
La harina de soja fermentada modificó la microbiota ruminal y aumentó el nivel sérico de prolactina

La harina de soja fermentada modificó la microbiota ruminal y aumentó el nivel sérico de prolactina en vacas Holstein lactantes
 Jiyou Zhang1Feng Guan1Shana Huang2Yumin Ma3Shibao Wen4
Jiyou Zhang1Feng Guan1Shana Huang2Yumin Ma3Shibao Wen4 Wei Jin1*
Wei Jin1* Shengyong Mao1
Shengyong Mao1- 1Laboratorio de Microbiología Gastrointestinal, Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Nanjing, Nanjing, China
- número arábigoCentro de Extensión de Tecnología de Agricultura y Ganadería de Mongolia Interior, Hohhot, China
- 3Jiangsu Jiahui Biotechnology Co., Ltd., Haian, China
- 4Shanghai Menon Animal Nutrition Technology Co., Ltd., Shanghai, China
Este estudio tuvo como objetivo investigar los efectos de la harina de soja fermentada (FSM) sobre la producción de leche, los parámetros sanguíneos y la fermentación ruminal y la comunidad microbiana en vacas lecheras. En este estudio, se utilizaron 48 vacas Holstein sanas (paridad, 3,0 ± 0,6; días en leche, 86,0 ± 6,7). Las vacas fueron asignadas aleatoriamente a cuatro grupos (CON, T-200, T-400 y T-600) con 12 vacas por grupo. Las vacas en CON no se suplementaron con FSM. Las vacas en T-200, T-400 y T-600 fueron suplementadas con 200, 400 y 600 g/cabeza/día FSM, respectivamente. Este estudio tuvo una duración de 5 semanas (1 semana de adaptación y 4 semanas de tratamiento). Los resultados mostraron que la FSM no afectó la producción de leche ni los componentes de la leche (p > 0,05). En el suero, FSM aumentó considerablemente la prolactina (p < 0,01) y se observó un efecto de dosificación. La aspartato aminotransferasa y la proteína total fueron las más altas en el T-400 (p < 0,05), y los triglicéridos fueron los más bajos en el T-200 (p < 0,05), y no hubo diferencia para las 3 mediciones entre los otros 3 grupos (p > 0,05). En el rumen, la FSM no afectó el pH, la proteína cruda microbiana, el acetato, el propionato, el butirato, el valerato, los ácidos grasos volátiles totales y la relación acetato:propionato (p > 0,05), solo cambió el NH3-N, el isobutirato y el isovalerato (p < 0,05). Los resultados de la secuenciación de los genes de ARNr 16S de las bacterias ruminales mostraron que el FSM disminuyó la riqueza (p < 0,05) y la uniformidad (p < 0,05) de las comunidades bacterianas. El análisis de PCoA mostró que la FSH alteró la comunidad bacteriana ruminal (ANOSIM, R = 0,108, p = 0,002). En la abundancia relativa de filos, FSM aumentó Firmicutes (p = 0.015) y Actinobacteriota (p < 0.01) y Patescibacteria (p = 0.012), disminuyó Bacteroidota (p = 0.024). En la abundancia relativa de los géneros, FSM aumentó el grupo Christensenellaceae R-7 (p = 0.011), Lactococcus (p < 0.01), Candidatus Saccharimonas (p < 0.01), Olsenella (p < 0.01), disminuyó Muribaculaceae_norank (p < 0.01). En conclusión, la FSM suplementada alteró los parámetros de fermentación ruminal y la comunidad bacteriana, y aumentó el nivel de prolactina sérica en vacas Holstein lactantes. Estos hallazgos pueden proporcionar un enfoque para mantener el pico de lactancia en las vacas lecheras.
1 Introducción
La harina de soja fermentada (FSM) es una fuente de proteína vegetal de alta calidad para los animales, que contiene probióticos, enzimas digestivas, péptidos bioactivos, antioxidantes y proporciona efectos inmunomoduladores (1). Muchos estudios informaron que la alimentación de animales (cerdos, pollos y terneros) con FSM mostró efectos positivos con una mejor digestibilidad de los nutrientes y la salud intestinal y el rendimiento de la producción (2-4). Debido a la prohibición del uso de antimicrobianos promotores del crecimiento en la producción animal, el uso de FSM en rumiantes ha atraído un gran interés.
En el estudio de Kim et al. (5), el FSM se había utilizado en un iniciador de terneros y mostró efectos positivos sobre la salud y el crecimiento de los terneros. Tal y como demuestran Feizi et al. (4), el FSM mejoró la ingesta de iniciadores en los terneros y alteró la fermentación ruminal y la microbiota. En otro estudio de Rezazadeh et al. (6), la alimentación con FSM ayudó a los terneros a adaptarse al estrés del destete durante el clima frío. Un estudio en vacas lactantes informó que la alimentación con FSM aumentó la producción de proteína de la leche, la producción de grasa de la leche y la leche corregida con grasa, y disminuyó el recuento de células somáticas de la leche (7). Los estudios también demostraron que la alimentación con FSM altera los parámetros de fermentación ruminal y la microbiota ruminal en vacas Holstein lactantes (7, 8). Sin embargo, los resultados no fueron consistentes. Según lo informado por Wang et al. (8), FSM redujo la concentración total de ácidos grasos volátiles, la proporción de acetato a propionato y aumentó el porcentaje de propionato. De acuerdo con Amin et al. (7), el FSM incrementó el pH ruminal, el porcentaje de acetato y la relación acetato/propionato. La mayoría de los estudios sobre el uso de FSM en rumiantes se han centrado en terneros, pocos estudios investigaron las vacas lactantes, especialmente para las vacas en etapa temprana de lactancia (1).
Planteamos la hipótesis de que la alimentación con FSM podría causar cambios en la fermentación ruminal y en los parámetros de la microbiota y la sangre, lo que podría conducir a una mejora en el rendimiento lácteo de las vacas lecheras. En este estudio, nuestro objetivo fue explorar los efectos de la alimentación con FSM en el rendimiento de la leche, los parámetros sanguíneos y la fermentación ruminal y la comunidad bacteriana en vacas lecheras en la etapa temprana de la lactancia. Los resultados ofrecerían una referencia para la aplicación de FSM en la industria de vacas lecheras.
2 Materiales y métodos
2.1 Animales, dietas y manejo
Este experimento se llevó a cabo entre noviembre de 2020 y diciembre de 2020 en la granja lechera Jinshan Yinan de Shanghái (Shanghái, China). En este estudio, se utilizaron 48 vacas Holstein sanas en etapas similares de paridad (3.0 ± 0.6) y lactancia (86.0 ± 6.7 días en leche) y producción de leche (41.0 ± 2.8 kg). Las vacas fueron asignadas aleatoriamente a 4 grupos (CON, T-200, T-400 y T-600) con 12 vacas por grupo. Las vacas en CON no se suplementaron con FSM. Las vacas en T-200, T-400 y T-600 fueron suplementadas con 200, 400 y 600 g/cabeza/día FSM, respectivamente. Este estudio tuvo una duración de 5 semanas (1 semana de adaptación y 4 semanas de tratamiento). FSM (sustancia granular amarilla, fermentada por Saccharomyces cerevisiae y Bacillus subtilis) utilizada en este estudio fue comprada a Shanghai Yuanyao Agriculture Co., Ltd. La dieta básica utilizada en este estudio se formuló con base en las directrices de NRC (2001) para vacas lactantes. La composición nutricional de FSM se mostró en la Tabla Suplementaria S1. Los ingredientes y la composición química de la dieta básica se mostraron en la Tabla Suplementaria S2. Todas las vacas se alojaron en un establo de amarre, se ordeñaron tres veces al día con una máquina de ordeño totalmente automatizada (02,30, 10,30, 16:30) y se alimentaron con ración total mezclada tres veces al día (03,30, 10,30, 16:30), asegurando que las vacas tuvieran al menos 20 horas de libre acceso al alimento por día y libre acceso a agua dulce.
2.2 Muestreo
La producción de leche se determinó con un caudalímetro tunecino (JHF-G17, Sichuan Jinhaifeng Animal Husbandry Technology Co., Ltd., Sichuan, China). Las muestras de leche se recolectaron en los últimos 2 días de cada semana y se conservaron con dicromato de potasio, a 4 °C. Las muestras de leche recogidas del ordeño diario por la mañana, la tarde y la noche se mezclaron en una proporción de 4:3:3 antes de determinar la composición de la leche utilizando un analizador de infrarrojo cercano (MilkoScanTM 7 RM, Foss Electric, Dinamarca).
Las muestras de sangre se recogieron a través de la vena de la cola de las vacas antes de la alimentación matutina del último día del ensayo. Las muestras de sangre recogidas se sumergieron en agua tibia (37 °C) durante 10 min inmediatamente antes de centrifugar a 3.500 r/min durante 15 min. El sobrenadante se colectó y almacenó a -20 °C para la determinación de los índices bioquímicos séricos.
Las muestras de contenido ruminal se recogieron a las 4 h después de la alimentación matutina utilizando una sonda ruminal oral (Wuhan Kelibao Co., Ltd., Wuhan, China) el último día del ensayo. Para evitar la contaminación de la saliva, se desecharon los primeros 200 mL de líquido ruminal. Una parte del contenido ruminal se almacenó en nitrógeno líquido para la medición de la comunidad microbiana. Otra porción se filtró a través de cuatro capas de gasa esterilizada y se almacenó a -20 °C para la determinación de proteína bruta microbiana, NH3-N y ácidos grasos volátiles.
2.3 Análisis químico
Los índices bioquímicos séricos se midieron utilizando un analizador bioquímico totalmente automatizado (Vital Scientific NV, Países Bajos) siguiendo el procedimiento estándar. La prolactina (PRL) es una hormona productora de leche, se midió utilizando un kit de ensayo de inmunoabsorción ligado a enzimas (kit ELISA, Shanghái, China).
El valor de pH del fluido ruminal se midió utilizando un medidor de pH portátil (HI 9024C; Instrumentos HANNA, Woonsock, RI). La concentración de NH3-N en el líquido ruminal se determinó mediante un método colorimétrico de hipoclorito de fenol sodio según Weatherburn (9). El contenido microbiano de proteína bruta (MCP) en el líquido ruminal se determinó utilizando un método colorimétrico de azul brillante de Coomassie según Makkar et al. (10). La concentración de ácidos grasos volátiles (AGV) en el líquido ruminal se determinó mediante cromatografía de gases (GC-2014B, Shimadzu, Japón) equipada con una columna capilar (temperatura de la columna: 110 °C, espesor de la película: 30 m × 0,32 mm × 0,25 μm) (11).
2.4 Extracción de ADN genómico microbiano ruminal
El ADN genómico microbiano del rumen se extrajo utilizando métodos de extracción de fenol-cloroformo y lisis celular (12). La concentración de ADN se midió mediante un espectrofotómetro Nanodrop (Nyxor Biotech; París, Francia) y se almacenó a -80 °C para su posterior secuenciación.
2.5 Secuenciación MiSeq
Se utilizaron un par de cebadores de PCR para amplificar la región V3-V4 de los genes de ARNr 16S de las bacterias ruminales (13). Los cebadores fueron 341F (5-CCTAYGGGGRBGCASCAG-3) y 806R (5-GGACTACNNGGGTATCTAAT-3). Los amplicones se secuenciaron en una plataforma Illumina MiSeq PE 300 (Illumina Inc., San Diego, California, Estados Unidos) en un laboratorio comercial (Shanghai Biozeron Technology Co., Ltd., Shanghai, China). Los datos brutos se almacenaron en la base de datos Sequence Read Archive (SRA), el número de acceso es PRJNA1162692.
2.6 Análisis de datos
Se utilizó el software Trimmomatic (v.0.33) para recortar adaptadores y secuencias de baja calidad. Se utilizó el software FLASH (1.2.7) para concatenar segmentos emparejados en una secuencia (14). Se utilizó un software (QIIME2 v1.9.0) para procesar los archivos fastq de Illumina en bruto (15). Se filtraron las bases con un valor medio de calidad inferior a 20. Se utilizó el software UPARSE para clasificar secuencias con un nivel de similitud ≥ 97% en OTUs (16). Se utilizó la base de datos SILVA para realizar la asignación taxonómica de las secuencias representativas de OTU (17). El análisis de coordenadas principales (PCoA) se llevó a cabo con base en las métricas de Bray-Curtis (18). Las diferencias entre los grupos se evaluaron mediante ANOSIM utilizando el paquete vegano en R.
2.7 Análisis estadístico
Para analizar los datos de este estudio se utilizó un programa SPSS 20.0 (SPSS Inc., Chicago, IL, Estados Unidos).
Los datos (producción de leche y componentes) se analizaron con mediciones repetidas utilizando un procedimiento MIXTO, y se ajustaron con los datos del período de adaptación como factor covariable. El modelo incluyó los efectos fijos del tratamiento (CON, T-200, T-400 y T-600), el tiempo (semana 1 a 4), el tiempo de × tratamiento y la covariable. El tiempo (semana) se utilizó como medida repetida con las vacas como sujeto.
Los datos (parámetros de fermentación ruminal, índices bioquímicos séricos) se analizaron mediante la prueba de ANOVA de una vía. La diferencia significativa entre los tratamientos se evaluó mediante la prueba de Duncan. Los datos sobre las comunidades bacterianas se analizaron mediante la prueba no paramétrica (Kruskal-Wallis). La significancia se declaró a p < 0,05. Todos los resultados se expresaron como media ± error estándar.
3 Resultados
3.1 Producción de leche y composición de la leche
Como se muestra en la Tabla 1, no hubo efectos del tratamiento (p > 0,05), el tiempo (p > 0,05) y el tratamiento por tiempo (p > 0,05) para la producción de leche, el porcentaje de grasa láctea, los sólidos totales de la leche y el recuento de células somáticas. Hubo efectos del tiempo (p < 0,05), pero no del tratamiento (p > 0,05) y del tratamiento por tiempo (p > 0,05) para el porcentaje de proteína láctea, el porcentaje de lactosa láctea o la concentración de nitrógeno ureico lácteo.
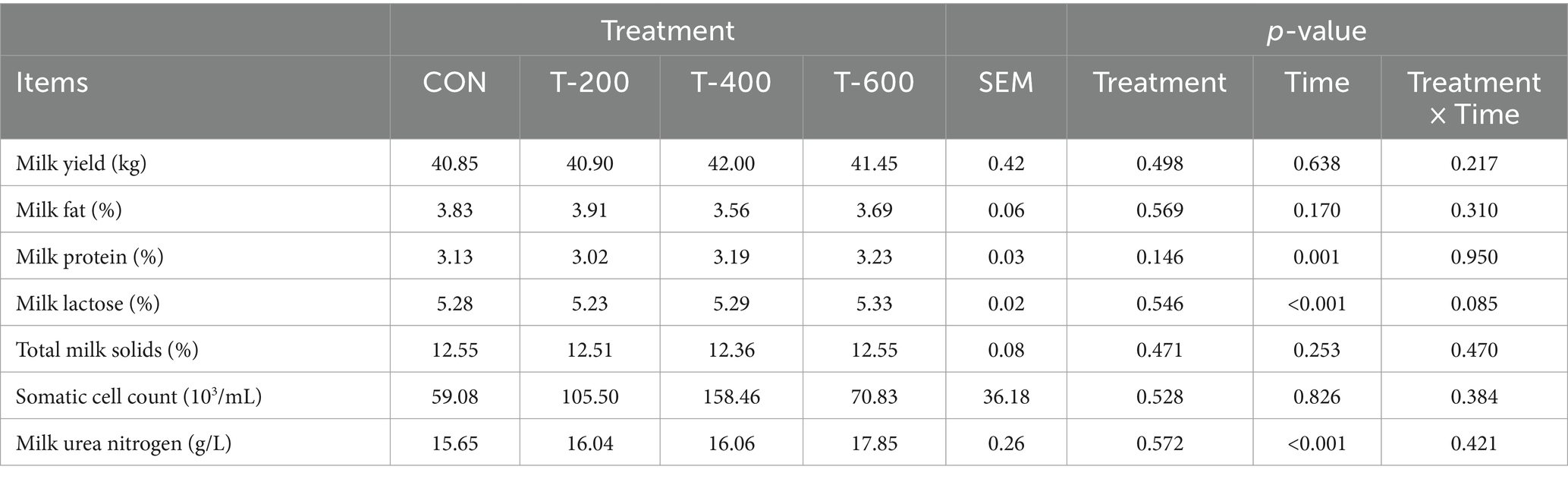
3.2 Índices bioquímicos séricos
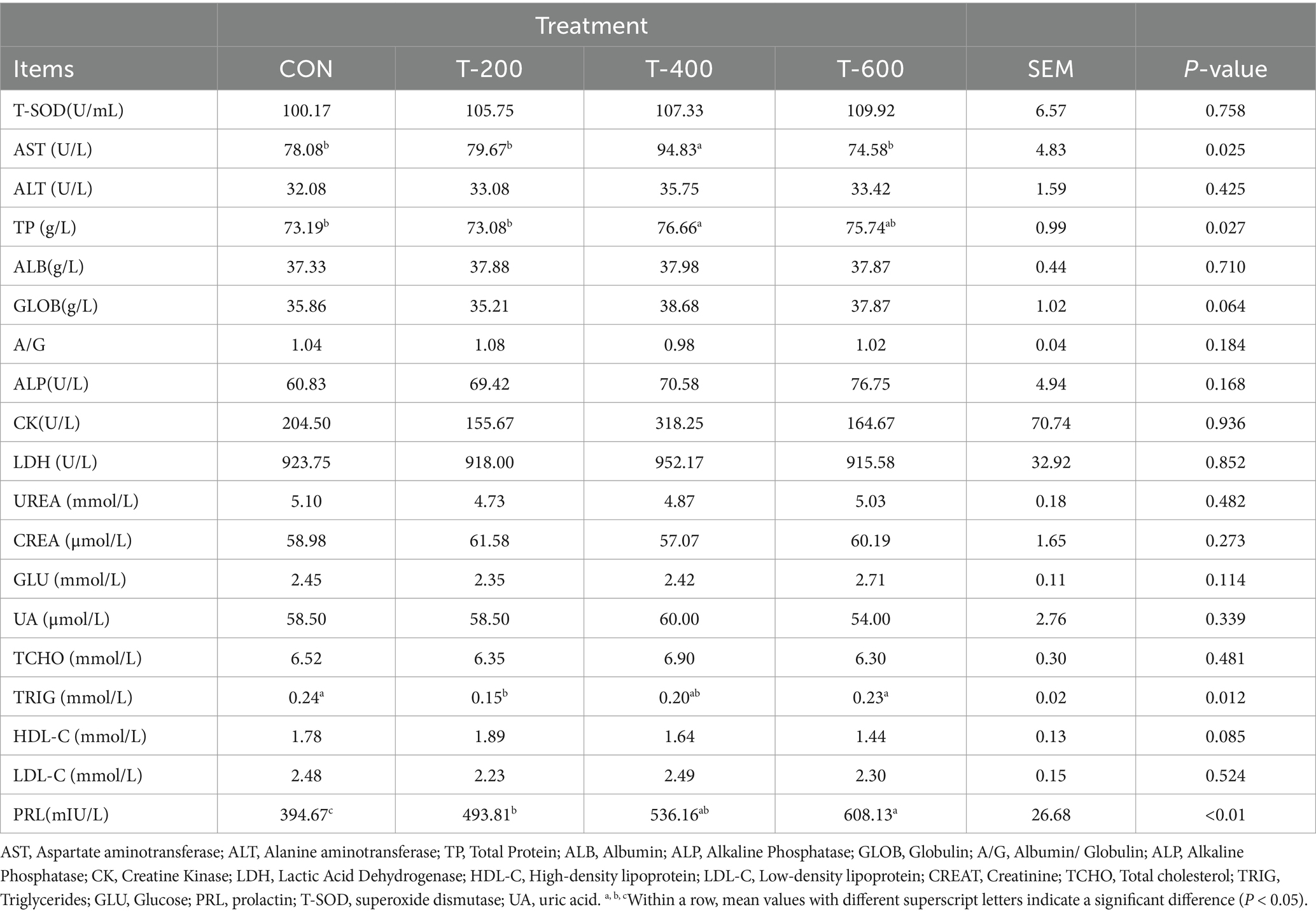
Como se muestra en la Tabla 2, hubo efectos del tratamiento para PRL (prolactina), AST (Aspartato aminotransferasa), TP (Proteína Total), TRIG (Triglicéridos) en el suero (p < 0,05). La PRL mostró un efecto dosiscórico, y aumentó con el aumento de la dosis de FSM (394,67, 493,81, 536,16 y 608,13 mUI/L, p < 0,01). La AST fue mayor en el T-400 (p < 0,05) y no difirió entre los otros 3 grupos (p > 0,05). La TP fue mayor en el T-400 que en el CON y el T-200 (p < 0,05), y el T-400 no difirió con el T-600 (p > 0,05). El TRIG fue menor en el T-200 que en el CON y el T-600 (p < 0,05), y el T-200 no difirió con el T-400 (p > 0,05). No hubo efectos del tratamiento para T-SOD (superóxido dismutasa), ALT (alanina aminotransferasa), ALB (albúmina), ALP (fosfatasa alcalina), GLOB (globulina), A/G (albúmina/globulina), ALP (fosfatasa alcalina), CK (creatina quinasa), LDH (ácido láctico deshidrogenasa), HDL-C (lipoproteína de alta densidad), LDL-C (lipoproteína de baja densidad), CREAT (creatinina), TCHO (colesterol total), GLU (glucosa) y UA (ácido úrico) en el suero (p > 0,05).
3.3 Parámetros de fermentación ruminal
Como se muestra en la Tabla 3, no hubo efectos del tratamiento para el pH ruminal, la proteína cruda microbiana, el acetato, el propionato, el butirato, el valerato, los ácidos grasos volátiles totales y la relación acetato a propionato (p > 0,05). Hubo efectos del tratamiento para la concentración de NH3-N, isobutirato e isovalerato (p < 0,05). La concentración de NH3-N fue mayor en T-400 que en los otros 3 grupos (p < 0,05). El isobutirato fue menor en T-200 y T-400 que en CON (p < 0,05), pero no difiere con el de T-600 (p > 0,05). El isovalerato fue menor en T-400 que en CON y T-600 (p < 0,05), pero no difiere con el de T-200 (p > 0,05).
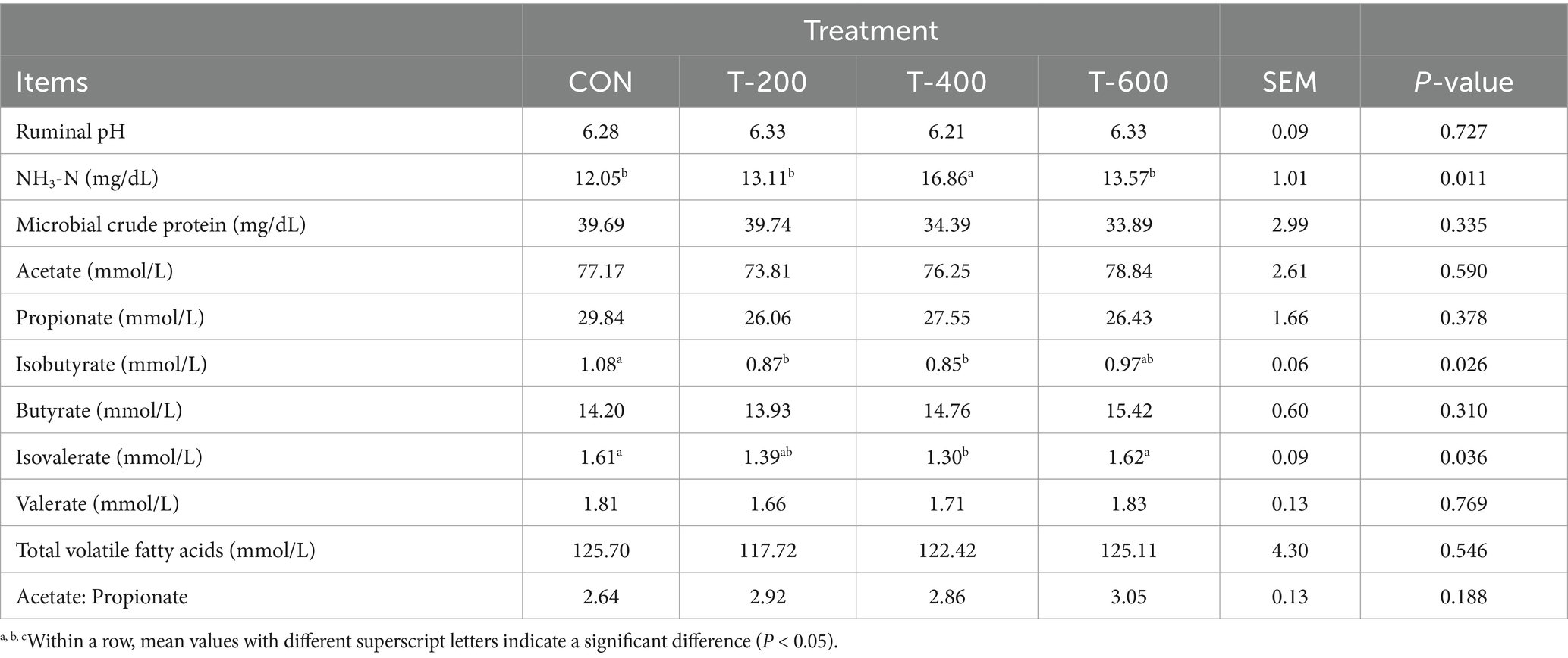
Tabla 3. Efectos de la alimentación con FSM sobre los parámetros de fermentación ruminal en vacas lactantes.
3.4 Comunidad bacteriana ruminal
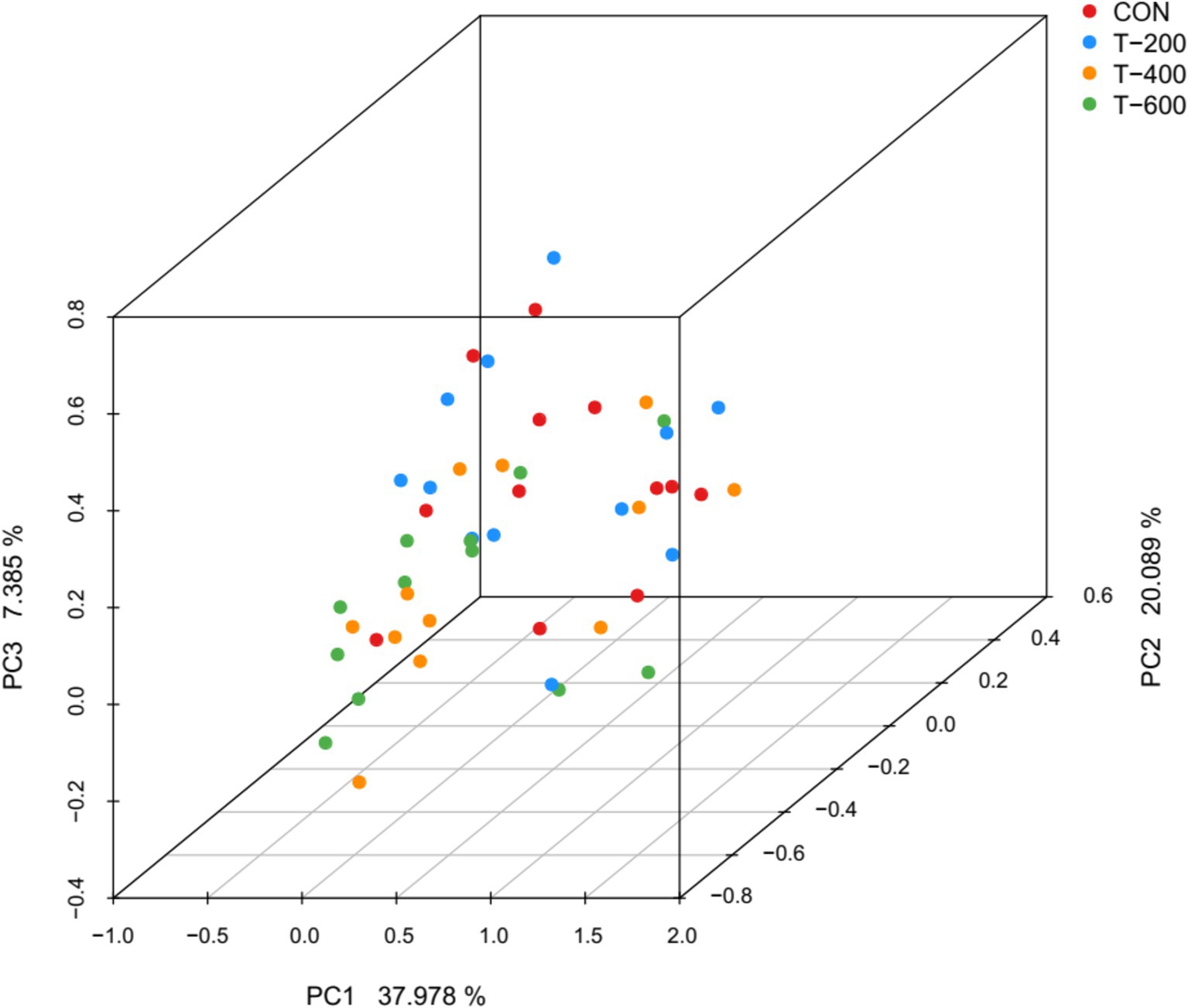
Se obtuvieron un total de 2.128.625 secuencias de alta calidad de 48 muestras, con un promedio de 44.346 secuencias para cada muestra. La curva de rarefacción tendió a aplanarse, lo que indica que la profundidad de secuenciación fue suficiente para analizar las comunidades bacterianas del rumen (Figura suplementaria S1). El PCoA basado en el algoritmo métrico de Bray Curtis mostró que el FSM alteró la estructura de la comunidad bacteriana ruminal (ANOSIM: R = 0,108, p = 0,002) (Figura 1). Se observaron diferencias significativas entre CON y T-400 (R = 0,074, p = 0,032); CON y T-600 (R = 0,109, p = 0,004); T-200 y T-400 (R = 0,076, p = 0,018); T-200 y T-600 (R = 0,103, p = 0,003). No hubo diferencias entre CON y T-200 (R = 0,042, p = 0,441); T-400 y T-600 (R = 0,039, p = 0,539).
Como se muestra en la Tabla 4, hubo efectos del tratamiento para el número de OTU (p < 0,01), el índice de Chao 1 (p = 0,043) y el índice de Shannon (p = 0,028). El número de OTUs y Chao 1 y Shannon fue menor en T-400 y T-600 que en CON (p < 0,05). No hubo efectos del tratamiento para Simpson (p = 0,051).
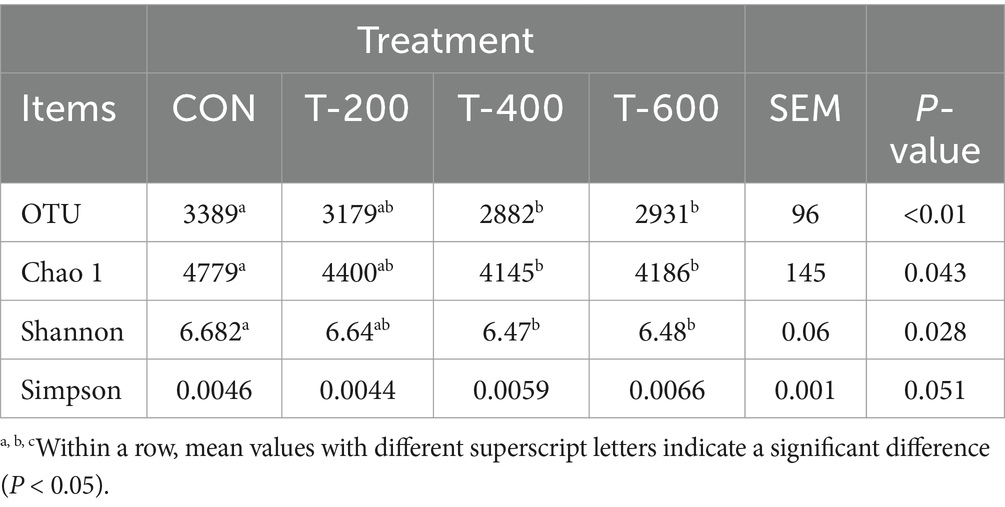
Tabla 4. Efectos de la alimentación con FSM sobre la diversidad alfa de la comunidad bacteriana ruminal en vacas lactantes.
Como se muestra en la abundancia relativa de filos en la Tabla 5, hubo efectos del tratamiento para Firmicutes (p = 0,015), Bacteroidota (p = 0,024), Actinobacterota (p < 0,01) y Patescibacteria (p = 0,012). Firmicutes fue mayor en T-400 y T-600 que en CON y T-200 (p < 0,05). Bacteroidota fue menor en T-400 y T-600 que en CON y T-200 (p < 0,05).
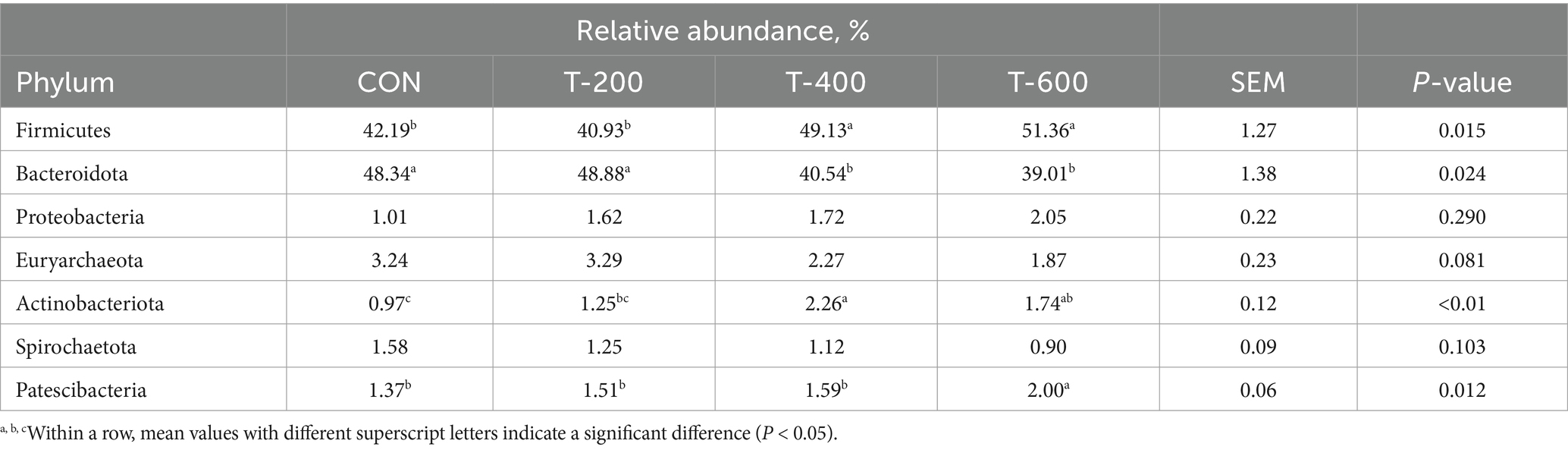
Actinobacterota fue mayor en T-400 y T-600 que en CON (p < 0,05). Patescibacteria fue mayor en T-600 que en los otros 3 grupos (p < 0,05).
Como se muestra en la abundancia relativa de los géneros en la Tabla 6, hubo efectos del tratamiento para Muribaculaceae_norank (p < 0.01), Christensenellaceae R-7 (p = 0.011), Lactococcus (p < 0.01), Candidatus Saccharimonas (p < 0.01) y Olsenella (p < 0.01). Muribaculaceae_norank fue menor en T-400 y T-600 que en CON (p < 0,05). El grupo R-7 de Christensenellaceae fue mayor en T-600 que en CON y T-200 (p < 0,05). Lactococcus y Candidatus Saccharimonas fueron mayores en T-600 que en los otros 3 grupos (p < 0,05). Olsenella fue mayor en T-400 y T-600 que en CON (p < 0,05).
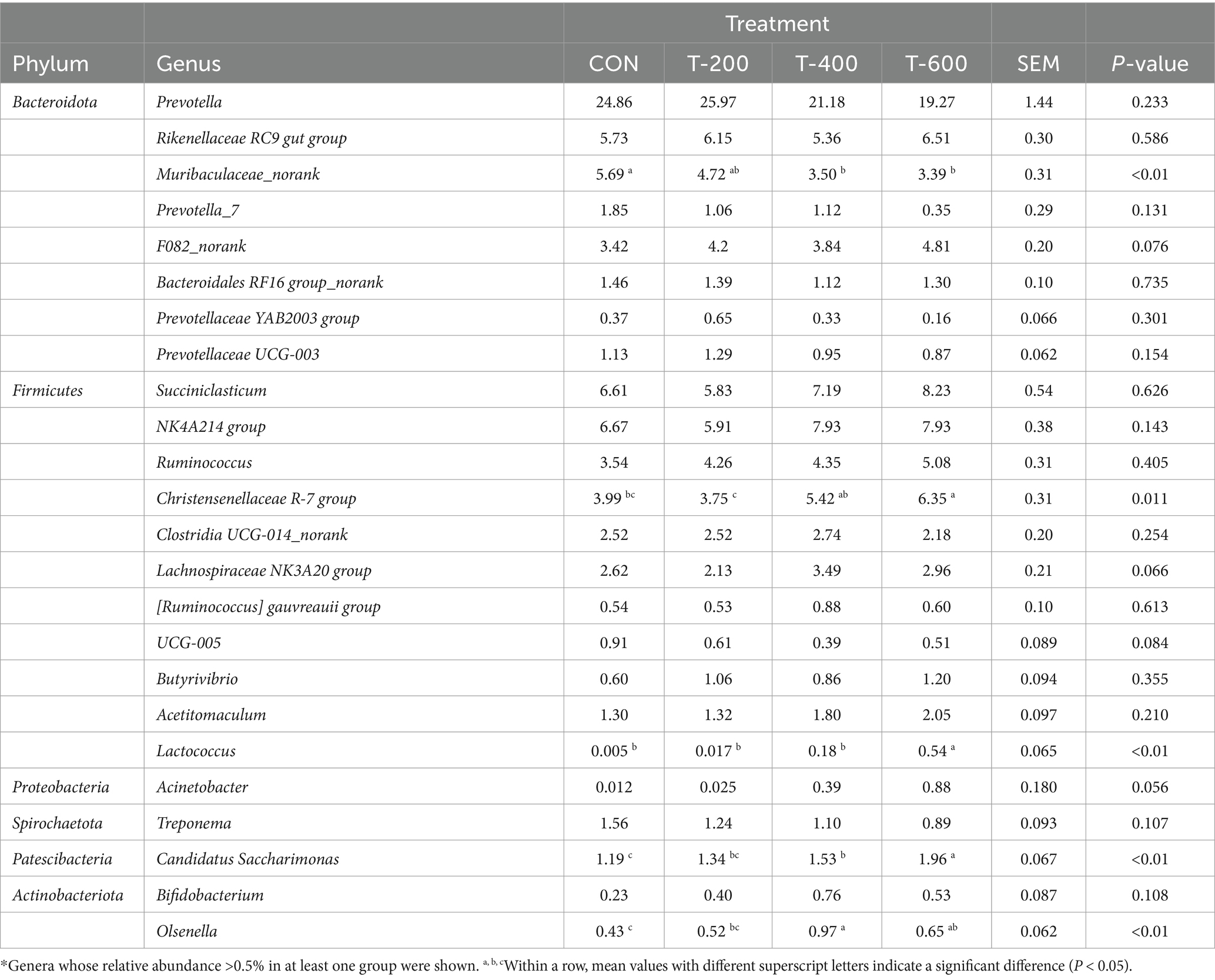
4 Discusión
FSM es una proteína de alta calidad, que contiene probióticos, enzimas digestivas, péptidos bioactivos, antioxidantes y factores antinutricionales bajos. La alimentación con FSM proporcionaría efectos positivos en las vacas lecheras, especialmente para las vacas en la etapa temprana de la lactancia, durante la cual las vacas sufren estrés múltiple. En este estudio, la alimentación con FSM no afectó la producción de leche ni la composición de la leche, solo se observó un aumento numérico en la producción de leche, el porcentaje de proteína láctea y el nitrógeno ureico de la leche. No es consistente con un estudio previo. Amin et al. (7) informaron que la alimentación con FSM aumentó la producción de proteína láctea, la producción de grasa láctea y la leche corregida con grasa, y disminuyó el recuento de células somáticas de la leche en vacas en la etapa temprana de lactancia (54 días en leche). Los diferentes resultados observados pueden atribuirse a las diferentes dosis de suplementación con FSM, al inóculo utilizado, a la composición de las dietas básicas y a las etapas de lactancia de las vacas involucradas en los diferentes estudios (1). Los estudios sobre FSM en vacas lactantes son muy pocos, por lo que se necesitan más trabajos para dilucidar el modo de acción de FSM en vacas lactantes.
Los índices bioquímicos séricos son indicadores indirectos de la salud y el estado metabólico del ganado. La alimentación con FSM causó un pequeño cambio en algunas mediciones de sangre, pero causó un gran aumento en la concentración sérica de prolactina. Estos cambios no se observaron o no se midieron en los estudios previos de Amin et al. (7) y Wang et al. (8). La prolactina es una hormona importante de la lactancia que desempeña un papel crucial en la promoción del desarrollo de la glándula mamaria, la síntesis de leche, la producción de leche y el mantenimiento de la lactancia (19). Durante la síntesis de la leche, la prolactina facilita la absorción de glucosa y aminoácidos, así como la síntesis de lactosa, grasa, caseína y lactoglobulina de la leche (20). El mecanismo que subyace al aumento de la prolactina sérica mediante la alimentación con FSM aún no está claro. En este estudio, el tratamiento con FSM solo duró 4 semanas, no se mostró el efecto de mantenimiento de la lactancia. Los estudios adicionales durarían 8 semanas o más para explorar el efecto de FSM en el mantenimiento de la lactancia en vacas lactantes. Sin embargo, el nuevo hallazgo podría proporcionar una nueva estrategia para la utilización de FSM en vacas lactantes, especialmente en la etapa temprana de lactancia.
La alimentación con FSM no afectó la concentración de acetato, propionato, butirato, ácidos grasos volátiles totales en el rumen, solo causó un pequeño cambio en la concentración de NH3-N, isobutirato e isovalerato. Dos estudios previos informaron que FSM cambió los parámetros de fermentación ruminal en vacas Holstein lactantes (7, 8). Wang et al. (8) informaron que FSM redujo la concentración total de ácidos grasos volátiles, la proporción de acetato a propionato y aumentó el porcentaje de propionato. Amin et al. (7) informaron que el FSM aumentó el pH ruminal, el porcentaje de acetato y la relación acetato a propionato. Los resultados de estos estudios no fueron consistentes. Las razones subyacentes aún no se han dilucidado.
La alimentación con FSM causó cambios en la comunidad bacteriana del rumen. Los estudios previos también informaron que la alimentación con FSM modificó las comunidades bacterianas del rumen en vacas Holstein lactantes (7, 8). Sin embargo, los cambios en estos estudios no fueron consistentes. En el estudio actual, la alimentación con FSM disminuyó el número de OTUs, los índices Chao 1 y Shannon, y aumentó la abundancia relativa de Firmicutes y disminuyó la abundancia relativa de Bacteroidota en los niveles del filo, que no se observaron en los dos estudios anteriores. Amin et al. (7) reportaron que FSM enriqueció el género de Muribaculaceae_norank, los cuales se redujeron en el presente estudio. Tanto en el presente estudio como en el de Amin et al. (7) se observó el enriquecimiento del género Christensenellaceae del grupo R-7 mediante la alimentación con FSM. El enriquecimiento de los géneros Lactococcus, Candidatus Saccharimonas y Olsenella no se observó en los dos estudios previos. Fernando et al. (21) informaron que los Firmicutes estaban más adaptados a la fermentación de fibras, mientras que los Bacteroidota eran más efectivos en la degradación del almidón. Wang et al. (22) informaron que Actinobacteriota tenía la capacidad de degradar polisacáridos. Sugirió que FSM mejoró la capacidad de fermentación de fibra en el rumen. Lagkouvardos et al. (23) informaron que los genomas de Muribaculaceae contenían un conjunto sustancial y versátil de enzimas activas en carbohidratos, lo que sugiere que los miembros de esta familia tenían la capacidad de degradar carbohidratos complejos, los autores también afirmaron que la aptitud de las especies de Muribaculaceae en la degradación de carbohidratos en la dieta probablemente explica la disminución de la presencia en los ensayos de alimentación utilizando dietas altas en calorías o enriquecidas con carbohidratos. Los lactococos son homofermentativos y se utilizan para la producción de ácido láctico L(+) a partir de la glucosa. En la industria láctea, las especies de Lactococcus se utilizan principalmente en la producción de ácido láctico a partir de lactosa, hidrólisis de caseína, lipólisis de grasa por actividades débiles de esterasa y fermentación de ácido cítrico (24). El enriquecimiento de Lactococcus puede deberse a la mejora de la degradación de la fibra o a los nutrientes proporcionados por FSM. El enriquecimiento de Candidatus Saccharimonas en el rumen se observó al alimentar con un producto de fermentación de Saccharomyces cerevisiae en vacas Holstein lactantes (25). Tong et al. (26) reportaron que la abundancia de Candidatus Saccharimonas se correlacionó positivamente con la concentración de propionato en el rumen de vacas lactantes. Ranilla et al. (27) observaron que un antioxidante (carvacrol) enriquecía a Candidatus Saccharimonas en un ensayo in vitro. Sugirió que el enriquecimiento de Candidatus Saccharimonas podría estar asociado con el antioxidante proporcionado por FSM. Los miembros de Olsenella podían utilizar almidón y glucógeno, produciendo lactato, acetato y formiato (28). Kim et al. (29) informaron que la abundancia relativa de Olsenella fue mayor en el rumen de las vacas Holstein alimentadas con una dieta alta en granos. McLoughlin et al. (30) informaron que la abundancia relativa de Olsenella en el rumen se asoció positivamente con la eficiencia alimenticia in ovejas. Elolimy et al. (31) observaron una mayor abundancia relativa de Olsenella en el intestino posterior de terneras Holstein con alta eficiencia alimenticia. Sin embargo, Ellison et al. (32) encontraron una mayor abundancia de Olsenella en el rumen de corderos de baja eficiencia alimenticia alimentados con una dieta concentrada. Sugirió que FSM aumentó la eficiencia alimenticia en el estudio actual. Desafortunadamente, la eficiencia alimenticia no se determinó en este estudio. Debe medirse en el estudio posterior.
FSM es una fuente de proteína vegetal de alta calidad que contiene más del 50% de proteína bruta. El estudio suplementó FSM directamente en las dietas sin modificar los niveles de proteínas dietéticas en los diversos grupos de tratamiento. Como resultado, los niveles de proteína bruta de la dieta en los grupos de tratamiento aumentaron aproximadamente entre un 0,7 y un 1,4% en comparación con el grupo de control. Se observó un ligero aumento en el valor numérico del porcentaje de proteína láctea y del nitrógeno ureico lácteo, pero este aumento fue estadísticamente insignificante. Los estudios adicionales ajustarían los niveles de proteínas de la dieta para que sean iguales en todos los tratamientos.
5 Conclusión
La alimentación con FSM a las vacas lactantes no afectó el rendimiento de la leche, pero aumentó los niveles séricos de prolactina, lo que ayudaría a las vacas a mantener la lactancia. Además, la alimentación con FSM solo provocó un cambio menor en los parámetros de fermentación ruminal, pero alteró en gran medida la microbiota ruminal, con el aumento de Firmicutes y la disminución de Bacteroidota en la abundancia relativa. Aunque se debe realizar más trabajo para demostrar los efectos de la FSM, estos hallazgos pueden proporcionar un enfoque para mantener el pico de lactancia en las vacas lecheras.
Declaración de disponibilidad de datos
Los conjuntos de datos presentados en este estudio se pueden encontrar en repositorios en línea. Los nombres del repositorio o repositorios y el número de acceso se pueden encontrar en el artículo/Material complementario.
Declaración ética
El estudio en animales fue aprobado por el Comité de Cuidado y Uso de Animales de la Universidad Agrícola de Nanjing. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
JZ: Curación de datos, Investigación, Visualización, Redacción – borrador original. FG: Redacción – borrador original, Investigación. SH: Investigación, Escritura – borrador original. YM: Escritura – revisión y edición, Investigación. SW: Escritura – revisión y edición, conceptualización. WJ: Redacción – borrador original, Redacción – revisión y edición. SM: Conceptualización, Obtención de fondos, Administración de proyectos, Supervisión, Redacción, revisión y edición.
Financiación
El (los) autor(es) declara(n) que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue apoyada por la Fundación Nacional de Ciencias Naturales de China (32072755).
Conflicto de intereses
YM fue empleado por Jiangsu Jiahui Biotechnology Co., Ltd. y SW fue empleado por Shanghai Menon Animal Nutrition Technology Co., Ltd.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1498639/full#supplementary-material
Referencias
1. Lambo, MT, Ma, H, Zhang, H, Song, P, Mao, H, Cui, G, et al. Mecanismo de acción, beneficios y brecha de investigación en la utilización de harina de soja fermentada como fuente de proteínas de alta calidad para el ganado y las aves de corral. Anim Nutr. (2024) 16:130–46. doi: 10.1016/j.aninu.2023.10.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Yuan, L, Chang, J, Yin, Q, Lu, M, Di, Y, Wang, P, et al. La harina de soja fermentada mejora el rendimiento del crecimiento, la digestibilidad de los nutrientes y la flora microbiana de los lechones. Anim Nutr. (2017) 3:19–24. doi: 10.1016/j.aninu.2016.11.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Jazi, V, Mohebodini, H, Ashayerizadeh, A, Shabani, A y Barekatain, R. La harina de soja fermentada mejora la infección por Salmonella Typhimurium en pollos de engorde jóvenes. Ciencia avícola. (2019) 98:5648–60. doi: 10.3382/ps/pez338
4. Feizi, LK, Zad, SS, Jalali, SAH, Rafiee, H, Jazi, MB, Sadeghi, K, et al. La harina de soja fermentada afecta la fermentación ruminal y la abundancia de especies bacterianas seleccionadas en terneros Holstein: un análisis multinivel. Sci Rep. (2020) 10:12062. doi: 10.1038/s41598-020-68778-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Kim, MH, Yun, CH, Lee, CH y Ha, JK. Los efectos de la harina de soja fermentada sobre los parámetros inmunofisiológicos y relacionados con el estrés en terneros Holstein después del destete. J Lácteos Sci. (2012) 95:5203–12. doi: 10.3168/jds.2012-5317
6. Rezazadeh, F, Kowsar, R, Rafiee, H y Riasi, A. La fermentación de la harina de soja mejora el rendimiento del crecimiento y la respuesta inmunitaria de los terneros Holstein destetados bruscamente durante el clima frío. Anim Feed Sci Technol. (2019) 254:114206. doi: 10.1016/j.anifeedsci.2019.114206
7. Amin, AB, Zhang, L, Zhang, J y Mao, S. La harina de soja fermentada modificó el microbioma ruminal para mejorar el rendimiento de los componentes de la leche en vacas Holstein. Aplicación Microbiol Biotechnol. (2022) 106:7627–42. doi: 10.1007/s00253-022-12240-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Wang, Z, Yu, Y, Li, X, Xiao, H, Zhang, P, Shen, W, et al. Sustituto de harina de soja fermentada en la dieta de vacas lecheras Holstein lactantes: fermentación ruminal modulada y microflora ruminal. Microbiol frontal. (2021) 12:625857. doi: 10.3389/fmicb.2021.625857
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Weatherburn, MW . Reacción de fenol-hipoclorito para la determinación de amoníaco. Química Anal. (1967) 39:971–4. doi: 10.1021/ac60252a045
10. Makkar, H, Sharma, O, Dawra, R y Negi, S. Determinación simple de la proteína microbiana en el licor ruminal. J Lácteos Sci. (1982) 65:2170–3. DOI: 10.3168/JDS. S0022-0302(82)82477-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Wang, Y, Xu, L, Liu, J, Zhu, W y Mao, S. Una dieta rica en cereales modificó dinámicamente la composición de la microbiota asociada a la mucosa e indujo lesiones de la mucosa en el colon de las ovejas. Microbiol frontal. (2017) 8:2080. doi: 10.3389/fmicb.2017.02080
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Mao, S, Zhang, R, Wang, D, y Zhu, W. La diversidad de la comunidad bacteriana fecal y su relación con la concentración de ácidos grasos volátiles en las heces durante la acidosis ruminal subaguda en vacas lecheras. BMC Vet Res. (2012) 8:237–13. doi: 10.1186/1746-6148-8-237
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Behrendt, L, Larkum, AW, Trampe, E, Norman, A, Sørensen, SJ y Kühl, M. Diversidad microbiana de las comunidades de biopelículas en microniches asociadas con la ascidia didemnídida Lissoclinum patella. ISME J. (2012) 6:1222–37. doi: 10.1038/ismej.2011.181
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Magoč, T, y Salzberg, SL. FLASH: ajuste rápido de la longitud de lecturas cortas para mejorar los ensamblajes del genoma. Bioinformática. (2011) 27:2957–63. doi: 10.1093/bioinformatics/btr507
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Campbell, BJ, Polson, SW, Hanson, TE, Mack, MC y Schuur, EA. El efecto de la deposición de nutrientes en las comunidades bacterianas en el suelo de la tundra ártica. Environ Microbiol. (2010) 12:1842–54. doi: 10.1111/j.1462-2920.2010.02189.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Edgar, RC . Búsqueda y agrupación en clústeres de órdenes de magnitud más rápido que BLAST. Bioinformática. (2010) 26:2460–1. doi: 10.1093/bioinformática/BTQ461
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Quast, C, Pruesse, E, Yilmaz, P, Gerken, J, Schweer, T, Yarza, P, et al. El proyecto SILVA ribosomal RNA gene database: mejora del procesamiento de datos y herramientas basadas en la web. Ácidos nucleicos Res. (2013) 41:D590-6. doi: 10.1093/nar/gks1219
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Mitter, EK, de Freitas, JR y Germida, JJ. Microbioma bacteriano de la raíz de las plantas que crecen en las cubiertas de recuperación de arenas bituminosas. Microbiol frontal. (2017) 8:849. doi: 10.3389/fmicb.2017.00849
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Lacasse, P, Ollier, S, Lollivier, V, y Boutinaud, M. Nuevos conocimientos sobre la importancia de la prolactina en los rumiantes lecheros. J Lácteos Sci. (2016) 99:864–74. doi: 10.3168/jds.2015-10035
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Kim, YJ . Papel fundamental de la prolactina y otras hormonas en la lactogénesis y la composición nutricional de la leche materna. Clin Exp Pediatr. (2020) 63:312–3. doi: 10.3345/cep.2020.00311
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Fernando, SC, Purvis, H, Najar, F, Sukharnikov, L, Krehbiel, C, Nagaraja, T, et al. Dinámica de la población microbiana ruminal durante la adaptación a una dieta alta en granos. Appl Environ Microbiol. (2010) 76:7482–90. doi: 10.1128/AEM.00388-10
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Wang, C, Dong, D, Wang, H, Müller, K, Qin, Y, Wang, H, et al. Análisis metagenómico de consorcios microbianos enriquecidos a partir de compost: nuevos conocimientos sobre el papel de las actinobacterias en la descomposición de la lignocelulosa. Biotecnol Biocombustibles. (2016) 9:1–17. doi: 10.1186/s13068-016-0440-2
23. Lagkouvardos, I, Lesker, TR, Hitch, TCA, Gálvez, EJC, Smit, N, Neuhaus, K, et al. El estudio de la secuencia y el cultivo de Muribaculaceae revela nuevas especies, preferencia de huéspedes y potencial funcional de esta familia aún no descrita. Microbioma. (2019) 7:28. doi: 10.1186/s40168-019-0637-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Kim, W . El género Lactococcus En: WH Holzapfel y BJB Wood, editores. Bacterias lácticas: biodiversidad y taxonomía John Wiley & Sons, Ltd. (2014) doi: 10.1002/9781118655252
25. Jiang, Q, Sherlock, DN, Elolimy, AA, Yoon, I, y Loor, JJ. La alimentación con un producto de fermentación de Saccharomyces cerevisiae durante un desafío de barrera intestinal en vacas Holstein lactantes afecta a la microbiota ruminal y al metaboloma. J Lácteos Sci. (2024) 107:4476–94. doi: 10.3168/jds.2023-24147
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Tong, J, Zhang, H, Yang, D, Zhang, Y, Xiong, B y Jiang, L. Análisis secuencial de Illumina de la microbiota ruminal en vacas lecheras lactantes de alto y bajo rendimiento. PLoS Uno. (2018) 13:e0198225. doi: 10.1371/journal.pone.0198225
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Ranilla, MJ, Andrés, S, Gini, C, Biscarini, F, Saro, C, Martín, A, et al. Efectos del aceite esencial de Thymbra capitata sobre los productos finales de fermentación in vitro y las comunidades bacterianas ruminales. Sci Rep. (2023) 13:4153. doi: 10.1038/s41598-023-31370-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Göker, M, Held, B, Lucas, S, Nolan, M, Yasawong, M, Glavina Del Rio, T, et al. Secuencia completa del genoma de la cepa tipo Olsenella uli (VPI D76D-27C). Stand Genomic Sci. (2010) 3:76–84. doi: 10.4056/sigs.1082860
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Kim, YH, Nagata, R, Ohkubo, A, Ohtani, N, Kushibiki, S, Ichijo, T, et al. Cambios en el pH ruminal y reticular y en las comunidades bacterianas en ganado Holstein alimentado con una dieta alta en granos. BMC Vet Res. (2018) 14:310. doi: 10.1186/s12917-018-1637-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. McLoughlin, S, Spillane, C, Claffey, N, Smith, PE, O’Rourke, T, Diskin, MG, et al. La composición del microbioma ruminal está alterada en las ovejas con divergencia en la eficiencia alimenticia. Microbiol frontal. (2020) 11:1981. doi: 10.3389/fmicb.2020.01981
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Elolimy, A, Alharthi, A, Zeineldin, M, Parys, C, y Loor, JJ. La divergencia de la ingesta de alimento residual durante el período previo al destete se asocia con perfiles únicos de microbioma y metaboloma del intestino posterior en terneras neonatales de vaquillonas Holstein. J Anim Sci Biotechnol. (2020) 11:13. doi: 10.1186/s40104-019-0406-x
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: harina de soja fermentada, vacas, rendimiento de la leche, índices bioquímicos séricos, fermentación ruminal, comunidad bacteriana
Cita: Zhang J, Guan F, Huang S, Ma Y, Wen S, Jin W y Mao S (2024) La harina de soja fermentada modificó la microbiota ruminal y aumentó el nivel de prolactina sérica en vacas Holstein lactantes. Frente. Vet. Sci. 11:1498639. doi: 10.3389/fvets.2024.1498639
Editado por:
Shengguo Zhao, Academia China de Ciencias Agrícolas, China
Revisado por:
Xiaodan Huang, Universidad de Lanzhou, China
Yanfeng Xue, Universidad Agrícola de Anhui, China
Derechos de autor © 2024 Zhang, Guan, Huang, Ma, Wen, Jin y Mao. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Wei Jin, jinwei@njau.edu.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía