos destetados, lo que se refleja en un aumento del contenido de propionato y butirato y una disminución de la concentración de NH3-N en el rumen. Además, la suplementación con GSE puede mejorar la capacidad antioxidante y la inmunidad de los corderos destetados al aumentar el contenido sérico de GSH-Px, T-AOC e IgG y el recuento de lactobacilos fecales, y disminuir las concentraciones séricas de MDA, IL-1β y TNF-α y los recuentos fecales de Escherichia coli y Salmonella. Por lo tanto, el GSE puede utilizarse como aditivo alimentario de corderos destetados. Estos datos proporcionaron apoyo teórico para la aplicación de GSE en la cría de corderos.
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán facilitados por los autores, sin reservas indebidas.
Declaración ética
Los procedimientos experimentales relacionados con el cuidado y manejo de corderos fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Oceánica de Guangdong (Zhanjiang, Guangdong, China; Código de homologación: SYXK-2023-024). El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
JM: Conceptualización, Investigación, Visualización, Escritura – borrador original, Escritura – revisión y edición. TL: Investigación, Visualización, Escritura – revisión y edición. LL: Investigación, Visualización, Escritura – revisión y edición. YL: Investigación, Escritura – revisión y edición. XC: Investigación, Escritura – revisión y edición. SL: Investigación, Redacción – revisión y edición. CD: Visualización, Escritura – revisión y edición. CW: Visualización, Escritura – revisión y edición. FY: Supervisión, redacción, revisión y edición. SG: Conceptualización, Curación de datos, Adquisición de fondos, Administración de proyectos, Supervisión, Redacción, revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue apoyada por el Programa de Fondos de Investigación Científica de la Universidad Oceánica de Guangdong (060302052308 y 060302052318) y la Fundación Nacional de Ciencias Naturales de China (32202712).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Vahmani, P, Ponnampalam, EN, Kraft, J, Mapiye, C, Bermingham, EN, Watkins, PJ, et al. Bioactividad y efectos sobre la salud de los lípidos cárnicos de rumiantes. Revisión invitada. Ciencia de la carne. (2020) 165:108114. doi: 10.1016/j.meatsci.2020.108114
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Ding, W, Lu, Y, Xu, B, Chen, P, Li, A, Jian, F, et al. Carne de oveja: información sobre la evaluación del ovino, valor nutritivo, factores influyentes e intervenciones. Agricultura. (2024) 14:1060. doi: 10.3390/agricultura14071060
3. Huang, Y, Liu, L, Zhao, M, Zhang, X, Chen, J, Zhang, Z, et al. Regímenes de alimentación que afectan la canal y los atributos de calidad de la carne de oveja y cabra: una revisión exhaustiva. Anim Biosci. (2023) 36:1314–26. doi: 10.5713/ab.23.0051
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Zhang, H, Zheng, Y, Zha, X, Liu, X, Ma, Y, Loor, JJ, et al. La suplementación dietética con N-carbamilglutamato y L-arginina mejora el estado redox y suprime la apoptosis en el colon de corderos lechales con retraso en el crecimiento intrauterino. Anim Nutr. (2022) 11:359–68. doi: 10.1016/j.aninu.2022.08.009
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Guo, H, Cui, J, Li, Q, Liang, X, Li, J, Yang, B, et al. Una evaluación multiómica de los mecanismos de los microbios intestinales utilizados para tratar la diarrea en corderos destetados precozmente. mSystems. (2024) 9:E0095323. doi: 10.1128/msystems.00953-23
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Abdelsattar, MM, Zhao, W, Saleem, AM, Kholif, AE, Vargas-Bello-Pérez, E y Zhang, N. Desarrollo físico, metabólico y microbiano del rumen en cabretos: una revisión sobre los desafíos y estrategias del destete temprano. Animales. (2023) 13:2420. doi: 10.3390/ani13152420
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Li, Y, Guo, YL, Zhang, CX, Cai, XF, Liu, P y Li, CL. Efectos de las formas físicas de los piensos iniciadores sobre el crecimiento, la digestibilidad de los nutrientes, la actividad de las enzimas gastrointestinales y la morfología de los corderos antes y después del destete. Animal. (2021) 15:100044. doi: 10.1016/j.animal.2020.100044
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Ghazwani, M, Hakami, AR, Sani, SS, Sultana, S, Sultana, T, Bashir, W, et al. Actividad antibacteriana del extracto acuoso y metanólico de Mentha piperita frente a bacterias omnipresentes aisladas de la Ovis vignei. Pak Vet J. (2023) 43:103–8. doi: 10.29261/pakvetj/2023.018
9. Tchoupou-Tchoupou, EC, Ndofor-Foleng, HM, Nwenya, JM, Okenyi, NJ, Ikeh Nnana, E, Ngwu, NR, et al. Efectos del extracto de hexano de ajo sobre parámetros hematológicos, bioquímicos e histológicos en pollitos cruzados F1 no infectados e infectados con Salmonella typhimurium. Int J Vet Sci. (2022) 11:435–42. doi: 10.47278/journal.ijvs/2022.135
10. Los extractos de Xu, D, Wang, X, Shi, W y Bao, Y. Lonicera flos y Curcuma longa L. mejoran el rendimiento del crecimiento, la capacidad antioxidante y la respuesta inmunitaria en pollos de engorde. Frente Vet Sci. (2024) 11:1388632. doi: 10.3389/fvets.2024.1388632
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Nawaz, M, Zhou, J, Khalid, I, Shamim, A, Hussain, A, Ahmed, Z, et al. Actividad antiparasitaria del extracto vegetal frente a nematodos gastrointestinales y Rhipicephalus microplus. Int J Vet Sci. (2022) 11:474–8. doi: 10.47278/journal.ijvs/2022.147
12. Velázquez-Antúnez, J, Olivares-Pérez, J, Olmedo-Juárez, A, Rojas-Hernández, S, Villa-Mancera, A, Romero-Rosales, T, et al. Actividad biológica de los compuestos secundarios de las hojas de Guazuma ulmifolia para inhibir la eclosión de los huevos de Haemonchus contortus. Pak Vet J. (2023) 43:55–60. doi: 10.29261/pakvetj/2022.075
13. Rehman, A, Hussain, K, Zaman, MA, Faurk, MAZ, Abbas, A, Mero, WMS, et al. Efecto de los extractos de coneflower, neem y tomillo sobre el rendimiento del crecimiento, la química sanguínea, la inmunidad y la población microbiana intestinal de pollos de engorde. Kafkas Univ Vet Fak. (2023) 29:407–13. doi: 10.9775/kvfd.2023.29625
14. Aljohani, ASM. Compuestos botánicos: un enfoque prometedor para el control de especies de Mycobacterium de importancia veterinaria y zoonótica. Pak Vet J. (2023) 43:633–42. doi: 10.29261/pakvetj/2023.107
15. Coelho, MG, da Silva, AP, de Toledo, AF, Cezar, AM, Tomaluski, CR, Barboza, RDF, et al. Suplementación con mezcla de aceites esenciales en el sustituto de leche de terneros lecheros: rendimiento y salud. PLoS Uno. (2023) 18:e0291038. doi: 10.1371/journal.pone.0291038
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Caliskan, GU, y Emin, N. Eficacia protectora de los aceites de ajo macerados frescos y envejecidos en aceite de cártamo contra las adherencias intraabdominales en ratas. Pak Vet J. (2023) 43:290–6. doi: 10.29261/pakvetj/2023.030
17. Ali, W, Khatyan, U, Sun, J, Alasmari, A, Alshahrani, MY, Qazi, IH, et al. Efecto mitigante del extracto de cáscara de granada contra la lesión testicular inducida por furano por apoptosis, enzimas esteroidogénicas y estrés oxidativo. Quimiosfera. (2024) 358:142086. doi: 10.1016/j.chemosphere.2024.142086
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Silva, JTP, Borges, MH, de Souza, CAC, Fávaro-Trindade, CS, Sobral, PJA, de Oliveira, AL, et al. Extracto rico en fenólicos y antocianinas de orujo de uva: producción por extracción líquida a presión en proceso intermitente y encapsulación por atomización. Seguridad alimentaria. (2024) 13:279. doi: 10.3390/alimentos13020279
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Dimitrina, RK, Yavor, I, Zlatina, RC y Tzonka, G. Potencial antimicrobiano, actividad antioxidante y contenido fenólico de extractos de semillas de uva de cuatro variedades de uva. Microorganismos. (2023) 11:395. doi: 10.3390/microorganismos11020395
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Gressner, DK, Koch, C, Romberg, FJ, Winkler, A, Dusel, G, Herzog, E, et al. El efecto de la semilla de uva y el extracto de harina de orujo de uva en el rendimiento de la leche y la expresión de genes de estrés e inflamación del retículo endoplásmico en el hígado de vacas lecheras al principio de la lactancia. J Dairy Sci. (2015) 98:8856–68. doi: 10.3168/jds.2015-9478
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Mu, C, Yang, W, Wang, P, Zhao, J, Hao, X y Zhang, J. Efectos de una dieta alta en concentración suplementada con proantocianidinas de semilla de uva sobre el rendimiento del crecimiento, la función hepática, la calidad de la carne y la actividad antioxidante en corderos de engorde. Anim Feed Sci Technol. (2020) 266:114518. doi: 10.1016/j.anifeedsci.2020.114518
22. Urkmez, E, y Biricik, H. Suplementación con extracto de semilla de uva en terneros lecheros predestete estresados por calor: I. Efectos sobre el estado antioxidante, la respuesta inflamatoria, los parámetros hematológicos y fisiológicos. Alimentación animada. Sci Technol. (2022) 292:115421. doi: 10.1016/j.anifeedsci.2022.115421
23. Nallathambi, R, Poulev, A, Zuk, JB y Raskin, I. El extracto de semilla de uva rico en proantocianidinas reduce la inflamación y el estrés oxidativo y restaura la función de barrera de unión estrecha en las células de colon caco-2. Nutrientes. (2020) 12:1623. doi: 10.3390/nu12061623
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Urkmez, E, y Biricik, H. Suplementación con extracto de semilla de uva en terneros lecheros predestete sometidos a estrés térmico: II. Efectos sobre el rendimiento del crecimiento, los metabolitos sanguíneos, las respuestas hormonales y los parámetros de fermentación fecal. Anim Feed Sci Technol. (2022) 292:115422. doi: 10.1016/j.anifeedsci.2022.115422
25. Li, Z, Wang, X, Wang, W, An, R, Wang, Y, Ren, Q, et al. Beneficios de la tributirina sobre el rendimiento del crecimiento, el desarrollo del tracto gastrointestinal, las bacterias ruminales y la formación de ácidos grasos volátiles en corderos Han de cola pequeña destetados. Anim Nutr. (2023) 15:187–96. doi: 10.1016/j.aninu.2023.08.006
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Thanh, LP, Kha, PTT, Loor, JJ y Hang, TTT. El extracto de tanino de semilla de uva y los ácidos grasos poliinsaturados afectan in vitro a la fermentación ruminal y a la producción de metano. J Anim Sci. (2022) 100:skac039. doi: 10.1093/jas/skac039
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Hulbert, LE, y Moisa, SJ. El estrés, la inmunidad y el manejo de los terneros. J Dairy Sci. (2016) 99:3199–216. doi: 10.3168/jds.2015-10198
28. Consejo Nacional de Investigaciones Científicas. Requerimientos de nutrientes de los pequeños rumiantes. Washington, DC: The National Academies Press (2007).
29. Erwin, ES, Marco, GJ, y Emery, EM. Análisis de ácidos grasos volátiles de sangre y líquido ruminal por cromatografía de gases. J Dairy Sci. (1961) 44:1768–71. DOI: 10.3168/JDS. S0022-0302(61)89956-6
30. Broderick, GA, y Kang, JH. Determinación simultánea automatizada de amoníaco y aminoácidos totales en fluido ruminal y medios in vitro. J Dairy Sci. (1980) 63:64–75. DOI: 10.3168/JDS. S0022-0302(80)82888-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Makkar, HPS, Sharma, OP, Dawra, RK y Negi, SS. Determinación simple de proteína microbiana en licor ruminal. J Dairy Sci. (1982) 65:2170–3. DOI: 10.3168/JDS. S0022-0302(82)82477-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. AOAC. Métodos oficiales de análisis. Washington, DC: Asociación de Químicos Analíticos Oficiales (2019).
33. Alimirzaei, M, Alijoo, YA, Dehghan-Banadaky, M, y Eslamizad, M. Los efectos de la alimentación con niveles altos o bajos de leche en los primeros años de vida sobre el rendimiento del crecimiento, el recuento microbiano fecal y el estado metabólico e inflamatorio de las crías hembras Holstein. Animal. (2020) 14:303–11. doi: 10.1017/S1751731119001691
34. Sanders, ER. Técnicas de laboratorio aséptico: métodos de enchapado. J Vis Exp. (2012) 63:3064. DOI: 10.3791/3064-V
35. Flinn, T, Kleemann, DO, Swinbourne, AM, Kelly, JM, Weaver, AC, Walker, SK, et al. Mortalidad neonatal de corderos: principales factores de riesgo y el posible papel mejorador de la melatonina. J Animal Sci Biotechnol. (2020) 11:107. DOI: 10.1186/s40104-020-00510-W
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Zhao, F, He, W, Wu, T, Elmhadi, M, Jiang, N, Zhang, A, et al. La suplementación con butirato de sodio recubierto alivió el estrés del destete y remodeló la flora microbiana en corderos destetados. Frente Vet Sci. (2024) 11:1423920. doi: 10.3389/fvets.2024.1423920
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Mao, HL, Ji, WW, Yun, Y, Zhang, YF, Li, ZF y Wang, C. Influencia de la suplementación con probióticos en el rendimiento del crecimiento, las variables plasmáticas y la comunidad bacteriana ruminal de corderos con retraso en el crecimiento. Microbiol frontal. (2023) 14:1216534. doi: 10.3389/fmicb.2023.1216534
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Hassan, FA, Mahrose, KM y Basyony, MM. Efectos del extracto de semilla de uva como antioxidante natural sobre el rendimiento del crecimiento, las características de la canal y el estado antioxidante de los conejos durante el estrés por calor. Arch Anim Nutr. (2016) 70:141–54. doi: 10.1080/1745039X.2016.1139609
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Andersone, A, Janceva, S, Lauberte, L, Ramata-Stunda, A, Nikolajeva, V, Zaharova, N, et al. Actividad antiinflamatoria, antibacteriana y antifúngica de proantocianidinas oligoméricas y extractos obtenidos a partir de residuos agrícolas lignocelulósicos. Moléculas. (2023) 28:863. doi: 10.3390/moléculas28020863
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Suescun-Ospina, ST, Vera, N, Astudillo, R, Yunda, C, Williams, P, Allende, R, et al. Efectos de la inclusión de orujo de uva País en dietas altas y bajas en forraje: fermentación ruminal, producción de metano y ácidos grasos volátiles. Ital J Anim Sci. (2022) 21:924–33. doi: 10.1080/1828051X.2022.2076620
41. Puchalska, J, Szumacher-Strabel, M, Patra, AK, Ślusarczyk, S, Gao, M, Petrič, D, et al. El efecto de diferentes concentraciones de polifenoles totales de hojas híbridas de Paulownia sobre la fermentación ruminal, la producción de metano y los microorganismos. Animales. (2021) 11:2843. doi: 10.3390/ani11102843
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Theodorou, MK, Kingston-Smith, AH, Winters, AL, Lee, MRF, Minchin, FR, Morris, P, et al. Los polifenoles y su influencia en la función intestinal y la salud de los rumiantes: una revisión. Environ Chem Lett. (2006) 4:121–6. doi: 10.1007/s10311-006-0061-2
43. Gäbel, G, y Sehested, J. Transporte de AGCC en el estómago anterior de rumiantes. Comp Biochem Physiol. (1997) 118:367–74. doi: 10.1016/S0300-9629(96)00321-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Aschenbacah, JR, Kristensen, NB, Donkin, SS, Hammon, HM y Penner, GB. Gluconeogénesis en vacas lecheras: el secreto de la elaboración de leche dulce a partir de masa agria. Int Union Biochem Mol Biolo Life. (2010) 62:869–77. DOI: 10.1002/iub.400
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Mentschel, J, Leiser, R, Mülling, C, Pfarrer, C y Claus, R. El ácido butírico estimula el desarrollo de la mucosa ruminal en el ternero principalmente mediante una reducción de la apoptosis. Arch Anim Nutr. (2001) 55:85–102. DOI: 10.1080/17450390109386185
46. Khiaosa-ard, R, Metzler-Zebeli, BU, Ahmed, S, Muro-Reyes, S, Deckardt, K, Chizzola, R, et al. La fortificación de granos secos de destilería más solubles con harina de semilla de uva en la dieta modula la mitigación del metano y la microbiota ruminal en Rusitec. J Dairy Sci. (2015) 98:2611–26. doi: 10.3168/jds.2014-8751
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Zhang, W, Sun, S, Zhang, Y, Zhang, Y, Wang, J, Liu, Z, et al. La suplementación con ácido benzoico mejora el rendimiento del crecimiento, la digestibilidad de los nutrientes y el metabolismo del nitrógeno de los corderos destetados. Frente Vet Sci. (2024) 11:1351394. doi: 10.3389/fvets.2024.1351394
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Ma, J, Zhu, Y, Wang, Z, Yu, X, Hu, R, Wang, X, et al. La suplementación con glutamina afectó a la comunidad bacteriana intestinal y a la fermentación, lo que mejoró la digestibilidad de los nutrientes en los yaks con retraso en el crecimiento. FEMS Microbiol Ecol. (2021) 97:FIAB084. DOI: 10.1093/femsec/fiab084
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Juráček, M, Vašeková, P, Massányi, P, Kováčik, A, Bíro, D, Šimko, M, et al. El efecto de la alimentación del orujo de uva pasajera sobre los nutrientes, la digestibilidad y el perfil bioquímico sérico de las hembras. Agricultura. (2021) 11:1194. doi: 10.3390/agricultura11121194
50. Xu, HJ, Zhang, QY, Wang, LH, Zhang, CR, Li, Y y Zhang, YG. Rendimiento del crecimiento, digestibilidad, metabolitos sanguíneos, fermentación ruminal y comunidades bacterianas en respuesta a la inclusión de ácido gálico en el alimento inicial de terneros lecheros antes del destete. J Dairy Sci. (2022) 105:3078–89. DOI: 10.3168/jds.2021-20838
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Nudda, A, Correddu, F, Marzano, A, Battacone, G, Nicolussi, P, Bonelli, P, et al. Efectos de las dietas que contienen semilla de uva, linaza o ambas sobre los rasgos de producción de leche, las actividades hepáticas y renales y la inmunidad de las ovejas lecheras lactantes. J Dairy Sci. (2015) 98:1157–66. doi: 10.3168/jds.2014-8659
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Moiseeva, K, Anipchenko, P, Vasil’eva, S, Karpenko, LY, Vasil’ev, R, Pilaeva, N, et al. Dinámica del colesterol y triglicéridos en el suero de vacas con lipidosis hepática. J Anim Sci. (2019) 97:208. doi: 10.1093/jas/skz258.427
53. Nousiainen, J, Shingfield, KJ y Huhtanen, P. Evaluación del nitrógeno ureico de la leche como diagnóstico de la alimentación con proteínas. J Dairy Sci. (2004) 87:386–98. DOI: 10.3168/JDS. S0022-0302(04)73178-1
54. Yin, X, Ji, S, Duan, C, Tian, P, Ju, S, Yan, H, et al. La sucesión de la comunidad bacteriana fecal y su correlación con los cambios de los indicadores inmunes séricos en corderos desde el nacimiento hasta los 4 meses. J Integr Agr. (2023) 22:537–50. doi: 10.1016/j.jia.2022.08.055
55. Hong, M, Cheng, L, Liu, Y, Wu, Z, Zhang, P y Zhang, X. Una fuente vegetal natural: los polifenoles del té, un medicamento potencial para mejorar la inmunidad y combatir los virus. Nutrientes. (2022) 14:550. doi: 10.3390/nu14030550
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Izuddin, WI, Loh, TC, Foo, HL, Samsudin, AA, Humam, AM y Postbiotic, L. Plantarum RG14 mejora el crecimiento del epitelio ruminal, el estado inmunológico y regula al alza la función de barrera intestinal en corderos post-destete. Sci Rep. (2019) 9:9938. doi: 10.1038/s41598-019-46076-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. Al-Ashy, B, Chakroun, I, El-Sabban, ME, y Homaidan, FR. El papel de NF-κB en la mediación de los efectos antiinflamatorios de la IL-10 en las células epiteliales intestinales. Citocina. (2006) 36:1–8. doi: 10.1016/j.cyto.2006.10.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Terra, X, Pallarès, V, Ardèvol, A, Bladé, C, Fernandez-Larrea, J, Pujadas, G, et al. Efecto modulador de las procianidinas de semilla de uva sobre la inflamación local y sistémica en ratas con obesidad inducida por la dieta. J Nutr Biochem. (2011) 22:380–7. doi: 10.1016/j.jnutbio.2010.03.006
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Du, X, Cheng, X, Ji, K, Degen, AA, Liang, Y, Wu, X, et al. Efectos del tomillo suplementario sobre las respuestas inmunitarias, los índices antioxidantes, las concentraciones de enzimas ruminales y la composición de bacterias ruminales en ovejas Hu. Anim Feed Sci Technol. (2023) 306:115828. doi: 10.1016/j.anifeedsci.2023.115828
60. Huang, T, Che, Q, Chen, X, Chen, D, Yu, B, He, J, et al. Los polifenoles de manzana mejoran la capacidad antioxidante intestinal y la función de barrera al activar la vía de señalización Nrf2/Keap1 en un modelo de cerdo. J Agr Químico de los Alimentos. (2022) 70:7576–85. doi: 10.1021/acs.jafc.2c02495
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Tuo, X, Wang, S, Cui, D, Wang, F, Wang, H, Liu, Y, et al. Perfiles de resistencia a antibióticos y marcadores de virulencia de cepas de Escherichia coli aisladas de corderos diarreicos en Gansu y Qinghai, China. Pak Vet J. (2020) 40:123–6. doi: 10.29261/pakvetj/2019.102
62. Yaqubi, AK, Astuti, SD, Zaidan, AH, Syahrom, A, y Nurdin, DZI. Efecto antibacteriano de nanopartículas rojas de plata activadas con láser rojo sintetizadas con extracto de semilla de uva contra Staphylococcus aureus y Escherichia coli. Láseres Med Sci. (2024) 39:47. doi: 10.1007/s10103-024-03991-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: extracto de semilla de uva, cordero, rendimiento de crecimiento, digestibilidad de nutrientes, inmunidad
Cita: Ma J, Li T, Lin L, Lu Y, Chen X, Li S, Du C, Wei C, Yin F y Gan S (2024) Efectos de la suplementación con extracto de semilla de uva en el rendimiento del crecimiento, la digestión de nutrientes y la inmunidad de los corderos destetados. Frente. Vet. Sci. 11:1402637. doi: 10.3389/fvets.2024.1402637
Editado por:
Izhar Hyder Qazi, Universidad Shaheed Benazir Bhutto de Ciencias Veterinarias y Animales, Pakistán
Revisado por:
Rao Zahid Abbas, Universidad de Agricultura, Faisalabad, Pakistán
Yuan-Tai Hung, Devenish Nutrition, Estados Unidos
Derechos de autor © 2024 Ma, Li, Lin, Lu, Chen, Li, Du, Wei, Yin y Gan. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Atribución (CC BY).
*Correspondencia: Shangquan Gan, gansq1977@126.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía