
La suplementación dietética con proteína Clostridium autoethanogenum mejora el rendimiento del crecimiento

La suplementación dietética con proteína Clostridium autoethanogenum mejora el rendimiento del crecimiento y promueve la síntesis de proteínas musculares mediante la activación de la vía de señalización mTOR del pollo de engorde
 Chunqiao Shan1†
Chunqiao Shan1†  Yan Liu2† Chaoxin Ma3
Yan Liu2† Chaoxin Ma3  Chuang Li3 Qiuchen Liu2 Sisi Liu4 Guotuo Jiang2*
Chuang Li3 Qiuchen Liu2 Sisi Liu4 Guotuo Jiang2*  Jing Tian1*
Jing Tian1*- 1Escuela de Ingeniería Biológica, Universidad Politécnica de Dalian, Dalian, China
- 2Instituto de Investigación Biotecnológica Dalian Sanyi, Dalian Sanyi Animal Medicine Co., Ltd., Dalian Liaoning, China
- 3Centro de Control de Calidad de Investigación, Jiangsu Sanyi Animal Nutrition Technology Co., Ltd., Xuzhou, China
- 4Academia de Ciencias Agrícolas de Harbin, Harbin Heilongjiang, China
El experimento tuvo como objetivo evaluar los efectos de diferentes proporciones de proteína Clostridium autoethanogenum (CAP) utilizadas en las dietas sobre el rendimiento del crecimiento, la calidad muscular, los índices séricos y la vía mTOR de pollos de engorde de plumas blancas. Cuatrocientos ochenta pollos de engorde Arbor Acres (AA) de 1 día de edad, que comprenden el mismo número de machos y hembras, fueron asignados aleatoriamente a uno de cuatro tratamientos, y cada tratamiento consistió en 12 réplicas de 10 aves. Se formularon cuatro dietas basadas en principios isoenergéticos e isonitrogenados. El grupo control (CAP 0) no recibió ningún CAP, mientras que los grupos experimentales recibieron 2% (CAP 2), 3% (CAP 3) y 4% (CAP 4) de CAP durante seis semanas. En comparación con el CAP0, (1) el índice de conversión alimenticia (FCR) fue menor (p < 0.05) y el rendimiento muscular de la pierna fue mayor (p < 0.05) en el CAP3 y CAP4; (2) Los niveles séricos de TP, ALB, T-AOC y SOD mejoraron en el CAP3 (p < 0,05); (3) La expresión del gen Lipin-1 se reguló a la baja y los genes AMPKɑ2, Akt y 4E-BP1 se regularon al alza en el grupo experimental (p < 0,05); (4) La inclusión de 3% de NAC en la dieta aumentó los niveles de fosforilación de 4E-BP1, S6K1, Akt y AMPKɑ2 mediante la modulación de la vía de señalización mTOR (p < 0,05). En conclusión, las dietas para pollos de engorde que contienen un 3% de NAC pueden activar la vía de señalización mTOR para promover la síntesis muscular y mejorar el rendimiento del crecimiento.
Introducción
La población mundial ha aumentado drásticamente en los últimos años y podría alcanzar los 10 mil millones en 2050 (1, 2). Como resultado, se espera que la demanda mundial de carne supere los 400 millones de toneladas, mientras que la demanda de productos lácteos superará los 800 millones de toneladas (3). La harina de soja se usa ampliamente en la industria de la cría debido a su alto contenido de proteínas y factores antinutricionales relativamente bajos. Sin embargo, China depende en gran medida de las importaciones de soja y de la inestabilidad de los precios, lo que limita gravemente el crecimiento de la industria ganadera. Por lo tanto, reducir el uso de proteína bruta (PC) en las dietas y desarrollar y utilizar nuevas fuentes de proteínas son esenciales para aliviar la escasez de recursos proteicos.
Las dietas bajas en proteínas se basan en el modelo de proteína ideal mediante la adición de ciertos aminoácidos sintéticos para compensar la reducción de proteínas, de modo que se cumplan el tipo, la cantidad y la proporción de aminoácidos necesarios para el crecimiento animal, lo que puede mejorar la conversión alimenticia, la deposición de nitrógeno animal y las emisiones, al tiempo que garantiza que se mantenga el rendimiento normal de la producción del ganado y las aves de corral (4-6). En comparación con las raciones tradicionales, las dietas bajas en proteínas pueden reducir los costos de alimentación, mejorar la salud de los animales y tener grandes ventajas en el área de la cría saludable y respetuosa con el medio ambiente (7, 8). Con el apoyo de las políticas nacionales, las dietas bajas en proteínas se han utilizado ampliamente en la industria de la cría de China.
Los microorganismos como levaduras, algas, hongos y bacterias pueden producir proteínas unicelulares (SCP) que contienen altas cantidades de proteínas, vitaminas y minerales. El SCP puede sustituir eficazmente las materias primas proteicas como la harina de pescado y la harina de soja (9, 10). Además, la producción de SCP no está limitada por el tiempo y la geografía y puede ser producida en fábrica a gran escala (11). Numerosos estudios han confirmado que el SCP se puede utilizar en dietas de pollos de engorde para reducir la necesidad de harina de soja. Además, la cantidad adecuada de SCP puede aumentar el rendimiento de los músculos de la pechuga y las patas, al tiempo que mejora el aumento de peso, la FCR y la eficiencia económica de los pollos de engorde (12-14).
La NAC producida por Clostridium autoethanogenum (CA) es un SCP con un perfil de aminoácidos relativamente equilibrado (15, 16). Tiene potencial como sustituto de las materias primas proteicas en el ganado, las aves de corral y los productos acuáticos. La investigación ha demostrado que la NAC puede servir como un reemplazo viable de la harina de pescado debido a su alto contenido de proteínas y aminoácidos, la falta de factores antinutricionales y el bajo contenido de aminas biógenas (17, 18). Además, la AC puede convertir la materia inorgánica en materia orgánica y disminuir la descarga de desechos industriales, lo que la convierte en un valioso contribuyente a la promoción de un desarrollo económico sostenible y respetuoso con el medio ambiente (19).
La vía mTOR es responsable de la detección de nutrientes y regula el crecimiento y el metabolismo de los organismos (20-22). La inhibición de la vía mTOR reduce la síntesis de proteínas, mientras que su activación promueve la síntesis de proteínas (23, 24). Además, promueve el crecimiento y la proliferación celular al aumentar la síntesis de proteínas y lípidos y disminuir la autofagia celular (25). Numerosos estudios en acuicultura han demostrado que la NAC puede regular la vía de señalización mTOR, mejorar el crecimiento de los organismos y mejorar el rendimiento del crecimiento (26-28). Sin embargo, hay una falta de investigación sobre la NAC en el ganado y las aves de corral, y los mecanismos de regulación del metabolismo y el crecimiento siguen sin estar claros. Por lo tanto, el estudio de los efectos y los mecanismos reguladores de la alimentación con NAC en pollos de engorde puede proporcionar apoyo teórico para la aplicación de NAC en pollos de engorde, lo cual es de gran valor de investigación e importancia teórica. Planteamos la hipótesis de que las proporciones adecuadas de NAC podrían regular el crecimiento de los pollos de engorde a través de la vía de señalización mTOR. Por lo tanto, se llevó a cabo un ensayo de alimentación utilizando cuatro gradientes de CAP (CAP0, CAP2, CAP3 y CAP4) para evaluar sus efectos sobre el rendimiento de los pollos de engorde, la calidad muscular, los índices séricos y la vía mTOR.
Materiales y métodos
Diseño experimental y dietas
En nuestro experimento, un total de 480 pollos de engorde Arbor Acres (AA) de un día de edad (peso corporal inicial de 43,84 ± 1,38 g) comprados a Zhenghe Poultry Industry Co., Ltd. en la ciudad de Shangqiu se dividieron aleatoriamente en cuatro grupos de tratamiento, con 12 réplicas de 10 aves por tratamiento (mitad macho y mitad hembra). Los pollos de engorde fueron alimentados con cuatro dietas isoenergéticas e isonitrogenadas que contenían NAC a 0 g/kg (Control, CAP0), 20 g/kg (CAP2), 30 g/kg (CAP3) y 40 g/kg (CAP4) respectivamente, y la formulación experimental del alimento cumplió con la DB22_T3207-2020 «Especificaciones técnicas para la dieta baja en proteínas para pollos de engorde». El experimento constó de dos etapas. La dieta de la etapa inicial se administró a los sujetos de 1 a 21 días, mientras que la dieta de la etapa posterior se administró de 22 a 42 días. Para conocer la composición nutricional y los ingredientes de las dietas, consulte las Tablas 1 y 2. La temperatura se mantuvo en 33 °C durante los tres primeros días, después de lo cual disminuyó gradualmente en 1 °C cada dos días hasta alcanzar los 20 °C. El período de luz consistió en tres días consecutivos después de la eclosión, seguidos de 23 h de luz y 1 h de oscuridad durante el resto del período experimental. Los pollos de engorde fueron vacunados de acuerdo con el esquema de inmunización y tuvieron libre acceso a alimento y agua fresca, mientras que la humedad relativa se mantuvo en 60%.
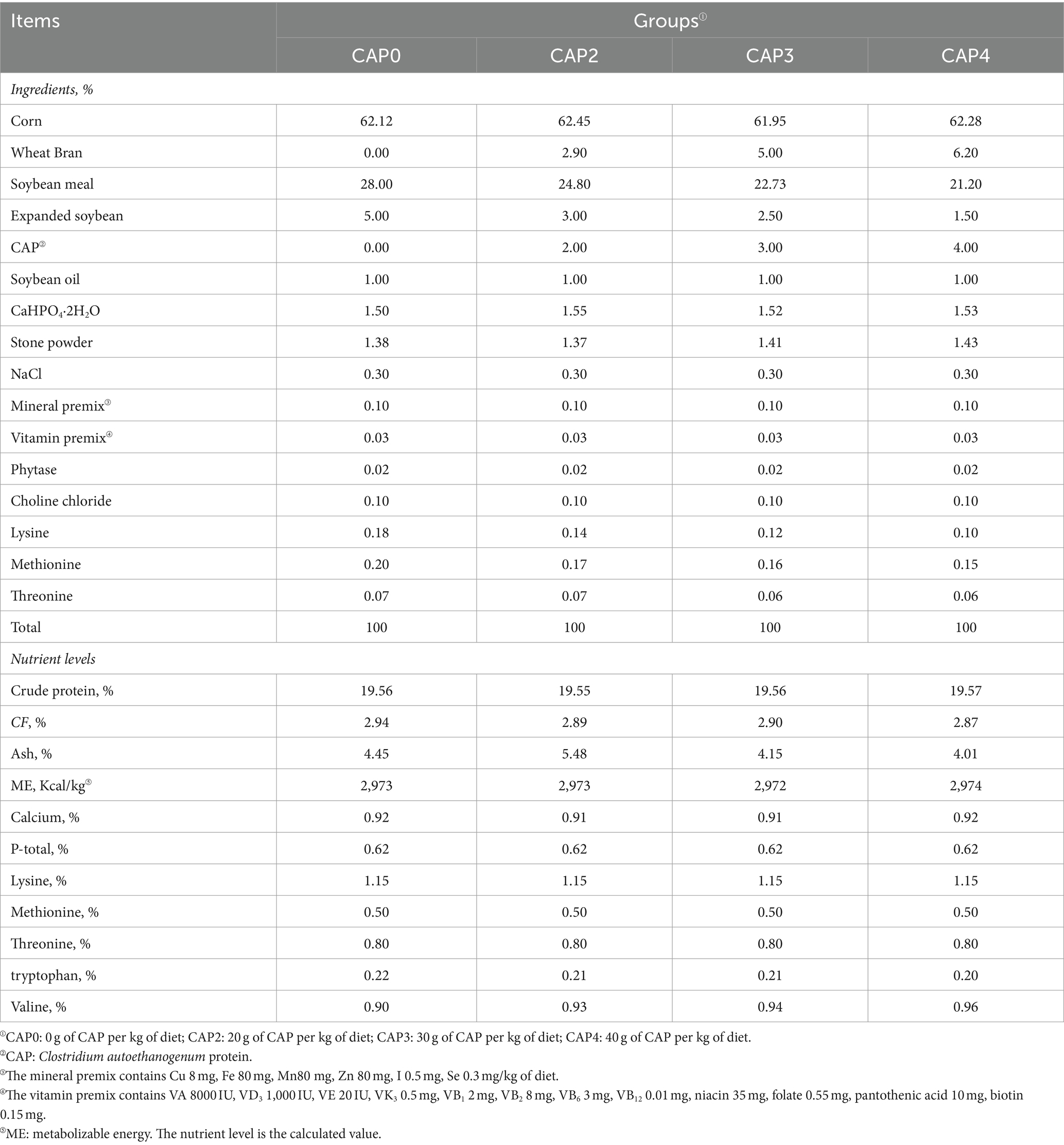 Tabla 1. Composición y contenido de nutrientes de las dietas para pollos de engorde de 1 a 21 d (base secado al aire) %.
Tabla 1. Composición y contenido de nutrientes de las dietas para pollos de engorde de 1 a 21 d (base secado al aire) %.
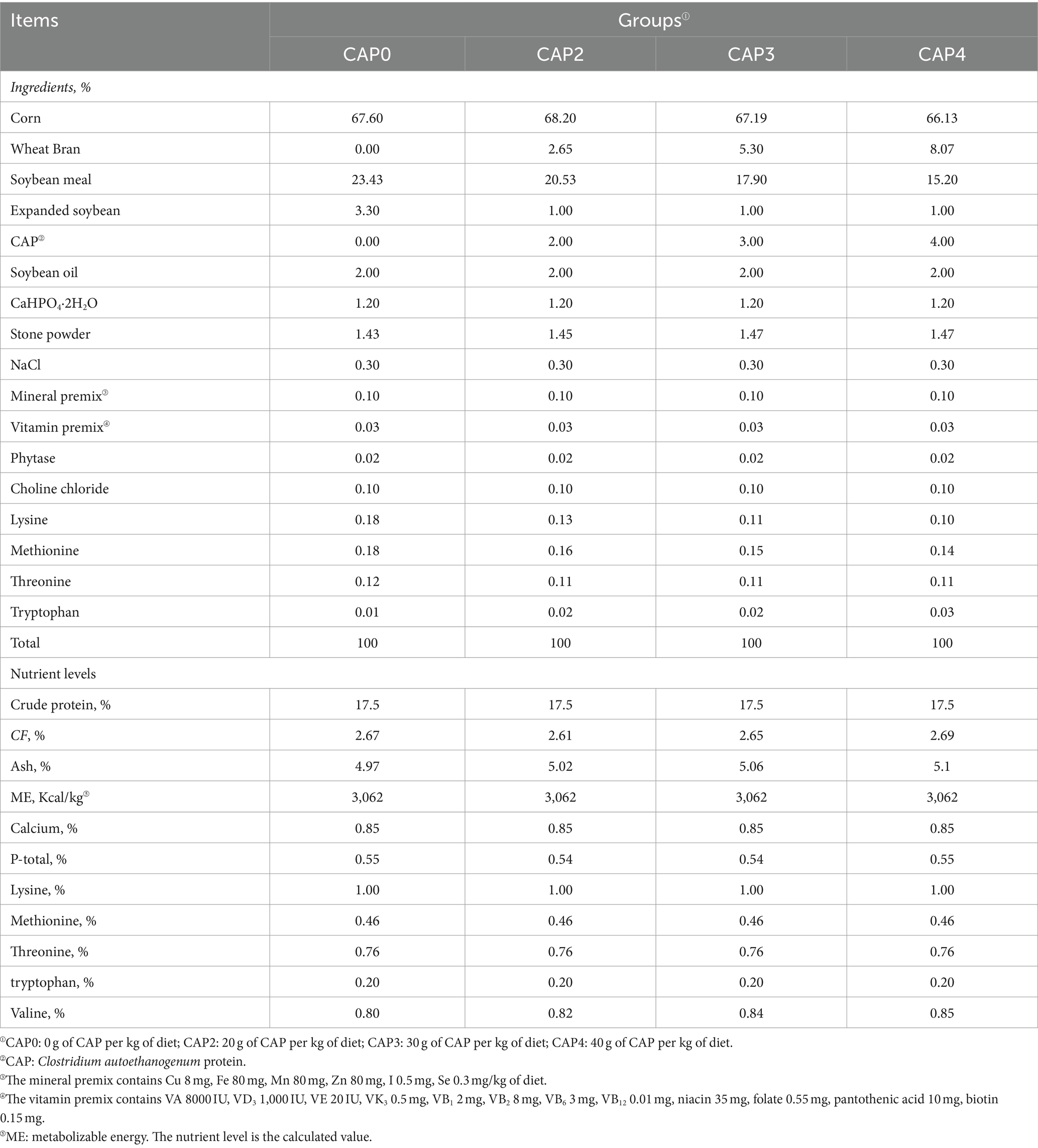 Tabla 2. Composición y contenido de nutrientes de las dietas para pollos de engorde de 22 a 42 días (base secada al aire) %.
Tabla 2. Composición y contenido de nutrientes de las dietas para pollos de engorde de 22 a 42 días (base secada al aire) %.
Rendimiento de la producción
Al final de 1, 21 y 42 días, se pesaron los pollos (después de un ayuno de 12 h) y se registró el consumo de alimento y la cantidad de pollos de engorde muertos para cada corral de tratamiento para el cálculo de la ganancia diaria promedio (GMD), la ingesta diaria promedio de alimento (IDF) y la FCR. A los 42 d, se recolectaron muestras de sangre de 24 pollos de engorde (6 pollos seleccionados al azar para cada tratamiento, mitades macho y hembra) a través del sangrado de la vena yugular. Tras la obtención de la muestra de sangre, se centrifugó a 3.000 g a 4°C durante 10 min. De esta forma, se obtiene el sérum. Después de la sangría de la vena yugular y la ejecución, el rendimiento del sacrificio se evaluó según el estándar NYT823-2020.
Composición muscular
Las muestras de músculo mamario se hornearon en un horno a 65 °C para hacer muestras secadas al aire, y luego se analizaron de acuerdo con el método estándar (29). Brevemente, las muestras se secaron en un horno a 105°C hasta peso constante para el cálculo de la materia seca. El contenido de PB se determinó por el método de nitrógeno Kjeldahl. El contenido de lípidos crudos se midió mediante extracción con éter utilizando un extractor Soxhlet. El contenido de cenizas se determinó mediante combustión a 550 °C en un horno de mufla durante 6 h.
Índices bioquímicos y antioxidantes séricos
La alanina aminotransferasa (ALT), la aspartato transaminasa (AST), la fosfatasa alcalina (ALP), la lactato deshidrogenasa (LDH), la amilasa (AMS), la proteína total (TP), la albúmina (ALB), el ácido úrico (UA), la glucosa (GLU), los triglicéridos (TG), el colesterol total (CT), las lipoproteínas de alta densidad (HDL), las lipoproteínas de baja densidad (LDL), la capacidad antioxidante total (T-AOC), la catalasa (CAT), el malondialdehído (MDA), la glutatión peroxidasa (GSH-Px) y la superóxido dismutasa (SOD) se determinaron mediante kits de ensayo (Nanjing Jiancheng Co., Nanjing, China).
Determinación de índices inmunológicos
La procalcitonina (PCT), la inmunoglobulina A (IgA), la inmunoglobulina G (IgG), la interleucina-1 (IL-1), la interleucina-6 (IL-6) y la interleucina-8 (IL-8) se determinaron mediante kits de ensayo (Shanghai Enzyme Link Biotechnology Co., Shanghai, China).
2.6 Extracción de ARN, síntesis de ADNc y PCR en tiempo real
El ARN se extrajo por primera vez de muestras de músculo mamario e hígado utilizando el kit RNAiso Plus (Takara, Japón). A continuación, se evaluaron las concentraciones y la OD260/OD280 del ARN utilizando un espectrofotómetro Micro-Drop (BIO-DL, Estados Unidos), y se sintetizó ADN complementario (ADNc) utilizando un kit de reactivos PrimeScript™ RT (Takara, Japón). El sistema de reacción se preparó de acuerdo con el kit de PCR cuantitativa fluorescente. Las siguientes fueron las condiciones de termociclado para los genes diana: 95 °C durante 30 s, 95 °C durante 15 s y 60 °C durante 30 s para recoger las señales de fluorescencia, y se realizaron 40 ciclos. La cuantificación de la expresión génica relativa se realizó utilizando 2-ΔΔCT. Los cebadores dirigidos para varios genes en este estudio se indican en la Tabla 3.
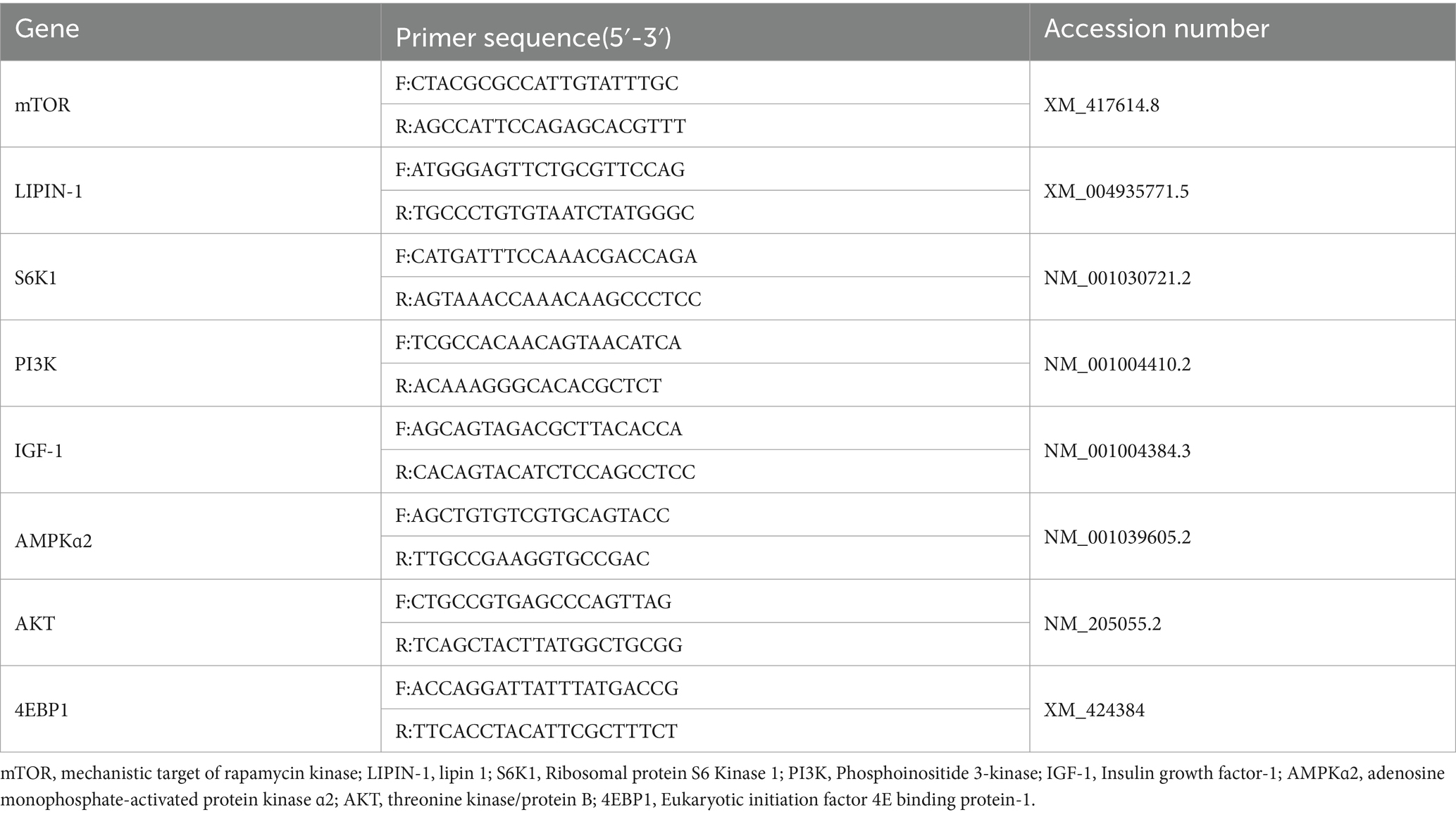 Tabla 3. Secuencias de cebadores.
Tabla 3. Secuencias de cebadores.
2.7 Extracción de proteínas y Western blot
De acuerdo con la masa y el volumen de cada muestra, agregue el volumen correspondiente de lisado, déjelo reposar durante 5 minutos, centrifugue a 12,000 r durante 10 minutos y el sobrenadante es el extracto de proteína. La concentración de proteínas de las muestras se determinó de acuerdo con las instrucciones del BCA Protein Concentration Assay Kit (Solarbio Beijing, China). Después de determinar la cantidad de proteína a cargar, diluir las muestras de proteína con tampón de carga de 5 × y PBS y hervirlas a 100 °C durante 5 min. Se cargaron cantidades iguales de proteína en los pocillos del gel SDS-PAGE, y las proteínas se transfirieron del gel a la membrana de PVDF. Posteriormente, se cerró la membrana con leche desnatada en polvo al 5% durante una hora. Después de la incubación con anticuerpos primarios y secundarios, la membrana de PVDF se colocó en un aparato de exposición química, rociada uniformemente con una solución quimioluminiscente ECL, y el tiempo de exposición se estableció para exponer la membrana. Los resultados se analizaron utilizando un sistema de procesamiento de imágenes en gel (ImageJ) para analizar los valores de la escala de grises de las bandas objetivo. Los anticuerpos utilizados en el presente estudio fueron: fosfo-mTOR (Ser2448), mTOR, 4EBP1, AKT y fosfo-AKT (Ser473) adquiridos al Grupo Proteintech; fosfo-p70S6 quinasa (Thr389), AMPK alfa-2 y fosfo-AMPK alfa-1,2 (Thr172, Thr183) compradas a Thermo Fisher Scientific; p70S6 Kinasa comprada a Affinity; phospho-4E-BP1 (Thr37/46) (236B4) comprado a la tecnología de señalización celular.
Análisis estadístico
Los datos experimentales se cotejaron con Excel 2021 y se realizó la prueba de Shapiro-Wilk, seguida de ANOVA de una vía (ANOVA de una vía) utilizando SPSS 26.0 y el método de Duncan para comparaciones múltiples. Los valores se presentaron como media ± SEM, y las diferencias se consideraron estadísticamente significativas a p < 0,05.
Resultados
Efectos del uso de NAC en las dietas sobre el rendimiento del crecimiento de los pollos de engorde
En la Tabla 4 se muestran los efectos de las diferentes proporciones de NAC añadidas a las dietas sobre el rendimiento del crecimiento de los pollos de engorde. A los 22-42 días, la FCR fue menor en el grupo CAP3 que en el grupo CAP0 (p < 0,05). A los 1-42 días, la FCR fue menor en los grupos CAP3 y CAP4 que en el grupo CAP0 (p < 0,05).
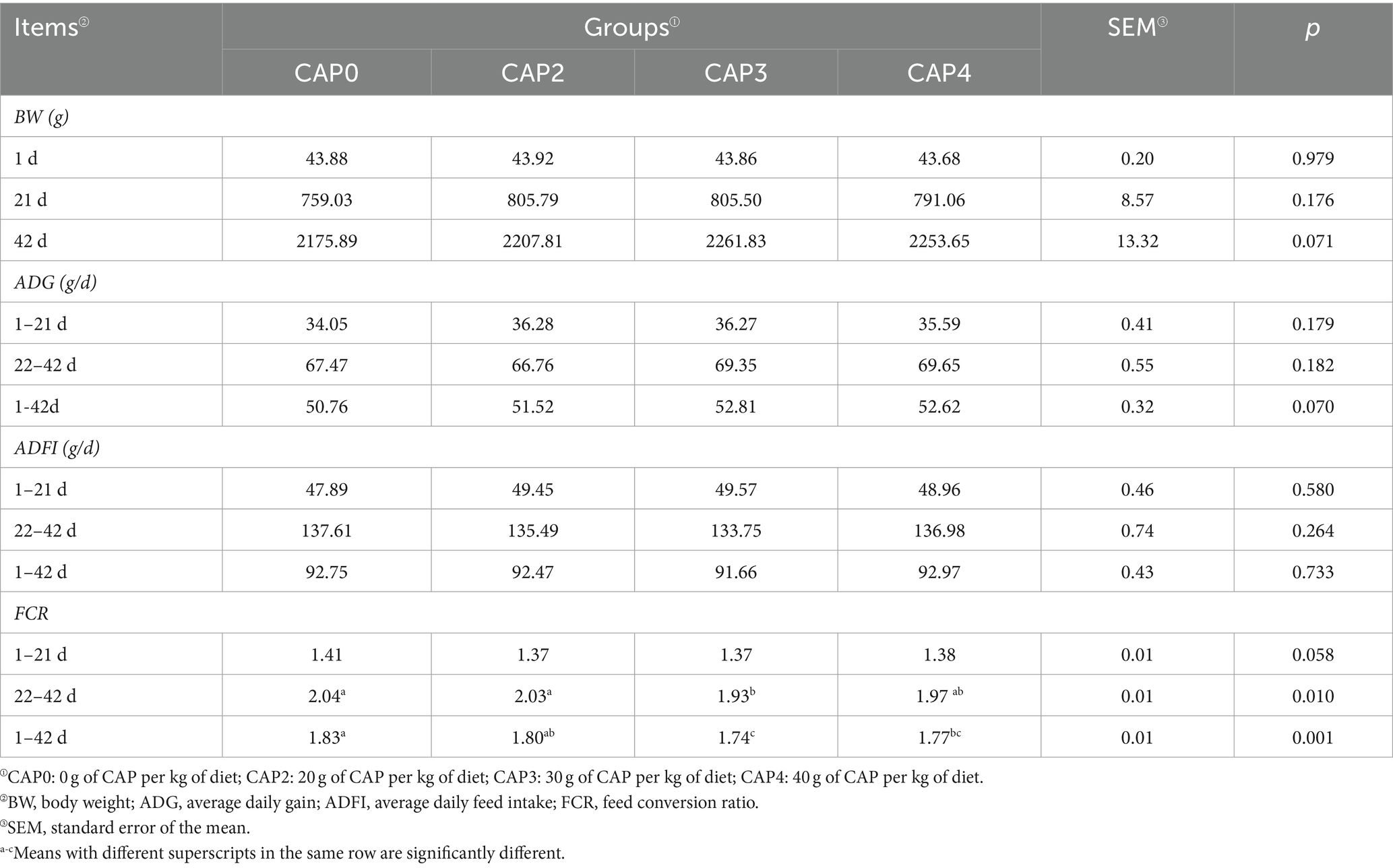 Tabla 4. Efectos del uso de NAC en dietas sobre el rendimiento del crecimiento de pollos de engorde.
Tabla 4. Efectos del uso de NAC en dietas sobre el rendimiento del crecimiento de pollos de engorde.
Efectos del uso de NAC en dietas sobre el rendimiento de sacrificio de pollos de engorde
Como se muestra en la Tabla 5, la NAC no tuvo un efecto significativo sobre el porcentaje de aderezo, el porcentaje de rendimiento semieviscerado y el porcentaje de rendimiento eviscerado (p > 0.05), pero sí tuvo un impacto en el rendimiento muscular del pecho y el rendimiento muscular de la pierna (p < 0.05). El rendimiento muscular de la pierna del grupo CAP3 fue mayor que el de los otros grupos (p < 0,05), y el rendimiento muscular de la pierna del grupo CAP0 fue menor que el de los grupos CAP3 y CAP4 (p < 0,05).
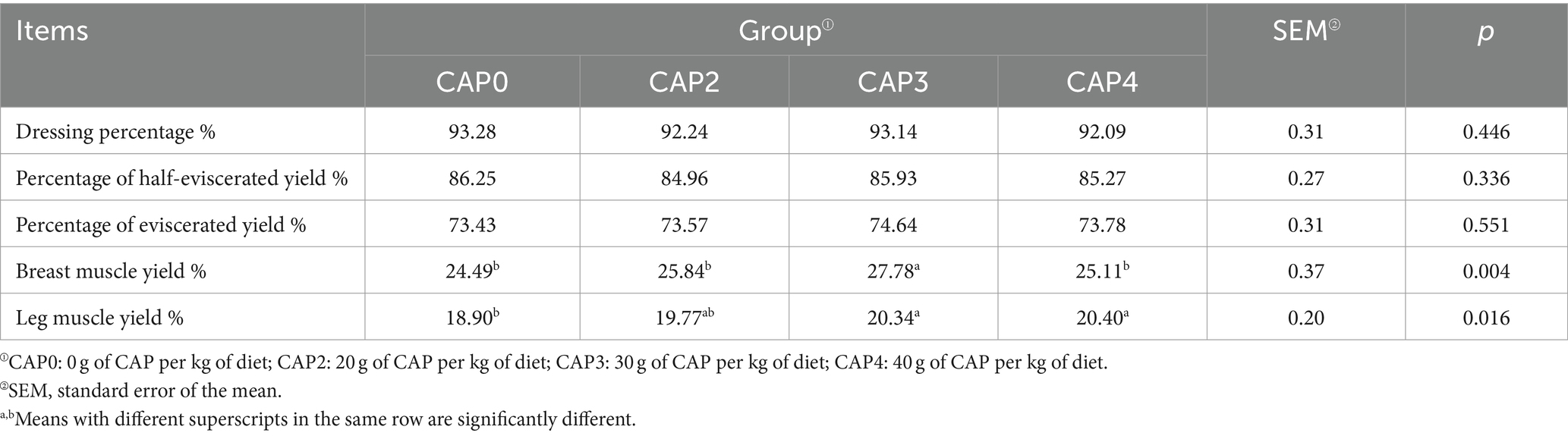 Tabla 5. Efectos del uso de NAC en dietas sobre el rendimiento del sacrificio de pollos de engorde.
Tabla 5. Efectos del uso de NAC en dietas sobre el rendimiento del sacrificio de pollos de engorde.
Efectos del uso de NAC en dietas sobre la composición muscular de pollos de engorde
En la Tabla 6 se describen los efectos de la adición de NAC sobre la calidad de la carne y la composición de los pollos de engorde. El contenido de proteína bruta muscular en el grupo CAP0 fue menor que en el grupo CAP3 (p < 0,05).
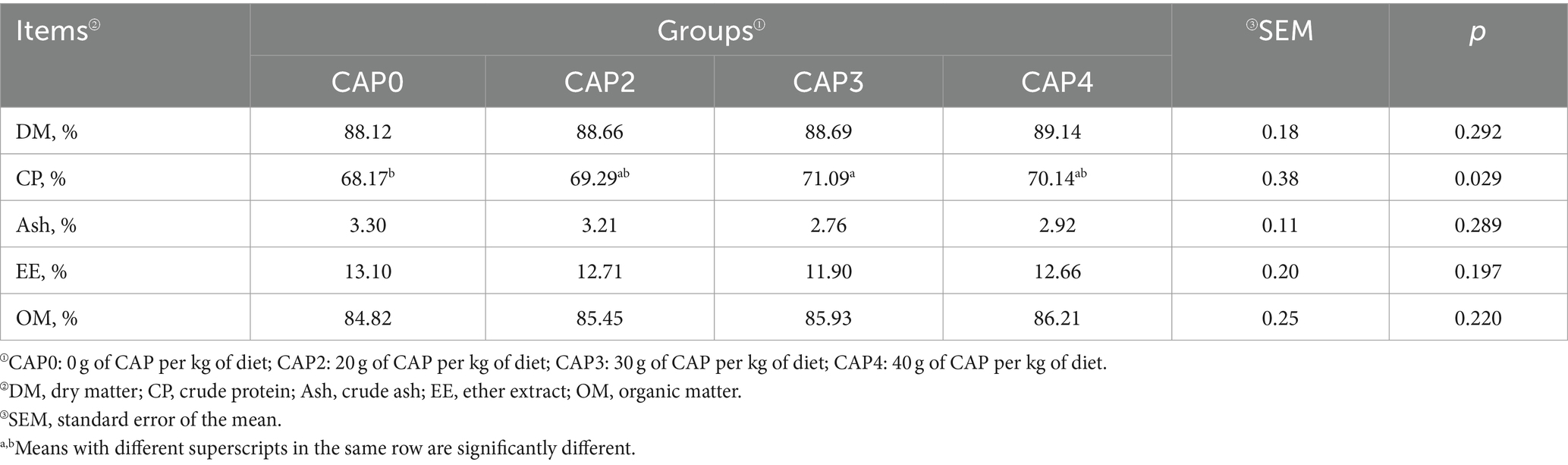 Tabla 6. Efectos del uso de NAC en dietas sobre la composición muscular de pollos de engorde.
Tabla 6. Efectos del uso de NAC en dietas sobre la composición muscular de pollos de engorde.
Efectos del uso de NAC en dietas sobre los índices bioquímicos séricos de pollos de engorde
Como se muestra en la Tabla 7, los niveles de inclusión de NAC en la dieta influyeron en los índices bioquímicos séricos de los pollos de engorde. Los grupos CAP0 y CAP4 presentaron niveles de TP más bajos que el grupo CAP3 (p < 0,05). Los pollos de engorde alimentados con dietas CAP0 y CAP2 tuvieron un menor contenido de UA en comparación con las dietas CAP3 y CAP4 (p < 0,05). El grupo CAP3 tuvo los niveles más bajos de TG, seguido por el grupo CAP4. La actividad de AST en CAP4 fue mayor que en los grupos CAP0 y CAP2 (p < 0,05).
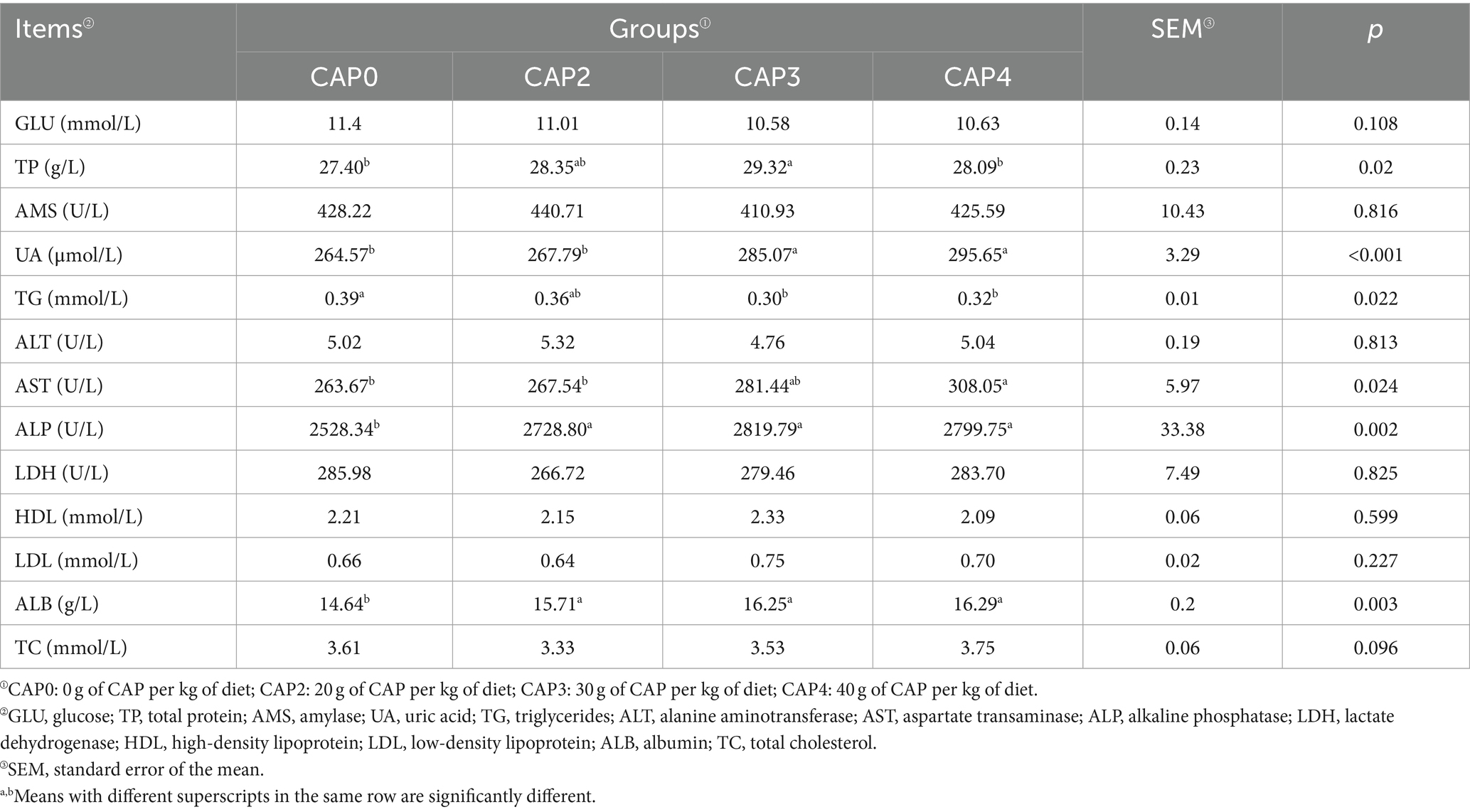 Tabla 7. Efectos del uso de NAC en dietas sobre los índices bioquímicos séricos de pollos de engorde.
Tabla 7. Efectos del uso de NAC en dietas sobre los índices bioquímicos séricos de pollos de engorde.
Efectos del uso de CAP en dietas sobre los índices antioxidantes en pollos de engorde
Como se muestra en la Tabla 8, el T-AOC en el grupo CAP0 fue menor que en los grupos CAP2 y CAP3 (p < 0,05). En cuanto al GSH-Px, los grupos CAP2, CAP3 y CAP4 fueron mayores que el grupo CAP0 (p < 0,05). La SOD fue mayor en el grupo CAP3 que en los grupos CAP0 y CAP4 (p < 0,05).
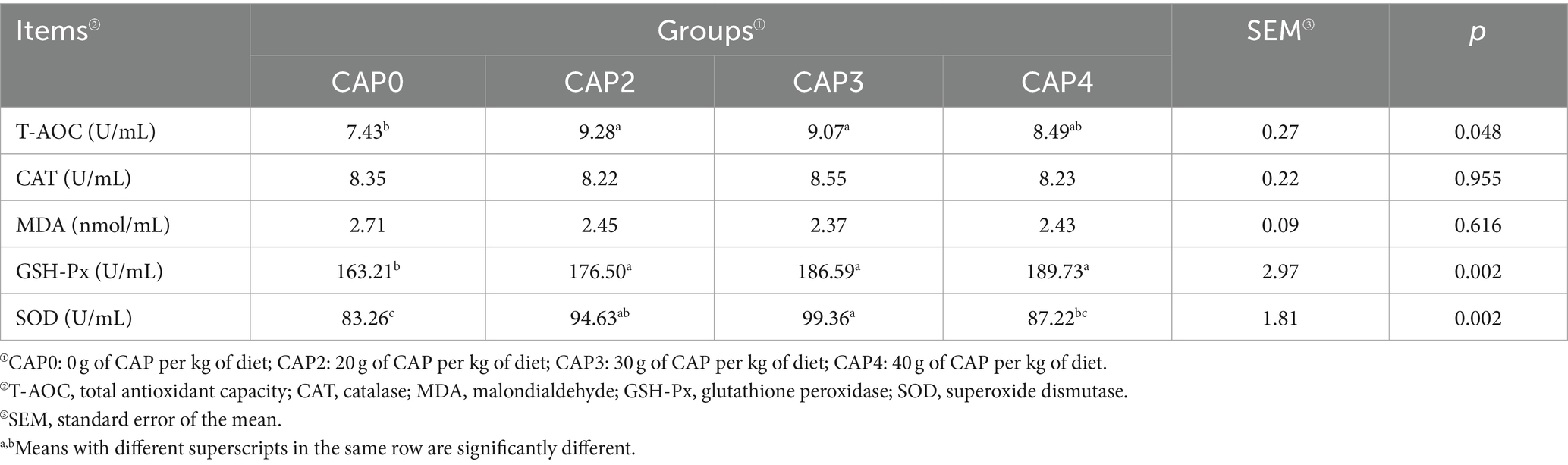 Tabla 8. Efectos del uso de NAC en dietas sobre los índices séricos de antioxidantes de pollos de engorde.
Tabla 8. Efectos del uso de NAC en dietas sobre los índices séricos de antioxidantes de pollos de engorde.
Efectos del uso de NAC en dietas sobre factores inmunes e inflamatorios en pollos de engorde
La Tabla 9 muestra los efectos de la adición de NAC sobre los factores inmunológicos e inflamatorios del pollo de engorde. Los pollos de engorde alimentados con dietas CAP3 y CAP4 tuvieron niveles significativamente más altos de IgG en comparación con los alimentados con dietas CAP0 y CAP2 (p < 0,05).
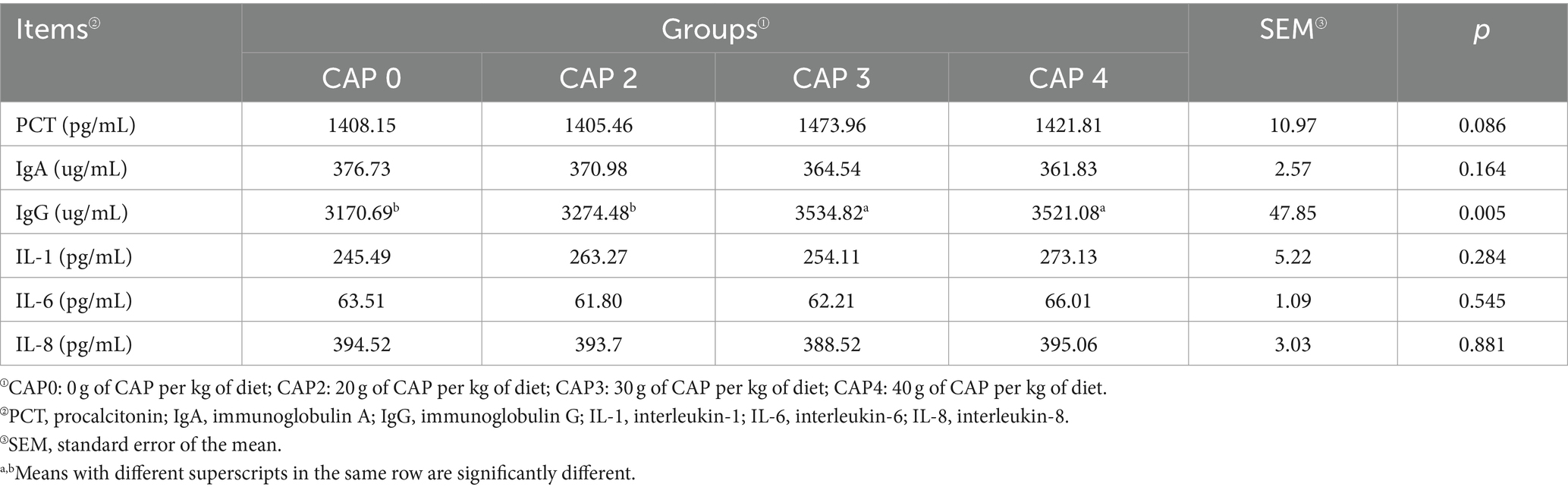 Tabla 9. Efectos del uso de NAC en dietas sobre los factores inmunes e inflamatorios séricos en pollos de engorde.
Tabla 9. Efectos del uso de NAC en dietas sobre los factores inmunes e inflamatorios séricos en pollos de engorde.
Expresión génica asociada a dianas de mamíferos de la vía de señalización mTOR en el músculo de la mama y el hígado
Los niveles de ARNm de los genes relacionados con la vía de señalización mTOR en el músculo pectoral se muestran en la Figura 1. El nivel de expresión de Lipin-1 fue menor en los grupos CAP2, CAP3 y CAP4 en comparación con el grupo CAP0, con el nivel más bajo observado en el grupo CAP3 (p < 0,05). Por el contrario, la expresión de AKT fue mayor en los grupos CAP4 que en los grupos CAP0 (p < 0,05). Las dietas alimentadas con pollos de engorde CAP4 tuvieron una mayor expresión de PI3K que las alimentadas con dieta CAP0 (p < 0,05). Los pollos de engorde alimentados con dietas CAP3 y CAP4 mostraron un mayor nivel de ARNm de AMPKɑ2 y 4EBP1 en comparación con las dietas alimentadas con pollos de engorde CAP0 y CAP2 (p < 0,05). Además, se observó una diferencia entre los grupos CAP3 y CAP4 en el gen AMPKɑ2 (p < 0,05).
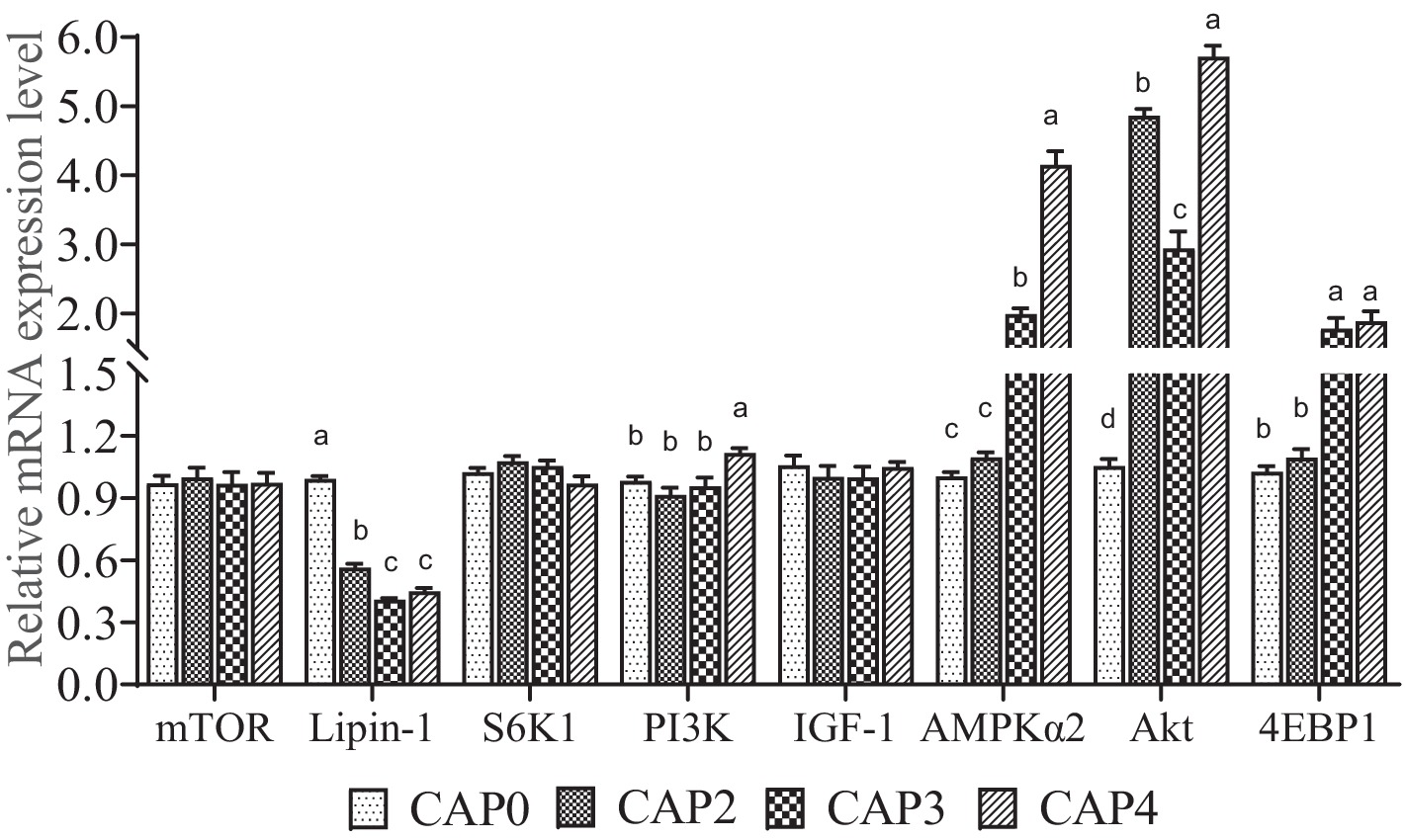 Figura 1. Efectos de la CAP en el patrón de expresión de genes clave de la vía mTOR en el músculo mamario. Los datos se presentan como medias y errores estándar (±EE) (n = 6).
Figura 1. Efectos de la CAP en el patrón de expresión de genes clave de la vía mTOR en el músculo mamario. Los datos se presentan como medias y errores estándar (±EE) (n = 6).
Los niveles de ARNm de los genes relacionados con la vía de señalización mTOR en el hígado se muestran en la Figura 2. Las dietas alimentadas con pollos de engorde CAP3 y CAP4 mostraron un nivel más alto de ARNm de mTOR que las dietas alimentadas con pollos de engorde CAP0 y CAP2 (p < 0,05). Además, el grupo CAP3 fue mayor que el CAP4 (p < 0,05). El nivel de ARNm de Lipin-1 en el grupo CAP0 fue mayor que el de los otros grupos (p < 0,05), y el grupo CAP4 tuvo una expresión significativamente mayor de AMPKɑ2 que los otros grupos (p < 0,05), lo que es consistente con la tendencia en el músculo pectoral. El nivel relativo de expresión de ARNm de Akt fue mayor en los grupos CAP2 y CAP4 en comparación con los grupos CAP0 y CAP3 (p < 0,05). Las expresiones de 4E-BP1 aumentaron en los grupos de tratamiento en comparación con el grupo control, y el grupo CAP3 fue mayor que los otros grupos (p < 0,05).
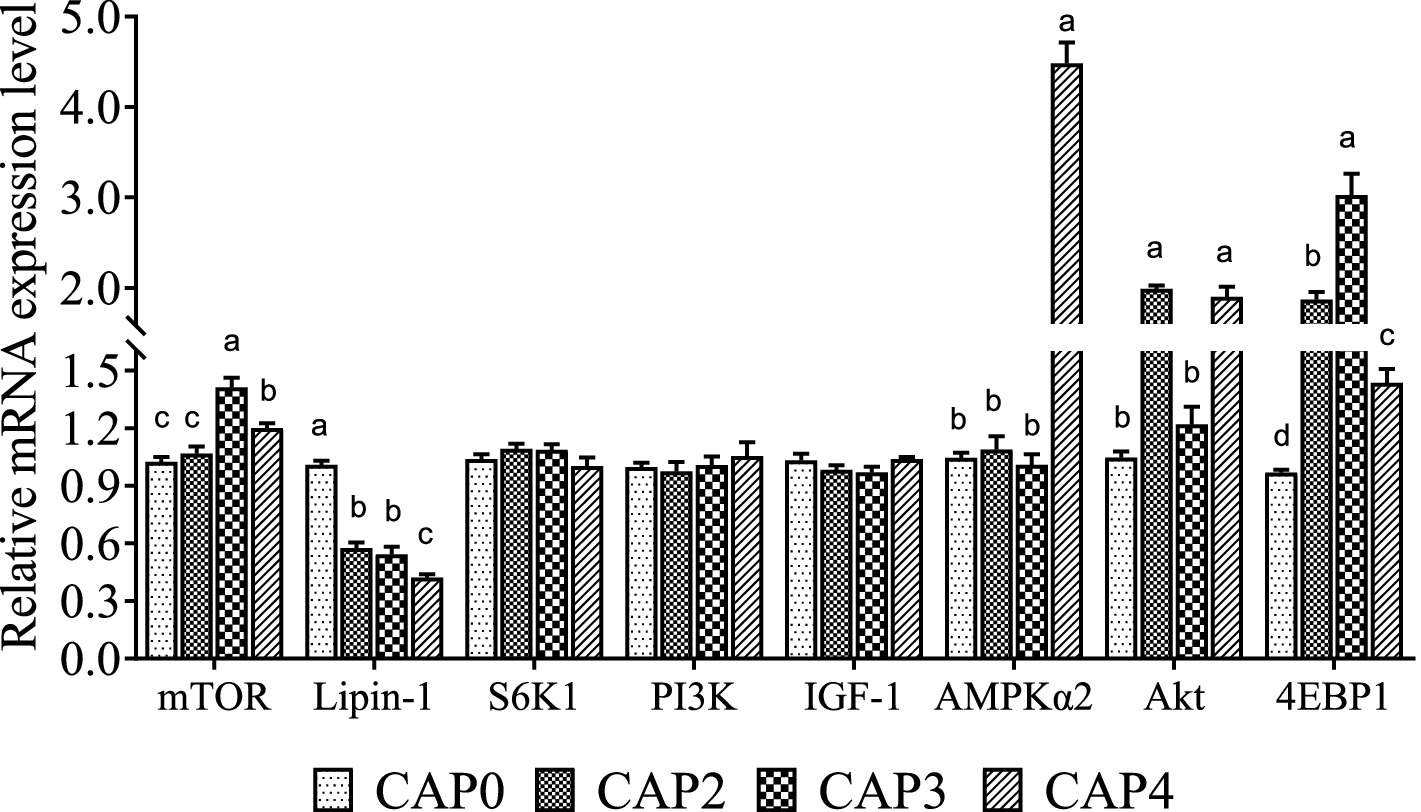 Figura 2. Efectos de la CAP en el patrón de expresión de genes clave de la vía mTOR en el hígado. Los datos se presentan como medias y errores estándar (±EE) (n = 6). Las diferencias significativas (p < 0,05) se indican mediante diferentes superíndices alfabéticos sobre las barras, mientras que la misma letra indica que no hay diferencia.
Figura 2. Efectos de la CAP en el patrón de expresión de genes clave de la vía mTOR en el hígado. Los datos se presentan como medias y errores estándar (±EE) (n = 6). Las diferencias significativas (p < 0,05) se indican mediante diferentes superíndices alfabéticos sobre las barras, mientras que la misma letra indica que no hay diferencia.
Expresión de proteínas asociadas a dianas de mamíferos de la vía de señalización mTOR en el músculo de la mama y el hígado
El presente estudio evaluó la fosforilación proteica de reguladores clave en la vía de señalización mTOR en el músculo mamario (Figura 3). Los niveles de fosforilación de 4E-BP1 y Akt mostraron la misma tendencia y fueron mayores en el grupo CAP3 que en los otros grupos (p < 0,05). Los niveles de fosforilación de AMPKɑ2 fueron mayores en los grupos CAP2 y CAP3 en comparación con el grupo control (p < 0,05).
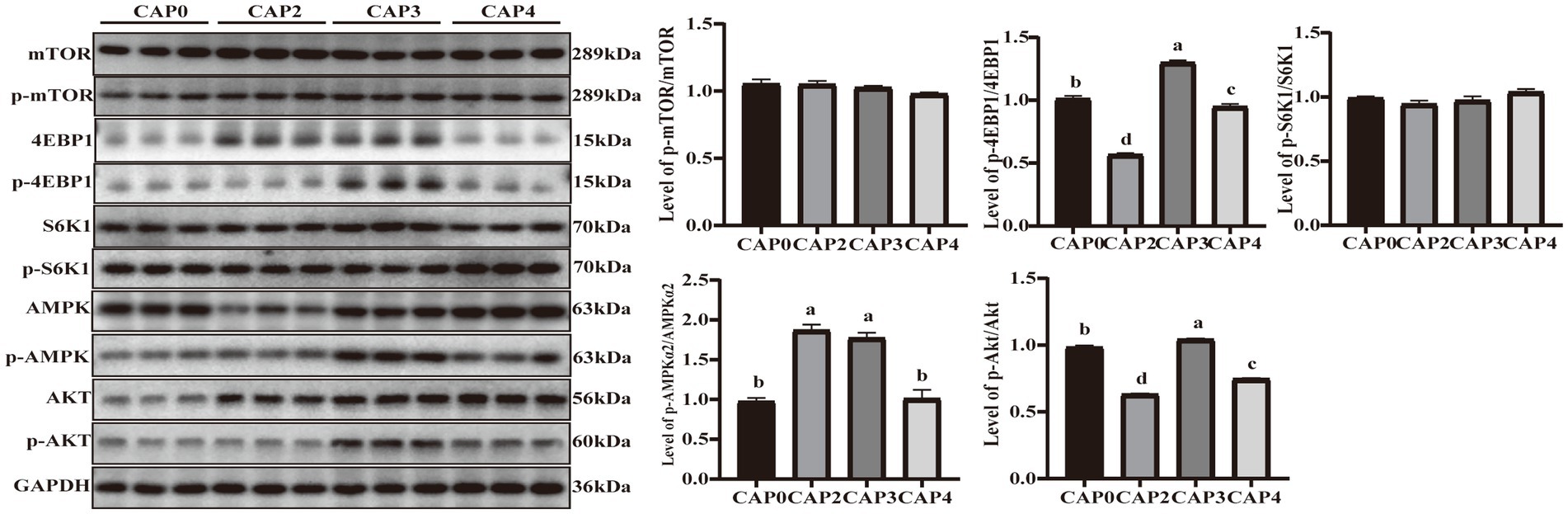 Figura 3. Niveles de expresión de proteínas y niveles de fosforilación de algunos genes seleccionados del músculo mamario implicados en la síntesis de proteínas y el metabolismo energético en pollos de engorde alimentados con dietas NAC. Los datos se presentan como medias y errores estándar (±EE) (n = 6). Las diferencias significativas (p < 0,05) se indican mediante diferentes superíndices alfabéticos sobre las barras, mientras que la misma letra indica que no hay diferencia.
Figura 3. Niveles de expresión de proteínas y niveles de fosforilación de algunos genes seleccionados del músculo mamario implicados en la síntesis de proteínas y el metabolismo energético en pollos de engorde alimentados con dietas NAC. Los datos se presentan como medias y errores estándar (±EE) (n = 6). Las diferencias significativas (p < 0,05) se indican mediante diferentes superíndices alfabéticos sobre las barras, mientras que la misma letra indica que no hay diferencia.
En la Figura 4 se presenta el nivel de fosforilación de proteínas de los reguladores clave en la señalización de mTOR en comparación con los diferentes tratamientos en el hígado. Los niveles de fosforilación de mTOR, 4EBP1 y S6K1 en los grupos CAP3 y CAP4 fueron mayores que en los grupos CAP0 y CAP2 (p < 0,05). El nivel de fosforilación de AMPKɑ2 fue mayor en el grupo CAP2 que en los otros grupos (p < 0,05).
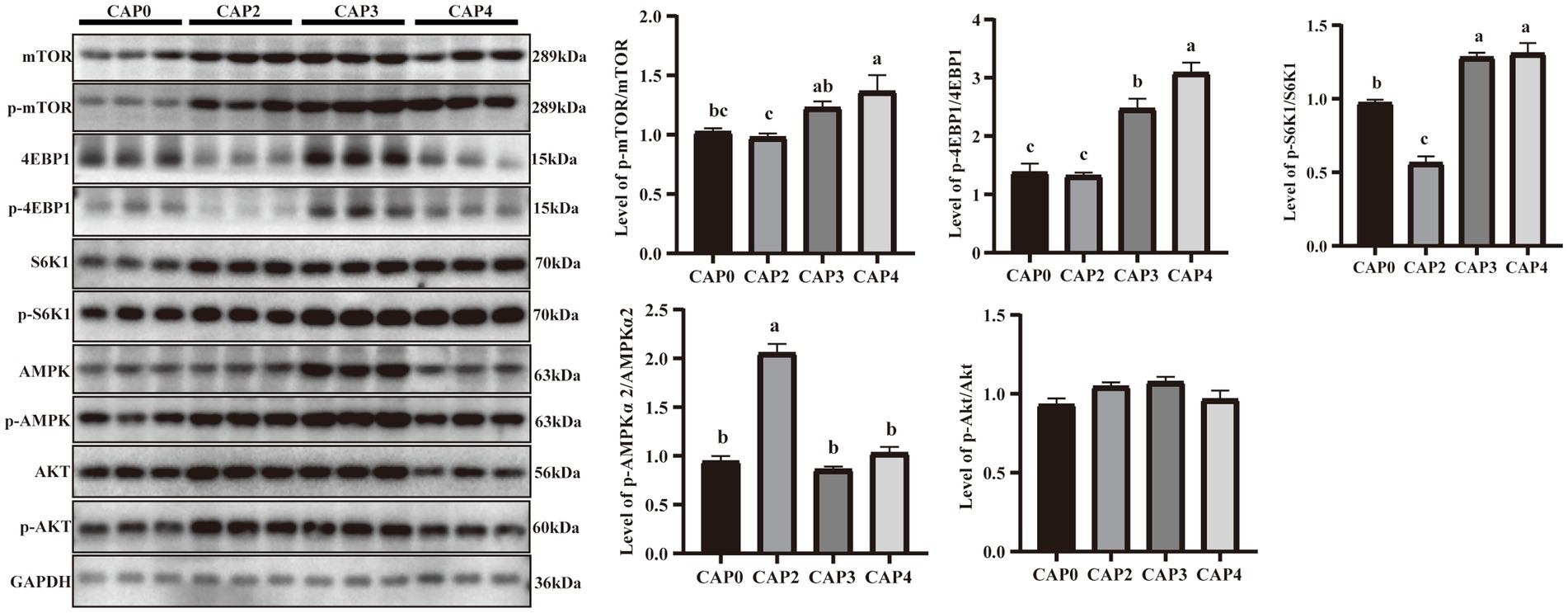 Figura 4. Niveles de expresión proteica y niveles de fosforilación de algunos genes hepáticos seleccionados implicados en la síntesis proteica y el metabolismo energético en pollos de engorde alimentados con dietas NAC. Los datos se presentan como medias y errores estándar (±EE) (n = 6). Las diferencias significativas (p < 0,05) se indican mediante diferentes superíndices alfabéticos sobre las barras, mientras que la misma letra indica que no hay diferencia.
Figura 4. Niveles de expresión proteica y niveles de fosforilación de algunos genes hepáticos seleccionados implicados en la síntesis proteica y el metabolismo energético en pollos de engorde alimentados con dietas NAC. Los datos se presentan como medias y errores estándar (±EE) (n = 6). Las diferencias significativas (p < 0,05) se indican mediante diferentes superíndices alfabéticos sobre las barras, mientras que la misma letra indica que no hay diferencia.
Discusión
En la actualidad, la investigación sobre la PAC en el ganado y las aves de corral es limitada, y la mayoría de los estudios se centran en los animales acuáticos. Varios estudios han demostrado que una proporción adecuada de PAC puede mejorar el crecimiento y el desarrollo de los animales, mejorar el rendimiento de la producción y promover la buena salud (26, 27, 30, 31). Sin embargo, una alta proporción de sustitución de NAC puede conducir a una disminución en el rendimiento de la producción animal (32, 33), así como a daños intestinales y hepáticos, y a un aumento de la mortalidad (15). Wu et al. (34) encontraron que agregar la cantidad adecuada de NAC en las dietas de pollos de engorde mejoró la tasa de crecimiento y la eficiencia de conversión alimenticia, lo cual es consistente con nuestros hallazgos. Por lo tanto, la función de la NAC puede estar relacionada con la facilidad de digestión (15, 34).
La evaluación del rendimiento del sacrificio es crucial para determinar la calidad de las canales de ganado y aves de corral. Por lo tanto, es importante tener en cuenta el rendimiento de los músculos del pecho y las piernas, ya que son indicadores cruciales del rendimiento de la producción de carne. Investigaciones previas han demostrado que el suministro de un nivel adecuado de SCP en las dietas de los pollos de engorde puede mejorar la calidad de la canal y aumentar el rendimiento muscular de los senos y las piernas (14), lo que es consistente con los resultados de este estudio. Otro estudio reportó que la suplementación de diferentes proporciones de SCP en dietas de pollos de engorde no afectó el porcentaje de aderezo, lo que se alinea con los hallazgos del presente estudio (35). Según el estudio, la inclusión de CAP en la dieta de los pollos de engorde aumentó el contenido de PB en el músculo, lo que sugiere una aceleración de la síntesis y deposición de proteínas. Estos hallazgos son consistentes con estudios previos realizados por Xue et al. (28) y Wu et al. (10) en animales acuáticos.
Los indicadores bioquímicos séricos cambian según la salud y el estado metabólico del cuerpo. La medición de estos indicadores puede reflejar visualmente la salud, la nutrición y el estado metabólico del cuerpo. La TP refleja el estado de absorción y síntesis de proteínas, y un alto contenido de TP indica un metabolismo vigoroso de las proteínas, que puede mejorar la función inmunológica del cuerpo y promover el crecimiento y el desarrollo (36, 37). El ALB desempeña un papel crucial en la estabilización de la osmolalidad sanguínea y en el transporte de sustancias metabólicas (38). La UA es un metabolito proteico producido principalmente en el hígado y excretado a través de los riñones y los intestinos. La hiperuricemia y la gota pueden ser causadas por la sobreproducción o la reducción de la excreción de UA (39). Estudios previos han demostrado que la inclusión de NAC en las dietas puede aumentar los niveles de TP y ALB, y los niveles de AU también tienden a aumentar con el aumento del contenido de NAC (34), lo que es consistente con los resultados del presente estudio. El CT y el TG son los principales lípidos en la sangre e importantes indicadores de la absorción, el metabolismo y el transporte de lípidos (40, 41). En el presente estudio, la NAC tuvo un efecto significativo sobre el metabolismo lipídico de los pollos de engorde, como lo demuestra la disminución de los niveles de TG. Este hallazgo es consistente con los resultados reportados por Wu et al. (34) pero difiere de los observados en animales acuáticos (11, 27), lo que puede atribuirse a las diferencias entre especies.
La AST y la ALT se utilizan a menudo como indicadores de lesión hepática (42). La fosfatasa alcalina actúa como desfosforilador y tiene una función biológica importante, comúnmente utilizada para evaluar la lesión y la enfermedad hepática (43-45). Vale la pena señalar que los niveles de ALP aumentan durante el desarrollo esquelético cuando el organismo está creciendo rápidamente (46). En el presente estudio, la AST y la ALP tendieron a aumentar cuando se agregó NAC a la dieta. Se sugirió que la NAC puede causar cierto grado de daño a la función hepática al tiempo que promueve un rápido crecimiento del organismo. De acuerdo con los resultados del presente estudio, estudios previos han mostrado una tendencia creciente en la AST con el aumento de la NAC (15, 33). Por su parte, Maulu et al. (31) demostraron que el 5 y el 10% de NAC aumentaron la actividad de la FA, lo que es similar a los resultados del presente estudio.
La SOD reduce el daño oxidativo al eliminar los radicales libres y generar moléculas de oxígeno y peróxido de hidrógeno. El T-AOC refleja la capacidad antioxidante total del cuerpo, y el GSH-Px convierte los peróxidos dañinos en compuestos hidroxilo inofensivos, protegiendo la estructura y función normales de las membranas celulares (47). En el presente estudio, la inclusión de CAP en la dieta aumentó las actividades de GSH-Px y SOD, así como T-AOC, lo que indica que la proporción adecuada de CAP puede mejorar la capacidad antioxidante del organismo. Este hallazgo es consistente con estudios previos que han demostrado la capacidad de la NAC para aumentar las actividades de las enzimas antioxidantes y mejorar la capacidad antioxidante del organismo (30, 31, 34). Por el contrario, una alta proporción de NAC puede perjudicar el sistema de defensa antioxidante del organismo, lo que conduce al daño oxidativo (11, 17). Por lo tanto, especulamos que esta situación podría deberse a la relación de sustitución, las diferencias entre especies y la ausencia de nutrientes específicos requeridos por ciertos animales en la NAC. Por ejemplo, se informó que la deficiencia de taurina da lugar a niveles elevados de daño oxidativo en los peces, lo que tiene un impacto perjudicial en la salud y el rendimiento económico. (48).
La IgG es la principal Ig en el organismo, representando alrededor del 75% de la cantidad total de Ig sérica (49), y la IgA es la segunda más abundante, representando el 10-20% de la Ig sérica total (50). Estudios previos han demostrado que la NAC puede regular al alza los indicadores relacionados con el sistema inmunitario y mejorar la función inmunitaria, lo que es consistente con los resultados del presente estudio (18, 26, 31). La IL-1, la IL-6 y la IL-8 son miembros de la familia de las interleucinas y desempeñan un papel crucial en la regulación de la inflamación y las respuestas inmunitarias en el cuerpo (51, 52). Maulu et al. (31) informaron que la NAC no tuvo ningún efecto adverso sobre la regulación de la inflamación y las respuestas inmunes, ya que los niveles de IL-8 no fueron significativamente diferentes del grupo control. Del mismo modo, el presente estudio no encontró diferencias significativas en los niveles de IL-8 entre los grupos, lo que indica que la NAC no causó una respuesta inflamatoria en el cuerpo.
La vía mTOR integra varias señales externas de energía, aminoácidos, factores inflamatorios y hormonas, con una función vital en la detección y regulación de nutrientes (53). Además, desempeña un papel importante en la deposición, el crecimiento y el desarrollo de proteínas, y la regulación metabólica en el cuerpo (54). El aumento de peso de los animales se debe principalmente a la deposición de músculo y grasa, en la que la deposición muscular depende principalmente de la síntesis de proteínas, y la aceleración de la síntesis de proteínas puede promover el rápido crecimiento del organismo (10, 55). Numerosos estudios han demostrado que los genes mTOR, 4E-BP1 y S6K estaban asociados con la síntesis de proteínas. La activación de mTOR se acompañó típicamente de cambios en la expresión de los genes 4E-BP1 y S6K, pero 4E-BP1 juega un papel más importante en la síntesis de proteínas que S6K (10, 27, 56, 57).
Estudios previos han demostrado que una proporción adecuada de NAC puede mejorar la expresión de genes asociados con la vía de señalización mTOR, estimular la síntesis de proteínas musculares y mejorar el rendimiento del crecimiento del organismo (10, 27, 28). Además, Li et al. (26) también encontraron que la inclusión dietética de NAC mejoró los niveles de expresión de genes relacionados con el crecimiento, los antioxidantes y la vía TOR. El presente estudio mostró que el gen Lipin-1 estaba regulado a la baja, y los genes AMPKɑ2, Akt y 4E-BP1 estaban regulados al alza en comparación con el grupo de control, lo que sugiere que la proporción adecuada de CAP activa la vía de señalización mTOR y promueve el crecimiento y desarrollo del organismo, lo que es consistente con estudios anteriores.
Los niveles de fosforilación de S6K1 y 4E-BP1 suelen ser indicadores del grado de activación de la vía mTOR, y la activación de la vía mTOR puede promover la proliferación y el crecimiento celular (56, 58, 59). En el presente estudio, el grupo CAP3 aumentó significativamente los niveles de fosforilación de 4E-BP1, S6K1, Akt y AMPKɑ2, lo que proporciona más evidencia a nivel de expresión de proteínas de que una proporción adecuada de CAP puede activar la vía de señalización de mTOR y regular el crecimiento del organismo. De acuerdo con los resultados de este estudio, Maulu et al. (27) informaron que los niveles de fosforilación de 4E-BP1, S6K1 y AMPK aumentaron significativamente en un 5 y 10% CAP. Sin embargo, a diferencia de los resultados de nuestro estudio, algunos investigadores encontraron que la adición de NAC a la dieta inhibe la vía mTOR y afecta negativamente el crecimiento y desarrollo del organismo (60, 61). Las discrepancias en estos estudios pueden deberse a la especificidad de la especie y a las variaciones en la cantidad de sustitución, y se necesita más investigación para determinar las razones exactas.
La activación de la vía mTOR está estrechamente relacionada con los niveles de aminoácidos (AA). Se ha demostrado que los AA, en particular los aminoácidos de cadena ramificada (BCAA), regulan la síntesis de proteínas a través de la vía mTOR mediante el control de las actividades de S6K1 y 4E-BP1 (62, 63). La leucina, el principal BCAA que regula la síntesis de proteínas, acelera la síntesis de proteínas en el organismo activando la vía mTOR, promoviendo así el crecimiento (20, 64, 65). Por lo tanto, especulamos que el efecto promotor del crecimiento de la NAC puede estar relacionado con su alto contenido de AA y digestibilidad (15, 66). Sin embargo, vale la pena señalar que la NAC tiene un bajo contenido de arginina y un alto contenido de ácidos nucleicos. La arginina es un importante regulador de la vía mTOR, y la falta de arginina conduce a una disminución de la síntesis de proteínas, lo que puede explicar por qué una alta proporción de NAC afecta negativamente el crecimiento y el desarrollo del organismo.
Conclusión
En este estudio, la adición de un 3% de NAC a las dietas de pollos de engorde redujo el FCR (22-42 días) y aumentó el rendimiento muscular de la pechuga y las patas, así como el contenido de PB de la carne de pollo. La adición de un 3% de CAP a las dietas de los pollos de engorde también mejoró las propiedades antioxidantes (T-AOC, GSH-Px y SOD) y mejoró la IgG del organismo. Además, la inclusión de un 3% de CAP en la dieta activó la vía mTOR y aumentó los niveles de fosforilación de 4E-BP1, S6K1, Akt y AMPKɑ2. En particular, la adición de un 4% de NAC a la dieta aumentó la UA, la AST y la FA séricas. Basándonos en los resultados anteriores, nuestro nivel de uso recomendado en dietas para pollos de engorde es del 3%.
Declaración de disponibilidad de datos
Los conjuntos de datos presentados en este estudio se pueden encontrar en repositorios en línea. Los nombres de los repositorios y los números de acceso se pueden encontrar en el artículo/Material complementario.
Declaración ética
Los estudios en animales fueron aprobados por las Directrices de Bienestar de Animales de Laboratorio de China. Los estudios se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
CS: Redacción – borrador original, Visualización, Validación, Metodología, Investigación, Análisis formal, Curación de datos, Conceptualización. YL: Redacción – borrador original, Validación, Software, Metodología, Análisis formal, Conceptualización. CM: Redacción – borrador original, validación, recursos, metodología. CL: Redacción – borrador original, software, recursos, metodología. QL: Redacción – borrador original, Validación, Recursos, Metodología, Investigación. SL: Redacción – borrador original, Software, Metodología, Análisis formal, Curación de datos. GJ: Redacción – revisión y edición, Visualización, Validación, Supervisión, Administración de proyectos, Obtención de fondos, Conceptualización. JT: Redacción – revisión y edición, Visualización, Validación, Supervisión, Administración de proyectos, Obtención de fondos, Conceptualización.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este trabajo de investigación fue apoyado por el proyecto «Plan de Talentos Xingliao» de la provincia de Liaoning (Proyecto No. XLYC1803014).
Conflicto de intereses
YL, QL y GJ fueron empleados por Dalian Sanyi Animal Medicine Co., Ltd. CM y CL fueron empleados por Jiangsu Sanyi Animal Nutrition Technology Co., Ltd.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1389738/full#supplementary-material
Referencias
1. Gaffney, J, Challender, M, Califf, K, y Harden, K. Construyendo puentes entre la innovación de los agronegocios y los pequeños agricultores: una revisión. Glob Food Sec. (2019) 20:60–5. doi: 10.1016/j.gfs.2018.12.008
2. Bastawrous, A, y Suni, A-V. Magnitud proyectada a treinta años (hasta 2050) de discapacidad visual de cerca y de lejos y el impacto económico si las soluciones existentes se implementan a nivel mundial. Epidemiol oftálmico. (2020) 27:115–20. doi: 10.1080/09286586.2019.1700532
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Boland, MJ, Rae, AN, Vereijken, JM, Meuwissen, MP, Fischer, AR, van Boekel, MA, et al. El futuro suministro de proteínas de origen animal para el consumo humano. Tendencias Food Sci Technol. (2013) 29:62–73. doi: 10.1016/j.tifs.2012.07.002
4. Woyengo, T, Knudsen, KB y Børsting, C. Dietas bajas en proteínas para pollos de engorde: conocimiento actual y estrategias potenciales para mejorar el rendimiento y la salud, y para reducir el impacto ambiental. Anim Feed Sci Technol. (2023) 115574. doi: 10.1016/j.anifeedsci.2023.115574
5. Honeyman, M. Cuestiones de sostenibilidad de la producción porcina estadounidense. J Anim Sci. (1996) 74:1410–7. doi: 10.2527/1996.7461410x
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Van Harn, J, Dijkslag, M, y Van Krimpen, M. Efecto de las dietas bajas en proteínas suplementadas con aminoácidos libres sobre el rendimiento del crecimiento, el rendimiento del sacrificio, la calidad de la camada y las lesiones de las almohadillas de los pollos de engorde machos. Poult Sci. (2019) 98:4868–77. doi: 10.3382/ps/pez229
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Attia, YA, Bovera, F, Wang, J, Al-Harthi, MA, y Kim, WK. Suplementación con múltiples aminoácidos en dietas bajas en proteínas: efecto sobre el rendimiento, el rendimiento de la canal, la calidad de la carne y la excreción de nitrógeno de los pollos de engorde en condiciones climáticas cálidas. Animales. (2020) 10:973. doi: 10.3390/ani10060973
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Wang, Y, Zhou, J, Wang, G, Cai, S, Zeng, X y Qiao, S. Avances en dietas bajas en proteínas para cerdos. J Anim Sci Biotechnol. (2018) 9:1–14. doi: 10.1186/s40104-018-0276-7
9. Bratosin, BC, Darjan, S, y Vodnar, DC. Proteína unicelular: un potencial sustituto en la nutrición humana y animal. Sostener para. (2021) 13:9284. doi: 10.3390/su13169284
10. Wu, Z, Yu, X, Guo, J, Fu, Y, Guo, Y, Pan, M, et al. Sustitución de la harina de pescado de la dieta con proteína Clostridium autoethanogenum sobre el rendimiento del crecimiento, la digestión, las vías mTOR y la calidad muscular del abulón Haliotis discus hannai. Acuicultura. (2022) 553:738070. doi: 10.1016/j.aquaculture.2022.738070
11. Chen, Y, Sagada, G, Xu, B, Chao, W, Zou, F, Ng, WK, et al. Sustitución parcial de harina de pescado por proteína unicelular de Clostridium autoethanogenum en la dieta de besugo juvenil (Acanthopagrus schlegelii). Aquac Res. (2020) 51:1000–11. doi: 10.1111/are.14446
12. Saied, J, Al-Jabary, Q y Thalij, K. Efecto del cultivo de levadura de suplementos dietéticos sobre el rendimiento de la producción y los parámetros hematológicos en pollos de engorde. Int J Poult Sci. (2011) 10:376–80. doi: 10.3923/ijps.2011.376.380
13. El-Naga, M. Efecto de la suplementación con levadura en la dieta sobre el rendimiento de los pollos de engorde. Egipto Poult Sci J. (2012) 32:95–106.
14. Hombegowda, GP, Suresh, BN, Shivakumar, MC, Ravikumar, P, Girish, BC, Rudrappa, SM, et al. Rendimiento de crecimiento, características de la canal y salud intestinal de pollos de engorde alimentados con dietas incorporadas con proteína unicelular. Anim Biosci. (2021) 34:1951–62. doi: 10.5713/ab.20.0844
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Wei, H, Yu, H, Chen, X, Chao, W, Zou, F, Chen, P, et al. Efectos de la harina de soja reemplazada por la proteína Clostridium autoethanogenum sobre el rendimiento del crecimiento, los índices bioquímicos plasmáticos y la histopatología hepatopáncreas e intestinal de la carpa herbívora (Ctenopharyngodon idllus). Chin J Anim Nutr. (2018) 30:4190–01. Disponible en: https://kns.cnki.net/kcms/detail/11.5461.S.20180731.1602.028.html
16. Norman, RO, Millat, T, Winzer, K, Minton, NP y Hodgman, C. Progreso hacia la producción de productos químicos de plataforma utilizando Clostridium autoethanogenum. Biochem Soc Trans. (2018) 46:523–35. doi: 10.1042/BST20170259
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Yang, P, Li, X, Song, B, He, M, Wu, C y Leng, X. El potencial de Clostridium autoethanogenum, una nueva proteína unicelular, en la sustitución de la harina de pescado en la dieta de la lobina negra (Micropterus salmoides): crecimiento, utilización del alimento e histología intestinal. Aquac Fisher. (2023) 8:67–75. doi: 10.1016/j.aaf.2021.03.003
18. Ma, S, Liang, X, Chen, P, Wang, J, Gu, X, Qin, Y, et al. Una nueva proteína unicelular de Clostridium autoethanogenum como proteína funcional para la lubina negra (Micropterus salmoides). Anim Nutr. (2022) 10:99–10. doi: 10.1016/j.aninu.2022.04.005
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Zhou, Y-M, Liu, Y, Liu, W y Shen, Y. Generación de piensos proteicos microbianos (MPF) a partir de residuos y su aplicación en la acuicultura en China. J Environ Chem Ing. (2023) 11:109297. doi: 10.1016/j.jece.2023.109297
20. Murgas Torrazza, R, Suryawan, A, Gazzaneo, MC, Orellana, RA, Frank, JW, Nguyen, HV, et al. La suplementación con leucina de una comida baja en proteínas aumenta la síntesis de proteínas del músculo esquelético y del tejido visceral en cerdos neonatos al estimular el inicio de la traducción dependiente de mTOR. J Nutr. (2010) 140:2145–52. doi: 10.3945/jn.110.128421
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Yao, K, Yin, Y-L, Chu, W, Liu, Z, Deng, D, Li, T, et al. La suplementación dietética con arginina aumenta la actividad de señalización de mTOR en el músculo esquelético de cerdos recién nacidos. J Nutr. (2008) 138:867–72. doi: 10.1093/jn/138.5.867
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Zhang, X, Zhao, F, Si, Y, Huang, Y, Yu, C, Luo, C, et al. GSK3β regula la síntesis de leche y la proliferación de células epiteliales mamarias de vacas lecheras a través de la vía de señalización mTOR / S6K1. Moléculas. (2014) 19:9435–52. doi: 10.3390/moléculas19079435
23. Dancey, J. Señalización de mTOR y desarrollo de fármacos en el cáncer. Nat Rev Clin Oncol. (2010) 7:209–19. doi: 10.1038/nrclinonc.2010.21
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Gundermann, DM, Walker, DK, Reidy, PT, Borack, MS, Dickinson, JM, Volpi, E, et al. La activación de la señalización de mTORC1 y la síntesis de proteínas en el músculo humano después del ejercicio de restricción del flujo sanguíneo es inhibida por la rapamicina. Am J Physiol Endocrinol Metab. (2014) 306:E1198–204. doi: 10.1152/ajpendo.00600.2013
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Laplante, M, y Sabatini, DM. Señalización mTOR de un vistazo. J Cell Sci. (2009) 122:3589–94. doi: 10.1242/jcs.051011
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Li, M, Liang, H, Xie, J, Chao, W, Zou, F, Ge, X, et al. La dieta suplementada con una nueva proteína Clostridium autoethanogenum tiene un efecto positivo en el rendimiento del crecimiento, el estado antioxidante y la inmunidad en la carpa Jian juvenil (Cyprinus carpio var Jian). Rep. Aquac (2021) 19:100572. doi: 10.1016/j.aqrep.2020.100572
27. Maulu, S, Liang, H, Ge, X, Yu, H, Huang, D, Ke, J, et al. Efecto de la proteína Clostridium autoethanogenum en la dieta sobre el crecimiento, la composición corporal, los parámetros plasmáticos y la expresión de genes hepáticos relacionados con el crecimiento y la vía de señalización AMPK/TOR/PI3K de los juveniles de tilapia de cultivo (GIFT: Oreochromis niloticus) mejorados genéticamente. Anim Feed Sci Technol. (2021) 276:114914. doi: 10.1016/j.anifeedsci.2021.114914
28. Xue, R, Li, H, Liu, S, Hu, Z, Wu, Q y Ji, H. Sustitución de harina de soja con proteína de Clostridium autoethanogenum en dietas de carpa herbívora (Ctenopharygodon idella): efectos sobre el rendimiento del crecimiento, la utilización del alimento, el valor nutricional muscular y las características sensoriales. Anim Feed Sci Technol. (2023) 295:115547. doi: 10.1016/j.anifeedsci.2022.115547
29. AOAC. Métodos Oficiales de Análisis de AOAC International. Asociación Internacional de Químicos Analíticos Oficiales (AOAC). MD Gaithersburg, 17ª ed. Rockville: AOAC. (2003).
30. Li, L, Liu, X, Wang, Y, Huang, Y y Wang, C. Efectos de la alimentación alterna entre la harina de pescado y las nuevas dietas proteicas en la salud intestinal de la lubina negra juvenil (Micropterus salmoides). Rep. Aquac (2022) 23:101023. doi: 10.1016/j.aqrep.2022.101023
31. Maulu, S, Hualiang, L, Ke, J, Ren, M, Ge, X, Huang, D, et al. La proteína dietética de Clostridium autoethanogenum modula la absorción intestinal, el estado antioxidante y la respuesta inmunitaria en juveniles GIFT (Oreochromis niloticus). Aquac Res. (2021) 52:5787–99. doi: 10.1111/are.15454
32. Yao, W, Yang, P, Zhang, X, Xu, X, Zhang, C, Li, X, et al. Efectos de la sustitución de la harina de pescado en la dieta por proteína de Clostridium autoethanogenum sobre el crecimiento y la calidad de la carne del camarón blanco del Pacífico (Litopenaeus vannamei). Acuicultura. (2022) 549:737770. doi: 10.1016/j.aquaculture.2021.737770
33. Jiang, X, Yao, W, Yang, H, Tan, S, Leng, X y Li, X. Efectos dietéticos de la proteína de Clostridium autoethanogenum que sustituye la harina de pescado sobre el crecimiento, la histología intestinal y la inmunidad del camarón blanco del Pacífico (Litopenaeus vannamei) según el análisis del transcriptoma. Pescados y mariscos Immunol. (2021) 119:635–44. doi: 10.1016/j.fsi.2021.10.005
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Wu, Y, Wang, J, Jia, M, Huang, S, Cao, Y, Yao, T, et al. Inclusión de la proteína Clostridium autoethanogenum en la dieta para pollos de engorde: mejora del rendimiento del crecimiento, el metabolismo de los lípidos y la microbiota intestinal. Front Vet Sci. (2022) 9:1028792. doi: 10.3389/fvets.2022.1028792
35. Najib, H. Valor alimenticio de la proteína unicelular producida a partir de frutos de palmera datilera (Phoenix dactylifera) para pollos de engorde. Ann Res Rev Biol. (2014) 4:2406–13. doi: 10.9734/ARRB/2014/9524
36. Li, H-H, Jiang, X-R y Qiao, J-Y. Efecto de Bacillus subtilis en la dieta sobre el rendimiento del crecimiento y los índices bioquímicos e inmunológicos séricos en lechones destetados. J Appl Anim Res. (2021) 49:83–8. doi: 10.1080/09712119.2021.1877717
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Torres-Pitarch, A, Hermans, D, Manzanilla, EG, Bindelle, J, Everaert, N, Beckers, Y, et al. Efecto de las enzimas alimentarias sobre la digestibilidad y el crecimiento en cerdos destetados: una revisión sistemática y metaanálisis. Anim Feed Sci Technol. (2017) 233:145–59. doi: 10.1016/j.anifeedsci.2017.04.024
38. Aldecoa, C, Llau, JV, Nuvials, X y Artigas, A. Papel de la albúmina en la preservación de la integridad del glicocálix endotelial y la microcirculación: una revisión. Ann Intens Care. (2020) 10:1–12. doi: 10.1186/s13613-020-00697-1
39. Zou, F, Zhao, X, y Wang, F. Una revisión sobre los componentes de la fruta que afectan el nivel de ácido úrico y sus mecanismos subyacentes. J Bioquímica de los alimentos. (2021) 45:E13911. doi: 10.1111/jfbc.13911
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Li, Y, Li, W, Luo, L, Ren, Y, Xing, W, Xu, G, et al. Los niveles de lípidos en la dieta afectan el rendimiento del crecimiento, el metabolismo de los lípidos, el estado antioxidante e inmunológico del esturión de Amur Acipenser schrenckii. Rep. Aquac (2023) 33:101796. doi: 10.1016/j.aqrep.2023.101796
41. Pan, S, Yan, X, Dong, X, Li, T, Suo, X, Liu, H, et al. Influencia de la suplementación con inositol en la dieta sobre el crecimiento, la histología hepática, el metabolismo de los lípidos y la expresión de genes relacionados en el mero híbrido juvenil ( ♀ epinephelus fuscoguttatus× ♂ e. lanceolatu) alimentado con dietas altas en lípidos. Aquac Nutr. (2022) 1–13. doi: 10.1155/2022/6743690
42. Amirabagya, F, Hapsari, RAF, y Wulandari, E. El efecto del extracto de semilla de Jatropha curcas L sobre la actividad de AST/ALT y el grosor de la vena central en el hígado. Pharm J. (2021) 13:66–72. doi: 10.5530/pj.2021.13.10
43. Tang, Z, Chen, H, He, H y Ma, C. Ensayos para la actividad de la fosfatasa alcalina: progreso y perspectivas. Tendencias de TrAC Química Anal. (2019) 113:32–43. doi: 10.1016/j.trac.2019.01.019
44. Jie, X, Wu, M, Yang, H y Wei, W. Sonda fluorescente de infrarrojo cercano rojo para la detección de fosfatasa alcalina in vivo resuelta en el tiempo con la ayuda de un nanocontenedor fotosensible. Química anal. (2019) 91:13174–82. doi: 10.1021/acs.analchem.9b03497
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Zhang, P, Fu, C, Zhang, Q, Li, S y Ding, C. Estrategia fluorescente radiométrica para localizar la actividad de la fosfatasa alcalina en las mitocondrias basada en el proceso ESIPT. Química anal. (2019) 91:12377–83. doi: 10.1021/acs.analchem.9b02917
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Wang, K, Wang, W, Zhang, X-Y, Jiang, A-Q, Yang, Y-S y Zhu, H-L. Sondas fluorescentes para la detección de fosfatasa alcalina en sistemas biológicos: avances recientes y perspectivas de futuro. Tendencias de TrAC Química Anal. (2021) 136:116189. doi: 10.1016/j.trac.2021.116189
47. Benhar, M. Papel de las enzimas glutatión peroxidasa y tiorredoxina reductasa de mamíferos en la respuesta celular al estrés nitrosativo. Gratis Radic Biol Med. (2018) 127:160–4. doi: 10.1016/j.freeradbiomed.2018.01.028
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Ma, Q-W, Guo, H-Y, Zhu, K-C, Guo, L, Liu, B-S, Zhang, N, et al. La ingesta dietética de taurina afecta el crecimiento y la regulación de la síntesis de taurina en el pámpano dorado, Trachinotus ovatus (Linnaeus 1758). Acuicultura. (2021) 530:735918. doi: 10.1016/j.aquaculture.2020.735918
49. Mahoma, Y. La mejor subclase de IgG para el desarrollo de fármacos terapéuticos con anticuerpos monoclonales y su producción comercial: una revisión. Inmunoma Res. (2020) 16:1–12. doi: 10.35248/1745-7580.20.16.173
50. Chen, K, Magri, G, Grasset, EK y Cerutti, A. Repensar las respuestas de anticuerpos de la mucosa: IgM, IgG e IgD se unen a IgA. Nat Rev Immunol. (2020) 20:427–41. doi: 10.1038/s41577-019-0261-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Bouquet, M, Passmore, MR, Hoe, LES, Tung, J-P, Simonova, G, Boon, A-C, et al. Desarrollo y validación de ELISAs para la cuantificación de interleucina (IL)-1β, IL-6, IL-8 e IL-10 en plasma ovino. J Métodos Immunol. (2020) 486:112835. doi: 10.1016/j.jim.2020.112835
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Hao, G, Duan, W, Sun, J, Liu, J y Peng, B. Efectos del febuxostat sobre las citocinas séricas IL-1, IL-4, IL-6, IL-8, TNF-α y COX-2. Exp Ther Med. (2019) 17:812–6. doi: 10.3892/etm.2018.6972
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Tang, Z-L, Zhang, K, Lv, S-C, Xu, G-W, Zhang, J-F y Jia, H-Y. LncRNA MEG3 suprime la vía de señalización PI3K/AKT/mTOR para mejorar la autofagia e inhibir la inflamación en queratinocitos tratados con TNF-α y ratones psoriásicos. Citoquina. (2021) 148:155657. doi: 10.1016/j.cyto.2021.155657
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Sabatini, DM. Veinticinco años de mTOR: descubriendo el vínculo entre los nutrientes y el crecimiento. Proc Natl Acad Sci. (2017) 114:11818–25. doi: 10.1073/pnas.1716173114
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Goldspink, D. La influencia de la inmovilización y el estiramiento en el recambio proteico del músculo esquelético de rata. J Physiol. (1977) 264:267–82. doi: 10.1113/jphysiol.1977.sp011667
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Liu, GY y Sabatini, DM. mTOR en el nexo de la nutrición, el crecimiento, el envejecimiento y la enfermedad. Nat Rev Mol Cell Biol. (2020) 21:183–03. doi: 10.1038/s41580-019-0199-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. She, Y, Deng, H, Cai, H y Liu, G. Regulación de la expresión de moléculas de señalización clave en la vía mTOR de células satélite del músculo esquelético en pollitos neonatos: efectos de la leucina y el péptido glicina-leucina. J Anim Physiol Anim Nutr. (2019) 103:786–90. doi: 10.1111/jpn.13090
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Zhong, H, Yuan, P, Li, Y, Batonon-Alavo, D, Deschamps, C, Feng, B, et al. La metionina protege las células mamarias contra el estrés oxidativo mediante la producción de S-adenosilmetionina para mantener la actividad de señalización de mTORC1. Oxidativa Med Cell Longev. (2021) 2021:5550196. doi: 10.1155/2021/5550196
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Batool, A, Majeed, ST, Aashaq, S, Majeed, R, Shah, G, Nazir, N, et al. El secuestro del factor de iniciación eucariota 4E (eIF4E) media la respuesta de 4E-BP1 a la rapamicina. Int J Biol Macromol. (2019) 125:651–9. doi: 10.1016/j.ijbiomac.2018.12.102
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Huang, B, Shi, M, Pang, A, Tan, B y Xie, S. Efectos del reemplazo de harina de pescado por harina de proteína de Clostridium Autoethanogenum sobre el metabolismo de los ácidos biliares del colesterol, la capacidad antioxidante, la salud hepática e intestinal del mero de genciana perla (Epinephelus fuscoguttatus♀× Epinephelus lanceolatus♂). Animales. (2023) 13:1090. doi: 10.3390/ani13061090
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Fan, Z, Li, C, Wu, D, Li, J, Wang, L, Cao, D, et al. Evaluación de cuatro nuevas fuentes de proteínas como alternativas a la harina de soja para dos especificaciones de la carpa herbívora (Ctenopharyngodon idellus) cultivada en jaulas: efecto sobre el rendimiento del crecimiento, la calidad de la carne y la expresión de genes relacionados con el músculo. Frente Mar Sci. (2022) 9:935651. doi: 10.3389/fmars.2022.935651
62. Komorowski, JR, Ojalvo, SP, Sylla, S, Tastan, H, Orhan, C, Tuzcu, M, et al. La adición de un complejo amilopectina/cromo a los aminoácidos de cadena ramificada mejora la síntesis de proteínas musculares en el músculo esquelético de rata. J Int Soc Sports Nutr. (2020) 17:26. doi: 10.1186/s12970-020-00355-8
63. Rezaei, R y Wu, G. Los aminoácidos de cadena ramificada regulan el recambio de proteínas intracelulares en las células epiteliales mamarias porcinas. Aminoácidos. (2022) 54:1491–04. doi: 10.1007/s00726-022-03203-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
64. Columbus, DA, Fiorotto, ML y Davis, TA. La leucina es un importante regulador de la síntesis de proteínas musculares en los recién nacidos. Aminoácidos. (2015) 47:259–70. doi: 10.1007/s00726-014-1866-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
65. Zou, T, Cao, SP, Xu, WJ, Han, D, Liu, HK, Yang, YX, et al. Efectos de los niveles de leucina en la dieta sobre el crecimiento, el contenido de proteínas tisulares y la expresión relativa de genes relacionados con la síntesis de proteínas en juveniles de carpa gibelí (Carassius auratus gibelio var. CAS III). Aquac Res. (2018) 49:2240–8. doi: 10.1111/are.13682
66. Li, X, Chen, Y, Zheng, C, Chi, S, Zhang, S, Tan, B, et al. Evaluación de seis nuevas fuentes de proteínas sobre la digestibilidad aparente en el camarón blanco del Pacífico, Litopenaeus vannamei. Aquac Nutr. (2022) 1–11. doi: 10.1155/2022/8225273
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: Proteína de Clostridium autoethanogenum, pollos de engorde, rendimiento de crecimiento, mTOR, actividad antioxidante, función inmune
Cita: Shan C, Liu Y, Ma C, Li C, Liu Q, Liu S, Jiang G y Tian J (2024) La suplementación dietética con proteína Clostridium autoethanogenum mejora el rendimiento del crecimiento y promueve la síntesis de proteínas musculares mediante la activación de la vía de señalización mTOR del pollo de engorde. Frente. Vet. Sci. 11:1389738. doi: 10.3389/fvets.2024.1389738
Editado por:
Yangchun Cao, Universidad A&F del Noroeste, China
Revisado por:
Sabreen Ezzat Fadl, Universidad de Matrouh, Egipto
Weiwei Wang, Academia de Administración Nacional de Alimentos y Reservas Estratégicas, China
Derechos de autor © 2024 Shan, Liu, Ma, Li, Liu, Liu, Jiang y Tian. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Guotuo Jiang, jgt600@126.com; Jing Tian, Tianjing@dlpu.edu.cn
†Estos autores han contribuido igualmente a este trabajo y comparten la primera autoría
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.