
Efectos de la taurina sobre el rendimiento del crecimiento, la diarrea, el estrés oxidativo y la función de barrera intestinal de los lechones

Efectos de la taurina sobre el rendimiento del crecimiento, la diarrea, el estrés oxidativo y la función de barrera intestinal de los lechones destetados

 Zichen Wu1Donghua Deng1Bin Wang2Xiaoling Zhou2Bingyu Zhou2Chunping Wang2Yan Zeng2*
Zichen Wu1Donghua Deng1Bin Wang2Xiaoling Zhou2Bingyu Zhou2Chunping Wang2Yan Zeng2*- 1Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Hunan, Changsha, China
- número arábigoInstituto de Microbiología de Hunan, Changsha, China
El daño oxidativo resultante del estrés del destete afecta significativamente el rendimiento del crecimiento y el estado de salud de los lechones. La taurina, un antioxidante dietético con diversas funciones, se investigó en este estudio por su papel protector contra el daño oxidativo inducido por el estrés del destete y su mecanismo subyacente. Cuarenta lechones machos castrados destetados de 28 días de edad fueron asignados aleatoriamente a cuatro grupos. El grupo de control recibió la dieta basal, mientras que los grupos experimentales fueron alimentados con la dieta basal suplementada con 0,1, 0,2% o 0,3% de taurina durante un período de 28 días. In vitro, se utilizó H2O2 para inducir daño oxidativo a la mucosa yeyunal de lechones a través de células IPEC-J2. Los resultados demostraron que la suplementación con taurina redujo la incidencia de diarrea en lechones en comparación con la del grupo control (p < 0,05); La adición de 0,2 y 0,3% de taurina condujo a un aumento de la ganancia media diaria y a una mejora de la eficiencia de la conversión alimenticia en lechones destetados, mostrando una correlación dosis-respuesta lineal (p < 0,05). La suplementación con taurina al 0,2 y al 0,3% mejoró las actividades séricas de CAT y GSH-Px, al tiempo que disminuyó los niveles séricos de NO, XOD, GSSG y MDA (p < 0,05). Además, elevó significativamente los niveles de GSS, Trx, POD, complejo I, mt-nd5 y mt-nd6, mejorando la capacidad de eliminación de aniones superóxido y la tasa de eliminación libre de hidroxilo en los hígados de lechones destetados, al tiempo que redujo los niveles de NO en el hígado (p < 0,05). Además, la suplementación con taurina al 0,2 y 0,3% disminuyó los niveles séricos de IL-6 y elevó las concentraciones de IgA, IgG e IL-10 en lechones destetados (p < 0,05). Los niveles de ocludina, claudina y ZO-1 en la mucosa del yeyuno de los lechones destetados aumentaron con la suplementación con taurina de 0,2 y 0,3% (p < 0,05). En las células IPEC-J2, el pretratamiento con 25 mM de taurina durante 24 h mejoró las actividades de SOD y TAC; redujo el contenido de MDA; reguló al alza la expresión de ARNm de varios genes, incluidos ZO-1, ocludina, claudina-1, Nrf2 y HO-1; y revirtió el daño oxidativo inducido por la exposición aH2O2 (p < 0,05). En general, los hallazgos sugieren que la inclusión de 2 y 3% de taurina en la dieta puede mejorar el rendimiento del crecimiento, reducir las tasas de diarrea, mejorar el estrés oxidativo y la inflamación, y promover la función de barrera intestinal en lechones destetados.
1 Introducción
En general, la cría de lechones desempeña un papel vital en el proceso de cría de cerdos. La alimentación y el manejo adecuados de los lechones tienen un impacto directo en métricas clave como el tiempo de sacrificio, la tasa y el peso, lo que influye en la eficiencia de la producción de la cría de cerdos (1, 2). El destete es una fase crítica en el ciclo de crecimiento del cerdo y es muy susceptible a los estímulos ambientales, la calidad del alimento, las micotoxinas y los microorganismos patógenos. Estos factores mencionados pueden dar lugar a una sobreproducción de especies reactivas de oxígeno (ROS) intestinales en los lechones, lo que conduce al estrés oxidativo. Este estrés oxidativo puede desencadenar posteriormente daños inflamatorios en la mucosa intestinal y comprometer la función de barrera. Como consecuencia, los lechones pueden experimentar una reducción del rendimiento del crecimiento, diarrea y resultados potencialmente fatales, lo que impone cargas económicas sustanciales a la industria porcina (3, 4).
El estrés oxidativo se produce cuando los animales liberan radicales libres oxidativos que superan las defensas antioxidantes del organismo, dando lugar a diversas manifestaciones fisiológicas o patológicas (5, 6). Las investigaciones indican que el proceso de destete altera el equilibrio redox en los lechones, reduciendo notablemente la actividad de las enzimas antioxidantes al tiempo que eleva los radicales libres y los productos de peróxido, induciendo esencialmente el estrés oxidativo (7, 8). Las células responden al estrés oxidativo activando varios factores de transcripción, incluido el factor 2 relacionado con el factor nuclear E2 (Nrf2). Nrf2 desempeña un papel crucial en la regulación del equilibrio redox celular, facilitando la expresión de antioxidantes protectores e iniciando reacciones de desintoxicación de fase I en mamíferos. Se erige como un objetivo clave para el cuerpo en el manejo del estrés oxidativo (9). Los antioxidantes dietéticos, como las antocianinas y la vitamina E, son potentes antioxidantes naturales capaces de eliminar eficazmente los radicales libres, protegiendo así las células contra el daño oxidativo y el estrés inmunológico (10, 11). En consecuencia, mejorar el estado redox de los lechones es una estrategia crucial para fomentar un desarrollo saludable.
La suplementación de las dietas de los lechones con componentes funcionales ha surgido como un enfoque factible y eficaz para mitigar el estrés oxidativo después del destete (7, 12). La taurina, un aminoácido no proteico que contiene azufre, desempeña un papel fundamental en los procesos celulares como la regulación del volumen, el ajuste de la presión osmótica, la fosforilación de proteínas, la estabilidad de la membrana, el metabolismo de los ácidos biliares, la modulación neuronal, la homeostasis del calcio y la desintoxicación, y exhibe propiedades antioxidantes y antiinflamatorias que son cruciales para mantener la homeostasis animal (13, 14). Investigaciones recientes subrayan la capacidad de la taurina para aliviar diversas dolencias relacionadas con el daño oxidativo y su participación en la modulación del sistema inmunológico, ofreciendo beneficios significativos para la producción animal (15-17). Por lo tanto, este estudio tuvo como objetivo evaluar cómo la taurina afecta el rendimiento del crecimiento, los niveles de estrés oxidativo, la función inmune y la integridad de la barrera intestinal en lechones destetados.
2 Materiales y métodos
Este estudio en animales fue revisado y aprobado por el Comité Institucional de Cuidado y Uso de Animales (202105) de la Universidad Agrícola de Hunan. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
2.1 Materiales
La taurina (pureza ≥99%) y el peróxido de hidrógeno (pureza 30%) se compraron a Shanghai Aladdin Biochemical Technology Co., Ltd.
2.2 Diseño experimental y dietas
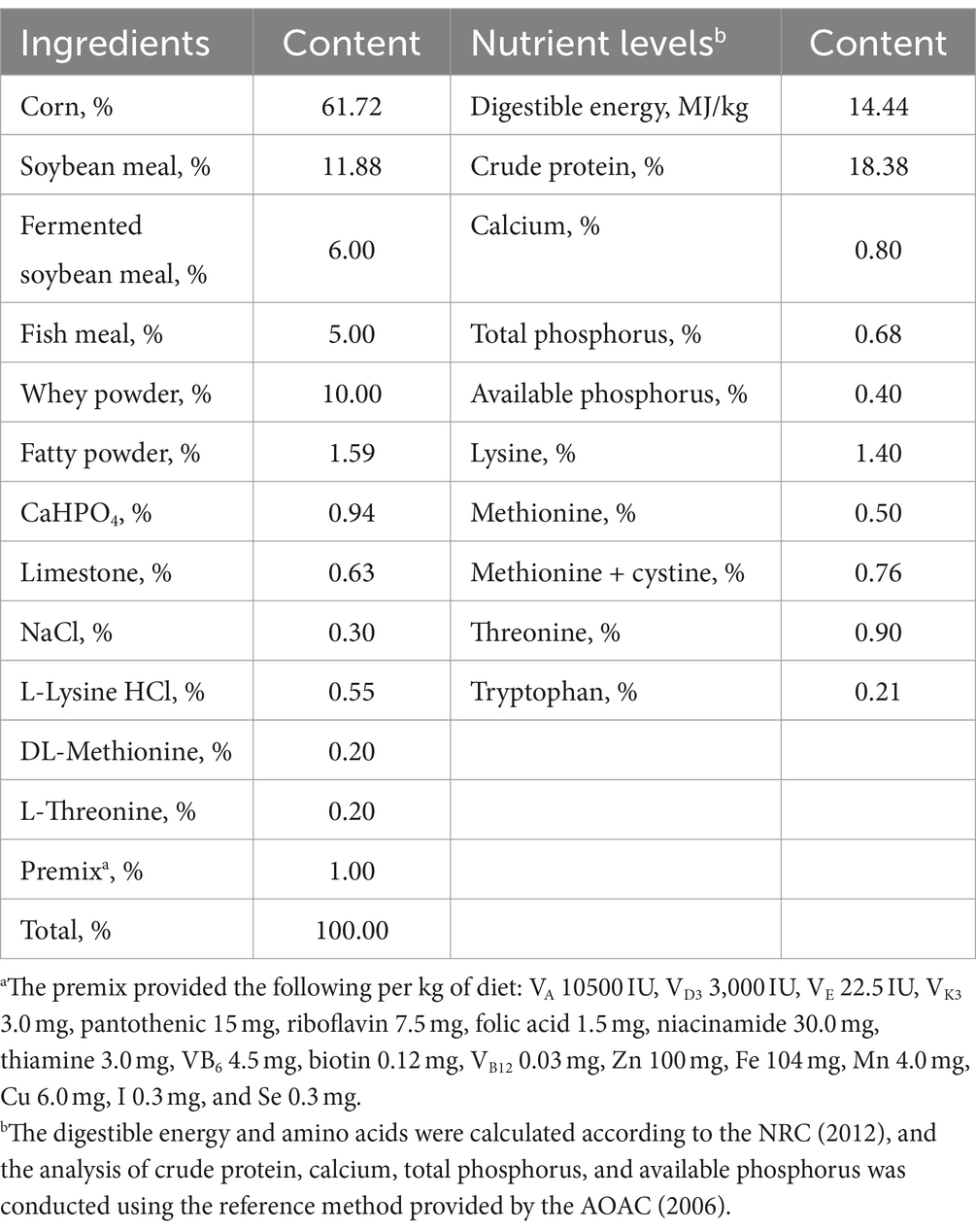
Cuarenta lechones Duroc × Landrace castrados sanos de 28 días de edad × grandes destetados se dividieron aleatoriamente en 4 grupos con 10 réplicas en cada grupo y 1 cerdo en cada réplica. Las plantas se alimentaron en un solo corral y se alimentaron durante 3 días. El grupo control fue alimentado con una dieta basal, y el grupo experimental fue alimentado con una dieta suplementada con 0,1, 0,2% o 0,3% de taurina. El período de prueba fue de 28 días. La dieta básica se preparó de acuerdo con la NRC 2012, y la composición del alimento y el nivel nutricional se muestran en la Tabla 1. El establo de cerdos se limpió y desinfectó antes del experimento. Durante el experimento, los animales fueron alimentados dos veces al día, se les proporcionó comida libremente y se les permitió beber libremente. Los lechones fueron sometidos a procedimientos de inmunización de rutina, y su estado de salud fue cuidadosamente observado y registrado todos los días.
2.3 Recogida de muestras
Después de 12 h de ayuno, se recolectó la vena cava anterior de los lechones en la mañana del día 29 del período experimental. La sangre entera se colocó en un recipiente ordinario de recolección de sangre al vacío. Después de permanecer a temperatura ambiente durante 30 min, la mezcla se centrifugó a 845 g y 4 °C durante 10 min, después de lo cual el sobrenadante se separó en un tubo EP de 1,5 mL. Después de una congelación rápida con nitrógeno líquido, las muestras se almacenaron a -80 °C. Los pacientes fueron anestesiados aleatoriamente con pentobarbital sódico (40 mg/kg) y sacrificados mediante sangría. Se abrieron la cavidad torácica y la cavidad abdominal, y se separaron el hígado y el yeyuno. Se extrajo una cantidad adecuada de hígado del tubo congelado en la posición fija y se almacenó a -80 °C después de una congelación rápida con nitrógeno líquido. Se extrajeron aproximadamente 20 cm de tejido del yeyuno medio y se enjuagó el tejido con solución salina normal. La capa mucosa se raspó con un portaobjetos, se recogió en un tubo de criopreservación para una congelación rápida con nitrógeno líquido y luego se almacenó a -80 °C.
2.4 Evolución del crecimiento
Los lechones se pesaron con el estómago vacío en las mañanas del día 1 y el día 29 del experimento. Durante el experimento, se registró la ingesta diaria de alimento y el peso residual del alimento para calcular la ganancia diaria promedio (ADG), la ingesta diaria promedio de alimento (ADFI) y la relación alimento-ganancia (F/G). La diarrea de los lechones se observó y registró a las 17 horas todos los días, y la tasa de diarrea (%) = número de lechones con diarrea × 100/28.
2.5 Parámetros séricos
Los contenidos de catalasa sérica (CAT), glutatión peroxidasa (GSH-Px), superóxido dismutasa (SOD), capacidad antioxidante total (T-AOC), capacidad de eliminación de aniones superóxido, tasa de eliminación de radicales hidroxilo, óxido nítrico (NO), actividad de xantina oxidasa (XOD), glutatión (GSH), glutatión oxidado (GSSG), malondialdehído (MDA) y peroxidasa (POD) fueron detectados por kits bioquímicos producidos por el Instituto de Bioingeniería de Nanjing Jianchen, China. La inmunoglobulina sérica A (IgA), la inmunoglobulina G (IgG), la inmunoglobulina M (IgM), la interleucina-4 (IL-4), la interleucina-10 (IL-10), la interleucina-1β (IL-1β), la interleucina-6 (IL-6) y el factor de necrosis tumoral α (TNF-α) se detectaron mediante kits de ELISA, que se compraron a Jiangsu Enzyme Immunoassay Industry Co., Ltd., China. Todos los procedimientos se realizaron en estricta conformidad con las instrucciones del kit.
2.6 Análisis del estado oxidativo hepático
Se midieron los niveles de glutatión sintasa (GSS), tiredoxina (Trx), NO, capacidad de eliminación de aniones superóxido, tasa de eliminación de radicales hidroxilo, POD, complejo I, NADH deshidrogenasa 5 codificada mitocondrialmente (mt-ND5) y NADH deshidrogenasa 6 codificada mitocondrialmente (mt-ND6). Todas las operaciones se llevaron a cabo en estricta conformidad con las instrucciones del kit producido por el Instituto de Biotecnología de Nanjing Jianchen, China.
2.7 Análisis de las proteínas de barrera de la mucosa yeyunal
Los niveles de claudina-1, proteína de unión estrecha 1 (ZO-1) y ocludina en la mucosa yeyunal se detectaron mediante kits de ELISA comprados a Jiangsu Enzyme Immunoassay Industry Co., Ltd., China. Todas las operaciones se llevaron a cabo estrictamente de acuerdo con las instrucciones del kit.
2.8 Cultivo celular
Las células epiteliales yeyunales porcinas (IPEC-J2) se cultivaron en DMEM/F12 completo suplementado con FBS al 10% (OPCEL, Inner Mongolia Opcel Biotechnology Co., Ltd., BS1101) y penicilina-estreptomicina al 1% (Gibco, 15140-122) en una incubadora de células con 5% de CO2 a 37 °C. Cuando las células alcanzaron el 80% de confluencia, se lavaron dos veces con PBS estéril y luego se subcultivaron o trataron con fármacos. Las proporciones de siembra celular para recipientes de cultivo de varios tamaños fueron las siguientes: 5 × 106 células/cm2 para placas de cultivo celular de 100 mm; 9 × 105 células/cm2 para placas de cultivo celular de 6 pocillos; y 3 × 104 células/cm2 para placas de cultivo celular de 96 pocillos.
2.9 Selección de la concentración de daidzeína
Para determinar la concentración óptima de taurina, las células IPEC-J2 se sembraron en placas de cultivo celular de 96 pocillos con seis repeticiones por tratamiento. Después de alcanzar aproximadamente el 80% de confluencia, las células se sometieron a dos lavados con PBS estéril. Posteriormente, se añadieron 100 μL de medio DMEM/F12 con diversas concentraciones de taurosulfonato (0, 10, 25, 50 y 100 mM, suplementados con 100 μg/mL de estreptomicina y 100 U/mL de penicilina) a cada pocillo durante un período de incubación de 24 h. Después de otra ronda de lavados con PBS, el grupo modelo recibió 100 μL del medio de tratamiento que contenía 0,6 mM deH2O2 según los métodos de Li et al. (18), mientras que el grupo en blanco fue tratado con el mismo volumen del medio de tratamiento sinH2O2. La evaluación de la viabilidad celular se llevó a cabo mediante el ensayo CCK8 después de 1 h de incubación.
2.10 Mediciones de la actividad de T-AOC, SOD y CAT y del contenido de MDA
Las células se cultivaron y trataron en placas de 6 pocillos. Las células cultivadas al 80% de confluencia se preincubaron con el medio de tratamiento, con o sin 25 mM de taurina, durante 24 h. Posteriormente, las células fueron expuestas a 0,6 mM H2O2 durante 1 h, cosechadas y analizadas siguiendo las directrices operativas de los kits bioquímicos para T-AOC (G0115W), SOD (G0101W), CAT (G0105W) y MDA (G0109W) proporcionados por Suzhou Grandis Biotechnology Co., Ltd., China.
2.11 RT-qPCR
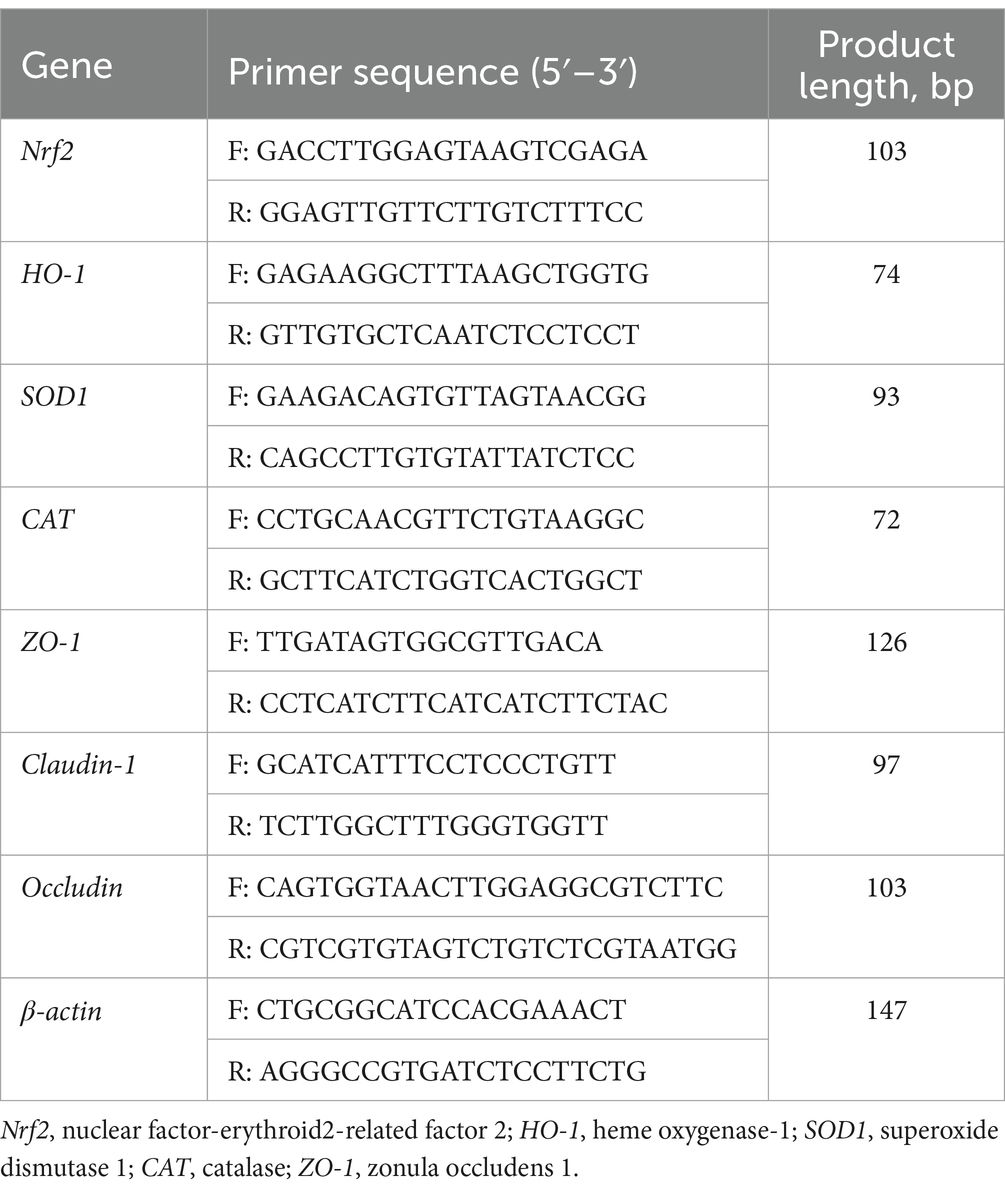
Las células se clasificaron en cuatro grupos en función de la presencia o ausencia de 25 mM de taurina y 0,6 mM deH2O2 según el método de cultivo descrito en la sección 2.10. Una vez finalizado el cultivo celular, las células se cosecharon utilizando un raspador celular, seguido de la extracción total de ARN mediante el método TRIzol. Posteriormente, se analizaron los niveles de expresión de ARNm de los genes Nrf2, HO-1, CAT, SOD, ZO-1, ocludina y claudina-1 mediante PCR cuantitativa en tiempo real. Los cebadores específicos para porcinos utilizados en este estudio se diseñaron a medida (Tabla 2). Los ciclos de PCR y los ensayos de expresión relativa se adhirieron a la metodología descrita en nuestro estudio de investigación previo realizado por Yin et al. (19).
2.12 Análisis estadístico
En los experimentos con animales, se utiliza un diseño aleatorio de un solo factor, en el que cada cerdo sirve como unidad estadística designada. Los datos distribuidos normalmente se sometieron a ANOVA de un solo factor, junto con comparaciones de grupos por pares mediante el método de Duncan. Se implementa el análisis de contraste polinómico para evaluar los impactos lineales y cuadráticos de los aditivos. Se realizó una prueba no paramétrica de Kruskal-Wallis si los datos se desviaban de una distribución normal, seguida de una prueba de Dunn post hoc para comparaciones intergrupales. Los gráficos se generaron utilizando el software GraphPad Prism 9.0.0. La significación estadística se determinó a nivel de p < 0,05. Los resultados se presentan como las medias y los errores estándar de las medias (SEM).
3 Resultados
3.1 Rendimiento del crecimiento
El impacto de la taurina en el rendimiento del crecimiento de los lechones destetados se detalla en la Tabla 3. En comparación con los del grupo control, la inclusión de 0,2 y 0,3% de taurina en la dieta mejoró significativamente el peso final medio y la GMD de los lechones, al tiempo que redujo notablemente la F/G. El peso final y la GMD de los lechones aumentaron proporcionalmente a la concentración de taurina, mientras que la relación F/G disminuyó linealmente. Además, la suplementación de las dietas de los lechones destetados con 0,1, 0,2% o 0,3% de taurina condujo a una reducción significativa en la incidencia de diarrea, con efectos lineales y cuadráticos.
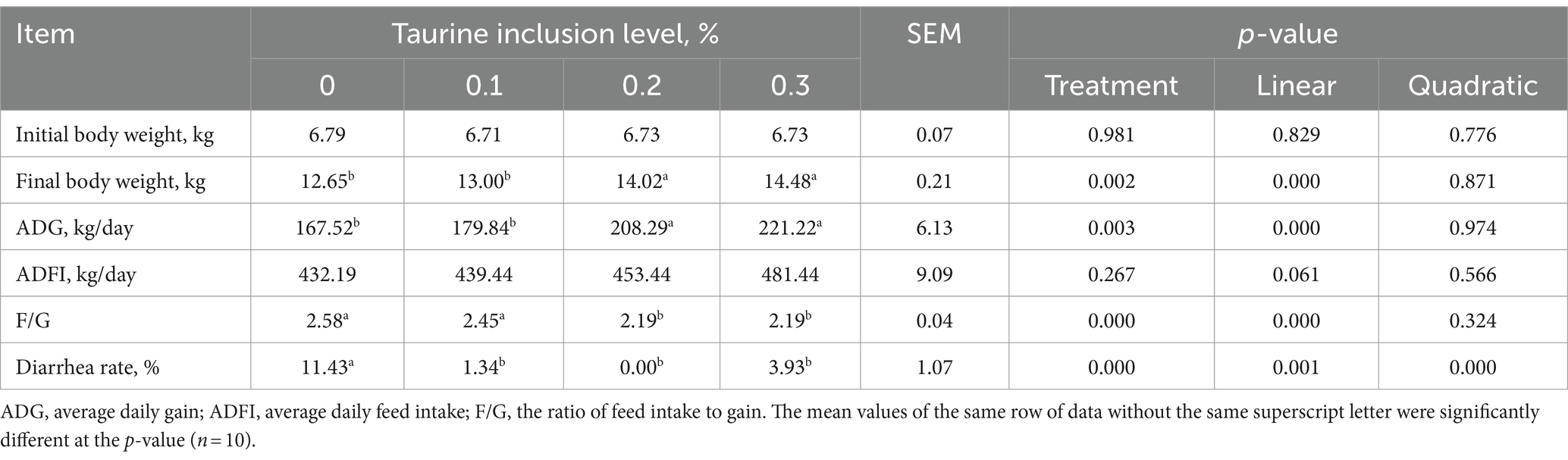
Tabla 3. Efecto de la taurina en el rendimiento de crecimiento de lechones destetados.
3.2 Parámetros antioxidantes séricos
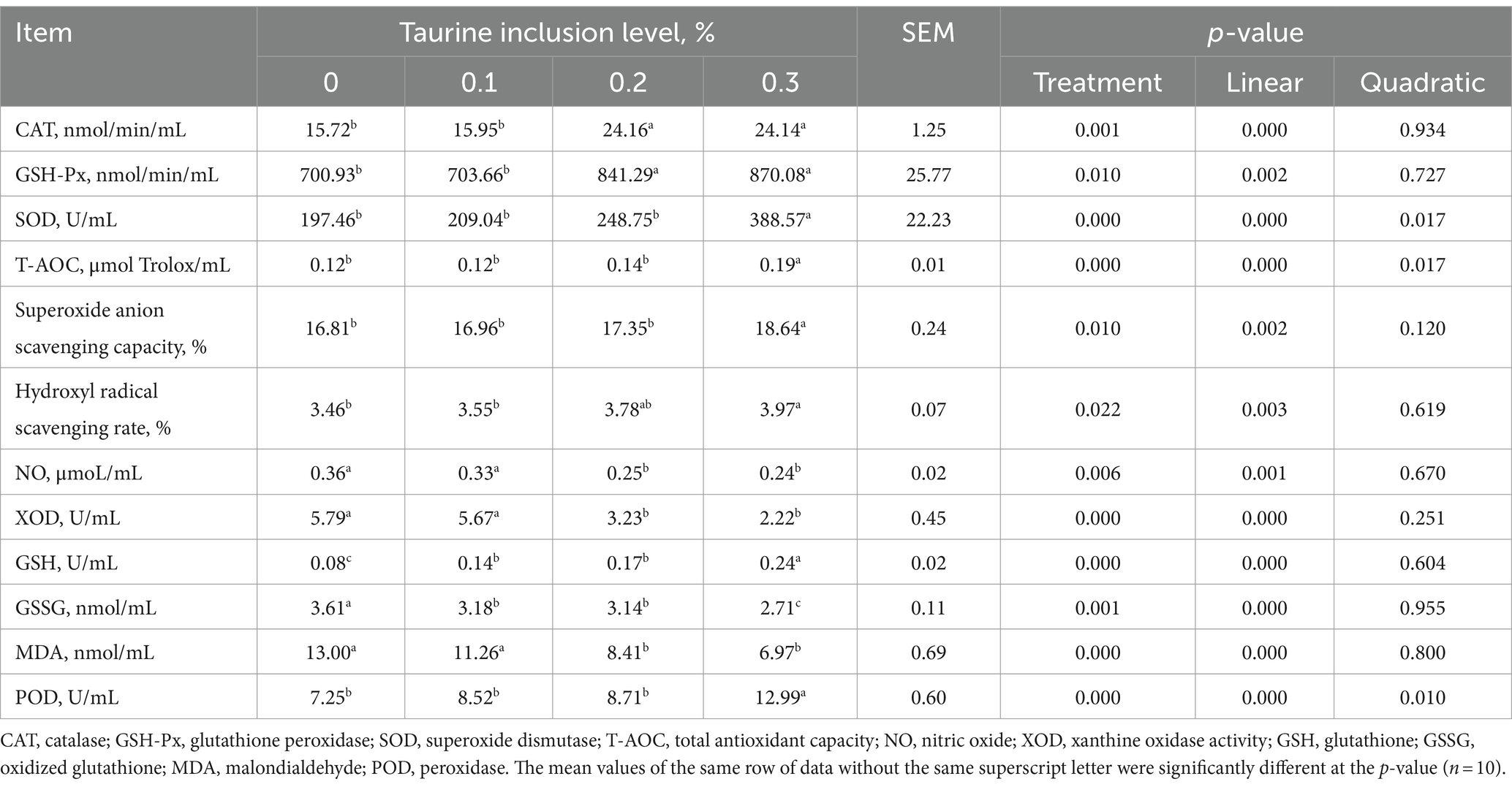
El análisis presentado en la Tabla 4 demuestra que la suplementación de las dietas de los lechones destetados con 0,1, 0,2 y 0,3% de taurina resultó en un aumento de los niveles séricos de GSH en comparación con los del grupo control. La inclusión de 0,2 y 0,3% de taurina condujo a una mayor actividad sérica de CAT y a una reducción de los niveles séricos de NO, XOD, GSSG y MDA en los lechones destetados. Además, la introducción de taurina al 0,3% mejoró las actividades de GSH-Px y SOD en el suero de lechones y aumentó la T-AOC y la POD, la capacidad de eliminación de aniones superóxido y la tasa de eliminación de radicales hidroxilo. El estudio también reveló un aumento lineal en CAT, GSH-Px, SOD, T-AOC, la capacidad de eliminación de aniones superóxido, la tasa de eliminación de radicales hidroxilo, GSH y POD con el aumento de la concentración de taurina. Por el contrario, hubo disminuciones lineales en los niveles de NO, XOD, GSSG y MDA.
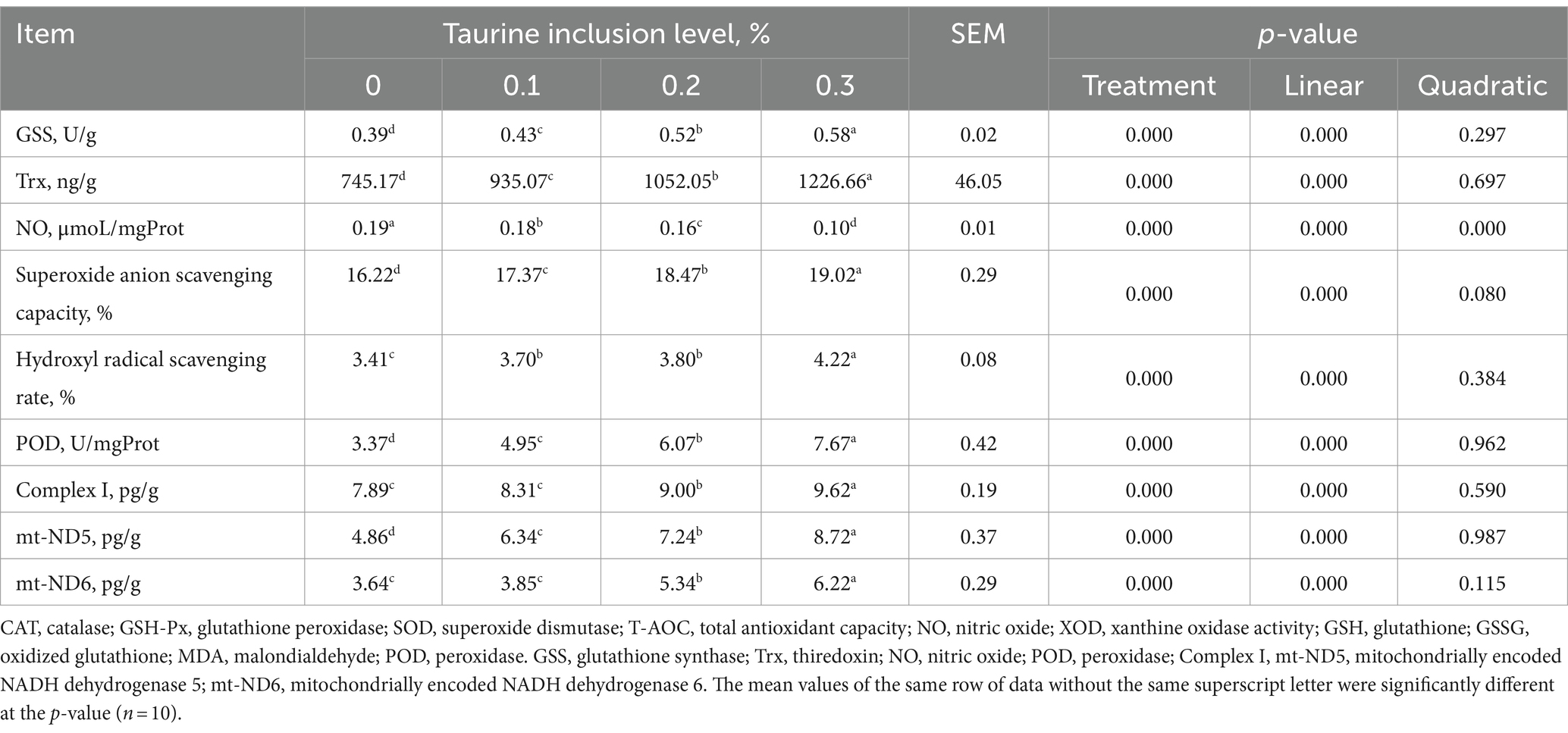
3.3 Parámetros antioxidantes hepáticos
Los hallazgos presentados en la Tabla 5 ilustran el impacto de la taurina sobre la capacidad antioxidante del hígado de lechones destetados. En comparación con los del grupo de control, la suplementación dietética con taurina al 0,1, 0,2 y 0,3% dio lugar a un aumento de los niveles de GSS, Trx, POD y mt-ND5 en el hígado; mayor capacidad de eliminación de aniones superóxido; y una elevada tasa de aclaramiento libre de hidroxilo, al tiempo que reduce los niveles de NO hepático en los lechones destetados. Además, la administración de taurina al 0,2 y al 0,3% condujo a un aumento de los niveles de complejo hepático I y mt-ND6 en lechones destetados. El presente estudio también reveló un aumento lineal en GSS, Trx, la capacidad de eliminación de aniones superóxido, la tasa de eliminación de radicales hidroxilo, POD, complejo I, mt-ND5 y mt-ND6 con concentraciones crecientes de taurina y una disminución lineal en los niveles de NO.
3.4 Parámetros inmunitarios séricos
La Tabla 6 muestra que la suplementación con 0,1, 0,2 y 0,3% de taurina en la dieta de los lechones destetados condujo a una reducción significativa de los niveles séricos de IL-6 y un aumento notable de los niveles de IgA e IgG en comparación con los del grupo control. Además, la adición de 0,2 y 0,3% de taurina elevó significativamente el nivel sérico de IL-10, mientras que la inclusión de taurina al 0,3% aumentó notablemente los niveles séricos de IL-4 e IgM. En particular, los niveles de IL-4, IL-10, IgA, IgG e IgM mostraron patrones de crecimiento lineales y cuadráticos en correlación con aumentos en la concentración de taurina, mientras que el nivel de IL-6 disminuyó linealmente.
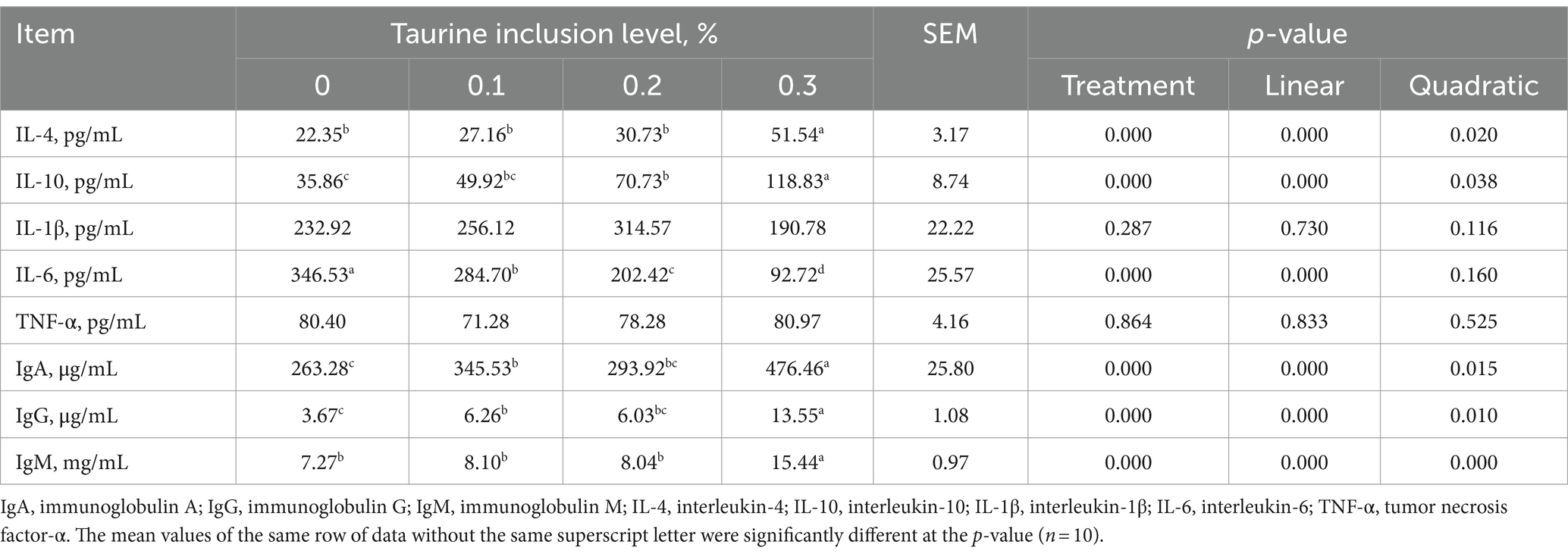
Tabla 6. Efecto de la taurina sobre los índices inmunes séricos en lechones destetados.
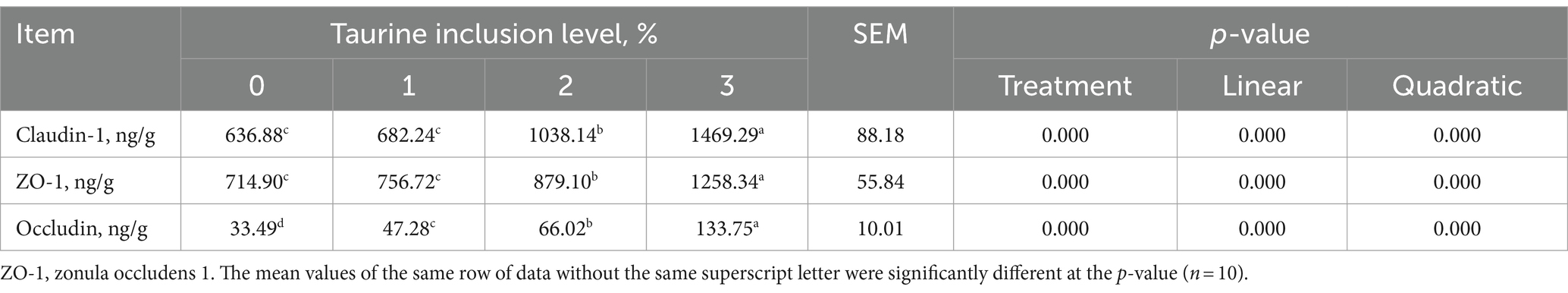
3.5 Proteínas de barrera de la mucosa yeyunal
La Tabla 7 muestra que, en comparación con los del grupo control, la suplementación de las dietas de los lechones destetados con 0,1, 0,2 y 0,3% de taurina condujo a un aumento notable de los niveles de ocludina en la mucosa yeyunal. Además, la inclusión de taurina al 0,2 y al 0,3% elevó notablemente los niveles de claudina y ZO-1. Al mismo tiempo, los niveles de ocludina, claudina y ZO-1 mostraron aumentos lineales y cuadráticos proporcionales al aumento de la concentración de taurina.
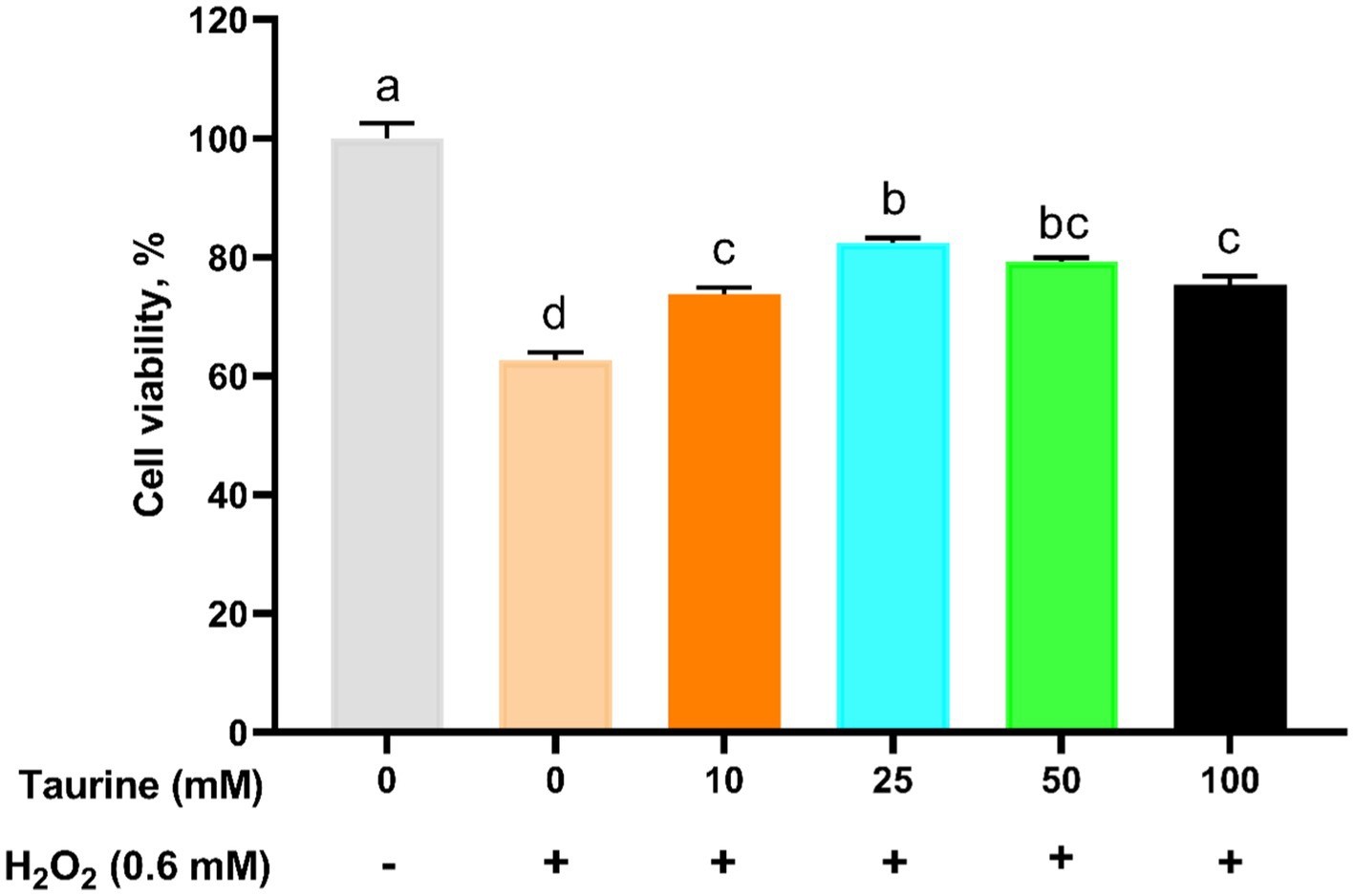
3.6 La concentración de taurina en las células IPEC-J2
Determinar los parámetros óptimos de tratamiento para la taurina. Las células se trataron previamente con varias concentraciones de taurina durante 24 h y posteriormente se expusieron o no a 0,6 mM H2O2 durante 1 h. Las evaluaciones de viabilidad celular revelaron que la tasa de viabilidad más alta ocurrió en el grupo de tratamiento con taurina de 25 mM (Figura 1). En consecuencia, seleccionamos 25 mM como la concentración óptima para todos los experimentos posteriores.
3.7 Actividad de SOD, actividad de CAT, contenido de T-AOC y MDA en células IPEC-J2 tratadas conH2O2
Este estudio se centró en investigar el impacto de la taurina en la actividad de las enzimas antioxidantes y el contenido de MDA en células IPEC-J2 expuestas a la catalasa. Para el análisis se utilizaron kits bioquímicos (Figura 2). La exposición aH2O2 disminuyó significativamente las actividades de CAT y SOD, así como de T-AOC, al tiempo que aumentó notablemente el contenido de MDA en las células en comparación con las del grupo control. Como era de esperar, el pretratamiento de 25 mM durante 24 h no solo aumentó significativamente la capacidad antioxidante de las células, sino que también mejoró el daño por estrés oxidativo causado por la exposición alH2O2. Este pretratamiento condujo a un aumento considerable de las actividades de CAT y SOD y T-AOC, junto con una reducción del contenido de MDA.
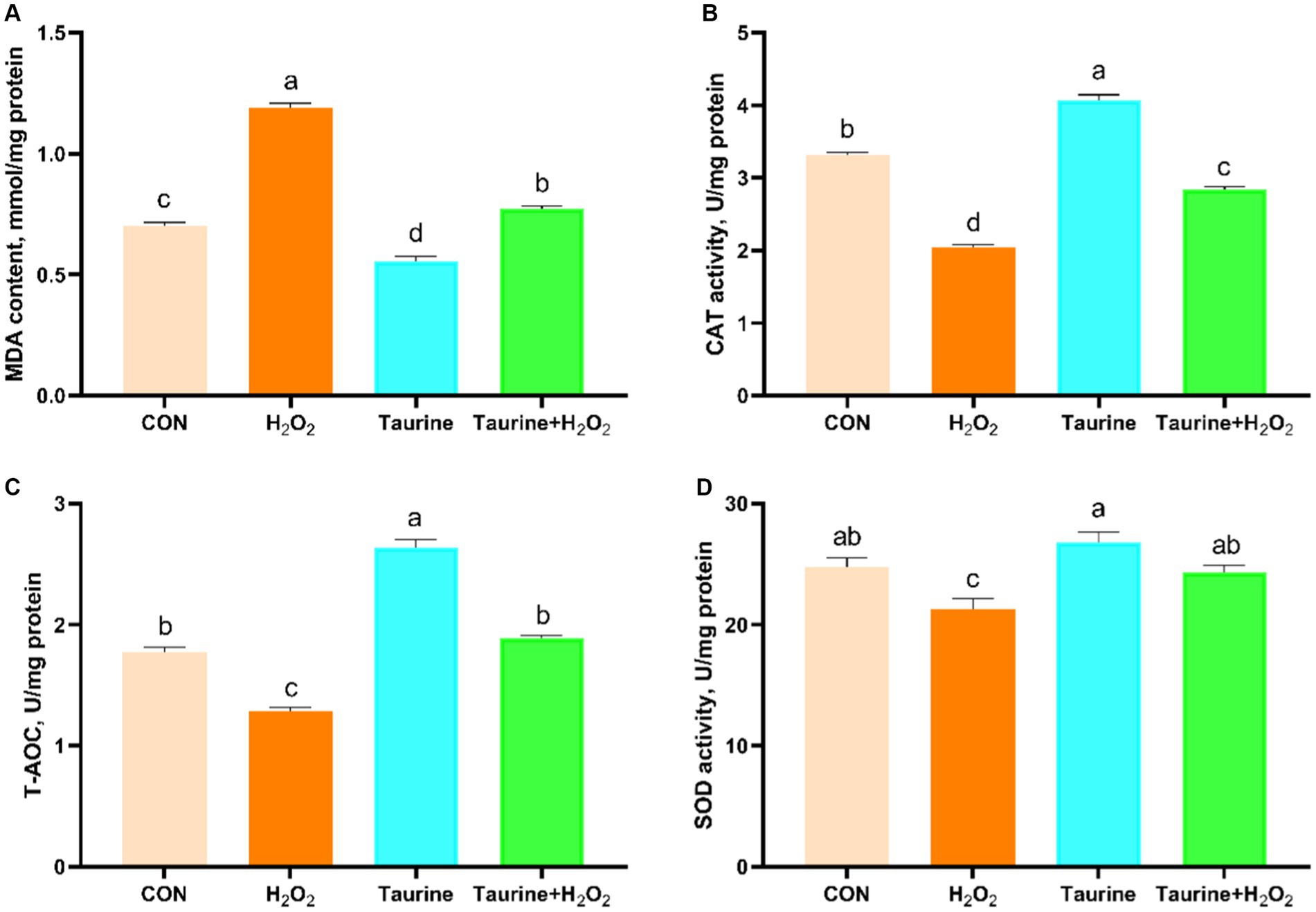
3.8 Efecto de la taurina sobre la expresión de genes implicados en la vía de señalización Nrf2 en células IPEC-J2
La vía de señalización Nrf2 sirve como un mecanismo regulador crucial para combatir el estrés oxidativo. Nuestro estudio exploró el impacto del pretratamiento con taurina en los niveles de expresión de los genes de la vía Nrf2 en células IPEC-J2 expuestas aH2O2 (Figura 3). En comparación con los del grupo control, la expresión de los genes Nrf2, HO-1 y CAT en el grupo tratado conH2O2fue notablemente menor. Por el contrario, el pretratamiento con 25 mM de taurina durante 24 h aumentó sustancialmente la expresión de los genes Nrf2, HO-1 y CAT, lo que condujo a una mejora significativa del daño oxidativo causado por la exposición aH2O2.
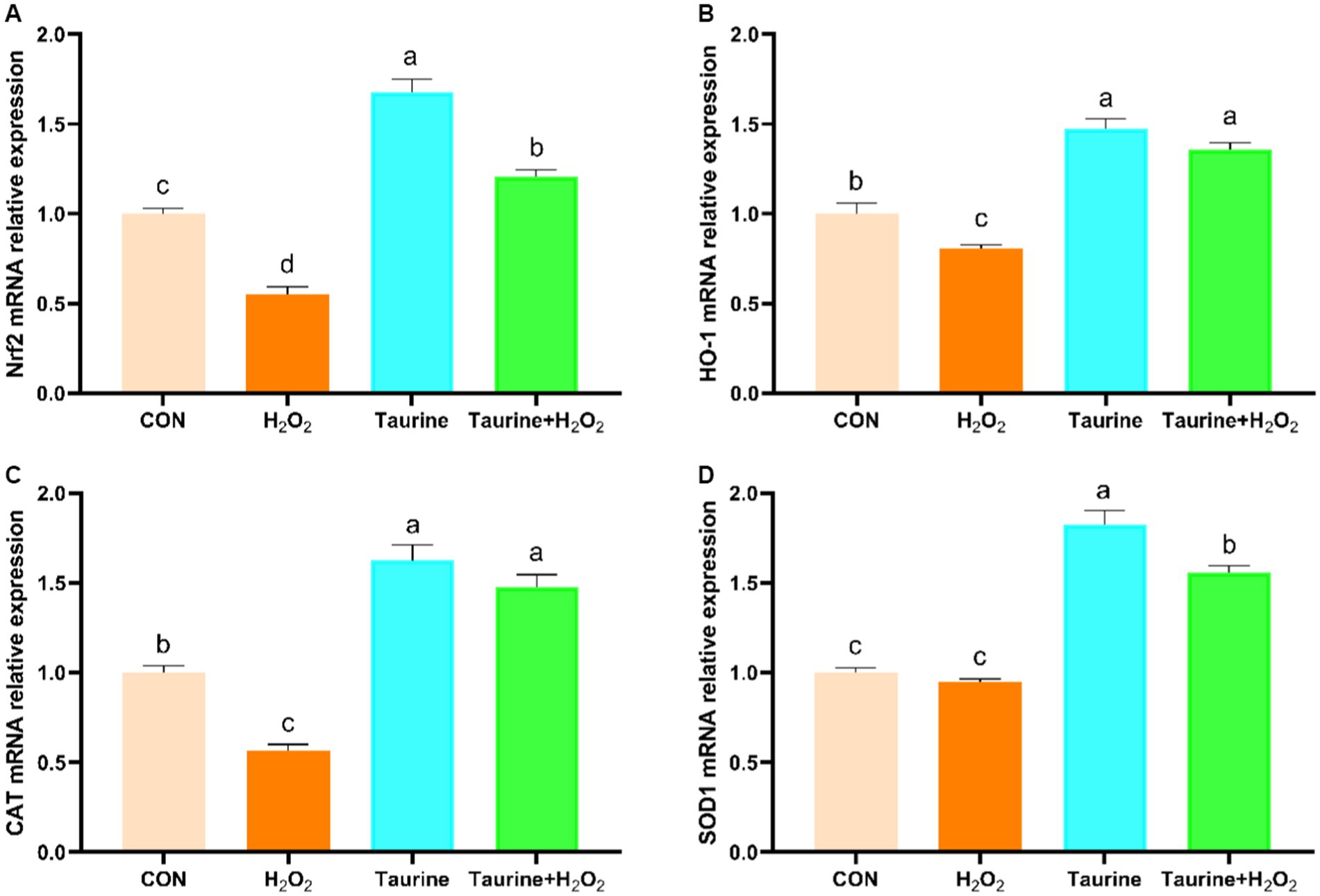
3.9 Efecto de la taurina sobre la expresión génica relacionada con la función de barrera en células IPEC-J2 expuestas aH2O2
En la Figura 4 se ilustra el impacto del pretratamiento con taurina en los niveles de expresión de genes relacionados con la unión estrecha en células IPEC-J2 expuestas aH2O2. En comparación con los del grupo de control, la exposición aH2O2 disminuyó notablemente la expresión de los genes ZO-1, ocludina y claudina-1. Por el contrario, el pretratamiento con 25 mM de taurina durante 24 h reguló significativamente la expresión de los genes ZO-1, ocludina y claudina-1, revirtiendo efectivamente la regulación negativa inducida porH2O2.
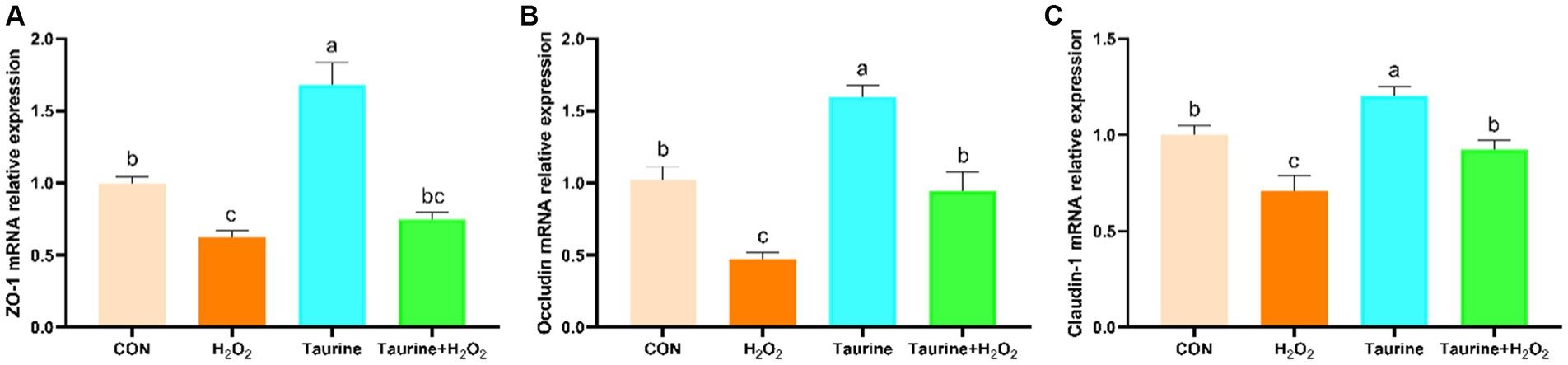
4 Discusión
El estrés del destete provoca diarrea, reducción del rendimiento del crecimiento, compromiso de la función intestinal y otros efectos nocivos, lo que resulta en notables pérdidas económicas para la industria de la cría de cerdos (20, 21). Este estudio tuvo como objetivo investigar los beneficios potenciales de la taurina sobre el rendimiento del crecimiento, el estrés oxidativo y la barrera intestinal de lechones destetados. En la actualidad, la suplementación de las dietas con antioxidantes es una estrategia eficaz para mitigar el estrés oxidativo y mejorar el rendimiento del crecimiento y el estado de salud de los lechones destetados (7). Además, investigaciones anteriores han indicado que los antioxidantes pueden servir como alternativas a los antibióticos para facilitar el destete suave de los lechones (22). Nuestros hallazgos revelaron que la adición de taurina a las dietas de los lechones destetados, particularmente a los niveles de 0,2 y 0,3%, aumentó significativamente tanto el peso final promedio como la ganancia diaria de los lechones, al tiempo que redujo la relación alimento-carne. De acuerdo con los hallazgos de un estudio reciente de Wang et al. (23), la suplementación con taurina al 0,2 y 0,4% mejoró notablemente el rendimiento del crecimiento de los lechones destetados. Además, estudios previos han sugerido que la taurina podría mitigar los impactos adversos del LPS en el rendimiento del crecimiento de lechones y pollos de engorde (16, 24). Durante el proceso de destete, los lechones a menudo experimentan diarrea debido a diversos factores estresantes externos y un desarrollo incompleto. Cada año, un número considerable de lechones destetados sucumben o son sacrificados debido a diarrea relacionada con el estrés del destete, lo que resulta en pérdidas significativas en el sector de la cría (3, 25). Nuestra investigación mostró que la taurina disminuyó efectivamente la tasa de diarrea en lechones destetados en comparación con la del grupo de control. Estudios posteriores revelaron que la taurina mejoró los niveles de estrés oxidativo y la función de barrera intestinal de los lechones, lo que corrobora aún más el potencial de la taurina como sustituto antioxidante de los antibióticos alimentarios para mejorar el rendimiento del crecimiento y la diarrea de los lechones destetados.
El estrés del destete no solo está estrechamente relacionado con posibles cambios en el sistema inmunológico y la función de la barrera intestinal, sino que también resulta en un estrés oxidativo severo (26). Las reacciones redox del cuerpo subyacen a varias vías bioquímicas y funciones celulares, que dependen principalmente del delicado equilibrio entre los sistemas oxidativo y antioxidante. Un desequilibrio debido a la producción excesiva de ROS o a la eliminación inadecuada de las ROS por parte de los antioxidantes induce estrés oxidativo, lo que conduce a la apoptosis celular, daño tisular, trastornos metabólicos, inflamación, diarrea y reducción del rendimiento productivo en los lechones (27, 28). Los estudios de Wen et al. (29) destacaron un aumento notable en los niveles de malondialdehído (MDA) en la sangre de lechones destetados, junto con una disminución de las actividades de SOD y GSH-Px. La taurina puede eliminar las especies reactivas de oxígeno y mitigar la peroxidación lipídica. Los niveles de SOD, GSH-Px y MDA en la sangre y los tejidos son biomarcadores comunes de estrés oxidativo. El MDA es un metabolito de la peroxidación autolipídica, y un aumento en los niveles de GSH y SOD refleja un aumento en la capacidad antioxidante del huésped. Nuestra investigación demostró que la suplementación con taurina mejora la actividad sérica de GSH-Px y los niveles de GSH y mejora las capacidades de eliminación de aniones superóxido y radicales hidroxilo, al tiempo que reduce los niveles séricos de NO, XOD y GSSG. Además, el estrés oxidativo está estrechamente relacionado con la ferroptosis, donde tanto GSH-Px como GSH desempeñan un papel crucial como reguladores al facilitar la reducción de ROS (30, 31). En conclusión, nuestros hallazgos se alinean con investigaciones previas que sugieren que la taurina mejora la capacidad antioxidante sérica de los lechones destetados.
El hígado, un órgano esencial, realiza varias funciones vitales, incluida la desintoxicación, el metabolismo, la secreción de bilis y la defensa inmunológica. Exhibe una alta sensibilidad a las sustancias exógenas, lo que lo hace susceptible al estrés oxidativo inducido por medicamentos, virus o toxinas. El estrés del destete puede desencadenar estrés oxidativo en el hígado de los lechones (33, 34). Investigaciones anteriores han demostrado que el destete disminuye las actividades de SOD, CAT y GSH-PX en el hígado de los lechones (35). Numerosos estudios avalan el papel protector de la taurina en la lesión hepática. En nuestro estudio, la taurina aumentó los niveles de GSS, Trx, POD, complejo I, mt-nd5 y mt-nd6 en el hígado de lechones destetados. Mejoró la capacidad de eliminación de aniones superóxido y la tasa de eliminación de radicales libres de hidroxilo y redujo notablemente los niveles de NO en el hígado. Wu et al. (36) informaron que la taurina inhibe el aumento de MDA y la disminución de la actividad de las enzimas antioxidantes en hígados de rata inducidos por la aflatoxina B1. Shi et al. (37) informaron de la eficacia de la taurina para aliviar el daño oxidativo hepático del aceite de pescado oxidado en bagres jóvenes. Nuestros hallazgos se alinean con estos estudios, sugiriendo el potencial de la taurina para mejorar la capacidad antioxidante de los hígados de los lechones destetados.
El estrés del destete suele provocar un compromiso de la función inmunitaria y una respuesta inflamatoria (38). Durante el destete, los lechones están expuestos a bacterias, toxinas y antígenos de la cavidad intestinal, que pueden infiltrarse en tejidos, órganos y torrente sanguíneo a través de la submucosa, lo que provoca alteraciones en el sistema inmunitario intestinal y desencadena inflamación (39, 40). El exceso de citocinas proinflamatorias puede exacerbar el daño a la integridad intestinal y la función epitelial. Los investigadores estudiaron la expresión génica de citocinas inflamatorias en lechones destetados y observaron un aumento significativo en los niveles de IL-6 en el tracto intestinal, al igual que los niveles de IL-1β y TNF-α (41, 42). La taurina exhibe propiedades antiinflamatorias y contribuye significativamente al manejo de enfermedades cardiovasculares y afecciones inflamatorias metabólicas como la diabetes mellitus y la enfermedad del hígado graso no alcohólico (15, 43, 44). Los neutrófilos son los principales contribuyentes al daño tisular en la mastitis. Los estudios han demostrado que la taurina puede mitigar la agregación de neutrófilos, dificultar la expresión de cxcl2 y aliviar la mastitis causada por Streptococcus uberis en vacas lecheras (45). Además, la investigación ha demostrado que la suplementación con taurina puede mejorar la hinchazón de las células hepáticas y la infiltración inflamatoria en lechones afectados por micotoxinas, reduciendo los niveles séricos de los factores proinflamatorios IL-1β, IL-6, IL-8 y TNF-α (46). Nuestro estudio corrobora estos hallazgos, demostrando que la incorporación de taurina en las dietas de lechones destetados aumenta los niveles séricos de IgA, IgG, IgM, IL-4 e IL-10 mientras reduce los niveles de IL-6. Este hallazgo sugiere que la taurina desempeña un papel crucial en la regulación inmunitaria, contrarrestando el deterioro de la función inmunitaria en lechones destetados debido al estrés del destete.
El yeyuno sirve como un órgano vital para la absorción de nutrientes del entorno externo y desempeña un papel crucial en la comunicación con el medio interno y externo (47, 48). Debido a su sensibilidad a factores internos y externos, el yeyuno puede experimentar estrés oxidativo, lo que provoca anomalías en su morfología y estructura, daños en las funciones de barrera y capacidades de digestión y absorción comprometidas (49). En consecuencia, el daño oxidativo a la mucosa yeyunal está estrechamente relacionado con el desarrollo de afecciones como el retraso del crecimiento y la diarrea en el ganado y las aves de corral (12, 50). En este estudio, la suplementación de lechones destetados con taurina elevó significativamente los niveles de claudina, ZO-1 y ocludina en la mucosa yeyunal. Investigaciones previas de Shi et al. (37) demostraron que la incorporación de taurina en una dieta que contenía aceite de pescado oxidado regulaba notablemente la expresión intestinal de ARNm de ZO-1 y ZO-2 en plántulas de bagre. Además, Zhao et al. (24) informaron que la taurina podría aumentar la expresión proteica de ZO-1, ocludina y claudina-1 en el colon de lechones desafiados con LPS. Estos hallazgos se alinean con los del presente estudio, reforzando la noción de que la taurina tiene el potencial de aliviar el deterioro de la función de barrera intestinal inducido por el estrés del destete.
La estructura intestinal está compuesta principalmente por vellosidades y criptas, que están envueltas por una sola capa de células epiteliales cilíndricas (51, 52). Estas células absorbentes, que constituyen del 90 al 95% de las células epiteliales, presentan microvellosidades apicales y albergan varias enzimas responsables de la descomposición y absorción de azúcares y proteínas (53-55). Además, las células epiteliales intestinales actúan como una interfaz crucial entre las células inmunitarias y los agentes ambientales, ayudando en la detección y respuesta a los alimentos, bacterias simbióticas y patógenos para protegerse contra la invasión de patógenos (56, 57). Por lo tanto, las células epiteliales intestinales desempeñan un papel fundamental en la función intestinal. El estrés oxidativo tiene el potencial de dañar la mucosa intestinal. En esta investigación, se utilizaron IPEC-J2 para replicar el daño oxidativo observado en la mucosa intestinal debido al estrés del destete en lechones a través de la exposición aH2O2. La presencia deH2O2 desencadena la generación de ROS y provoca daño lipídico, lo que conduce a la producción de biomarcadores de degradación lipídica (MDA). El presente estudio reveló que el pretratamiento de las células IPEC-J2 con 25 mM de taurina durante 24 h mejoró significativamente las actividades de las enzimas antioxidantes y redujo el contenido de MDA. Además, el pretratamiento con taurina durante 24 h mitigó sustancialmente el daño oxidativo inducido por la exposición a H2O2en células IPEC-J2, revirtiendo las disminuciones en las actividades de SOD y CAT y el aumento en el contenido de MDA causado por la exposición aH2O2. La SOD está ampliamente distribuida en la matriz mitocondrial de las células animales y desempeña un papel crucial en la eliminación de los radicales libres de oxígeno para proteger las células de la mucosa intestinal del daño oxidativo, lo que la convierte en un antioxidante vital en el cuerpo (58). Además, el pretratamiento con taurina durante 24 h revirtió la regulación negativa de los genes que codifican proteínas de barrera inducida porH2O2, lo que subraya el papel de la taurina en la mejora de la lesión mucosa inducida por H2O2.
La vía de señalización Nrf2, que es crucial para la respuesta al estrés oxidativo, desempeña un papel vital en el mantenimiento de la homeostasis redox del cuerpo. La producción excesiva de ROS puede impedir la expresión de Nrf2, lo que conduce a un desequilibrio redox (59, 60
). Nuestro estudio investigó la expresión de genes relacionados con la vía Nrf2 en células IPEC-J2 mediante qPCR. Observamos que la taurina mejoró significativamente la expresión de los genes Nrf2 y HO-1, revirtiendo la regulación negativa inducida por la exposición aH2O2. El HO-1, un objetivo de Nrf2, regula directamente la actividad del promotor del HO-1 a través del Nrf2, exhibiendo potentes efectos antioxidantes al eliminar las ROS y defenderse contra sustancias nocivas como peróxidos y radicales libres (61, 62). El pretratamiento con taurina también aumentó la expresión de los genes SOD y CAT en las células IPEC-J2. De acuerdo con investigaciones anteriores, la taurina se muestra prometedora para aliviar el estrés oxidativo. Los investigadores han demostrado que la taurina, a través de la activación de Nrf2, protege las células epiteliales de la glándula mamaria del cerdo del estrés oxidativo inducido porH2O2 (63). Además, los estudios de Wang et al. (64) revelaron que la taurina podría mitigar la regulación negativa de Nrf2 y HO-1 inducida por el 5-fluorouracilo, mejorando la inflamación de la mucosa en los tejidos de yeyuno y colon de ratón. Nuestros hallazgos respaldan el potencial de la taurina como suplemento dietético para combatir el estrés oxidativo.
5 Conclusión
Este estudio demostró que la taurina mejora el rendimiento del crecimiento de los lechones destetados, reduce la incidencia de diarrea, alivia el estrés oxidativo y mitiga las respuestas inflamatorias asociadas con el destete. La suplementación con taurina puede aumentar la expresión de genes y proteínas relacionadas con las uniones estrechas en el epitelio intestinal yeyunal. Además, la taurina tiene el potencial de mejorar el daño oxidativo en las células epiteliales intestinales a través de la vía Nrf2/HO-1. En conclusión, la taurina se ha convertido en un complemento alimenticio prometedor para mitigar el daño oxidativo en lechones destetados.
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán puestos a disposición por los autores, sin reservas indebidas.
Declaración de ética
Los estudios con animales fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales (202105) de la Universidad Agrícola de Hunan. Los estudios se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
MZ: Curación de datos, Análisis formal, Investigación, Metodología, Software, Redacción – borrador original, Redacción – revisión y edición. ZW: Análisis formal, Investigación, Metodología, Software, Redacción – borrador original, Redacción – revisión y edición. DD: Análisis formal, Investigación, Software, Redacción – borrador original, Redacción – revisión y edición. BW: Investigación, Software, Redacción – borrador original, Redacción – revisión y edición. XZ: Investigación, Software, Escritura – borrador original, Escritura – revisión y edición. BZ: Investigación, Software, Redacción – borrador original, Redacción – revisión y edición. CW: Investigación, Software, Escritura – borrador original, Escritura – revisión y edición. YZ: Conceptualización, Investigación, Administración de proyectos, Recursos, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declaran/n que no se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Ko, H-L, López-Vergé, S, Chong, Q, Gasa, J, Manteca, X, y Llonch, P. Comunicación corta: la socialización previa al destete y el enriquecimiento ambiental afectan el rendimiento a corto plazo después de reagruparse en cerdos criados comercialmente. Animal. (2021) 15:100115. doi: 10.1016/j.animal.2020.100115
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Douglas, SL, Edwards, SA, y Kyriazakis, I. Estrategias de manejo para mejorar el rendimiento de los cerdos de bajo peso al nacer hasta el destete y sus consecuencias a largo plazo. J Anim Sci. (2014) 92:2280–8. doi: 10.2527/jas.2013-7388
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Canibe, N, Højberg, O, Kongsted, H, Vodolazska, D, Lauridsen, C, Nielsen, TS, et al. Revisión de las medidas preventivas para reducir la diarrea post-destete en lechones. Animales. (2022) 12:2585. doi: 10.3390/ani12192585
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Li, M, Chen, L, Zhao, Y, Sun, H y Zhao, L. Investigación sobre el mecanismo de HRP para aliviar el estrés inmunológico de las células IPEC-J2 basada en el análisis de secuenciación del transcriptoma. Nuerca delantera. (2022) 9:944390. doi: 10.3389/fnut.2022.944390
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Wu, K, El Zowalaty, AE, diciendo, VI, y Papagiannakopoulos, T. Las funciones pleiotrópicas de las especies reactivas de oxígeno en el cáncer. Nat Cáncer. (2024) 5:384–99. doi: 10.1038/s43018-024-00738-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Rauf, A, Khalil, AA, Awadallah, S, Khan, SA, Abu-Izneid, T, Kamran, M, et al. Especies reactivas de oxígeno en sistemas biológicos: vías, enfermedades asociadas y posibles inhibidores: una revisión. Ciencia de la Alimentación Nutr. (2024) 12:675–93. doi: 10.1002/fsn3.3784
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Qiao, L, Dou, X, Song, X, Chang, J, Yi, H y Xu, C. Dirigirse a las mitocondrias con nutrientes antioxidantes para la prevención y el tratamiento de la diarrea postdestete en lechones. Anim Nutr. (2023) 15:275–87. doi: 10.1016/j.aninu.2023.09.002
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Xu, X, Wei, Y, Hua, H, Jing, X, Zhu, H, Xiao, K, et al. Polifenoles procedentes de Ilex latifolia Thunb. Aliviar la lesión intestinal mediante la modulación de la ferroptosis en lechones destetados bajo estrés oxidativo. Antioxidantes. (2022) 11:966. doi: 10.3390/antiox11050966
9. Yu, Z, Zhao, L, Zhao, J-L, Xu, W, Guo, Z, Zhang, A-Z, et al. El polisacárido dietético de Taraxacum mongolicum mejora el crecimiento, la respuesta inmune y el estado antioxidante en asociación con NF-κB, Nrf2 y TOR en la carpa Jian (Cyprinus carpio var. Jian). Acuicultura. (2022) 547:737522. doi: 10.1016/j.aquaculture.2021.737522
10. Niu, X-T, Sun, C, Zhao, L, Chen, X-M, Wang, G-Q y Li, M-Y. El papel principal del receptor de glucocorticoides (GR) en la astaxantina alivia el estrés inmunológico en el linfocito Channa argus. Acuicultura. (2024) 584:740637. doi: 10.1016/j.aquaculture.2024.740637
11. Nishida, Y, Nawaz, A, Hecht, K y Tobe, K. La astaxantina como nuevo regulador mitocondrial: un nuevo aspecto de los carotenoides, más allá de los antioxidantes. Nutrientes. (2021) 14:107. doi: 10.3390/nu14010107
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Wang, L, Wang, C, Peng, Y, Zhang, Y, Liu, Y, Liu, Y, et al. Avances de la investigación sobre estrategias de nutrición antiestrés en cerdos. Anim Nutr. (2023) 13:342–60. doi: 10.1016/j.aninu.2023.03.006
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Surai, PF, Earle-Payne, K, y Kidd, MT. La taurina como antioxidante natural: de los efectos antioxidantes directos a la acción protectora en varios modelos toxicológicos. Antioxidantes. (2021) 10:1876. doi: 10.3390/antiox10121876
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. García-Ayuso, D, Di Pierdomenico, J, Martínez-Vacas, A, Vidal-Sanz, M, Picaud, S, y Villegas-Pérez, MP. Taurina: un nutracéutico prometedor en la prevención de la degeneración de la retina. Neural Regen Res. (2024) 19:606–10. doi: 10.4103/1673-5374.380820
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Bae, M, Ahmed, K y Yim, J-E. Efectos beneficiosos de la taurina sobre los parámetros metabólicos en animales y humanos. J Obes Metab Syndr. (2022) 31:134–46. doi: 10.7570/jomes21088
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Han, H, Zhang, J, Chen, Y, Shen, M, Yan, E, Wei, C, et al. La suplementación dietética con taurina atenúa las respuestas inflamatorias inducidas por lipopolisacáridos y el estrés oxidativo de los pollos de engorde a una edad temprana. J Anim Sci. (2020) 98:98. doi: 10.1093/jas/skaa311
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Wu, S, Li, J, Deng, Y, Fang, P, Lei, W, Luo, A, et al. Efectos de los polifenoles de manzana y taurina sobre el rendimiento del crecimiento, la morfología de los tejidos y el metabolismo de los lípidos y la glucosa en anguila de arrozal (Monopterus albus) alimentada con aceite de pescado de alta oxidación. Aquac Nutr. (2023) 2023:4912141. doi: 10.1155/2023/4912141
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Li, Y, Jiang, X, Cai, L, Zhang, Y, Ding, H, Yin, J, et al. Efectos de la daidzeína sobre la capacidad antioxidante en cerdos destetados y células IPEC-J2. Anim Nutr. (2022) 11:48–59. doi: 10.1016/j.aninu.2022.06.014
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Yin, J, Li, Y, Tian, Y, Zhou, F, Ma, J, Xia, S, et al. La microbiota derivada del cerdo obeso de Ningxiang reconfigura el metabolismo de la carnitina para promover la deposición de ácidos grasos musculares en cerdos magros DLY. Innovación. (2023) 4:100486. doi: 10.1016/j.xinn.2023.100486
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Li, Y, Shi, P, Yao, K, Lin, Q, Wang, M, Hou, Z, et al. Diarrea inducida por una absorción insuficiente de grasa en lechones destetados: causas y regulación nutricional. Anim Nutr. (2024) 16:299–305. doi: 10.1016/j.aninu.2023.12.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Zhao, X, Pang, J, Zhang, W, Peng, X, Yang, Z, Bai, G, et al. Metabolismo del triptófano y diarrea de lechones: dónde estamos y los desafíos futuros. Anim Nutr. (2024) 17:123–33. doi: 10.1016/j.aninu.2024.03.005
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Li, Q, Zheng, T, Ding, H, Chen, J, Li, B, Zhang, Q, et al. Explorando los beneficios de los probióticos en la inflamación intestinal y la diarrea, desde una perspectiva antioxidante. Antioxidantes. (2023) 12:1342. doi: 10.3390/antiox12071342
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Wang, L, Jiang, L, Chu, Y, Feng, F, Tang, W, Chen, C, et al. La taurina dietética mejora el rendimiento del crecimiento y la salud intestinal a través del sistema antioxidante GSH/GSSG y la vía de señalización Nrf2/ARE en lechones destetados. Antioxidantes. (2023) 12:1852. doi: 10.3390/antiox12101852
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Zhao, D-D, Gai, Y-D, Li, C, Fu, Z-Z, Yin, D-Q, Xie, M, et al. Efecto de la taurina dietética sobre la función de barrera intestinal, la microbiota colónica y los metabolitos en lechones destetados inducidos por LPS. Microbiol frontal. (2023) 14:1259133. doi: 10.3389/fmicb.2023.1259133
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Yin, Z, Wang, K, Liu, Y, Li, Y, He, F, Yin, J, et al. Lactobacillus johnsonii mejora la función de barrera intestinal y reduce la diarrea post-destete en lechones: participación del sistema endocannabinoide. Animales. (2024) 14:493. doi: 10.3390/ani14030493
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Correa, F, Luise, D, Palladino, G, Scicchitano, D, Brigidi, P, Martelli, PL, et al. Influencia de la gravedad de las lesiones corporales en el estado oxidativo y la microbiota intestinal de cerdos destetados. Animal. (2023) 17:100818. doi: 10.1016/j.animal.2023.100818
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Yu, J, Zheng, C, Zheng, J, Duan, G, Guo, Q, Zhang, P, et al. Desarrollo de lesiones intestinales y restauración de lechones destetados bajo estrés inmunológico crónico. Antioxidantes. (2022) 11:2215. doi: 10.3390/antiox11112215
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Wang, M, Huang, H, Wang, L, Yin, L, Yang, H, Chen, C, et al. El ácido tánico atenúa el daño oxidativo intestinal mejorando la capacidad antioxidante y la barrera intestinal en lechones destetados y células IPEC-J2. Nuerca delantera. (2022) 9:1012207. doi: 10.3389/fnut.2022.1012207
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Wen, C, Li, F, Guo, Q, Zhang, L, Duan, Y, Wang, W, et al. Efectos protectores de la taurina frente al daño muscular inducido por el diquat en lechones destetados a 35 días. J Anim Sci Biotechnol. (2020) 11:56. doi: 10.1186/s40104-020-00463-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Li, M-Y, Shi, Y-C, Xu, W-X, Zhao, L y Zhang, A-Z. Explorando la lesión de la barrera hematoencefálica inducida por Cr(VI) y la neurotoxicidad en el pez cebra y el pez cabeza de serpiente, e inhibiendo los efectos tóxicos de la astaxantina. Contaminación ambiental. (2024) 355:124280. doi: 10.1016/j.envpol.2024.124280
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Zhao, L, Duan, H, Liu, Y, Wang, Y, Li, M y Li, M. Exposición a largo plazo del pez cebra (Danio rerio) al Cr(VI): toxicidad reproductiva y neurotoxicidad. Reg Stud Mar Sci. (2024) 74:103559. doi: 10.1016/j.rsma.2024.103559
32. Tan, Y, An, K y Su, J. Revisión: mecanismo de los herbívoros que metabolizan sinérgicamente plantas tóxicas a través de la microbiota hepática e intestinal. Comp Biochem Physiol C. (2024) 281:109925. doi: 10.1016/j.cbpc.2024.109925
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Yu, C, Wang, D, Shen, C, Luo, Z, Zhang, H, Zhang, J, et al. Remodelación del metabolismo hepático de la glucosa en respuesta al destete precoz en lechones. Animales. (2024) 14:190. doi: 10.3390/ani14020190
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Cai, L, Ming, D, Chen, W, Zhao, Y, Li, Y, Sun, W, et al. La silibina alivió la lesión hepática al regular el equilibrio redox, la respuesta inflamatoria y la función mitocondrial en lechones destetados bajo estrés oxidativo inducido por paraquat. Antioxidantes. (2024) 13:324. doi: 10.3390/antiox13030324
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Yu, L, Li, H, Peng, Z, Ge, Y, Liu, J, Wang, T, et al. El destete precoz afecta a la función antioxidante del hígado en los lechones. Animales. (2021) 11:2679. doi: 10.3390/ani11092679
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Wu, G, San, J, Pang, H, Du, Y, Li, W, Zhou, X, et al. La taurina atenúa la lesión hepática inducida por AFB1 al aliviar el estrés oxidativo y regular la apoptosis mediada por mitocondrias. Toxicon. (2022) 215:17–27. doi: 10.1016/j.toxicon.2022.06.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Shi, Y, Hu, Y, Wang, Z, Zhou, J, Zhang, J, Zhong, H, et al. El efecto protector de la taurina sobre el estrés oxidativo hepático inducido por el aceite de pescado oxidado y el deterioro de la función de barrera intestinal en juveniles de Ictalurus punctatus. Antioxidantes. (2021) 10:1690. doi: 10.3390/antiox10111690
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Liu, B, Jiang, X, Cai, L, Zhao, X, Dai, Z, Wu, G, et al. La putrescina mitiga la atrofia intestinal mediante la supresión de la respuesta inflamatoria en lechones destetados. J Anim Sci Biotechnol. (2019) 10:69. doi: 10.1186/s40104-019-0379-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Tang, X, Xiong, K, Fang, R y Li, M. Estrés por destete y salud intestinal de los lechones: una revisión. Frente Immunol. (2022) 13:1042778. doi: 10.3389/fimmu.2022.1042778
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Huting, AM, Middelkoop, A, Guan, X y Molist, F. Uso de estrategias nutricionales para dar forma al tracto gastrointestinal de lechones lactantes y destetados. Animales. (2021) 11:402. doi: 10.3390/ani11020402
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Tang, W, Wei, Y, Ni, Z, Hou, K, Luo, XM y Wang, H. El control mediado por IgA de la interacción huésped-microbiano durante la reacción de destete influye en la inflamación intestinal. Microbios intestinales. (2024) 16:2323220. doi: 10.1080/19490976.2024.2323220
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Yan, H, Xing, Q, Xiao, X, Yu, B, He, J, Mao, X, et al. Efecto de los postbióticos y el aceite esencial de Saccharomyces cerevisiae sobre el rendimiento del crecimiento y la salud intestinal de cerdos destetados durante la infección por K88 ETEC. J Anim Sci. (2024) 102:skae007. doi: 10.1093/jas/skae007
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Qaradakhi, T, Gadanec, LK, McSweeney, KR, Abraham, JR, Apostolopoulos, V, y Zulli, A. El efecto antiinflamatorio de la taurina en las enfermedades cardiovasculares. Nutrientes. (2020) 12:2847. doi: 10.3390/nu12092847
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Ouyang, G, Wang, N, Tong, J, Sun, W, Yang, J y Wu, G. Alivio de la taurina en la lesión hepática de ratas diabéticas tipo 2 mediante la mejora de la capacidad antioxidante y antiinflamatoria. Heliyon. (2024) 10:e28400. doi: 10.1016/j.heliyon.2024.e28400
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Li, M, Wang, Z, Fu, S, Sun, N, Li, W, Xu, Y, et al. La reducción taurina de las lesiones por infiltración de neutrófilos mejora la mastitis inducida por Streptococcus uberis. Int Immunopharmacol. (2023) 124:111028. doi: 10.1016/j.intimp.2023.111028
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Ji, X, Tang, Z, Zhang, F, Zhou, F, Wu, Y y Wu, D. La suplementación dietética con taurina contrarresta la lesión hepática inducida por el deoxinivalenol al aliviar el estrés oxidativo, la disfunción mitocondrial, la apoptosis y la inflamación en lechones. Ecotoxicol Environ Saf. (2023) 253:114705. doi: 10.1016/j.ecoenv.2023.114705
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Rao, MC. Fisiología del transporte de electrolitos en el intestino: implicaciones para la enfermedad. Compr Physiol. (2019) 9:947–1023. doi: 10.1002/cphy.c180011
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Cheng, Y, Ding, S, Azad, MA, Song, B y Kong, X. Comparación de las razas de cerdos en la morfología del intestino delgado y las funciones digestivas a diferentes edades. Metabolitos. (2023) 13:132. doi: 10.3390/metabo13010132
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Hu, W, Du, L, Shao, J, Qu, Y, Zhang, L, Zhang, D, et al. Respuestas moleculares y metabólicas al estrés inmune en el yeyuno de pollos de engorde: análisis transcriptómico y metabolómico. Sci. (2024) 103:103621. doi: 10.1016/j.psj.2024.103621
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Jin, S, Xu, H, Yang, C y Karmin, O. Regulación del estrés oxidativo en el intestino de los lechones después de la infección por Escherichia coli enterotoxigénica (ETEC). Biochim Biophys Acta. (2024) 1871:119711. doi: 10.1016/j.bbamcr.2024.119711
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Lema, I, Araújo, JR, Rolhion, N, y Demignot, S. Jejunum: el lugar de encuentro poco estudiado de los lípidos de la dieta y la microbiota. Biochimie. (2020) 178:124–36. doi: 10.1016/j.biochi.2020.09.007
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Steinbach, E, Masi, D, Ribeiro, A, Serradas, P, Le Roy, T y Clément, K. Microbioma del intestino delgado superior en la obesidad y trastornos metabólicos relacionados: un nuevo campo de investigación. Metabolismo. (2024) 150:155712. doi: 10.1016/j.metabol.2023.155712
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Cui, C, Wang, X, Li, L, Wei, H y Peng, J. Implicaciones multifacéticas de las células de Paneth en diversas enfermedades dentro del intestino y sistémicamente. Frente Immunol. (2023) 14:1115552. doi: 10.3389/fimmu.2023.1115552
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Atanga, R, Singh, V, e In, JG. Células enteroendocrinas intestinales: dianas farmacológicas presentes y futuras. Int J Mol Sci. (2023) 24:8836. doi: 10.3390/ijms24108836
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Wiarda, JE, Becker, SR, Sivasankaran, SK y Loving, CL. Diversidad de células epiteliales regionales en el intestino delgado de cerdos. J Anim Sci. (2023) 101:101. doi: 10.1093/jas/skac318
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Wallaeys, C, García-González, N, y Libert, C. Las células de Paneth como piedras angulares de la salud intestinal y del organismo: un cebador. EMBO Mol Med. (2023) 15:E16427. doi: 10.15252/emmm.202216427
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. Seo, K, Seo, J, Yeun, J, Choi, H, Kim, Y-I y Chang, S-Y. El papel de las barreras mucosas en la salud intestinal humana. Arch Pharm Res. (2021) 44:325–41. doi: 10.1007/s12272-021-01327-5
58. Chen, Y, Li, B, Li, K y Lin, Y. Nanoenzimas de superóxido dismutasa: estado actual y perspectivas futuras sobre el tratamiento y diagnóstico de enfermedades cerebrales. Chem Commun. (2024) 60:4140–7. DOI: 10.1039/d3cc06288k
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Morgenstern, C, Lastres-Becker, I, Demirdöğen, BC, Costa, VM, Daiber, A, Foresti, R, et al. Biomarcadores de señalización de NRF2: estado actual y retos futuros. Redox Biol. (2024) 72:103134. doi: 10.1016/j.redox.2024.103134
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Egbujor, MC, Olaniyan, OT, Emeruwa, CN, Saha, S, Saso, L, y Tucci, P. Una visión del papel de los aminoácidos como antioxidantes a través de la activación de NRF2. Aminoácidos. (2024) 56:23. doi: 10.1007/s00726-024-03384-8
61. Yang, W, Huang, J, Xiao, B, Liu, Y, Zhu, Y, Wang, F, et al. La taurina protege a los espermatocitos de ratón del daño inducido por la radiación ionizante a través de la activación de la señalización Nrf2/HO-1. Célula Physiol Biochem. (2017) 44:1629–39. doi: 10.1159/000485762
Resumen de PubMed | Texto completo de Crossref | Google Académico
62. Wang, H, Cheng, Q, Bao, L, Li, M, Chang, K y Yi, X. Papel citoprotector de la hemo oxigenasa-1 en la quimiorresistencia del cáncer: enfoque en propiedades antioxidantes, antiapoptóticas y pro-autofagia. Antioxidantes (Basilea). (2023) 12:1217. doi: 10.3390/antiox12061217
Resumen de PubMed | Texto completo de Crossref | Google Académico
63. Xu, M, Che, L, Gao, K, Wang, L, Yang, X, Wen, X, et al. La taurina alivia el estrés oxidativo en las células epiteliales mamarias porcinas al estimular la vía de señalización Nrf2-MAPK. Ciencia de la Alimentación Nutr. (2023) 11:1736–46. doi: 10.1002/fsn3.3203
Resumen de PubMed | Texto completo de Crossref | Google Académico
64. Wang, J, Wei, L, Liu, C, Wang, L, Zheng, W, Liu, S, et al. El tratamiento con taurina alivia la mucositis intestinal inducida por el 5-fluorouracilo en ratones. Alimentos vegetales Hum Nutr. (2022) 77:399–404. doi: 10.1007/s11130-022-00980-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: lechones destetados, taurina, estrés oxidativo, salud intestinal, vía de señalización Nrf2/HO-1
Cita: Zhou M, Wu Z, Deng D, Wang B, Zhou X, Zhou B, Wang C y Zeng Y (2024) Efectos de la taurina en el rendimiento del crecimiento, la diarrea, el estrés oxidativo y la función de barrera intestinal de los lechones destetados. Frente. Vet. Sci. 11:1436282. doi: 10.3389/fvets.2024.1436282
Editado por:
Moyosore Joseph Adegbeye, Universidad de África, Estado de Bayelsa, Nigeria
Revisado por:
Lei Zhao, Universidad Agrícola de Heilongjiang Bayi, China
Hongbin Si, Universidad de Guangxi, China
Derechos de autor © 2024 Zhou, Wu, Deng, Wang, Zhou, Zhou, Wang y Zeng. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Yan Zeng, zengyan@hnwyy.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía