
La ración mezclada total fermentada mejora la digestibilidad de los nutrientes y modula los componentes de la leche
 La ración mezclada total fermentada mejora la digestibilidad de los nutrientes y modula los componentes de la leche y la comunidad microbiana fecal en vacas lecheras Holstein lactantes
La ración mezclada total fermentada mejora la digestibilidad de los nutrientes y modula los componentes de la leche y la comunidad microbiana fecal en vacas lecheras Holstein lactantes
 Lijun Wang1
Lijun Wang1 Sanjun Jin1
Sanjun Jin1 Ping Wang1
Ping Wang1 Xinxin Li1Chaoqi Liu1Siying Sun2Guangning Zhang3
Xinxin Li1Chaoqi Liu1Siying Sun2Guangning Zhang3 Juan Chang1
Juan Chang1 Qingqiang Yin1*Haiyang Zhang1*Qun Zhu4
Qingqiang Yin1*Haiyang Zhang1*Qun Zhu4- 1Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Henan, Zhengzhou, China
- número arábigoInstituto de Educación Internacional, Universidad Agrícola de Henan, Zhengzhou, China
- 3Facultad de Ciencia y Tecnología Animal, Universidad Agrícola del Noreste, Harbin, China
- 4Henan Delin Productos Biológicos Co., Ltd., Xinxiang, China
La ración mezclada total fermentada (FTMR) es un método eficaz para preservar subproductos de alta humedad con mayor estabilidad aeróbica después de la fermentación. La FTMR tiene el potencial de satisfacer las necesidades nutricionales diarias del ganado y mejorar su rendimiento productivo. El objetivo de esta investigación fue examinar la influencia de la FTMR en el rendimiento de la lactancia, la digestibilidad aparente del tracto total, las comunidades de microbiota fecal y los perfiles de fermentación en vacas lecheras lactantes. Un total de 12 vacas fueron asignadas aleatoriamente a dos grupos: el grupo TMR y el grupo FTMR. El grupo de TMR fue alimentado con una dieta de ración total mixta (TMR), y el grupo de FTMR fue alimentado con una dieta de FTMR. El FTMR no afectó la producción de leche en las vacas lecheras a pesar de una disminución en la ingesta de materia seca, lo que aumentó la eficiencia del alimento. A diferencia del grupo TMR, el contenido de grasa láctea en el grupo FTMR fue mayor. El grupo FTMR mostró una mayor digestibilidad de la fibra detergente neutro (FDN), la materia orgánica (MO), la materia seca (MS), la proteína bruta (PC) y la fibra detergente ácida (FDA) en el tracto digestivo total que el grupo TMR. La FTMR aumentó la concentración de butirato en la materia fecal y redujo el pH de las heces. Los índices de Chao1, ACE y Shannon de la comunidad de arqueas en las heces de las vacas lecheras fueron significativamente más altos en las vacas alimentadas con la FTMR en comparación con las alimentadas con la TMR. El análisis de LefSe reveló niveles más altos de Oscillospira, Lactobacillus, Prevotella y Dehalobacterium en las heces de las vacas lecheras alimentadas con FTMR que en las alimentadas con TMR. Sin embargo, las abundancias de Roseburia, rc4-4, Bulleidia y Sharpea mostraron la tendencia opuesta. Las abundancias de Halobacterias, Halobacteriales y Halobacteriaceae, que son biomarcadores para distinguir las arqueas fecales en la TMR de la FTMR, fueron sustancialmente mayores en las heces de las vacas lecheras que consumieron la TMR que en las que consumieron la FTMR. Por lo tanto, la FTMR puede mejorar el contenido de grasa de la leche, la eficiencia de la digestibilidad aparente del alimento en el tracto total y la diversidad de arqueas en las heces. Además, este trabajo proporciona una base teórica para la factibilidad de la alimentación con FTMR para vacas lecheras.
1 Introducción
Uno de los principales desafíos a los que se enfrenta la industria ganadera es la escasez de recursos proteicos y forrajeros de alta calidad en China (1). A pesar de la presencia de abundantes recursos de piensos no convencionales y residuos de cultivos en China, sus tasas de digestión y utilización como recursos de piensos siguen siendo bajas (2). La tecnología de ración mezclada total fermentada (FTMR) representa un enfoque de alimentación novedoso desarrollado a partir de la modulación de TMR y la tecnología de procesamiento (3). Utiliza eficazmente los recursos alimentarios no convencionales y los residuos de cultivos como fuentes de piensos, minimizando el desperdicio, lo que ayuda a mitigar la escasez de recursos alimentarios y a reducir los costos de reproducción (4). Una dieta de alta calidad con una nutrición completa y equilibrada y un almacenamiento a largo plazo se logra mezclando y fermentando minuciosamente varios materiales de alimentación, incluidos forraje, concentrado y minerales esenciales, con bacterias de fermentación (5). Después de la fermentación, el alimento demuestra altos niveles de proteína bruta, excelente palatabilidad y un valor nutritivo significativo (6, 7). FTMR puede utilizar eficazmente los recursos de piensos no convencionales y los residuos de cultivos como fuentes de piensos para reducir el desperdicio, aliviando así la escasez de recursos de piensos y reduciendo los costes de cría.
Debido a las intrincadas condiciones de la fermentación anaeróbica en FTMR, su composición microbiana es muy diversa. Por lo tanto, la composición microbiana del alimento juega un papel crítico no solo en la calidad de la fermentación, sino también en la estabilidad aeróbica del alimento (8). Con los avances en la tecnología microbiana aplicada en la industria de los piensos, el uso de enzimas y bacterias se ha convertido en una estrategia común para mejorar la calidad de los piensos para ensilaje (9). Las combinaciones óptimas de enzimas y bacterias pueden estimular eficazmente la fermentación de los materiales de los piensos, mejorando tanto la calidad como la eficiencia de la alimentación (10-12). La celulasa descompone la celulosa, la hemicelulosa y la lignina en el alimento en azúcares accesibles que las bacterias del ácido láctico pueden utilizar, aumentando su sustrato de fermentación, acelerando la reducción del pH y mejorando la calidad de la fermentación del ensilado (13, 14). Lactobacillus buchneri, una bacteria de fermentación heterotípica, no solo mejora la estabilidad aeróbica y prolonga la vida útil, sino que también mejora la salud intestinal y la inmunidad de los animales, y aumenta la producción de leche en las vacas (15).
En los rumiantes, los microbios gastrointestinales de las vacas lecheras son fundamentales para adquirir nutrientes y energía (16, 17). La investigación ha demostrado que la dieta determina en gran medida la composición de las bacterias en las heces y la estructura de la comunidad de arqueas en las vacas lecheras, y el microbioma fecal sirve como indicador de los cambios microbianos del intestino posterior (18-20). Las dietas altas en concentrados disminuyen significativamente las poblaciones de bacterias celulolíticas como Ruminococcus, Fibrobacter y Ruminiclostridium, así como metanógenos como Methanosarcina, Methanobrevibacter y Methanosphaera (21). Al-Azzawi et al. demostraron que la adición de carbón activado en polvo a la dieta reduce la abundancia de Proteobacteria y Methanobrevibacter en las heces (21). Además, las arqueas del intestino posterior son responsables de la producción de metano, que contribuye significativamente al calentamiento global (22-24). Sin embargo, existe información limitada sobre los efectos de la TMR de cofermentación con Lactobacillus brucei y celulasa sobre los parámetros de fermentación fecal y la diversidad microbiana. Este estudio tuvo como objetivo evaluar y comparar los impactos de las raciones mezcladas totales (TMR) y las raciones mezcladas totales fermentadas (FTMR) en el rendimiento de la lactancia, los parámetros de fermentación fecal y la microbiota fecal en el ganado lechero, proporcionando información valiosa sobre el uso de FTMR para el manejo de las vacas lecheras.
2 Materiales y métodos
2.1 Animales y dietas
Todos los procedimientos experimentales con animales fueron aprobados por el Comité de Cuidado y Uso de Animales de la Universidad Agrícola de Henan (número de aprobación: HENAU-2021-025).
Las vacas lecheras experimentales se obtuvieron de la granja lechera Ruiya ubicada en Zhengzhou, Henan, China. Un total de doce vacas Holstein de parto 2 (peso corporal promedio = 616 ± 13,4 kg, período de lactancia promedio = 106 ± 7,55 días) fueron asignadas aleatoriamente a los grupos de ración mezclada total (TMR, n = 6) o ración mezclada total fermentada (FTMR, n = 6) en función de su producción diaria de leche y período de lactancia. Cada grupo incluyó 6 réplicas (corrales) con una vaca por corral. El estudio se llevó a cabo durante un período de 14 semanas, con las dos semanas iniciales designadas como período de adaptación.
Tanto las dietas TMR como FTMR consistieron en 50% de forraje y 50% de concentrado en base a materia seca, formuladas para satisfacer los requerimientos nutricionales utilizando el modelo Cornell-Penn-Miner Dairy versión 3.08.01. Las composiciones de TMR y FTMR fueron idénticas (en base a materia seca): heno de alfalfa (11.80%), alimento húmedo de gluten de maíz (6.86%), rastrojo de maíz (3.43%), ensilaje de maíz (27.80%), maíz molido (24.60%), harina de soya (9.80%), harina de semilla de algodón (4.91%), DDGS (7.35%), soya expandida (0.95%) y premezcla (2.50%). El contenido de humedad se ajustó a 48.0%. Se añadieron bacterias lácticas (BAL) y celulasa al FTMR. El BAL incluyó una mezcla de Lactobacillus plantarum (CGMCC 1.12934, obtenida del Centro General de Recolección de Cultivos Microbiológicos de China) y Lactobacillus brucei (BNCC189797, obtenida de Beina Bio, Beijing, China) en una proporción de 1:1, aplicada a 1 × 1011 ufc/g de material fresco. Se añadió celulasa (10.000 U/g, XS Biotechnology Co., Ltd., Pekín, China) a 10 g/kg de material fresco. Los aditivos se disolvieron en agua y se rociaron uniformemente sobre la mezcla con un rociador. La mezcla fermentada se envasó con film estirable de polietileno con una compactación de 800cm3 y se fermentó al aire libre durante 60 días a 16-30°C utilizando una empacadora de ensilaje (Takakita MW1010H, Japón). En la Tabla 1 se presentan las composiciones de nutrientes de ambos alimentos. Las vacas fueron alimentadas ad libitum dos veces al día a intervalos de 12 h (6:30 y 18:30) con libre acceso al agua en establos individuales. Las vacas lecheras fueron ordeñadas dos veces al día a las 6:00 y a las 18:00.
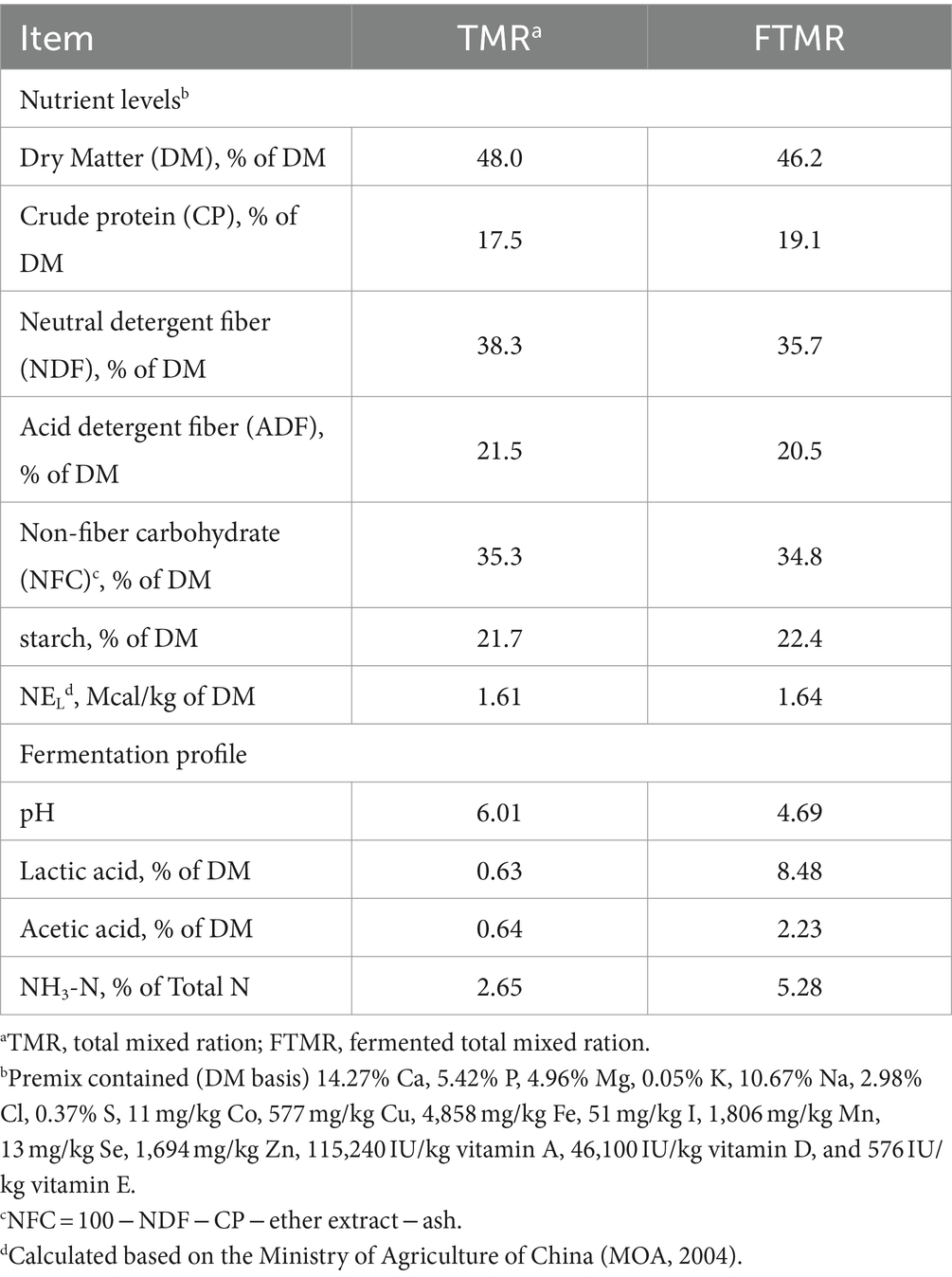
2.2 Análisis de la producción de leche y sus componentes
En las semanas 7 y 13 del experimento, se registró la producción de leche y se recolectaron muestras de leche durante 6 días consecutivos. Cada día, en el Centro de Pruebas de Mejora del Rebaño Lechero de Henan en Zhengzhou, China, se mezcló una alícuota de 50 mL de leche, proporcional a la producción diaria real (mañana y tarde), utilizando un analizador de leche automatizado de infrarrojo cercano (MilkoScan, Foss Electric, Hillerød, Dinamarca). Se añadió dicromato de potasio para conservar las muestras de leche, que se almacenaron a 4°C hasta el análisis. La lactosa, la proteína, la grasa, el nitrógeno ureico de la leche (MUN), los sólidos totales (TS) y el recuento de células somáticas (SCC) se determinaron utilizando los métodos de análisis infrarrojo descritos por Laporte y Paquin (25).
2.3 Recogida de muestras fecales
Durante las semanas 7 y 13 del experimento, se recogieron unos 500 g de muestras fecales puntuales de los rectos de las vacas utilizando guantes estériles. Estas muestras se compusieron para cada vaca a las 6:00 y a las 18:00 durante tres días consecutivos. Para medir el pH, se mezclaron 5 g de material fecal con 250 mL de agua destilada, y el pH se midió rápidamente con un medidor de pH portátil (PHB-5, ShanghaiLeici, Shanghai, China).
Se utilizó un conjunto de muestras para la digestibilidad aparente del tracto total. Las muestras fecales se secaron a 60 °C en un horno de aire forzado y luego se molieron con una pantalla de 1 mm en una máquina trituradora de microplantas (FZ102, Taisite Instrument Co., Ltd., Tianjin, China). Posteriormente, se analizaron la materia seca (MS), la proteína cruda (PC), las cenizas, el almidón, la fibra detergente neutra (FDN), la fibra detergente ácida (FDA) y la FDN no digerible (FNDi). El otro conjunto de muestras se utilizó para el análisis de microorganismos fecales. Las muestras se congelaron inmediatamente con nitrógeno líquido. Se combinaron cantidades iguales de muestras congeladas de cada vaca y punto de tiempo para garantizar la uniformidad, utilizando un homogeneizador de grifo estéril (Shanghai Hannuo Ltd., Shanghai, China). Estas muestras mezcladas se almacenaron a -80 °C para minimizar la actividad microbiana, preparándolas para la posterior extracción de ADN.
2.4 Análisis de muestras fecales
Las muestras de alimento se analizaron para cenizas, materia seca (MS), proteína bruta (PC) y almidón siguiendo los procedimientos 942.05, 934.01, 976.05 y 982.30, respectivamente, según lo descrito por la Asociación de Químicos Analíticos Oficiales (AOAC, 1990). Las concentraciones de fibra detergente neutro (FDN) y fibra detergente ácida (FDA) se determinaron consecutivamente utilizando un analizador de fibra Ankom A200 (Ankom Technology, Macedonia, NY). La FDN no digerible (FDNi) sirvió como marcador indirecto, y la digestibilidad aparente de los nutrientes del tracto total se calculó en consecuencia. El marcador iNDF se identificó a través del análisis in vitro descrito por Goeser y Combs (26). Para las muestras fecales, la materia prima se diluyó 1:4 con agua destilada para medir el pH fecal utilizando un medidor de pH Accumet AB150 (Fisher, Canadá).
El contenido de ácidos grasos volátiles (AGV) fecales se determinó mediante cromatografía líquida de alta resolución (HPLC, Waters 600, Milford, Massachusetts, Estados Unidos). La preparación de cada muestra consistió en diluir 1 g de material fecal con 1 mL de agua. A continuación, se añadieron 300 μL de un patrón interno (ácido 4-metilvalérico, Sigma-Aldrich, St. Louis, MO) y 200 μL de ácido fosfórico al 25%, se mezclaron bien y luego se centrifugaron a 12.000 × g durante 15 min a 4 °C. El sobrenadante resultante se transfirió a un nuevo tubo para su análisis mediante HPLC, siguiendo el método detallado por Wang et al. (27).
2.5 Puntuación de la condición corporal
Durante las semanas 7 y 13 del experimento, los veterinarios evaluaron la puntuación de la condición corporal (BCS) de las vacas experimentales poco después de la sesión de ordeño matutina. La evaluación utilizó una escala de 5 puntos (1-5 puntos, con incrementos de 0,25) según lo descrito por Vasseur et al. (28).
2.6 Amplificación de ARNr 16S y secuenciación de Illumina MiSeq
Para la extracción de ADN genómico de muestras fecales se utilizó el TIANamp Bacteria DNA Kit (TIANGEN, Pekín, China) siguiendo el protocolo del fabricante. La verificación de la concentración e integridad del ADN se llevó a cabo utilizando electroforesis en gel de agarosa y un espectrofotómetro NanoDrop 2000 (Thermo Fisher Scientific, Massachusetts, Estados Unidos). Posteriormente, el ADN se amplificó por triplicado empleando el sistema de ADN polimerasa de alta fidelidad Q5 (New England Biolabs (Beijing) LTD, Beijing, China). La región V3-V4 del gen 16S rRNA para el análisis bacteriano se amplificó utilizando los cebadores 338F (5′-ACTCCTRCGGGAGGCAGCAGCAG-3′) y 806R (5′-GGACTACCVGGGTATCTAAT-3′) (23), mientras que la región V3-V4 del gen 16S rRNA para el análisis de arqueas se amplificó utilizando los cebadores 524F (5′-TGYCAGCCGCCGCGGCGGTAA-3′) y 958R (5′-YCCGGCGTTGAVTCCAATT-3′). La amplificación por PCR consistió en una desnaturalización inicial a 95 °C durante 3 min, seguida de 25 ciclos de desnaturalización a 95 °C, recocido a 60 °C para bacterias/55 °C para arqueas, y elongación a 72 °C, concluyendo con un paso final de extensión a 72 °C durante 10 min. Los productos de PCR se analizaron mediante electroforesis en gel de agarosa al 2%. La purificación utilizó el kit de extracción de gel de ADN AxyPrep (Axygen Bioscience, Union City, CA, Estados Unidos), seguido de la cuantificación mediante el kit de ensayo Quant-iT PicoGreen dsDNA (Thermo Fisher Scientific, Massachusetts, Estados Unidos). Las bibliotecas de secuenciación se validaron utilizando un Agilent Bioanalyzer (Agilent Technologies, Palo Alto, CA, Estados Unidos), y su cuantificación fue confirmada por un Agilent Bioanalyzer (Agilent Technologies, Palo Alto, CA, Estados Unidos) y Promega QuantiFluor-ST™ (Promega, Madison, WI, Estados Unidos). La secuenciación fue realizada por Frasergen Bioinformatics Technology Co., Ltd. (Wuhan, China) en una plataforma Illumina MiSeq (Illumina, Inc., San Diego, CA, Estados Unidos), y los datos originales de RNA-seq se han depositado en el NCBI Sequence Read Archive (SRA), número de acceso: PRJNA1119071.
2.7 Análisis estadísticos y bioinformáticos
El procesamiento de secuencias de la plataforma MiSeq se realizó utilizando QIIME (versión 1.8.0) (29). Se mantuvieron las lecturas que cumplían con los criterios de puntuación de calidad promedio ≥ 25 y una longitud entre 220 y 250 nt. Las secuencias superpuestas (superposición de >10 pb) se ensamblaron utilizando FLASH v1.2.7. Para obtener etiquetas limpias de alta calidad, las lecturas en bruto se sometieron a condiciones de filtrado específicas a través del control de calidad QIIME (v1.8.0): se excluyeron las secuencias ≤160 pb o con homopolímeros de ≥8 pb. Las unidades taxonómicas operacionales (OTUs) se agruparon con una identidad del 97% utilizando UCLUST (30), y las secuencias quiméricas se eliminaron con USEARCH (v5.2.236, http://www.drive5.com/usearch/). La secuencia más prevalente dentro de cada OTU (bacterias y arqueas) se definió como la «secuencia representativa» y se alineó con la base de datos de bacterias SILVA (versión 119) (31), la base de datos de protozoos NCBI-nt (32), la base de datos ITS de hongos Unite (versión 7.0) (33) y la base de datos de arqueas SILVA (31) utilizando PyNAST (29) con parámetros estándar. Los índices de diversidad alfa (ACE, Chao1, Shannon, Simpson) se calcularon mediante muestras enrarecidas en QIIME para evaluar la diversidad y abundancia de las comunidades bacterianas y de arqueas. Se utilizó el análisis de componentes principales (PCA) para la evaluación de la diversidad beta. El análisis del tamaño del efecto del análisis discriminante lineal (LefSe) en la plataforma en línea Galaxy (34) identificó biomarcadores funcionales discriminativos, empleando un umbral de tamaño-efecto de 2,0 en la puntuación logarítmica LDA para determinar los principales módulos abundantes en los grupos TMR y FTMR.
Se utilizaron pruebas t de muestras independientes (para datos distribuidos normalmente) o pruebas U de Mann-Whitney (para datos distribuidos no normalmente) para determinar diferencias sustanciales en la abundancia relativa de los 10 filos y géneros principales y en los índices de diversidad alfa entre los dos grupos. P < 0,05 y p < 0,01 indicaron diferencias estadísticamente significativas y extremadamente significativas, respectivamente.
3 Resultados
3.1 Efecto de la TMR y la FTMR en el rendimiento de la lactancia de las vacas lecheras
En la Tabla 2 se presentan los resultados de la producción de leche y de los componentes, además de la ingesta de alimento. Las vacas lecheras que consumieron la dieta FTMR mostraron una DMI reducida, una mayor concentración de grasa láctea, una ECM más alta y una mejor eficiencia alimenticia en comparación con las que seguían la dieta TMR. Se observaron diferencias significativas en la eficiencia alimenticia entre los grupos TMR y FTMR.
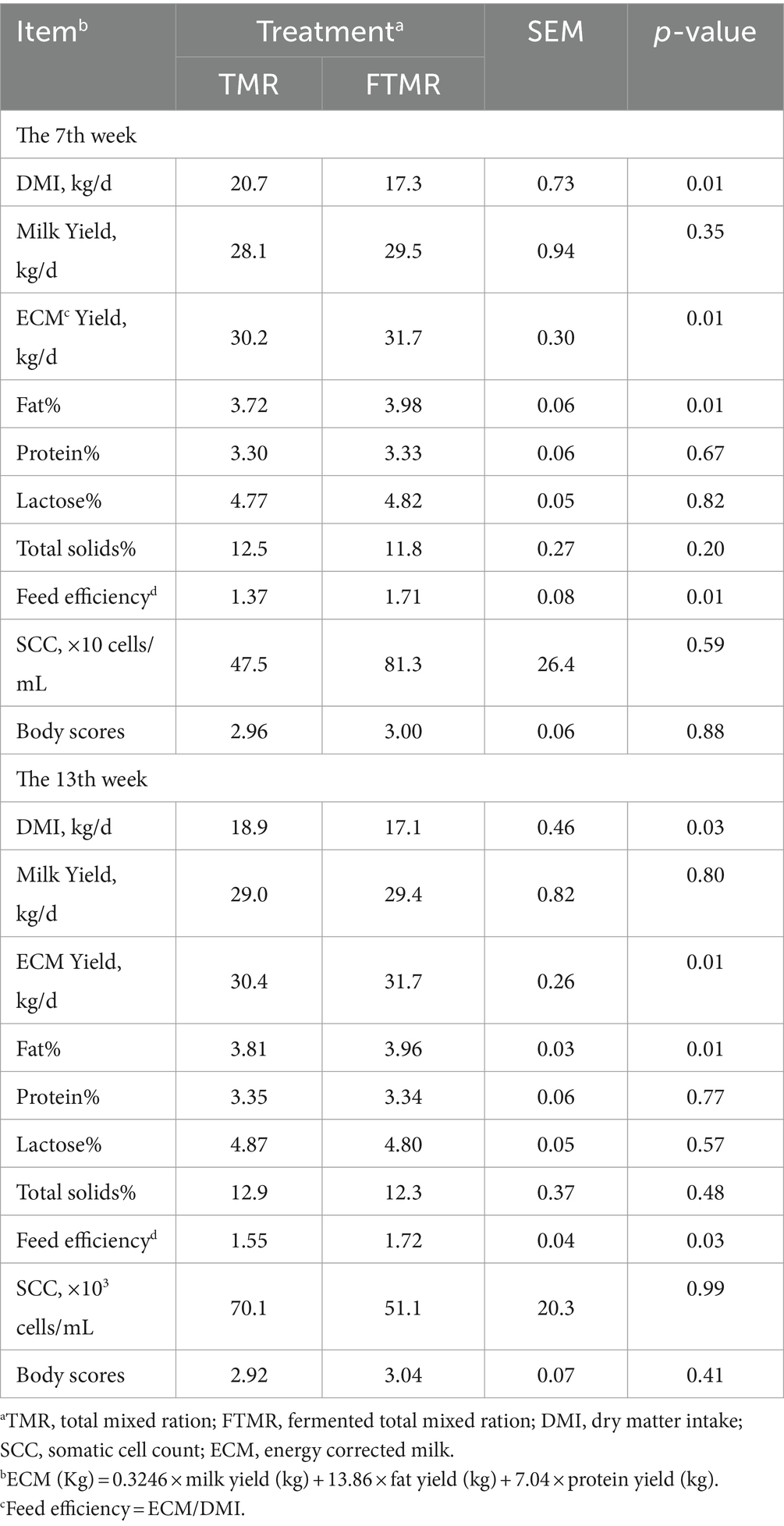
3.2 Efecto de TMR y FTMR en la digestibilidad aparente del tracto total de las heces de vacas lecheras
La Tabla 3 muestra la digestibilidad aparente de los nutrientes del tracto total de las dos dietas de tratamiento, que fue mayor para la dieta FTMR que para la dieta TMR para DM, OM, PC, NDF y ADF.
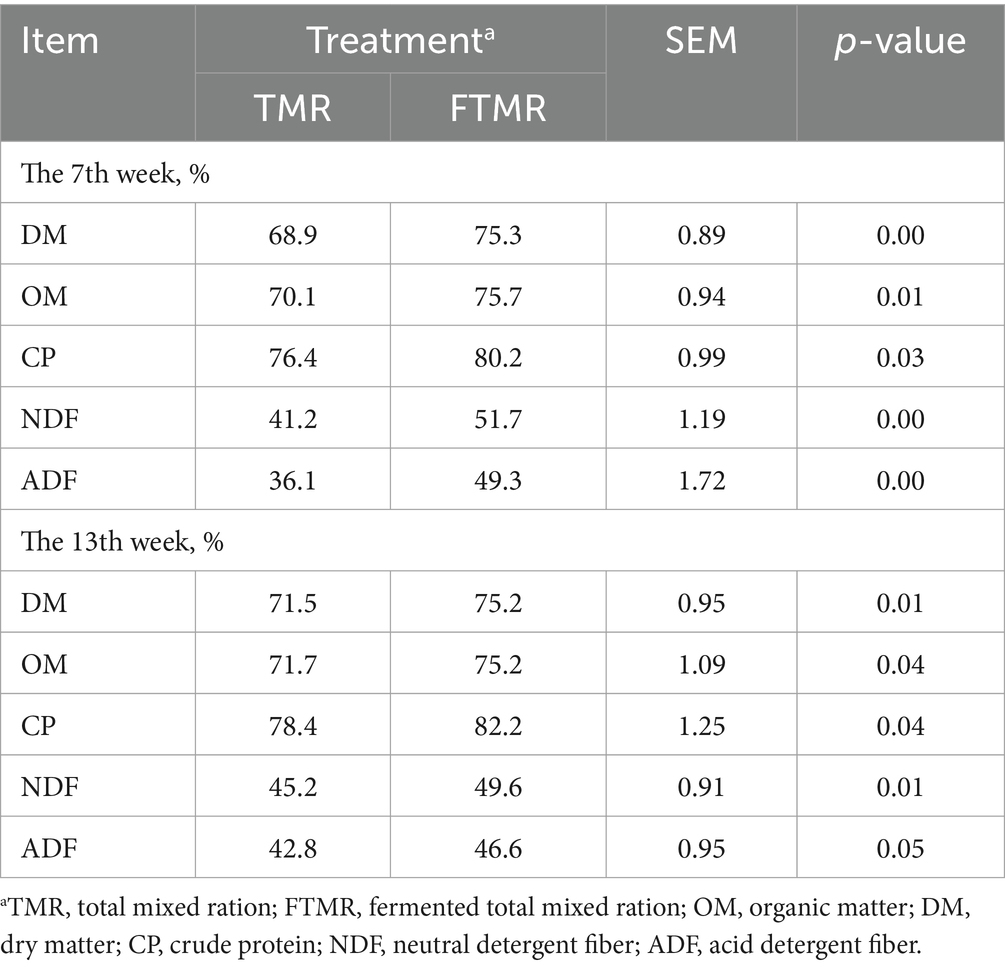
3.3 Efecto de TMR y FTMR sobre el patrón de AGV fecal y el pH de las vacas lecheras
El pH fecal, acetato y propionato fueron menores (p < 0,05) en las muestras tratadas con FTMR que en las muestras tratadas con TMR (Tabla 4). El contenido de butirato en las heces de las vacas alimentadas con FTMR fue mayor que el de las heces de las vacas alimentadas con TMR. No se observaron efectos significativos para la concentración total de AGV en heces, el isobutirato, el valerato, el isovalerato y la relación acetato/propionato.
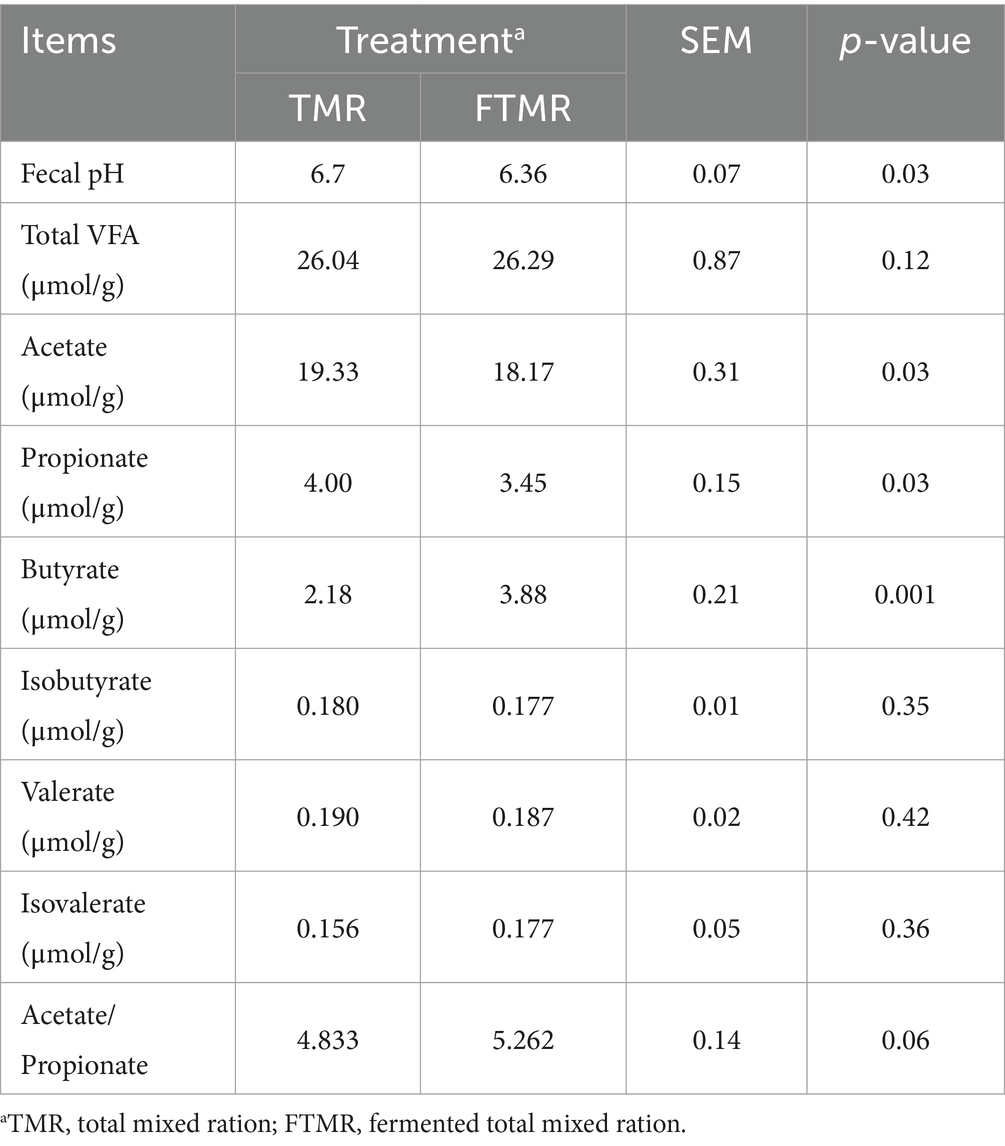
3.4 Análisis de la secuenciación del ARNr 16S de bacterias y arqueas en las heces de vacas lecheras alimentadas con TMR y FTMR
La Figura 1A muestra las especies bacterianas (OTU) que se encuentran en TMR y FTMR, y la Figura 1C ilustra las especies de arqueas (OTU). Las vacas alimentadas con FTMR exhibieron significativamente más especies observadas que las alimentadas con TMR (p < 0.05).
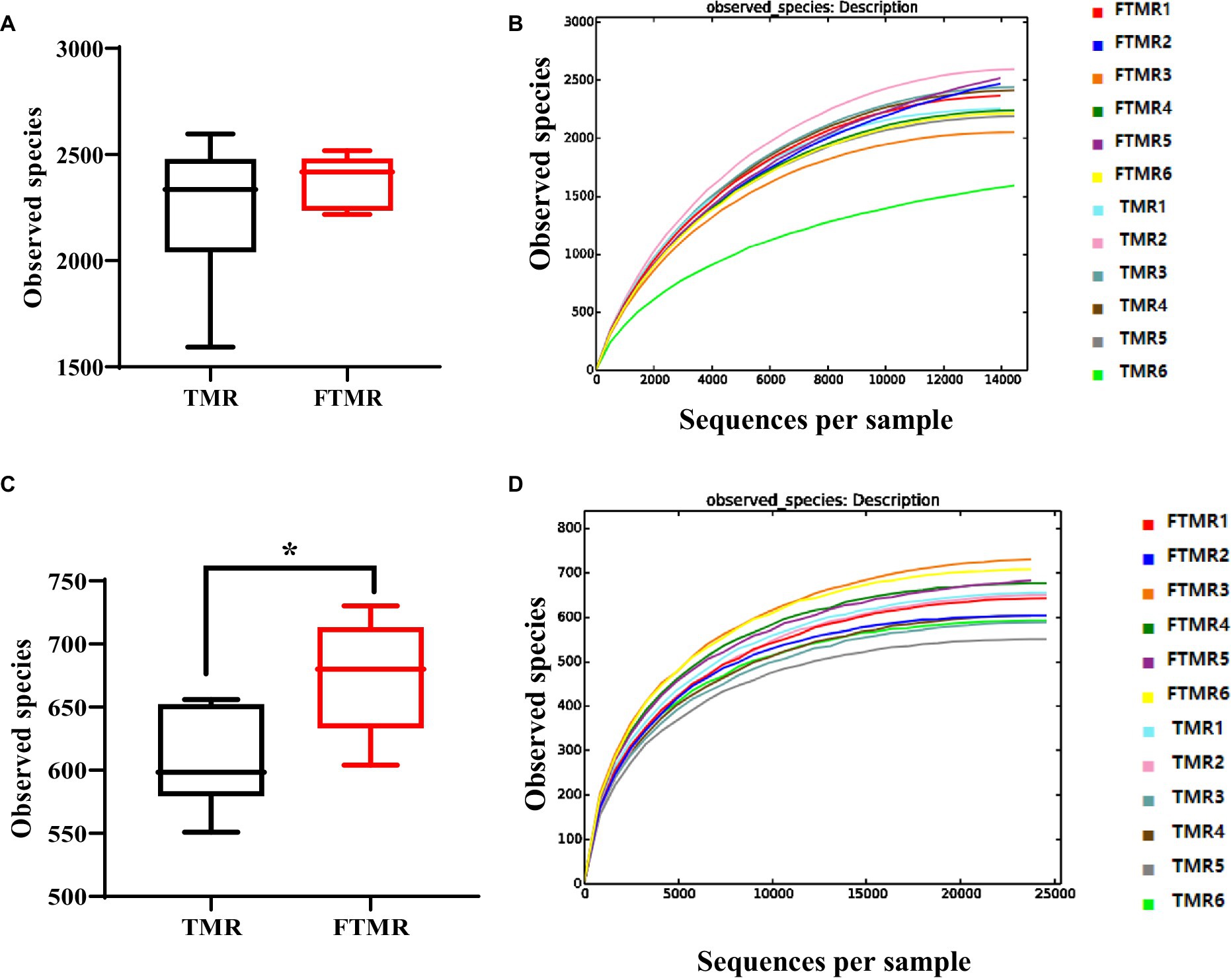
Las curvas de dispersión de las OTUs identificadas en este estudio indicaron que a medida que aumentaba la profundidad de secuenciación, se detectaban más especies. Con un mayor número de secuencias analizadas, los bordes de las curvas de dispersión se aplanaron, lo que sugiere una cobertura completa de los datos de secuenciación (Figuras 1B, D). La cobertura de Good, que evalúa qué tan bien se representan las muestras mediante la secuenciación, se acercó a casi el 99%, lo que indica una detección completa de los tipos bacterianos en las muestras.
3.5 Diferencias en la diversidad bacteriana y arquearia entre las heces de vacas lecheras alimentadas con TMR y FTMR
Los índices de diversidad alfa comúnmente empleados incluyeron índices de abundancia (Chao1 y ACE) e índices de diversidad (Simpson y Shannon). El análisis de las bacterias fecales entre los dos grupos no indicó diferencias estadísticamente significativas en los índices Chao1, ACE, Shannon o Simpson, como se indica en la Tabla 5 (p > 0,05). Además, la RMFT incrementó significativamente los índices de abundancia de arqueas fecales (índices Chao y ACE) y los índices de diversidad (índice de Shannon) (p < 0,05).
Se utilizaron evaluaciones de diversidad beta para analizar las similitudes en la estructura de la comunidad entre los dos grupos. El análisis de NMDS puso de manifiesto diferencias significativas en las comunidades bacterianas fecales, distinguiendo claramente entre los grupos TMR y FTMR a nivel de OTU. De manera similar, los resultados del NMDS mostraron una variabilidad sustancial en las comunidades de arqueas fecales, con una separación considerable observada entre los dos grupos, lo que indica composiciones distintas de comunidades de arqueas en los grupos TMR y FTMR (Ver Figura 2).
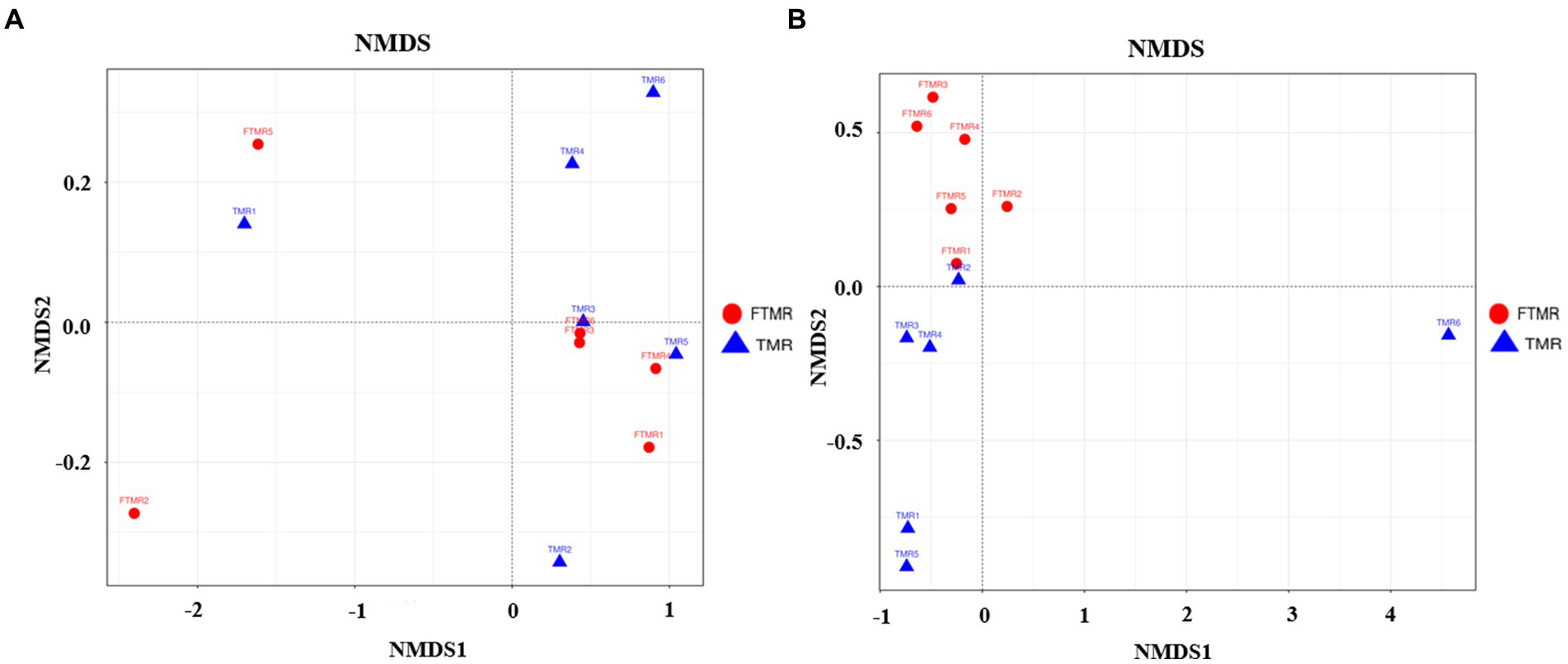
Figura 2. NMDS de las comunidades microbianas fecales. NMDS ponderado por bacterias fecales (A) y arqueas (B). TMR: ración total mezclada; FTMR: ración mezclada total fermentada.
3.6 Abundancia relativa y microbiota central de bacterias y arqueas en las heces de vacas lecheras alimentadas con TMR y FTMR
La Figura 3 muestra las distribuciones de los 10 y 20 principales filos y géneros de bacterias fecales en los grupos TMR y FTMR. Se consideraron predominantes los filos bacterianos con una abundancia relativa superior al 1%. A nivel de filo (Figura 3A), Firmicutes, Bacteroidetes y Actinobacteria fueron prevalentes en ambos grupos. Entre los géneros con una abundancia relativa superior al 1% (Figura 3B), las bacterias dominantes incluyeron Unclassified_Ruminococcaceae, Unclassified_Bacteroidales, Unclassified_Clostridiales, Oscillospira, Unclassified_Peptostreptococcaceae, Unclassified_Lachnospiraceae, Bifidobacterium, Unclassified_Rikenellaceae, Unclassified_S24-7, Dorea, Unclassified_RF16, CF231, Clostridium y Unclassified_Clostridiaceae. Oscillospira mostró una diferencia notable entre los grupos TMR y FTMR (p = 0,01) (Figura 3E).
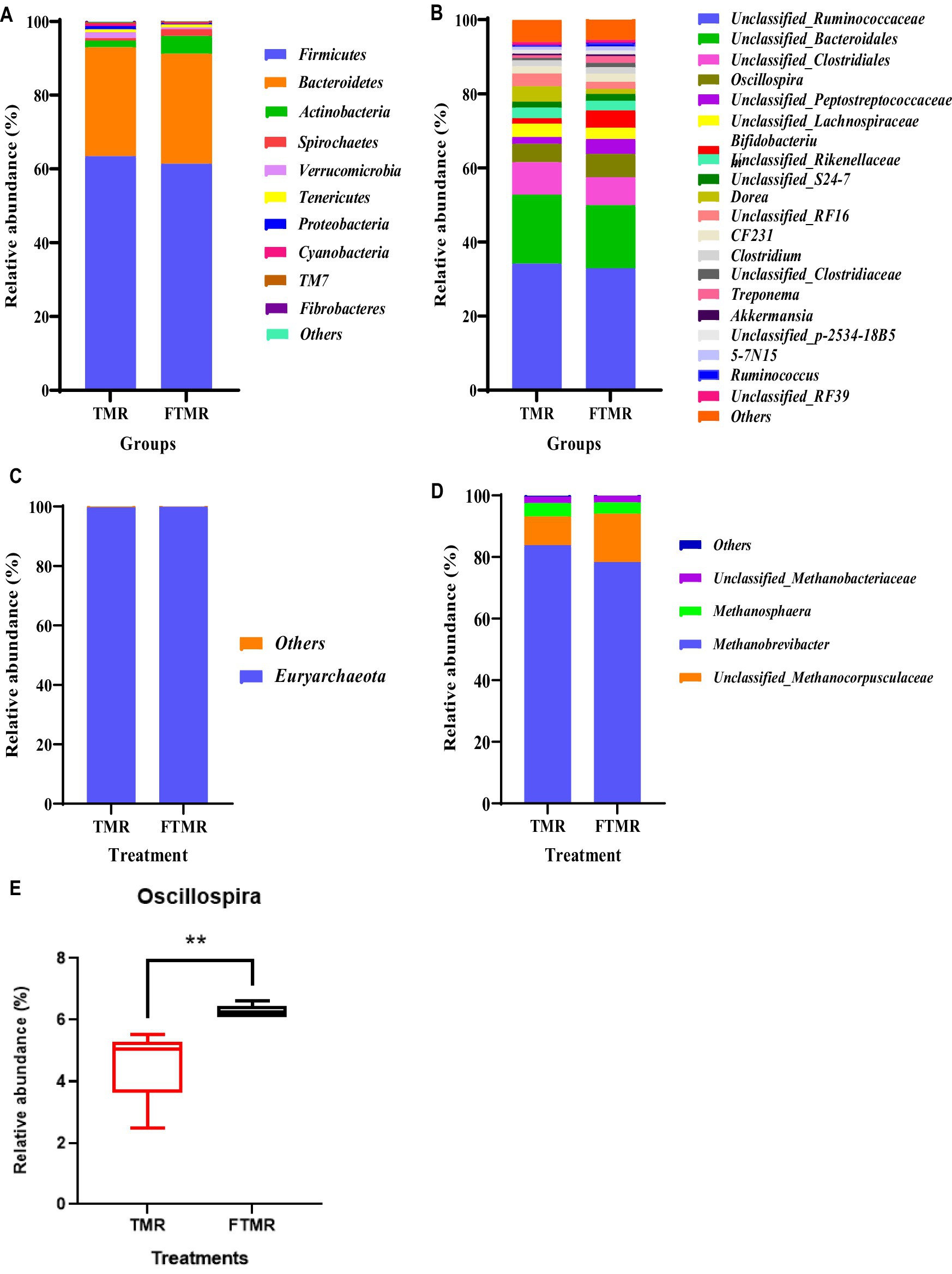
Figura 3. Abundancia relativa de microbios fecales a nivel de filo y género. La abundancia de filos de bacterias (A) y arqueas (C). La abundancia de géneros de bacterias (B) y arqueas (D). Género Oscillospira (E) entre ambos grupos. Los diagramas de caja pequeños muestran los percentiles 25, 50 y 75, y los bigotes muestran los valores extremos de los datos. TMR: ración total mezclada; FTMR, ración mixta fermentada.
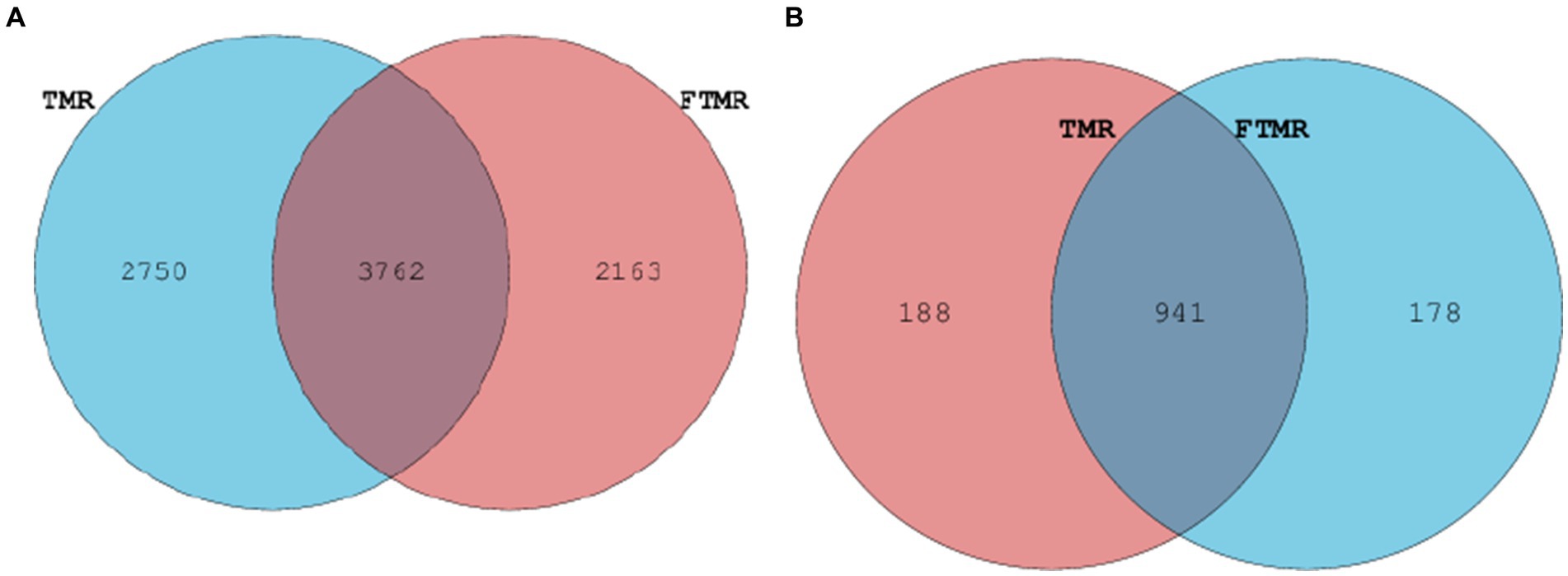
En la Figura 3 se presentan las abundancias relativas de arqueas fecales en los grupos TMR y FTMR tanto a nivel de filo como de género. Los filos de arqueas con una abundancia relativa superior al 1% se clasificaron como dominantes. A nivel de filo (Figura 3C), Euryarchaeota fueron las arqueas más prevalentes en ambos grupos. Entre los géneros de arqueas con abundancias relativas superiores al 1% (Figura 3D), las arqueas dominantes incluyeron Methanobrevibacter, Unclassified_Methanocorpusculaceae, Methanosphaera y Unclassified_Methanobacteriaceae. Los taxones compartidos entre los grupos TMR y FTMR se consideraron parte de la comunidad microbiana central. El número de OTU compartidas para las bacterias (Figura 4A) fue de 3.762, y para las arqueas (Figura 4B), fue de 947.
3.7 Análisis del tamaño del efecto LDA entre los grupos TMR y FTMR
Las clasificaciones de los módulos de enriquecimiento se determinaron mediante el análisis LEfSe. El cladograma (Figura 5A) confirmó visualmente las diferencias en 16 taxones bacterianos entre los grupos TMR y FTMR. La Figura 5B, el gráfico LEfSe, muestra las diferentes puntuaciones de LDA de los taxones bacterianos entre estos grupos. Los biomarcadores significativos para las bacterias incluyeron Roseburia, rc4-4, Bulleidia y Sharpea a nivel de género. Del mismo modo, los biomarcadores notables para las arqueas entre los grupos fueron Lactobacillus, Prevotella, Dehalobacterium y Oscillospira.
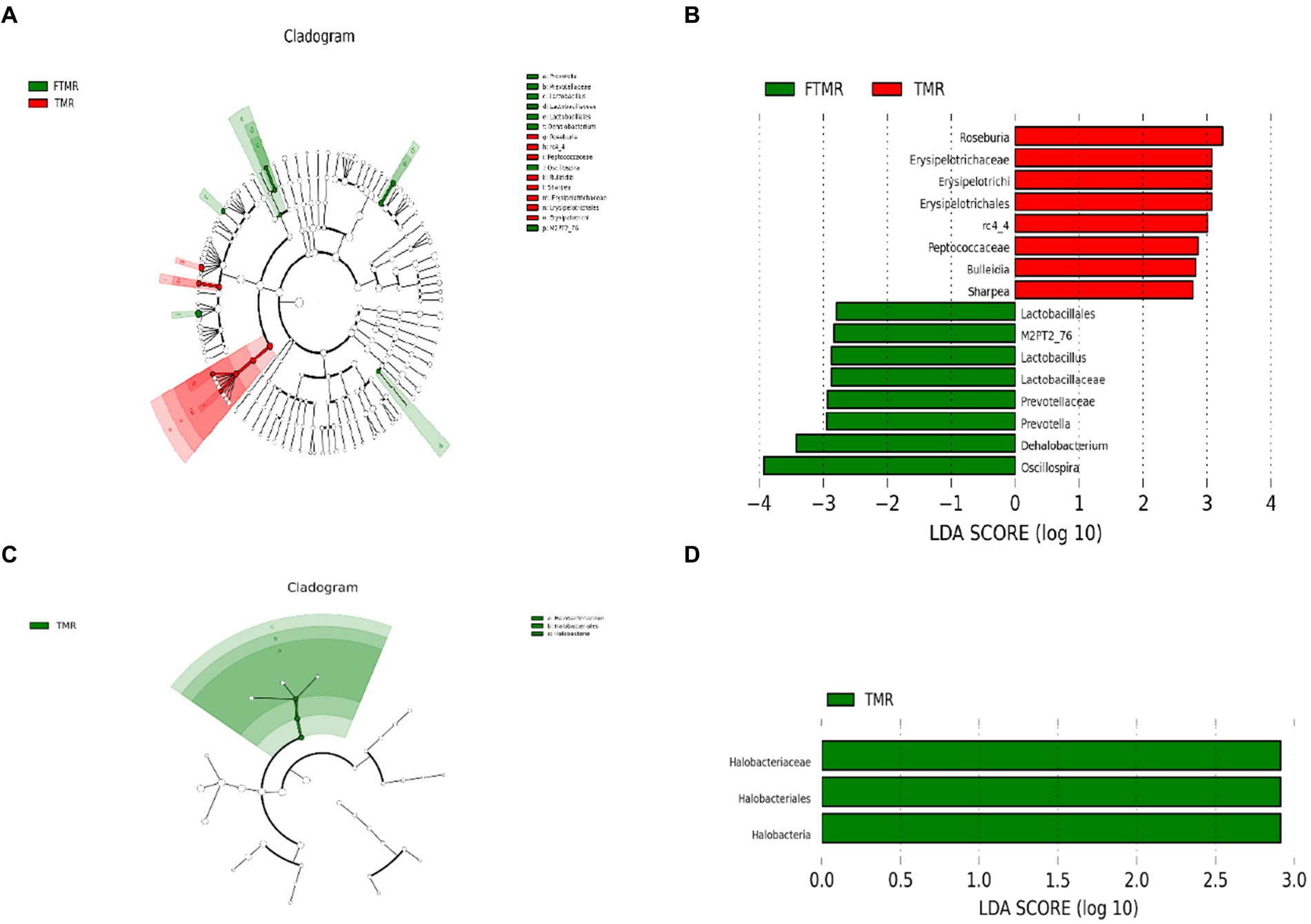
La Figura 5C muestra las variaciones entre los tres taxones de arqueas entre los grupos TMR y FTMR en un cladograma. En la Figura 5D, el gráfico de análisis LEfSe ilustra diferencias significativas en las puntuaciones de LDA de los taxones de arqueas entre estos grupos. Se evidenciaron biomarcadores significativos que distinguieron entre los grupos a nivel de clase y orden, específicamente Halobacteria y Halobacteriales, respectivamente. Además, a nivel familiar, Halobacteriaceae mostró diferencias significativas entre los grupos TMR y FTMR.
4 Discusión
4.1 Rendimiento de la lactancia, digestibilidad aparente del tracto total, pH fecal y AGV de las vacas lecheras
El DMI es un factor crucial que influye en el rendimiento de la producción de vacas lecheras (35). En este estudio, se observó una disminución del IMD en el grupo FTMR, mientras que la producción de leche no se vio afectada. Esto indica que la dieta FTMR demuestra una eficiencia alimenticia superior. La concentración de ácido acético en relación con los ácidos totales se asoció negativamente con la ingesta de ensilaje (36). Investigaciones anteriores han demostrado que las concentraciones de ácido acético superiores al 1,7% de MS reducen significativamente la ingesta de rumiantes (37). Además, la concentración de amoníaco-N en el ensilado de pasto se ha correlacionado inversamente con la ingesta de materia seca en vacas lecheras lactantes (38). Como resultado, el grupo alimentado con FTMR exhibió niveles más altos de ácidos orgánicos y nitrógeno amoniacal, junto con una menor ingesta de materia seca en comparación con el grupo alimentado con TMR. Sin embargo, la FTMR no tuvo ningún impacto en el DMI en las ovejas Ujumqin, probablemente debido a las diferencias entre especies y la calidad de la fermentación de la dieta (39). A pesar de la disminución de la DMI en la dieta FTMR, la producción de leche no se vio afectada debido a la mejora de la eficiencia alimenticia y al aumento de la ECM en comparación con la dieta TMR. Investigaciones previas indican que las enzimas fibrolíticas exógenas mejoran la digestibilidad de la fibra, aumentando así la ingesta de energía disponible y mejorando la eficiencia alimentaria (40). En comparación con la dieta TMR, la FTMR demostró una mayor digestibilidad de los nutrientes y eficiencia alimenticia, aumentando notablemente la digestibilidad de la MS (41). Cao et al. observaron un aumento de la digestibilidad de la MS con TMR fermentada en ovejas en comparación con el consumo regular de TMR (39). Además, la digestibilidad de la PC y la FDN mejoró en la dieta de FTMR, lo que concuerda con estudios recientes que demuestran una mejor digestibilidad de la PC y la FDN en ovejas alimentadas con FTMR, atribuida a un aumento de los niveles digeribles de PB y FDN (42). Se ha reportado que los niveles de enzimas impactan significativamente la digestibilidad de la MS y la FDN en el tracto total de manera cúbica (43), elevando así de manera equivalente la digestibilidad de la MO en vacas alimentadas con la dieta FTMR. El acetato es reconocido como un precursor primario para la síntesis de grasa láctea en la glándula mamaria durante la lactancia (44). Estudios previos han demostrado que las ovejas alimentadas con FTMR exhibieron mayores concentraciones de acetato en comparación con las alimentadas con TMR fresca (39), lo que contribuye a una mayor producción de grasa láctea debido a una mayor digestibilidad de la fibra de las enzimas fibrolíticas agregadas (43).
La fermentación alteró la composición de carbohidratos en el FTMR, afectando los patrones de fermentación del intestino posterior. A diferencia del grupo TMR, el grupo FTMR mostró un pH fecal más bajo. La adición de almidón (carbohidrato soluble) a las dietas de las vacas lecheras puede reducir significativamente el pH fecal, lo que podría aumentar la derivación de nutrientes en el rumen y acelerar las tasas de paso (45). Los microbios del intestino posterior en las vacas lecheras degradan del 5 al 10% de los carbohidratos, incluido el almidón, las pequeñas partículas que escapan de la fermentación ruminal y los componentes del rumen no digeridos (46, 47). Después del ensilado, la TMR mostró un contenido reducido de carbohidratos no fibrosos, lo que sugiere que la FTMR aumenta las concentraciones de carbohidratos no fibrosos y solubles, acelera las tasas de paso de alimento y aumenta las sustancias fermentables en el intestino posterior. Los estudios han reportado que las dietas altas en granos (ricas en carbohidratos sin fibra) aumentan significativamente los niveles de ácido butírico fecal y reducen el pH en las vacas (48), lo cual es consistente con los hallazgos del estudio actual. El grupo FTMR mostró concentraciones de butirato fecal marcadamente más altas, lo que se corresponde con una mayor abundancia relativa de bacterias productoras de butirato clave como Ruminococcaceae, Oscilospira y Bacteroidales. El acetato y el propionato son fuentes de energía cruciales para los rumiantes. La investigación sugiere que la fermentación dietética promueve la fermentación y absorción de propionato en el rumen (49, 50), lo que podría influir en las poblaciones microbianas que producen estos ácidos tanto en el rumen como en el intestino posterior.
4.2 Composición de la comunidad bacteriana fecal de las vacas lecheras
El histograma de abundancia relativa de los filos indicó que Firmicutes, junto con Bacteroidetes, eran las bacterias fecales predominantes, lo que concuerda con estudios previos (51-53). Investigaciones anteriores han relacionado la proporción de Firmicutes a Bacteroidetes con la extracción de energía en humanos y ratones (54, 55). Nuestros hallazgos no mostraron cambios en la proporción de Firmicutes a Bacteroidetes, lo que sugiere que esta proporción no se ve afectada en gran medida por las dietas de los rumiantes (56). Esto podría atribuirse a que la producción de energía primaria ocurre en el intestino anterior en lugar del intestino posterior (20).
En las heces, los géneros dominantes fueron Unclassified_Bacteroidales, Unclassified_Ruminococcaceae, Unclassified_Clostridiales y Oscillospira. Estudios anteriores han destacado que Unclassified_Ruminococcaceae y Unclassified_Bacteroidales son partes integrales del «microbioma bacteriano central» del rumen (57). Estos géneros desempeñan un papel crucial en la descomposición de las fibras vegetales dentro del tracto gastrointestinal (57).
En este estudio, la abundancia de Unclassified_Clostridiales, considerado un componente central de las bacterias fecales, no mostró diferencias significativas entre los grupos TMR y FTMR, lo cual es consistente con investigaciones previas (58). Esto indica que Unclassified_Clostridiales puede contribuir significativamente al ecosistema microbiano fecal independientemente de las diferencias dietéticas. Investigaciones adicionales que involucren el aislamiento e identificación de cepas representativas de Unclassified_Clostridiales podrían proporcionar información más profunda sobre su papel en la función del intestino posterior (59).
Nuestros hallazgos revelaron una abundancia significativamente mayor de Oscillospira en el grupo FTMR en comparación con otros grupos a nivel de género. El contenido de Oscillospira en el rumen del ganado alimentado con una dieta alta en almidón (carbohidratos solubles) aumentó (60). Zhao et al. sugirieron que la fermentación aumenta la estructura cristalina de los granos de almidón, promoviendo la producción de almidón resistente (61). Por lo tanto, los niveles elevados de Oscillospira en el grupo FTMR pueden atribuirse al mayor contenido de almidón resistente en el intestino posterior de las vacas alimentadas con FTMR. Mackie et al. observaron que Oscillospira es prevalente en el rumen de las vacas lecheras y está influenciado por factores dietéticos (62), lo que indica su capacidad de respuesta a la dieta en el tracto gastrointestinal. Oscillospira es conocida como un potencial productor de butirato, que contribuye a la salud intestinal (63), lo que sugiere que las dietas FTMR pueden mejorar la salud gastrointestinal a través de una mayor abundancia de Oscillospira. Aunque Oscillospira no se ha cultivado en forma pura (9), su potencial como probiótico para regular el tracto gastrointestinal de las vacas lecheras merece consideración.
A nivel de género, Roseburia, rc4-4, Bulleidia y Sharpea fueron significativamente más abundantes en el grupo TMR en comparación con el grupo FTMR, lo que concuerda con los hallazgos de Kim et al. (58). Esto indica que los taxones bacterianos menos comunes responden mejor a los cambios en la dieta que los que se encuentran más comúnmente. Las especies de Roseburia son conocidas por su capacidad para descomponer los polisacáridos de la dieta, produciendo butirato (64), lo que se correlaciona con nuestra observación de niveles más altos de ácido butírico en el grupo FTMR que en el grupo TMR. El género rc4-4 se asocia con dietas ricas en fibra (65), lo que sugiere que la dieta TMR puede tener un mayor contenido de fibra en el intestino posterior en comparación con la dieta FTMR. Sharpea desempeña un papel fundamental en la producción de ácido láctico (66, 67), que aumenta durante la acidosis ruminal en las vacas (68) y varía con la composición de la dieta en las ovejas (69). Si bien el ácido láctico no es un producto primario de la fermentación normal del rumen, las dietas altas en carbohidratos y azúcares solubles pueden conducir a su acumulación (70). Por lo tanto, la mayor abundancia de Sharpea en el grupo TMR puede deberse a un mayor flujo de carbohidratos y niveles de azúcar soluble desde el rumen hacia el intestino posterior.
Identificamos cuatro géneros, Lactobacillus, Prevotella, Dehalobacterium y Oscillospira, como biomarcadores distintivos entre los grupos FTMR y TMR. Lactobacillus es reconocido por su papel probiótico en la salud gastrointestinal de los rumiantes y se ha asociado con la reducción de las tasas de diarrea en los terneros (71-73). Nuestros hallazgos mostraron que Lactobacillus constituía menos del 1% de la microbiota fecal y no se consideraba una comunidad central, en consonancia con Tang et al. (74). Su abundancia varía con la dieta, pasando de la dominancia de la lactosa en los animales más jóvenes a dietas más ricas en carbohidratos complejos a medida que maduran (75-77). Si bien Lactobacillus prospera en el ensilaje, su supervivencia a través del tracto gastrointestinal, particularmente en el rumen, es limitada (50, 74). No obstante, nuestros resultados indicaron niveles significativamente más altos de Lactobacillus en las heces del grupo FTMR en comparación con el grupo TMR, lo que sugiere que el ensilaje podría servir como vehículo para la entrega de probióticos (50). Prevotella fue abundante en los grupos de TMR y FTMR, lo que concuerda con su presencia generalizada en las heces de vaca (56, 78, 79) y su dominio en el rumen (57, 71, 80). Su prevalencia en las heces (81) varía con la dieta, como niveles más altos en las dietas a base de maíz en comparación con las que tienen granos húmedos de destilería (82), lo que indica su presencia relacionada con la dieta en las heces del ganado (58, 83, 84). Las especies de Prevotella son conocidas por sus capacidades enzimáticas en la degradación de proteínas, almidón y hemicelulosa para producir succinato y acetato (85-87). Dehalobacterium mostró una mayor abundancia en las heces de FTMR en comparación con TMR, lo que sugiere que la dieta influye en su presencia (88). Se ha relacionado con dietas ricas en glucosa, aunque se necesita más investigación sobre su papel en el intestino posterior de las vacas lecheras (89).
4.3 Composición de la comunidad fecal de arqueas de vacas lecheras
Nuestros resultados mostraron que el índice de arqueas Chao1 en el grupo FTMR fue marcadamente mayor que el del grupo TMR, lo que confirma que la diversidad arqueal en el grupo FTMR fue marcadamente mayor que la de los otros grupos. El filo arqueológico predominante identificado en las heces fue Euryarchaeota. Investigaciones previas han indicado que Euryarchaeota es el filo bacteriano de arqueas más prevalente en el rumen (22, 85, 90), lo que subraya su función crítica en el sistema gastrointestinal. Además, se confirmó que Methanobrevibacter es el género de bacterias arqueales más prevalente en las heces, alineándose con estudios previos (22, 91), que establecieron su capacidad para generar CH4 a partir de hidrógeno y ácido fórmico (91, 92). Sin embargo, no se observaron diferencias significativas en la abundancia de Methanobrevibacter entre los grupos FTMR y TMR, lo que implica que FTMR puede no disminuir la producción de metano en el intestino posterior. Zhang et al. (93) observaron que las raciones altas en concentrado disminuyeron significativamente la abundancia de Fibrobacter y Methanobrevibacter, demostrando que un aumento en la concentración de estas bacterias en la dieta podría inhibir la síntesis de metano (94, 95). Por el contrario, este estudio no alteró la relación concentración/aproximación entre TMR y FTMR. Además, existe una interacción simbiótica entre bacterias y metanógenos en el intestino animal, y las bacterias celulolíticas en el rumen de los rumiantes muestran una correlación positiva con el número de metanógenos (96). Cabe destacar que no se detectaron alteraciones significativas en las abundancias relativas de Fibrobacteres y Fibrobacter en este estudio entre los grupos TMR y FTMR. Las halobacterias, las halobacterias y las Halobacteriaceae sirvieron como biomarcadores para diferenciar las TMR de la FTMR, siendo sus abundancias relativas considerablemente mayores en el grupo de TMR que en el grupo de FTMR, posiblemente debido a las variaciones en el contenido de cloruro de las células del intestino posterior entre los dos grupos (97).
5 Conclusión
En conclusión, se demostró que la FTMR aumenta el contenido de grasa láctea, mejora la digestibilidad de los nutrientes, eleva la proporción de butirato en la materia fecal y disminuye el pH fecal. Estos hallazgos indican que la FTMR puede aumentar la fermentación del intestino posterior. Además, la FTMR mejoró significativamente la diversidad de las comunidades de arqueas fecales y aumentó la abundancia relativa del género Oscillospira, lo que sugiere una mejor salud del intestino posterior en comparación con la TMR. Si bien se encontró que Methanobrevibacter es el género de arqueas más común en muestras fecales, no se observaron disparidades significativas entre los grupos de TMR y FTMR, lo que sugiere que FTMR puede no tener un impacto significativo en la producción de metano en el intestino posterior. Este estudio proporciona una base teórica para investigar el impacto de la FTMR en las comunidades bacterianas y arqueales en el intestino posterior, lo que indica que la alimentación con FTMR puede afectar beneficiosamente la salud del intestino posterior en las vacas lecheras.
Declaración de disponibilidad de datos
Los datos presentados en el estudio se depositan en el NCBI Sequence Read Archive (SRA), número de acceso: PRJNA1119071.
Declaración de ética
El estudio en animales fue aprobado por el Comité Asesor de Cuidado Animal de la Universidad Agrícola de Henan. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
LW: Escritura – borrador original, Escritura – revisión y edición, Conceptualización. SJ: Investigación, Software, Escritura – revisión y edición. PW: Curación de datos, Redacción – borrador original. XL: Metodología, Redacción – revisión y edición. CL: Análisis formal, Supervisión, Redacción – revisión y edición. SS: Análisis formal, Redacción – revisión y edición. GZ: Administración de proyectos, Redacción – revisión y edición. JC: Recursos, Redacción – revisión y edición. QY: Adquisición de fondos, Redacción – revisión y edición. HZ: Recursos, Visualización, Escritura – borrador original. QZ: Supervisión, Redacción – revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue financiada por la Investigación: Proyecto Clave de Ciencia y Tecnología de la Provincia de Henan (232102110103), la Fundación de Ciencias Naturales de Henan (242300421573), la Fundación de Ciencias Naturales de la Provincia de Heilongjiang para el Proyecto de Guía Conjunta (LH2022C030), los Proyectos de Orientación del Programa Clave de Investigación y Desarrollo de la Provincia de Heilongjiang (GZ20230020), el Proyecto Científico y Tecnológico Clave de Henan (201111311100), el Proyecto Clave de Investigación y Desarrollo de Henan (241111113700).
Reconocimientos
Nos gustaría agradecer al personal de nuestro laboratorio por su continua asistencia.
Conflicto de intereses
QZ fue empleado de Henan Delin Biological Products Co., Ltd.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Li, Y, Wu, Q, Lv, J, Jia, X, Gao, J, Zhang, Y, et al. Asociaciones de las estructuras moleculares de las proteínas con sus características de aporte de nutrientes y biodegradación en diferentes subproductos de la calabaza utilizada con semillas. Animales. (2022) 12:956. doi: 10.3390/ani12080956
2. Saleem, AM, Ribeiro, GO, Sanderson, H, Alipour, D, Brand, T, Hünerberg, M, et al. Efecto de las enzimas fibrolíticas exógenas y la expansión de la fibra de amoníaco en la fermentación de la paja de trigo en un sistema ruminal artificial (RUSITEC)1. J Anim Sci. (2019) 97:3535–49. doi: 10.1093/jas/skz224
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Schingoethe, DJ . Una revisión de 100 años: Alimentación total con raciones mixtas de vacas lecheras. J Dairy Sci. (2017) 100:10143–50. doi: 10.3168/jds.2017-12967
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Supapong, C, Cherdthong, A, Wanapat, M, Chanjula, P y Uriyapongson, S. Efectos de los niveles de azufre en la ración mezclada total fermentada que contiene raíz de yuca fresca en la utilización del alimento, las características del rumen, la síntesis de proteínas microbianas y los metabolitos sanguíneos en el ganado vacuno nativo tailandés. Animales. (2019) 9:261. doi: 10.3390/ani9050261
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Zhao, J, Dong, Z, Chen, L, Wang, S y Shao, T. El reemplazo de maíz de planta entera con cáscara de brote de bambú en la calidad de la fermentación, la composición química, la estabilidad aeróbica y la digestibilidad in vitro del ensilado de ración mezclada total. Anim Feed Sci Tech. (2020) 259:114348. doi: 10.1016/j.anifeedsci.2019.114348
6. Weinberg, ZG, Chen, Y, Miron, D, Raviv, Y, Nahim, E, Bloch, A, et al. Conservación de raciones totales mezcladas para vacas lecheras en fardos envueltos con film estirable de polietileno, un experimento a escala comercial. Anim Feed Sci Tech. (2011) 164:125–9. doi: 10.1016/j.anifeedsci.2010.11.016
7. Xie, Y, Xu, S, Li, W, Wang, M, Wu, Z, Bao, J, et al. Efectos de la aplicación de inoculante Lactobacillus plantarum y sorbato de potasio sobre la calidad de fermentación, digestibilidad in vitro y estabilidad aeróbica de ensilaje de ración mixta total a base de ensilaje de alfalfa. Animales. (2020) 10:2229. doi: 10.3390/ani10122229
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Chang, L, Mu, G, Wang, M, Zhao, T, Tuo, Y, Zhu, X, et al. Diversidad microbiana y propiedades fisicoquímicas relacionadas con la calidad de la col picante en el noreste de China y análisis de su correlación. Alimentos. (2022) 11:1511. doi: 10.3390/foods11101511
9. Liu, Q, Li, X, Desta, ST, Zhang, J-G y Shao, T. Efectos de Lactobacillus plantarum y la enzima fibrolítica en la calidad de la fermentación y la digestibilidad in vitro del ensilado total de raciones mezcladas, incluida la paja de colza. J Integr Agr. (2016) 15:2087–96. doi: 10.1016/S2095-3119(15)61233-3
10. Kondo, M, Shimizu, K, Jayanegara, A, Mishima, T, Matsui, H, Karita, S, et al. Cambios en la composición de nutrientes y fermentación ruminal in vitro de ensilaje de ración mixta total almacenado a diferentes temperaturas y períodos. J Sci Food Agr. (2016) 96:1175–80. doi: 10.1002/jsfa.7200
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Wang, F y Nishino, N. Resistencia al deterioro aeróbico del ensilado de ración mezclada total: efecto de la formulación de la ración, la infiltración de aire y el período de almacenamiento sobre las características de fermentación y la estabilidad aeróbica. J Sci Food Agr. (2008) 88:133–40. doi: 10.1002/jsfa.3057
12. Gusmão, J, Danés, M, Casagrande, D, y Bernardes, T. Ensilado de ración mixta total que contiene pasto de elefante para granjas lecheras a pequeña escala. Ciencia de pasto y forraje. (2018) 73:717–26. doi: 10.1111/gfs.12357
13. Bai, B, Qiu, R, Wang, Z, Liu, Y, Bao, J, Sun, L, et al. Efectos de la celulasa y las bacterias lácticas sobre el rendimiento del ensilado y la comunidad bacteriana del ensilado de caragana korshinskii. Microorganismos. (2023) 11:337. DOI: 10.3390/microorganismos11020337
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Sun, Q, Gao, F, Yu, Z, Tao, Y, Zhao, S y Cai, Y. Calidad de fermentación y composición química del ensilado de arbustos tratado con inoculantes de bacterias lácticas y aditivos de celulasa. Anim Sci J. (2012) 83:305–9. doi: 10.1111/j.1740-0929.2011.00962.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Arriola, KG, Vyas, D, Kim, D, Agarussi, MCN, Silva, VP, Flores, M, et al. Efecto de Lactobacillus hilgardii, Lactobacillus buchneri, o su combinación sobre la fermentación y el valor nutritivo del ensilado de sorgo y maíz. J Dairy Sci. (2021) 104:9664–75. DOI: 10.3168/jds.2020-19512
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Xu, H, Huang, W, Hou, Q, Kwok, L-y, Sun, Z, Ma, H, et al. Los efectos de la administración de probióticos sobre la producción de leche, los componentes de la leche y la microbiota de bacterias fecales de las vacas lecheras. Boletín Sci. (2017) 62:767–74. doi: 10.1016/j.scib.2017.04.019
17. Liu, J, Taft, DH, Maldonado-Gómez, MX, Johnson, D, Treiber, ML, Lemay, DG, et al. El resistoma fecal del ganado lechero se asocia con la dieta durante la lactancia. Na Commun. (2019) 10:4406. doi: 10.1038/s41467-019-12111-x
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Faulkner, MJ, Wenner, BA, Solden, LM y Weiss, WP. La fuente de cobre, zinc y manganeso en la dieta suplementaria afecta la abundancia relativa de microbios fecales en las vacas lecheras lactantes. J Dairy Sci. (2017) 100:1037–44. doi: 10.3168/jds.2016-11680
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Welch, CB, Lourenco, JM, Krause, TR, Seidel, DS, Fluharty, FL, Pringle, TD, et al. Evaluación de las comunidades bacterianas fecales de novillos angus con eficiencias alimenticias divergentes a lo largo de la vida desde el destete hasta el sacrificio. Fronteras de la Veterinaria. (2021) 8:694. doi: 10.3389/fvets.2021.597405
20. Zhang, J, Shi, H, Wang, Y, Cao, Z, Yang, H y Li, S. Efecto de dietas alimentadas limitadas con diferentes proporciones de forraje a concentrado en la composición de la comunidad fecal bacteriana y de arqueas en novillas Holstein. Microbiol frontal. (2018) 9:976. doi: 10.3389/fmicb.2018.00976
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Al-Azzawi, M, Bowtell, L, Hancock, K y Preston, S. Adición de carbón activado a la dieta del ganado para mitigar las emisiones de GEI y mejorar la producción. Sostener para. (2021) 13:8254. doi: 10.3390/su13158254
22. Kumar, S, Indugu, N, Vecchiarelli, B, y Pitta, DW. Patrones asociativos entre hongos anaeróbicos, arqueas metanogénicas y comunidades bacterianas en respuesta a cambios en la dieta y la edad en el rumen de vacas lecheras. Microbiol frontal. (2015) 6:781. doi: 10.3389/fmicb.2015.00781
23. Zhang, J, Shi, H, Wang, Y, Li, S, Cao, Z, Ji, S, et al. Efecto de las proporciones de forraje a concentrado en la dieta, los cambios en el perfil dinámico y las interacciones de la microbiota ruminal y los metabolitos en novillas Holstein. Microbiol frontal. (2017) 8:2206. doi: 10.3389/fmicb.2017.02206
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Ramin, M, Chagas, JC, Smidt, H, Expósito, RG y Krizsan, SJ. Emisiones entéricas y fecales de metano de vacas lecheras alimentadas con dietas de ensilado de pasto o maíz suplementadas con aceite de colza. Animales. (2021) 11:1322. doi: 10.3390/ani11051322
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Laporte, MF y Paquin, P. Análisis de infrarrojo cercano de grasa, proteína y caseína en leche de vaca. J Agric Food Chem. (1999) 47:2600–5. doi: 10.1021/jf980929r
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Goeser, JP, y Combs, DK. Un método alternativo para evaluar la digestibilidad de la fibra detergente neutro ruminal in vitro durante 24 horas. J Dairy Sci. (2009) 92:3833–41. doi: 10.3168/jds.2008-1136
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Wang, L, Zhang, G, Li, Y y Zhang, Y. Efectos de una dieta alta en forraje/concentrado sobre la producción de ácidos grasos volátiles y los microorganismos involucrados en la producción de AGV en el rumen de las vacas. Animales. (2020) 10:223. doi: 10.3390/ani10020223
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Vasseur, E, Gibbons, J, Rushen, J y de Passillé, AM. Desarrollo e implementación de un programa de entrenamiento para asegurar una alta repetibilidad de la puntuación de la condición corporal de las vacas lecheras. J Dairy Sci. (2013) 96:4725–37. doi: 10.3168/jds.2012-6359
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Caporaso, JG, Kuczynski, J, Stombaugh, J, Bittinger, K, Bushman, FD, Costello, EK, et al. QIIME permite el análisis de datos de secuenciación comunitaria de alto rendimiento. Métodos Nat. (2010) 7:335–6. doi: 10.1038/nmeth.f.303
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Edgar, RC . Búsqueda y agrupación en órdenes de magnitud más rápida que BLAST. Bioinformática. (2010) 26:2460–1. doi: 10.1093/bioinformatics/btq461
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Pruesse, E, Quast, C, Knittel, K, Fuchs, BM, Ludwig, WG, Peplies, J, et al. SILVA: un recurso integral en línea para datos de secuencia de ARN ribosómico alineados y de calidad verificada compatibles con ARB. Ácidos nucleicos Res. (2007) 35:7188–7196. doi: 10.1093/nar/gkm864
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Wolf, MA, Sfriso, A, y Moro, I. Contaminación térmica y asentamiento de nuevas especies exóticas tropicales: el caso de Grateloupia yinggehaiensis (Rhodophyta) en la laguna de Venecia. Plataforma Costera del Estuario S. (2014) 147:11–16. doi: 10.1016/j.ecss.2014.05.020
33. Kõljalg, U, Larsson, KH, Abarenkov, K, Nilsson, RH, Alexander, IJ, Eberhardt, U, et al. UNITE: una base de datos que proporciona métodos basados en la web para la identificación molecular de hongos ectomicorrícicos. Nuevo Phytol. (2005) 166:1063–1068. doi: 10.1111/j.1469-8137.2005.01376.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Segata, N, Izard, J, Waldron, L, Gevers, D, Miropolsky, L, Garrett, WS, et al. Descubrimiento y explicación de biomarcadores metagenómicos. Genoma Biol. (2011) 12:R60. doi: 10.1186/gb-2011-12-6-r60
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Ma, J, Sun, G, Shah, AM, Fan, X, Li, S y Yu, X. Efectos de diferentes etapas de crecimiento del ensilado de amaranto en la degradación ruminal de las vacas lecheras. Animales. (2019) 9:793. doi: 10.3390/ani9100793
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Wilkins, RJ, Hutchinson, KJ, Wilson, RF y Harris, CE. La ingesta voluntaria de ensilado por parte de las ovejas: I. Interrelaciones entre la composición y la ingesta de ensilaje. JAS. (1971) 77:531–7. doi: 10.1017/S0021859600064613
37. Gerlach, K, Jobim, JLP, Nussio, CC, y Gustavo, L. Análisis de datos sobre el efecto del ácido acético en la ingesta de materia seca en ganado lechero. Anim Feed Sci Technol. (2021) 272:114782. doi: 10.1016/j.anifeedsci.2020.114782
38. Huhtanen, P, Khalili, H, Nousiainen, JI, Rinne, M, Jaakkola, S, Heikkil, T, et al. Predicción del potencial de ingesta relativa de ensilado de pasto por parte de las vacas lecheras. Livest Prod Sci. (2002) 73:111–30. doi: 10.1016/S0301-6226(01)00279-2
39. Cao, Y, Zang, Y, Jiang, Z, Han, Y, Hou, JJ, Liu, H, et al. Calidad de la fermentación y valor nutritivo de las raciones mezcladas totales frescas y fermentadas que contienen centeno silvestre chino o rastrojo de maíz. Grassl Sci. (2016) 62:213–23. doi: 10.1111/grs.12134
40. Tewoldebrhan, TA, Appuhamy, JADRN, Lee, JJ, Niu, M, Seo, S, Jeong, S, et al. La β-mananasa exógena mejora la eficiencia de la conversión alimenticia y reduce el recuento de células somáticas en el ganado lechero. J Dairy Sci. (2017) 100:244–52. doi: 10.3168/jds.2016-11017
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Ferraretto, LF, Crump, PM y Shaver, RD. Efecto de los métodos de recolección y procesamiento del tipo de grano de cereal y grano de maíz en la ingesta, digestión y producción de leche por vacas lecheras a través de un metaanálisis. J Dairy Sci. (2013) 96:533–50. doi: 10.3168/jds.2012-5932
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Cao, Y, Takahashi, T, Horiguchi, KI, Yoshida, N y Cai, Y. Emisiones de metano de ovejas alimentadas con ración mezclada total fermentada o no fermentada que contiene arroz integral y salvado de arroz. Anim Feed Sci Tech. (2010) 157:72–8. doi: 10.1016/j.anifeedsci.2010.02.004
43. Refat, B, Christensen, DA, Mckinnon, JJ, Yang, W, Beattie, AD, Mcallister, TA, et al. Efecto de las enzimas fibrolíticas sobre el rendimiento de la lactancia, el comportamiento alimentario y la digestibilidad en vacas lecheras de alta producción alimentadas con una dieta basada en ensilado de cebada. J Dairy Sci. (2018) 101:7971–9. doi: 10.3168/jds.2017-14203
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Smith, SB . Contribución del ciclo de las pentosas a la lipogénesis en el tejido adiposo bovino. Arch Biochem Biophys. (1983) 221:46–56. doi: 10.1016/0003-9861(83)90120-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Neubauer, V, Petri, RM, Humer, E, Kröger, I, Reisinger, N, Baumgartner, W, et al. La dieta rica en almidón indujo acidosis ruminal y disbiosis del intestino posterior en vacas lecheras de diferentes lactancias. Animales. (2020) 10:1727. doi: 10.3390/ani10101727
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Gressley, TF, Hall, MB y Armentano, LE. Simposio sobre nutrición de rumiantes: productividad, digestión y respuestas saludables a la acidosis del intestino posterior en rumiantes. J Anim Sci. (2011) 89:1120–30. doi: 10.2527/jas.2010-3460
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Lyu, J, Yang, Z, Wang, E, Liu, G, Wang, Y, Wang, W, et al. Posibilidad de utilizar subproductos con alto contenido de FDN para alterar los perfiles fecales de ácidos grasos de cadena corta, la comunidad bacteriana y la digestibilidad de las vacas lecheras lactantes. Microorganismos. (2022) 10:1731. doi: 10.3390/microorganismos10091731
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Mao, S, Zhang, R, Wang, D y Zhu, W. La diversidad de la comunidad bacteriana fecal y su relación con la concentración de ácidos grasos volátiles en las heces durante la acidosis ruminal subaguda en vacas lecheras. BMC Vet Res. (2012) 8:237. doi: 10.1186/1746-6148-8-237
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Gastelen, S, Dijkstra, J, Gerrits, WJJ, Gilbert, MS y Bannink, A. Comunicación breve: cuantificación de la fermentación del almidón postruminal en vacas Holstein-Frisonas de lactancia temprana. Animal. (2023) 17:100974. doi: 10.1016/j.animal.2023.100974
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Han, H, Ogata, Y, Yamamoto, Y, Nagao, S y Nishino, N. Identificación de bacterias del ácido láctico en el rumen y las heces de vacas lecheras alimentadas con ensilaje de ración mixta total para evaluar la supervivencia de las bacterias del ensilado en el intestino. J Dairy Sci. (2014) 97:5754–62. doi: 10.3168/jds.2014-7968
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Rice, WC, Galyean, ML, Cox, SB, Dowd, SE, y Cole, NA. Influencia de las dietas húmedas de granos de destilería en la estructura de la comunidad bacteriana fecal del ganado vacuno. BMC Microbiol. (2012) 12:25. doi: 10.1186/1471-2180-12-25
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Li, S, Khafipour, E, Krause, DO, Kroeker, A, Rodriguez-Lecompte, JC, Gozho, GN, et al. Efectos de los desafíos de la acidosis ruminal subaguda sobre la fermentación y las endotoxinas en el rumen y el intestino posterior de las vacas lecheras. J Dairy Sci. (2012) 95:294–303. doi: 10.3168/jds.2011-4447
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Plaizier, JC, Li, S, Danscher, AM, Derakshani, H, Andersen, PH y Khafipour, E. Cambios en la microbiota en la digesta ruminal y las heces debido a un desafío de acidosis ruminal subaguda (SARA) a base de granos. Microb Ecol. (2017) 74:485–95. doi: 10.1007/s00248-017-0940-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Ley, RE, Turnbaugh, PJ, Klein, S y Gordon, JI. Microbios intestinales humanos asociados con la obesidad. Naturaleza. (2006) 444:1022–3. doi: 10.1038/4441022a
55. Turnbaugh, PJ, Ley, RE, Mahowald, MA, Magrini, V, Mardis, ER, y Gordon, JI. Un microbioma intestinal asociado a la obesidad con mayor capacidad de recolección de energía. Naturaleza. (2006) 444:1027–31. doi: 10.1038/nature05414
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Li, Y, Meng, Q, Zhou, B y Zhou, Z. Efecto de las hojas de morera ensiladas y el orujo de morera secado al sol en la composición de la comunidad bacteriana fecal en novillos de engorde. BMC Microbiol. (2017) 17:97. doi: 10.1186/s12866-017-1011-9
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. Henderson, G, Cox, F, Ganesh, S, Jonker, A, Young, W y Janssen, PH. La composición de la comunidad microbiana del rumen varía con la dieta y el huésped, pero se encuentra un microbioma central en una amplia gama geográfica. Sci Rep. (2015) 5:1–15. doi: 10.1038/srep14567
58. Kim, M, Kim, J, Kuehn, LA, Bono, JL, Berry, ED, Kalchayanand, N, et al. Investigación de la diversidad bacteriana en las heces de bovinos alimentados con diferentes dietas 1. J Anim Sci. (2014) 92:683–94. doi: 10.2527/jas.2013-6841
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Kim, M, Morrison, M y Yu, Z. Estado del censo de diversidad filogenética de los microbiomas ruminales. FEMS Microbiol Ecol. (2011) 76:49–63. doi: 10.1111/j.1574-6941.2010.01029.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Myer, PR, Wells, JE, Smith, TP, Kuehn, LA, y Freetly, HC. Perfiles de la comunidad microbiana del colon de novillos que difieren en eficiencia alimenticia. Springerplus. (2015) 4:454. doi: 10.1186/s40064-015-1201-6
61. Zhao, T, Li, X, Zhu, R, Ma, Z, Liu, L, Wang, X, et al. Efecto de la fermentación natural sobre la estructura y propiedades fisicoquímicas del almidón de trigo. Carbohidrato Polím. (2019) 218:163–9. doi: 10.1016/j.carbpol.2019.04.061
Resumen de PubMed | Texto completo de Crossref | Google Académico
62. Mackie, RI, Aminov, RI, Hu, W, Klieve, AV, Ouwerkerk, D, Sundset, MA, et al. Ecología de especies de Oscillospira no cultivadas en el rumen de ganado vacuno, ovino y reno evaluadas por microscopía y enfoques moleculares. Appl Environ Microbiol. (2003) 69:6808–15. doi: 10.1128/AEM.69.11.6808-6815.2003
Resumen de PubMed | Texto completo de Crossref | Google Académico
63. Walters, WA, Xu, Z y Knight, R. Metaanálisis de microbios intestinales humanos asociados con la obesidad y la EII. FEBS Lett. (2014) 588:4223–33. doi: 10.1016/j.febslet.2014.09.039
Resumen de PubMed | Texto completo de Crossref | Google Académico
64. Rodríguez, J, Neyrinck, AM, Zhang, Z, Seethaler, B, Nazaré, JA, Robles Sánchez, C, et al. El perfil de metabolitos revela la interacción de la quitina-glucano con la microbiota intestinal. Microbios intestinales. (2020) 12:1810530. doi: 10.1080/19490976.2020.1810530
Resumen de PubMed | Texto completo de Crossref | Google Académico
65. Zhong, Y, Nyman, M y Fåk, F. Modulación de la microbiota intestinal en ratas alimentadas con dietas ricas en grasas mediante el procesamiento de cebada integral a malta de cebada. Mol Nutr Food Res. (2015) 59:2066–76. doi: 10.1002/mnfr.201500187
Resumen de PubMed | Texto completo de Crossref | Google Académico
66. Marketta, R, Marcia, F, Ilma, T, Tomasz, S, Ali-Reza, B y Päivi, M. Efectos del tipo de aditivo de ensilado de pasto y el método de conservación del grano de cebada en la fermentación ruminal, la comunidad microbiana y la producción de leche de las vacas lecheras. Agricultura. (2022) 12:1–18. doi: 10.3390/agricultura12020266
67. Lin, L, Xie, F, Sun, D, Liu, J, Zhu, W y Mao, S. La diafonía entre el microbioma ruminal y el huésped estimula el desarrollo del epitelio ruminal en un modelo de cordero. Microbioma. (2019) 7:83. doi: 10.1186/s40168-019-0701-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
68. Plaizier, JC, Li, S, Tun, HM y Khafipour, E. Los modelos nutricionales de la acidosis ruminal subaguda inducida experimentalmente (SARA) difieren en su impacto sobre las comunidades bacterianas del rumen y del intestino posterior en vacas lecheras. Microbiol frontal. (2017) 7:2128. doi: 10.3389/fmicb.2016.02128
69. Kamke, J, Kittelmann, S, Soni, P, Li, Y, Tavendale, M, Ganesh, S, et al. Los análisis del metagenoma y el metatranscriptoma ruminal de ovejas de bajo rendimiento de metano revelan un microbioma enriquecido con Shardea caracterizado por la formación y utilización de ácido láctico. Microbioma. (2016) 4:56. doi: 10.1186/s40168-016-0201-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
70. Dewanckele, L, Toral, PG, Vlaeminck, B y Fievez, V. Revisión invitada: papel de los intermedios de biohidrogenación ruminal y los microbios ruminales en la depresión de la grasa láctea inducida por la dieta: una actualización. J Dairy Sci. (2020) 103:7655–81. doi: 10.3168/jds.2019-17662
Resumen de PubMed | Texto completo de Crossref | Google Académico
71. Koike, S, Ueno, M, Ashida, N, Imabayashi, T y Kobayashi, Y. Efecto de la suplementación con Bacillus subtilis C-3102 en sustituto de leche sobre el crecimiento y la microbiota ruminal en terneros predestetados. Anim Sci J. (2021) 92:e13580. doi: 10.1111/asj.13580
Resumen de PubMed | Texto completo de Crossref | Google Académico
72. Timmerman, HM, Mulder, L, Everts, H, Van Espen, D, Van Der Wal, E, Klaassen, G, et al. Salud y crecimiento de terneros alimentados con sustitutos de leche con o sin probióticos. J Dairy Sci. (2005) 88:2154–65. doi: 10.3168/jds. S0022-0302(05)72891-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
73. Nocek, J y Kautz, W. Suplementación microbiana de alimentación directa sobre la digestión ruminal, la salud y el rendimiento del ganado lechero antes y después del parto. J Dairy Sci. (2006) 89:260–6. doi: 10.3168/jds. S0022-0302(06)72090-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
74. Tang, MT, Han, H, Yu, Z, Tsuruta, T y Nishino, N. Variabilidad, estabilidad y resiliencia de la microbiota fecal en vacas lecheras alimentadas con ensilado de maíz de cultivo entero. Aplicación Microbiol Biotechnol. (2017) 101:6355–64. doi: 10.1007/s00253-017-8348-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
75. Hennessy, ML, Indugu, N, Vecchiarelli, B, Bender, J, Pappalardo, C, Leibstein, M, et al. Cambios temporales en la comunidad bacteriana fecal en terneros lecheros Holstein desde el nacimiento hasta la transición a una dieta sólida. PLoS UNO. (2020) 15:e0238882. doi: 10.1371/journal.pone.0238882
Resumen de PubMed | Texto completo de Crossref | Google Académico
76. Gómez, D, Arroyo, L, Costa, M, Viel, L y Weese, J. Caracterización de la microbiota bacteriana fecal de terneros lecheros sanos y diarreicos. J Veterinario Pasante Med. (2017) 31:928–39. doi: 10.1111/jvim.14695
Resumen de PubMed | Texto completo de Crossref | Google Académico
77. Yang, Q, Huang, X, Wang, P, Yan, Z, Sun, W, Zhao, S, et al. Desarrollo longitudinal de la microbiota intestinal en lechones sanos y diarreicos inducido por cambios en la dieta relacionados con la edad. Microbiología. (2019) 8:E923. DOI: 10.1002/mbo3.923
Resumen de PubMed | Texto completo de Crossref | Google Académico
78. Dowd, SE, Callaway, TR, Wolcott, RD, Sun, Y, McKeehan, T, Hagevoort, RG, et al. Evaluación de la diversidad bacteriana en las heces del ganado bovino mediante pirosecuenciación de pirosecuenciación de amplicones FLX codificada por etiquetas bacterianas de ADNr 16S (bTEFAP). BMC Microbiol. (2008) 8:125–8. doi: 10.1186/1471-2180-8-125
79. Albonico, F, Barelli, C, Albanese, D, Manica, M, Partel, E, Rosso, F, et al. La leche cruda y la microbiota fecal de las vacas lecheras alpinas comerciales varían según el rebaño, el contenido de grasa y la dieta. PLoS Uno. (2020) 15:e0237262. doi: 10.1371/journal.pone.0237262
Resumen de PubMed | Texto completo de Crossref | Google Académico
80. Xue, M, Sun, H, Wu, X, Guan, L y Liu, J. Evaluación de las bacterias del rumen en vacas lecheras con un rendimiento variado de proteína láctea. J Dairy Sci. (2019) 102:5031–41. doi: 10.3168/jds.2018-15974
Resumen de PubMed | Texto completo de Crossref | Google Académico
81. Liu, J, Ml, Z, Ry, Z, Wy, Z y Sy, M. Estudios comparativos de la composición de la microbiota bacteriana asociada con el contenido ruminal, el epitelio ruminal y las heces de vacas lecheras lactantes. Microb Biotechnol. (2016) 9:257–68. doi: 10.1111/1751-7915.12345
Resumen de PubMed | Texto completo de Crossref | Google Académico
82. Durso, L, Wells, J, Harhay, G, Rice, W, Kuehn, L, Bono, J, et al. Comparación de las comunidades bacterianas en heces de ganado vacuno alimentado con dietas que contienen maíz y grano de destilería húmedo con solubles. Lett Appl Microbiol. (2012) 55:109–14. doi: 10.1111/j.1472-765X.2012.03265.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
83. Shanks, OC, Kelty, CA, Archibeque, S, Jenkins, M, Newton, RJ, McLellan, SL, et al. Estructuras comunitarias de bacterias fecales en bovinos de diferentes explotaciones de alimentación animal. Appl Environ Microb. (2011) 77:2992–3001. doi: 10.1128/AEM.02988-10
Resumen de PubMed | Texto completo de Crossref | Google Académico
84. Durso, LM, Harhay, GP, Smith, TP, Bono, JL, DeSantis, TZ, Harhay, DM, et al. Variación de animal a animal en la diversidad microbiana fecal entre el ganado vacuno. Appl Environ Microb. (2010) 76:4858–62. doi: 10.1128/AEM.00207-10
Resumen de PubMed | Texto completo de Crossref | Google Académico
85. Danielsson, R, Dicksved, J, Sun, L, Gonda, H, Müller, B, Schnürer, A, et al. La producción de metano en las vacas lecheras se correlaciona con la estructura de la comunidad metanogénica y bacteriana del rumen. Microbiol frontal. (2017) 8:226. doi: 10.3389/fmicb.2017.00226
86. Stevenson, DM, y Weimer, PJ. La dominancia de Prevotella y la baja abundancia de especies bacterianas ruminales clásicas en el rumen bovino se revelaron mediante PCR en tiempo real de cuantificación relativa. Appl Environ Microb. (2007) 75:165–74. doi: 10.1007/s00253-006-0802-y
87. Cota, MA . Interacción de las bacterias ruminales en la producción y utilización de maltooligosacáridos a partir del almidón. Appl Environ Microb. (1992) 58:48–54. doi: 10.1128/aem.58.1.48-54.1992
Resumen de PubMed | Texto completo de Crossref | Google Académico
88. Lau, SKP, Teng, JLL, Chiu, TH, Chan, E, Tsang, AKL, Panagiotou, G, et al. Comunidades microbianas diferenciales de ganado omnívoro y herbívoro en el sur de China. Comput Struct Biotechnol J. (2018) 16:54–60. doi: 10.1016/j.csbj.2018.02.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
89. O’Connor, A, Quizon, PM, Albright, JE, Lin, FT y Bennett, BJ. La capacidad de respuesta de la microbiota cardiometabólica a la dieta está influida por la genética del huésped. Genoma de Mamm. (2014) 25:583–99. doi: 10.1007/s00335-014-9540-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
90. Zhou, M, Hernández-Sanabria, E, y Guan, LL. Evaluación de la ecología microbiana de los metanógenos ruminales en el ganado con diferentes eficiencias alimenticias. Appl Environ Microbiol. (2009) 75:6524–33. doi: 10.1128/AEM.02815-08
Resumen de PubMed | Texto completo de Crossref | Google Académico
91. Holman, DB, Hao, X, Topp, E, Yang, HE y Alexander, TW. Efecto del cocompostaje de estiércol de ganado con residuos de construcción y demolición sobre la microbiota de arqueas, bacterias y hongos, y sobre los determinantes de la resistencia a los antimicrobianos. PLoS Uno. (2016) 11:e0157539. doi: 10.1371/journal.pone.0157539
Resumen de PubMed | Texto completo de Crossref | Google Académico
92. Zhang, T, Mu, Y, Zhang, D, Lin, X, Wang, Z, Hou, Q, et al. Determinación de características microbiológicas en el tracto digestivo de diferentes especies de rumiantes. Microbiología Abierta. (2018) 8:E00769. DOI: 10.1002/mbo3.769
93. Zhang, R, Liu, J, Jiang, L, Wang, X y Mao, S. Los efectos remodeladores de las dietas altas en concentración sobre la composición y función microbiana en el intestino posterior de las vacas lecheras. Nuerca delantera. (2021) 8:809406. doi: 10.3389/fnut.2021.809406
94. Mc Geough, E, O’kiely, P, Hart, K, Moloney, A, Boland, T y Kenny, D. Emisiones de metano, ingesta de alimento, rendimiento, digestibilidad y fermentación ruminal del ganado vacuno de engorde de carne de engorde ofrecido a ensilados de trigo integral que difieren en el contenido de grano. J Anim Sci. (2010) 88:2703–16. doi: 10.2527/jas.2009-2750
Resumen de PubMed | Texto completo de Crossref | Google Académico
95. Knapp, JR, Laur, G, Vadas, PA, Weiss, WP y Tricarico, JM. Revisión invitada: metano entérico en la producción de ganado lechero: cuantificación de las oportunidades y el impacto de la reducción de emisiones. J Dairy Sci. (2014) 97:3231–61. doi: 10.3168/jds.2013-7234
Resumen de PubMed | Texto completo de Crossref | Google Académico
96. Morvan, B, Bonnemoy, F, Fonty, G y Gouet, P. Determinación cuantitativa de bacterias acetogénicas y reductoras de sulfato que utilizan H2 y arqueas metanogénicas del tracto digestivo de diferentes mamíferos. Curr Microbiol. (1996) 32:129–33. doi: 10.1007/s002849900023
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: ración mezclada total fermentada, vacas lecheras, digestibilidad, microbiota fecal, patrón de fermentación fecal
Cita: Wang L, Jin S, Wang P, Li X, Liu C, Sun S, Zhang G, Chang J, Yin Q, Zhang H y Zhu Q (2024) La ración mezclada total fermentada mejora la digestibilidad de los nutrientes y modula los componentes de la leche y la comunidad microbiana fecal en vacas lecheras Holstein lactantes. Frente. Vet. Sci. 11:1408348. doi: 10.3389/fvets.2024.1408348
Editado por:
Izhar Hyder Qazi, Shaheed Benazir Bhutto, Universidad de Ciencias Veterinarias y Animales, Pakistán
Revisado por:
Yu Pi, Academia China de Ciencias Agrícolas, China
Ruangyote Pilajun, Universidad Ubon Ratchathani, Tailandia
Derechos de autor © 2024 Wang, Jin, Wang, Li, Liu, Sun, Zhang, Chang, Yin, Zhang y Zhu. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Qingqiang Yin, qqy1964@henau.edu.cn; Haiyang Zhang, dahai0806702@126.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía