
Impacto de la reducción de la proteína bruta dietética en la fase inicial sobre el desarrollo inmunológico

Impacto de la reducción de la proteína bruta dietética en la fase inicial sobre el desarrollo inmunológico y la respuesta de los pollos de engorde a lo largo del período de crecimiento
 Mohammad Kamely1
Mohammad Kamely1 Wanwei He2
Wanwei He2 Jeremy Wakaruk1Rosa Whelan3Víctor Naranjo3
Jeremy Wakaruk1Rosa Whelan3Víctor Naranjo3 Daniel R. Barreda1,2*
Daniel R. Barreda1,2*- 1Departamento de Ciencias Agrícolas, Alimentarias y Nutricionales, Universidad de Alberta, Edmonton, AB, Canadá
- número arábigoDepartamento de Ciencias Biológicas, Universidad de Alberta, Edmonton, AB, Canadá
- 3Evonik Nutrition & Care GmbH, Hanau, Alemania
Los niveles de proteína bruta (PC) en las dietas comerciales de pollos de engorde (Gallus gallus), optimizados para la producción de rendimiento máximo frente al costo del alimento, solo han comenzado a evaluarse para determinar su impacto en la función inmunológica. Con el fin de estudiar los efectos inmunológicos de los niveles de PC en la dieta, se administraron diferentes dietas de fase inicial (día 1-14) a 230 pollos de engorde machos Ross 708 asignados aleatoriamente a 1 día de edad en dos grupos de tratamiento. Grupo 1: La dieta estándar (ETS) contenía 3.000 kcal de AMEn/kg de energía y 23,78% de PC; y Grupo 2: La dieta reducida en proteína bruta (PCR) contenía 3.000 kcal de AMEn/kg de energía y 21,23% de PC. A partir del día 15 al 35 se asignó a ambos grupos una dieta estándar común para el engorde (3.150 kcal AMEn/kg de energía y 22,18% de PC). El zymosan, un glicano derivado de las paredes celulares de la levadura que se une a TLR 2 y Dectina-1, se utilizó para el desafío intraabdominal. Los resultados demostraron que una dieta inicial reducida de proteína cruda (21,23 vs. 23,78% PC) entre los 1 y los 14 años, mientras se mantenían los mismos niveles de energía metabolizable y aminoácidos esenciales, no afectó el rendimiento del crecimiento de los pollos de engorde ni el peso de los órganos linfoides (P > 0,05). Curiosamente, los niveles basales de leucocitos en el grupo RCP aumentaron significativamente (P < 0,01) en el compartimento sanguíneo a d35 en las aves no desafiadas. También se detectaron mejoras significativas en la infiltración de leucocitos en la cavidad abdominal después del desafío inmune con zymosan (día 14 y día 35; P < 0,01). Los niveles posteriores al desafío de las células de expresión génica TNF-α, IL-1β y CXCL8 recolectadas de la cavidad abdominal no se vieron afectados por las dietas (P > 0,05). Además, los tratamientos dietéticos no influyeron en el porcentaje de células productoras de ROS en la cavidad abdominal (P > 0,05). Hasta donde sabemos, este es el primer estudio que informa sobre los impactos de una dieta reducida en proteínas brutas en la respuesta inmune innata de las aves de corral a un modelo de inflamación aguda en la cavidad abdominal. En general, nuestros resultados destacan que las dietas reducidas en proteínas brutas se pueden utilizar sin afectar negativamente el rendimiento de los pollos de engorde y pueden mejorar la capacidad de los pollos de engorde para reclutar leucocitos tras la infección.
Introducción
Los pollos de engorde han estado expuestos a una intensa selección genética para mejorar los rasgos de rendimiento, principalmente por el crecimiento rápido y la eficiencia de la utilización del alimento, y por lo tanto pueden tener efectos perjudiciales sostenidos en el sistema inmunológico (1, 2). Ya se han detectado diferencias en el sistema inmune adaptativo (3, 4). Por ejemplo, la cepa de pollos de engorde de 1957 tiene niveles más altos de anticuerpos anti-SRBC (Ig e IgM totales) que la cepa de 1991 cuando se desafió con SRBC a las 2 semanas de edad (5). Las respuestas inmunitarias innatas, cruciales para limitar la difusión del patógeno tras la entrada en un huésped (6), son menos conocidas en lo que respecta a los cambios en la funcionalidad debidos a la selección genética. En un ejemplo, la respuesta a la proteína en fase aguda se mantuvo sin cambios entre los pollos de engorde de 1970, 1990 y los pollos de engorde modernos (7). Por el contrario, la función de los heterófilos mostró una alta variabilidad entre líneas genéticamente distintas con respecto a la fagocitosis, las trampas extracelulares de heterófilos y la expresión de citocinas (8, 9).
Otro factor importante que modula la función inmunitaria, al tiempo que interactúa con la composición genética, es la composición de la dieta. Las fuentes de proteínas dietéticas suelen ser un componente de alto costo en las dietas de las aves de corral. Dado que los animales requieren aminoácidos específicos, las formulaciones que requieren aminoácidos específicos a menudo conducen a niveles de proteína bruta (PC) que son demasiado costosos y dan como resultado que el exceso de nitrógeno se excreta como contaminante ambiental (10). Por lo tanto, la industria ha dado pasos para formular dietas sobre una base de aminoácidos digeribles, en lugar de formular sobre la base de niveles de proteína bruta, que son mucho menos precisos (11). La reducción resultante de la proteína bruta es posible gracias a la suplementación de la dieta con fuentes de aminoácidos cristalinos para suministrar suficientes aminoácidos esenciales limitantes (11-13). Si bien se han llevado a cabo investigaciones exhaustivas para determinar los requisitos estándar de aminoácidos digeribles ileales para todas las fases de alimentación de los pollos de engorde en condiciones saludables (14), los requisitos para un desarrollo inmunológico óptimo, o durante un desafío, siguen siendo en gran medida desconocidos (15). Además, dado que las tasas de inclusión de nutrientes en las dietas comerciales no suelen ser mayores de lo necesario para maximizar la eficiencia de la producción, generalmente no se sabe si esto es suficiente para mantener un sistema inmunológico robusto.
La observación de las respuestas inmunitarias después de la inyección intraperitoneal/intraabdominal in vivo de zymosan ya ha contribuido a nuestra comprensión del estado inmunitario en estudios comparativos con animales que incluyen peces, cerdos, ratones y pollos (16-18). En este modelo, el zymosan sirve como un estimulador inmunológico fúngico y, en condiciones de resolución automática, permite examinar las fases de inducción y control de la inflamación aguda. La cavidad abdominal de las aves proporciona un entorno adecuado para estudiar las contribuciones de los leucocitos migratorios a la inducción y regulación de las respuestas inmunitarias antimicrobianas. El fenotipo y el número de leucocitos infiltrantes se pueden examinar después de un desafío inmunológico in vivo, y los leucocitos infiltrantes se pueden recolectar fácilmente para exámenes ex vivo. Para este estudio, se utilizó este modelo para evaluar el crecimiento y la función inmune de los pollos de engorde, comparando los pollos de engorde criados con una dieta inicial reducida de proteína bruta frente a una dieta comercial estándar de nivel de proteína bruta para la fase inicial. Se monitorizó el crecimiento y la función inmunitaria durante 35 días.
Materiales y métodos
Animales y Diseño Experimental
Un total de 230 pollos de engorde machos Ross 708 machos (Sofina Foods, División Lilydale, Hatchery de Edmonton) fueron asignados aleatoriamente a 1 día de edad en dos grupos de tratamiento dietético: proteína cruda estándar (STD) y proteína bruta reducida (RCP). Con el fin de maximizar la calidad de los datos y minimizar el impacto asociado al estrés, se utilizaron plumas paralelas para las mediciones inmunológicas y de rendimiento. Se utilizaron ocho réplicas de corrales en total, con seis corrales de 15 aves (tres corrales por tratamiento) para las mediciones del rendimiento del crecimiento (total de 90 aves) y dos corrales de 70 aves (un corral por tratamiento) para parámetros inmunológicos (total de 140 aves). Los grupos de tratamiento dietético se establecieron en un patrón de corral alterno dentro del establo. Todas las aves fueron alojadas en el Centro de Investigación Avícola del Departamento de Ciencias Agrícolas, Alimentarias y Nutricionales de la Universidad de Alberta. Todos los animales fueron mantenidos de acuerdo con las pautas especificadas por el Consejo Canadiense de Cuidado Animal, y los protocolos fueron aprobados por el Comité de Cuidado y Uso de Animales de la Universidad de Alberta.
Las dietas (iniciador de crumble, cultivador granulado) y agua se proporcionaron ad libitum durante todo el experimento. Del día 1 al 14 se asignaron dos dietas de inicio diferentes con diferentes niveles de PC, con proporciones de aminoácidos equilibradas según las recomendaciones de Evonik (AMINOChick®). La dieta RCP contenía 3.000 kcal AMEn/kg y 21,23% PC, mientras que la ETS contenía 3.000 kcal AMEn/kg y 23,78% PC. De los 15 a los 35 días se asignó a todos los grupos la misma dieta estándar de engorde/engorde (3.150 kcal AMEn/kg y 22,18% PC). En la Tabla 1 se muestran la formulación del alimento y la composición nutricional de las dietas de inicio (1-14 días de edad) y de engorde (15-35 días de edad).
Ganancia de peso corporal, ingesta de alimento y tasa de conversión alimenticia
El peso corporal se midió semanalmente para cada corral. La ingesta de alimento se determinó como la diferencia entre la cantidad de alimento ofrecido y la cantidad no consumida en la fase de inicio y crecimiento. La ingesta diaria de alimento (IDF) se calculó dividiendo el alimento consumido de cada corral en la fase de inicio y crecimiento por el número total real de aves. El índice de conversión alimenticia (FCR: g de alimento/g de ganancia de peso corporal) se calculó dividiendo la ingesta diaria de alimento por la ganancia diaria de peso corporal.
Pesos de los órganos linfoides
El fémur, el timo, la bolsa de Fabricio y el peso del bazo se midieron a los 7, 14, 28 y 35 días de edad de cinco aves por grupo. Se pesaron estos órganos y los pollitos correspondientes, y el peso relativo de los órganos se expresó como porcentaje del peso corporal (PC).
Inyecciones intraabdominales
La inducción de una inflamación aguda intraabdominal se derivó de un protocolo personalizado previamente validado en nuestro laboratorio (18). Brevemente, el zymosan A de Saccharomyces cerevisiae (2,5 mg, Sigma-Aldrich) se resuspendió en 500 ml de PBS−/− (sin calcio/sin magnesio) y se administró mediante inyección en la cavidad abdominal para inducir una respuesta inflamatoria. Las aves que no recibieron una inyección a las 0 h se utilizaron como grupo control. Esta dosis inyectable permitió el establecimiento de condiciones de autoresolución, donde se pudieron observar fases de inducción temprana y resolución tardía de la inflamación aguda.
Aislamiento de sangre periférica, médula ósea y leucocitos intraabdominales
Se tomaron muestras de sangre periférica completa mediante venopunción de venopura de vena alar. Con el fin de determinar los impactos inmunológicos a corto y largo plazo de las dietas, se midió el número total de leucocitos, heterófilos, monocitos/macrófagos y linfocitos a las 0, 4 y 12 h después de las inyecciones intraabdominales de zymosan en los días 14 y 35 en la médula ósea y la cavidad abdominal. Los leucocitos de médula ósea se obtuvieron después de la recolección del fémur, donde los huesos se lavaron con 20 ml de medio estéril RPMI-1640. Se extrajeron glóbulos rojos de muestras de sangre y médula ósea mediante tratamiento con un tampón de lisis de glóbulos rojos (ACK Gibco, EE. UU.) (proporción 1:10). Los leucocitos intraabdominales se recuperaron inyectando 10 y 20 ml de medio RPMI-1640 los días 14 y 35, respectivamente. Las células se recolectaron y mantuvieron a 4 °C. Los pollos que no recibieron inyección se utilizaron como control negativo (aves 0 h). Los leucocitos totales en sangre se contaron en una cámara de hemocitómetro utilizando la solución de Natt y Herrick para obtener una dilución de sangre 1:200 (19).
Microscopía y tinción celular
Los leucocitos (1 × 105 células) de sangre periférica, médula ósea e intraabdominal se centrifugaron en portaobjetos de vidrio a 55 g durante 6 min a temperatura ambiente utilizando una citocentrífuga (Thermo Scientific ™ Cytospin ™ 4) y se tiñeron con el set de tinción Hema3 (Fisher Scientific) de acuerdo con las especificaciones del fabricante. Las imágenes se generaron utilizando un microscopio DM1000 (Leica, Wetzler, Alemania) y un objetivo de campo claro de 100× (aumento total de 1.000×).
Oxidación y tinción de anticuerpos de CellROX
La producción de ROS se determinó con base en el procedimiento de oxidación CellROX (Molecular Probes). Brevemente, se incubaron 1 × 106 leucocitos intraabdominales con 5 μM de reactivo CellROD, seguido de una incubación de 30 minutos a 41 °C. Los leucocitos se lavaron dos veces con PBS−/− y se fijaron en formaldehído al 1%. Los datos se adquirieron en un citómetro de flujo de imágenes multiespectrales ImageStream MKII (Amnis Corporation, EMD Millipore, Seattle, WA, EE. UU.) y se analizaron utilizando el software INSPIRE (Amnis Corporation, EMD Millipore; Seattle, WA, EE.UU.).
Expresión génica
La expresión génica se realizó como se describió anteriormente (17, 20). Brevemente, las células de lavado abdominal se recolectaron con RPMI 1640 y se centrifugaron y mantuvieron dentro de 1 mL de reactivo de Trizol (Invitrogen Life Technologies) en nitrógeno líquido. El ARN total se extrajo y se transcribió inversamente en ADNc utilizando la transcriptasa inversa SMARTScribe (Clontech) de acuerdo con los protocolos del fabricante. La PCR cuantitativa se ejecutó en el sistema ABI 7500 (Applied Biosystems, Carlsbad, CA, USA). La concentración y pureza del ARN se determinaron mediante el uso de un espectrofotómetro NanoDrop y un bioanalizador 2100. Los datos se normalizaron a pollo a partir del tiempo 0 h. El gen ARN ribosómico 28S se utilizó como interno y de normalización. Cada muestra se realizó por triplicado. Los niveles de expresión relativa de los genes diana se calcularon según el método 2-ΔΔCt. Las secuencias de cebadores para cada gen se muestran en la Tabla 2.

Análisis estadístico
La evaluación de la significancia se basó en análisis de análisis de varianza de dos vías (ANOVA) con el punto de tiempo y el tratamiento como efectos principales, utilizando las pruebas de comparación múltiple de Sidak en el software Prism 8 (GraphPad Software, La Jolla, CA, USA). También se realizaron pruebas post hoc adicionales de Tukey para analizar las diferencias dentro de los tratamientos y el tiempo después de la provocación intraabdominal. Se analizaron los datos de la bolsa de Fabricio, timo, fémur, bazo, peso corporal, ingesta de alimento y índice de conversión alimenticia mediante t-test. Los estadísticos con P < 0,05 se consideraron significativos.
Resultados y Discusión
Rendimiento y peso de los órganos linfoides
La reducción de la proteína cruda de la dieta en la fase de inicio de los pollos de engorde no afectó significativamente la ganancia de peso corporal, la ingesta de alimento o la tasa de conversión alimenticia en la fase de inicio o más allá hasta los 35 días de edad (P > 0.05; Tabla 3). Los pollos de engorde alimentados con una dieta RCP tendieron a tener una menor ganancia de peso corporal entre d 1 y d 28, pero compensada entre d 28 y d 35 (Tabla 3). Esto es consistente con investigaciones previas que han demostrado que las dietas reducidas en proteínas en la fase inicial no tienen un impacto negativo en el rendimiento, y además, el menor peso corporal se compensará después de la cuarta semana (19, 21, 22). Esto también es consistente con informes anteriores que señalan la suplementación con aminoácidos específicos en lugar de CP como lo más crítico para el rendimiento de los pollos de engorde. El mantenimiento de proporciones óptimas de aminoácidos para los aminoácidos esenciales y niveles suficientes de equivalente de glicina parecen ser lo más importante (13, 14, 21, 23-25). Con base en esto, la alimentación con dietas RCP formuladas para cumplir con los requisitos de aminoácidos digeribles podría ser una forma eficiente de reducir la excreción de nitrógeno al medio ambiente y disminuir el costo de la alimentación sin afectar el rendimiento del crecimiento.
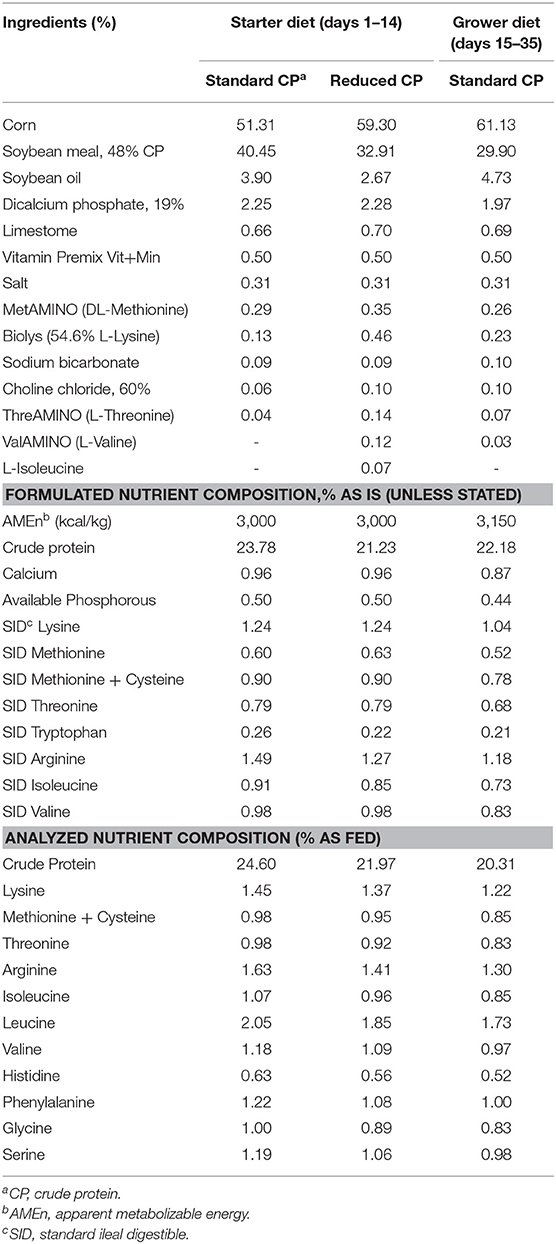
La médula ósea y el bazo constituyen órganos inmunorreticantes relevantes que, entre otros, albergan reservas de poblaciones maduras de leucocitos naïve que pueden desplegarse rápidamente tras la infección (5, 6). Los órganos linfoides primarios de los pollos (bursa y timo) proporcionan el entorno de desarrollo para las células B y T, respectivamente (26), y se ha demostrado que la deficiencia de aminoácidos o el exceso de proteínas dietéticas cambian la respuesta inmunitaria en las aves de corral (27). Sin embargo, la suplementación con la dieta RCP no afectó el desarrollo de los órganos linfoides (Figura 1). No se observaron diferencias significativas entre la bolsa, el timo, el bazo o el fémur, lo que sugiere que no hay deficiencia en los componentes nutricionales necesarios para el desarrollo adecuado de los órganos linfoides. Esto es consistente con investigaciones previas que mostraron que la disminución moderada en el nivel de PC (rango entre 18 y 23%) no tuvo ningún efecto sobre el peso de la bursa, el timo y el bazo (19, 28, 29).
Reclutamiento de leucocitos en el sitio de desafío abdominal y producción de ROS después del desafío con zymosan
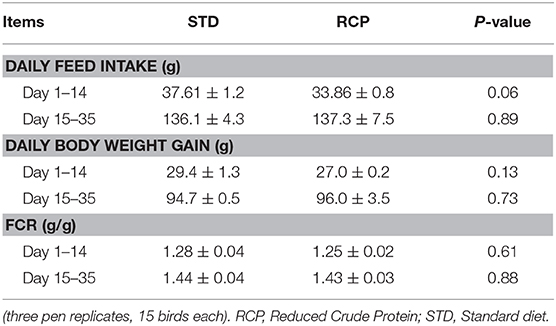
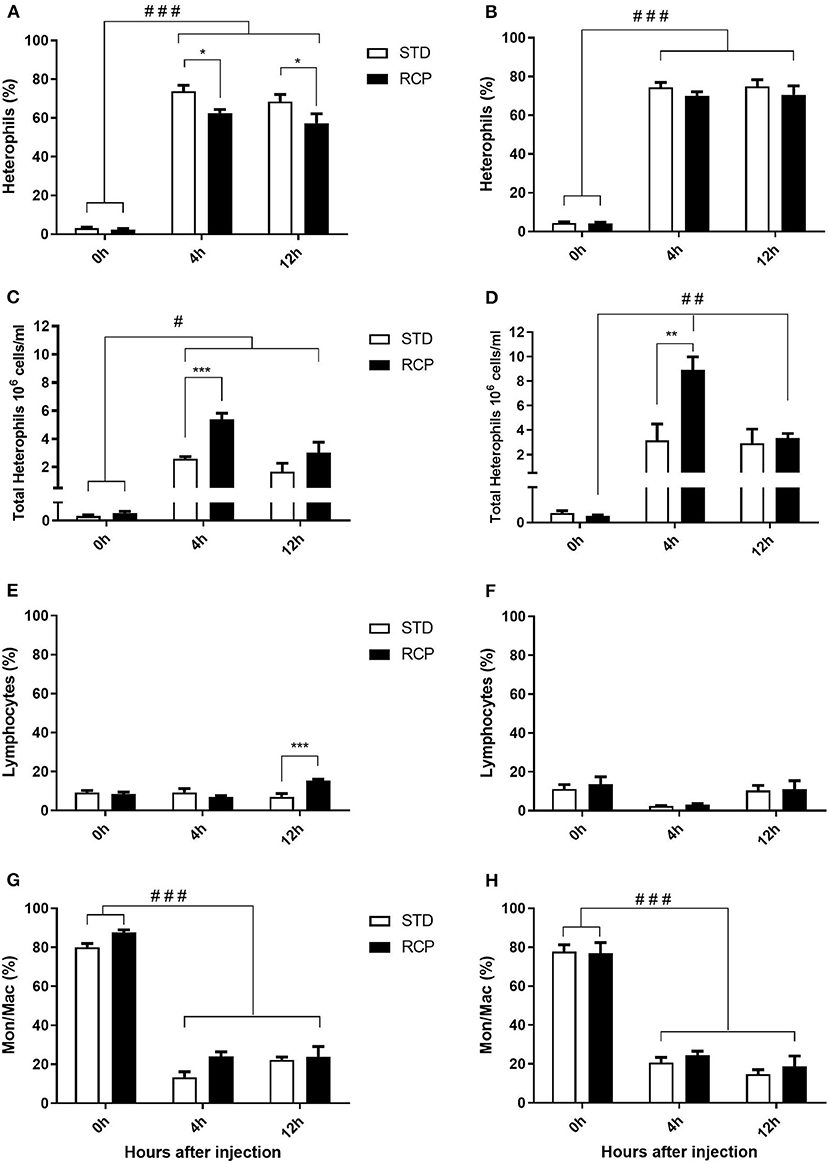
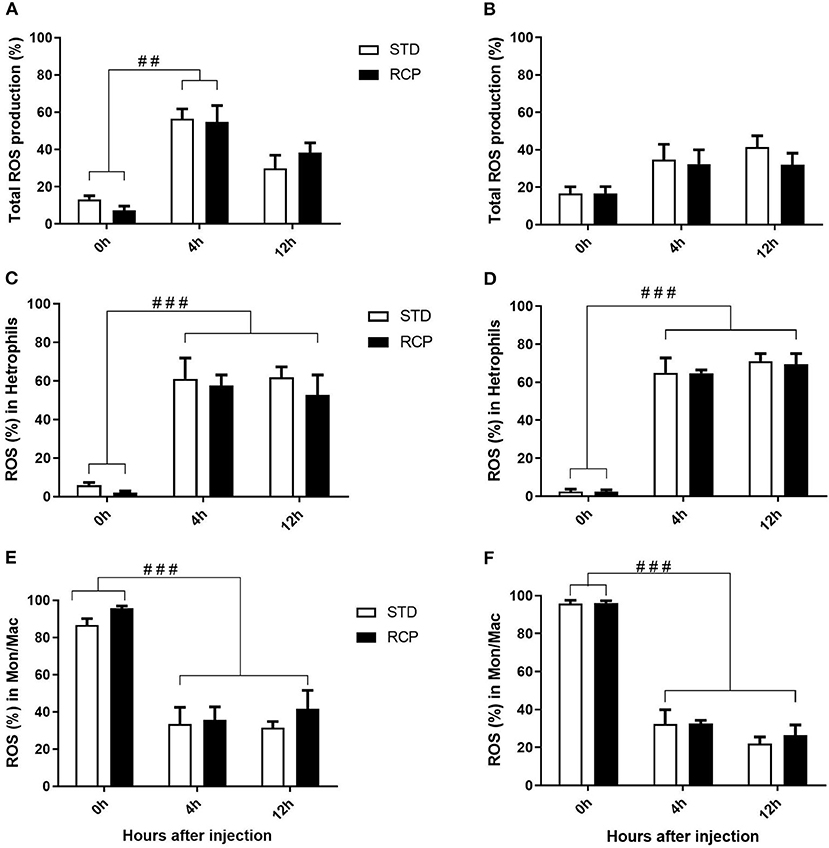
El desafío inmunitario in vivo con zymosan como imitador fúngico dio lugar a un aumento significativo en el reclutamiento de leucocitos a la cavidad abdominal entre las aves suplementadas con RCP en la fase inicial (P < 0,01 en comparación con el tratamiento con ETS; Figuras 2A, B). Además, se observó una interacción significativa entre el tiempo y el tratamiento (P < 0,05), evidente tanto en el día 14 como en el día 35 (Figuras 2A-D). Hemos demostrado previamente que los heterófilos son el leucocito dominante reclutado en la cavidad abdominal después de la exposición a zymosan en pollos (18). Estas células cumplen un papel como las primeras respondedoras contra las infecciones bacterianas, fúngicas y virales (30). Los heterófilos transmigran al sitio de la inflamación en grandes cantidades para neutralizar los patógenos y promover la eliminación de los desechos celulares por fagocitosis (31-34). Como era de esperar, encontramos que la infiltración leucocitaria estuvo dominada por heterófilos (Figuras 3A,B; P < 0,01). La mayor migración de leucocitos en el grupo RCP a d 14 se correlacionó con niveles basales más altos de neutrófilos maduros naïve dentro del grupo de almacenamiento de médula ósea (0 h; Figuras 2C, 6C), así como tasas significativamente más altas de salida de leucocitos (Figura 2C). La mayor migración de leucocitos en el grupo RCP a d 35 se correlacionó con un mayor número basal de leucocitos totales en sangre a d 35 (Figura 2F). El porcentaje de leucocitos productores de ROS no se vio afectado por los tratamientos a las 0, 4 y 12 h después de la provocación con zymosan (Figuras 4A,B).
Producción de ROS en heterófilos y subconjuntos de monocitos/macrófagos
La investigación es limitada con respecto a los efectos del nivel de PC en la producción de ROS en leucocitos de pollo. Un estudio previo no reportó diferencias en la producción de ROS en leucocitos polimorfonucleares en sangre de pollos de engorde de 4 semanas alimentados con 16 o 18% de PB (35) y no hubo diferencias en la producción de NO en macrófagos abdominales en reproductoras de pollos de engorde a la edad de 65 semanas alimentadas con una dieta de 16 o 18% de PB (36). Se puso poco énfasis en esto dado que el dogma de larga data era que los heterófilos aviares dependían principalmente de mecanismos independientes del oxígeno para la actividad antimicrobiana (33, 34). Sin embargo, nuestro grupo demostró recientemente que este no es el caso, sino que proporciona evidencia de una producción significativa de ROS por parte de los heterófilos de pollo utilizando una peritonitis autoresolutiva inducida por la inyección de zymosan (18). De acuerdo con estos resultados, nuestro estudio actual encontró que los heterófilos dominaron el reclutamiento de leucocitos en la cavidad abdominal después de un desafío de zymosan equivalente (Figura 3). Además, los heterófilos fueron los principales contribuyentes a la producción de ROS durante el proceso inflamatorio agudo inducido por la inyección de zymosan (Figura 4). Aunque la producción de ROS se mantuvo constante por célula (Figura 4), el mayor número de heterófilos observados infiltrándose en el sitio de desafío inmunológico de las aves suplementadas con RCP apunta a una mayor capacidad general de producción de ROS por parte de estas células en pollos de engorde.
Médula ósea y poblaciones de leucocitos de sangre periférica
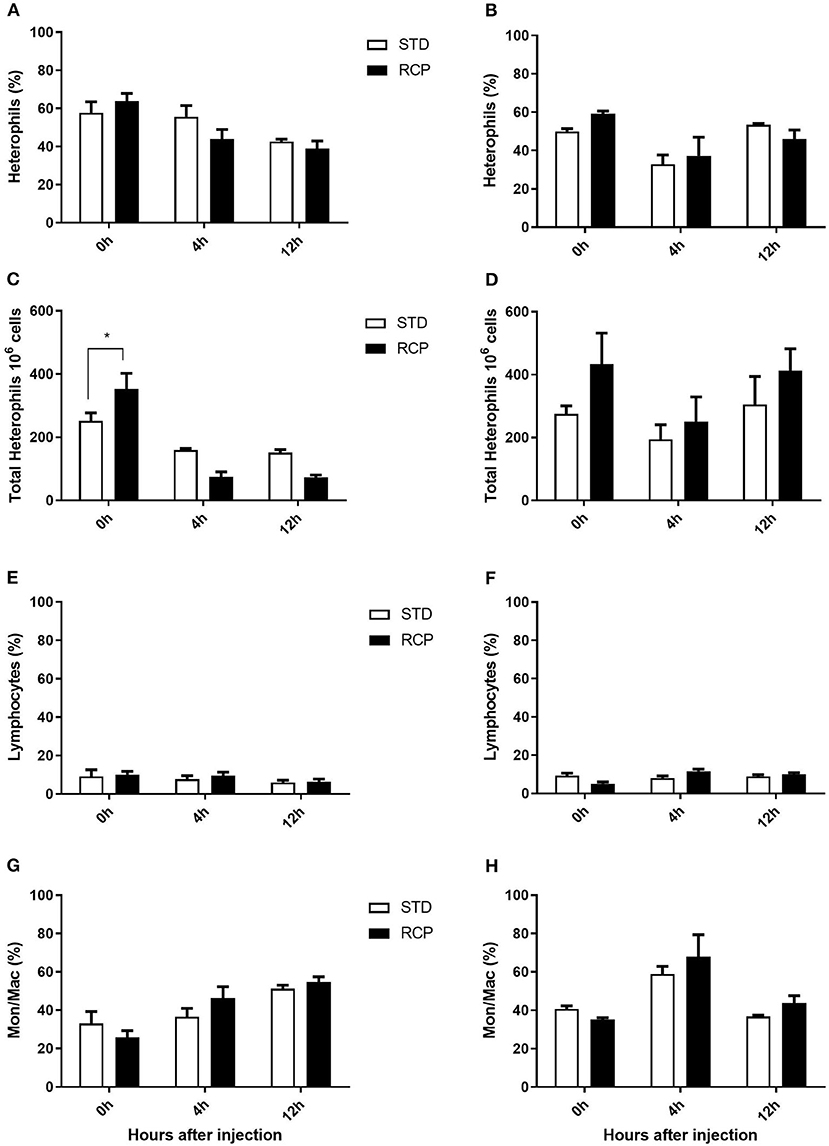
La médula ósea ofrece un compartimento hematopoyético donde las células inmunitarias se desarrollan inicialmente a partir de progenitores tempranos y se mantienen las reservas de leucocitos maduros vírgenes, antes de su liberación a la circulación sanguínea (5, 6, 18). Por lo tanto, estos representan sitios de órganos relevantes para evaluar la distribución y migración de glóbulos blancos ante un desafío inmunológico. El impacto de una dieta reducida en proteína bruta en la composición de leucocitos de sangre periférica y médula ósea después de un desafío con zymosan in vivo se muestra en las Figuras 5 y 6, respectivamente. Los pollos alimentados con dietas de RCP y ETS durante 14 días mostraron números basales equivalentes de leucocitos (es decir, pre-desafío inmunológico) en la cavidad abdominal, la médula ósea y la sangre periférica (Figuras 2A, C, E). Los números basales se mantuvieron constantes en la cavidad abdominal y la médula ósea después de 35 días de crecimiento (Figuras 2B, D). Sin embargo, se identificaron números basales significativamente más altos de leucocitos en la sangre en el día 35 (P < 0,01; Figura 2F). De acuerdo con estos resultados, el examen de mayor resolución de distintos subconjuntos de heterófilos, linfocitos y monocitos/macrófagos mostró un número significativamente mayor de heterófilos en la sangre después de 35 días de crecimiento (Figura 5). Tras la provocación con timosano, el grupo de PCR mostró un menor número total de leucocitos, pero ningún cambio en la proporción de subgrupos individuales en la médula ósea a los 14 días (Figuras 2C, 6 respectivamente) después de 4 h de provocación intraabdominal (P < 0,01). Esto probablemente se asoció con una mayor salida de leucocitos desde este compartimento hematopoyético en ruta hacia el sitio de desafío abdominal (Figura 2). Sin embargo, este cambio significativo en el leucocitos totales de la médula ósea no se observó en el día 35 (Figura 2D). Curiosamente, detectamos una mayor capacidad de reclutamiento total de leucocitos en la cavidad abdominal a d 35 en comparación con d 14 (Figuras 2B vs. 2A), lo que se correlacionó aún más con niveles basales más altos de leucocitos en la sangre durante los mismos períodos de tiempo (Figuras 2F vs. 2E). Una posibilidad es que la sangre pueda desempeñar un papel más importante en el mantenimiento de los leucocitos que pueden ser reclutados a un sitio de infección en etapas posteriores del desarrollo de las aves. Por lo tanto, una mayor disponibilidad de leucocitos en sangre y la posterior infiltración en el sitio de desafío pueden reflejar un potencial para una mayor protección entre las aves RCP.
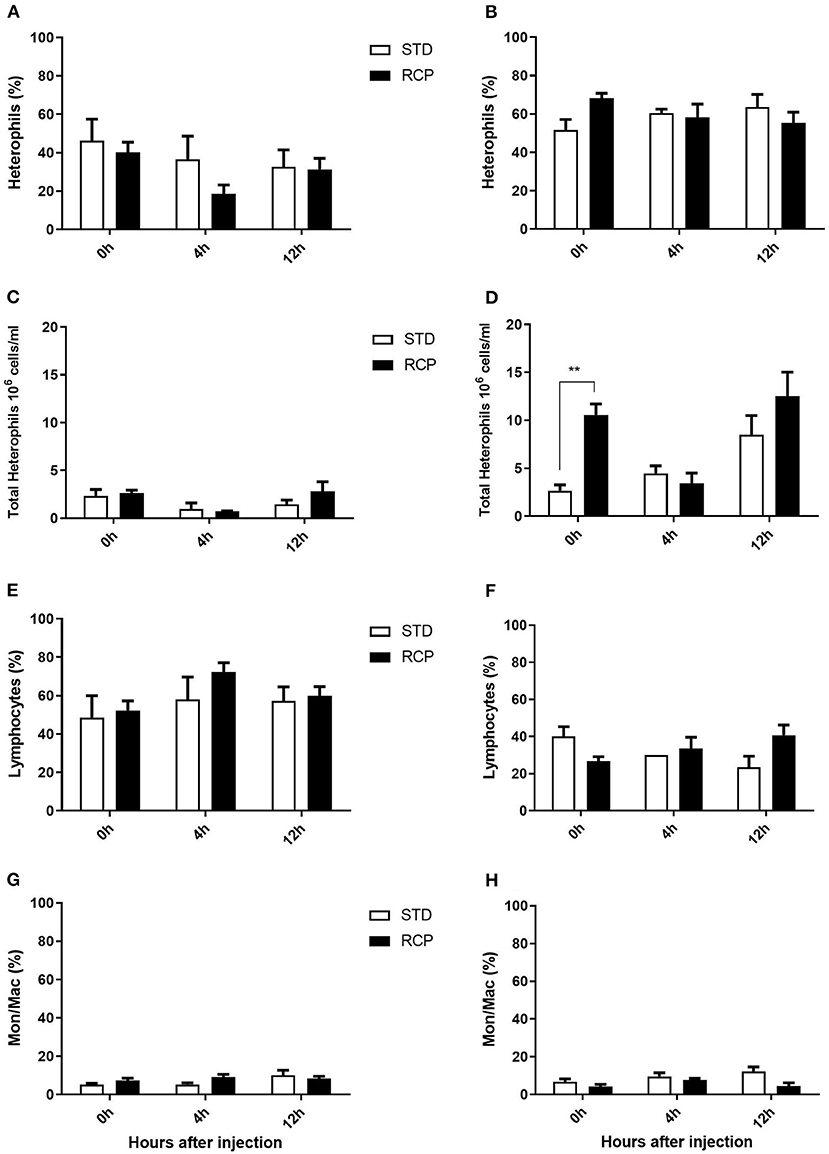
I
Las influencias hormonales indirectas de las proteínas dietéticas pueden ser un factor que contribuye al aumento de los niveles de leucocitos en el grupo RCP. Se ha demostrado que los cambios en el sistema endocrino y las actividades metabólicas, que están directamente influenciadas por la dieta, afectan el estado inmunológico (37). Muchos componentes del sistema endocrino, como la hormona del crecimiento, las hormonas tiroideas y los niveles de corticosterona, pueden regularse bajo condiciones de restricción proteica o energética (19, 37, 38). Además, se ha demostrado que la liberación de heterófilos de la médula ósea a la sangre circulatoria aumenta bajo la estimulación de corticosterona (39). Por lo tanto, es razonable sugerir que el aumento de leucocitos y heterófilos en la médula ósea, la sangre y, posteriormente, una mayor infiltración de leucocitos en la cavidad abdominal pueden ser las consecuencias de cambios en el sistema endocrino en respuesta al nivel más bajo de proteína bruta, aunque se necesitan estudios futuros para aclarar la relación entre el nivel de PC y el nivel de PC. sistema endocrino y modulación inmunológica.
Expresión génica de citocinas proinflamatorias
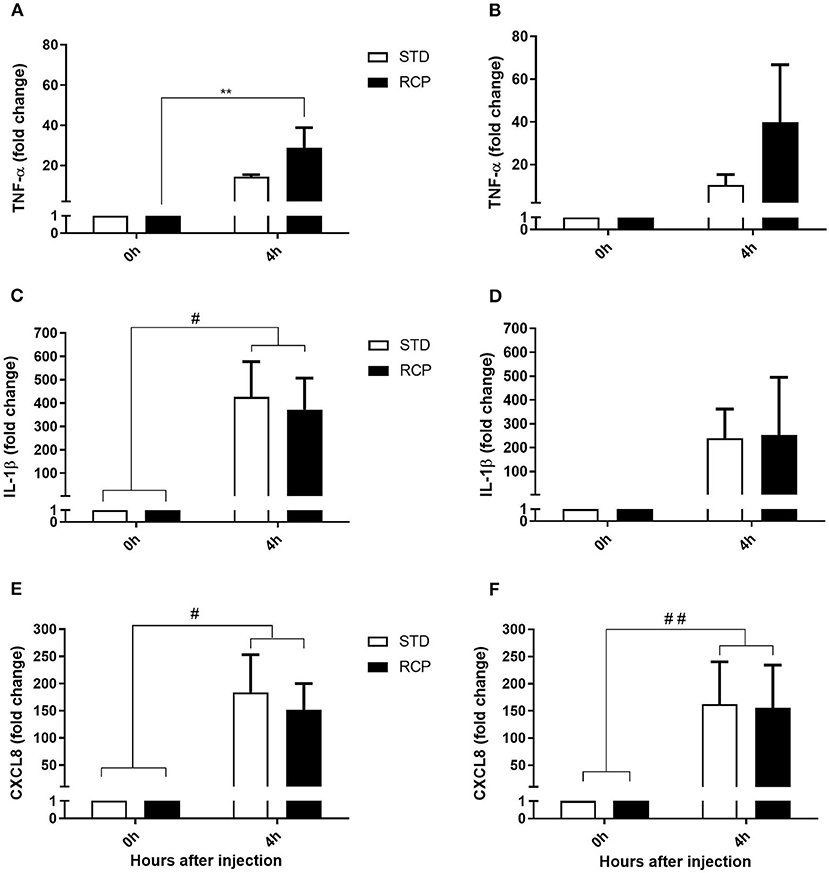
La fase temprana de la inflamación aguda se manifiesta por una funcionalidad multifacética que implica la liberación de mediadores lipídicos como la prostaglandina E2, el leucotrieno C4, el tromboxano A2, quimiocinas como CXCL8 y citocinas proinflamatorias como TNF-α, IL-1 β, IL-6, IL-17, que en última instancia contribuyen al reclutamiento de leucocitos al sitio de la inflamación (17, 18). TNF-α e IL-1β representan activadores tempranos de citocinas de la inflamación aguda, mientras que CXCL8 se encuentra entre las primeras quimiocinas que contribuyen al reclutamiento de leucocitos. Por lo tanto, estos tres genes se utilizaron como marcadores moleculares de la respuesta inflamatoria aguda del pollo. No se detectaron diferencias en la expresión basal de estos tres genes (Figura 7) entre el grupo STD y RCP (P > 0,05). Por el contrario, se detectaron niveles más altos de expresión de TNF-α (Figura 7A) en el grupo RCP después de la provocación intraabdominal con zymosan en el día 14 (P < 0,01). Los niveles aumentados de expresión de TNF-α son consistentes con los niveles más altos de infiltración de leucocitos descritos anteriormente en el grupo RCP después de la provocación intraabdominal.
Conclusiones
En resumen, las reducciones moderadas de la PC dietética en la fase de inicio de los pollos de engorde no afectaron significativamente el rendimiento del crecimiento ni el peso relativo de los órganos en la fase de inicio o más allá hasta los 35 días de edad. Esto era de esperar, ya que las dietas se formularon para cumplir con todos los requisitos estándar de aminoácidos esenciales digeribles ileales en proporciones óptimas. Sin embargo, en comparación con el tratamiento de las ETS, la RCP en la fase inicial aumentó significativamente el reclutamiento de leucocitos a la cavidad abdominal en respuesta a un desafío inmunitario in vivo que utilizaba zymosan como imitador de hongos. Esto se observó durante la fase inicial hasta el día 14 de edad, después de lo cual todas las aves fueron alimentadas con una dieta estándar común de productores de proteína cruda. Además, los niveles totales de leucocitos en la sangre a los 35 días en los tratamientos no desafiados fueron más altos en los grupos de RCP en comparación con los grupos de ETS. Estos cambios que persisten en el período de crecimiento pueden indicar cambios epigenéticos que afectan el desarrollo inmunológico en respuesta a los cambios en la nutrición que ocurren directamente después de la eclosión durante la fase de inicio del desarrollo. Queda por investigar si los cambios observados en el desarrollo inmunitario son una consecuencia directa de los niveles de proteínas de la dieta, o reflejan diferencias en la disponibilidad de aminoácidos libres frente a aminoácidos unidos a proteínas entre las dietas, y/o se deben a las diferencias en las proporciones de almidón:lípidos de las dietas como resultado de los cambios en la inclusión de ingredientes crudos para reducir la PC. Si bien no se pudo observar ninguna diferencia en los mediadores inflamatorios o en la producción de ROS, se plantea la hipótesis de que los puntos de tiempo para observar estos cambios potencialmente ocurrieron antes de los primeros muestreos. La investigación futura podría incluir puntos de tiempo más tempranos, así como marcadores inflamatorios adicionales para comprender mejor los impulsores y los cambios posteriores en la actividad inmunitaria, en respuesta a los niveles de PC en la dieta en la fase inicial. La industria también se beneficiaría de más investigación, para investigar cómo los cambios inmunológicos que ocurren a partir de RCP en la fase inicial pueden beneficiar las estrategias de vacunación, la resistencia a enfermedades a diferentes tipos de patógenos relevantes en el campo o la recuperación del rendimiento del crecimiento después del desafío. En conclusión, este estudio proporciona nuevas y emocionantes pruebas que respaldan los beneficios potenciales de las dietas RCP en fase inicial para el desarrollo y la respuesta del sistema inmunitario en pollos de engorde.
Declaración de disponibilidad de datos
Los conjuntos de datos generados para este estudio están disponibles previa solicitud al autor de correspondencia.
Declaración de Ética
Todos los animales fueron mantenidos de acuerdo con las pautas especificadas por el Consejo Canadiense de Cuidado Animal, y los protocolos fueron aprobados por el Comité de Cuidado y Uso de Animales de la Universidad de Alberta.
Contribuciones del autor
MK, JW, RW, VN y DB concibieron y diseñaron el estudio. MK y WH realizaron los experimentos. MK analizó e interpretó los datos, preparó las figuras y redactó el manuscrito. Todos los autores participaron en la revisión del manuscrito y aprobaron la versión final del mismo. Todos los autores contribuyeron al artículo y aprobaron la versión presentada.
Conflicto de intereses
RW y VN fueron empleados de Evonik Nutrition & Care GmbH.
Este trabajo contó con el apoyo de la beca Evonik Nutrition & Care GmbH RES0037933. El financiador contribuyó al diseño del estudio y a la revisión de manuscritos. El financiador no participó en la recopilación, análisis o interpretación de datos.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Abreviaturas
AMEn, Energía metabolizable aparente corregida por nitrógeno; CP: Proteína bruta; FCR: Índice de conversión alimenticia; Mon/Mac, Monocitos/macrófagos; RCP: Dieta reducida en proteína bruta; ROS: Especies reactivas de oxígeno; SID: digestible ileal estándar; ETS, dieta estándar.
Referencias
1. Erf GF, Bottje WG, Bersi TK. CD4, CD8 and TCR defined T-cell subsets in thymus and spleen of 2-and 7-week old commercial broiler chickens. Vet Immunol Immunop. (1998) 62:339–48. doi: 10.1016/S0165-2427(97)00070-6
2. Khajavi M, Rahimi SH, Hassan ZM, Kamali MA, Mousavi T. Effect of feed restriction early in life on humoral and cellular immunity of two commercial broiler strains under heat stress conditions. Bri Poult Sci. (2003) 44:490–7. doi: 10.1080/000071660310001598328
3. Cheema MA, Qureshi MA, Havenstein GB. A comparison of the immune profile of commercial broiler strains when raised on marginal and high protein diets. Int J Poult Sci. (2003) 2:300–12. doi: 10.3923/ijps.2003.300.312
4. Qureshi MA, Marsh JA, Dietert RR, Sung YJ, Nicolas-Bolnet C, Petitte JN. Perfiles de las funciones efectoras de los macrófagos de pollo. Poult Sci. (1994) 73:1027–34. doi: 10.3382/ps.0731027
Resumen de PubMed | Texto completo de CrossRef | Google Académico
5. Erf GF. Función y desarrollo del sistema inmunológico en pollos de engorde. Poult Sci. (1997) 76:109–23.
6. Kogut MH, Iqbal M, He H, Philbin V, Kaiser P, Smith A. Expresión y función de los receptores tipo Toll en heterófilos de pollo. Dev Comp Immunol. (2005) 29:791–807. DOI: 10.1016/j.dci.2005.02.002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
7. O’Reilly EL, Bailey RA, Eckersall PD. Un estudio comparativo de las concentraciones de proteínas en fase aguda en líneas de cría de pollos de engorde históricas y modernas. Poult Sci. (2018) 97:3847–53. doi: 10.3382/ps/pey272
8. Chuammitri P, Redmond SB, Kimura K, Andreasen CB, Lamont SJ, Palić D. Las respuestas funcionales de los heterófilos a los inmunomoduladores dietéticos varían en líneas de pollos genéticamente distintas. Veterinario Immunol Immunopathol. (2011) 142:219–27. doi: 10.1016/j.vetimm.2011.05.019
9. Redmond SB, Chuammitri P, Andreasen CB, Palić D, Lamont SJ. Chicken heterophils from commercially selected and non-selected genetic lines express cytokines differently after in vitro exposure to Salmonella enteritidis. Vet Immunol Immunopathol. (2009) 132:129–34. doi: 10.1016/j.vetimm.2009.05.010
10. Sigolo S, Zohrabi Z, Gallo A, Seidavi A, Prandini A. Effect of a low crude protein diet supplemented with different levels of threonine on growth performance, carcass traits, blood parameters, and immune responses of growing broilers. Poult Sci. (2017) 96:2751–60. doi: 10.3382/ps/pex086
11. Hilliar M, Hargreave G, Girish CK, Barekatain R, Wu SB, Swick RA. Using crystalline amino acids to supplement broiler chicken requirements in reduced protein diets. Poult Sci. (2020) 99:1551–63. doi: 10.1016/j.psj.2019.12.005
12. Alleman F, Michel J, Chagneau AM, Leclercq B. The effects of dietary protein independent of essential amino acids on growth and body composition in genetically lean and fat chickens. Br Poult Sci. (2000) 41:214–8. doi: 10.1080/713654902
13. Hofmann P, Siegert W, Kenéz Á, Naranjo VD, Rodehutscord M. La proteína cruda muy baja y las concentraciones variables de glicina en la dieta afectan el rendimiento del crecimiento, las características de la excreción de nitrógeno y el metaboloma sanguíneo de los pollos de engorde. J Nutr. (2019) 149:1122–32. doi: 10.1093/jn/nxz022
Resumen de PubMed | Texto completo de CrossRef | Google Académico
14. Chrystal PV, Moss AF, Khoddami A, Naranjo VD, Selle PH, Liu SY. Impactos de las dietas reducidas en proteína bruta en parámetros clave en pollos de engorde machos ofrecidos por dietas a base de maíz. Poult Sci. (2020) 99:505–16. doi: 10.3382/ps/pez573
Resumen de PubMed | Texto completo de CrossRef | Google Académico
15. Klasing KC. Nutrición y sistema inmunológico. Bri Poult Sci. (2007) 48:525–37. doi: 10.1080/00071660701671336
Resumen de PubMed | Texto completo de CrossRef | Google Académico
16. Doherty NS, Poubelle P, Borgeat P, Beaver TH, Westrich GL, Schrader NL. La inyección intraabdominal de zymosan en ratones induce dolor, inflamación y la síntesis de peptidoleukotrienos y prostaglandina E2. Prostaglandinas. (1985) 30:769–89. doi: 10.1016/0090-6980(85)90006-1
Resumen de PubMed | Texto completo de CrossRef | Google Académico
17. Havixbeck JJ, Rieger AM, Wong ME, Hodgkinson JW, Barreda DR. Contribuciones de los neutrófilos a la inducción y regulación de la respuesta inflamatoria aguda en peces teleósteos. J Leucocitos Biol. (2016) 99:241–52. doi: 10.1189/jlb.3HI0215-064R
Resumen de PubMed | Texto completo de CrossRef | Google Académico
18. Más Bayona JA, Karuppannan AK, Barreda DR. Contribución de los leucocitos a la inducción y resolución de la respuesta inflamatoria aguda en pollos. Dev Comp Immunol. (2017) 74:167–77. doi: 10.1016/j.dci.2017.04.018
Resumen de PubMed | Texto completo de CrossRef | Google Académico
19. Kamely M, Karimi Torshizi MA, Rahimi S. Incidencia del síndrome de ascitis y respuesta hematológica relacionada en pollos de engorde con restricción de alimento a corto plazo criados a baja temperatura ambiente. Poult Sci. (2015) 94:2247–56. doi: 10.3382/ps/pev197
20. Kamely M, Karimi Torshizi MA, Wideman RF, West J. Regulación positiva de SERT y ADORA1 en pollos de engorde con insuficiencia ventricular derecha aguda. Res Vet Sci. (2017) 95:2673–78.
21. Han Y, Suzuki H, Parsons CM, Baker DH. Fortificación de aminoácidos de una dieta baja en proteínas de harina de maíz y soja para maximizar la ganancia de peso y la eficiencia alimenticia del pollito. Poult Sci. (1992) 71:1168–78. doi: 10.3382/ps.0711168
Resumen de PubMed | Texto completo de CrossRef | Google Académico
22. Ospina-Rojas IC, Murakami AE, Eyng C, Nunes RV, Duarte CRA, Vargas MD. Suplementación con aminoácidos disponible comercialmente en dietas bajas en proteínas para pollos de engorde con diferentes proporciones de glicina digerible + serina: lisina. Poult Sci. (2012) 91:3148–3155. doi: 10.3382/ps.2012-02470
23. Aletor VA, Hamid II, Nieß E, Pfeffer E. Dietas suplementadas con aminoácidos bajos en proteínas en pollos de engorde: efectos sobre el rendimiento, las características de la canal, la composición de todo el cuerpo y la eficiencia de la utilización de nutrientes. J Sci Alimentos Agric. (2000) 80:547–54. doi: 10.1002/(SICI)1097-0010(200004)80:5<547::AID-JSFA531>3.0.CO; 2-C
24. Lemme A, Hiller P, Klahsen M, Taube V, Stegemann J, Simon I. Reduction of dietary protein in broiler diets not only reduces n-emissions but is also accompanied by several further benefits. J Appl Poult Res. (2019) 28:867–80. doi: 10.3382/japr/pfz045
25. McGill E, Kamyab A, Firman JD. Dietas bajas en proteína cruda de maíz y harina de soja con suplementación de aminoácidos para pollos de engorde en el período inicial. 1. Efectos de la alimentación con un 15% de proteína bruta. Int J Poult Sci. (2012) 11:161–5. doi: 10.3923/ijps.2012.161.165
26. Cooper MD, Raymond DA, Peterson RD, Sur de MA, Buena RA. Las funciones del sistema timo y el sistema bursa en el pollo. J Exp Med. (1966) 123:75–102. doi: 10.1084/jem.123.1.75
Resumen de PubMed | Texto completo de CrossRef | Google Académico
27. Bunchasak C, Poosuwan K, Nukraew R, Markvichitr K, Choothesa A. Efecto de la proteína dietética en la producción de huevos y las respuestas inmunitarias de las gallinas ponedoras durante el período de máxima producción. Int J Poult Sci. (2005) 4:701–8. doi: 10.3923/ijps.2005.701.708
28. Kidd MT, Gerard PD, Heger J, Kerr BJ, Rowe D, Sistani, et al. Respuestas de treonina y proteína bruta en pollos de engorde. Anim Feed Sci Tech. (2001) 94:57–64. doi: 10.1016/S0377-8401(01)00301-7
29. Rao SR., Praharaj NK, Reddy MR, Sridevi B. Competencia inmunológica, resistencia a Escherichia coli y crecimiento en pollitos padres de pollos de engorde machos alimentados con diferentes niveles de proteína bruta. Vet Res Commun. (1999) 23:323–36.
30. Rankin SM. La médula ósea: un sitio de aclaramiento de neutrófilos. J Hervor de leucocitos. (2010) 88:241–51. doi: 10.1189/jlb.0210112
Resumen de PubMed | Texto completo de CrossRef | Google Académico
31. Freire MO, Van Dyke TE. Resolución natural de la inflamación. Periodoncia 2000. (2013) 63:149–64. doi: 10.1111/prd.12034
Resumen de PubMed | Texto completo de CrossRef | Google Académico
32. Genovese KJ, He H, Swaggerty CL, Kogut MH. El heterófilo aviar. Dev Comp Immunol. (2013) 41:334–40. doi: 10.1016/j.dci.2013.03.021
Resumen de PubMed | Texto completo de CrossRef | Google Académico
33. Harmon BG. Heterófilos aviares en la inflamación y la resistencia a enfermedades. Ciencia avícola. (1998) 77:972–7. doi: 10.1093/PS/77.7.972
Resumen de PubMed | Texto completo de CrossRef | Google Académico
34. Mitchell EB, Johns J. Hematología aviar y trastornos relacionados. Veterinario Clin Exot Anim. (2008) 11:501–22. doi: 10.1016/j.cvex.2008.03.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
35. Papp Z, Dahiya JP, Warren T, Widyaratne G, Drew MD, Smits JE. Respuesta de quimioluminiscencia de sangre total en pollos de engorde en diferentes dietas experimentales y desafiados con clostridium perfringens. Bri Poult Sci. (2009) 50:57–65. DOI: 10.1080/00071660802613278
Resumen de PubMed | Texto completo de CrossRef | Google Académico
36. Khan RU, Rahman ZU, Javed I, Muhammad F. Efecto de las vitaminas, el nivel de proteínas y los probióticos en la respuesta inmunitaria de los reproductores de pollos de engorde machos en muda. J Anim Physiol Anim Nutr. (2014) 98:620–7. doi: 10.1111/jpn.12087
Resumen de PubMed | Texto completo de CrossRef | Google Académico
37. Malheiros RD, Moraes VM, Collin A, Janssens GP, Decuypere E, Buyse J. Macronutrientes dietéticos, funcionamiento endocrino y metabolismo intermediario en pollos de engorde: sustituciones por pares entre proteínas, grasas y carbohidratos. Nutr Res. (2003) 23:567–78. doi: 10.1016/S0271-5317(03)00022-8
38. Rajman M, Juráni M, Lamošová D, Máčajová M, Sedlačková M, Koštál L, et al. The effects of feed restriction on plasma biochemistry in growing meat type chickens (Gallus gallus). Comp Biochem Physiol A Mol Integr Physiol. (2006) 145:363–71. doi: 10.1016/j.cbpa.2006.07.004
Keywords: broiler, cytokines, dietary crude protein, inflammation, leukocyte, innate immunity
Citation: Kamely M, He W, Wakaruk J, Whelan R, Naranjo V and Barreda DR (2020) Impact of Reduced Dietary Crude Protein in the Starter Phase on Immune Development and Response of Broilers Throughout the Growth Period. Front. Vet. Sci. 7:436. doi: 10.3389/fvets.2020.00436
Received: 03 March 2020; Accepted: 16 June 2020;
Published: 07 August 2020.
Edited by:
Fulvia Bovera, University of Naples Federico II, Italy
Reviewed by:
Samuel J. Rochell, University of Arkansas, United States
Alireza Seidavi, Islamic Azad University, Iran
Derechos de autor © 2020 Kamely, He, Wakaruk, Whelan, Naranjo y Barreda. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Atribución (CC BY).
*Correspondencia: Daniel R. Barreda, d.barreda@ualberta.ca
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía