
Influencia de los ácidos grasos de cadena media en microbios seleccionados y en la fermentación ruminal

Influencia de los ácidos grasos de cadena media en microbios seleccionados y en la fermentación ruminal in vitro de ensilado de maíz expuesto al aire
 Jaime Salinas-Chavira1
Jaime Salinas-Chavira1 Claudio Arzola-Álvarez2
Claudio Arzola-Álvarez2 Michael E. Hume3
Michael E. Hume3 Mozart Fonseca4Óscar Ruiz-Barrera2Yamicela Castillo-Castillo2
Mozart Fonseca4Óscar Ruiz-Barrera2Yamicela Castillo-Castillo2 Marina Ontiveros-Magadán5Bárbara Jones6 añosTawni L. Crippen3
Marina Ontiveros-Magadán5Bárbara Jones6 añosTawni L. Crippen3 Toni L. Poole3Aracely Zúñiga-Serrano1
Toni L. Poole3Aracely Zúñiga-Serrano1 Robin C. Anderson3*
Robin C. Anderson3*- 1Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Tamaulipas, Ciudad Victoria, Tamaulipas, México
- número arábigoFacultad de Ciencia y Ecología Animal, Universidad Autónoma de Chihuahua, Chihuahua, México
- 3Departamento de Agricultura de los Estados Unidos, Servicio de Investigación Agrícola, Unidad de Investigación sobre la Inocuidad de los Alimentos y los Piensos, College Station, TX, Estados Unidos
- 4Departamento de Agricultura, Nutrición y Ciencias Veterinarias, Universidad de Nevada, Reno, NV, Estados Unidos
- 5Departamento de Ciencias Veterinarias, Universidad Autónoma de Ciudad Juárez, Ciudad Juárez, Chihuahua, México
- 6Departamento de Ciencia Animal y Tecnología Veterinaria, Universidad Estatal de Tarleton, Stephenville, TX, Estados Unidos
Se evaluaron in vitro varios ácidos grasos de cadena media y diferentes formas químicas de estos ácidos como tratamientos de ensilado de maíz expuesto aeróbicamente contra el deterioro y los microbios patógenos y para determinar sus efectos sobre la fermentación ruminal. Los tratamientos fueron control (sin aditivo), miristato (MY), laurato (LA), monolaurina (MLA), laurato de éster metílico (MELA), una mezcla de mono-, di- y triglicéridos de laurato (BLA) y monocaprilato (MC). Después de 24 h de incubación aeróbica (37 °C), el crecimiento de levaduras y mohos no se vio influenciado (P > 0,05) por los tratamientos, mientras que el crecimiento neto de las bacterias lácticas disminuyó, aunque ligeramente, en comparación con el de los controles no tratados (P < 0,01) por todos los tratamientos del ensilado de maíz expuesto al aire. En comparación con los controles, los enterococos de tipo natural disminuyeron (P < 0,01) en MLA, MELA y BLA. Staphylococcus aureus se redujo (P < 0.01) con MLA, MELA, BLA y MC. Los aerobios totales mostraron reducciones (P < 0.01) con MLA, BLA y MC. Los números de Listeria monocytogenes se redujeron (P < 0.01) con MELA. La incubación anaeróbica (24 h; 39°C) de líquido ruminal (10 mL) con 0,2 g de ensilado de maíz expuesto al aire y tratado con AGCM reveló mayores acumulaciones de hidrógeno (P < 0,01) con MLA y MC durante el tratamiento control. El metano disminuyó (P < 0,01) únicamente por MLA. Hubo un aumento (P < 0,01) de acetato con MELA y MC; de propionato con MELA o por BLA; y de butirato con MLA, MELA, BLA y MC. AGV totales, hexosa fermentada y amoníaco se incrementaron (P < 0,01) con MELA, BLA y MC. La relación acetato:propionato se incrementó (P < 0,01) con MC. Los resultados mostraron que el tratamiento del ensilado de maíz expuesto al aire con AGCM esterificado no tuvo ningún efecto sobre las levaduras y los mohos, pero evitó la propagación o redujo las poblaciones de algunas bacterias no deseadas y potencialmente deseables. Se observó una modesta reducción de metano durante la incubación in vitro de suspensiones de fluido ruminal con ensilado tratado con MLA, y las acumulaciones de amoníaco aumentaron en el ensilado esterificado tratado con MCFA. Se observaron pocos o ningún otro efecto perjudicial sobre las características beneficiosas de la fermentación ruminal.
1 Introducción
La apertura del sello anaeróbico de los ensilajes durante la alimentación inicia cambios aeróbicos irreversibles que alteran la calidad nutricional y microbiológica del alimento, y este proceso continúa en el comedero. Las levaduras y los mohos son las principales fuentes de microorganismos de deterioro aeróbico, y algunas fuentes bacterianas actúan como contribuyentes secundarios (1). La seguridad microbiana del ensilado podría verse comprometida durante la fase aeróbica, con el crecimiento de bacterias patógenas como Staphylococcus aureus, Listeria monocytogenes, Enterobacteriaceae (Escherichia coli y Salmonella spp.) así como Clostridium botulinum (2, 3). Además, algunos mohos pueden producir micotoxinas con el potencial de perturbar la producción y la salud de los animales (1).
Los ácidos grasos de cadena media (AGCM) octanoicos (C8, caprilatos), decanoicos (C10, capratos) y dodecanoicos (C12, lauratos), y sus monoglicéridos, tienen la capacidad de reducir el número de bacterias Gram-positivas (4, 5). Además, el caprilato inhibió el crecimiento de bacterias enteropatógenas (6-10). Durante la preparación del ensilado, los AGCM también han mostrado influencias en la fermentación. McDonald y Henderson (11) observaron una reducción de la fermentación con ácido hexanoico (C6, caproatos), mientras que Abel et al. (12) encontraron una reducción del contenido de lactato en el ensilado de hierba tratado con caprilato. Además, se ha reducido la proliferación de levaduras y mohos en ensilajes tratados con ácido propiónico (C3, propionato) (13).
Los AGCM han reducido consistentemente la producción de metano durante la fermentación ruminal (14-16). Sin embargo, cuando se han añadido AGCM en el momento del ensilado, no se ha reducido la producción de metano tras la ingestión y posterior fermentación en el rumen (12). Existe información limitada sobre los efectos del AGCM cuando se aplica en ensilajes en el momento de la apertura como un posible tratamiento para inhibir o retrasar el deterioro del ensilado o promover el control de bacterias patógenas durante la fase aeróbica. Además, hay informes limitados sobre los efectos residuales de los AGCM aplicados en la apertura del ensilado sobre las características de fermentación ruminal. Ontiveros-Magadan et al. (17), con ensilaje expuesto al aire tratado con laurato (LA) o monolaurina (MLA), observaron una reducción del número de bacterias Gram-positivas, levaduras y mohos, aunque el número de bacterias lácticas (BAL) se redujo después de unas horas de incubación; Además, la producción de metano también se redujo durante las incubaciones ruminales in vitro. En un estudio similar, Arzola-Álvarez et al. (18) utilizaron AGCM (C6; C8; C10; una mezcla C8:10; y una mezcla C6:12) y encontraron reducciones de enterococos y BAL, aunque Staphylococcus aureus y Listeria monocytogenes no se vieron afectados por el tratamiento, y la levadura y el moho se redujeron únicamente con la mezcla C8:10. Esos autores informaron pequeños cambios en las características de la fermentación ruminal.
Como se ha señalado, la información sobre el tratamiento del ensilado de maíz expuesto al aire con AGCM es limitada e inconsistente en lo que respecta a los cambios microbianos durante la fase aeróbica y a los posibles efectos residuales sobre las características de la fermentación ruminal. Además, hasta donde sabemos, no hay información sobre la aplicación en el ensilado de laurato de éster metílico, mezclas de mono-, di- y triglicéridos de laurato o monocaprilato. Teniendo en cuenta la importancia de mantener la calidad nutricional y reducir la proliferación de microorganismos no deseados durante la exposición aeróbica del ensilado, el objetivo del presente experimento fue probar nuestra hipótesis de que el tratamiento del ensilado de maíz expuesto aeróbicamente con MCFA y ésteres de MCFA evitará la propagación de microbios patógenos y de deterioro seleccionados sin tener efectos adversos sobre la fermentación ruminal cuando se incuba in vitro bajo condiciones de simulación de hábitat ruminal.
2 Materiales y métodos
2.1 Fuentes de microbios
La cepa Scott A de Listeria monocytogenes (serotipo 4b) fue proporcionada por el Dr. J. F. Frank, de la Universidad de Georgia, Athens, GA, EE.UU. El difunto Dr. Max Paape proporcionó una cepa de Staphylococcus aureus resistente a múltiples fármacos y resistente a la meticilina (MDR) ATCC 49521, que demostró anteriormente ser resistente a la eritromicina, linezolid, penicilina, quinupristina/dalfopristina y vancomicina (19). Se utilizaron cultivos madre cultivados durante la noche a 37 °C en caldo de soja tríptico aeróbico (Becton Dickinson and Company, Sparks, MD, EE. UU.) como inóculos para las incubaciones con ensilado expuesto al aire. El ensilado de maíz (38 ± 5.3% de materia seca y pH 4.73) se recolectó en el Centro Regional de Productos Lácteos del Suroeste de la Universidad Estatal de Tarleton (Stephenville, TX, EE. UU.) y se almacenó a temperatura ambiente hasta su uso ~ 24 h después.
El líquido ruminal utilizado para las pruebas in vitro de los posibles efectos de arrastre de MCFA en el ensilado tratado se recolectó la mañana del experimento de una vaca Jersey canulada que pastaba en pastizales de Bermuda con acceso ad libitum al heno de alfalfa en el Centro de Investigación Agrícola de las Llanuras del Sur del USDA, College Station, TX. Todos los procedimientos para el cuidado y uso de esta vaca fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales del Centro de Investigación Agrícola de las Llanuras del Sur.
2.2 Ensayos con poblaciones mixtas de bacterias del ensilaje y del rumen
Porciones de cuatro gramos de ensilado expuesto al aire se colocaron en juegos separados de triplicado (n = 3/tratamiento) tubos centrífugos cónicos de 50 mL que habían sido cargados previamente sin o con 30 mg del MCFA de prueba. Para este estudio, los AGCM utilizados fueron miristato (MY; ácido tetradecanoico), laurato (LA; ácido dodecanoico), monolaurina (MLA; monolaurato de glicerol), laurato de éster metílico (MELA), una mezcla de mono-, di- y triglicéridos de laurato (BLA) o monocaprilato (MC; monocaprilato de glicerol). Todos los AGCM utilizados en este estudio fueron suministrados por Berg + Schmidt GmbH & Co. KG (Hamburgo, Alemania). Se añadió agua destilada estéril (10 mL) a cada uno de los tubos preparados anteriormente y cada tubo se inoculó con 10 μL de L. monocytogenes Scott A y S. aureus ATCC 49521. Cada inóculo se cultivó aeróbicamente durante 20 h a 37°C en caldo de soja tríptico (Becton Dickinson and Company) antes de la inoculación. A continuación, las preparaciones de ensilado expuestas al aire se taparon y se incubaron a 30 °C mientras estaban expuestas al aire. Las muestras líquidas recogidas al inicio y finalización del período de incubación de 24 horas se diluyeron en serie (10 veces) en tampón de fosfato de sodio 0,1 M (pH 6,5) y luego se colocaron en un medio selectivo y diferencial solidificado para el recuento de células viables de poblaciones microbianas seleccionadas. Las cepas de S. aureus y L. monocytogenes inoculadas experimentalmente se enumeraron respectivamente utilizando sal de manitol BBL™ (Becton Dickinson and Company) y agar selectivo de Listeria suplementado con suplemento selectivo de Listeria oxoide™ (Oxford LTD, Basingstoke, Hampshire, Inglaterra). Las bacterias lácticas (BAL) y los enterococos de tipo salvaje se enumeraron utilizando agares Rogosa SL y m Enterococcus, respectivamente (cada uno procedente de Becton Dickinson and Company). Los recuentos respectivos de aerobios totales y levaduras y mohos se enumeraron utilizando las placas de recuento aeróbico de Petrifilm de 3M™ y las placas de recuento de levaduras y mohos de Petrifilm™ de 3M™ (3M Petrifilm, St. Paul, MN, EE. UU.).™ Las placas inoculadas y el petrifilm se utilizaron de acuerdo con las instrucciones del fabricante.
Para examinar los posibles efectos de arrastre del AGCM en el ensilado tratado expuesto al aire en la fermentación ruminal, el ensilado de maíz expuesto al aire que había sido tratado individualmente sin o con MY, LA, MLA, MELA, BLA o MC se incubó in vitro en condiciones de hábitat ruminal simulando utilizando líquido ruminal recolectado recién recolectado a las 10:00 de la mañana del estudio de un novillo Jersey canulado ruminalmente que pastaba en pasturas de pasto Bermuda. El líquido ruminal, que contenía poblaciones viables de microbios ruminales, se obtuvo exprimiendo el contenido ruminal recogido a través de la cánula a través de un colador de pintura de nailon (20) en un termo de 250 ml que se tapó inmediatamente después del llenado y se devolvió al laboratorio dentro de los 30 minutos posteriores a la recolección. El fluido ruminal recolectado se distribuyó en volúmenes de 10 mL bajo un flujo continuo de dióxido de carbono en 18 tubos superiores de cultivo de engarce de 150 mm (3 tubos replicados/tratamiento) que habían sido precargados sin o con ensilado de 0,2 gramos expuesto aeróbicamente que habían sido tratados individualmente sin o con MY, LA, MLA, MELA, BLA o MC. A continuación, los tubos se taponaron, sellaron e incubaron durante 24 h a 39 °C. Los fluidos de cultivo y los gases de cabeza recogidos al principio y al final de la incubación se sometieron a cromatografía de gases para la determinación de ácidos grasos volátiles (AGV) (21) y para el contenido de hidrógeno y metano (22). Los volúmenes de gas en el espacio de cabeza al final de la incubación se midieron mediante el desplazamiento del volumen utilizando una jeringa de vidrio. Las concentraciones de amoníaco se midieron colorimétricamente (23). Las estimaciones estequiométricas de las cantidades de hexosa fermentadas se calcularon como (12 acetato + 12 propionato + butirato + valerato) y la eficiencia de fermentación se calculó como (0,62 acetato + 1,09 propionato + 0,78 butirato)/(acetato + propionato + butirato), Chalupa (24). La lejía, el nitroferricianuro, el fenol y el hidróxido de sodio utilizados en el ensayo de amoníaco se obtuvieron de CloroxTM (Oakland, CA, EE. UU.), Fisher (Fair Lawn, NJ, EE. UU.), Sigma-Aldrich (St. Louis, MO, EE. UU.) y Argos Organics (Geel, Bélgica). El fosfato mono y disódico utilizado para preparar el tampón de fosfato 0,1 M se compró a Sigma.
2.3 Análisis estadístico
Los cultivos control y tratados se incubaron por triplicado, y cada réplica sirvió como unidad experimental independiente. Los cambios netos en los recuentos de bacterias viables, expresados como log10 unidades formadoras de colonias (UFC)/g de ensilaje, las acumulaciones netas de productos de fermentación ruminal y las estimaciones estequiométricas de las cantidades de hexosa fermentada se determinaron como la diferencia entre las mediciones realizadas al comienzo y al final de la incubación, y se analizaron los posibles efectos del tratamiento utilizando un análisis de varianza completamente aleatorizado con una comparación múltiple de Dunnett bilateral con los controles (Statistix10 Software analítico, Tallahassee, FL, EE. UU.).
3 Resultados
3.1 Concentraciones microbianas seleccionadas
En la Tabla 1 se muestran los efectos de los diferentes AGCM en el cambio neto, que representa la diferencia entre las concentraciones iniciales y finales, en las concentraciones microbianas durante la incubación. En el caso de S. aureus, la disminución de 1,51 a 1,90 log-fold observada en el ensilado expuesto aeróbicamente tratado con MC, BLA, MLA y MELA difirió (P < 0,05) del aumento de 0,14 log10 UFC/g observado con los controles no tratados. El cambio neto en las concentraciones de S. aureus observado con MY y LA no difirió (P > 0.05) del observado con los controles. De manera similar, las concentraciones aerobias totales mostraron reducciones desde sus niveles iniciales cuando el ensilaje expuesto al aire fue tratado con MLA, BLA o MC, pero la magnitud de la disminución fue mucho más modesta que la observada con S. aureus. Sin embargo, el cambio neto en los aerobios totales fue significativamente diferente con el ensilado tratado con MLA, BLA y MC (P < 0.01) que el cambio neto observado con los controles no tratados (Tabla 1). Sin embargo, las concentraciones de aerobios totales fueron similares (P > 0,05) entre el grupo control y los tratados con LA, MY o MELA.
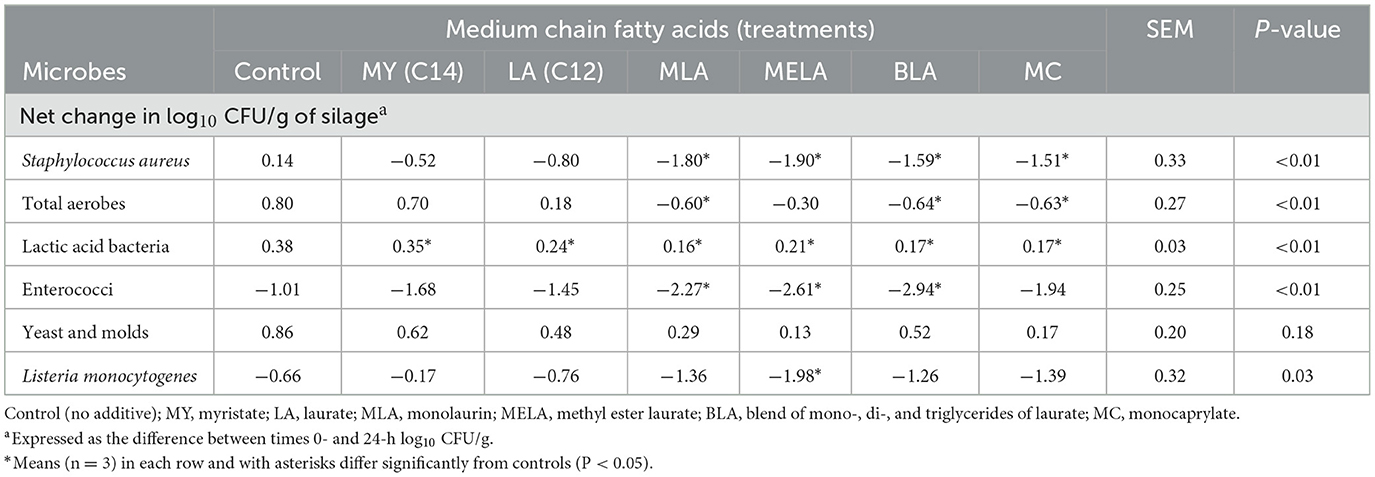
Las poblaciones de BAL aumentaron ligeramente, en 0,17-0,38 log10 UFC/g en todos los ensilajes tratados, así como en el ensilado no tratado, pero el aumento fue menor que en el control no tratado para todos los ensilajes tratados (P < 0,01), incluidos aquellos con las formas libres no esterificadas o las formas esterificadas del AGCM (Tabla 1). Las poblaciones de enterococos disminuyeron de 10 a 100 veces con respecto a sus niveles iniciales en los ensilajes no tratados y tratados, pero el cambio neto con respecto a los niveles iniciales en los ensilajes tratados con MLA, MELA y BLA fue significativamente diferente (P < 0.01) del cambio neto en los controles. El cambio neto en las concentraciones de levadura y mohos, cuando se comparó con el cambio neto observado en los controles no tratados, no difirió (P > 0.05), y las poblaciones aumentaron más variablemente de 0.13 a 1.39 log10 UFC/g (Tabla 1). La población Scott A inoculada experimentalmente con L. monocytogenes también respondió de manera variable a los tratamientos con AGCM en el ensilado de maíz expuesto al aire, con poblaciones disminuidas desde sus concentraciones iniciales en todos los ensilajes tratados y no tratados (Tabla 1). Sin embargo, la disminución de 1.98 log10 UFC/g observada en el ensilado tratado con MELA fue el único ensilado tratado que difirió significativamente (P < 0.05) que el cambio neto de -0.66 log10 UFC/g observado con los controles no tratados (Tabla 1).
3.2 Fermentación ruminal in vitro
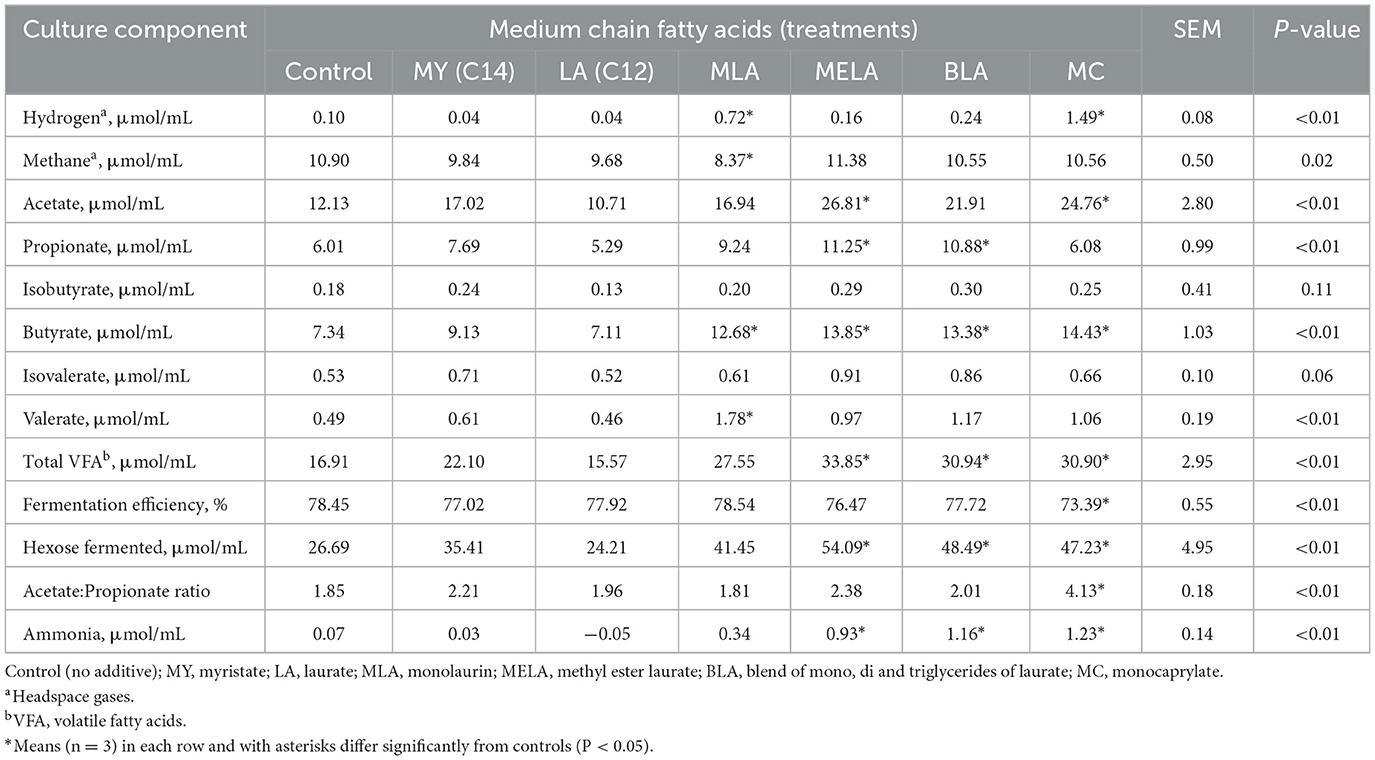
En la Tabla 2 se muestra un resumen de los efectos de los AGCM sobre las características de la fermentación ruminal in vitro. Las acumulaciones de hidrógeno en la suspensión de fluido ruminal in vitro incubada con ensilaje que habían sido tratadas con MLA o MC se incrementaron (P < 0.01) en comparación con las acumulaciones observadas en las suspensiones de fluido ruminal control incubadas in vitro con ensilaje que no habían sido tratadas con ninguna de las preparaciones de AGCM (Tabla 2). El metano disminuyó (P < 0.01) en un 23% en las suspensiones de fluido ruminal incubadas in vitro con la adición de MLA en comparación con lo producido por las suspensiones ruminales control incubadas con ensilaje no tratado (Tabla 2). Las acumulaciones de acetato aumentaron (P < 0.01) más de 2 veces en las suspensiones de fluido ruminal incubadas in vitro con ensilaje tratado con MELA o MC en comparación con la cantidad producida por las suspensiones de fluido ruminal control incubadas con el ensilado no tratado (Tabla 2). Las acumulaciones de propionato aumentaron (P < 0,01) más de 1,8 veces en las suspensiones ruminales incubadas con MELA o BLA cuando se compararon las acumulaciones en las suspensiones de líquido ruminal control (Tabla 2). Las acumulaciones de isobutirato e isovalerato en las suspensiones de líquido ruminal incubadas con los ensilajes tratados no difirieron (P > 0.05) en comparación con las acumulaciones observadas en las incubaciones de líquido ruminal control (Tabla 2). Sin embargo, las acumulaciones de butirato en las suspensiones de líquido ruminal incubadas con MLA, MELA, BLA o MC aumentaron (P < 0,01) de 1,7 a 2 veces en comparación con las incubaciones de control (Tabla 2). La acumulación de valerato en las suspensiones de líquido ruminal incubadas con MLA se incrementó (P < 0,01) 3,6 veces en comparación con las incubaciones de líquido ruminal control. Debido al aumento de las acumulaciones de ácidos grasos individuales observadas anteriormente, las acumulaciones de AGV totales y las estimaciones estequiométricas de la hexosa fermentada en las incubaciones in vitro en el fluido ruminal aumentaron (P < 0,01) de 1,8 a 2 veces debido al ensilado tratado con MELA, BLA o MC (Tabla 2). La relación acetato:propionato se incrementó (P < 0.05) 2 veces en las suspensiones de fluido ruminal incubadas con ensilaje tratado con MC en comparación con las incubaciones de fluido ruminal control, lo que consecuentemente contribuyó a una disminución del 6.4% (P < 0.05) en la eficiencia de fermentación estimada de las incubaciones de ensilaje tratadas con MC (Tabla 2). Las acumulaciones de amoníaco aumentaron (P < 0.01) de 13 a casi 18 veces en las suspensiones de líquido ruminal incubadas con MELA, BLA o MC en comparación con las incubaciones de líquido ruminal de control (Tabla 2).
4 Discusión
4.1 Concentraciones microbianas seleccionadas
El laureado o el miristato no influyeron en el crecimiento de S. aureus; sin embargo, el número de esta bacteria se redujo de 10 a 100 veces cuando los ensilajes se trataron con los monoglicéridos de los ácidos caprílico o láurico, así como con MELA. En concordancia, Arzola-Álvarez et al. (18) no reportaron cambios en el crecimiento de S. aureus en ensilaje de maíz expuesto al aire tratado con AGCM. Sin embargo, estos resultados contrastan con los de Ontiveros-Magadan et al. (17) en un estudio similar con ensilado de maíz expuesto al aire. Observaron reducciones en las poblaciones de S. aureus en ensilajes tratados con LA o con MLA (un glicérido de laureado). Probablemente, estos diferentes resultados podrían atribuirse a las diferentes dosis de AGCM utilizadas. En el presente estudio y en el estudio de Arzola-Álvarez et al. (18) se utilizaron 0,3 mg/kg de AGCM, mientras que Ontiveros-Magadan et al. (17) utilizaron 0,5 mg/kg de AGCM. Además de la dosis, un tema que requiere más investigación se relaciona con la eficacia de las formas químicas de los AGCM para inhibir o reducir el crecimiento de S. aureus en el ensilado de maíz expuesto al aire, considerando que en el presente estudio, los glicéridos de los AGCM fueron más efectivos que los ácidos grasos libres (laureado o miristato). Por ejemplo, Kelsey et al. (25), en cultivos puros de S. aureus, informaron una disminución similar del crecimiento con tratamientos con el laurato MCFA, el caprato, el miristato y el monolaurato de glicerol.
El laureado, el miristato o el ácido láurico de éster metílico tuvieron poca o ninguna influencia negativa en el crecimiento total de los aerobios; Sin embargo, los monoglicéridos de los ácidos caprílico y láurico redujeron las poblaciones totales de aerobios, pero la disminución fue <10 veces. Como se ha revisado en otros casos (26, 27), no es inesperado que los monoglicéridos de los ácidos grasos de cadena media, que poseen características de surfactante y formación de micelas más fáciles, puedan exhibir más potencia que los ácidos grasos libres contra ciertos microbios, pero no necesariamente todos. Arzola-Álvarez et al. (18) no reportaron cambios en el crecimiento aerobio total en ensilado de maíz expuesto al aire tratado con AGCM en forma libre. Sin embargo, estos resultados contrastan con los de Ontiveros-Magadan et al. (17) quienes encontraron reducciones aerobias totales cuando los ensilajes fueron tratados con LA o con MLA. Como se ha mencionado anteriormente, las dosis utilizadas en los diferentes estudios podrían estar relacionadas con las diferencias en los resultados.
Todos los ácidos grasos redujeron moderadamente el crecimiento neto de las BAL, en comparación con los controles, lo que indica un efecto bacteriostático modesto, en lugar de bactericida, este último que se manifestaría como una disminución de las concentraciones con respecto a su nivel inicial. De acuerdo con Ellis et al. (28), las bacterias del ácido láctico generalmente consisten en múltiples géneros diferentes de bacterias Gram-positivas, como ciertas especies de Bacillus, así como especies pertenecientes a Enterococcus, Lactobacillus, Lactococcus, Pediococcus y Weissella (27). En consecuencia, la disminución de BAL en el ensilado tratado puede deberse, al menos parcialmente, a la disminución observada de enterococos que se analiza a continuación. Los resultados actuales son consistentes con los de Arzola-Álvarez et al. (18) quienes observaron reducciones de BAL con tratamientos de AGCM en ensilado de maíz expuesto al aire. En cultivos puros de microorganismos relacionados con el ensilaje, Woolford (29) reportó que el aumento en el tamaño de la cadena de ácidos grasos tuvo un mayor efecto antimicrobiano. Además, informaron que los ácidos caprílico (octanoico), cáprico (decanoico) y láurico (dodecanoico) fueron efectivos contra las BAL, levaduras, mohos y bacterias Gram-positivas, mientras que las bacterias Gram-negativas fueron menos sensibles que las bacterias Gram-positivas a los ácidos grasos analizados. Las bacterias grampositivas, que carecen de una capa de membrana externa, generalmente se reconocen como más susceptibles al daño de la pared celular causado por los AGCM o sus ésteres que las bacterias gramnegativas, presumiblemente debido a la capacidad limitada de los compuestos para pasar a través de la capa de membrana externa de las bacterias gramnegativas (26). En diferentes resultados, Ontiveros-Magadan et al. (17) no encontraron cambios en el crecimiento de BAL a las 24 h de incubación en ensilaje de maíz expuesto al aire tratado con LA o MLA. Las BAL son bacterias deseadas que mantienen las condiciones ácidas en los ensilados; sin embargo, cuando el ensilado está expuesto al aire, las BAL exhiben un crecimiento reducido y, en el estudio de Ontiveros-Magadan et al. (17), las BAL estuvieron ausentes en el grupo control y solo tuvieron pequeños cambios en los ensilajes expuestos al aire tratados con LA o MLA.
El laureado y el miristato no influyeron en el crecimiento de los enterococos, sin embargo, el éster metílico y los glicéridos del ácido láurico sí redujeron su número. Se ha demostrado que el crecimiento de enterococos está reducido en el ensilado de maíz expuesto al aire tratado con AGCM (18). Además, Bunkova et al. (5) observaron el crecimiento inhibitorio in vitro de bacterias Gram-positivas (Bacillus cereus, B. subtilis, E. faecalis, Micrococcus luteus y S. aureus) tratadas con monoglicéridos que contienen AGCM, incluidos C8, C10, C12 y otros ácidos grasos. También informaron que las bacterias gramnegativas analizadas (Citrobacter freundii, E. coli, Proteus mirabilis, Pseudomonas aeruginosa y Salmonella enterica) eran más resistentes a los mismos monoglicéridos.
En el estudio actual, los tratamientos no influyeron en el crecimiento de levaduras y mohos en el ensilado de maíz expuesto al aire. La inhibición del crecimiento de estos microorganismos es un objetivo deseable debido al efecto de deterioro de los ensilados durante la fase aeróbica después de abrir el silo. No hay un efecto consistente de los AGCM sobre el crecimiento de levaduras y mohos. Woolford (29), utilizando cultivos puros, informó reducciones en el número de levaduras y mohos asociados con los ensilajes cuando se trataron con ácidos grasos C10 y C12. Arzola-Álvarez et al. (18) encontraron reducciones en el número de levaduras y mohos mediante una mezcla de tratamiento de ácidos grasos C8:C10 en ensilado expuesto al aire; sin embargo, no hubo reducciones cuando se utilizaron los mismos ácidos grasos por separado. Ontiveros-Magadan et al. (17), utilizando ensilado de maíz expuesto al aire tratado con LA (C12), encontraron reducciones de levaduras y mohos; Y los mismos autores señalaron que, desde una perspectiva práctica, esas reducciones serían modestas e intrascendentes.
Las poblaciones de Listeria monocytogenes solo disminuyeron con el MELA. En concordancia, Arzola-Álvarez et al. (18) no reportaron cambios en el crecimiento de L. monocytogenes en ensilado de maíz expuesto al aire tratado con AGCM en formas libres. En cultivos puros, L. monocytogenes tenía poblaciones reducidas cuando se trataba con MLA (30, 31). Ontiveros-Magadan et al. (17), utilizando ensilado de maíz expuesto al aire, observaron reducciones en las poblaciones de L. monocytogenes tratadas con LA o con MLA (un glicérido de laureado), aunque las reducciones fueron más evidentes con LA que con MLA. Como se indicó anteriormente, las concentraciones de AGCM podrían explicar estas diferentes respuestas al crecimiento de L. monocytogenes.
4.2 Fermentación ruminal in vitro
La producción de hidrógeno se incrementó mediante la monolaurina y los glicéridos monocaprílicos, mientras que el LA y el MY no influyeron en la producción de hidrógeno. De acuerdo, investigaciones previas con ensilado de maíz expuesto al aire mostraron que el AGCM tenía una influencia mínima en la fermentación del rumen (17, 18). En el presente estudio, solo el MC tuvo una acumulación de hidrógeno que se asoció con efectos negativos sobre la eficiencia de la fermentación, la relación acetato:propionato o la producción de amoníaco.
En el estudio actual, el AGCM tuvo una influencia mínima en la producción de metano, solo siendo reducido por la monolaurina. En concordancia, Arzola-Álvarez et al. (18) utilizando ensilado de maíz expuesto al aire reportaron una influencia mínima de los AGCM en la producción de metano ruminal. En contraste, Ontiveros-Magadan et al. (17) encontraron reducciones significativas de metano con LA y MLA en ensilado de maíz expuesto al aire; como se ha dicho, esta diferencia se debe a la mayor dosis de AGCM en el estudio de Ontiveros-Magadan et al. (17). El ácido láurico mostró una reducción de metano en altas concentraciones de LA (14). Goel et al. (32) observaron una disminución de la producción de metano ruminal in vitro con ácido cáprico a 400 y 600 mg/l, pero no se observaron reducciones a 200 mg/l.
El éster metílico, el ácido láurico y los glicéridos monocaprílicos aumentaron la producción de acetato. Este resultado coincidió con los de Arzola-Álvarez et al. (18), quienes utilizaron ensilaje de maíz expuesto al aire, quienes reportaron una mínima influencia de los AGCM en la producción de acetato ruminal. Jordan et al. (33) también encontraron un efecto mínimo del aceite de coco (rico en AGCM) sobre las proporciones de acetato, propionato y butirato, con raciones mixtas totales para el ganado vacuno. Abel et al. (12) observaron mayor contenido de acetato y metano en los ensilajes tratados con ácido caprílico (C8) que en los ensilados tratados con cáprico (C10).
El éster metílico, el ácido láurico y la mezcla de ácido láurico aumentaron el propionato. Este resultado coincidió con los de Arzola-Álvarez et al. (18) que utilizaron ensilaje de maíz expuesto al aire, y quienes reportaron una influencia mínima de los AGCM en formas libres sobre la producción de propionato ruminal. Jordan et al. (33) también encontraron un efecto mínimo del aceite de coco (rico en AGCM) sobre las proporciones de acetato, propionato y butirato, con raciones mixtas totales para el ganado vacuno. Abel et al. (12) no observaron mayor contenido de propionato en los ensilajes tratados con ácido caprílico (C8) que en los ensilados tratados con cáprico (C10).
El contenido de butirato se incrementó por MLA, MC, MELA y por el BLA. Estos resultados concuerdan con los de Arzola-Álvarez et al. (18), quienes utilizaron ensilaje de maíz expuesto al aire, quienes reportaron una mínima influencia de AGCM en formas libres sobre la producción de butirato ruminal. Jordan et al. (33) también encontraron un efecto mínimo del aceite de coco (rico en AGCM) sobre las proporciones de acetato, propionato y butirato con las raciones mixtas totales para el ganado vacuno. Abel et al. (12) no observaron mayor contenido de butirato en los ensilajes tratados con ácido caprílico (C8) que en los ensilados tratados con cáprico (C10).
El contenido de isobutirato e isovalerato no se vio influenciado por los tratamientos, y el contenido de valerato solo aumentó con el MLA. Estos resultados concuerdan con los de Arzola-Álvarez et al. (18) utilizando ensilado de maíz expuesto al aire. Abel et al. (12) no observaron mayores niveles de isobutirato en los ensilajes tratados con ácido caprílico (C8) que en los ensilados tratados con cáprico (C10).
Los AGV totales, la hexosa fermentada y el amoníaco fueron incrementados por MELA, BLA y por MC. Estos resultados concuerdan con los de Arzola-Álvarez et al. (18) trabajando con ensilaje de maíz expuesto al aire y reportaron una influencia mínima de los AGCM en las mismas variables de las fermentaciones ruminales. Jordan et al. (33) reportaron resultados diferentes, quienes observaron reducciones del contenido total de AGV en las raciones mezcladas para el ganado vacuno. Abel et al. (12) observaron mayores reducciones en el contenido de amoníaco en los ensilajes tratados con ácido caprílico (C8) que en los ensilajes tratados con cáprico (C10).
El glicérido monocaprílico redujo la eficiencia de la fermentación y aumentó la relación acetato:propionato. Estas son características de fermentación no deseadas en el rumen y podrían estar relacionadas con las acumulaciones de hidrógeno observadas con este tratamiento. En el presente estudio, la eficiencia de la fermentación no se vio influenciada por LA o MY; sin embargo, en contraste, Arzola-Álvarez et al. (18) encontraron una eficiencia fermentativa reducida con AGCM. Se justifica más investigación sobre este tema, en particular sobre el efecto residual en los ensilajes tratados de los diferentes AGCM sobre las características de la fermentación ruminal.
La producción de amoníaco ruminal no fue modificada por LA, MY, ni por MLA. Por el contrario, el contenido de amoníaco fue mayor, aunque no necesariamente significativamente, en las suspensiones de fluido ruminal incubadas con ensilaje tratado con el AGCM esterificado que con los ácidos grasos libres. Teniendo en cuenta que los tratamientos con AGCM esterificados de ensilado expuesto al aire dieron lugar a disminuciones apreciables en las concentraciones bacterianas de estafilococos, enterococos y lactato, es razonable sospechar que estas poblaciones bacterianas pueden estar correlacionadas con una absorción apreciable de amoníaco dentro de la microbiota ruminal. La falta de respuesta en el ensilado de maíz expuesto al aire a los AGCM sobre la producción de amoníaco ruminal coincidió con los resultados de Arzola-Álvarez et al. (18). Sin embargo, consistentemente, la producción ruminal de amoníaco se ha reducido con la suplementación con LA, aunque las cantidades de ácido láurico administradas al ganado en esos estudios (240 g/vaca por día) probablemente habrían excedido la cantidad, 0,15 mg/mL de base de líquido ruminal, incluida en nuestras incubaciones in vitro (34, 35). Suponiendo un volumen ruminal de 60 litros de las vacas de 660-680 kg, la concentración ruminal máxima habría sido de ~4 mg de laurato/mL. La reducción del amoníaco ruminal es un efecto deseable del LA debido a la reducción de la proteólisis microbiana y la reducción de las poblaciones microbianas, con el efecto neto de la mejora del nitrógeno por parte del animal rumiante. Las indicaciones de nuestro estudio son que una dosis más alta de ácidos grasos puede causar una reducción en el contenido de amoníaco en la fermentación ruminal.
5 Conclusión
Los resultados del presente estudio apoyan nuestra hipótesis de que el tratamiento del ensilado de maíz expuesto aeróbicamente con preparaciones de MCFA DE MLA, MELA, BLA o MC no solo evitó la propagación de S. aureus inoculado experimentalmente, sino que logró reducciones de casi 2 log10 veces en el recuento de colonias de este patógeno causante de mastitis. Sin embargo, estos tratamientos también causaron disminuciones significativas, aunque leves (<1 log10 veces), en las poblaciones potencialmente beneficiosas de las poblaciones autóctonas de bacterias lácticas, así como reducciones de casi 2 log10 veces en las poblaciones enterocócicas. Los efectos de estas preparaciones de AGCM, así como de las formas libres de MY o LA, sobre los aerobios totales, levaduras y mohos y sobre L. monocytogenes inoculados experimentalmente fueron variables. Las pruebas de los posibles efectos adversos del ensilado tratado con AGCM en la fermentación ruminal durante la incubación in vitro en condiciones de simulación de hábitat ruminal revelaron pocos o ningún efecto perjudicial sobre las características beneficiosas de la fermentación ruminal, con la excepción de aumentos significativos en las acumulaciones de amoníaco en las suspensiones de fluidos ruminales incubadas con MELA, BLA y MC, lo que implica un posible efecto sobre la asimilación de amoníaco dentro de la población mixta de rumen Microbios.
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán facilitados por los autores, sin reservas indebidas.
Contribuciones de los autores
JS-C: Conceptualización, Análisis formal, Investigación, Metodología, Supervisión, Validación, Redacción – borrador original, Redacción – revisión y edición. CA-A: Conceptualización, Análisis formal, Investigación, Validación, Redacción – borrador original, Redacción – revisión y edición. MH: Conceptualización, Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. MF: Análisis formal, Metodología, Redacción – borrador original, Redacción – revisión y edición. OR-B: Conceptualización, Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. YC-C: Conceptualización, Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. MO-M: Conceptualización, Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. BJ: Conceptualización, Análisis formal, Metodología, Redacción – borrador original, Redacción – revisión y edición. TC: Conceptualización, Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. TP: Conceptualización, Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. AZ-S: Análisis formal, Investigación, Metodología, Redacción – borrador original, Redacción – revisión y edición. RA: Conceptualización, Análisis formal, Investigación, Metodología, Administración de proyectos, Recursos, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este proyecto fue financiado por fondos de investigación asignados por el Departamento de Agricultura de los Estados Unidos.
Reconocimientos
La asistencia técnica experta de Jackie Kotzur es muy apreciada.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Pahlow G, Muck RE, Driehuis F, Oude Elferink SJWH, Spoelstra SF. Microbiología del ensilado. En:Buxton DR, Muck RE, Harrison HJ, , editores. Ciencia y Tecnología del Ensilaje (Serie Agronomía Nº 42). Madison, WI: Sociedad Americana de Agronomía (2003). págs. 31-93.
2. Driehuis F, Oude Elferink SJWH, Van Wikleaar PG. Lactobacillus buchneri mejora la estabilidad aeróbica del maíz de cultivo entero a escala de laboratorio y granja, pero no afecta la ingesta de alimento y la producción de leche de las vacas lecheras. En: Proc. 12th Intl. Silage Conf., Uppsala, Suecia. Univ. Agric. Sci. Uppsala (1999). págs. 106-7.
3. Borreani, G, Fernandes Bernardes T, Tabacco E. El deterioro aeróbico influye en la calidad fermentativa, microbiológica y nutricional de los ensilados de maíz y sorgo en las granjas en cadenas de producción de leche y queso de alta calidad. R Bras Zootec. (2008) 37:68–77. doi: 10.1590/S1516-35982008001300009
4. Galbraith H, Miller TB, Paton AM, Thompson JK. Actividad antibacteriana de los ácidos grasos de cadena larga y la reversión con calcio, magnesio, ergocalciferol y colesterol. J Appl Bacteriol. (1971) 34:803–13. doi: 10.1111/j.1365-2672.1971.tb01019.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Bunkova L, Bunka F, Janis R, Krejci J, Dolezalkova I, Pospisil Z, et al. Comparación del efecto antibacteriano de siete 1-monoglicéridos sobre patógenos transmitidos por los alimentos o bacterias de deterioro. Acta Vet Brno. (2011) 80:29–39. doi: 10.2754/avb201180010029
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Nair MKM, Vasudevan P, Hoagland T, Venkitanarayanan K. Inactivación de Escherichia coli O157: H7 y Listeria monocytogenes en leche por ácido caprílico y monocaprilina. Microbiol alimentario. (2004) 21:611–6. doi: 10.1016/j.fm.2004.01.003
7. Skrivanová E, Savka OG, Marounek M. Efecto in vitro de los ácidos grasos C2-C18 sobre Salmonellas. Folia Microbiol. (2004) 49:199–202. doi: 10.1007/BF02931402
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. de los Santos FS, Donoghue AM, Venkitanarayanan K, Dirain ML, Reyes-Herrera I, Blore PJ, et al. El ácido caprílico suplementado en los piensos reduce la colonización entérica por Campylobacter jejuni en pollos de engorde de diez días de edad. Poult Sci. (2008) 87:800–4. doi: 10.3382/ps.2007-00280
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Skrivanová E, Molatová Z, Skrivanová V, Marounek M. Actividad inhibitoria de la leche de conejo y los ácidos grasos de cadena media contra Escherichia coli O128 enteropatógena. Microbiol veterinario. (2009) 135:358–62. doi: 10.1016/j.vetmic.2008.09.083
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Cenesiz AA, Çiftci I. Efectos moduladores de los ácidos grasos de cadena media en la nutrición y salud avícola. Mundos Poult Sci J. (2020) 76:234–48. doi: 10.1080/00439339.2020.1739595
11. McDonald P, Henderson AR. El uso de ácidos grasos como aditivos para el ensilado de hierba. J Sci Alimentos Agric. (1974) 25:791–5. doi: 10.1002/jsfa.2740250708
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Abel H, Immig I, Harman E. Efecto de la adición de ácido caprílico y cáprico a la hierba sobre las características de fermentación durante el ensilado y en el sistema ruminal artificial RUSITEC. Anim Feed Sci Tech. (2002) 99:65–72. doi: 10.1016/S0377-8401(02)00084-6
13. Queiroz OCM, Ogunade IM, Weinberg Z, Adesogan AT. Revisión de ensilajes: patógenos transmitidos por alimentos en ensilaje y su mitigación por aditivos de ensilaje. J Dairy Sci. (2018) 101:4132–42. doi: 10.3168/jds.2017-13901
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Dohme F, Machmüller A, Wasserfallen A, Kreuzer M. Metanogénesis ruminal influenciada por ácidos grasos individuales suplementados para dietas completas de rumiantes. Lett Appl Microbiol. (2001) 32:47–51. doi: 10.1046/j.1472-765x.2001.00863.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Soliva CR, Hindrichsen IK, Meile L, Kreuzer M, Machmuller A. Efectos de las mezclas de ácido láurico y mirístico sobre los metanógenos ruminales y la metanogénesis in vitro. Lett Appl Microbiol. (2003) 37:35–9. doi: 10.1046/j.1472-765X.2003.01343.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Joch M, Vadronová M, Cešpiva M, Zabloudilová P, Výborná A, Tyrolová Y, et al. La mezcla de ácido cáprico y láurico disminuyó la producción de metano en el rumen, mientras que la combinación con nitrato no tuvo ningún beneficio adicional en la reducción de metano. Ann Anim Sci. (2023) 23:799–808. doi: 10.2478/aoas-2023-0010
17. Ontiveros-Magadan M, Anderson RC, Ruiz-Barrera O, Arzola-Álvarez C, Salinas-Chavira J, Hume ME, et al. Evaluación de compuestos antimicrobianos para inhibir el crecimiento de bacterias patógenas grampositivas o resistentes a los antimicrobianos en ensilado expuesto al aire. Can J Anim Sci. (2022) 102:75–84. doi: 10.1139/cjas-2021-0061
18. Arzola-Álvarez C, Ruiz-Barrera O, Castillo-Castillo Y, Ontiveros M, Fonseca M, Jones BW, et al. Efectos en el ensilado de maíz expuesto al aire de ácidos grasos de cadena media sobre microbios de deterioro seleccionados, patógenos zoonóticos y fermentación ruminal in vitro. J Environ Sci Health B. (2023) 58:45–50. doi: 10.1080/03601234.2023.2168449
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Božic A, Anderson RC, Arzola-Álvarez C, Ruiz-Barrera O, Corral-Luna A, Castillo-Castillo Y, et al. Inhibición de estafilococos multirresistentes por clorato de sodio y compuestos selectos de ácidos grasos nitro y de cadena media. J Appl Microbiol. (2019) 128:1508–16. doi: 10.1111/jam.14232
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Leyendecker SA, Callaway TR, Anderson RC, Nisbet DJ. Nota técnica sobre un método muy simplificado para recolectar líquido ruminal usando un colador de pintura de nylon. J Sci Alimentos Agric. (2004) 84:387–9. doi: 10.1002/jsfa.1673
21. Cagle CM, Batista LFD, Anderson RC, Fonseca MA, Cravey MD, Julien C, et al. Evaluación de diferentes niveles de inclusión de levaduras vivas secas, impactos sobre diversos parámetros ruminales y digestibilidades in situ de materia seca y fibra detergente neutra en ganado vacuno de engorde y engorde. J Anim Sci. (2019) 97:4987–98. doi: 10.1093/jas/skz342
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Allison MJ, Mayberry WR, McSweeney CS, Stahl DA. Synergistes jonesii, gen. nov, sp nov: bacteria ruminal que degrada los piridinodioles tóxicos. Syst Appl Microbiol. (1992) 15:522–9. doi: 10.1016/S0723-2020(11)80111-6
23. Chaney AL, Marbach EP. Reactivos modificados para la determinación de urea y amoníaco. Clin Chem. (1962) 8:130–2. doi: 10.1093/clinchem/8.2.130
24. Chalupa W. Manipulación de la fermentación ruminal. J Anim Sci. (1977) 46:585–99. doi: 10.2527/jas1977.453585x
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Kelsey JA, Bayles KW, Shafii B, McGuire MA. Los ácidos grasos y los monoacilgliceroles inhiben el crecimiento de Staphylococcus aureus. Lípidos. (2006) 41:951–61. DOI: 10.1007/S11745-006-5048-Z
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Barker LA, Bakkum BW, Chapman C. El uso clínico de la monolaurina como suplemento dietético: una revisión de la literatura. J Quiropract Med. (2019) 18:305–10. doi: 10.1016/j.jcm.2019.02.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Jackman JA, Boyd RD, Elrod CC. Ácidos grasos de cadena media y monoglicéridos como aditivos alimentarios para la producción porcina: hacia la mejora de la salud intestinal y la mitigación de patógenos en los piensos. J Anim Sci Biotechnol. (2020) 11:44. doi: 10.1186/s40104-020-00446-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Ellis JK, Hindrichsen IK, Klop G, Kinley RD, Milora N, Bannink A, et al. Efectos de la inoculación de ensilaje de bacterias lácticas sobre la emisión de metano y la productividad del ganado lechero Holstein Friesian. J Dairy Sci. (2016) 99:7159–74. doi: 10.3168/jds.2015-10754
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Woolford MK. Cribado microbiológico de los ácidos grasos de cadena lineal (Cl-C12) como posibles aditivos de ensilaje. J Sci Alimentos Agric. (1975) 26:219–28. doi: 10.1002/jsfa.2740260213
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Razavi-Rohani SM, Griffiths MW. El efecto del laurato de mono y poliglicerol sobre el deterioro y las bacterias patógenas asociadas a los alimentos. J Alimentos Saf. (1994) 14:131–51. doi: 10.1111/j.1745-4565.1994.tb00590.x
31. Wang LL, Johnson EA. Inhibición de Listeria monocytogenes por ácidos grasos y monoglicéridos. Appl Environ Microbiol. (1992) 58:624–9. DOI: 10.1128/AEM.58.2.624-629.1992
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Goel G, Arvidsson K, Vlaeminck B, Bruggeman G, Deschepper K, Fievez, V. Efectos del ácido cáprico sobre la metanogénesis ruminal y la biohidrogenación del ácido linoleico y α-linolénico. Animal. (2009) 3:810–6. doi: 10.1017/S1751731109004352
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Jordan E, Lovett DK, Monahan FJ, Callan J, Flynn B, O’mara FP. Efecto del aceite de coco refinado o harina de copra en la producción de metano y en la ingesta y rendimiento de novillas de carne. J Ciencia Animal. (2006) 84:162–70. doi: 10.2527/2006.841162x
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Hristov AN, Pol MV, Agle M, Zaman S, Schneider C, Ndegwa P, et al. Efecto del ácido láurico y el aceite de coco sobre la fermentación ruminal, la digestión, las pérdidas de amoníaco del estiércol y la composición de ácidos grasos de la leche en vacas lactantes. J Dairy Sci. (2009) 92:5561–82. doi: 10.3168/jds.2009-2383
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Hristov AN, Lee C, Cassidy T, Long M, Heyler K, Corl B, et al. Efectos de los ácidos láurico y mirístico sobre la fermentación ruminal, la producción y la composición de ácidos grasos de la leche en vacas lecheras lactantes. J Dairy Sci. (2011) 94:382–95. doi: 10.3168/jds.2010-3508
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: ácidos grasos, fase aeróbica, ensilado de maíz, microbios, rumen
Cita: Salinas-Chavira J, Arzola-Álvarez C, Hume ME, Fonseca M, Ruiz-Barrera O, Castillo-Castillo Y, Ontiveros-Magadan M, Jones B, Crippen TL, Poole TL, Zúñiga-Serrano A y Anderson RC (2024) Influencia de los ácidos grasos de cadena media en microbios seleccionados y en la fermentación ruminal in vitro de ensilado de maíz expuesto al aire. Frente. Vet. Sci. 11:1416695. doi: 10.3389/fvets.2024.1416695
Recibido: 12 de abril de 2024; Aceptado: 12 de agosto de 2024;
Publicado: 11 de septiembre de 2024.
Editado por:
Matteo Dell’Anno, Universidad de Milán, Italia
Revisado por:
Roberto Senas Cuesta, Universidad de Arkansas, Estados
Unidos Burarat Phesatcha, Universidad Tecnológica de Rajamangala Isan, Tailandia
Copyright © 2024 Salinas-Chavira, Arzola-Alvarez, Hume, Fonseca, Ruiz-Barrera, Castillo-Castillo, Ontiveros-Magadan, Jones, Crippen, Poole, Zúñiga-Serrano y Anderson. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Atribución (CC BY). S
*Correspondencia: Robin C. Anderson, robin.anderson@usda.gov
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía