
La hibridación promueve el rendimiento del crecimiento al alterar la microbiota ruminal y los metabolitos en las ovejas

La hibridación promueve el rendimiento del crecimiento al alterar la microbiota ruminal y los metabolitos en las ovejas
 Rui Zhang1,2,3
Rui Zhang1,2,3 Liwa Zhang1,2,3Xuejiao An1,2,3Jianye Li1,2,3Chune Niu1,2,3Jinxia Zhang4Zhiguang Geng4Tao Xu1,2,3,5Bohui Yang1,2,3Zhenfei Xu1,2,3*
Liwa Zhang1,2,3Xuejiao An1,2,3Jianye Li1,2,3Chune Niu1,2,3Jinxia Zhang4Zhiguang Geng4Tao Xu1,2,3,5Bohui Yang1,2,3Zhenfei Xu1,2,3* Yaojing Yue1,2,3*
Yaojing Yue1,2,3*- 1Instituto de Ganadería y Ciencias Farmacéuticas de Lanzhou, Academia China de Ciencias Agrícolas, Lanzhou, China
- número arábigoCentro de Investigación de Tecnología de Ingeniería de Cría de Ovejas de la Academia China de Ciencias Agrícolas, Lanzhou, China
- 3Laboratorio Clave de Genética y Cría Animal en la Meseta Tibetana, Ministerio de Agricultura y Asuntos Rurales, Lanzhou, China
- 4Instituto de Investigación de Ciencias Agrícolas de Qingyang, Qingyang, China
- 5Centro de Servicios Integrales Agrícolas y Rurales del municipio de Gengwan, Qingyang, China
La hibridación puede mejorar sustancialmente el rendimiento del crecimiento. Este estudio utilizó la metagenómica y la secuenciación del metaboloma para examinar si la microbiota ruminal y sus metabolitos contribuían a este fenómeno. Se seleccionaron 48 corderos machos ♂de aproximadamente 3 meses de edad Hu × ♀Hu (HH, n = 16), ♂Poll Dorset × ♀Hu (DH, n = 16) y ♂Southdown × ♀Hu (SH, n = 16) con peso corporal similar. Las ovejas fueron alimentadas individualmente bajo las mismas condiciones nutricionales y de manejo durante 95 días. Una vez finalizado el ensayo, se sacrificaron siete ovejas cercanas al peso medio por grupo para recoger muestras de tejido y contenido ruminal con el fin de medir los parámetros epiteliales del rumen, los patrones de fermentación, la microbiota y los perfiles de metabolitos. Los valores de peso corporal final (FBW), ganancia diaria promedio (ADG) e ingesta de materia seca (DMI) en los grupos DH y SH fueron significativamente más altos y la relación alimentación-ganancia (F/G) significativamente menor que el valor en el grupo HH; además, la altura de la papila en el grupo DH fue mayor que en el grupo HH. Las concentraciones de acetato, propionato y ácidos grasos volátiles totales (AGV) en el grupo DH fueron mayores que las del grupo HH, mientras que la concentración de NH3-N disminuyó en los grupos DH y SH. El análisis metagenómico reveló que varias especies de Prevotella y Fibrobacter eran significativamente más abundantes en el grupo DH, lo que contribuye a una mayor capacidad para degradar la celulosa de la dieta y enriquecer sus funciones en las enzimas involucradas en la descomposición de los carbohidratos. La bacteria Bacteroidaceae fue mayor en el grupo SH, lo que indica una mayor capacidad para digerir la fibra dietética. El análisis metabolómico reveló que las concentraciones de metabolitos del rumen (principalmente lisofosfatidiletanolaminas [LPE]) fueron mayores en el grupo DH, y el análisis de metabolitos relacionado con el microbioma indicó que Treponema bryantii y Fibrobacter succinogenes se correlacionaron positivamente con los LPE. Además, encontramos que el sulfóxido de metionina y el ácido N-metil-4-aminobutírico fueron metabolitos característicos en los grupos DH y SH, respectivamente, y están relacionados con el estrés oxidativo, lo que indica que la adaptabilidad ambiental de las ovejas cruzadas debe mejorarse aún más. Estos hallazgos profundizan sustancialmente la comprensión general de cómo la hibridación promueve el rendimiento del crecimiento desde la perspectiva de la microbiota ruminal, esto es vital para el cultivo de nuevas especies y la formulación de estrategias de nutrición de precisión para las ovejas.
1 Introducción
La hibridación es un método importante de cría de ovejas que puede producir heterosis, lo que da como resultado una nueva raza que es más fuerte o se comporta mejor que sus padres (1). La hibridación binaria en el cultivo de nuevas variedades de ovejas ovinas se ha reportado con frecuencia en China (2). A pesar de que el cruzamiento tripartito se utiliza habitualmente en la producción porcina y en el desarrollo de nuevas razas (3, 4), la investigación pertinente se encuentra todavía en las primeras fases de selección de especies adecuadas en la industria ovina (5). En comparación con el primero, el segundo podría utilizar eficazmente la heterosis de tres individuos híbridos, produciendo un efecto significativamente más fuerte. Hu es una raza ovina autóctona con estro perenne, alta fecundidad, alta lactancia, buenos rasgos de crianza y gran adaptabilidad; Por lo tanto, a menudo se utilizan como ovejas cruzadas por ovejas ovinas, sin embargo, su tasa de producción de carne y porcentaje de aderezo son insatisfactorios (6, 7). Poll Dorset es originario de Australia y Nueva Zelanda, tiene las características de crecimiento rápido, madurez temprana y estro perenne, y se considera el principal progenitor de la hibridación en la industria china de cría de ovejas (8). Southdown es considerada la mejor raza por su calidad de carne en el Reino Unido debido a su estructura de carne ideal, madurez temprana, fácil cría, buena calidad de canal y porcentaje de aderezo > 55% (9, 10). Southdown y Poll Dorset son pequeñas especies de ovejas que crecen rápidamente en las primeras etapas y son adecuadas para producir corderos gordos de alta calidad, lo que se alinea con la demanda actual del mercado de cordero en China. Por lo tanto, intentamos cultivar una nueva raza de ovejas utilizando Poll Dorset y Southdown como sementales, cruzándolas con ovejas Hu y monitoreando los rasgos de producción de la generación F1.
El rendimiento del crecimiento es el principal rasgo de selección para la cría de nuevas variedades de ovejas. La microbiota ruminal está asociada con el crecimiento y la salud de los animales (11). El rumen es colonizado por bacterias, protozoos, arqueas, hongos y virus, que degradan fibras vegetales complejas y polisacáridos para producir ácidos grasos volátiles (AGV), proteínas microbianas y vitaminas, que a su vez proporcionan nutrientes para satisfacer los requisitos del huésped para el mantenimiento y el crecimiento (12, 13). Aunque la dieta desempeña un papel importante en la configuración de la microbiota gastrointestinal, pruebas recientes han revelado que la genética del huésped es un factor importante en la determinación de la composición de la microbiota intestinal en el ganado bovino (13-15). Sin embargo, la investigación sobre los efectos de la genética del huésped en la composición de la microbiota ruminal de ovejas híbridas es limitada.
Por lo tanto, en este estudio, planteamos la hipótesis de que la composición y la función de la microbiota ruminal en las ovejas se ven afectadas por la genética del huésped, lo que a su vez afecta al rendimiento del crecimiento. Evaluamos la microbiota ruminal y los perfiles de metabolitos de diferentes ovejas híbridas criadas en el mismo entorno de granja, con el objetivo de dilucidar los mecanismos moleculares subyacentes desde la perspectiva de la microbiota ruminal y, por lo tanto, proporcionar una base teórica y práctica para mejorar el rendimiento del crecimiento de las ovejas híbridas.
2 Materiales y métodos
2.1 Animales, diseño experimental y recolección de muestras
Este estudio se llevó a cabo en la Compañía de Cría de Ovejas Cordero de Qinghuan en el condado de Huanxian, Gansu, China, desde octubre de 2022 hasta enero de 2023. El diseño experimental, los procedimientos y los métodos fueron aprobados por el Comité de Ética y Administración Animal del Instituto de Ganadería y Ciencias Farmacéuticas de Lanzhou de la Academia China de Ciencias Agrícolas con el número de permiso 2022-018 y siguieron las normas chinas para el uso y cuidado de los animales. Se seleccionaron cuarenta y ocho corderos machos ♂de aproximadamente 3 meses de edad Hu × ♀Hu (HH, n = 16), ♂Poll Dorset × ♀Hu (DH, n = 16) y ♂Southdown × ♀Hu (SH, n = 16) y se criaron en un solo corral bajo las mismas condiciones nutricionales y de manejo durante 95 días (incluido un período de preprueba de 15 días). Se eligieron ovejas Hu de 216 corderos machos de aproximadamente 3 meses de edad con un peso corporal promedio de 22,98 ± 6,37 kg, y se seleccionaron ovejas de la generación F1 de peso corporal similar. Los sementales Poll Dorset y Southdown (de 31 meses de edad) generados a partir del trasplante de embriones y los sementales Hu (de 24 meses de edad) se cruzaron con ovejas Hu (paridad uno o dos) por sincronización de celo e inseminación artificial. Se confirmó que las crías (una, en el caso de los gemelos) provenían de diferentes padres. Todas las ovejas fueron alimentadas con la misma proporción de mezcla total compuesta por heno de avena (6.08% MS), Leymus chinensis (13.04% MS), ensilaje de maíz (13.53% MS), maíz (32.78% MS), megalac (6.08% MS) y mezclas de granos (28.49% MS) dos veces al día a las 08:00 y 15:00 (Tabla Suplementaria S1). Durante el período experimental, todos los animales tuvieron acceso a alimento y agua ad libitum. La ingesta de alimento se registró diariamente en función del alimento ofrecido y los rechazos para calcular la ingesta promedio de materia seca (MSI). El peso corporal se registró cada 20 días utilizando una báscula digital para ganado (TCS Electronic Platform Scale, Rongcheng, China) para calcular la ganancia diaria promedio (ADG) y la relación alimento-ganancia (F/G).
Al final del experimento, se seleccionaron siete ovejas cercanas al peso promedio por grupo y se ayunaron durante 12 h antes de la cosecha. La cosecha se basaba en procedimientos comerciales estándar, restringidos, desangüados, despellejados y eviscerados individualmente. Las muestras de tejido ruminal de aproximadamente 2 cm × 2 cm se separaron cuidadosamente del saco dorsal izquierdo para fijarlas en una solución de paraformaldehído al 4% para la tinción de hematoxilina-eosina (H&E). A continuación, se recogieron dos porciones de 5 mL de contenido ruminal de cada animal, se transfirieron a un tubo estéril, se congelaron inmediatamente con nitrógeno líquido y se almacenaron a -80 °C para la extracción de ADN y metabolitos. Se tomaron muestras de otros 15 mL de contenido ruminal y se almacenaron en un recipiente esterilizado a -20 °C para evaluar los parámetros de fermentación. El contenido ruminal de todos los animales se recolectó del saco dorsal izquierdo como una mezcla de componentes líquidos y sólidos. Todas las muestras se recogieron dentro de los 30 minutos posteriores al sacrificio.
2.2 Parámetros epiteliales del rumen
Los parámetros epiteliales del rumen, incluyendo la altura de la papila, el ancho de la papila y el grosor muscular, se obtuvieron mediante la técnica de tinción H&E (16). Los tejidos ruminales fijos se deshidrataron en alcohol, se aclararon en xileno y se incrustaron en parafina. A continuación, las muestras de concreción de enfriamiento se seccionaron a 5 μm de espesor y se montaron en portaobjetos de vidrio. Las secciones incrustadas en parafina se desparafinaron con xileno, se pasaron a través de una serie de etanol graduado para eliminar el xileno, se enjuagaron con agua destilada y se tiñeron con H&E. Posteriormente, la parafina se selló con goma neutra después de la deshidratación y se examinó inmediatamente. Por último, se adquirieron dos portaobjetos digitales de cada animal utilizando un dispositivo Slide Viewer (3D HISTECH Ltd.) con doble aumento; a continuación, se eligieron cinco papilas (excluyendo las dañadas) al azar de cada portaobjetos digital para medir la altura de la papila utilizando el software ImageJ (National Institutes of Health, Bethesda, MD, Estados Unidos). Dentro de las papilas medidas para la altura, se identificaron tres regiones diferentes (apical, media y basal) para medir el ancho de la papila. Cada portaobjetos digital se dividió en cinco partes iguales para medir el grosor muscular.
2.3 Parámetros de fermentación ruminal
El pH del rumen se midió inmediatamente después del sacrificio de las ovejas utilizando un medidor de acidez portátil PHS-10 de Ark Technology (Chengdu, China). La concentración de NH3-N se determinó por colorimetría. En resumen, 2 g de contenido de rumen se pesaron con precisión y se mezclaron con agua desionizada en una proporción de 1:5. La solución mezclada se agitó durante 1 h a 105 r/min y se centrifugó a 12.000 rpm a 4 °C durante 20 min. A continuación, el sobrenadante se pasó a través de una membrana de filtro de fibra polimérica de 0,45 μm y se almacenó para su posterior análisis. Posteriormente, se midió la concentración de NH3-N utilizando un kit (Biosino Biotechnology Co. Ltd., Pekín, China) de acuerdo con las instrucciones del fabricante y un lector de microplacas (DR-200BS; Hiwell-Diatek, Wuxi, China). Las concentraciones de AGV se midieron como se describió anteriormente (16). Brevemente, el contenido ruminal se centrifugó a 5.400 rpm durante 10 min. Posteriormente, mezclamos uniformemente 1 mL del sobrenadante resultante y una solución de metafosfato al 25% de 0,2 mL que contenía ácido 2-etilbutírico como patrón interno en un nuevo tubo de centrífuga. A continuación, este tubo de reacción se sumergió en un baño de hielo (30 min) y se centrifugó a 10.000 rpm durante otros 10 min. El sobrenadante se pasó a través de una membrana filtrante de fase orgánica de 0,22 μm y se almacenó en frascos de 2 mL para su posterior análisis. Un cromatógrafo de gases (GC 7890A; Agilent Technologies, Santa Clara, CA, Estados Unidos) equipado con una columna capilar AT-FFAP (50 m × 0,32 mm × 0,25 μm; Agilent Technologies) para determinar las concentraciones de AGV. La temperatura de la columna se mantuvo a 60 °C durante 1 min, se elevó 5 °C/min a 115 °C y se incrementó 15 °C/min a 180 °C. En particular, las temperaturas del detector y del inyector fueron de 260 °C y 250 °C, respectivamente.
2.4 Secuenciación del metagenoma y análisis bioinformático
El ADN genómico total se extrajo de la muestra de contenido ruminal utilizando el E.Z.N.A. Soil DNA Kit (Omega Biotek, Norcross, GA, Estados Unidos). La concentración y pureza del ADN se determinaron utilizando el minifluorómetro TBS-380 (Promega, Madison, WI, Estados Unidos) y el espectrofotómetro NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, Estados Unidos), respectivamente. El ADN se fragmentó a aproximadamente 400 pb para la construcción de bibliotecas de extremos emparejados utilizando un Covaris M220 (Gene Company Limited, Hong Kong, China). La secuenciación de las bibliotecas del metagenoma se realizó utilizando una plataforma Illumina NovaSeq 6,000 (Illumina Inc., San Diego, CA, Estados Unidos).
El control de calidad de las lecturas de la secuencia metagenómica se realizó utilizando el software Fastp (versión 0.20.0) para recortar los extremos 3′ y 5′ de las lecturas, cortar las bases de baja calidad y eliminar las lecturas cortas y los registros «N» (17). A continuación, las lecturas filtradas por calidad se alinearon con el genoma de referencia de Ovis aries utilizando BWA v 0.7.1 para filtrar el ADN del huésped (18). El resto de las lecturas se ensamblaron utilizando Multiple Megahit (versión 1.1.2) (19), y se seleccionaron contigs con longitudes ≥300 pb como resultado final del ensamblaje para la predicción de marcos de lectura abiertos (ORF) utilizando MetaGene v 0.3.38 (20). Los contigs no redundantes se identificaron mediante CD-HIT con una identidad de secuencia del 95% y una cobertura del 90% (21). Las lecturas de secuenciación originales se mapearon a los genes predichos para estimar su abundancia utilizando SOAPAligner v. 2.21 (22). Posteriormente, el catálogo de genes no redundantes se alineó con la base de datos de secuencias de proteínas no redundantes del NCBI utilizando BLASTP (versión 2.2.28+) para obtener anotaciones taxonómicas y abundancias de especies (23). Los perfiles taxonómicos se realizaron a nivel de dominio, filo, género y especie, y se calcularon abundancias relativas. Se realizó el PCoA basado en matrices de disimilitud de Bray-Curtis a nivel de dominio. Se analizaron bacterias con una abundancia relativa >0,01% y arqueas con una abundancia relativa >0,001%. Por último, se anotó una búsqueda BLAST (versión 2.2.28+) con un criterio de corte de optimización de 1e-5 en la base de datos de la Enciclopedia de Genes y Genomas de Kioto (KEGG) (24). La anotación de la enzima activa en carbohidratos (CAZyme) se llevó a cabo utilizando hmmscan1 contra la base de datos CAZy versión 5.02 con un valor de corte e de 1e-5.
2.5 Secuenciación del metaboloma y análisis bioinformático
La extracción de metabolitos de muestras ruminales se basó en procedimientos publicados previamente (16). El metaboloma del contenido ruminal se analizó mediante cromatografía líquida de ultra resolución (UPLC; Paquete de cuñas UFLC Shimadzu CBM30A; Shimadzu, Kyoto, Japón) y espectrometría de masas en tándem (MS/MS, QTRAP® 6.500+, SCIEX, Framingham, MA, Estados Unidos) (25). Después de obtener los datos de cromatografía líquida-espectrometría de masas de las muestras, los picos de cromatografía iónica extraída de todos los metabolitos se integraron utilizando el software MultiQuant (Applied Biosystems, Foster City, CA, Estados Unidos). Los picos cromatográficos de los metabolitos en diferentes muestras se corrigieron mediante integración (26). Las concentraciones relativas de metabolitos ruminales se examinaron mediante cambio de pliegue (FC) (FC ≥ 2 o FC ≤ 0,5) y variable de importancia en proyección (VIP) (VIP ≥ 1) para identificar los diferentes metabolitos. Los metabolitos identificados se anotaron utilizando la base de datos de compuestos KEGG (27).
2.6 Análisis estadístico
Todos los datos se comprobaron para determinar la normalidad y los valores atípicos utilizando el programa SPSS versión 26.0 (IBM Corp., Armonk, NY, Estados Unidos) antes de realizar cualquier análisis estadístico. Se utilizó ANOVA de una vía y prueba post-hoc de LSD para analizar los datos de rendimiento de crecimiento, así como los parámetros epiteliales y de fermentación del rumen. Para los análisis de correlación se utilizó la prueba de correlación de Spearman. La significación estadística se estableció en p < 0,05. GraphPad Prism versión 8.5 (GraphPad Software, La Jolla, CA, Estados Unidos) dibujó los mapas estadísticos.
3 Resultados
3.1 La hibridación promovió el rendimiento del crecimiento en ovejas Hu
En la Tabla 1 se presenta un resumen de los datos de crecimiento y rendimiento. El peso corporal inicial (MCI) no difirió significativamente entre los tres grupos (p = 0,156). Los grupos DH y SH presentaron mayor GMD que el grupo HH (p = 0,008), pero no difirieron entre sí. El peso corporal final (FBW) fue mayor en los grupos DH y SH que en el grupo HH (p < 0,001). El IMD también fue mayor en los grupos DH y SH, resultando en un valor F/G menor que en el grupo HH (p = 0,013).
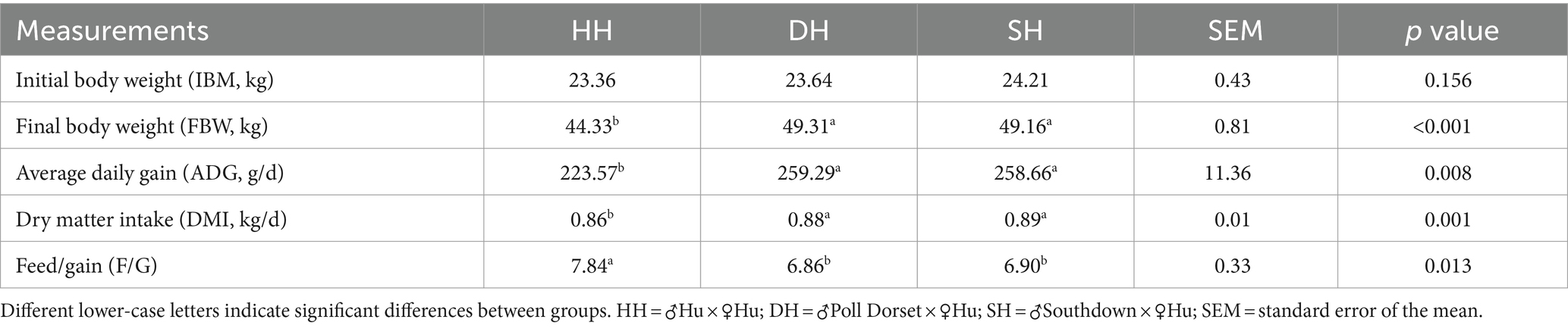
Tabla 1. La hibridación promovió el rendimiento del crecimiento en ovejas Hu.
3.2 La hibridación aumentó el desarrollo epitelial ruminal en ovejas Hu
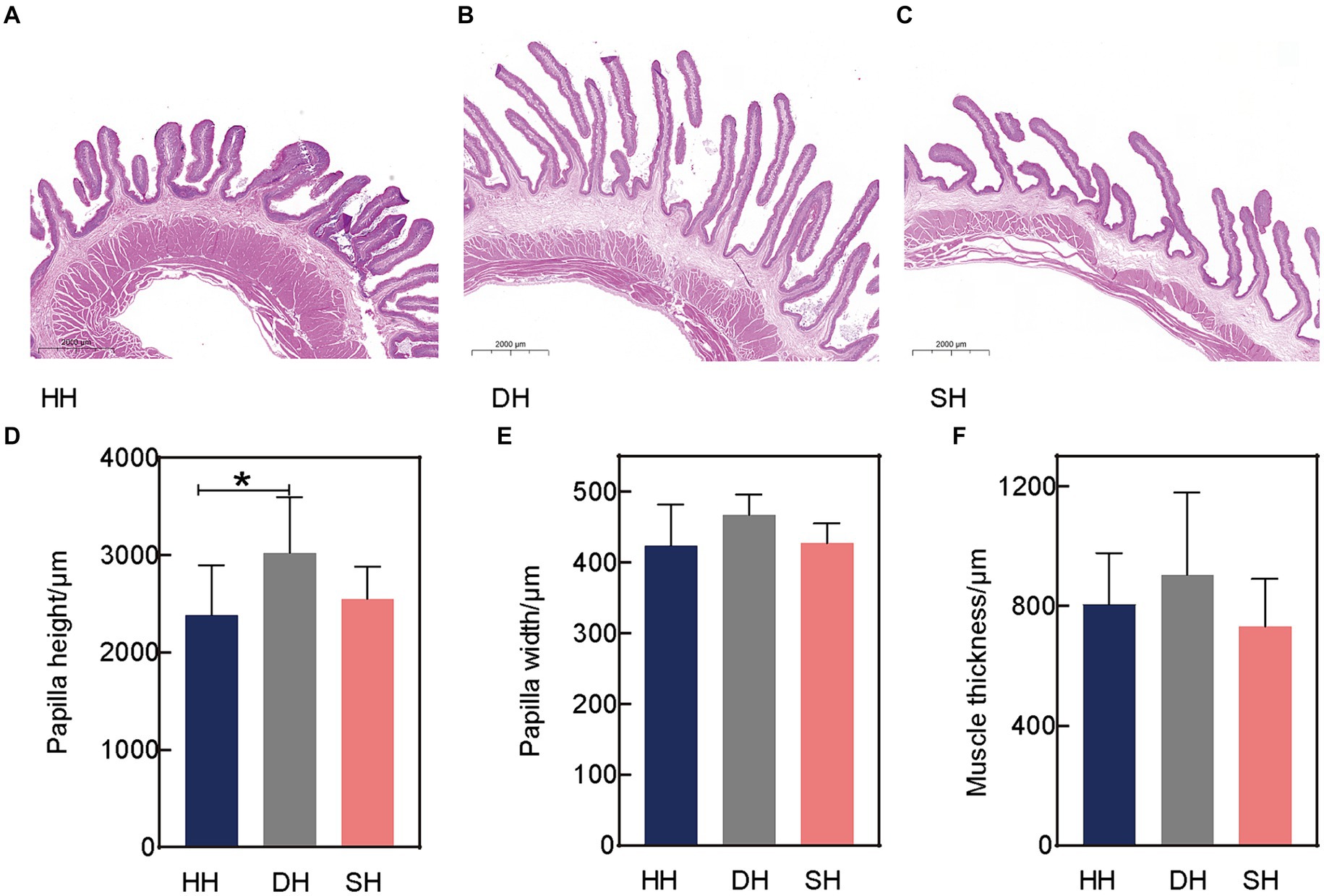
Se midieron tres índices de desarrollo epitelial ruminal, incluyendo la altura de la papila, el ancho de la papila y el grosor muscular, utilizando tinción de H&E (Figuras 1A-C). La hibridación no tuvo efecto sobre el ancho de la papila ruminal ni sobre el grosor muscular (p > 0,128; Figuras 1E,F). La mayor diferencia se observó en la altura de la papila, el grupo DH presentó una mayor altura que el grupo HH (p = 0,046; Figura 1D).
3.3 La hibridación alteró los parámetros de fermentación ruminal en ovejas Hu
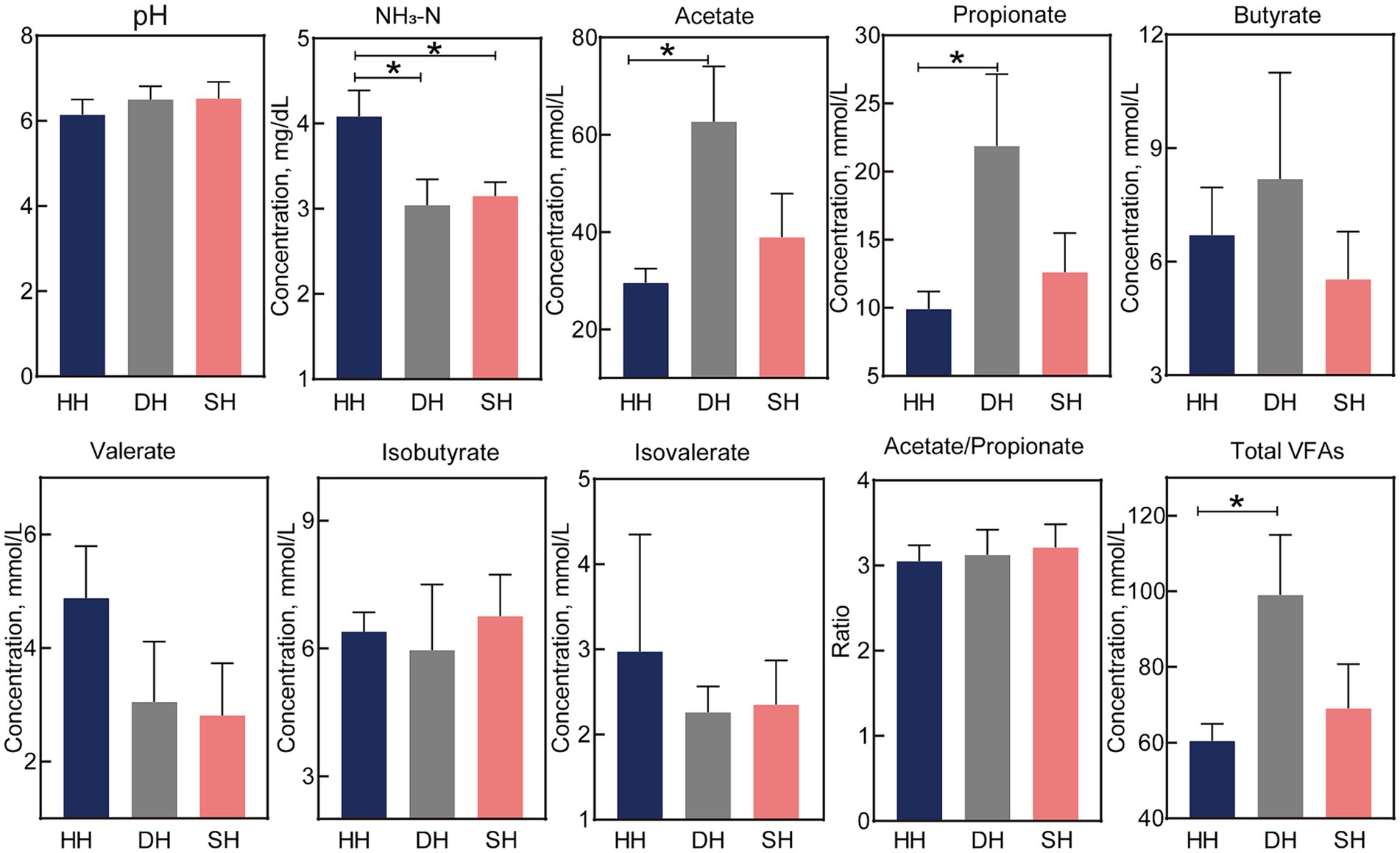
La concentración de NH3-N fue significativamente menor en los grupos SH y DH que en el grupo HH (p = 0,023; Figura 2). Las concentraciones de acetato y propionato en el grupo DH fueron significativamente mayores que las del grupo HH (p < 0,041), lo que resultó en AGV totales más altos (p = 0,048). El valor de pH, así como las concentraciones de butirato, valerato, isobutirato, isovalerato y acetato/propionato permanecieron sin cambios.
3.4 Perfil del metagenoma ruminal
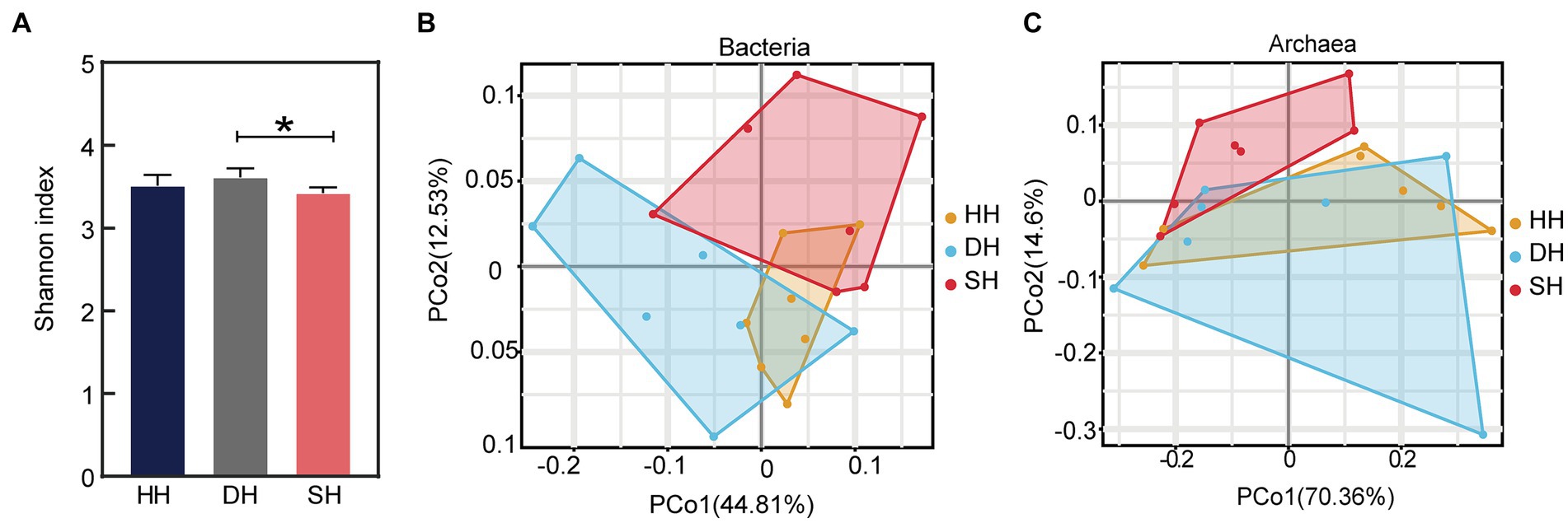
La secuenciación del metagenoma generó un total de 1.443.800.424 lecturas brutas, con una media de 68.752.401 ± 5.953.935 lecturas (media ± DE) por muestra. Después del control de calidad y la eliminación de los genes del huésped, se retuvieron un total de 1.284.320.034 lecturas, es decir, 61.158.097 ± 5.264.516 por muestra. Después del montaje, se generaron un total de 19.962.258 contigs (con una longitud media de N50 de 825 ± 60), con 950.584 ± 91.196 por muestra (Tabla suplementaria S2). Los resultados indicaron que los datos de secuenciación eran creíbles y podían utilizarse para investigaciones bioinformáticas posteriores. El análisis de α-diversidad mediante la prueba de Kruskal-Wallis reveló que el índice de Shannon aumentó significativamente en el grupo DH en comparación con el grupo SH (p = 0,016; Figura 3A), mientras que los índices Ace y Simpson no difirieron entre los tres grupos (Tabla Suplementaria S3). El metagenoma ruminal estaba compuesto por un 97,18% de bacterias (591.631.456 secuencias), un 1,60% de arqueas (9.758.146 secuencias), un 0,43% de eucariotas (2.601.458 secuencias) y un 0,78% de virus (4.731.066 secuencias). El análisis de coordenadas principales (PCoA) a nivel de dominio reveló una separación entre los grupos HH y DH y los grupos HH y SH basados en bacterias (Figura 3B), así como entre los grupos SH y HH y los grupos SH y DH basados en arqueas (Figura 3C), no se observó ninguna separación basada en eucariotas o virus (Figura Suplementaria S1). Por lo tanto, el análisis comparativo de los taxones microbianos del rumen entre los tres grupos se centró únicamente en las bacterias y las arqueas.
3.5 Perfiles composicionales del microbioma ruminal y diferencias taxonómicas
Los filos bacterianos dominantes en el rumen fueron Bacteroidetes (HH: 44,39%; DH: 44,87%; SH: 45,53%) y Firmicutes (HH: 42,72%; DH: 40,33%; SH: 43,24%), seguido de Spirochaetes (HH: 2,81%; DH: 4,64%; SH: 2,55%); sin embargo, no se encontraron diferencias significativas entre los tres grupos (p > 0,05; Figura complementaria S2A). Se identificaron un total de 5.413 géneros. Entre ellos, los bacteroidales no clasificados (HH: 19,53%; DH: 14,50%; SH: 20,21%), Prevotella (HH: 14,31%; DH: 19,93%; SH: 14,87%) y Clostridia no clasificada (HH: 8,33%; DH: 7,03%; SH: 8,83%) fueron las bacterias predominantes en las muestras ruminales (Figura suplementaria S2B). Se encontraron un total de 20.943 especies, entre las cuales las bacterias dominantes fueron la bacteria Bacteroidales (HH: 19,35%; DH: 14,42%; SH: 20,00%), Prevotella sp. (HH: 12,37%; DH: 16,29%; SH: 12,71%) y la bacteria Clostridia (HH: 8,33%; DH: 7,03%; SH: 8,83%) (Figura suplementaria S2C).
El análisis diferencial reveló un total de 28 especies bacterianas significativamente diferentes entre los grupos DH y HH. Entre ellos, 18 exhibieron abundancias significativamente mayores en el grupo DH. Las abundancias relativas de Treponema bryantii, Fibrobacter sp., Prevotella sp. tc2-28, Spirochaetia bacterium y Alloprevotella sp. fueron las cinco principales que aumentaron significativamente. Además, las abundancias relativas de Prevotella copri, Prevotella sp. BP1-148, Prevotella sp. E13-3, Fibrobacter succinogenes, Fibrobacter sp. UWB4, Fibrobacter sp. UWB2, Fibrobacter sp. UWR2, Fibrobacter sp. UWOV1 y Fibrobacter sp. UWB1 también se incrementó significativamente en el grupo DH. Diez bacterias se enriquecieron significativamente en el grupo HH, principalmente la bacteria Bacteroidales, la bacteria Anaerolineaceae, Sarcina sp., Sarcina sp. DSM 11001 y Flexilinea sp. (Figura 4A). En comparación con los niveles en el grupo HH, solo una bacteria (bacteria Bacteroidaceae) se enriqueció significativamente en el grupo SH, mientras que tres (Succiniclasticum ruminis, Sarcina sp. DSM 11001 y Ruminococcus sp. M6 (2020)) fueron significativamente más abundantes en el grupo HH (Figura 4B). Cuando se compararon con los niveles en el grupo DH, 19 abundancias bacterianas fueron significativamente diferentes, incluyendo seis (Bacteroidales bacterium, Pyramidobacter sp., Clostridiaceae bacterium, Bacteroidales bacterium WCE2004, Bacteroidetes bacterium y p Firmicutes no clasificado) con mayores abundancias en el grupo SH y 13 (Treponema sp., Acidaminococcaceae bacterium, Fibrobacter sp., Treponema bryantii, Schwartzia sp., Prevotella sp.tc2-28, Bacteria Spirochaetia, Schwartzia succinivorans, Fibrobacter succinogenes, Fibrobacter sp. UWB2, Fibrobacter sp. UWB3, Eubacterium ruminantium y Fibrobacter sp. UWB12) con mayores abundancias en el grupo DH (Figura suplementaria S3A).
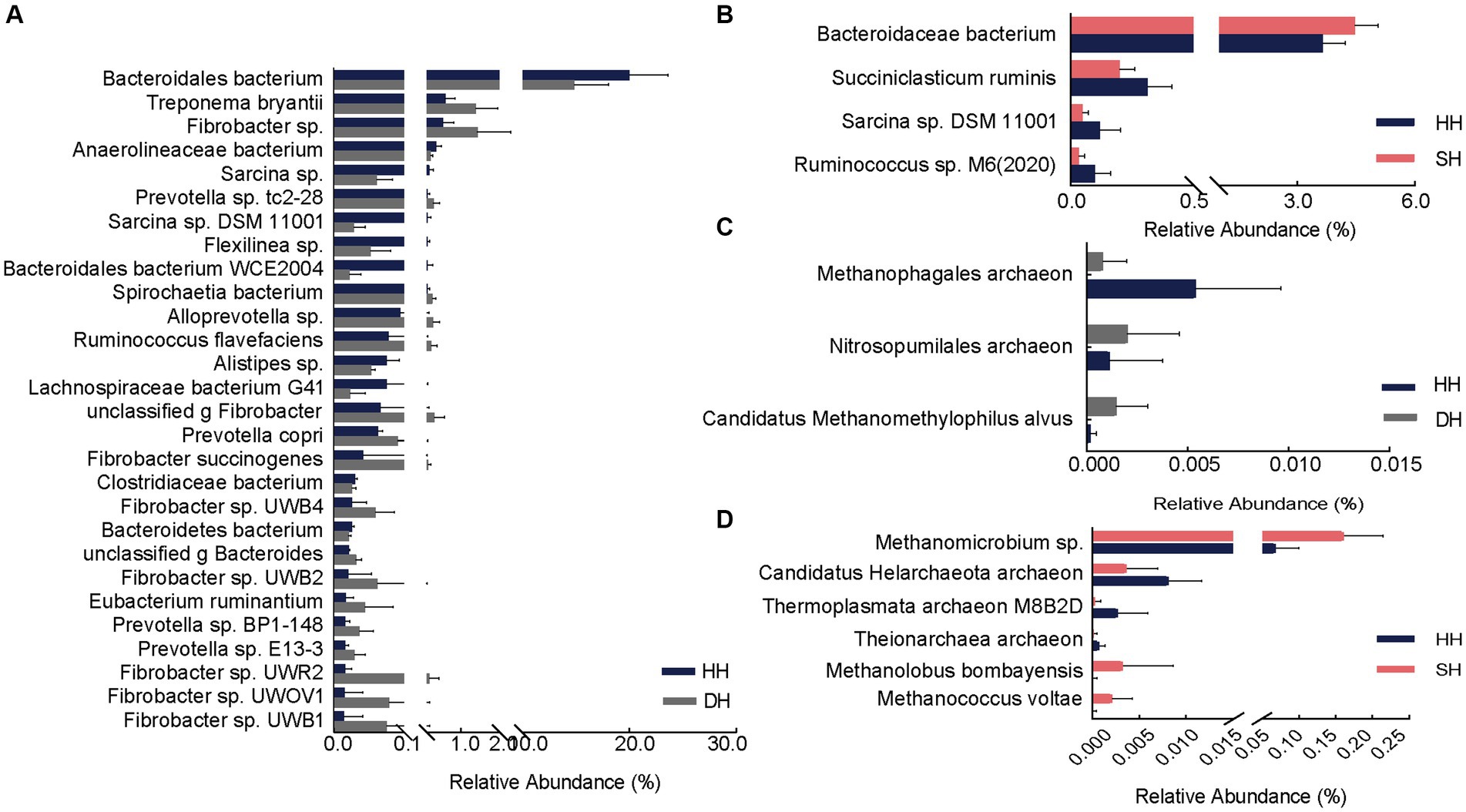
Figura 4. Especies ruminales diferenciales de bacterias y arqueas. (A) Especies bacterianas significativamente diferentes entre los grupos HH y DH. (B) Especies bacterianas significativamente diferentes entre los grupos HH y SH. (C) Especies de arqueas significativamente diferentes entre los grupos HH y DH. (D) Especies de arqueas significativamente diferentes entre los grupos HH y SH. n = 7 individuos/grupo.
A nivel de especies, la abundancia de dos especies de arqueas (Nitrosopumilales archaeon y Candidatus Methanomethylophilus alvus) se enriqueció significativamente y una (Methanophagales archaeon) disminuyó significativamente en el grupo DH en comparación con los niveles en el grupo HH (Figura 4C). La abundancia de tres arqueas (Methanomicrobium sp., Methanolobus bombayensis y Methanococcus voltae) se enriqueció significativamente y tres (Candidatus Helarchaeota archaeon, Thermoplasmata archaeon M8B2D y Theionarchaea archaeon) disminuyeron en el grupo SH en comparación con los niveles en el grupo HH (Figura 4D). Cuando se compararon con los niveles en el grupo DH, las abundancias de Methanocorpusculum sp. y Candidatus Altiarchaeales archaeon WOR SM1 86-2 fueron significativamente mayores, mientras que las abundancias de Candidatus Methanomethylophilaceae archaeon, Candidatus Methanomethylophilus sp., Methanomicrobium sp., Methanobacterium sp. ERen5, g Methanobacterium sin clasificar y Methanolobus bombayensis fueron significativamente más bajos que los del grupo SH (Figura suplementaria S3B).
3.6 Perfiles funcionales y diferencias del microbioma ruminal
Las funciones del microbioma ruminal se determinaron utilizando perfiles KEGG y genes que codifican CAZymes. Para los perfiles KEGG, se anotaron un total de seis vías en el primer nivel entre los tres grupos, incluyendo el «metabolismo» (HH: 49,78%; DH: 50,16%; SH: 50,85%), «procesamiento de la información genética» (HH: 16,78%; DH: 16,71%; SH: 17,05%), «procesamiento de información ambiental» (HH: 12,92%; DH: 12,98%; SH: 12,77%), «procesos celulares» (HH: 9,71%; DH: 9,68%; SH: 9,42%), «enfermedades humanas» (HH: 6,67%; DH: 6,47%; SH: 6,22%) y «sistemas organísicos» (HH: 4,15%; DH: 4,00%; SH: 3,70%). En el segundo nivel, obtuvimos un solo perfil, «metabolismo de cofactores y vitaminas», que se enriqueció significativamente en el grupo DH en comparación con el nivel en los grupos HH (Figura suplementaria S4A). Nueve perfiles KEGG se enriquecieron significativamente en el grupo SH en comparación con los niveles en el grupo HH, entre ellos, «replicación y reparación», «metabolismo de nucleótidos» y «biosíntesis de otros metabolitos secundarios» fueron los más dominantes (Figura suplementaria S4B). Siete perfiles KEGG fueron significativamente diferentes entre los grupos DH y SH. «biosíntesis y metabolismo de glicanos» y «metabolismo de nucleótidos» fueron los más enriquecidos en el grupo SH, mientras que la «transducción de señales» se enriqueció en el grupo DH (Figura suplementaria S4C). Al comparar las vías KEGG identificadas en el tercer nivel, la «formación de biopelículas: pseudomonas aeruginosa», «quimiotaxis bacteriana» y «metabolismo de la riboflavina» se enriquecieron significativamente en el grupo DH, mientras que las vías de «lisosoma» y «biosíntesis de varios metabolitos secundarios de plantas» disminuyeron significativamente en comparación con los niveles en el grupo HH (Figura 5A). Se identificaron un total de 21 vías diferenciales en los grupos HH y SH. Entre ellos, 13 se enriquecieron significativamente en el grupo SH, incluyendo principalmente «biosíntesis de aminoácidos», «recombinación homóloga», «biosíntesis de peptidoglicanos», «metabolismo de nucleótidos», «metabolismo de pirimidina», «reparación de desajustes», «replicación del ADN», «metabolismo de alanina, aspartato y glutamato», «vías de fijación de carbono en procariotas», «resistencia a betalactámicos», «biosíntesis de lisina», «metabolismo de glicina, serina y treonina» y «un grupo de carbono por folato»; mientras que ocho se enriquecieron significativamente en el grupo HH (Figura 5B). Se identificaron veinte vías diferenciales en los grupos DH y SH. De ellos, cinco se enriquecieron significativamente en el grupo DH y 15 en el grupo SH. «Metabolismo del carbono», «biosíntesis de aminoacil-ARNt», «metabolismo de nucleótidos», «metabolismo de pirimidina» y «degradación de otros glicanos» fueron las cinco vías principales más enriquecidas en el grupo SH (Figura suplementaria S5A).
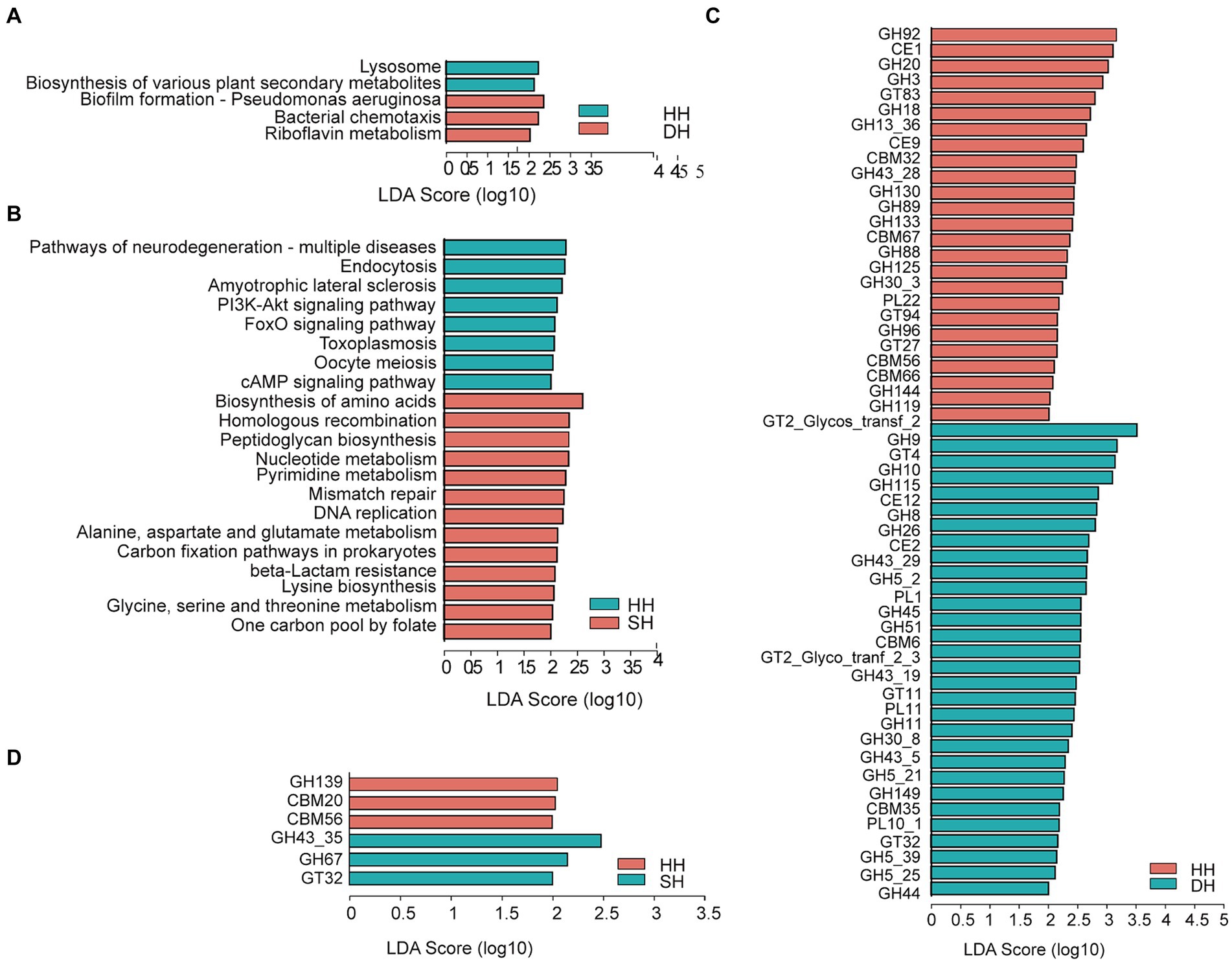
Para los perfiles de CAZyme, se identificaron 55 genes expresados diferencialmente que codifican CAZymes entre los grupos HH y DH a nivel familiar (Figura 5C). Entre ellos, 23 implicados en la descomposición de los hidratos de carbono (incluyendo celulosa, hemicelulosa, almidón, proteínas y lignina) se enriquecieron en el grupo DH (2 esterasas de carbohidratos [CE], 18 glucósidos hidrolasas [GH] y 3 polisacáridos liasas [PL]), y 18 en el grupo HH (2 CE, 15 GH y 1 PL). Cinco glicosiltransferasas (GTs) implicadas en la síntesis de carbohidratos se enriquecieron en el grupo DH, mientras que tres se enriquecieron en el grupo HH. En la degradación de los hidratos de carbono complejos como parte de los módulos de unión a los hidratos de carbono (CBM) participaron varios CAZymes no catalíticos, incluidos dos enriquecidos en el grupo DH y cuatro en el grupo HH. Se observaron un total de seis genes expresados diferencialmente que codifican CAZymes entre los grupos HH y SH (Figura 5D). Entre estos, 2 GH y 1 GT se enriquecieron en el grupo SH, mientras que 1 GH y 2 CBM se enriquecieron en el grupo HH. Se obtuvieron 41 genes diferenciales que codifican CAZymes entre los grupos DH y SH (Figura suplementaria S5B). Entre los implicados en la descomposición de los hidratos de carbono, 21 estaban enriquecidos en el grupo SH (2 CEs, 15 GHs, 2 GLs y 2 PLs), mientras que 12 estaban enriquecidos en el grupo DH (2 CEs, 8 GH y 2 PLs). Además, se enriquecieron 3 GT en el grupo DH.
3.7 Análisis de correlación entre fenotipo y microbioma
El análisis del coeficiente de correlación de Spearman identificó correlaciones entre el rendimiento del crecimiento, los parámetros epiteliales del rumen, los parámetros de fermentación y microbiomas significativamente diferentes (|r| > 0,5; p < 0,05). Cuando comparamos los grupos de HH y DH, la abundancia relativa de Alloprevotella sp. se correlacionó positivamente con FBW, así como la abundancia relativa de Fibrobacter sp. UWR2 y Fibrobacter sp. se correlacionaron positivamente con DMI, y la abundancia relativa de Alistipes sp. se correlacionó negativamente con FBW, ADG y DMI (Figura 6A). Cuando comparamos los grupos HH y SH, la abundancia relativa de la bacteria Bacteroidaceae se correlacionó positivamente con FBW y DMI; sin embargo, las abundancias relativas de Ruminococcus sp. M6(2020) y Theionarchaea archaeon se correlacionaron negativamente con la concentración de NH3-N (Figura 6B). Cuando comparamos los grupos DH y SH, encontramos que Pyramidobacter sp. se correlacionó negativamente con las concentraciones de acetato, propionato y butirato, las abundancias relativas de Candidatus Methanomethylophilus sp. y Candidatus Methanomethylophilaceae archaeon se relacionaron negativamente con las concentraciones de acetato, propionato y AGV totales (Figura suplementaria S6).
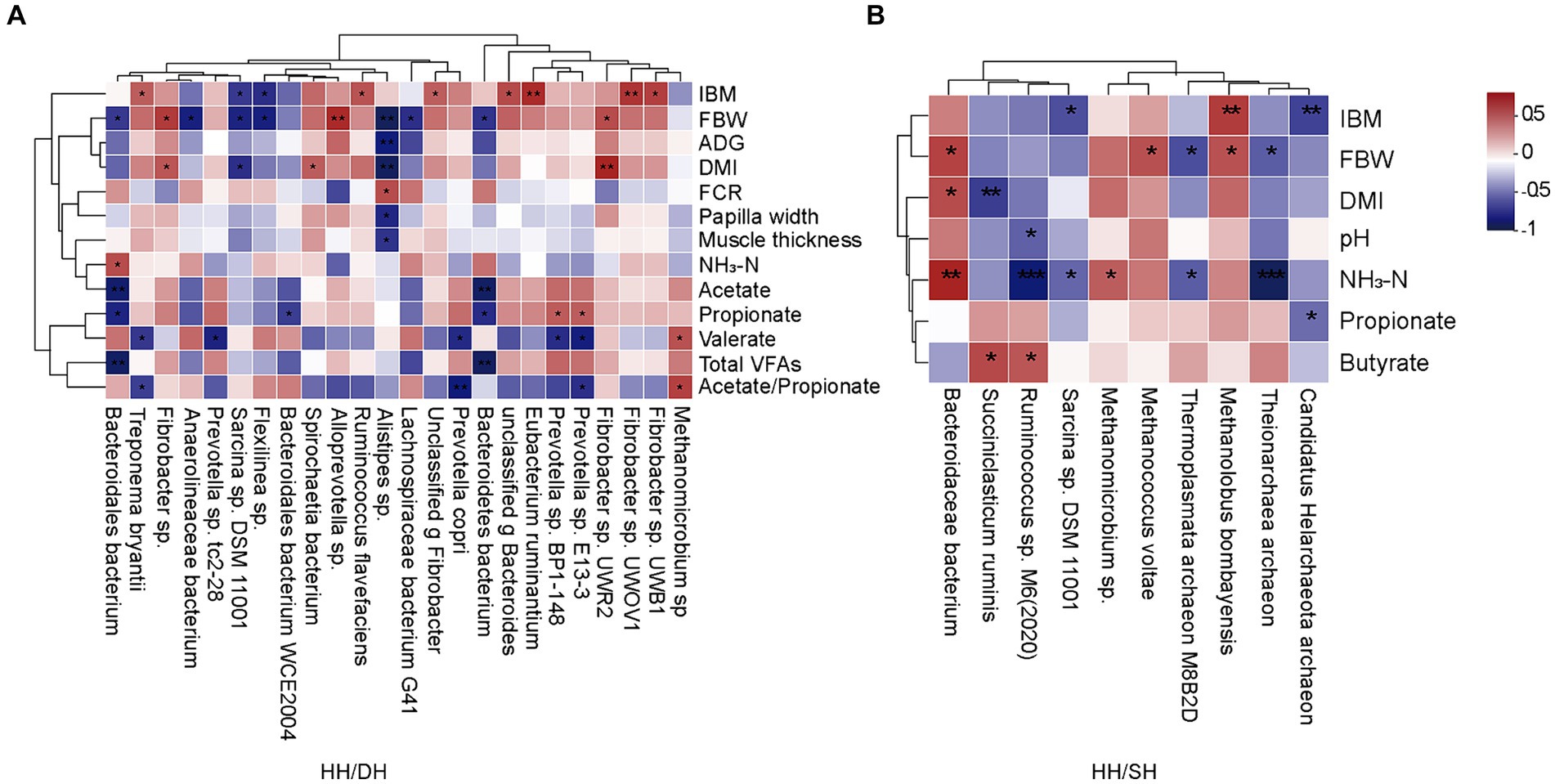
Figura 6. Análisis de correlación entre el rendimiento del crecimiento, los parámetros epiteliales del rumen, los parámetros de fermentación y microbiomas significativamente diferentes. (|r| > 0,5). (A) Entre los grupos HH y DH. (B) Entre los grupos HH y SH. n = 7 individuos/grupo. *p < 0,05, **p < 0,01 y ***p < 0,001.
3.8 Análisis del metaboloma ruminal
En total, se identificaron 947 metabolitos en las 21 muestras de rumen. Los gráficos de puntuación de las proyecciones ortogonales al análisis discriminante de estructuras latentes (OPLS-DA) mostraron una buena separación de metabolitos entre HH y DH (R2X = 0,419, R2Y = 0,994, Q2 = 0,345), HH y SH (R2X = 0,495, R2Y = 0,992, Q2 = 0,580) y DH y SH (R2X = 0,451, R2Y = 0,991, Q2 = 0,315) (Figura 7A). Después de examinar las concentraciones relativas de metabolitos ruminales por FC (FC ≥ 2 y FC ≤ 0,5) y VIP (VIP ≥ 1), obtuvimos 14 metabolitos enriquecidos de manera significativamente diferente en los grupos HH y DH, incluidos nueve metabolitos regulados al alza y cinco regulados a la baja (Figura 7B). Los niveles de lisofosfatidiletanolamina (LPE) (P-18:1), (P-17:0), (O-17:1), (P-17:1) y sulfóxido de metionina aumentaron significativamente en el grupo DH, mientras que thr-tyr, ciclo (his-pro) y heparina disminuyeron significativamente. Curiosamente, el sulfóxido de metionina solo estaba presente en el grupo DH. Un total de 35 metabolitos se enriquecieron de manera significativamente diferente entre los grupos HH y SH, con 19 regulados al alza y 16 a la baja (Figura 7C). Los niveles de 6 de ácidos orgánicos y sus derivados, 3 de acilos grasos (AG) y 2 de aminoácidos y sus derivados se incrementaron significativamente en el grupo SH, mientras que los niveles de 6 de nucleótidos y sus metabolitos, 2 de compuestos heterocíclicos, 3 de aminoácidos y sus derivados disminuyeron significativamente. El ácido N-metil-4-aminobutírico se identificó solo en el grupo SH. Entre los grupos DH y SH se observaron 21 metabolitos con diferencias significativas, incluyendo 3 regulados al alza y 18 regulados a la baja. Los niveles de ácido 3-hidroxi-3-metilbutírico, 7-cetocolesterol y lisofosfatidilcolina (LPC) (14:0/0:0) aumentaron significativamente, mientras que 4 de nucleótidos y sus metabolitos, 10 de glicerofosfolípidos (GP), 2 de alcohol y metabolitos de aminas disminuyeron significativamente (Figura suplementaria S7).
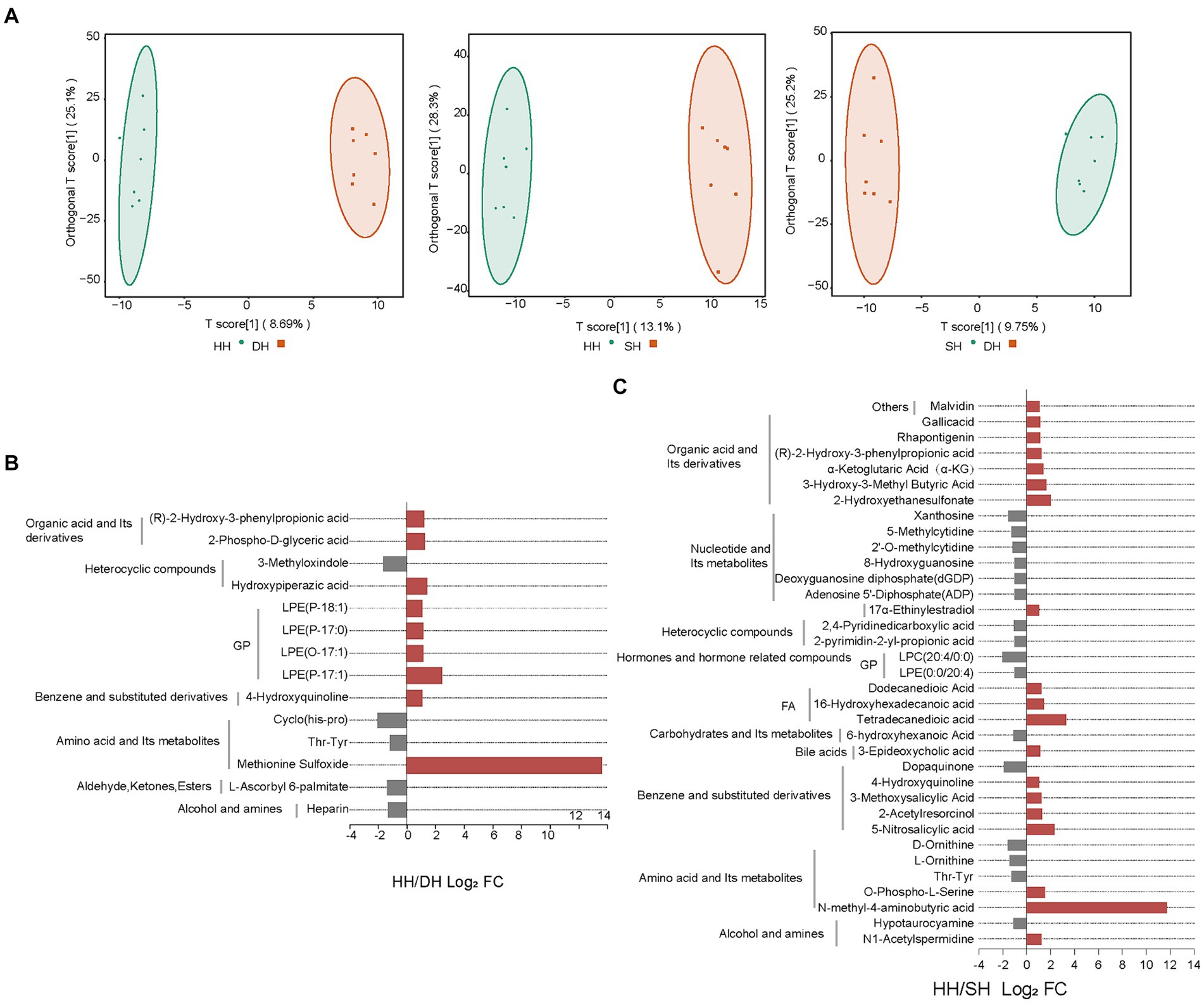
Figura 7. Cambio en los niveles de metabolitos del contenido ruminal. (A) Análisis OPLS-DA. (B) Metabolitos significativamente diferenciales entre los grupos HH y DH. (C) Metabolitos significativamente diferenciales entre los grupos HH y SH. n = 7 individuos/grupo.
El análisis de la vía KEGG reveló que de estos metabolitos diferenciales, seis fueron anotados en 16 vías entre los grupos HH y DH (Figura 8A). «Metabolismo de la cisteína y la metionina», «metabolismo de la fenilalanina», «metabolismo de la glicina, la serina y la treonina», «biosíntesis de aminoácidos», «glucólisis/gluconeogénesis» y «vía de las pentosas fosfato» se enriquecieron significativamente en el grupo DH, sin embargo, la «vía de señalización Fc epsilon RI» se enriqueció en el grupo HH. Se anotaron doce metabolitos en 48 vías entre los grupos HH y SH (Figura 8B). Entre ellos, relacionados con el metabolismo de los aminoácidos, el metabolismo de los ácidos biliares y las vías del ciclo del ácido tricarboxílico se enriquecieron principalmente en el grupo SH, por ejemplo, «metabolismo de la fenilalanina», «metabolismo de la histidina», «metabolismo de la glicina, la serina y la treonina», «metabolismo de la taurina y la hipotaurina», «secreción de bilis» y «ciclo del citrato (ciclo del TCA)». El grupo HH enriqueció el metabolismo de nucleótidos y transportadores ABC relacionados con el metabolismo de las purinas. Se anotaron siete metabolitos en 12 vías entre los grupos DH y SH (Figura suplementaria S8). «Digestión y absorción de grasas», «metabolismo de glicerolípidos» y «digestión y absorción de vitaminas» se enriquecieron principalmente en el grupo DH, mientras que «degradación de valina, leucina e isoleucina» y «metabolismo de colina en cáncer» se enriquecieron principalmente en el grupo SH.
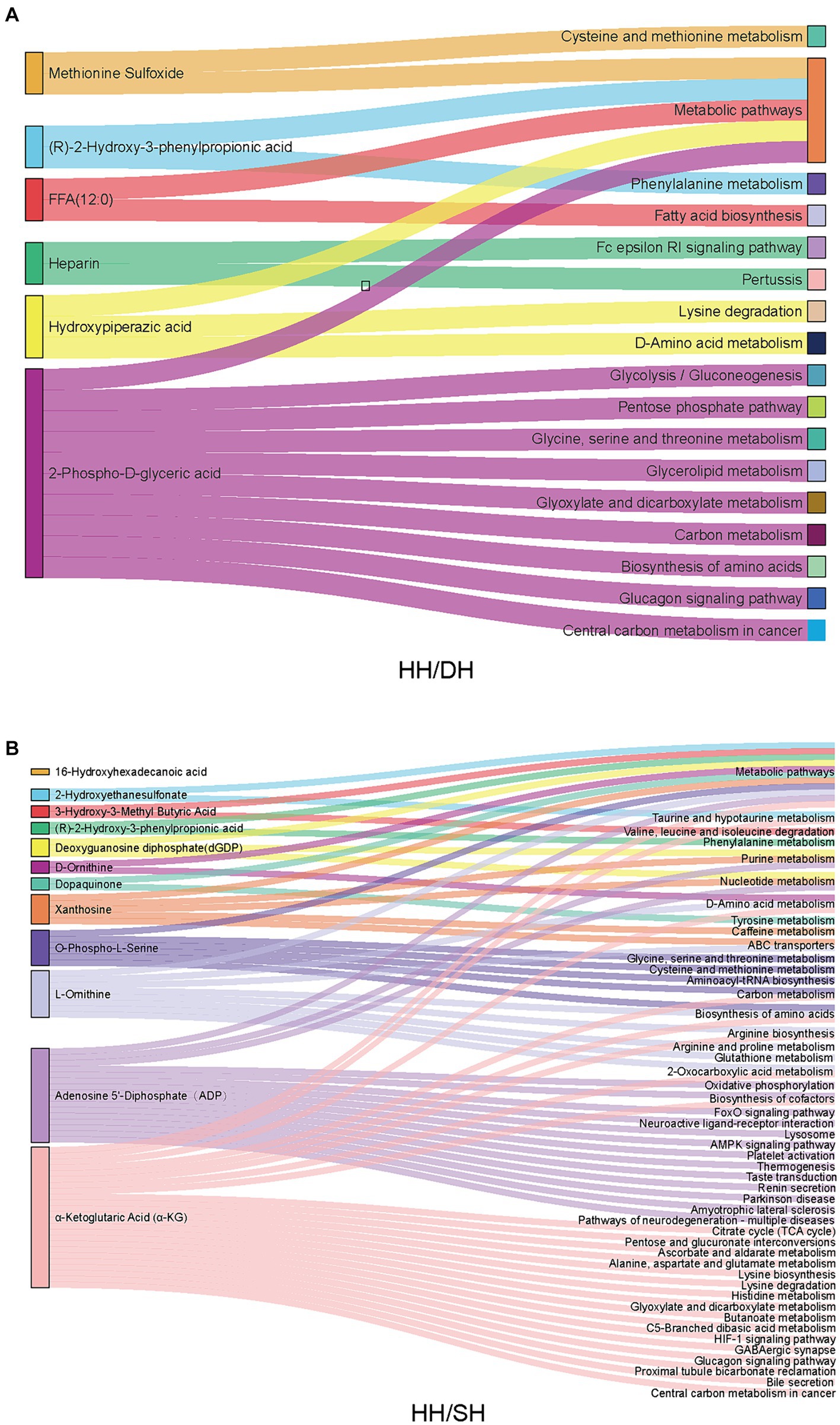
3.9 Análisis de metabolitos relacionados con el microbioma
El análisis del coeficiente de correlación de Spearman identificó correlaciones entre los microbiomas diferenciales y los metabolitos (|r| > 0,8; p < 0,05). Los niveles de Treponema bryantii, Fibrobacter succinogenes, Fibrobacter sp. UWB4 y Fibrobacter sp. se correlacionaron positivamente con LPE (P-17:0); Alloprevotella sp. se correlacionó positivamente con 4-hidroxiquinoleína; y Alistipes sp. se correlacionó negativamente con el ácido hidroxipipelázico cuando comparamos los grupos HH y DH (Figura 9A). Bacteroidales bacterium y Ruminococcus sp. M6 (2020) se correlacionaron positivamente con el ácido 3-hidroxi-3-metilbutírico y la dopaquinona, respectivamente; mientras que Theionarchaea archaeon y Thermoplasmata archaeon M8B2D se correlacionaron negativamente con el ácido tetradecanodioico en los grupos HH y SH (Figura 9B). Candidatus Altiarchaeales archaeon WOR SM1 86-2 se correlacionó positivamente con LPE (16:0/0:0), LPE (0:0/16:0), LPE (O-18:2), fosfatidilcolina (PC) (O-1:0/O-16:0), lisofosfatidilserina (LPS) (20:0), ácido lisofosfatídico (LPA) (22:6); El 7-cetocolesterol se correlacionó positivamente con Candidatus Methanomethylophilaceae archaeon, Candidatus Methanomethylophilus sp. y Bacteroidales bacterium entre los grupos DH y SH (Figura suplementaria S8).
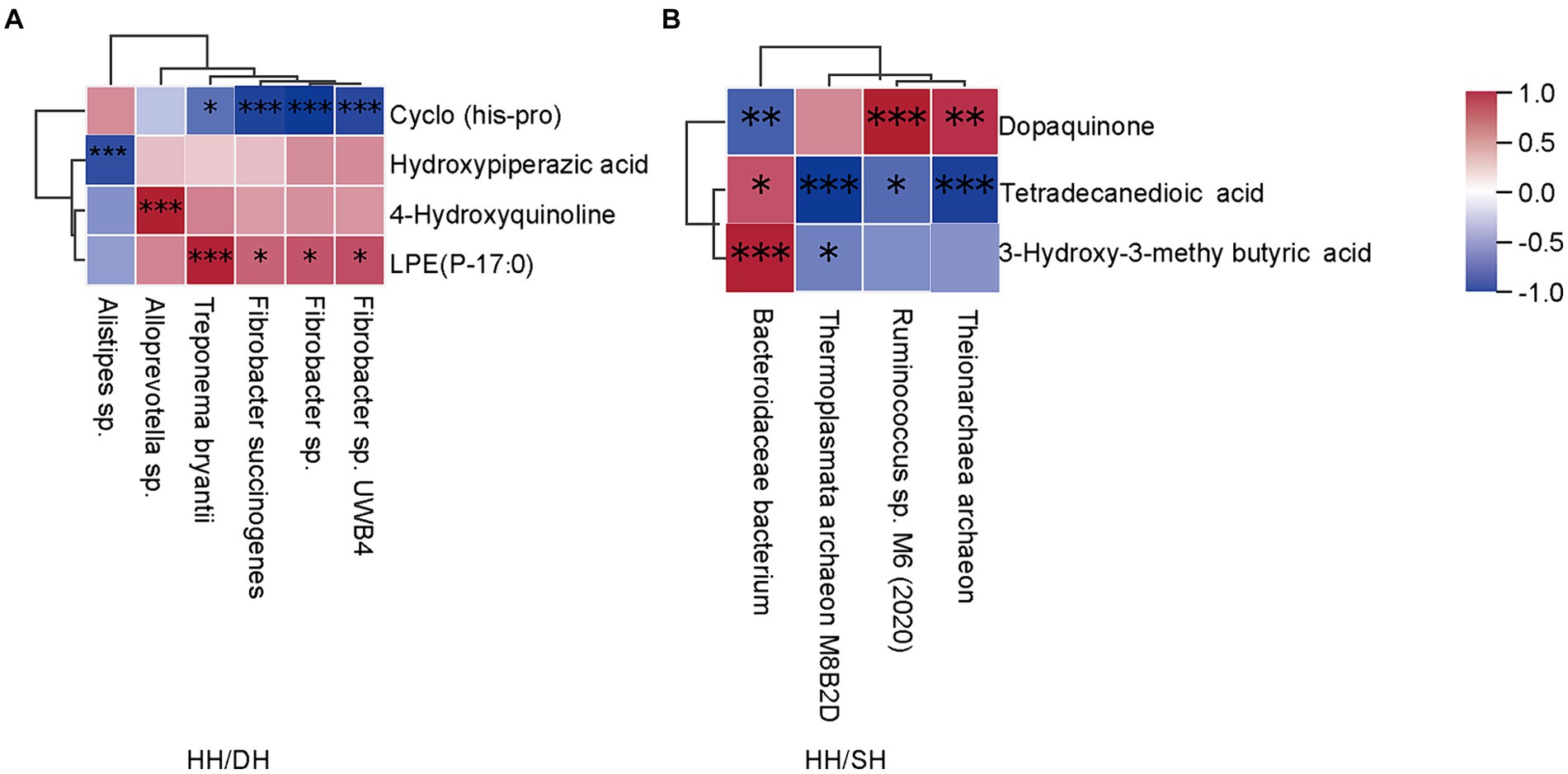
4 Discusión
Desde el punto de vista de la genética y el mejoramiento, la introducción de un gen exógeno favorable puede ser una mejor opción para mejorar la heredabilidad baja a moderada de los rasgos (28). En este estudio, seleccionamos ovejas Poll Dorset y Southdown como sementales para mejorar el rendimiento del crecimiento de las ovejas Hu, centrándonos en cómo la hibridación afecta el rendimiento del crecimiento e investigando el mecanismo de regulación del microbioma ruminal y sus metabolitos.
La hibridación da lugar a la heterosis y mejora el crecimiento de los animales. Como se esperaba, nuestros datos mostraron que la hibridación aumentó el DMI y el ADG, lo que resultó en un mayor FBW y un menor F/G. Nuestro estudio previo indicó que la descendencia híbrida F1 creció más rápido y se hizo más grande que las ovejas Hu, particularmente durante los últimos períodos (29). Otro estudio sobre un sistema de cruzamiento de tres vías entre Angus, Qaidam y yak indicó que las crías de F2 eran significativamente mejores que las del yak y 1/2 yak (28). Estos resultados sugieren la importancia de la hibridación para mejorar el rendimiento del crecimiento de los animales.
La microecología ruminal está estrechamente asociada con el crecimiento animal. En este estudio, la hibridación cambió significativamente el patrón de fermentación ruminal en ovejas, especialmente la concentración de NH3-N. El NH3-N se deriva principalmente de la desaminación de aminoácidos mediada por microbios y la hidrólisis de nitrógeno no proteico en el rumen (30). El microbioma ruminal utiliza NH3-N para proporcionar precursores primarios de la síntesis de proteínas para el huésped. El NH3-N también tiene un efecto de retroalimentación sobre la estructura de la microbiota ruminal y la función epitelial, afectando a la fermentación ruminal y a la salud del huésped (31). La concentración de NH3-N fue significativamente menor en los grupos SH y DH que en el grupo HH, lo que indica que la descendencia híbrida tuvo una mayor eficiencia de utilización de las fuentes de nitrógeno para sintetizar proteínas bacterianas. Los AGV son la principal fuente de energía para los rumiantes, que fermentan materiales vegetales en AGV que regulan una variedad de funciones fisiológicas en el rumen (32). El acetato, el propionato y el butirato son los tres principales AGV y proporcionan aproximadamente el 70% de los requisitos energéticos. La concentración de acetato se ve afectada principalmente por la degradación microbiana de las sustancias fibrosas. Los informes han demostrado que la concentración de acetato ruminal aumenta cuando los rumiantes se alimentan con una dieta rica en fibra (33) o cuando la microbiota ruminal está dominada por Prevotella (34). Del acetato en el rumen, el 67% se utiliza para la energía oxidativa, mientras que el resto se utiliza para la síntesis de grasa corporal. El propionato contribuye a la suplementación energética a través de la vía gluconeogénica para promover la síntesis de glucosa (35). En este estudio, los niveles de acetato y propionato en el grupo DH aumentaron significativamente, lo que resultó en AGV totales más altos, lo que indica un mayor suministro de energía al huésped y un mejor rendimiento de crecimiento.
El epitelio ruminal es un indicador importante del desarrollo ruminal y es responsable de la absorción y el metabolismo de los nutrientes y los subproductos microbianos. La evidencia ha demostrado que los AGV se absorben a través del epitelio ruminal y que su capacidad de absorción está relacionada principalmente con el área superficial y la expresión de vectores transportadores de AGV en el epitelio (36). El epitelio ruminal es abundante en las mitocondrias, lo que impulsa el metabolismo epitelial; Un epitelio más grueso indica una utilización más eficiente del alimento en el ganado (37). Además, el butirato estimula la proliferación celular epitelial (38). Nuestros resultados indican que la hibridación aumentó la altura de la papila, mientras que no se observaron cambios en el ancho de la papila ni en el grosor del músculo, lo que sugiere una mayor superficie de contacto con los nutrientes para triturar completamente el alimento para promover la digestión y la eficiencia alimenticia (39). Además, se considera que estas variaciones en las estructuras físicas del epitelio ruminal pueden influir en la microbiota ruminal.
Utilizando PCoA, el presente estudio encontró diferencias significativas en el microbioma ruminal entre la descendencia híbrida F1 y las ovejas Hu. La hibridación puede producir un nuevo flujo génico y diferentes patrones de expresión génica en la descendencia. El ciervo Sika y el ciervo alce tienen una microbiota ruminal diferente a la de sus crías híbridas; Los ratones puros e híbridos tienen diferentes características de agrupamiento microbiano, lo que sugiere un efecto significativo de la genética del huésped en el microbioma ruminal que puede resultar de la transmisión vertical (40-42). Además, encontramos que Bacteroidetes y Firmicutes fueron las bacterias dominantes a nivel de filo, sin embargo, la hibridación no tuvo un efecto considerable sobre sus abundancias relativas. La evidencia ha demostrado que los Bacteroidetes tienen estimaciones de heredabilidad bajas, mientras que los Firmicutes tienen estimaciones de heredabilidad moderadas, lo que sugiere que se ven afectados en gran medida por factores ambientales, como la dieta (15, 43). Al mismo tiempo, las diferentes estimaciones de heredabilidad indican que los efectos del huésped no son iguales para los diferentes filotipos microbianos del rumen. A nivel de especies, observamos que las bacterias relacionadas con la degradación de la fibra fueron más abundantes en el grupo DH, Treponema bryantii, Prevotella copri y Fibrobacter succinogenes fueron particularmente abundantes. Treponema bryantii, cuando está presente con especies celulolíticas como Bacteroides succinogenes, puede promover la digestión de materiales celulósicos para aumentar la eficiencia alimenticia (44). Se encontró una mayor abundancia de Treponema bryantii y Bacteroides sp. en el ciego de cerdos con alta eficiencia alimenticia (45). Las Prevotella se consideran las bacterias más importantes para la degradación y fermentación de polisacáridos en función de sus perfiles genómicos de glucósidos hidrolasas, expresión génica y abundancia en el rumen (46). Prevotella copri podría ser la especie bacteriana clave asociada con la ingesta de alimento del huésped y el metabolismo de las grasas, y la evidencia muestra que su abundancia aumenta en cerdos con una ingesta diaria de alimento promedio alta (47) y promueve la acumulación de grasa en cerdos alimentados con dietas de fórmula (48). Los estudios en humanos indican que Prevotella copri aumenta en gran medida el potencial microbiano para la biosíntesis de aminoácidos de cadena ramificada (49), lo que es beneficioso para el rendimiento de la proteína láctea en el ganado lechero y el crecimiento animal (50, 51). Además, Prevotella copri es hereditaria (h2 > 0,15) en el ciego y las heces de los cerdos (52). Fibrobacter succinogenes es considerada como la bacteria más celulolítica en los microbiomas ruminales y se especializa en la producción de succinato, acetato y formiato (53). Un aumento en la población de Fibrobacter succinogenes en el rumen se atribuye a un aumento en la degradabilidad de la materia seca (54). En el presente estudio, las abundancias relativas de Fibrobacter sp. UWR2 y Fibrobacter sp. se relacionaron positivamente con DMI, lo que sugiere su papel esencial en la DMI y la degradabilidad de la materia seca. En el grupo SH, la abundancia de la bacteria Bacteroidaceae fue mayor que la del grupo HH. La bacteria Bacteroidaceae también es una bacteria celulolítica del rumen que puede convertir el succinato en propionato como elLe fuente de energía (55). Por lo tanto, la mayor abundancia de bacterias degradadoras de polisacáridos en el rumen de las ovejas híbridas mejora la degradación de los carbohidratos de la dieta, promoviendo así la capacidad digestiva y el rendimiento del crecimiento. Esto puede atribuirse a la genética del huésped porque excluimos los efectos de los factores ambientales. Sin embargo, esta especulación debe verificarse mediante el seguimiento de la microbiota ruminal de las ovejas Poll Dorset y Southdown. El grupo DH carecía de arqueas relacionadas con la digestión y absorción de nutrientes, sin embargo, la abundancia de bacterias metanogénicas (Methanomicrobium sp., Methanolobus bombayensis y Methanococcus voltae) aumentó en el grupo SH en comparación con los niveles del grupo HH, y la producción de metano es un proceso intensivo en energía, lo que resulta en un bajo rendimiento de producción. Sin embargo, en este estudio, la abundancia de estas bacterias metanogénicas fue bastante baja. También encontramos que la DMI y la eficiencia de la utilización de proteínas para piensos en el grupo SH fueron mayores que las del grupo HH, lo que sugiere que las contribuciones de DMI y eficiencia en la utilización de proteínas para piensos superaron la tasa de producción de metano, lo que puede ser la razón de la ganancia de FBW en el grupo SH. Además, Xue et al. encontraron que algunas bacterias metanogénicas con baja abundancia estaban enriquecidas en los rumenes de vacas lecheras con altos rendimientos de proteína láctea (50); Sin embargo, se necesitan más estudios para validar nuestras hipótesis.
El análisis taxonómico funcional de KEGG reveló que la biosíntesis y el metabolismo de los aminoácidos se enriquecieron en el grupo SH. Los productos metabólicos como la lisina, el aspartato, la glicina, la alanina, el glutamato, la serina y la treonina son importantes contribuyentes a la síntesis de proteínas microbianas del rumen, constituyendo el 90% de las proteínas microbianas que llegan al intestino delgado y proporcionan una fuente de proteínas para el huésped (56). Nuestro estudio previo identificó que el contenido de proteínas musculares era mayor en el grupo SH que en el grupo HH (29), lo que está en línea con los resultados de la microbiota ruminal. Los genes que codifican CAZymes involucrados en la deconstrucción de carbohidratos (GH, CE, PL y CBM) se enriquecieron en el grupo DH, lo que indica que estas ovejas eran más capaces de degradar sustratos complejos para promover la fermentación microbiana, lo que es consistente con nuestros resultados de VFA.
Además de afectar la composición y función del microbioma ruminal, la hibridación altera los metabolitos ruminales y las vías metabólicas. En primer lugar, los LPEs son componentes de la membrana celular y son los segundos más abundantes in vivo después de los LPCs (57, 58). Según se informa, los LPE inducen la activación de la señalización de la proteína quinasa activada por mitógenos y exhiben efectos antiapoptóticos en las células de feocromocitoma (59). LPE (P-18:1) tiene ácido oleico como su cadena de acilo, estimulando el crecimiento de neuritas y efectos neuroprotectores contra la excitotoxicidad inducida por glutamato (60). Además, los LPEs están implicados en la formación de gotas lipídicas al suprimir la lipólisis y la biosíntesis de ácidos grasos, promoviendo así la absorción de sustancias lipídicas (61). Esta evidencia indica que los LPEs enriquecidos en el grupo DH se beneficiaron de la deposición de grasa, lo que resultó en un aumento de FBW. Curiosamente, el sulfóxido de metionina solo estaba presente en el grupo DH. La exposición de las proteínas a especies reactivas de oxígeno y peróxido de hidrógeno puede conducir a la oxidación de la metionina libre y de los residuos de metionina, formando sulfóxido de metionina (62). Los niveles de sulfóxido de metionina se elevan durante el estrés oxidativo, el envejecimiento y la inflamación. Los resultados de este estudio implican que las ovejas del grupo DH tenían poca adaptabilidad ambiental. Sin embargo, el sulfóxido de metionina puede ser reparado diastereoselectivamente por la metionina sulfóxido reductasa para producir metionina (63). La metionina aumenta la capacidad antioxidante endógena de las proteínas del arroz al estimular la expresión de metionina sulfóxido reductasa y mejorar la síntesis de glutatión a través de la vía Nrf2-ARE (64). Los efectos beneficiosos de la suplementación con metionina sobre la actividad antioxidante de las aves estresadas por calor se han demostrado previamente (65). El presente estudio muestra que, aunque las ovejas del grupo DH se encontraban en un estado de estrés oxidativo, este fenómeno puede mejorarse mediante la regulación del sistema de metionina sulfóxido reductasa, que será nuestra próxima dirección de investigación. El ácido N-metil-4-aminobutírico solo estuvo presente en el grupo SH. El producto metabólico ácido aminobutírico actúa como un neurotransmisor inhibidor clave del sistema nervioso central y funciona a través de la sedación, aliviando la fiebre y disminuyendo la producción de calor (66). El ácido aminobutírico reduce el estrés causado por diversas condiciones ambientales y mejora la productividad (67). Por ejemplo, el ácido aminobutírico mejora el rendimiento del crecimiento, la calidad de la carne y la ingesta de alimento al promover la activación de enzimas características y mejorar la absorción y la función inmune de la mucosa intestinal en condiciones de estrés térmico (68, 69). Los metabolitos característicos en los grupos DH y SH se relacionaron con el estrés oxidativo, lo que indica que la adaptabilidad ambiental de las ovejas de la generación F1 necesita mejoras, lo que esperamos que ocurra en la generación F2. El ácido α-cetoglutárico es un metabolito intermedio del ciclo del ácido tricarboxílico y se genera a partir de glucosa y glutamina, proporcionando sustratos para la síntesis de carbohidratos, aminoácidos, grasas y otras biomoléculas (70). En este estudio, el α-cetoglutarato aumentó significativamente en el grupo SH en comparación con los niveles en el grupo HH, mejorando aún más la capacidad del metabolismo de los aminoácidos. Nuestros resultados fueron consistentes con un informe previo de Kong (71), quien confirmó que las vías del ciclo del ácido tricarboxílico mejoraron en el músculo de las ovejas de la generación Southdown × Hu F1.
Cuatro metabolitos y seis microbiomas mostraron relaciones reguladoras en el rendimiento del crecimiento entre los grupos DH y HH. Alloprevotella sp. puede fermentar carbohidratos para producir acetato y succinato como metabolitos finales y se correlaciona positivamente con la 4-hidroxiquinoleína, que tiene efectos antibacterianos y antiinflamatorios que mantienen la salud del rumen (72). Treponema bryantii, Fibrobacter succinogenes, Fibrobacter sp. y Fibrobacter sp.UWB4 se asocian con la degradación de la fibra y la producción de propionato, que se correlacionan positivamente con LPE (P-17:0). El propionato puede promover la síntesis de glucosa a través de la vía gluconeogénica en el rumen, y la glucosa es un precursor de la forma de glicerol y ácidos grasos, que ayudan aún más a formatear los LPE. Los LPE se originan en parte a partir de los lípidos de la dieta, pero también de la síntesis microbiana intestinal. Los niveles de fosfolípidos en las células huésped de ratones libres de gérmenes difieren de los de las células huésped de ratones convencionales, lo que sugiere una relación entre los microbios intestinales y los glicerofosfolípidos (73). La abundancia relativa de Faecalibacterium y Prevotella está relacionada con los lípidos circulantes, en particular los LEP y el fosfatidilglicerol (74); sin embargo, encontramos en el presente estudio que Fibrobacter también se relacionó con los niveles de LPE (P-17:0). La bacteria Bacteroidales se correlacionó positivamente con el ácido 3-hidroxi-3-metilbutírico entre los grupos SH y HH. El ácido 3-hidroxi-3-metilbutírico es un metabolito de la leucina que estimula el eje de la hormona del crecimiento / factor de crecimiento similar a la insulina-1 (75). Estos resultados indican que la microbiota ruminal de las ovejas del grupo DH estuvo implicada principalmente en la deposición de grasa corporal mediante el aumento de la DMI y la degradabilidad de la materia seca, mientras que la del grupo SH participó principalmente en el metabolismo de los aminoácidos para proporcionar energía. Sin embargo, la relación causal entre la microbiota y los metabolitos requiere más investigación. Además, los datos de microbiomas y metabolitos del rumen de las ovejas Poll Dorset y Southdown deben considerarse y dilucidarse más a fondo en estudios futuros.
5 Conclusión
Los resultados de este estudio respaldan la viabilidad de utilizar programas de cruzamiento para promover el crecimiento y aumentar la eficiencia alimenticia mediante la alteración de la microbiota ruminal y los metabolitos en ovejas. En el grupo DH, los niveles de Treponema bryantii, Prevotella copri y Fibrobacter succinogenes aumentaron, y sus funciones se enriquecieron principalmente a nivel de CAZyme, contribuyendo a niveles más altos de acetato, propionato, AGV totales y LPE para promover aún más el crecimiento. La bacteria Bacteroidaceae se enriqueció en el grupo SH y está involucrada en el metabolismo de los aminoácidos, satisfaciendo la demanda de proteínas microbianas ruminales que son utilizadas por los huéspedes para promover el crecimiento. Además, el sulfóxido de metionina y el ácido N-metil-4-aminobutírico fueron metabolitos característicos en los grupos DH y SH, respectivamente, lo que indica que las ovejas cruzadas de la generación F1 tenían poca adaptabilidad ambiental. En conjunto, estos hallazgos proporcionan nuevas ideas y estrategias nutricionales para el cultivo de nuevas razas de ovejas.
Declaración de disponibilidad de datos
Los datos presentados en el estudio se depositan en el repositorio de la SRA, número de acceso PRJNA1111660.
Declaración ética
El estudio en animales fue aprobado por el Comité de Administración y Ética Animal del Instituto de Ganadería y Ciencias Farmacéuticas de Lanzhou de la Academia China de Ciencias Agrícolas. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
RZ: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Visualización, Redacción – borrador original, Redacción – revisión y edición. LZ: Curación de datos, Investigación, Metodología, Redacción – borrador original. XA: Curación de datos, Investigación, Metodología, Redacción – revisión y edición. JL: Curación de datos, Metodología, Recursos, Redacción – revisión y edición. CN: Investigación, Validación, Visualización, Redacción – Revisión y Edición. JZ: Curación de datos, Metodología, Visualización, Redacción, revisión y edición. ZG: Curación de datos, Metodología, Visualización, Redacción – revisión y edición. TX: Curación de datos, investigación, redacción, revisión y edición. POR: Conceptualización, Análisis formal, Escritura – revisión y edición. ZX: Conceptualización, Investigación, Administración de proyectos, Supervisión, Redacción – borrador original, Redacción – revisión y edición. YY: Conceptualización, Obtención de fondos, Investigación, Administración de proyectos, Supervisión, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declara(n) que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio fue financiado por la Tarea Principal de Investigación Científica del Proyecto de Innovación Científica y Tecnológica de la Academia China de Ciencias Agrícolas (CAAS-ZDRW202106), el Fondo de Investigación Basal de la Institución Científica Central de Interés Público (1610322023014, 1610322024020) y el Programa de Ciencia y Tecnología de la provincia de Gansu (22ZD6NA037, 21ZD11NM001).
Reconocimientos
Estamos agradecidos con la Compañía de Cría de Ovejas Cordero de Qinghuan por su trabajo, incluido el cuidado y la alimentación de los animales.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1455029/full#supplementary-material
Notas
Referencias
1. Bunning, H, Wall, E, Chagunda, M, Baños, G y Simm, G. Heterosis en esquemas de cruzamiento de ganado en regiones tropicales: metaanálisis de los efectos de la combinación de razas, el tipo de rasgo y el clima en el nivel de heterosis. J Anim Sci. (2019) 97:29–34. doi: 10.1093/jas/sky406
2. Liu, A, Chen, X, Liu, M, Zhang, L, Ma, X y Tian, S. Expresión diferencial y análisis funcional de CircRNA en los ovarios de ovejas Hanper de baja y alta fecundidad. Animales (Basilea). (2021) 11:71863. doi: 10.3390/ani11071863
3. Yang, Y, Shen, L, Gao, H, Ran, J, Li, X, Jiang, H, et al. Comparación de la composición de la microbiota cecal en cerdos híbridos a partir de dos cruzamientos separados de tres vías. Anim Biosci. (2021) 34:1202–9. doi: 10.5713/ab.20.0681
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Christensen, OF, Legarra, A, Lund, MS y Su, G. Evaluación genética para el cruzamiento de tres vías. Genet Sel Evol. (2015) 47:98. doi: 10.1186/s12711-015-0177-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Quan, K, Li, J, Han, H, Wei, H, Zhao, J, Si, HA, et al. Revisión de la oveja Huang-Huai, una nueva raza de ovejas multíparas identificada por primera vez en China. Trop Anim Salud Prod. (2020) 53:35. DOI: 10.1007/S11250-020-02453-W
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Lu, Z, Yue, Y, Shi, H, Zhang, J, Liu, T, Liu, J, et al. Efectos de los sementales de ovejas sobre las características de la fibra muscular, la composición de ácidos grasos y los compuestos de sabor volátiles en corderos cruzados F1. Seguridad alimentaria. (2022) 11:4076. doi: 10.3390/alimentos11244076
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Zhao, L, Zhang, D, Li, X, Zhang, Y, Zhao, Y, Xu, D, et al. La proteómica comparativa revela los mecanismos genéticos del peso corporal en ovejas Hu y ovejas Dorper. J Proteoma. (2022) 267:104699. doi: 10.1016/j.jprot.2022.104699
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Quan, F, Zhang, Z, An, Z, Hua, S, Zhao, X y Zhang, Y. Múltiples factores que afectan la superovulación en Dorset en China. Reprod Domest Anim. (2011) 46:39–44. doi: 10.1111/j.1439-0531.2009.01551.x
9. Salón, SJG . Conservación y desarrollo de razas minoritarias británicas de ovejas: el ejemplo de Southdown. J Agric Sci. (1989) 112:39–45. doi: 10.1017/S0021859600084082
10. Dai, R, Zhou, H, Fang, Q, Zhou, P, Yang, Y, et al. Variación en DGAT1 ovino y su asociación con rasgos musculares de la canal en ovejas de Southdown. Genes (Basilea). (2022) 13:13. doi: 10.3390/genes13091670
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Cheng, J, Zhang, X, Xu, D, Zhang, D, Zhang, Y, Song, Q, et al. Relación entre las diferencias y rasgos microbianos del rumen entre las ovejas Hu, las ovejas Tan y las ovejas Dorper. J Anim Sci. (2022) 100:100. doi: 10.1093/jas/skac261
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Firkins, JL y Yu, Z. Simposio de nutrición de rumiantes: cómo utilizar los datos sobre el microbioma ruminal para mejorar nuestra comprensión de la nutrición de los rumiantes. J Anim Sci. (2015) 93:1450–70. doi: 10.2527/jas.2014-8754
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Li, F, Hitch, T, Chen, Y, Creevey, CJ y Guan, LL. Los análisis metagenómicos y metatranscriptómicos comparativos revelan el efecto de la raza en el microbioma ruminal y sus asociaciones con la eficiencia alimentaria en el ganado vacuno. Microbioma. (2019) 7:6. doi: 10.1186/s40168-019-0618-5
14. Cersosimo, LM, Bainbridge, ML, Wright, AD y Kraft, J. La raza y la etapa de lactancia alteran los perfiles de ácidos grasos protozoarios del rumen y las estructuras de la comunidad en el ganado lechero primíparo. J Agric Food Chem. (2016) 64:2021–9. doi: 10.1021/acs.jafc.5b05310
15. Li, F, Li, C, Chen, Y, Liu, J, Zhang, C, Irving, B, et al. La genética del huésped influye en la microbiota ruminal y las características microbianas hereditarias del rumen se asocian con la eficiencia alimenticia en el ganado. Microbioma. (2019) 7:92. doi: 10.1186/s40168-019-0699-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Zhang, R, Wu, J, Lei, Y, Bai, Y, Jia, L, Li, Z, et al. Los aceites esenciales de orégano promueven la capacidad digestiva del rumen al modular el desarrollo epitelial y la composición de la microbiota en el ganado vacuno. Tuerca delantera. (2021) 8:722557. doi: 10.3389/fnut.2021.722557
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Chen, S, Zhou, Y, Chen, Y y Gu, J. Fastp: un preprocesador FASTQ todo en uno ultrarrápido. Bioinformática. (2018) 34:i884–90. doi: 10.1093/bioinformática/bty560
18. Li, H y Durbin, R. Alineación de lectura corta rápida y precisa con transformada de excavadoras-ruedas. Bioinformática. (2009) 25:1754–60. doi: 10.1093/bioinformática/BTP324
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Li, D, Liu, CM, Luo, R, Sadakane, K y Lam, TW. Megahit: una solución ultrarrápida de un solo nodo para el ensamblaje de metagenómica grande y compleja a través de un grafo sucinto de Bruijn. Bioinformática. (2015) 31:1674–6. doi: 10.1093/bioinformática/BTV033
20. Noguchi, H, Park, J y Takagi, T. MetaGene: hallazgo de genes procariotas a partir de secuencias de escopeta del genoma ambiental. Ácidos nucleicos Res. (2006) 34:5623–30. doi: 10.1093/nar/gkl723
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Fu, L, Niu, B, Zhu, Z, Wu, S y Li, W. CD-HIT: acelerado para agrupar los datos de secuenciación de próxima generación. Bioinformática. (2012) 28:3150–2. doi: 10.1093/bioinformática/BTS565
22. Li, R, Li, Y, Kristiansen, K y Wang, J. SOAP: programa corto de alineación de oligonucleótidos. Bioinformática. (2008) 24:713–4. doi: 10.1093/bioinformática/BTN025
23. Altschul, SF, Madden, TL, Schaffer, AA, Zhang, J, Zhang, Z, Miller, W, et al. Gapped BLAST y PSI-BLAST: una nueva generación de programas de búsqueda de bases de datos de proteínas. Ácidos nucleicos Res. (1997) 25:3389–402. doi: 10.1093/nar/25.17.3389
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Altschul, SF, Gish, W, Miller, W, Myers, EW y Lipman, DJ. Herramienta básica de búsqueda de alineaciones locales. J Mol Biol. (1990) 215:403–10. doi: 10.1016/S0022-2836(05)80360-2
25. Sun, HZ, Wang, DM, Wang, B, Wang, JK, Liu, HY, Guan, LL, et al. Metabolómica de cuatro biofluidos de vacas lecheras: potenciales biomarcadores para la producción y calidad de la leche. J Proteoma Res. (2015) 14:1287–98. DOI: 10.1021/PR501305G
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. De Almeida, R, Do, PR, Porto, C, Dos, SG, Huws, SA, y Pilau, EJ. Exploración del metaboloma del fluido ruminal mediante cromatografía líquida, espectrometría de masas de alta resolución y redes moleculares. Sci Rep. (2018) 8:17971. doi: 10.1038/s41598-018-36196-4
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Kanehisa, M, y Goto, S. KEGG: Enciclopedia de genes y genomas de Kioto. Ácidos nucleicos Res. (2000) 28:27–30. doi: 10.1093/nar/28.1.27
28. Cao, XK, Cheng, J, Huang, YZ, Wang, XG, Ma, YL, Peng, SJ, et al. Evaluaciones del rendimiento del crecimiento y la calidad de la carne en ganado cruzado de tres vías desarrollado para la meseta tibetana y su comprensión molecular mediante análisis ómicos integrativos. J Agric Food Chem. (2019) 67:541–50. doi: 10.1021/acs.jafc.8b05477
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Zhang, R, An, X, Li, J, Lu, Z, Niu, C, Xu, Z, et al. Análisis comparativo del rendimiento del crecimiento, la productividad de la carne y la calidad de la carne en ovejas Hu y sus híbridos. Acta Pratacul Sin. (2024) 33:186–97. doi: 10.11686/cyxb2023157
30. Schwab, CG, y Broderick, GA. Una revisión de 100 años: nutrición de proteínas y aminoácidos en vacas lecheras. J Dairy Sci. (2017) 100:10094–112. doi: 10.3168/jds.2017-13320
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Shen, J, Zheng, W, Xu, Y y Yu, Z. La inhibición de la fermentación ruminal in vitro con alto contenido de amoníaco depende del pH. Frente Vet Sci. (2023) 10:163021. doi: 10.3389/fvets.2023.1163021
32. Shen, H, Xu, Z, Shen, Z y Lu, Z. La regulación de los ácidos grasos ruminales de cadena corta sobre las funciones de las barreras ruminales. Fisiola delantera. (2019) 10:1305. doi: 10.3389/fphys.2019.01305
33. Li, QS, Wang, R, Ma, ZY, Zhang, XM, Jiao, JZ, Zhang, ZG, et al. La selección dietética de microorganismos metabólicamente distintos impulsa el metabolismo del hidrógeno en los rumiantes. ISME J. (2022) 16:2535–46. doi: 10.1038/s41396-022-01294-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Wang, D, Tang, G, Wang, Y, Yu, J, Chen, L, Chen, J, et al. Identificación de grupos bacterianos ruminales y su influencia en los metabolitos ruminales y el rendimiento del crecimiento de cabras jóvenes. Anim Nutr. (2023) 15:34–44. doi: 10.1016/j.aninu.2023.05.013
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. De Vadder, F, Kovatcheva-Datchary, P, Zitoun, C, Duchampt, A, Bäckhed, F y Mithieux, G. El succinato producido por la microbiota mejora la homeostasis de la glucosa a través de la gluconeogénesis intestinal. Célula Metab. (2016) 24:151–7. doi: 10.1016/j.cmet.2016.06.013
36. Chai, J, Lv, X, Diao, Q, Usdrowski, H, Zhuang, Y, Huang, W, et al. La dieta sólida manipula la microbiota epitelial ruminal y sus interacciones con la transcriptómica del huésped en rumiantes jóvenes. Environ Microbiol. (2021) 23:6557–68. doi: 10.1111/1462-2920.15757
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Lam, S, Munro, JC, Zhou, M, Guan, LL, Schenkel, FS, Steele, MA, et al. Asociaciones de los parámetros ruminales con la eficiencia alimenticia y la rutina de muestreo en bovinos de carne. Animal. (2018) 12:1442–50. DOI: 10.1017/S1751731117002750
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Zhang, K, Xu, Y, Yang, Y, Guo, M, Zhang, T, Zong, B, et al. Los metabolitos derivados de la microbiota intestinal contribuyen negativamente al desarrollo de la función de barrera del intestino posterior en el modelo de cabra de destete temprano. Anim Nutr. (2022) 10:111–23. doi: 10.1016/j.aninu.2022.04.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Liu, H, Hu, J, Mahfuz, S y Piao, X. Efectos de los taninos hidrolizables como sustitutos del óxido de zinc sobre el estado antioxidante, la función inmunológica, la morfología intestinal y las actividades de las enzimas digestivas en lechones destetados. Animales. (2020) 10:757. doi: 10.3390/ani10050757
40. Li, Z, Wright, ADG, Si, H, Wang, X, Qian, W, Zhang, Z, et al. Los cambios en el microbioma ruminal y los metabolitos revelan el efecto de la genética del huésped en los cruces híbridos. Environ Microbiol Rep. (2016) 8:1016–23. doi: 10.1111/1758-2229.12482
41. Kreisinger, J, Ížková, D, Vohánka, J y Piálek, J. Microbiota gastrointestinal de individuos salvajes y endogámicos de dos subespecies de ratones domésticos evaluada mediante pirosecuenciación paralela de alto rendimiento. Mol Ecol. (2014) 23:5048. doi: 10.1111/mec.12909
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Wang, J, Kalyan, S, Steck, N, Turner, LM, Harr, B, Künzel, S, et al. El análisis de la microbiota intestinal en ratones domésticos híbridos revela una divergencia evolutiva en el hologenoma de un vertebrado. Nat Commun. (2015) 6:6440. doi: 10.1038/ncomms7440
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Goodrich, JK, Davenport, ER, Beaumont, M, Jackson, MA, Knight, R, Ober, C, et al. Determinantes genéticos del microbioma intestinal en gemelos del Reino Unido. Microbio huésped celular. (2016) 19:731–43. doi: 10.1016/j.chom.2016.04.017
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Kudo, H, Cheng, KJ, y Costerton, JW. Interacciones entre Treponema bryantii y bacterias celulolíticas en la degradación in vitro de celulosa de paja. Can J Microbiol. (1987) 33:244–8. doi: 10.1139/m87-041
45. Quan, J, Wu, Z, Ye, Y, Peng, L, Wu, J, Ruan, D, et al. Caracterización metagenómica de regiones intestinales en cerdos con eficiencia alimenticia contrastante. Microbiol frontal. (2020) 11:32. doi: 10.3389/fmicb.2020.00032
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Shinkai, T, Ikeyama, N, Kumagai, M, Ohmori, H, Sakamoto, M, Ohkuma, M, et al. Prevotella lacticifex sp. Nov., aislado del rumen de las vacas. Int J Syst Evol Microbiol. (2022) 72:5278. doi: 10.1099/ijsem.0.005278
47. Yang, H, Yang, M, Fang, S, Huang, X, He, M, Ke, S, et al. Evaluación del profundo efecto del microbioma intestinal en el apetito del huésped en cerdos. BMC Microbiol. (2018) 18:215. doi: 10.1186/s12866-018-1364-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Chen, C, Fang, S, Wei, H, He, M, Fu, H, Xiong, X, et al. Prevotella copri aumenta la acumulación de grasa en cerdos alimentados con dietas de fórmula. Microbioma. (2021) 9:175–5. doi: 10.1186/s40168-021-01110-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Pedersen, HK, Gudmundsdottir, V, Nielsen, HB, Hyotylainen, T, Nielsen, T, Jensen, BAH, et al. Los microbios intestinales humanos influyen en el metaboloma sérico del huésped y en la sensibilidad a la insulina. Nature (Londres). (2016) 535:376–81. doi: 10.1038/nature18646
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Xue, MY, Sun, HZ, Wu, XH, Liu, JX y Guan, LL. La multiómica revela que el microbioma del rumen y su metaboloma, junto con el metaboloma del huésped, contribuyen al rendimiento individualizado de las vacas lecheras. Microbioma. (2020) 8:64. doi: 10.1186/s40168-020-00819-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Pezeshki, A . El papel de la microbiota intestinal en la regulación del crecimiento y el metabolismo de los aminoácidos de cadena ramificada. J Anim Sci. (2024) 102:199. doi: 10.1093/jas/skae102.222
52. Chen, C, Huang, X, Fang, S, Yang, H, He, M, Zhao, Y, et al. Contribución de la genética del huésped a la variación de la composición microbiana del ciego, la luz y las heces en cerdos. Microbiol frontal. (2018) 9:2626. doi: 10.3389/fmicb.2018.02626
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Suen, G, Weimer, PJ, Stevenson, DM, Aylward, FO, Boyum, J, Deneke, J, et al. La secuencia completa del genoma de Fibrobacter succinogenes S85 revela un especialista celulolítico y metabólico. PLoS Uno. (2011) 6:E18814. doi: 10.1371/journal.pone.0018814
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Astuti, WD, Ridwan, R, Fidriyanto, R, Rohmatussolihat, R, Sari, NF, Sarwono, KA, et al. Cambios en la fermentación ruminal y perfiles bacterianos después de administrar Lactiplantibacillus plantarum como probiótico. Mundo Veterinario. (2022) 15:1969–74. doi: 10.14202/vetworld.2022.1969-1974
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Van Gylswyk, NO . Succiniclasticum ruminis gen. Nov., Sp. Nov., una bacteria ruminal que convierte el succinato en propionato como el único mecanismo de producción de energía. Int J Syst Bacteriol. (1995) 45:297–300. doi: 10.1099/00207713-45-2-297
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Li, Z, Shi, J, Lei, Y, Wu, J, Zhang, R, Zhang, X, et al. La castración altera la microbiota cecal e inhibe el crecimiento en el ganado Holstein. J Anim Sci. (2022) 100:100. doi: 10.1093/jas/skac367
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. Koistinen, KM, Suoniemi, M, Simolin, H y Ekroos, K. Lisofosfolipidómica cuantitativa en plasma y piel humanos por LC–MS/MS. Química bioanal anal. (2015) 407:5091–9. doi: 10.1007/s00216-014-8453-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Ukanović, N, Obradović, S, Zdravković, M, Urašević, S, Stojković, M, Tosti, T, et al. Lípidos y antiagregación plaquetaria: consideraciones importantes y perspectivas de futuro. Int J Mol Sci. (2021) 22:3180. doi: 10.3390/ijms22063180
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Inoue, N, Sakurai, T, Yamamoto, Y, Chiba, H y Hui, SP. Perfil de especies moleculares de lisofosfatidiletanolamina en suero humano y predicción in silico del sitio de unión a la albúmina. Biofactores. (2022) 48:1076–88. doi: 10.1002/biof.1868
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Hisano, K, Yoshida, H, Kawase, S, Mimura, T, Haniu, H, Tsukahara, T, et al. La abundante oleoil-lisofosfatidiletanolamina en el cerebro estimula el crecimiento de neuritas y protege contra la toxicidad del glutamato en las neuronas corticales cultivadas. J Biochem. (2021) 170:327–36. doi: 10.1093/jb/mvab046
61. Yamamoto, Y, Sakurai, T, Chen, Z, Inoue, N, Chiba, H y Hui, S. La lisofosfatidiletanolamina afecta la acumulación de lípidos y el metabolismo en una línea celular derivada del hígado humano. Nutrientes. (2022) 14:579. doi: 10.3390/nu14030579
62. Moskovitz, J y Smith, A. El sulfóxido de metionina y el sistema de metionina sulfóxido reductasa como moduladores de las vías de transducción de señales: una revisión. Aminoácidos. (2021) 53:1011–20. doi: 10.1007/s00726-021-03020-9
63. Sharov, VS y Schoneich, C. Oxidación diastereoselectiva de la proteína metionina por especies reactivas de oxígeno y reparación diastereoselectiva por metionina sulfóxido reductasa. Gratis Radic Biol Med. (2000) 29:986–94. doi: 10.1016/s0891-5849(00)00400-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
64. Li, H, Cai, L, Liang, M, Wang, Z, Zhang, Y, Wu, Q, et al. La metionina aumenta la capacidad antioxidante endógena de la proteína de arroz a través de la estimulación del sistema antioxidante MSR y la activación de la vía Nrf2-are en ratas adultas y en crecimiento. Eur Food Res Technol. (2020) 246:1051–63. doi: 10.1007/s00217-020-03464-5
65. De Freitas, DA, de Souza, KA, Alcalde, CR, Gasparino, E, y Feihrmann, AC. La suplementación con metionina libre o dipéptido de metionina mejora la calidad de la carne en pollos de engorde expuestos a estrés térmico. J Food Sci Technol. (2021) 58:205–15. doi: 10.1007/s13197-020-04530-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
66. Hu, H, Bai, X, Shah, AA, Wen, AY, Hua, JL, Che, CY, et al. La suplementación dietética con glutamina y ácido γ-aminobutírico mejora el rendimiento del crecimiento y los parámetros séricos en pollos de engorde de 22 a 35 días de edad expuestos a ambientes cálidos. J Anim Physiol Anim Nutr. (2016) 100:361–70. doi: 10.1111/jpn.12346
Resumen de PubMed | Texto completo de Crossref | Google Académico
67. Park, K, Oh, M, Joo, Y y Han, J. Efectos del ácido gamma aminobutírico en el rendimiento, células sanguíneas de pollos de engorde sometidos a entornos de estrés múltiple. Anim Biosci. (2023) 36:248–55. doi: 10.5713/ab.22.0031
68. Dai, SF, Gao, F, Zhang, WH, Song, SX, Xu, XL y Zhou, GH. Efectos de la glutamina dietética y el ácido gamma-aminobutírico sobre el rendimiento, las características de la canal y los parámetros séricos en pollos de engorde bajo estrés térmico circular. Anim Feed Sci Technol. (2011) 168:51–60. doi: 10.1016/j.anifeedsci.2011.03.005
69. Chen, Z, Xie, J, Wang, B y Tang, J. Efecto del ácido gamma-aminobutírico sobre las enzimas digestivas, la función de absorción y la función inmune de la mucosa intestinal en pollos estresados por calor. Poult Sci. (2014) 93:2490–500. doi: 10.3382/ps.2013-03398
70. Íñigo, M, Deja, S.A., y Burgess, C.C. Entresijos del ciclo del TCA: el papel central de la anaplerosis. Annu Rev Nutr. (2021) 41:19–47. doi: 10.1146/annurev-nutr-120420-025558
71. Kong, L, Yue, Y, Li, J, Yang, B, Chen, B, Liu, J, et al. La transcriptómica y la metabolómica revelan un mejor rendimiento de las ovejas Hu en la hibridación con las ovejas Southdown. Alimentos Res Int. (2023) 173:113240. doi: 10.1016/j.foodres.2023.113240
72. Oien, NL, Brideau, RJ, Hopkins, TA, Wieber, JL, Knechtel, ML, Shelly, JA, et al. Actividades antiherpes de amplio espectro de las carboxamidas de 4-hidroxiquinoleína, una nueva clase de inhibidores de la polimerasa del herpesvirus. Agentes antimicrobianos quimioterápicos. (2002) 46:724–30. doi: 10.1128/AAC.46.3.724-730.2002
Resumen de PubMed | Texto completo de Crossref | Google Académico
73. Schoeler, M, y Caesar, R. Lípidos dietéticos, microbiota intestinal y metabolismo lipídico. Rev Endocr Metab Dis. (2019) 20:461–72. doi: 10.1007/s11154-019-09512-0
74. Brown, EM, Clardy, J y Xavier, RJ. Metabolismo lipídico del microbioma intestinal y su impacto en la fisiología del huésped. Microbio huésped celular. (2023) 31:173–86. doi: 10.1016/j.chom.2023.01.009
Palabras clave: hibridación, rumen, metagenoma, rendimiento de crecimiento, ovejas
Cita: Zhang R, Zhang L, An X, Li J, Niu C, Zhang J, Geng Z, Xu T, Yang B, Xu Z y Yue Y (2024) La hibridación promueve el rendimiento del crecimiento al alterar la microbiota ruminal y los metabolitos en las ovejas. Frente. Vet. Sci. 11:1455029. doi: 10.3389/fvets.2024.1455029
Editado por:
Qingbiao Xu, Universidad Agrícola de Huazhong, China
Revisado por:
Kaizhen Liu, Universidad Agrícola de Henan, China
Shaobin Li, Universidad Agrícola de Gansu, China
Derechos de autor © 2024 Zhang, Zhang, An, Li, Niu, Zhang, Geng, Xu, Yang, Xu y Yue. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Atribución (CC BY).
*Correspondencia: Zhenfei Xu, 236xuzhenfei@163.com; Yaojing Yue, yueyaojing@caas.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía