
Efectos del nanoselenio en el rendimiento, los índices sanguíneos y los metabolitos de la leche de las vacas lecheras durante el período pico de lactancia
Efectos del nanoselenio en el rendimiento, los índices sanguíneos y los metabolitos de la leche de las vacas lecheras durante el período pico de lactancia

 Yajing Wang2
Yajing Wang2 Manlin Wei1*
Manlin Wei1* Wen Peng1Yuxiang Wang1
Wen Peng1Yuxiang Wang1 Runze Zhang1Yongjie Zheng1
Runze Zhang1Yongjie Zheng1 Ji Ju1
Ji Ju1 Chenyang Dong1Liu Du1Meili Bao1
Chenyang Dong1Liu Du1Meili Bao1- 1Facultad de Ciencia y Tecnología Animal, Universidad Minzu de Mongolia Interior, Tongliao, China
- número arábigoFacultad de Ciencia y Tecnología Animal, Universidad Agrícola de China, Pekín, China
Para comparar el impacto del nanoselenio y el selenito de sodio en el rendimiento, los índices sanguíneos y los metabolitos de la leche de las vacas lecheras durante el período pico de lactancia, se seleccionaron dos grupos de vacas lecheras en las mismas condiciones: grupo control (grupo CON) y grupo de tratamiento (grupo NSe) para un experimento de 38 días (10 días para la adaptación y 28 días para el muestreo). Al grupo control (CON) se le proporcionó una dieta basal +3,3 g/d de selenito de sodio (pureza 1%), mientras que al grupo de nanoselenio (NSe) se le ofreció la misma dieta +10 mL/d de nanoselenio (concentración de selenio 1.500 mg/L). Los resultados mostraron que la NSe aumentó significativamente la producción de leche, el contenido de selenio en la leche y la eficiencia alimenticia (p < 0.05), pero no tuvo un efecto significativo sobre otros componentes de la leche (p > 0.05). La NSe aumentó significativamente el nitrógeno ureico en sangre (BUN) y la fosfatasa alcalina (ALP) (p < 0,05), pero no tuvo efectos significativos sobre el malondialdehído (MDA), la superóxido dismutasa (SOD), la glutatión peroxidasa (GSH-Px), la capacidad antioxidante total en sangre (T-AOC) o el selenio en sangre (p > 0,05). Además, se determinó la metabolómica no dirigida de la leche mediante la tecnología LC-MS, y se examinaron los metabolitos diferencialmente abundantes y sus vías de enriquecimiento. De acuerdo con estos hallazgos, NSe aumentó considerablemente el contenido de cetilmanósido, ácido undecilenoico, ácido 3-hidroxipentadecanoico, ácido 16-hidroxipentadecanoico, ácido treónico, etc., pero disminuyó el contenido de ácido galactárico, ácido mesacónico, CDP-glucosa, etc. Además, las vías metabólicas enriquecidas que se examinaron con un valor de impacto superior a 0,1 incluyeron el metabolismo de la niacina y la niacinamida, el piruvato, el ciclo del citrato, la riboflavina, el glicerofosfolípido, el butanoato y la tirosina. El análisis de correlación de Pearson también reveló una relación entre diferentes metabolitos de la leche y el selenio en sangre, así como entre el selenio de la leche y los índices bioquímicos de la sangre. En conclusión, en comparación con el selenito de sodio, el nanoselenio mejora la producción de leche, la eficiencia alimenticia y el contenido de selenio de leche de las vacas lecheras y regula los metabolitos de la leche y las vías metabólicas relacionadas en las vacas lecheras Holstein durante el período de lactancia máxima, lo que tiene ciertas perspectivas de aplicación en la producción lechera.
1 Introducción
El selenio (Se), un oligoelemento importante para los animales, está ampliamente involucrado en los procesos metabólicos fisiológicos y es de suma importancia para las funciones reproductivas, el crecimiento animal y la inmunidad (1, 2). El selenio existe en varias formas en los animales, pero principalmente como selenoproteínas, incluida la glutatión peroxidasa (GSH-Px), la tiroxina desiodinasa (DIO) y la tiorredoxina reductasa (TrxR) (3). Debido a que el selenio a menudo está estrechamente relacionado con la actividad inmune y el rendimiento reproductivo de los animales (4), la deficiencia de selenio puede provocar muchas enfermedades, incluido el estrés oxidativo, la miocardiopatía y la retención de placenta en los animales. Sin embargo, cantidades excesivas también pueden provocar una intoxicación crónica o aguda por selenio (5, 6), lo que resulta en un rendimiento de producción reducido.
El selenio en la naturaleza suele existir en dos formas: inorgánica y orgánica (7). Durante mucho tiempo, el selenito de sodio ha sido un suplemento dietético de uso común, pero su utilización se ha vuelto limitada debido a su baja biodisponibilidad, facilidad de sobredosis, fuerte toxicidad y contaminación ambiental (8). En comparación, se considera que el selenio orgánico posee alta eficiencia, baja toxicidad y contaminación, una alta tasa de absorción (9) y seguridad (10). Para garantizar la ingesta efectiva de selenio, la adición excesiva generalmente se adopta en la producción animal. Debido a la relativa proximidad de las dosis requeridas y tóxicas, tanto el selenio inorgánico como el selenio orgánico pueden causar fácilmente intoxicación. El tipo de selenio tiene una gran influencia en su toxicidad: los selenidos son altamente tóxicos y el selenio elemental tiene baja toxicidad porque no es fácilmente absorbido por los animales. Sin embargo, el nanoselenio es mucho más seguro porque su toxicidad se encuentra entre el selenio elemental y el selenio orgánico (11). Desde un punto de vista químico, el nanoselenio es selenio elemental rojo (valencia cero) con un tamaño de partícula de 20-60 nm que se dispersa alrededor de la proteína (12). Sin embargo, la capacidad de absorción y utilización del nanoselenio en el intestino delgado puede verse reforzada por su estructura química única y sus efectos a nanoescala (13). Esto puede atribuirse a sus efectos beneficiosos en el duodeno, como lo demuestra un aumento en la longitud, así como en el íleon y el yeyuno, medido por un aumento en la altura de las vellosidades, el área superficial y la densidad de las células caliciformes (14). Los estudios han demostrado que el nanoselenio puede reducir o prevenir el estrés oxidativo (15); También tiene efectos antivirales o puede mejorar la eficacia de la vacunación. Además, la toxicidad aguda y subaguda del nanoselenio es mucho menor que la del selenio inorgánico (16). Por el contrario, el nanoselenio puede aliviar la fibrosis hepática inducida por cadmio en pollos (17), interfiere con la malformación de espermatozoides inducida por cadmio-cloro y tiene efectos antivirales (18). Los estudios en animales de producción también revelaron mejoras en el rendimiento del crecimiento, la composición de la canal y la función inmune de los pollos de engorde suplementados con nanoselenio (19, 20), que también puede regular la fermentación ruminal de las ovejas y mejorar la salud de las cabras (21, 22). Además, tiene el potencial de aumentar la expresión de enzimas que contienen selenio en la glándula mamaria de las vacas lecheras y la actividad de GSH-Px en la sangre (23). Por lo tanto, el nanoselenio tiene ciertas aplicaciones.
Hay más de 40 países en el mundo con diversos grados de deficiencia de Se en el suelo (24). En la actualidad, el suelo de la mayoría de las provincias de China también está en riesgo de deficiencia de selenio (0,125 ≤ Se < 0,175 mg/kg), y el contenido de selenio en sangre de la población en la mayoría de las provincias está cerca del límite inferior del estado nutricional clínico (100-200 μg/L) cuando se excluye la suplementación adicional (25). En comparación con el consumo de medicamentos suplementarios de selenio, las personas prefieren obtener selenio a través de los alimentos. La ingesta de alimentos o dietas ricas en selenio es útil para mejorar o resolver los problemas nutricionales y de salud causados por la deficiencia de selenio en humanos y animales (26). En los últimos años, varios alimentos ricos en selenio, como la leche, el té, el arroz y los huevos enriquecidos con selenio, han sido favorecidos por las personas (27), y los métodos de suplementación de selenio en la alimentación animal para producir productos ricos en selenio son valorados por los profesionales.
Los productos lácteos son los alimentos más comunes y son una buena fuente de minerales. Los estudios han demostrado que aumentar el contenido de selenio en la leche es una forma factible de satisfacer las necesidades de selenio de las personas (28). El selenio dietético se puede secretar en la leche para obtener leche rica en selenio (29). Sin embargo, como un nuevo tipo de suplemento de selenio, el nanoselenio rara vez se ha estudiado en vacas lecheras, especialmente la eficiencia de la transferencia de nanoselenio del alimento a la sangre y la leche de los animales, y los metabolitos en la leche también necesitan más investigación. Por lo tanto, se estudiaron los efectos del nanoselenio sobre el rendimiento, los índices bioquímicos sanguíneos, los niveles de selenio y los metabolitos de la leche de las vacas lecheras durante el período pico de lactancia. Este estudio proporciona una referencia para la aplicación de un suplemento de selenio seguro y eficiente en la producción láctea.
2 Materiales y métodos
2.1 Diseño experimental y manejo de la alimentación animal
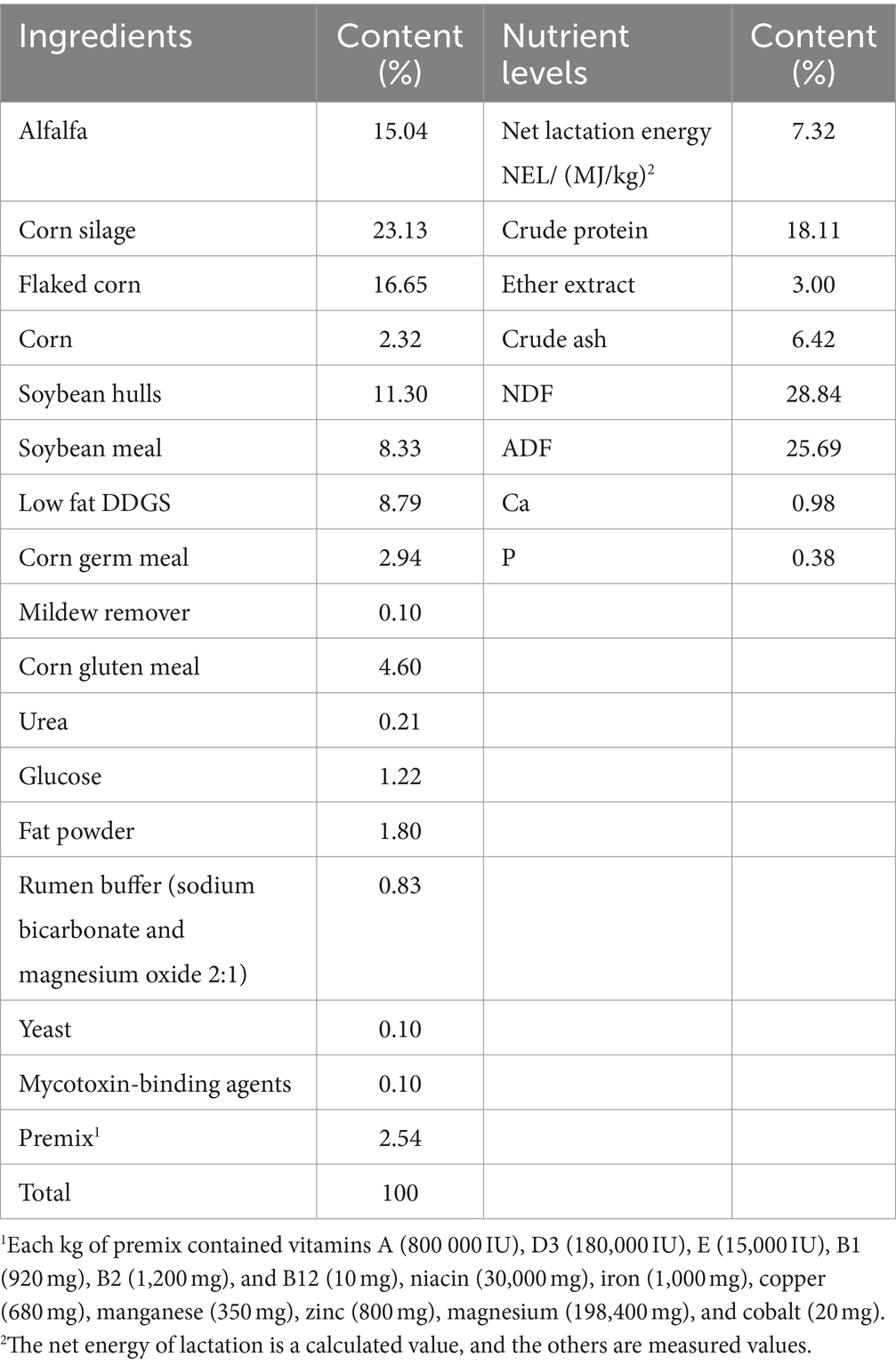
Se seleccionaron dos grupos de vacas Holstein sanas con pesos corporales, paridades y períodos de lactancia similares de 45 a 60 días como grupo de control (grupo CON) y grupo de tratamiento (grupo NSe), con 339 y 185 vacas, respectivamente, para el experimento de 38 días (adaptación de 10 días y medición de 28 días). La dieta basal se formuló con base en los Requerimientos de Nutrientes del Ganado Lechero (30) (Tabla 1). Al grupo CON se le administró una dieta basal +3,3 g/d de selenito de sodio (pureza del 1%), mientras que al grupo NSe se le administró una dieta basal +10 mL/d de nanoselenio (concentración de selenio 1.500 mg/L). El selenito de sodio y el nanoselenio se mezclaron previamente con harina de maíz y luego se mezclaron en dietas de TMR suplementadas con otras materias primas para piensos. El contenido de selenio en las dietas de ambos grupos fue de 15 mg por día. Las vacas fueron alimentadas dos veces (07:00 y 15:00), ordeñadas tres veces (07:00, 15:00 y 23:00) todos los días y tuvieron acceso gratuito a agua potable. La producción de leche y la ingesta de alimento se registraron diariamente y se evaluaron las indicaciones clínicas de intoxicación por selenio (Anexo 1 para los criterios de evaluación). Al final del ensayo, se recolectaron aleatoriamente muestras de leche de 27 vacas de cada grupo para analizar la composición de la leche, mientras que las muestras de sangre de 10 vacas de cada grupo se recolectaron aleatoriamente para determinar los índices bioquímicos, y se utilizaron 6 muestras de cada grupo para determinar el contenido de selenio en la sangre y la leche.
2.2 Registro de ingesta de alimento y análisis de muestras de alimento
La cantidad total y residual de la dieta TMR de cada grupo se registró diariamente. Se tomaron muestras de dietas representativas de TMR todos los días y se mezclaron en cantidades iguales para el análisis de nutrientes (monitoreo diario de la humedad). Después de secarse a 65 °C a un peso constante y triturarse, las muestras se pasaron por un tamiz de 1 mm. La materia seca (MS) se evaluó secándola a un peso constante a 105 °C y se utilizó para calcular la ingesta de materia seca (MSI) sobre la base de la ingesta individual de alimento. Se utilizaron métodos de AOAC (2005) para analizar los componentes de los piensos, incluyendo la proteína bruta (PC), la ceniza bruta (Ash) y el extracto de éter (EE) (31). Se empleó el método de Van Soest para examinar los contenidos de fibra detergente neutra (FDN) y fibra detergente ácida (FDA) (32).
2.3 Análisis de la producción y composición de la leche
Durante el período experimental, se registró la producción diaria de leche y se calculó la producción de leche corregida con grasa láctea (FCM) al 4%. Al final del ensayo, se determinó la composición de grasa, proteína, lactosa y cenizas en las muestras de leche mixta (leche matutina, media y vespertina = 4:3:3) de 27 vacas de cada grupo utilizando un analizador de composición de leche LM2 (Harrod Beijing Technology Co., Ltd.).
2.4 Toma de muestras y análisis de sangre
Al final del experimento, se recogieron muestras de sangre de 10 vacas de cada grupo de forma aleatoria para determinar los índices bioquímicos. Se tomaron muestras de sangre (10 mL) de la vena de la raíz de la cola 1 h antes de la alimentación matutina, se dejaron reposar a 4 °C durante 4 h y luego se centrifugaron (3.000 r/min, 10 min). Se colectó suero para determinar índices bioquímicos, índices de antioxidantes y contenido de selenio.
Los índices de antioxidantes sanguíneos superóxido malondialdehído (MDA), superóxido dismutasa (SOD), glutatión peroxidasa (GSH-Px) y capacidad antioxidante total (T-AOC) se determinaron utilizando kits de detección bioquímica (Instituto de Bioingeniería Nanjing Jiancheng, China) en un instrumento de etiquetado enzimático (Huawei Delang DR-200BS, Wuxi, China). Además, los niveles de glucosa (GLU), creatinina (CRE), nitrógeno ureico (BUN), alanina aminotransferasa (ALT), lactato deshidrogenasa (LDH), aspartato aminotransferasa (AST), γ-glutamil aminotransferasa (γ-GT), fosfatasa alcalina (ALP) y colinesterasa (CHE) también se midieron con el analizador bioquímico semiautomático A6 (Beijing Matsushige Technology, Beijing, China).
2.5 El contenido de selenio en la sangre y la leche
Se recogieron aleatoriamente muestras de sangre y leche de 6 vacas de cada grupo para determinar el contenido de selenio mediante colorimetría espectrofotométrica de 3,3′-diaminobenzidina (DAB). En los tubos en blanco y estándar, se agregaron 100 μL del material estándar y agua destilada, respectivamente. Se añadió tampón de ácido acético (400 μL) simultáneamente a los tubos en blanco, patrón y muestra, seguido de la adición de 200 μL de solución de EDTA-NA2. A continuación, se añadieron 200 μL de solución reveladora de color al tubo de muestra, seguido de una mezcla completa de todos los componentes. En la oscuridad, se añadieron 400 microlitros de solución alcalina a un baño de agua a 60 °C durante 20 minutos. Después de agitar, la mezcla se dejó durante 5 min, se retiraron 200 microlitros y se colocó la solución en una placa marcadora enzimática para determinar la densidad óptica (DO).
2.6 Análisis metabolómico no dirigido
La metabolómica no dirigida de la leche se determinó mediante LC-MS, y se examinaron los metabolitos diferencialmente abundantes y sus vías de enriquecimiento. Una vez que la muestra de leche se descongeló naturalmente a temperatura ambiente, se transfirieron 200 μL de muestra de leche a un tubo de centrífuga de 1,5 mL. A continuación, se introdujeron 400 μL de igual volumen de un disolvente de extracción compuesto de acetonitrilo y metanol. La muestra se sometió a vórtice durante 30 s y se extrajo por ultrasonidos a baja temperatura durante 30 min (5°C, 40 kHz). Después de exponer la muestra a -20 °C durante 30 min, se centrifugó a 4 °C durante 10 min a una velocidad de 13.000 rpm. A continuación, se desechó el sobrenadante y el sedimento se volvió a disolver en 100 μL de solución compleja (agua: acetonitrilo = 1:1). Posteriormente, la solución se extrajo mediante ultrasonidos a baja temperatura (5°C) durante 5 min a 40 kHz y se centrifugó a la misma velocidad a 4°C durante 5 min. Mediante intubación interna, el sobrenadante se vertió en un vial con una cánula interna para prepararlo para el análisis de la máquina. La detección se llevó a cabo mediante cromatografía líquida de ultra alta resolución-espectrometría de masas por transformada de Fourier en tándem (UPLC-FT-MS) en una plataforma LC-MS (sistema UHPLC-Q Exactive HF-X). Las condiciones de la máquina utilizadas incluyeron una columna cromatográfica (ACQUITY UPLC HSS T3, 100 mm × 2,1 mm de diámetro interior, 1,8 μm; Waters, Milford, EE.UU.), y la temperatura de la columna fue de 40°C; las fases móviles A y B estaban compuestas por agua (95%) y acetonitrilo (5%) con ácido fórmico (0,1%) y acetonitrilo (47,5%), alcohol isopropílico (47,5%) y agua (5%) con ácido fórmico (0,1%), respectivamente. El volumen de la muestra fue de 3 μL y la temperatura de la columna fue de 40 °C. Condiciones del espectro de masas: ionización por electrospray y modos de barrido de iones negativos y positivos para la captación de señales de espectro de masas, respectivamente.
2.7 Análisis estadístico
Los datos de rendimiento de producción, índice sanguíneo y contenido de leche se introdujeron en Excel (2016) y se analizaron mediante la prueba t de Student utilizando el software JMP13.0 (SAS Institute, Japón). Los resultados se presentan como los errores estándar y los valores medios. Las diferencias se consideraron significativas y extremadamente significativas cuando p < 0,05 y p < 0,01, respectivamente. Además, se llevó a cabo un análisis de diferencias de los archivos de la matriz después del preprocesamiento de los datos metabolómicos. La estabilidad del modelo se evaluó mediante validación interactiva de 7 ciclos, y se adoptó el análisis discriminante de mínimos cuadrados parciales (PLS-DA) utilizando el paquete de software R ropls (versión 1.6.2). También se realizaron análisis de diferencias múltiples y la prueba t de Student. Se utilizó el valor de p y la variable influencia en la proyección (VIP) del modelo de análisis discriminante de mínimos cuadrados parciales (PLS-DA) para evaluar los metabolitos diferencialmente abundantes. Cuando VIP > 1, p < 0.05 y FC > 1 o FC < 1, los metabolitos diferencialmente abundantes se identificaron a partir de la base de datos KEGG 1 e involucrados en las vías que fueron analizadas por el paquete de software Python SciPy. Se utilizó la prueba exacta de Fisher para determinar los datos estadísticos y las vías biológicas más relevantes. Se realizó un análisis de correlación de Pearson entre los metabolitos de la leche, el contenido de selenio en sangre, el contenido de selenio en la leche y los índices bioquímicos en sangre.
3 Resultados
3.1 DMI, producción de leche y composición de la leche
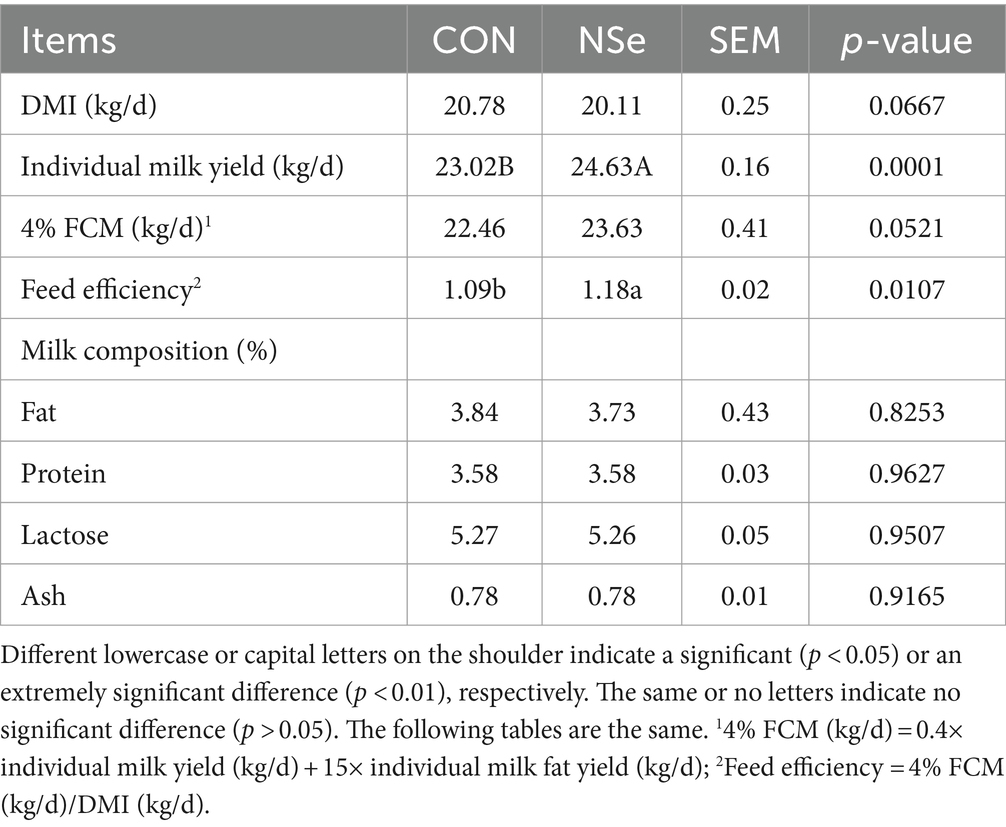
Como se muestra en la Tabla 2, la producción de leche y la eficiencia alimenticia aumentaron significativamente en el grupo NSe en comparación con los del grupo CON (p < 0,05). Sin embargo, los rendimientos de MSI, FCM y los contenidos de proteína, grasa, cenizas y lactosa de la leche no difirieron significativamente (p > 0,05). Además, no se observó ningún indicio de envenenamiento durante el experimento.
3.2 Índices de antioxidantes séricos
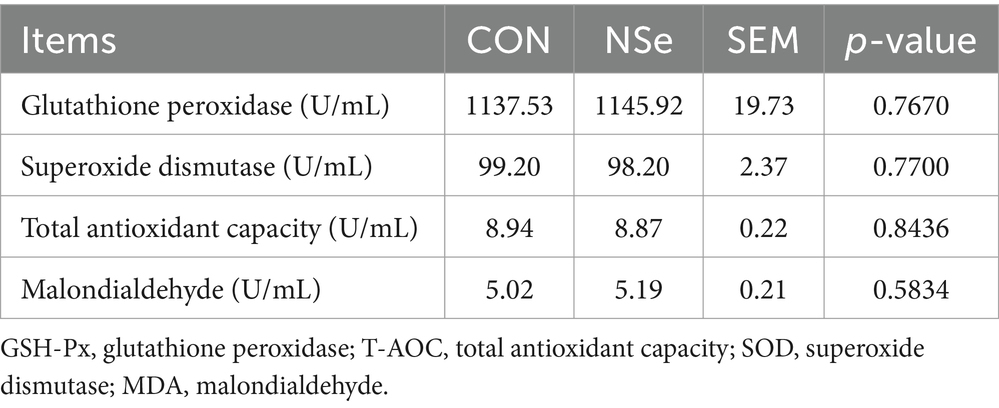
La Tabla 3 muestra que el nanoselenio no tuvo un impacto estadísticamente significativo en los niveles séricos de GSH-Px, T-AOC, SOD o MDA en vacas lecheras (p > 0.05).
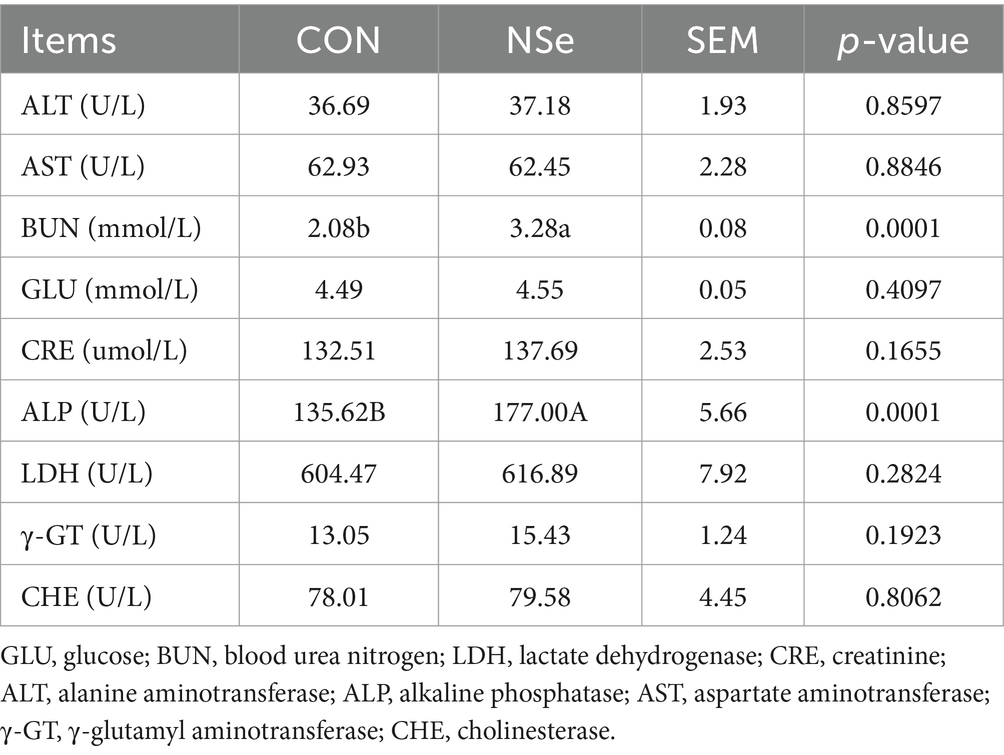
3.3 Índices bioquímicos séricos
Como se muestra en la Tabla 4, en comparación con el grupo CON, la NSe aumentó significativamente los niveles séricos de BUN y ALP (p < 0,05), pero no tuvo un impacto significativo en otros índices bioquímicos séricos (p > 0,05).
3.4 Contenido de selenio en la sangre y la leche
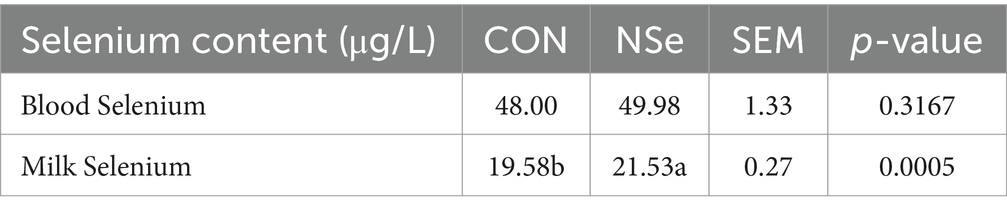
La Tabla 5 muestra que la NSe aumentó significativamente el contenido de selenio de la leche en un 9,96% (p < 0,05) en comparación con el del grupo CON, pero no afectó el contenido de selenio de la sangre (p > 0,05).
3.5 Metabolitos de la leche
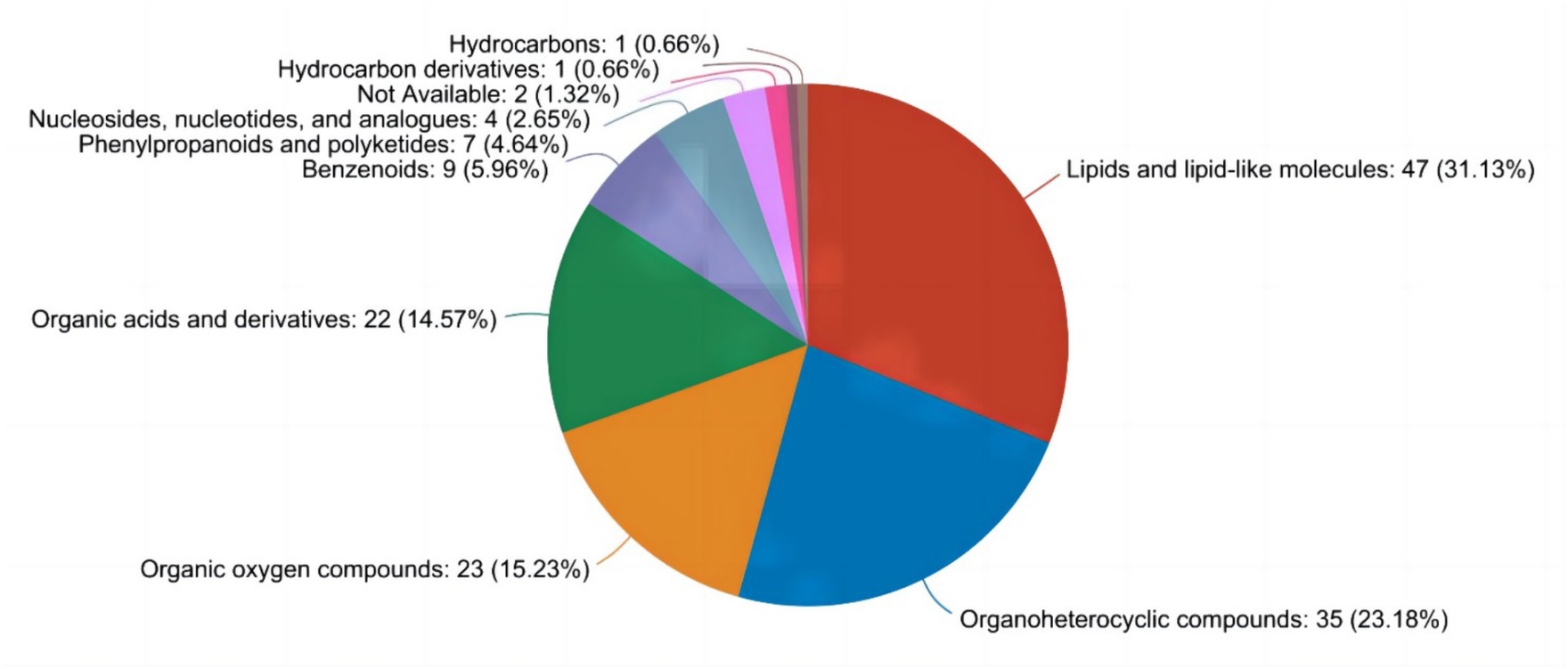
Como se ilustra en la Figura 1, a través del análisis metabolómico no dirigido de muestras de leche, se identificaron un total de nueve tipos distintos de metabolitos. Entre estos, los compuestos orgánicos heterocíclicos (24,14%), los lípidos y las moléculas lipídicas (33,10%) y los ácidos orgánicos y sus derivados (17,24%) constituyeron las tres principales categorías de compuestos.
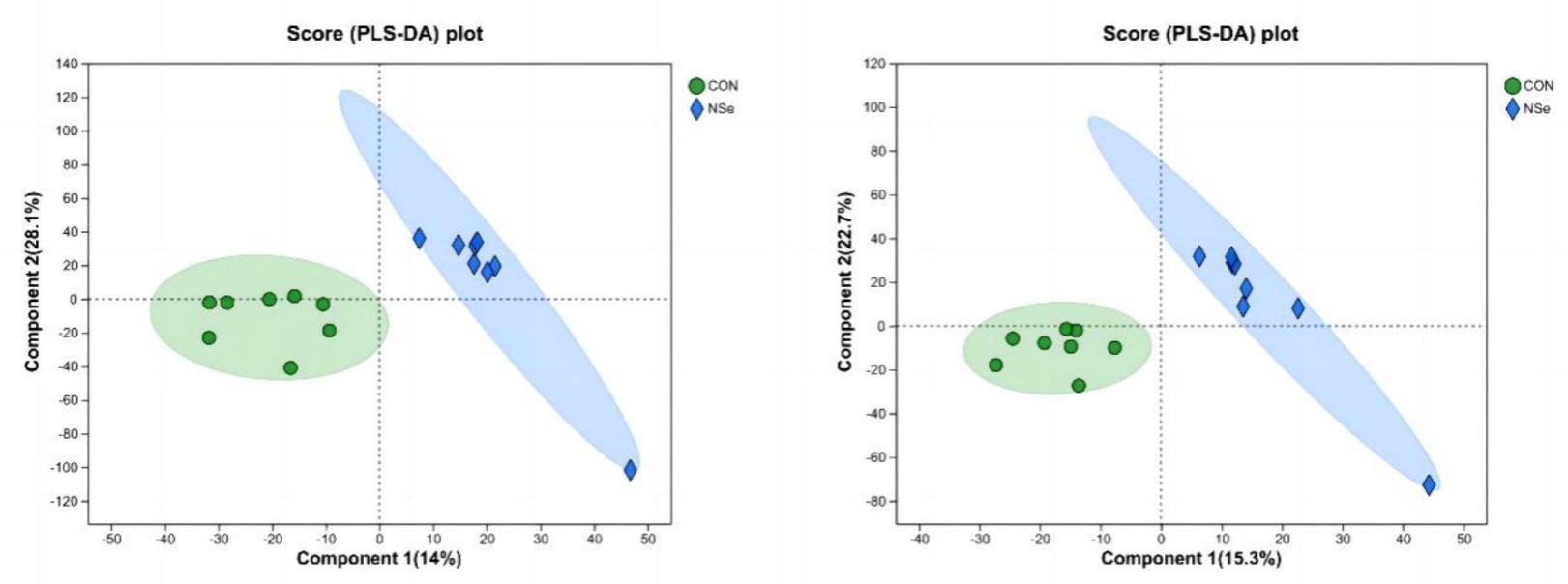
La tabla de puntuación PLS-DA (Figura 2) mostró que los metabolitos de la leche en ambos grupos podían separarse significativamente. En el modo de iones positivos, el 14% de la varianza fue explicada por el componente principal, y el 28,1% de la varianza fue explicada por el segundo componente principal. En el modo de iones negativos, el primer componente principal explicó el 15,3% y el segundo componente principal el 22,7%. Todas las muestras estaban dentro de los intervalos de confianza del 95%, lo que indica que hubo poca diferencia en el número de muestras en cada grupo entre los grupos, y el área de proyección del grupo NSe fue significativamente diferente de la del grupo CON.
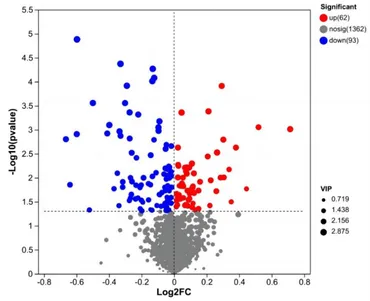
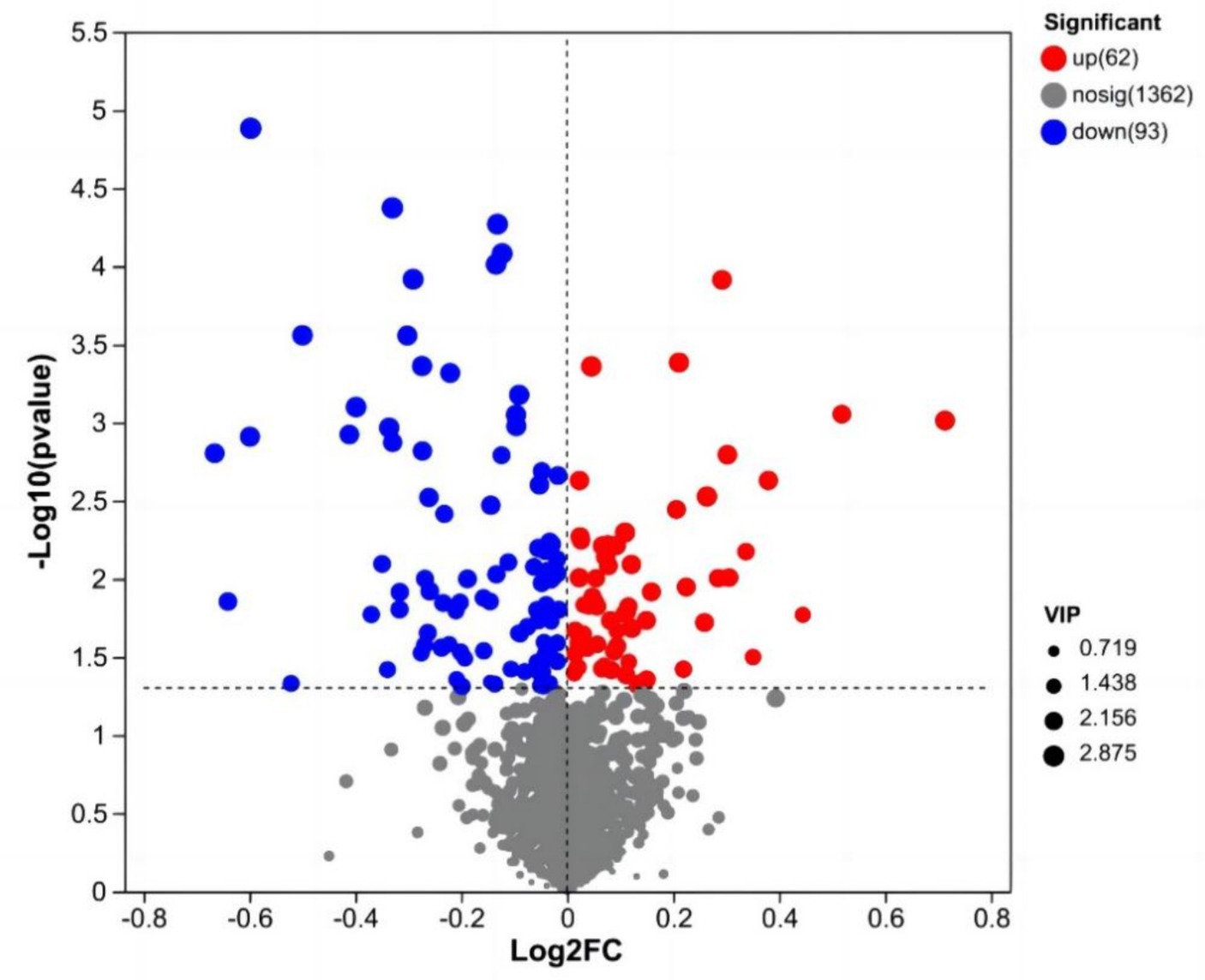
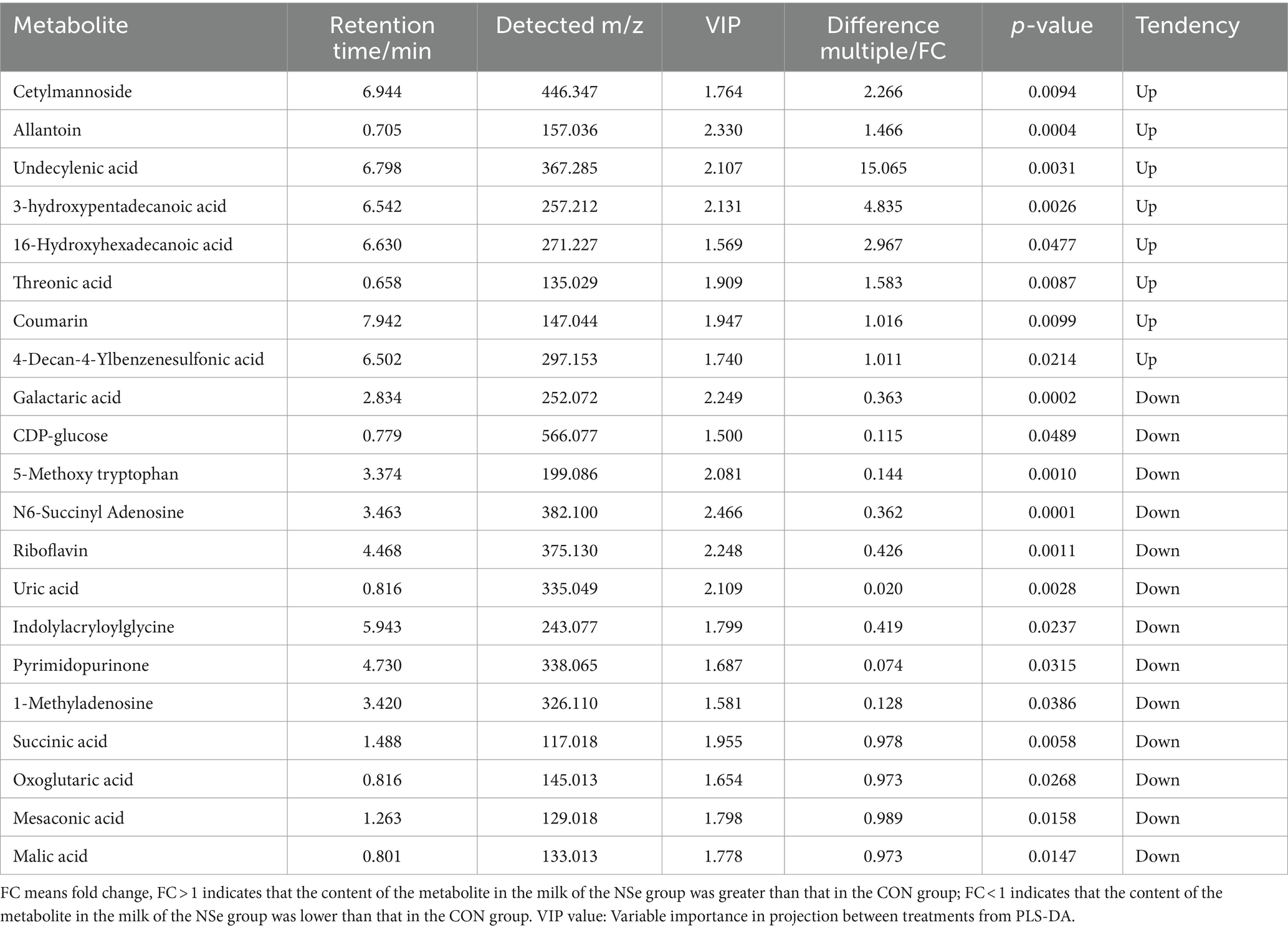
Después del preprocesamiento de los datos, se realizó un análisis de la prueba t de Student y se generó un gráfico volcánico de acuerdo con los valores de p y FC, como se muestra en la Figura 3. Se evaluaron un total de 155 metabolitos diferencialmente abundantes utilizando los parámetros de tamizaje FC > 1 o FC < 1, VIP > 1 y p < 0,05. Entre estos metabolitos, 62 estaban regulados a la baja, mientras que 93 estaban regulados al alza. Los resultados de algunos metabolitos diferencialmente abundantes se muestran en la Tabla 6.
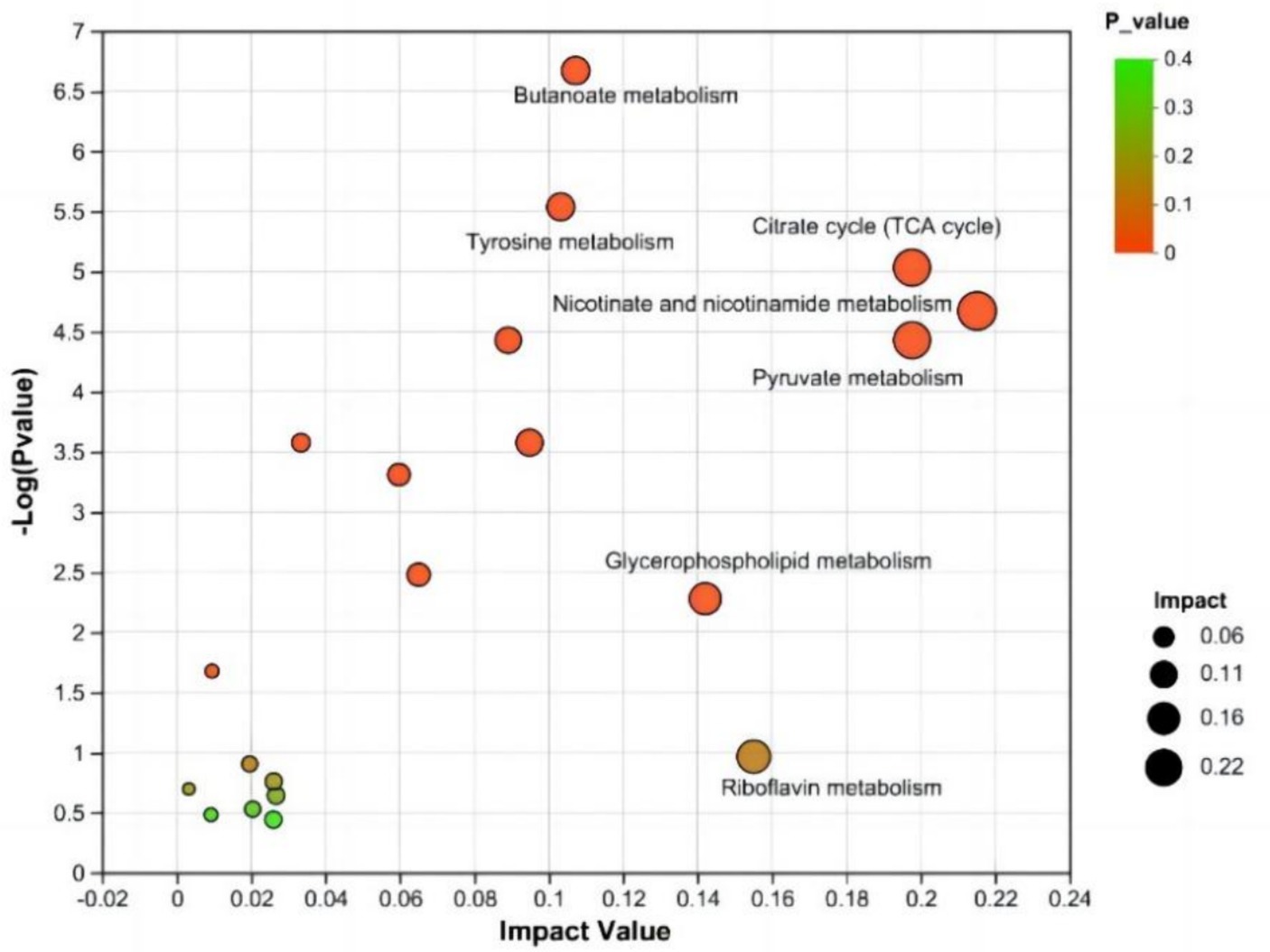
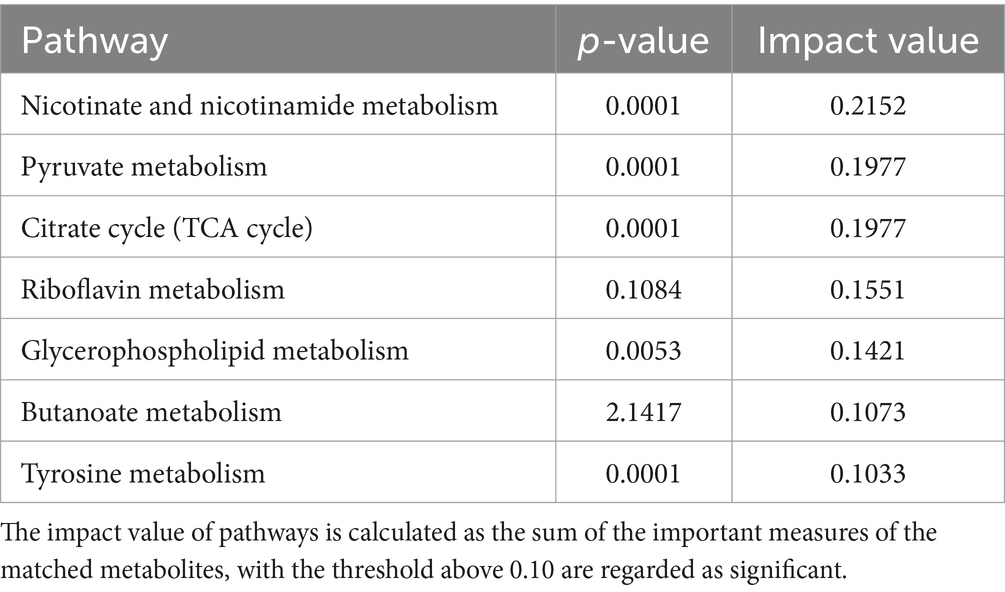
Se realizó un análisis de enriquecimiento de las vías metabólicas asociadas con los metabolitos diferencialmente abundantes entre los grupos CON y NSe. Los hallazgos demostraron la participación de 31 vías metabólicas que involucran 155 metabolitos diferencialmente abundantes identificados. Las vías metabólicas afectadas por metabolitos diferencialmente abundantes se examinaron utilizando un valor de impacto superior a 0,1. Las principales vías metabólicas implicadas fueron el metabolismo de la niacina y la niacinamida, el piruvato, el ciclo del citrato (ciclo del TCA), el glicerofosfolípido, la riboflavina, el butanoato y la tirosina. Los resultados se muestran en la Figura 4 y en la Tabla 7.
Se realizó un análisis de correlación de Pearson para examinar las relaciones entre el contenido de selenio en sangre, el contenido de selenio en leche y los índices bioquímicos en sangre. La Figura 5 muestra una correlación positiva estadísticamente significativa entre los niveles de selenio sérico y lácteo (p < 0,05). Además, también se detectó una correlación negativa estadísticamente significativa entre la concentración sérica de selenio y el T-AOC en sangre (p < 0,05). El contenido de selenio en la leche se correlacionó positivamente con BUN y ALP (p < 0.01), y γ-GT se correlacionó negativamente con AST (p < 0.05) y positivamente se correlacionó con SOD (p < 0.05).
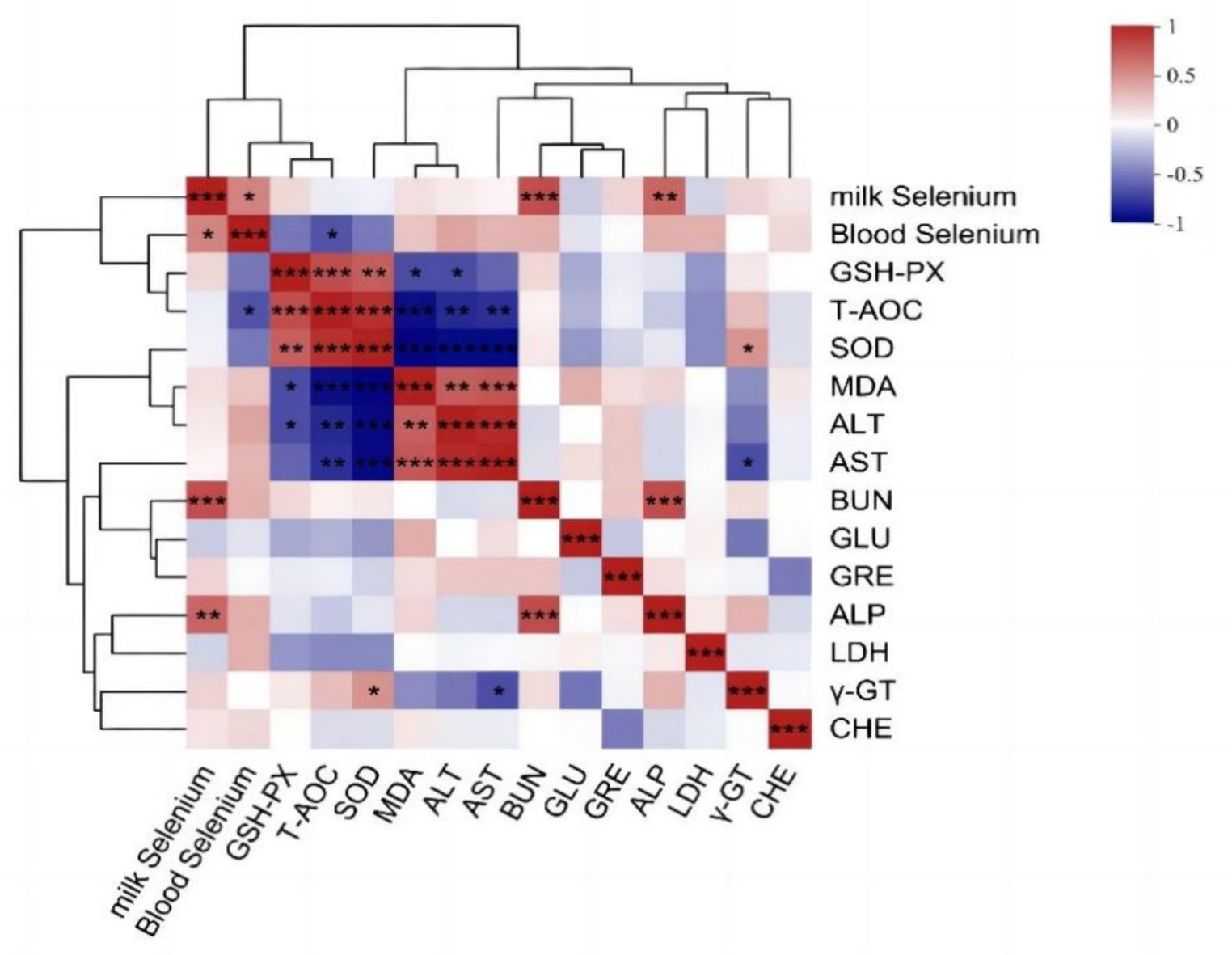
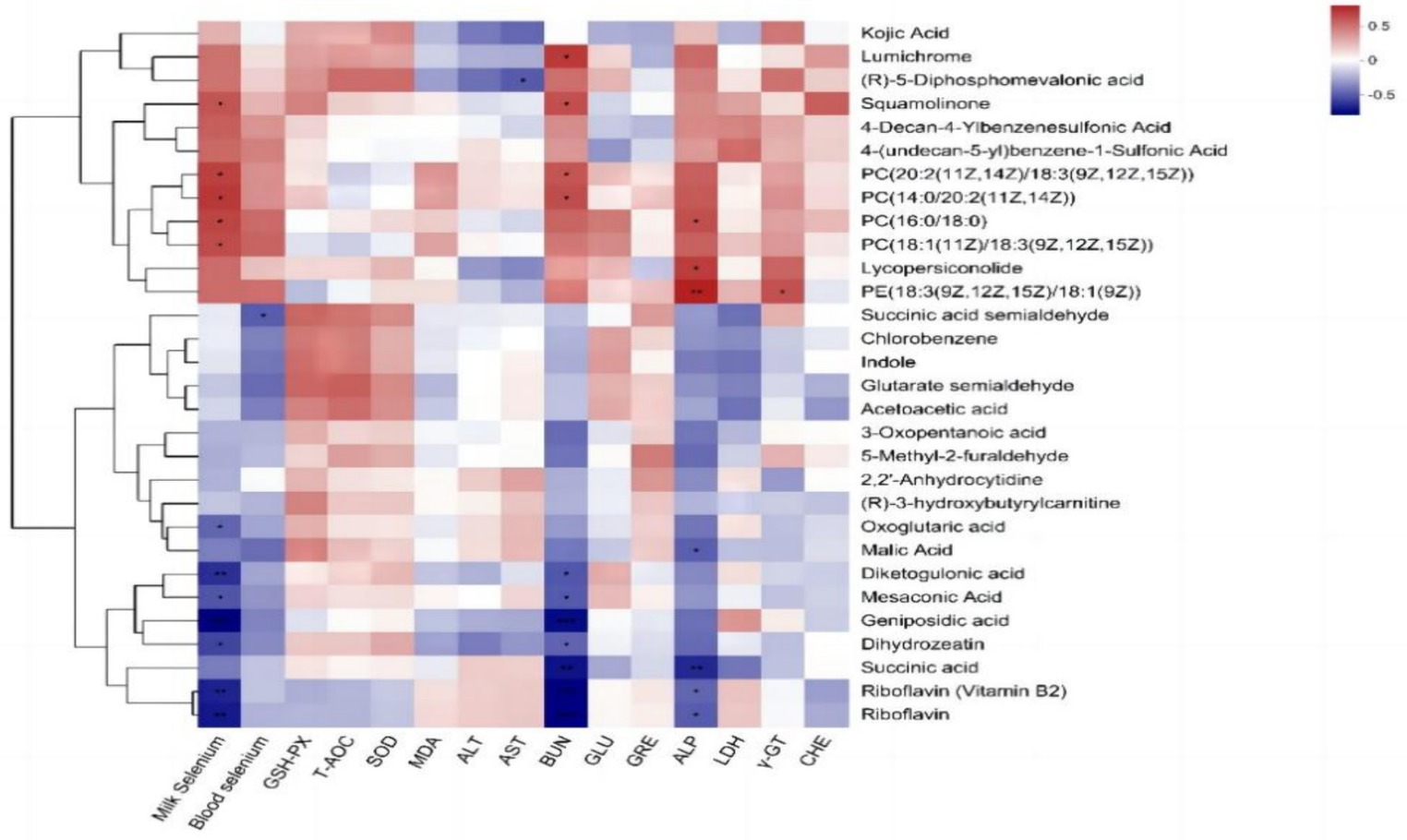
El análisis de correlación de Pearson de los metabolitos de la leche con el contenido de selenio en sangre, el contenido de selenio en la leche y los índices bioquímicos en sangre (Figura 6) reveló que el contenido de selenio en la leche se correlacionó significativamente positivamente con los metabolitos de la leche escamolinona, PC [20:2 (11Z, 14Z)/18:3 (9Z, 12Z, 15Z)], PC [14:0/20:2 (11Z, 14Z)], PC (16:0/18:0) y PC [18:1 (11Z)/18:3 (9Z, 12Z, 15Z)] y se correlacionó negativamente de manera extremadamente significativa con la riboflavina (vitamina B2), la riboflavina, el ácido dicetogulónico y el ácido geniposídico (p < 0,01), pero se correlacionó significativamente negativamente con el ácido oxoglutárico, el ácido mesocónico y la dihidrozeatina (p < 0,05). El contenido de selenio en sangre se correlacionó significativamente negativamente con la cantidad de semialdehído de ácido succínico entre los metabolitos de la leche (p < 0,05). La AST se correlacionó negativamente con el ácido (R)-5-difosfomevalónico (p < 0,05). El BUN mostró una correlación positiva significativa con lumicromo, escamolinona, PC (20:2 (11Z, 14Z)/18:3 (9Z, 12Z, 15Z) y PC [14:0/20:2 (11Z, 14Z)] (p < 0,05) y una correlación negativa altamente significativa con riboflavina (vitamina B2), riboflavina, ácido succínico y ácido geniposídico (p < 0,01). Además, el BUN se correlacionó significativamente negativamente con el ácido dicetogulónico, el ácido mesacónico y la dihidrozeatina (p < 0,05). La FA mostró una correlación positiva altamente significativa con la EP [18:3 (9Z, 12Z, 15Z)/18:1 (9Z)] (p < 0,01), una correlación positiva significativa con las concentraciones de lumicromo y escamolinona (p < 0,05) y una correlación negativa significativa con riboflavina, riboflavina (vitamina B2) y ácido málico (p < 0,05). Se observó una correlación positiva entre γ-GT y EP [18:3 (9Z, 12Z, 15Z)/18:1 (9Z)] (p < 0,05).
4 Discusión
Tanto el exceso como la insuficiencia de selenio en las dietas de los animales pueden reducir el rendimiento de los animales (33, 34). De acuerdo con los Requerimientos de Nutrientes de las Vacas Lecheras (30, 35), la cantidad de selenio en las dietas oscila entre aproximadamente 0,3 mg/kg MS y 3 mg/kg MS. La cantidad de selenio añadido en este experimento fue de aproximadamente 0,75 mg/kg MS. De acuerdo con los resultados de la evaluación de las indicaciones clínicas de intoxicación por selenio (Anexo 1 para los criterios de evaluación) y el contenido de selenio en sangre de las vacas lecheras, no se observaron indicaciones anormales durante todo el experimento, lo que indica que el contenido de selenio en las dietas estaba dentro de un rango aceptable.
Los diferentes tipos de suplementos de selenio tienen diversos efectos en el rendimiento de las vacas lecheras. Como se indica en la Tabla 2, el IMD no se ve afectado por las fuentes de selenio, lo que es consistente con los resultados de las investigaciones de Sun et al. (36) y Juniper et al. (37). Además, también se confirmó que la suplementación con la fuente de selenio no parecía tener ningún efecto sobre el IMD de las vacas lecheras, el ganado vacuno, los terneros y los corderos (38). Sin embargo, se encontró que la producción de leche de las vacas lecheras durante las fases media y tardía de la lactancia aumentaba mediante la administración de suplementos dietéticos que contenían levadura de selenio (SY) e hidroxi-selenometionina (OH-SeMet) (39). El aumento en la producción de leche en las vacas puede o no estar directamente relacionado con los efectos positivos de los suplementos de selenio (39, 40), ya que está influenciado por varios factores, incluida la composición nutricional, el estado de lactancia, la función ruminal y la salud en general. Sin embargo, en este experimento, sobre la base de que no hubo cambios significativos en el DMI, el aumento en la producción de leche de las vacas lecheras fue muy cercano al aumento en la eficiencia alimenticia. Por lo tanto, es razonable pensar que existe una cierta correlación entre la producción de leche y la eficiencia alimentaria. Wang et al. (41) han reportado resultados similares en el sentido de que la producción de leche aumentó con la suplementación con selenio-levadura sin cambios en el IMD, pero mejoró la fermentación ruminal. Las vacas lecheras tienen un metabolismo fuerte y una alta intensidad de producción cuando están en el pico del período de lactancia. Lo que significa que las vacas pueden obtener más nutrientes de los piensos y utilizarlos de forma más eficiente (42), lo que proporciona la base material para la mejora de la producción de leche. Por otro lado, la suplementación con selenio puede fortalecer la distribución de los capilares mamarios, y el área vascular mamaria tiende a aumentar (43), lo que proporciona condiciones organizativas para la mejora de la producción de leche. Además, el nanoselenio también tiene un efecto de tamaño único (nanoescala), lo que lo hace más absorbible y más biodisponible en el intestino delgado que el selenio inorgánico (41, 44). Esto también explica el aumento del contenido de selenio en la leche; es decir, aumentó la conversión de selenio alimenticio en selenio lechero. En cuanto a la composición de la leche, la suplementación con nanoselenio no tuvo efectos significativos sobre la grasa, la proteína, la lactosa o las cenizas de la leche, lo que estuvo en línea con los resultados de estudios anteriores (41). Esto indica que el selenio tiene poco efecto sobre la estructura y síntesis de las células acinares de la glándula mamaria (45), pero este hallazgo debe ser confirmado por estudios adicionales.
La glutatión peroxidasa (GSH-Px) es un indicador importante de la capacidad antioxidante de los animales. Puede proteger la estructura y la función de las membranas celulares al catalizar específicamente la reducción del peróxido de hidrógeno a través de la reducción de GSH (46), y el selenio es su componente activo. Por lo tanto, la suplementación con selenio en la dieta de las vacas lecheras podría, en teoría, mejorar la actividad de GSH-Px en sangre, mejorando en última instancia su capacidad antioxidante. Sin embargo, se informó que los efectos de la suplementación con selenio en la sangre GSH-Px fueron diferentes. Los estudios en cerdos de engorde, pollos de engorde y vacas lecheras han demostrado que la adición de cantidades adecuadas de selenio puede mejorar la actividad de GSH-Px en el suero y que el efecto del selenio orgánico es mayor que el del selenio inorgánico (47-49). Sin embargo, algunas investigaciones han demostrado que la suplementación con selenito de sodio y levadura de selenio en vacas lecheras no tiene un efecto significativo sobre la glutatión peroxidasa en sangre (GSH-Px) (50, 51). Estos resultados están en línea con los resultados de esta investigación, que no mostraron cambios significativos en los niveles séricos de SOD, T-AOC o MDA. El T-AOC, SOD y MDA también son parámetros importantes que reflejan el estado antioxidante de los animales. La resistencia del cuerpo al estrés oxidativo se puede medir con el T-AOC. La SOD es un indicador significativo de la capacidad antioxidante en los animales. Funciona eliminando los radicales libres, defendiendo contra los efectos nocivos de los radicales libres de oxígeno y reparando rápidamente las células dañadas. El MDA es un producto importante de la peroxidación de los lípidos de membrana y sirve como indicador del daño tisular causado por la peroxidación. La suplementación con selenio no necesariamente mejora el nivel de antioxidantes de los animales, y los estudios han demostrado que la capacidad antioxidante del cuerpo puede incluso reducirse con la suplementación con selenio (52). El impacto del selenio en la capacidad antioxidante del cuerpo depende de varios factores, como el tiempo y la cantidad de suplementos de selenio, la etapa de lactancia, el suministro de nutrientes, el estrés y la genética.
Los índices bioquímicos sanguíneos son indicadores importantes que reflejan el metabolismo fisiológico y el estado de salud de los animales (53). Cuando el cuerpo carece de selenio, se produce la inmunosupresión. Por el contrario, la suplementación excesiva con selenio en la dieta conducirá a la intoxicación por selenio en las vacas lecheras e incluso causará necrosis hepática y renal, que a menudo se reflejan en cambios en los parámetros bioquímicos de la sangre (54, 55). En la actualidad, se ha encontrado que la suplementación adecuada de selenio en la dieta puede mejorar la función inmunológica de las vacas lecheras, promover la producción de inmunoglobulina, aumentar los títulos de anticuerpos y reducir las enfermedades (56). Los resultados de este ensayo demostraron que la NSe aumentó significativamente los niveles de BUN y ALP en la sangre. Pero tuvo un impacto mínimo en otros indicadores bioquímicos, que fueron consistentes con estudios previos (4, 57). El BUN, un metabolito de aminoácidos y proteínas, puede servir como indicador del equilibrio del metabolismo de proteínas y aminoácidos dentro de un animal (58). El BUN de los rumiantes se deriva principalmente del amoníaco producido por la proteína degradable del rumen (RDP) y los metabolitos de la proteína no degradable del rumen (UDP) que ingresan al estómago y al intestino delgado. El BUN se envía al hígado a través de la vena porta para sintetizar urea, que se excreta por el riñón o regresa al rumen a través de la saliva y el epitelio ruminal. El nivel de BUN se ve afectado por muchos factores, como la proteína bruta (PC) de la dieta, la PDR, el tiempo de muestreo, la excreción renal y la reabsorción renal. Los estudios han demostrado que el contenido de BUN en el ganado aumenta primero después de la alimentación y luego disminuye a los niveles previos a las comidas 8 h después de la alimentación (59). La extracción de sangre en este estudio se realizó 1 h antes de la alimentación matutina, cuando el nivel de BUN estaba cerca del nivel más bajo en un día y dentro del rango normal [generalmente 4-16 mg/dL (60) o 1,4-5,7 mmol/L], lo que indica una cantidad suficiente de proteínas en la dieta. Dado que ambos grupos tenían niveles similares de creatinina sérica (ERC) y la relación BUN/CRE estaba dentro del rango normal (generalmente <20), se podía esperar una función de excreción renal normal. Por lo tanto, es más probable que el aumento de BUN en el grupo NSe esté relacionado con el aumento del nitrógeno en el tracto digestivo o la reabsorción de riñón, los cuales podrían ser buenos para la utilización de proteínas. Un estudio previo ha demostrado que el BUN de las vacas de alto rendimiento es mayor que el de las vacas de bajo rendimiento (61). Por otro lado, la actividad de la fosfatasa alcalina (ALP) en la sangre refleja la actividad metabólica y el estado funcional de los tejidos y órganos animales (62). La fosfatasa alcalina suele estar elevada cuando los animales desarrollan enfermedades o tumores hepáticos y óseos; Sin embargo, a veces también se eleva por razones fisiológicas, como en animales jóvenes, hembras gestantes y lactantes, o la ingesta de dietas ricas en grasas (63). Se ha reportado que el daño causado por la intoxicación por selenio ocurre principalmente en el hígado (64), pero con base en las concentraciones normales de AST y ALT, los indicadores de lesión y gravedad de los hepatocitos (57), se especuló que puede estar relacionado con una alta producción de leche o mastitis subclínica, en lugar de funciones hepáticas anormales. La razón es que la fosfatasa alcalina está estrechamente asociada con la reducción del calcio en la sangre y la promoción de la deposición ósea. Cuando la movilización de calcio es excesiva en el cuerpo, la proliferación compensatoria de los osteoblastos aumenta el nivel y la actividad de la FA sérica para mantener la estabilidad del contenido de calcio en el cuerpo para asegurar la deposición de calcio en el cuerpo (65). Cuando la producción de leche aumentó en las vacas del grupo NSe, los animales se movilizaron y consumieron más calcio, y el aumento del contenido de ALP en sangre permitió a las vacas mantener una deposición normal de calcio.
Después de que el selenio se absorbe en el tracto intestinal de un animal, puede ingresar a la sangre o transferirse a la leche, por lo que el contenido de selenio en sangre y selenio en la leche puede reflejar el estado de uso del selenio en las vacas. El contenido de selenio en la sangre a menudo está relacionado con la capacidad de unión del selenio, y los estudios han demostrado que agregar selenio orgánico a la dieta tiene un mayor efecto en el aumento de la concentración de selenio en la sangre que agregar selenio inorgánico (66, 67). Esto podría deberse al aumento de la capacidad de unión del selenio orgánico a la α y β globulina, LDL (lipoproteína de baja densidad), VLDL (lipoproteína de muy baja densidad) y albúmina en la sangre (68). El nanoselenio también puede tener un mayor grado de unión que el selenito de sodio, ya que también se ha informado del aumento del selenio en sangre (como el glutatión), el selenio en la leche, así como la estimulación de la expresión génica de la selenoproteína en las glándulas mamarias de las vacas lecheras mediante la suplementación con nanoselenio (23). La correlación positiva entre el selenio en la sangre y en la leche puede estar relacionada con la unión del selenio a las proteínas, y se considera que el selenio se une primero a los aminoácidos o proteínas en la sangre (69) y posteriormente es absorbido e integrado por la glándula mamaria para sintetizar las proteínas de la leche (70). Sin embargo, el nivel de selenio en sangre en el grupo NSe en este ensayo fue solo numéricamente más alto que en el grupo CON, pero no estadísticamente significativo (p > 0,05), solo los niveles de selenio en leche aumentaron significativamente (p < 0,05). Debido al pequeño tamaño de las partículas de nanoselenio (nanoescala), se necesitan más pruebas para confirmar si el nanoselenio no se une completamente a las proteínas de la sangre, sino que se transfiere rápidamente de los capilares mamarios a la leche.
Como la fuente más común de nutrientes para los seres humanos, los productos lácteos no solo proporcionan a las personas nutrientes básicos, sino que también tienen una mayor concentración de metabolitos de moléculas pequeñas, como ácidos orgánicos, ácidos grasos, aminoácidos, nucleótidos y péptidos bioactivos (71). Por lo tanto, los metabolitos de la leche de las vacas Holstein en el período pico de lactancia se analizaron mediante metabolómica no dirigida (LC-MS) en este estudio. Los resultados demostraron que la NSe aumentó significativamente el contenido de ácido treónico, cetilmanósido, ácido undecilenoico, ácido 3-hidroxipentadecanoico, ácido 16-hidroxipentadecanoico, etc. El ácido treónico es un metabolito activo del catabolismo del ácido ascórbico, también puede derivarse de proteínas glicosiladas (72), y se ha demostrado que es beneficioso para la absorción de calcio y para complementar el ácido ascórbico (73). Esto ayuda a garantizar una alta producción de leche. El ácido 3-hidroxipentadecanoico y el ácido 16-hidroxipentadecanoico son ácidos grasos hidroxilo que se encontraron presentes en la leche (74). El cetilmanósido activa el sistema del complemento en forma de liposomas (75). Mientras que se demostró que el ácido undecilenoide tiene actividad antifúngica (76). En este estudio, los niveles de ácido galactárico, ácido mesacónico y CDP-glucosa, etc. en la leche del grupo NSe disminuyeron. El ácido galactárico es un producto de la descomposición microbiana de la pectina, que ayuda a mantener la estabilidad de los alimentos (como el yogur) (77, 78). El ácido mesacónico es el producto de degradación del ácido cítrico en la leche (79). La CDP-glucosa participa en la síntesis de lactosa (80). Aunque la concentración de glucosa en la leche de todos los mamíferos (excepto los humanos) es baja (0,1-0,3 mmol/L), la concentración de glucosa en la leche puede reflejar la concentración de glucosa en las células mamarias. Cuando la eficiencia de la síntesis de lactosa en las células mamarias disminuye, la concentración de glucosa en la leche disminuye (81).
En las vías metabólicas donde se concentran metabolitos diferencialmente abundantes, la niacina y la niacinamida, el piruvato y el ciclo del citrato (ciclo del TCA) se relacionan principalmente con metabolitos como el ácido mesacónico, el ácido succínico, el ácido oxoglutárico y el ácido málico en la leche. La riboflavina interviene principalmente en el metabolismo de la riboflavina. La vía metabólica de los glicerofosfolípidos está relacionada principalmente con el metabolismo de los lípidos, como la PE [18:1 (9Z)/18:3 (9Z, 12Z, 15Z)] y la PC [18:1 (11Z)/18:3 (9Z, 12Z, 15Z)], y el metabolismo de las vías del butanoato y la tirosina está relacionado principalmente con metabolitos como el semialdehído del ácido succínico, el ácido acetoacético, el ácido succínico y el ácido maleico.
5 Conclusión
En condiciones experimentales a corto plazo, NSe aumentó significativamente la producción de leche, el contenido de selenio de la leche y la tasa de conversión alimenticia (p < 0,05). Sin embargo, no se observó un impacto significativo para otros componentes de la leche (p > 0,05). La NSe aumentó significativamente los niveles de BUN y ALP en sangre (p < 0,05), pero no tuvo un impacto significativo en los niveles sanguíneos de T-AOC, GSH-Px, SOD o MDA ni en el contenido de selenio en sangre (p > 0,05). En general, la eficiencia de la conversión de nanoselenio de selenio alimentario a selenio de leche es mayor que la del selenito de sodio, y el nanoselenio tiene ciertos efectos reguladores sobre los índices bioquímicos de la sangre, los metabolitos de la leche y las vías metabólicas relacionadas. Tiene potencial de aplicación en la producción de vacas.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio se incluyen en el artículo/material complementario, las consultas adicionales pueden dirigirse al autor correspondiente.
Declaración ética
El estudio en animales fue aprobado por todos los animales utilizados en este estudio fueron revisados y aprobados por el Comité de Ética de la Universidad Minzu de Mongolia Interior. (Número de Revisión de Ética: 2022092016001). El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
MX: Escritura – borrador original. YaW: Escritura – borrador original, Escritura – revisión y edición. MW: Redacción – borrador original, Redacción – revisión y edición. WP: Redacción – borrador original. YuW: Escritura – borrador original. RZ: Redacción – borrador original. YZ: Escritura – borrador original. JJ: Escritura – borrador original. CD: Escritura – borrador original. LD: Escritura – borrador original. MB: Escritura – borrador original.
Financiación
El/los autor/es declaran/n que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio obtuvo el «Proyecto Integrado de Demostración de Tecnologías Clave de Cría y Cría de Bovinos y Ovinos en la Zona de Transición de la Agricultura y Ganadería» (Nos. 16190050, 16200158 y 16210096); Apoyo al Proyecto de Apoyo a la Ciencia y la Tecnología de Mongolia Interior (Nº 2022YFXZ0015). Gracias por el apoyo financiero del proyecto «Desarrollo y demostración de tecnología de apoyo integral para la utilización eficiente y segura del forraje para vacas lecheras y saludables y de alto rendimiento».
Reconocimientos
En primer lugar, me gustaría agradecer al personal de la granja lechera experimental en Horqin Right Wing Front Banner, condado de Xing’an, Región Autónoma de Mongolia Interior, China, por su ayuda en el proceso de alimentación y recolección de muestras. Gracias a YW de la Universidad Agrícola de China por proporcionar condiciones experimentales y apoyo, gracias a MW de la Universidad Minzu de Mongolia Interior por su orientación en el experimento y el proceso de escritura. También me gustaría agradecer a los estudiantes del laboratorio por su ayuda en la recolección de muestras y la determinación del índice.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Notas
1. ^ https://www.kegg.jp/kegg/pathway.html
Referencias
1. Shen, X, Huo, B y Gan, S. Efectos del nano-selenio sobre la capacidad antioxidante en la gacela tibetana privada de se-se-(Procapra picticaudata) en la meseta Qinghai-Tíbet. Biol Trace Elem Res. (2021) 199:981–8. doi: 10.1007/s12011-020-02206-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Huang, Q, Wang, S, Yang, X, Han, X, Liu, Y, Khan, NA, et al. Efectos del selenio orgánico e inorgánico sobre la biodisponibilidad del selenio, el rendimiento del crecimiento, el estado antioxidante y la calidad de la carne de un ganado vacuno local en China. Front Vet Sci. (2023) 10:1171751. doi: 10.3389/fvets.2023.1171751
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Sordillo, LM. Estrategias nutricionales para optimizar la inmunidad del ganado lechero. J Dairy Sci. (2016) 99:4967–82. doi: 10.3168/jds.2015-10354
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Sun, LL, Gao, ST, Wang, K, Xu, JC, Sanz-Fernandez, MV, Baumgard, LH, et al. Efectos de la fuente sobre la biodisponibilidad del selenio, el estado antioxidante y el rendimiento en vacas lecheras lactantes durante condiciones de estrés oxidativo. J Dairy Sci. (2018) 102:311–9. doi: 10.3168/jds.2018-14974
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Mehdi, Y, y Dufrasne, I. Selenio en el ganado: una revisión. Moléculas. (2016) 21:545. doi: 10.3390/moléculas21040545
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Wang, S, Liu, X, Lei, L, Wang, D y Liu, Y. La deficiencia de selenio induce apoptosis, desequilibrio dinámico mitocondrial y respuestas inflamatorias en el hígado de ternero. Biol Trace Elem Res. (2022) 200:4678–89. doi: 10.1007/s12011-021-03059-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Hadrup, N, y Ravn-Haren, G. Absorción, distribución, metabolismo y excreción (ADME) de selenio oral de fuentes orgánicas e inorgánicas: una revisión. J Trace Elem Med Biol. (2021) 67:126801. doi: 10.1016/j.jtemb.2021.126801
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Kim, JH y Kil, DY. Comparación de los efectos tóxicos del selenio orgánico o inorgánico en la dieta y predicción de la ingesta de selenio y las concentraciones de selenio en los tejidos en pollos de engorde utilizando las concentraciones de selenio de las plumas. Poult Sci. (2020) 99:6462–73. doi: 10.1016/j.psj.2020.08.061
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Ye, R, Huang, J, Wang, Z, Chen, Y y Dong, Y. El oligoelemento selenio alivia eficazmente las enfermedades intestinales. Int J Mol Sci. (2021) 22:11708. doi: 10.3390/ijms222111708
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Zhang, L, Liu, XR, Liu, JZ, An, XP, Zhou, ZQ, Cao, BY, et al. El selenio orgánico e inorgánico suplementado afecta el rendimiento de la leche y la concentración de selenio en la leche y los tejidos de la cabra lechera de Guanzhong. Biol Trace Elem Res. (2018) 183:254–60. doi: 10.1007/s12011-017-1112-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Ferrari, L, Cattaneo, DM, Abbate, R, Manoni, M, Ottoboni, M, Luciano, A, et al. Avances en la suplementación con selenio: desde levaduras enriquecidas con selenio hasta posibles insectos enriquecidos con selenio y nanopartículas de selenio. Anim Nutr. (2023) 14:193–203. doi: 10.1016/j.aninu.2023.05.002
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Zhang, JS, Gao, XY, Zhang, LD y Bao, YP. Efectos biológicos de un nano selenio elemental rojo. Biofactores. (2001) 15:27–38. doi: 10.1002/biof.5520150103
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Wang, J, Chen, R, Xiang, L y Komarneni, S. Síntesis, propiedades y aplicaciones de nanomateriales de ZnO con vacantes de oxígeno: una revisión. Ceram Int. (2018) 44:7357–77. doi: 10.1016/j.ceramint.2018.02.013
14. Ahmadi, M, Ahmadian, A, Poorghasemi, M, Dong, X y Zou, X. Efectos del nanoselenio en las características del duodeno, yeyuno, íleon y colon en pollitos: un modelo animal. Int J Nano Dimensión. (2019) 10:225–229.
15. Yue, D, Zeng, C, Okyere, SK, Chen, Z y Hu, Y. El nanoselenio de glicina previene el estrés oxidativo cerebral y las anomalías neuroconductuales causadas por MPTP en ratas. J Trace Elem Med Biol. (2021) 64:126680. doi: 10.1016/j.jtemb.2020.126680
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Shakibaie, M, Shahverdi, AR, Faramarzi, MA, Hassanzadeh, GR, Rahimi, HR, Sabzevari, O, et al. Toxicidad aguda y subaguda de nuevas nanopartículas biogénicas de selenio en ratones. Pharm Biol. (2013) 51:58–63. doi: 10.3109/13880209.2012.710241
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Sun, XH, Lv, MW, Zhao, YX, Zhang, H, Saleem, MAU, Zhao, Y, et al. El nano-selenio antagonizó la fibrosis hepática inducida por cadmio en pollo. J Agric Food Chem. (2022) 71:846–56. doi: 10.1021/acs.jafc.2c06562
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Tian, J, Zhang, Y, Zhu, R, Wu, Y, Liu, X y Wang, X. El selenio elemental rojo (Se0) mejora la inmunoactividad de las células EPC, la carpa cruciana y el pez cebra contra la viremia primaveral del virus de la carpa. J Fish Biol. (2021) 98:208–18. doi: 10.1111/jfb.14571
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Cai, SJ, Wu, CX, Gong, LM, Song, T, Wu, H y Zhang, LY. Efectos del nano-selenio sobre el rendimiento, la calidad de la carne, la función inmunitaria, la resistencia a la oxidación y el contenido de selenio en los tejidos en pollos de engorde. Poult Sci. (2012) 91:2532–9. doi: 10.3382/ps.2012-02160
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Ahmadi, M, Ahmadian, A, y Seidavi, AR. Efecto de diferentes niveles de nano-selenio sobre el rendimiento, los parámetros sanguíneos, la inmunidad y las características de la canal de los pollos de engorde. Poult Sci J. (2018) 6:99–108. doi: 10.22069/psj.2018.13815.1276
21. Shi, L, Xun, W, Yue, W, Zhang, C, Ren, Y, Liu, Q, et al. Efecto del nano-selenio elemental sobre la digestibilidad del alimento, la fermentación ruminal y los derivados de purinas en ovejas. Anim Feed Sci Technol. (2011) 163:136–42. doi: 10.1016/j.anifeedsci.2010.10.016
22. Shi, L, Xun, W, Yue, W, Zhang, C, Ren, Y, Shi, L, et al. Efecto del selenito de sodio, la levadura de Sed y el selenio nanoelemental sobre el rendimiento del crecimiento, la concentración de Se y el estado antioxidante en cabras macho en crecimiento. Pequeña Rumin Res. (2011) 96:49–52. doi: 10.1016/j.smallrumres.2010.11.005
23. Han, L, Pang, K, Fu, T, Phillips, CJ y Gao, T. La suplementación con nano-selenio aumenta los perfiles de expresión génica de selenoproteína (Sel) y la concentración de selenio en leche en vacas lecheras lactantes. Biol Trace Elem Res. (2021) 199:113–9. doi: 10.1007/s12011-020-02139-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Yang, C, Yao, H, Wu, Y, Sun, G, Yang, W, Li, Z, et al. Estado y riesgos de la deficiencia de selenio en un área tradicionalmente deficiente de selenio en el noreste de China. Sci Total Environ. (2021) 762:144103. doi: 10.1016/j.scitotenv.2020.144103
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Dinh, QT, Cui, Z, Huang, J, Tran, TA, Wang, D, Yang, W, et al. Distribución del selenio en el medio ambiente chino y su relación con la salud humana: una revisión. Environ Int. (2018) 112:294–309. doi: 10.1016/j.envint.2017.12.035
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Hu, T, Li, L, Hui, G, Zhang, J, Li, H, Wu, W, et al. Biofortificación con selenio y su efecto sobre el cambio multielemento en Auricularia auricular. Químico de los alimentos. (2019) 295:206–13. doi: 10.1016/j.foodchem.2019.05.101
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Zhang, L, Gao, R, Zhang, H, Luo, X y Song, Z. Estudio sobre la influencia de la intención de compra de los consumidores de productos agrícolas ricos en selenio. Appl Sci. (2023) 13:1859. doi: 10.3390/app13031859
28. Knowles, SO, Grace, ND, Knight, TW, McNabb, WC y Lee, J. Adición de valor nutricional a la carne y la leche de ganado alimentado con pastos. Zelandia Vet J. (2004) 52:342–51. doi: 10.1080/00480169.2004.36450
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Ferreira, GM, y Petzer, I-M. Selenio orgánico e inorgánico inyectable en vacas lecheras: efectos sobre la leche, la sangre y los niveles de recuento de células somáticas. Onderstepoort J Vet Res. (2019) 86:e1–8. doi: 10.4102/ojvr.v86i1.1664
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Consejo Nacional de Investigación, Comité de Nutrición Animal y Subcomité de Nutrición del Ganado Lechero. (2001) Requerimientos de nutrientes del ganado lechero. Editorial de las Academias Nacionales.
31. AOAC International ed. Métodos oficiales de análisis. 18ª ed. Gaithersburg, MD, EE.UU.: AOAC International. (2005).
32. Soest, PV, Robertson, J y Lewis, B. Métodos para la fibra dietética, la fibra detergente neutra y los polisacuridos sin almidón en relación con la nutrición animal. J Dairy Sci. (1991) 74:3583–97. doi: 10.3168/jds. S0022-0302(91)78551-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Tinggi, U. Esencialidad y toxicidad del selenio y su estado en Australia: una revisión. Toxicol Lett Toxicol Lett. (2003) 137:103–10. doi: 10.1016/s0378-4274(02)00384-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Tufarelli, V, y Laudadio, V. La suplementación dietética con selenio y vitamina E mejora la producción de leche, la composición y las propiedades reológicas de las cabras lecheras Jonica. J Dairy Res. (2011) 78:144–8. doi: 10.1017/S0022029910000907
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Consejo Superior de Investigaciones Científicas. Requerimientos de nutrientes del ganado lechero. Washington DC: National Academies Press (2001).
36. Sun, P, Wang, J, Liu, W, Bu, DP, Liu, SJ y Zhang, KZ. Hidroxi-selenometionina: una nueva fuente orgánica de selenio que mejora el estado antioxidante y las concentraciones de selenio en la leche y el plasma de vacas lecheras a mitad de lactancia. J Dairy Sci. (2017) 100:9602–10. doi: 10.3168/jds.2017-12610
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Juniper, DT, Phipps, RH, Jones, AK y Bertin, G. Suplementación con selenio de vacas lecheras lactantes: efecto sobre la concentración de selenio en sangre, leche, orina y heces. J Dairy Sci. (2006) 89:3544–51. doi: 10.3168/jds. S0022-0302(06)72394-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Juniper, DT, Phipps, RH, Givens, DI, Jones, AK, Green, C y Bertin, G. Tolerancia de los animales rumiantes a la administración de altas dosis en el alimento de una levadura enriquecida con selenio. J Anim Sci. (2008) 86:197–204. doi: 10.2527/jas.2006-773
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Hachemi, MA, Sexton, JR, Briens, M, y Whitehouse, NL. Eficacia de la alimentación con hidroxi-selenometionina en plasma y selenio lácteo en vacas lecheras a mitad de lactancia. J Dairy Sci. (2023) 106:2374–85. doi: 10.3168/jds.2022-22323
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Zhang, ZD, Wang, C, Du, HS, Liu, Q, Guo, G, Huo, WJ, et al. Efectos del selenito de sodio y el selenito de sodio recubierto sobre el rendimiento de la lactancia, la digestión de nutrientes del tracto total y la fermentación ruminal en vacas lecheras Holstein. Animal. (2020) 14:2091–9. doi: 10.1017/S1751731120000804
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Wang, C, Liu, Q, Yang, WZ, Dong, Q, Yang, XM, He, DC, et al. Efectos de la levadura de selenio sobre la fermentación ruminal, el rendimiento de la lactancia y la digestibilidad del alimento en vacas lecheras lactantes. Livest Sci. (2009) 126:239–44. doi: 10.1016/j.livsci.2009.07.005
42. Gross, JJ. Fisiología de las vacas lecheras y límites de producción. Frente de animación. (2023) 13:44–50. doi: 10.1093/af/vfad014
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Vonnahme, KA, Wienhold, CM, Borowicz, PP, Neville, TL, Redmer, DA, Reynolds, LP, et al. El selenio supranutricional aumenta la vascularización de la glándula mamaria en corderas posparto. J Dairy Sci. (2011) 94:2850–8. doi: 10.3168/jds.2010-3832
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Sizova, Е, Miroshnikov, S, Lebedev, S, Usha, B y Shabunin, S. Uso de metales a nanoescala en la dieta de las aves de corral como aditivo mineral para piensos. Frente de animación. (2020) 6:185–91. doi: 10.1016/j.aninu.2019.11.007
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Neville, TL, Meyer, AM, Reyaz, A, Borowicz, PB, Redmer, DA, Reynolds, LP, et al. Crecimiento de la glándula mamaria y vascularización durante el parto y durante la lactancia en ovejas primíparas alimentadas con diferentes niveles de selenio y plano nutricional durante la gestación. J Anim Sci Biotechnol. (2013) 4:6. doi: 10.1186/2049-1891-4-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Gong, J y Xiao, M. Selenio y estado antioxidante en vacas lecheras en diferentes etapas de lactancia. Biol Trace Elem Res. (2015) 171:89–93. doi: 10.1007/s12011-015-0513-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Mahan, DC, Azain, M, Crenshaw, TD, Cromwell, GL, Dove, CR y Kim, SW. Suplementación de selenio orgánico e inorgánico a dietas que utilizan granos cultivados en varias regiones de los Estados Unidos con diferentes concentraciones naturales de se y alimentados con cerdos de engorde-engorde. J Anim Sci. (2014) 92:4991–7. doi: 10.2527/jas.2014-7735
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Gong, J y Xiao, M. Efecto de la suplementación con selenio orgánico sobre el estado del selenio, el estrés oxidativo y el estado antioxidante en vacas lecheras adecuadas al selenio durante el período periparto. Biol Trace Elem Res. (2018) 186:430–40. doi: 10.1007/s12011-018-1323-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Lin, X, Yang, T, Li, H, Ji, Y, Zhao, Y y He, J. Interacciones entre diferentes compuestos de selenio y oligoelementos esenciales involucrados en el sistema antioxidante de las gallinas ponedoras. Biol Trace Elem Res. (2019) 193:252–60. DOI: 10.1007/s12011-019-01701-x
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Ortman, K, y Pehrson, B. Efecto del selenato como suplemento alimenticio para las vacas lecheras en comparación con el selenito y la levadura de selenio. J Anim Sci. (1999) 77:3365–70. doi: 10.2527/1999.77123365x
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Calamari, L, Petrera, F y Bertin, G. Efectos de la suplementación con selenito de sodio o levadura de sodio (Sc CNCM I-3060) sobre el estado del selenio y las características de la leche en vacas lecheras. Livest Sci. (2010) 128:154–65. doi: 10.1016/j.livsci.2009.12.005
52. Stewart, WC, Bobe, G, Pirelli, GJ, Mosher, WD y Hall, JA. Selenio orgánico e inorgánico: III. Rendimiento de las ovejas y la progenie. J Anim Sci. (2012) 90:4536–43. doi: 10.2527/jas.2011-5019
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Rayman, diputado. Ingesta de selenio, estado y salud: una relación compleja. Hormonas. (2020) 19:9–14. doi: 10.1007/s42000-019-00125-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Prabhu, KS, y Lei, XG. Selenio. Adv Nutr. (2016) 7:415–7. doi: 10.3945/an.115.010785
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Avery, J y Hoffmann, P. Selenio, selenoproteínas e inmunidad. Nutrientes. (2018) 10:1203. doi: 10.3390/nu10091203
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Hall, JA, Bobe, G, Vorachek, WR, Estill, CT, Mosher, WD, Pirelli, GJ, et al. Efecto de la suplementación supranutricional con selenio materno y calostro sobre la absorción pasiva de inmunoglobulina G en terneros lecheros repletos de selenio. J Dairy Sci. (2014) 97:4379–91. doi: 10.3168/jds.2013-7481
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. Phipps, RH, Grandison, AS, Jones, AK, Juniper, DT, Ramos-Morales, E y Bertin, G. Suplementación con selenio de vacas lecheras lactantes: efectos sobre la producción de leche y el contenido total de selenio y especiación en sangre, leche y queso. Animal. (2008) 2:1610–8. doi: 10.1017/S175173110800298X
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Zhang, X, Zuo, Z, Liu, Y, Wang, C, Peng, Z, Zhong, J, et al. Efecto de los análogos de metionina sobre el rendimiento del crecimiento, los parámetros bioquímicos séricos, los aminoácidos libres séricos y la fermentación ruminal de los yaks. Animales. (2022) 12:3175. doi: 10.3390/ani12223175
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Wang, BM y Li, HX. Efectos de dietas con diferentes niveles de nitrógeno sobre el nitrógeno ureico plasmático en yaks. Gansu Anim Husbandry Vet Med. (2015) 45:44–7. doi: 10.15979/j.cnki.cn62-1064/s.2015.06.030
60. Zulu, VC, Sawamukai, Y, Nakada, K, Kida, K y Moriyoshi, M. Relación entre el factor de crecimiento similar a la insulina-I, los metabolitos sanguíneos y la función ovárica posparto en vacas lecheras. J Vet Med Sci. (2002) 64:879–85. doi: 10.1292/jvms.64.879
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Sosa, I, Leyton, L, Corea, E, et al. Correlación entre la leche y el nitrógeno ureico en sangre en vacas lecheras de alto y bajo rendimiento. Int Atómica. FAO. (2010):79–82.
62. Bayssa, M, Yigrem, S, Betsha, S, y Tolera, A. Producción, reproducción y algunas características de adaptación de la raza bovina Boran bajo clima cambiante: una revisión sistemática y meta-análisis. PLoS Uno. (2021) 16:e0244836. doi: 10.1371/journal.pone.0244836
Resumen de PubMed | Texto completo de Crossref | Google Académico
63. Fernández, Nueva Jersey, y Riñón, BA. Fosfatasa alcalina: más allá del hígado. Veterinario Clin Pathol. (2007) 36:223–33. doi: 10.1111/j.1939-165x.2007.tb00216.x
64. Diskin, CJ, Tomasso, CL, Alper, JC, Glaser, ML y Fliegel, SE. Exposición al selenio a largo plazo. Arch Intern Med. (1979) 139:824–6. doi: 10.1001/archinte.1979.03630440082028
65. Shao, H, Tao, M, Fan, Y, Jing, J y Lu, J. Niveles de vitamina D y otros factores relacionados con la densidad mineral ósea durante el embarazo. Aust N Z J Obstet Gynaecol. (2012) 52:571–5. doi: 10.1111/j.1479-828X.2012.01477.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
66. Gunter, SA, Beck, PA, y Hallford, DM. Efectos de la fuente suplementaria de selenio en los parámetros sanguíneos en vacas de carne y sus terneros lactantes. Biol Trace Elem Res. (2013) 152:204–11. doi: 10.1007/s12011-013-9620-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
67. Juniper, DT, Rymer, C y Briens, M. Bioeficacia de la hidroxi-selenometionina como suplemento de selenio en novillas lecheras preñadas y sobre el estado de selenio de sus terneros. J Dairy Sci. (2019) 102:7000–7010. doi: 10.3168/jds.2018-16065
Resumen de PubMed | Texto completo de Crossref | Google Académico
68. Schrauzer, GN. Selenometionina: una revisión de su importancia nutricional, metabolismo y toxicidad. J Nutr. (2000) 130:1653–6. doi: 10.1093/jn/130.7.1653
Resumen de PubMed | Texto completo de Crossref | Google Académico
69. Jafarzadeh, H, Allymehr, M, Talebi, A, Asri-Rezaie, S, y Soleimanzadeh, A. Los efectos del nano-selenio y el selenito de sodio sobre la concentración sérica de selenoproteína P y GPx en machos reproductores de pollos de engorde. Anim Sci Res. (2023). doi: 10.22034/AS.2023.51270.1660
70. Givens, DI, Allison, R, Cottrill, B, y Blake, JS. Aumento del contenido de selenio de la leche bovina BUN a través de la alteración de la forma y concentración de selenio en la dieta de la vaca lechera. J Sci Food Agric. (2004) 84:811–7. doi: 10.1002/jsfa.1737
71. Haug, A, Høstmark, AT, y Harstad, OM. La leche bovina en la nutrición humana: una revisión. Lípidos Salud Dis. (2007) 6:25. doi: 10.1186/1476-511X-6-25
Resumen de PubMed | Texto completo de Crossref | Google Académico
72. Harding, JJ, Hassett, P, Rixon, KC, Bron, AJ y Harvey, DJ. Azúcares, incluidos los ácidos eritrónico y treónico, en el humor acuoso humano. Curr Eye Res. (1999) 19:131–6. doi: 10.1076/ceyr.19.2.131.5334
Resumen de PubMed | Texto completo de Crossref | Google Académico
73. Aguilar, F, Autrup, H, Barlow, S, Castle, L, Crebelli, R, Dekant, W, et al. Ascorbato de calcio con contenido en treonato para su uso como fuente de vitamina C en complementos alimenticios. EFSA J. (2007) 5:1–10. doi: 10.2903/j.efsa.2007.491
74. Kokotou, MG, Mantzourani, C, Asimina, B, Mountanea, OG, y Kokotos, G. Método de cromatografía líquida y espectrometría de masas de alta resolución (LC-HRMS) para la determinación de hidroxiácidos grasos libres en leche de vaca y cabra. Moléculas. (2020) 25:3947. doi: 10.3390/moléculas25173947
Resumen de PubMed | Texto completo de Crossref | Google Académico
75. Funato, K, Yamashita, C, Kamada, J, Tominaga, S y Kiwada, H. Factor plasmático que desencadena la activación de la vía alternativa del complemento por liposomas. Pharm Res. (1994) 11:372–6. doi: 10.1023/A:1018952718496
Resumen de PubMed | Texto completo de Crossref | Google Académico
76. Shi, D, Zhao, Y, Yan, H, Fu, H, Shen, Y, Lu, G, et al. Efectos antifúngicos del ácido undecilénico en la formación de biopelículas de Candida albicans. Int J Clin Pharmacol Ther. (2016) 54:343–53. doi: 10.5414/CP202460
Resumen de PubMed | Texto completo de Crossref | Google Académico
77. Martins, LC, Monteiro, CC, Semedo, PM y Sá-Correia, I. Valorización de residuos agroindustriales ricos en pectina por levaduras: potencial y desafíos. Aplicación Microbiol Biotechnol. (2020) 104:6527–47. doi: 10.1007/s00253-020-10697-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
78. McSweeney, PL, y Fox, PF. Química láctea avanzada volumen 3: lactosa, agua, sales y vitaminas. Springer Science & BusinessMedia. Nueva York: Springer New York (2013).
79. Velioglu, YS. 10 ácidos alimentarios: ácidos orgánicos, ácidos orgánicos volátiles y ácidos fenólicos. Adv Food Biochem. (2009):313. doi: 10.1201/9781420007695-c10
80. Hirabayashi, J. Sobre el origen de las hexosas elementales. Q Rev Biol. (1996) 71:365–80. doi: 10.1086/419443
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: vaca, nanoselenio, indicadores sanguíneos, selenio sanguíneo, selenio lechero, metabolitos lácteos
Cita: Xiao M, Wang Y, Wei M, Peng W, Wang Y, Zhang R, Zheng Y, Ju J, Dong C, Du L y Bao M (2024) Efectos del nanoselenio en el rendimiento, los índices sanguíneos y los metabolitos de la leche de las vacas lecheras durante el período pico de lactancia. Frente. Vet. Sci. 11:1418165. doi: 10.3389/fvets.2024.1418165
Editado por:
Panagiotis E. Simitzis, Universidad Agrícola de Atenas, Grecia
Revisado por:
Shuanlin Zhang, Universidad Agrícola de Shanxi, China
Hangshu Xin, Universidad Agrícola del Noreste, China
Derechos de autor © 2024 Xiao, Wang, Wei, Peng, Wang, Zhang, Zheng, Ju, Dong, Du y Bao. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Manlin Wei, weimanlin@163.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía