
Las diferencias metabólicas del anestro, el celo, el embarazo, el pseudoembarazo y la lactancia en 800 perras

Las diferencias metabólicas del anestro, el celo, el embarazo, el pseudoembarazo y la lactancia en 800 perras
 Claudia Ottka1,2,3,4*
Claudia Ottka1,2,3,4* Katariina Vapalahti1,2,3
Katariina Vapalahti1,2,3 Sebastian P. Arlt5
Sebastian P. Arlt5 Alejandro Bartel6
Alejandro Bartel6 Hannes Lohi1,2,3,4
Hannes Lohi1,2,3,4- 1PetBiomics Ltd., Helsinki, Finlandia
- número arábigoDepartamento de Biociencias Veterinarias, Universidad de Helsinki, Helsinki, Finlandia
- 3Departamento de Genética Médica y Clínica, Universidad de Helsinki, Helsinki, Finlandia
- 4Centro de Investigación Folkhälsan, Helsinki, Finlandia
- 5Clínica de Medicina Reproductiva, Facultad Vetsuisse, Universidad de Zúrich, Zúrich, Suiza
- 6Instituto de Epidemiología y Bioestadística Veterinaria, Facultad de Medicina Veterinaria, Freie Universität Berlin, Berlín (Alemania)
Introducción: La reproducción provoca importantes cambios hormonales y fisiológicos en el cuerpo femenino. Sin embargo, los cambios metabólicos que ocurren durante la reproducción canina son escasamente estudiados.
Métodos: En este estudio transversal, evaluamos los efectos metabólicos del estado reproductivo canino utilizando una plataforma metabolómica de RMN 1H optimizada y validada para uso canino. La población de estudio consistió en un total de 837 perras sanas e intactas en edad reproductiva, de las cuales 663 perras estaban en anestro, 78 en celo, 43 estaban pseudoembarazadas, 15 estaban embarazadas y 38 estaban lactando. Las diferencias en los perfiles de metabolitos entre estos estados se estudiaron mediante la prueba de Kruskal-Wallis con pruebas post-hoc realizadas mediante la prueba de Dunn, y se visualizaron mediante diagramas de caja y un mapa de calor. La capacidad del perfil de metabolitos para diferenciar perras gestantes de las no gestantes se evaluó mediante la creación de un modelo de regresión logística de Firth multivariado utilizando selección escalonada hacia adelante.
Resultados: La lactancia, el embarazo y el celo se asociaron con cambios metabólicos distintos; El embarazo causó cambios importantes en las concentraciones de glicoproteína acetils, albúmina y creatinina, y cambios más pequeños en varios lípidos, citrato, glutamina y alanina. El pseudoembarazo, por otro lado, metabólicamente se parecía en gran medida al anestro. La lactancia causó cambios importantes en las concentraciones de aminoácidos y cambios más pequeños en varios lípidos, albúmina, citrato, creatinina y acetilos glicoproteicos. El calor, refiriéndose al proestro y al celo, afectó el metabolismo del colesterol y el LDL, y aumentó el tamaño de las partículas de HDL. La albúmina y la glicoproteína acetilos fueron los metabolitos incluidos en el modelo multivariado final para la detección de embarazo, y pudieron diferenciar perras gestantes de las no gestantes con excelente sensibilidad y especificidad.
Discusión: Estos resultados aumentan nuestra comprensión de las consecuencias metabólicas de la reproducción canina, con la posibilidad de mejorar la salud materna y garantizar el éxito reproductivo. Los metabolitos identificados podrían utilizarse para confirmar el embarazo canino.
1. Introducción
El tiempo alrededor del parto es un desafío para el cuerpo femenino debido a los cambios hormonales, el parto, la lactancia de los cachorros, la producción de leche y la involución uterina (1). En este contexto, la capacidad de transformación del cuerpo de la perra es notable. Pasa de acomodar a un individuo a una cámara de incubación para uno o más individuos adicionales, y a un proveedor eficaz de nutrientes para los individuos recién nacidos a través de la lactancia. Este proceso provoca grandes alteraciones en el cuerpo femenino, y está gobernado por complejas interacciones hormonales y neurológicas. También es un proceso delicado, y múltiples factores internos y externos pueden causar daños graves a la hembra embarazada, a sus crías o a ambos (2).
Dado que la salud de la madre está estrechamente relacionada con el éxito reproductivo, comprender los cambios metabólicos que ocurren durante la reproducción es esencial tanto para la salud de la madre como para la salud y el desarrollo normal de la descendencia. Por ejemplo, las necesidades nutricionales de la madre, tanto de micro como de macronutrientes, cambian durante la reproducción, y no satisfacerlas puede causar graves daños a la madre y a su descendencia (3, 4). Además, diferenciar los cambios fisiológicos de los fisiopatológicos es esencial para controlar la salud de la madre. Por ejemplo, las concentraciones más bajas de creatinina durante el embarazo podrían enmascarar la disfunción renal (5), y la elevación fisiológica de varios parámetros inflamatorios durante el embarazo podría llevar a una interpretación errónea entre el embarazo y la enfermedad inflamatoria (6-12).
El perfil metabólico se ha utilizado con éxito en humanos para caracterizar cambios específicos que ocurren durante el embarazo (12-17). Sin embargo, los efectos metabólicos del ciclo reproductivo, la gestación y la lactancia han sido escasamente estudiados en perros, y los métodos metabolómicos no se han utilizado en estudios en reproducción canina. Las características reproductivas de los perros difieren de las de los humanos en varios aspectos: los perros son animales monocíclicos que entran en celo solo cada 6-7 (5-12) meses (18, 19). Además, los cuerpos lúteos persisten alrededor de 2 meses después de la ovulación espontánea en celo, independientemente de si la perra queda embarazada o no (19). El entorno hormonal de las perras no embarazadas imita aproximadamente el del embarazo y la lactancia canina (20, 21), y puede causar síntomas similares al embarazo y la lactancia en un estado llamado pseudoembarazo (19, 22, 23). La implantación de los embriones se produce relativamente tarde, alrededor de 18 días después del pico de LH, es decir, durante el primer trimestre tardío (6). Debido a estas diferencias, las similitudes metabólicas en la reproducción canina y humana deben estudiarse para evaluar si los hallazgos de estas especies se pueden extrapolar entre sí. Si bien el embarazo y el pseudoembarazo caninos comparten entornos hormonales y síntomas similares (20-23), se sabe poco sobre las diferencias metabólicas entre estos dos estados. Debido a los síntomas similares de estas afecciones, es necesario utilizar pruebas diagnósticas para confirmar el embarazo (2). Todavía no se ha evaluado la utilidad de las medidas metabólicas en las pruebas de embarazo de perros.
Comprender los efectos metabólicos del ciclo reproductivo femenino y las características del metabolismo en etapas específicas es crucial para monitorear la salud durante estas fases sensibles. Diferenciar los cambios patológicos de las adaptaciones fisiológicas ayudará a detectar trastornos a tiempo, y comprender las necesidades metabólicas del perro puede ayudar a proporcionar una nutrición óptima y, por lo tanto, garantizar el éxito reproductivo. Este estudio tiene como objetivo caracterizar las diferencias en el metabolismo de perras sanas no esterilizadas en anestro, celo, pseudoembarazo, embarazo y lactancia. Además, nuestro objetivo es evaluar si los datos de metabolitos se pueden utilizar para diferenciar a las perras embarazadas de las no gestantes.
2. Materiales y métodos
Las muestras y los datos del estudio se derivaron de un conjunto de muestras previamente recolectado de 6,306 muestras de suero/plasma de perros domésticos. Los detalles de este conjunto de muestras, su recolección y análisis de RMN se describen en otra parte (24). En resumen, las muestras se recolectaron por venopunción cefálica de perros de propiedad privada en toda Finlandia en 2017-2018. Las muestras se enfriaron inmediatamente después de la separación de las células sanguíneas y se almacenaron a -80 °C hasta el análisis de espectroscopia de resonancia magnética nuclear de protones (RMN 1H). Los dueños de los perros completaron cuestionarios que cubrían la señalización del perro, su salud actual, la dieta, el ejercicio, el estrés y la fase del ciclo reproductivo. La participación fue voluntaria y los dueños de los perros dieron su consentimiento informado firmado. La toma de muestras de sangre siguió la legislación nacional y de la Unión Europea y fue aprobada por el Comité de Ética Animal de la Oficina Provincial Estatal del Sur de Finlandia, número de permiso: ESAVI/7482/04.10.07/2015, fecha de aprobación: 9.10.2015.
2.1. Metabolómicade RMN 1 H
Las muestras se analizaron utilizando una plataforma metabolómica de RMN 1H validada y específica para perros que cuantificó 123 mensurandos en unidades absolutas (24). Las mediciones cuantificadas se presentan en la Figura 1. Los detalles sobre el método y su validación se proporcionan en otra parte (24-27).

Brevemente, el método utiliza un espectrómetro de RMN Bruker AVANCE III HD 500 equipado con un cabezal de sonda Prodigy de gradiente z de triple canal (1H, 13C, 15N) de 5 mm y un cambiador de muestras refrigerado de alto rendimiento SampleJet (Bruker Corp., Billerica, Massachusetts, EE. UU.). La preparación de la muestra comienza con una mezcla ligera de la muestra y la centrifugación para eliminar el posible precipitado (25). A continuación, el proceso altamente automatizado continúa mediante la transferencia de cada muestra a tubos de RMN individuales y la mezcla con tampón de fosfato de sodio utilizando una estación de trabajo automatizada PerkinElmer JANUS equipada con un brazo dispensador de 8 puntas con Varispan (PerkinElmer Inc., Waltham, Massachusetts, EE. UU.) (26). Los espectros de RMN se adquieren automáticamente a partir de cada muestra utilizando parámetros estandarizados y se procesan automáticamente utilizando scripts internos optimizados para perros (24-27). La generación de resultados se basa en modelos de regresión e incluye control de calidad integrado (27).
2.2. Inclusión de muestras
En la Figura 2 se presenta una visión general de las muestras utilizadas, los criterios de inclusión, el preprocesamiento de los datos y los análisis estadísticos.
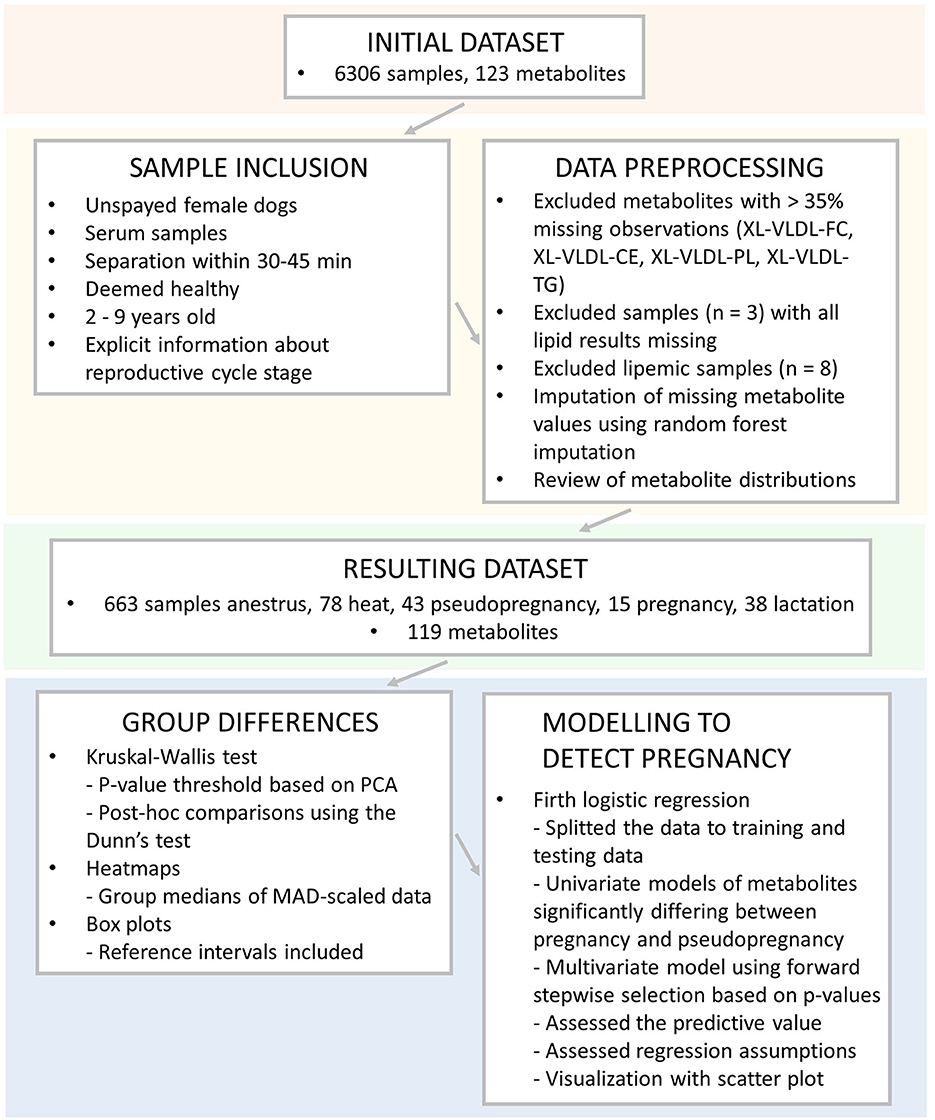
Se realizó una revisión de la inclusión de muestras para análisis estadísticos en Microsoft Office Excel (Microsoft Corp., Redmond, WA, EE. UU.). Solo se utilizaron muestras de perras no esterilizadas del conjunto de datos recopilado previamente (24). Solo se incluyeron muestras de suero para disminuir los efectos de los diferentes tipos de muestras en los resultados. Las muestras para las que no se había realizado la separación del suero de las células sanguíneas de acuerdo con las recomendaciones del fabricante del tubo de muestra (separación del suero dentro de los 30-45 minutos posteriores a la toma de muestras de sangre) se eliminaron del conjunto de datos.
El estado de salud de los perros reportado por el dueño se revisó cuidadosamente, y solo se incluyeron en el estudio los perros considerados sanos. Por ejemplo, los perros con enfermedades oculares menores, como distiquiasis o displasia articular leve o moderada observadas en radiografías rutinarias relacionadas con la reproducción sin síntomas ni tratamiento, se consideraron sanos. Para evaluar solo los datos de perros en edad reproductiva típica, incluimos muestras de animales de más de 2 a menos de 9 años. La información reportada por el propietario sobre el estado de la etapa del ciclo reproductivo (en celo, embarazada, lactante o pseudoembarazada) en el momento de la recolección de la muestra fue revisada cuidadosamente. Solo se incluyeron muestras de perros con información específica sobre las etapas del ciclo. Se excluyeron muestras de perros con etapas de ciclo poco claras o estado reproductivo desconocido, como perras apareadas sin diagnóstico de embarazo. Las perras que no informaron estar en celo, preñadas, lactantes o pseudoembarazadas se incluyeron en el grupo de anestro.
Finalmente, se verificaron los datos para detectar diferencias en la edad y la duración del ayuno en el R Project for Statistical Computing (RRID:SCR_001905) (28) utilizando la prueba de Kruskal-Wallis utilizando el paquete «stats», versión 3.6.2 (28). Las medianas y las desviaciones medias absolutas (MAD) se calcularon utilizando el paquete «stats», versión 3.6.2 (28), con la ayuda del paquete «dplyr», versión 1.0.7 (29). No se observaron diferencias significativas en la edad o la duración del ayuno en los grupos de anestro, celo, pseudoembarazo, embarazo y lactancia. El conjunto de datos final incluyó un total de 837 muestras. De estas muestras, 663 eran de perras en anestro, 78 de perras en celo, 43 de pseudogestantes, 15 de gestantes y 38 de perras lactantes.
2.3. Pretratamiento de datos y métodos estadísticos
El preprocesamiento de los datos y el análisis estadístico se realizaron en R 4.1.2 (28). El preprocesamiento de los datos se inició mediante la revisión de las observaciones faltantes. Se eliminaron los mensurandos de metabolitos con más del 35% de observaciones faltantes. Estas mediciones incluyeron los siguientes medidores de lipoproteínas de muy baja densidad (XL-VLDL): colesterol libre XL-VLDL, colesterol esterificado XL-VLDL, fosfolípidos XL-VLDL y triglicéridos XL-VLDL. Tres perros que no obtuvieron todos los resultados de lípidos y fueron eliminados del estudio. Se revisaron los datos en busca de valores atípicos y se extrajeron ocho muestras lipémicas con colesterol total superior a 15 mmol/l o triglicéridos totales superiores a 1,5 mmol/l.
La mayoría de los restantes mesurandos de metabolitos tenían menos del 1% (rango: 0-27%) de observaciones faltantes. Los valores de metabolitos faltantes restantes se imputaron utilizando la imputación de bosque aleatorio (RF) utilizando el paquete «missForest» versión 1.4 (30). Se utilizaron los valores predeterminados de la función missForest con el número máximo de iteraciones establecido en diez y el número de árboles establecido en 100. Se utilizó el valor del error cuadrático medio normalizado (NRMSE) para comprobar el rendimiento de la imputación de RF. El verdadero error de imputación fue 3.39e-13, lo que indica un rendimiento excelente. Las distribuciones de todos los metabolitos medidos se verificaron visualmente de forma individual para cada medidor. La mayoría de los mensurandos no se distribuyeron normalmente.
El umbral de significación para la prueba de hipótesis se obtuvo a partir de la corrección de Bonferroni basada en el número de componentes principales que explican más del 95% de la variación en los datos de metabolitos (27). Se utilizó este método ya que muchos de los resultados de los metabolitos están intercorrelacionados, y la corrección de Bonferroni debe realizarse sobre la base de comparaciones individuales. El análisis de los componentes principales se llevó a cabo en el conjunto de datos utilizando el paquete «stats», versión 3.6.2 (28). Diecinueve componentes principales explicaron el 95% de la variación de los datos, resultando en un umbral de significancia de p < 0,0026 (0,05/19).
Las diferencias en las mediciones de metabolitos entre los cinco grupos de estado reproductivo se analizaron mediante la prueba de Kruskal-Wallis utilizando el paquete «stats», versión 3.6.2 (28). En estos análisis se utilizó el umbral de significación de p < 0,0026. Las comparaciones post-hoc se llevaron a cabo utilizando la prueba de Dunn utilizando el paquete «FSA» (versión 0.8.20) (31). Los valores p se ajustaron mediante la corrección de Bonferroni basada en el número de comparaciones individuales. El umbral del valor p se estableció en p < 0,05.
La visualización de datos incluyó diagramas de caja y un mapa de calor. Se crearon diagramas de caja para visualizar las distribuciones en cada grupo, y se realizaron utilizando los paquetes «ggplot2» (32) y «ggpubr» (versión 0.4.0) (33). Para comprender la magnitud de las diferencias, se incorporaron a los diagramas de caja los intervalos de referencia séricos de todos los perros del método de RMN (24). Se creó un mapa de calor para visualizar las diferencias generales en los valores de metabolitos en los grupos estudiados. En primer lugar, los datos de metabolitos se utilizaron a escala MAD. Se calcularon las medianas de los datos a escala MAD para todos los grupos, y se incorporaron en el mapa de calor las medidas de metabolitos significativas en la prueba de Kruskal-Wallis. En la creación del mapa de calor se utilizaron los paquetes «ComplexHeatmap» (34), «grid» (28), «dplyr», versión 1.0.7 (29), «BiocManager», versión 1.30.16 (35) y «RColorBrewer» versión 1.1-2 (36). El mapa de calor y los diagramas de caja se finalizaron en Inkscape (37).
Para estudiar si los metabolitos podían diferenciar a las perras embarazadas de las no gestantes, creamos modelos de regresión logística de Firth (38) utilizando el paquete «logistf», versión 1.24.1 (39). Se utilizó la regresión logística de Firth debido a los grupos pequeños y desequilibrados. El estado de embarazo fue la variable dependiente y el metabolito la variable independiente. Para crear estos modelos, los datos de perras embarazadas y pseudoembarazadas se dividieron en datos de entrenamiento y prueba utilizando los paquetes «caret», versión 6.0-90 (40) y «tidyverse» (41). Los datos de entrenamiento cubrieron el 80% y los datos de prueba el 20% de las observaciones. La construcción del modelo se realizó utilizando los datos de entrenamiento.
En primer lugar, creamos modelos univariados de regresión logística de Firth para cada mensurando que muestran diferencias significativas en los valores de metabolitos entre embarazo y pseudoembarazo en la prueba de Kruskal-Wallis. Los modelos univariados se muestran en los datos complementarios S5.
A continuación, creamos un modelo de regresión logística de Firth multivariante con selección escalonada hacia delante añadiendo secuencialmente los mensurandos que recibieron el valor p más bajo cuando se añadieron al modelo, hasta que ninguna medida de metabolitos adicional alcanzó un umbral de valor p de <0,05. Solo los metabolitos que mostraron diferencias significativas entre el embarazo y el pseudoembarazo en la prueba de Kruskal-Wallis se utilizaron en la construcción del modelo. Antes de modelar, probamos si los metabolitos tenían interacciones significativas para ser incluidos en la construcción del modelo. El modelo multivariante y el proceso de selección escalonada hacia adelante se muestran en los datos complementarios S6.
Evaluamos cuidadosamente si el modelo cumplía con los supuestos de regresión logística y si el modelo se ajustaba adecuadamente. La multicolinealidad del modelo se verificó mediante el factor de inflación de la varianza (VIF) con un punto de corte de 10, utilizando el paquete «regclass», versión 1.6 (42). La linealidad del logit se comprobó añadiendo al modelo el término de interacción de la variable independiente y su logaritmo natural. Si el término de interacción contribuyó significativamente al modelo (p < 0.05), se consideró que la linealidad del logit no estaba presente. La presencia de observaciones influyentes se evaluó mediante el cálculo de los residuos con la ayuda del paquete «cmvnorm», versión 1.0-7 (43).
Los datos de las pruebas se utilizaron para evaluar qué tan bien el modelo puede diferenciar a las perras embarazadas de las pseudoembarazadas en muestras no incluidas en la creación del modelo. Esto se llevó a cabo utilizando el paquete «pROC» (44). Se generó una matriz de confusión para observar qué cantidad de perras preñadas y pseudopreñadas fueron clasificadas correctamente por el modelo, y se evaluó la sensibilidad y especificidad del modelo. Dado que el conjunto de datos de prueba era pequeño, queríamos evaluar más a fondo qué tan bien el modelo diferencia a las perras embarazadas de las no gestantes en un mayor número de perras. Para ello, creamos el conjunto de datos de EA con las muestras de perros en anestro (n = 663) y las 15 perras embarazadas en los datos, incluyendo tanto a las perras gestantes en los datos de entrenamiento como de pruebas. De manera similar a los datos de las pruebas, se generó una matriz de confusión y se evaluó la sensibilidad y especificidad del modelo en los datos de EA. Se generó y evaluó un gráfico que combina las curvas de características operativas del receptor (ROC) y su área bajo la curva (AUC) de los datos de prueba, entrenamiento y EA. La trama se finalizó en Inkscape (37).
Para visualizar cómo funcionaba el modelo multivariante final y cómo se agrupan las muestras de embarazadas, pseudoembarazadas y anestros utilizando los metabolitos incluidos en el modelo multivariante final, creamos un diagrama de dispersión utilizando el paquete «tidyverse» (41). El diagrama de dispersión se finalizó en Inkscape (37).
3. Resultados
3.1. Características del grupo
Las características de los grupos de estudio se presentan en la Tabla 1. No se observaron diferencias significativas en las edades ni en la duración del ayuno entre los grupos de la etapa reproductiva. Todos los grupos estaban formados por una gran variedad de razas diferentes.
3.2. El celo, el embarazo y la lactancia se asocian con características metabólicas distintas
Las concentraciones de múltiples lípidos, como las concentraciones molares de los ácidos grasos ácido palmítico (PalA), ácido oleico (OleA), ácido linoleico (LA) y ácido docosapentaenoico (DPA), así como el colesterol libre, el colesterol de lipoproteínas de baja densidad (LDL), las partículas y lípidos de LDL, así como el tamaño de partícula de lipoproteínas de alta densidad (HDL) aumentaron durante el calor en comparación con el anestro (Figura 3; Datos complementarios S2, S3). En un análisis más detallado de las subclases de lipoproteínas, se aumentaron las partículas de XL-HDL, todos los subtipos de partículas de LDL y sus componentes, excluyendo los triglicéridos. Además, se incrementó la concentración de partículas de lipoproteínas de muy baja densidad (S-VLDL) y las concentraciones de los aminoácidos tirosina y alanina.
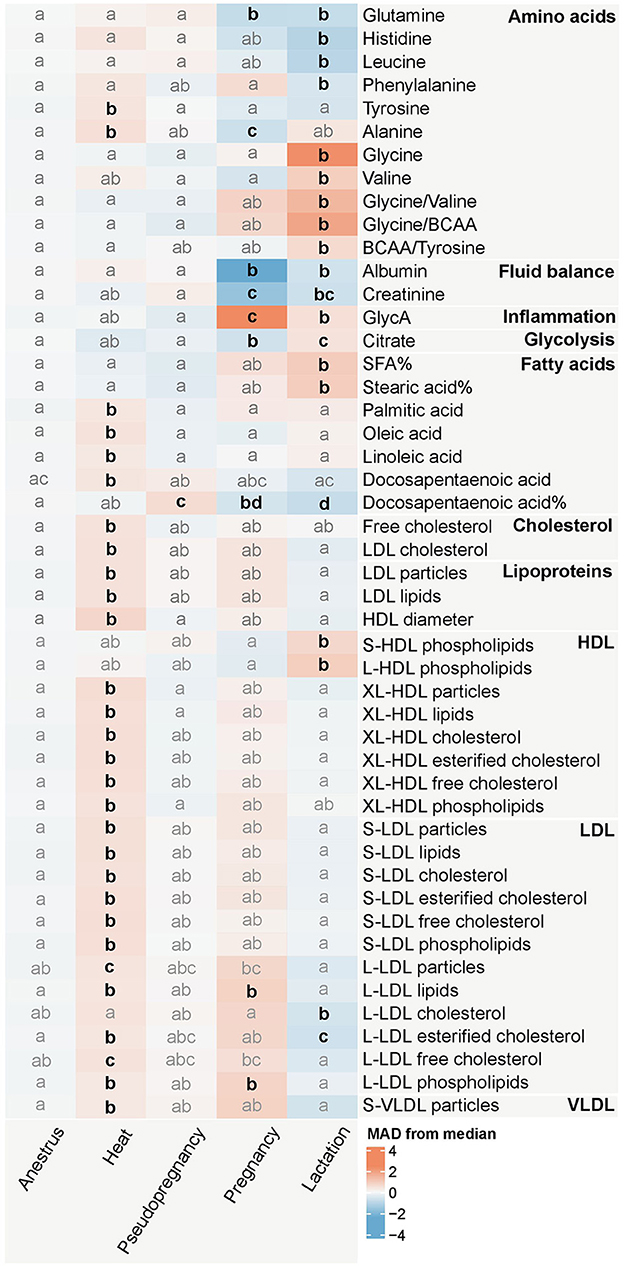
Se observaron diferencias mínimas en comparación con el anestro durante el pseudoembarazo, con solo aumento de la concentración relativa de DPA (Figura 3; Datos complementarios S2, S3). Las concentraciones de los lípidos y aminoácidos modificados por el calor se acercaron a las concentraciones observadas en el anestro.
Por el contrario, el embarazo causó cambios con algunas de las magnitudes más grandes, con las concentraciones de albúmina y GlycA difiriendo de la mediana del conjunto de datos con más de 2 MAD:s (Figura 3; Datos complementarios S2, S3). Además, las concentraciones de los aminoácidos glutamina y alanina, creatinina, citrato y la concentración relativa de DPA, disminuyeron durante el embarazo en comparación con el anestro. Las concentraciones de lípidos y fosfolípidos de las partículas grandes de LDL (L-LDL) permanecieron elevadas después del celo durante el embarazo.
La lactancia se asoció con cambios en múltiples medidores en múltiples grupos de metabolitos (Figura 3; Datos complementarios S2, S3). Las diferencias con respecto al anestro incluyeron la disminución de las concentraciones de los aminoácidos glutamina, histidina, leucina y fenilalanina, albúmina, creatinina, la concentración relativa de DPA y la concentración de colesterol esterificado en las partículas de L-LDL. Por el contrario, las concentraciones de los aminoácidos glicina y valina, así como las proporciones de aminoácidos glicina/valina y glicina/aminoácidos de cadena ramificada (BCAA), GlycA, citrato, la concentración relativa de ácidos grasos saturados y ácido esteárico (SteA), así como los fosfolípidos en las partículas S-LDL y L-LDL, aumentaron en comparación con el anestro.
3.3. Las concentraciones de metabolitos rara vez se sitúan fuera de los intervalos de referencia
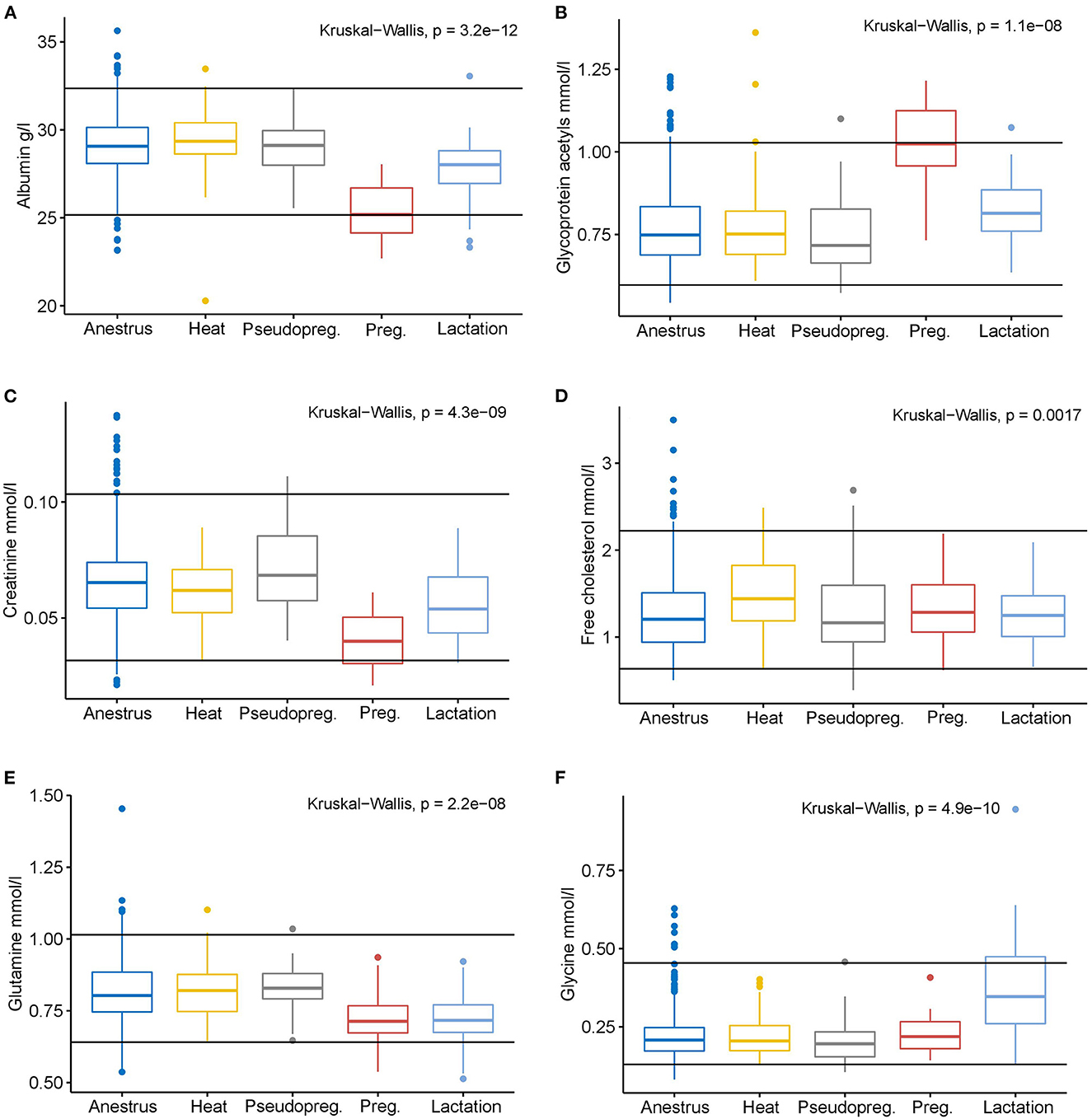
Si bien las diferentes fases del ciclo reproductivo mostraron claras diferencias metabólicas, las concentraciones de metabolitos en cada grupo rara vez excedieron o socavaron los intervalos de referencia sérica de todos los perros del método de RMN (Figura 4; Datos complementarios S3). Solo la mediana de perras gestantes para albúmina y GlycA alcanzó los límites de referencia inferior y superior del método, respectivamente. Otros cambios fueron más sutiles en magnitud, especialmente en comparación con los intervalos de referencia del método. Sin embargo, hay que tener en cuenta que 152 (18,2%) de los perros incluidos en este estudio se utilizaron en la generación de los intervalos de referencia de todos los perros (24).
3.4. El glycA y la albúmina pueden diferenciar a las perras gestantes de las no gestantes
En los modelos univariados de regresión logística de Firth, los mensurandos que más contribuyeron a predecir la inclusión a los grupos pseudoembarazadas y embarazadas fueron la albúmina (p < 0,001), la creatinina (p < 0,001), la GlycA (p < 0,001), la glutamina (p < 0,001) y la alanina (p = 0,008). La concentración relativa de DPA (p = 0,734) no fue un predictor adecuado para la inclusión del grupo en estos modelos univariados (Datos Suplementarios S5).
El modelo multivariado final creado por selección escalonada hacia adelante incluyó albúmina y GlycA como predictores (Datos Suplementarios S7). Ninguno de los términos de interacción alcanzó significación estadística (p < 0,05), y la edad y la duración del ayuno no tuvieron efecto significativo en el modelo, por lo que estos términos no se incluyeron en el modelo final. El modelo cumplió con el supuesto de linealidad del logit y no se observó multicolinealidad. El AUC del modelo sobre los datos de entrenamiento fue del 98,6% (Figura 5), lo que se considera excelente. El modelo funcionó exquisitamente en los datos de las pruebas con todas las perras pseudogestantes (n = 8) y gestantes (n = 3) clasificadas correctamente (Datos Suplementarios S8), haciendo que la sensibilidad, especificidad y AUC de los datos de las pruebas fuera del 100% (Figura 5). El modelo también funcionó bien en la clasificación de perras gestantes (n = 15) y perras en anestro (n = 663), con un AUC del 99,0% (Figura 5), una sensibilidad del 93,3% y una especificidad del 97,3% (Datos Suplementarios S9).
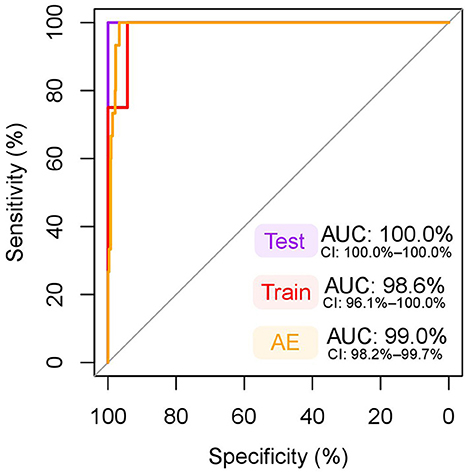
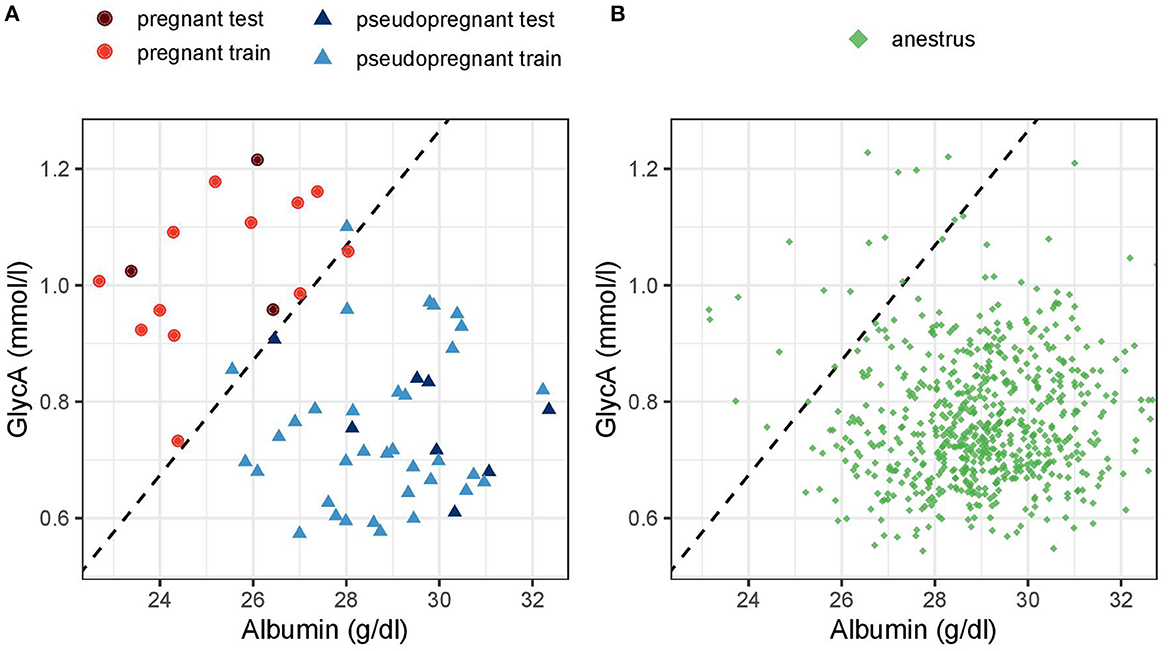
La Figura 6 destaca el agrupamiento distintivo de las muestras de perras embarazadas frente a perras pseudopreñadas y perras en anestro en función de las concentraciones séricas de albúmina y GlycA y visualiza cómo funciona el modelo multivariante.
4. Discusión
Este estudio identificó varias diferencias metabólicas en perras sanas y no esterilizadas durante el anestro, el celo, el pseudoembarazo, el embarazo y la lactancia, muchas de las cuales imitan las observadas en los humanos. Descubrir estos cambios aumenta nuestra comprensión de las consecuencias metabólicas de la reproducción femenina y es crucial para interpretar con precisión los valores de laboratorio en las perras. Además, desarrollamos un modelo de regresión logística de Firth multivariante basado en estos metabolitos que funcionó de manera excelente en la detección de perras embarazadas. Este hallazgo sugiere que las concentraciones de metabolitos podrían usarse como un método indirecto adicional de prueba de embarazo en perros sanos con una alta sensibilidad y especificidad.
El calor, referido al proestro y al estro, se caracterizó por varios cambios en las concentraciones de lípidos, como aumentos en las medidas de LDL, medidas de partículas XL-HDL, partículas S-VLDL, colesterol libre, el ácido graso saturado PalA, el ácido graso monoinsaturado OleA y los ácidos grasos poliinsaturados LA y DPA. Los estudios previos sobre los cambios en las concentraciones de lípidos durante el ciclo estral han tenido resultados contradictorios, con algunos estudios que no informan cambios y otros que observan varios cambios en los lípidos durante la fase folicular (16, 17, 45-49). Estos resultados pueden explicarse en parte por diferencias metodológicas, como la medición de diferentes parámetros lipídicos. Además, la magnitud de los cambios lipídicos que observamos fue relativamente pequeña, y todos los cambios se mantuvieron dentro de los intervalos de referencia, por lo que la observación de estos cambios requiere grandes tamaños de muestra. En este estudio, las concentraciones de los aminoácidos tirosina y alanina también aumentaron durante el celo. Cambios similares en los aminoácidos se han reportado previamente en humanos durante la fase folicular, la fase folicular se caracteriza por niveles más altos de aminoácidos que la fase lútea (16, 17, 50). Se ha sugerido que estos cambios ocurren debido a cambios en la actividad anabólica y catabólica durante el ciclo menstrual (16, 17). Además, se ha recomendado la alimentación con tirosina durante el celo, y aunque no se han documentado beneficios de su uso (51), el posible aumento de la ingesta de este aminoácido podría afectar su concentración plasmática.
El embarazo influyó en gran medida en las concentraciones de los marcadores inflamatorios GlycA y albúmina. Se sabe que el embarazo causa una carga importante en el sistema inmunológico femenino, que tiene que proporcionar una respuesta inmune adecuada para proteger tanto a la hembra como a su descendencia, y tolerar las reacciones inflamatorias causadas por la implantación, la invasión endometrial, el desarrollo de la placenta y el crecimiento del feto (2, 6, 11). Se ha demostrado que diferentes marcadores inflamatorios alcanzan su punto máximo en varias etapas del embarazo canino, pero todos los cambios parecen ocurrir después de la implantación (6). Dado que los marcadores inflamatorios se utilizan de forma rutinaria para detectar la inflamación, los efectos del embarazo siempre deben tenerse en cuenta en la interpretación de los resultados, y se deben determinar intervalos de referencia específicos del embarazo para ayudar en este desafío de interpretación (10).
La disminución de la concentración de creatinina durante el embarazo debido al aumento de la tasa de filtración glomerular es un fenómeno bien conocido y tiene importantes implicaciones prácticas (2, 5). Dado que las tasas de filtración glomerular basadas en las mediciones de creatinina en sangre se utilizan comúnmente para diagnosticar la enfermedad renal (52, 53), el cambio fisiológico en su concentración durante el embarazo puede enmascarar la lesión renal y conducir a enfermedades renales posiblemente graves debido a un diagnóstico insuficiente durante el embarazo (5). Por lo tanto, el estado reproductivo debe tenerse en cuenta al evaluar las concentraciones de creatinina en sangre. Los niveles ligeramente más bajos de citrato sérico observados en perras embarazadas probablemente sean causados por la mayor tasa de filtración glomerular durante el embarazo, lo que provoca un aumento de la excreción urinaria de citrato (54-56). Sin embargo, estudios previos no han identificado una disminución de las concentraciones plasmáticas de citrato durante el embarazo (12).
Las concentraciones de los aminoácidos glutamina y alanina también disminuyeron durante el embarazo. Los cambios en las concentraciones de aminoácidos durante el embarazo se han observado previamente en varias especies, con una tendencia general a la disminución de los niveles de aminoácidos durante el embarazo (12, 57-63). Las recomendaciones para la ingesta de proteínas son más altas durante el embarazo canino (4), y se cree que la disminución de los niveles de aminoácidos surge del aumento de la insulina postprandial (61) y la alta absorción de aminoácidos placentarios (63).
Se sabe que ciertos ácidos grasos poliinsaturados (AGPI), como el ácido docosahexaenoico (DHA), el ácido araquidónico (AA) y el ácido eicosapentaenoico (EPA), son necesarios para el desarrollo óptimo del neonato, especialmente en lo que respecta al sistema neuronal (3, 4, 64-66). En este estudio, las concentraciones molares de todos los AGPI estudiados, incluidos DHA, DPA, AA y LA en suero de perras embarazadas y lactantes, fueron similares al anestro, y solo la concentración relativa de DPA fue menor durante el embarazo y la lactancia que durante el anestro. Este hallazgo sugiere que las concentraciones de PUFA se conservan relativamente bien en el plasma canino durante el embarazo y la lactancia. Se necesitan más estudios para evaluar si las perras con bajo DPA sérico o sus crías se beneficiarían de la suplementación con PUFA.
En las perras lactantes, las concentraciones de GlycA, albúmina y creatinina se acercaban a los mismos valores que durante el anestro después de los cambios sorprendentes durante el embarazo y se mantuvieron solo ligeramente diferentes del anestro y dentro de los intervalos de referencia establecidos. Hay que tener en cuenta que después del parto también se produce la involución uterina y la reparación de los antiguos sitios placentarios en el útero (67). Estos fenómenos pueden dar lugar a marcadores inflamatorios elevados durante un tiempo determinado, incluso en perros con un puerperio normal. Al final de la lactancia, la concentración de GlycA o albúmina que cae fuera de los intervalos de referencia en las perras lactantes puede no ser fisiológica y debe evaluarse más a fondo. Los perros lactantes también tuvieron varias diferencias en las concentraciones séricas de aminoácidos en comparación con los perros en anestro: la glutamina, la histidina, la leucina y la fenilalanina disminuyeron, mientras que la glicina y la valina, así como las proporciones de glicina a valina y glicina a BCAA totales, aumentaron. Es probable que estos cambios reflejen la alta utilización de aminoácidos para satisfacer las altas concentraciones de proteínas de la leche secretada (68-70). Una ingesta suficiente de proteínas es esencial en las perras lactantes para satisfacer las altas demandas de proteínas de la lactancia (4). Se necesitan más estudios para evaluar si las concentraciones séricas de aminoácidos podrían reflejar los requisitos proteicos de la lactancia. Sin embargo, en este estudio, las concentraciones de aminoácidos de las perras lactantes rara vez cayeron fuera de los intervalos de referencia establecidos.
Además, la lactancia provocó múltiples cambios en el metabolismo energético, y la mayoría de los cambios en las concentraciones de lípidos durante el embarazo se revirtieron durante la lactancia. Se sabe que se producen cambios en el metabolismo de los lípidos y la glucosa en las madres humanas que amamantan, y las madres que amamantan generalmente tienen niveles más bajos de lípidos que las madres que no amamantan (71). El metabolismo energético durante la lactancia se caracteriza por una mayor tasa metabólica, un mayor gasto energético y la movilización de las reservas de grasa para satisfacer las necesidades lipídicas de la producción de leche, mientras que los hidratos de carbono siguen siendo los preferidos como fuente de energía (3, 4, 72-76). La leche es rica en lípidos (68-70) y los ácidos grasos saturados dominan su composición de ácidos grasos (68). El aumento de la producción y movilización de ácidos grasos saturados podría causar las concentraciones relativas más altas de ácidos grasos saturados en el suero de perros lactantes observadas en este estudio.
A excepción de la albúmina y el GlycA durante la gestación, los cambios en las concentraciones de metabolitos en los estados reproductivos estudiados no quedaron fuera de los intervalos de referencia de todos los perros. Por lo tanto, los intervalos de referencia establecidos para la mayoría de los metabolitos parecen ser válidos para las perras en todas las fases de la reproducción. Sin embargo, se debe prestar especial atención a la interpretación de los resultados de GlycA, albúmina y creatinina.
Curiosamente, el pseudoembarazo apenas se asoció con cambios metabólicos en comparación con el anestro. Este hallazgo sugiere que los cambios hormonales por sí solos no causan cambios metabólicos masivos, pero los cambios metabólicos durante el embarazo y la lactancia son causados específicamente por la implantación, el crecimiento fetal y la lactancia de los cachorros. Sin embargo, dado que se desconocían los días exactos del ciclo de las perras, el grupo de pseudoembarazo probablemente incluye tanto a las perras en metestro tardío como en anestro temprano, posiblemente con diferentes antecedentes hormonales.
De hecho, la mayor limitación de este estudio fue que tanto la salud del perro como el estado reproductivo fueron reportados por el propietario. Si bien la información de salud reportada por el propietario fue evaluada por un profesional veterinario, es posible que se haya producido un informe incorrecto del estado de enfermedad de los perros. Además, es posible que se produzcan clasificaciones erróneas del estado reproductivo. Para minimizar el posible sesgo, se eliminaron del conjunto de datos los casos poco claros basados en el estado informado por el propietario. Sin embargo, no se realizaron exámenes clínicos ni de laboratorio para la estadificación del ciclo. Dado que no se informó información sobre los síntomas relacionados con el calor o el día del ciclo, no se puede diferenciar el estro y el proestro, y las posibles dependencias, como la relación con el día de la ovulación o las interacciones con el aumento de las concentraciones de progesterona, no se pueden evaluar en este estudio. Dado que no se proporcionó una definición específica del término pseudoembarazo para los propietarios, no está claro si los propietarios clasificaron a sus perras como pseudoembarazadas en función del momento o de los signos evidentes de pseudoembarazo. Por lo tanto, las perras reportadas como pseudoembarazadas pueden incluir perras en metestro tardío y anestro temprano, y las perras reportadas en anestro pueden incluir perras en metestro. Además, la duración de la gestación y la lactancia era en su mayoría desconocida, y se desconocía la paridad y el tamaño de la camada. Por lo tanto, no es evidente en qué momento del embarazo y la lactancia se producen los cambios observados y si la paridad o el tamaño de la camada los afectan. Algunos cambios pueden incluso enmascararse, por ejemplo, si los parámetros cambian significativamente solo en una fase específica del embarazo o la lactancia. Por lo tanto, estudios posteriores con una fase del ciclo del celo definitivamente determinada e información detallada sobre la camada pueden revelar los efectos de la reproducción femenina sobre el metabolismo con mayor detalle. Sin embargo, los cambios observados en cada grupo se alinean con la literatura previa sobre otras especies.
El modelo de regresión logística multivariante de Firth creado fue capaz de diferenciar perras embarazadas sanas de perras no gestantes. Si bien su sensibilidad y especificidad fueron buenas en este conjunto de datos, el modelo tiene limitaciones. Al igual que otras pruebas de embarazo basadas en proteínas de fase aguda, solo son utilizables para la determinación del embarazo en individuos definitivamente sanos (2, 6, 11). Las afecciones inflamatorias, como la piometra, que es un diagnóstico diferencial grave para el embarazo y el pseudoembarazo, afectan a las proteínas de la fase aguda y pueden producirse diagnósticos erróneos graves si no se confirma el estado de salud de las perras (2, 6, 11). Del mismo modo, otras pruebas de embarazo basadas en proteínas de fase aguda solo son utilizables para la determinación del embarazo en individuos definitivamente sanos (2, 6, 11). Además, se debe estudiar más a fondo si el modelo funciona igual de bien en muestras de perras embarazadas y no embarazadas tomadas en días de ciclo similares. Por último, se debe estudiar durante qué período de tiempo del embarazo aumenta la GlycA y cuándo disminuye la albúmina.
Este estudio identificó varios cambios metabólicos en perras sanas durante el anestro, el celo, el pseudoembarazo, el embarazo y la lactancia. Comprender estos cambios es crucial para interpretar con precisión los valores de laboratorio y aumenta nuestra comprensión sobre las consecuencias metabólicas de la reproducción femenina. Un modelo de regresión logística de Firth multivariado basado en los marcadores inflamatorios albúmina y glicoproteína acetilos fue capaz de diferenciar perras gestantes de las no gestantes. Este hallazgo sugiere que, después de un estudio adicional, un modelo basado en datos de metabolitos podría usarse como método de prueba de embarazo en perros sanos.
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán puestos a disposición por los autores, sin reservas indebidas.
Declaración ética
El estudio en animales fue revisado y aprobado por el Comité de Ética Animal de la Oficina Provincial del Estado del Sur de Finlandia. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
CO: conceptualización, curación de datos, análisis formal, investigación, metodología, visualización, redacción, borrador original y redacción, revisión y edición. KV y SA: metodología y redacción, revisión y edición. AB: metodología, visualización y redacción: revisión y edición. NS: conceptualización, obtención de fondos, administración de proyectos, recursos, supervisión y redacción, revisión y edición. Todos los autores contribuyeron al artículo y aprobaron la versión presentada.
Financiación
La publicación en acceso abierto contó con el apoyo de la Universidad de Helsinki.
Reconocimientos
Jenni Puurunen fue reconocida por contribuir en los análisis iniciales de estos datos y por proporcionar la Figura 1 de los mensurandos de metabolitos analizados por la plataforma de RMN canina.
Conflicto de intereses
Este estudio recibió financiación de PetBiomics Ltd. El financiador tuvo la siguiente participación en el estudio: los salarios de CO y KV fueron financiados por PetBiomics Ltd. CO y KV eran empleados y HL el director de la junta y propietario de PetBiomics Ltd., una empresa que proporcionó la plataforma metabolómica de RMN usada para perros.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2023.1105113/full#supplementary-material
Referencias
1. Arlt SP. La perra alrededor del parto. Teriogenología. (2020) 150:452–7. doi: 10.1016/j.theriogenology.2020.02.046
Resumen de PubMed | Texto completo de CrossRef | Google Académico
2. Rootear Kustritz MV. Diagnóstico de embarazo y anomalías del embarazo en la perra. Teriogenología. (2005) 64:755–65. doi: 10.1016/j.theriogenology.2005.05.024
Resumen de PubMed | Texto completo de CrossRef | Google Académico
3. Fontaine E. Ingesta de alimentos y nutrición durante la gestación, lactancia y destete en la madre y las crías. Reprod Domest Anim. (2012) 47 Supl 6:326-30. doi: 10.1111/rda.12102
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Greco DS. Suplementos nutricionales para perras gestantes y lactantes. Teriogenología. (2008) 70:393–6. doi: 10.1016/j.theriogenology.2008.04.013
Resumen de PubMed | Texto completo de CrossRef | Google Académico
5. Wiles K, Bramham K, Seed PT, Nelson-Piercy C, Lightstone L, Chappell LC. Creatinina sérica en el embarazo: una revisión sistemática. Kidney Int Rep. (2019) 4:408–19. doi: 10.1016/j.ekir.2018.10.015
Resumen de PubMed | Texto completo de CrossRef | Google Académico
6. Vannucchi CI, Mirandola RM, Oliveira CM. Perfil proteico de fase aguda durante la gestación y diestro: propuesta de una prueba de embarazo precoz en perras. Anim Reprod Sci. (2002) 74:87–99. doi: 10.1016/s0378-4320(02)00160-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
7. Holst BS, Gustavsson MH, Johannisson A, Hillström A, Strage E, Olsson U, et al. Cambios inflamatorios durante el embarazo canino. Teriogenología. (2019) 125:285–92. doi: 10.1016/j.theriogenology.2018.11.016
Resumen de PubMed | Texto completo de CrossRef | Google Académico
8. Kuribayashi T, Shimada T, Matsumoto M, Kawato K, Honjyo T, Fukuyama M, et al. Determinación de proteína C reactiva (PCR) sérica en perros beagle sanos de diversas edades y perras beagle preñadas. Exp Anim. (2003) 52:387–90. doi: 10.1538/expanim.52.387
Resumen de PubMed | Texto completo de CrossRef | Google Académico
9. Kuribayashi T, Shimizu M, Shimada T, Honjyo T, Yamamoto Y, Kuba K, et al. Niveles de glicoproteína alfa 1 ácida (AAG) en perras beagle sanas y preñadas. Exp Anim. (2003) 52:377–81. doi: 10.1538/expanim.52.377
Resumen de PubMed | Texto completo de CrossRef | Google Académico
10. Larsson A, Palm M, Hansson L-O, Basu S, Axelsson O. Valores de referencia para la glicoproteína α1-ácida, α1-antitripsina, albúmina, haptoglobina, proteína C reactiva, IgA, IgG e IgM durante el embarazo. Acta Obstet Gynecol Scand. (2008) 87:1084–8. doi: 10.1080/00016340802428146
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Ulutas PA, Musal B, Kiral F, Bildik A. Niveles de proteínas en fase aguda en el embarazo y el ciclo de celo en perras. Res Vet Sci. (2009) 86:373–6. doi: 10.1016/j.rvsc.2008.09.001
Resumen de PubMed | Texto completo de CrossRef | Google Académico
12. Wang Q, Würtz P, Auro K, Mäkinen V-P, Kangas AJ, Soininen P, et al. Perfil metabólico del embarazo: evidencia transversal y longitudinal. BMC Med. (2016) 14:205. doi: 10.1186/s12916-016-0733-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
13. Handelman SK, Romero R, Tarca AL, Pacora P, Ingram B, Maymon E, et al. El metaboloma plasmático de las mujeres al principio del embarazo difiere del de las mujeres no embarazadas. PLoS UNO. (2019) 14:1–23. doi: 10.1371/journal.pone.0224682
Resumen de PubMed | Texto completo de CrossRef | Google Académico
14. Orczyk-Pawilowicz M, Jawien E, Deja S, Hirnle L, Zabek A, Mlynarz P. Metabolómica del líquido amniótico humano y el plasma materno durante el embarazo normal. PLoS UNO. (2016) 11:e0152740. doi: 10.1371/journal.pone.0152740
Resumen de PubMed | Texto completo de CrossRef | Google Académico
15. Pinto J, Barros AS, Domingues MRM, Goodfellow BJ, Galhano E, Pita C, et al. Después de un embarazo saludable por RMN, metabolómica del plasma y correlación con la orina. J Proteoma Res. (2015) 14:1263–74. doi: 10.1021/pr5011982
Resumen de PubMed | Texto completo de CrossRef | Google Académico
16. Wallace M, Hashim YZH-Y, Wingfield M, Culliton M, McAuliffe F, Gibney MJ, et al. Efectos de la fase del ciclo menstrual sobre los perfiles metabolómicos en mujeres premenopáusicas. Zumbido Reprod. (2010) 25:949–56. doi: 10.1093/humrep/deq011
Resumen de PubMed | Texto completo de CrossRef | Google Académico
17. Draper CF, Duisters K, Weger B, Chakrabarti A, Harms AC, Brennan L, et al. Rítmica del ciclo menstrual: patrones metabólicos en mujeres sanas. Sci Rep. (2018) 8:14568. doi: 10.1038/s41598-018-32647-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
18. de Gier J, Beijerink NJ, Kooistra HS, Okkens AC. Fisiología del anoestro canino y métodos para la manipulación de su longitud. Reprod Domest Anim. (2008) 43 Supl 2: 157-64. doi: 10.1111/j.1439-0531.2008.01156.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
19. Concannon PW. Ciclos reproductivos de la perra doméstica. Anim Reprod Sci. (2011) 124:200–10. doi: 10.1016/j.anireprosci.2010.08.028
Resumen de PubMed | Texto completo de CrossRef | Google Académico
20. Hinderer J, Lüdeke J, Riege L, Haimerl P, Bartel A, Kohn B, et al. Concentraciones de progesterona durante el embarazo canino. Anim MDPI. (2021) 11:3369. doi: 10.3390/ani11123369
Resumen de PubMed | Texto completo de CrossRef | Google Académico
21. Kooistra HS, Okkens AC. Secreción de prolactina y hormona del crecimiento en relación con la actividad ovárica en el perro. Reprod Domest Anim. (2001) 36:115–9. doi: 10.1046/j.1439-0531.2001.00311.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
22. Gobello C. Revisión de la pseudociesis canina. Teriogenología. (2021) 167:94–8. doi: 10.1016/j.theriogenology.2021.03.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
23. Root AL, Parkin TD, Hutchison P, Warnes C, Yam PS. Pseudoembarazo canino: una evaluación de la prevalencia y los protocolos de tratamiento actuales en el Reino Unido. BMC Vet Res. (2018) 14:170. doi: 10.1186/s12917-018-1493-1
Resumen de PubMed | Texto completo de CrossRef | Google Académico
24. Ottka C, Vapalahti K, Puurunen J, Vahtera L, Lohi H. Una nueva plataforma metabolómica basada en espectroscopia magnética nuclear canina: validación y manejo de muestras. Veterinario Clin Pathol. (2021) 2021:410–26. doi: 10.1111/vcp.12954
Resumen de PubMed | Texto completo de CrossRef | Google Académico
25. Soininen P, Kangas AJ, Wurtz P, Tukiainen T, Tynkkynen T, Laatikainen R, et al. Metabonómica de RMN sérica de alto rendimiento para estudios holísticos rentables sobre el metabolismo sistémico. Analista. (2009) 134:1781–5. DOI: 10.1039/B910205A
Resumen de PubMed | Texto completo de CrossRef | Google Académico
26. Soininen P, Kangas AJ, Würtz P, Suna T, Ala-Korpela M. Metabolómica cuantitativa de resonancia magnética nuclear sérica en epidemiología y genética cardiovascular. Circ Cardiovasc Genet. (2015) 8:192–206. doi: 10.1161/CIRCGENETICS.114.000216
Resumen de PubMed | Texto completo de CrossRef | Google Académico
27. Würtz P, Kangas AJ, Soininen P, Lawlor DA, Davey Smith G, Ala-Korpela M. Metabolómica cuantitativa de resonancia magnética nuclear sérica en epidemiología a gran escala: un manual de tecnologías ómicas. Am J Epidemiol. (2017) 186:1084–96. doi: 10.1093/aje/kwx016
Resumen de PubMed | Texto completo de CrossRef | Google Académico
28. Equipo central R. R: Un lenguaje y un entorno para la computación estadística. (2021). Disponible en línea en: https://www.r-project.org/ (consultado el 16 de noviembre de 2021).
29. Wickham H, François R, Henry L, Müller K. dplyr: Una gramática de la manipulación de datos. (2021). Disponible en línea en: https://cran.r-project.org/package=dplyr
30. Stekhoven DJ, Bühlmann P. MissImputación de valores faltantes no paramétricos para datos de tipo mixto. Bioinformática. (2012) 28:112–8. doi: 10.1093/bioinformática/BTR597
Resumen de PubMed | Texto completo de CrossRef | Google Académico
31. Ogle DH, Doll JC, Wheeler P, Dinno A. FSA: Análisis de las poblaciones de pesquerías. (2021). Disponible en línea en: https://github.com/droglenc/FSA
33. Kassambara A. ggpubr: Gráficos listos para publicaciones basadas en «ggplot2«. Paquete de R versión 0.4.0 (2020).
34. Gu Z, Eils R, Schlesner M. Los mapas de calor complejos revelan patrones y correlaciones en datos genómicos multidimensionales. Bioinformática. (2016) 32:2847–9. doi: 10.1093/bioinformatics/BTW313
Resumen de PubMed | Texto completo de CrossRef | Google Académico
35. Morgan M. BiocManager: Acceda al repositorio de paquetes del proyecto Bioconductor. (2021). Disponible en línea en: https://cran.r-project.org/package=BiocManager
36. Neuwirth E. RColorBrewer: Paletas ColorBrewer. (2014). Disponible en línea en: https://cran.r-project.org/package=RColorBrewer
Google Académico
37. Proyecto Inkscape. Paisaje de tinta. (2020). Disponible en línea en: https://inkscape.org (consultado el 20 de septiembre de 2022).
38. Firth D. Reducción del sesgo de las estimaciones de máxima verosimilitud. Biometría. (1993) 80:27–38. doi: 10.1093/biomet/80.1.27
39. Heinze G, Ploner M, Jiricka L. Regresión logística reducida por sesgo de Firth. (2022). Disponible en línea en: https://cran.r-project.org/package=logistf
40. Kuhn M. caret: Entrenamiento en clasificación y regresión. (2021). Disponible en línea en: https://cran.r-project.org/package=caret
41. Wickham H, Averick M, Bryan J, Chang W, McGowan L, François R, et al. Bienvenidos al Tidyverse. J Softw de código abierto. (2019) 4:1686. doi: 10.21105/joss.01686
42. Petrie A. regclass: Tools for an Introductory Class in Regression and Modeling. (2020). Available online at: https://cran.r-project.org/package=regclass
43. Hankin RKS. cmvnorm: The Complex Multivariate Gaussian Distribution. (2022). Available online at: https://cran.r-project.org/package=cmvnorm
44. Robin X, Turck N, Hainard A, Tiberti N, Lisacek F, Sanchez J-C, et al. pROC: un paquete de código abierto para R y S+ para analizar y comparar curvas ROC. BMC Bioinform. (2011) 12:77. doi: 10.1186/1471-2105-12-77
Resumen de PubMed | Texto completo de CrossRef | Google Académico
45. Schijf CP, van der Mooren MJ, Doesburg WH, Thomas CM, Rolland R. Diferencias en lípidos séricos, lipoproteínas, globulina fijadora de hormonas sexuales y testosterona entre la fase folicular y la lútea del ciclo menstrual. Acta Endocrinol (Copenh). (1993) 129:130–3. doi: 10.1530/acta.0.1290130
Resumen de PubMed | Texto completo de CrossRef | Google Académico
46. Bergström H, Ekström L, Warnqvist A, Bergman P, Björkhem-Bergman L. Variaciones en los biomarcadores de dislipidemia y disbiosis durante el ciclo menstrual: un estudio piloto en voluntarias sanas. BMC Salud de la Mujer. (2021) 21:166. doi: 10.1186/s12905-021-01306-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
47. Barnett JB, Woods MN, Lamon-Fava S, Schaefer EJ, McNamara JR, Spiegelman D, et al. Niveles plasmáticos de lípidos y lipoproteínas durante las fases folicular y lútea del ciclo menstrual. J Clin Endocrinol Metab. (2004) 89:776–82. doi: 10.1210/jc.2003-030506
Resumen de PubMed | Texto completo de CrossRef | Google Académico
48. Woods M, Schaefer EJ, Morrill A, Goldin BR, Longcope C, Dwyer JD, et al. Efecto de la fase del ciclo menstrual sobre los lípidos plasmáticos. J Clin Endocrinol Metab. (1987) 65:321–3. doi: 10.1210/jcem-65-2-321
Resumen de PubMed | Texto completo de CrossRef | Google Académico
49. Willson CJ, Chandra SA, Kimbrough CL, Jordan HL. Efecto de la fase del ciclo estral sobre los valores de patología clínica en perros beagle. Veterinario Clin Pathol. (2012) 41:71–6. doi: 10.1111/j.1939-165X.2011.00392.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
50. Moller SE, Moller BM, Olesen M, Fjalland B. Efectos de los anticonceptivos orales sobre los aminoácidos neutros plasmáticos y el colesterol durante un ciclo menstrual. Eur J Clin Pharmacol. (1996) 50:179–84. doi: 10.1007/s002280050089
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Spankowsky S, Heuwieser W, Arlt SP. ¿La administración oral del aminoácido tirosina afecta la concentración de estradiol-17( y el comportamiento sexual en la perra? Rec. Veterinario. (2013) 172:212. doi: 10.1136/vr.101240
Resumen de PubMed | Texto completo de CrossRef | Google Académico
52. Sociedad Internacional de Interés Renal (IRIS). Estadificación IRIS de la ERC (modificado en 2019). (2019). Disponible en línea en: http://www.iris-kidney.com/pdf/IRIS_Staging_of_CKD_modified_2019.pdf (consultado el 27 de diciembre de 2022).
53. Sociedad Internacional de Interés Renal (IRIS). Graduación de la Lesión Renal Aguda (modificado 2016). (2016). Disponible en línea en: http://www.iris-kidney.com/pdf/4_ldc-revised-grading-of-acute-kidney-injury.pdf (consultado el 27 de diciembre de 2022).
54. Semins MJ, Matlaga BR. Manejo de la urolitiasis en el embarazo. Int J Salud de la Mujer. (2013) 5:599–604. doi: 10.2147/IJWH. S51416
Resumen de PubMed | Texto completo de CrossRef | Google Académico
55. Maikranz P, Holley JL, Parks JH, Lindheimer MD, Nakagawa Y, Coe FL. La hipercalciuria gestacional causa sobresaturaciones patológicas de oxalato de calcio en orina. Riñón Int. (1989) 36:108–13. doi: 10.1038/ki.1989.168
56. Resim S, Ekerbicer HC, Kiran G, Kilinc M. Are changes in urinary parameters during pregnancy clinically significant? Urol Res. (2006) 34:244–8. doi: 10.1007/s00240-006-0051-7
57. Kerr GR. The free amino acids of serum during development of Macaca mulatto II. During Pregnancy and fetal life. Pediatr Res. (1968) 2:493–500. doi: 10.1203/00006450-196811000-00007
58. Felig P, Kim YJ, Lynch V, Hendler R. Amino acid metabolism during starvation in human pregnancy. J Clin Invest. (1972) 51:1195–202. doi: 10.1172/JCI106913
59. Okame R, Nakahara K, Murakami N. Plasma amino acid profiles at various reproductive stages in female rats. J Vet Med Sci. (2015) 77:815–21. doi: 10.1292/jvms.15-0095
60. Pastor-Anglada M, López-Tejero D, Remesar X. Free amino acid pools in some tissues of the pregnant rat. Horm Metab Res. (1986) 18:590–4. doi: 10.1055/s-2007-1012382
61. Metzger BE, Unger RH, Freinkel N. Carbohydrate metabolism in pregnancy. XIV Relationships between circulating glucagon, insulin, glucose and amino acids in response to a “mixed meal” in late pregnancy. Metabolism. (1977) 26:151–6. doi: 10.1016/0026-0495(77)90050-6
62. Schoengold DM, deFiore RH, Parlett RC. Free amino acids in plasma throughout pregnancy. Am J Obstet Gynecol. (1978) 131:490–9. doi: 10.1016/0002-9378(78)90108-4
63. Palou A, Arola L, Alemany M. Plasma amino acid concentrations in pregnant rats and in 21-day foetuses. Biochem J. (1977) 166:49–55. doi: 10.1042/bj1660049
64. Innis SM. Perinatal biochemistry and physiology of long-chain polyunsaturated fatty acids. J Pediatr. (2003) 143:S1–8. doi: 10.1067/s0022-3476(03)00396-2
65. Heinemann KM, Bauer JE. Docosahexaenoic acid and neurologic development in animals. J Am Vet Med Assoc. (2006) 228:655, 700–5. doi: 10.2460/javma.228.5.700
Resumen de PubMed | Texto completo de CrossRef | Google Académico
66. Heird WC, Lapillonne A. El papel de los ácidos grasos esenciales en el desarrollo. Annu Rev Nutr. (2005) 25:549–71. doi: 10.1146/annurev.nutr.24.012003.132254
Resumen de PubMed | Texto completo de CrossRef | Google Académico
67. Schulze LS-C, Heuwieser W, Arlt SP. Temperatura corporal de las perras en la primera semana después del parto medida por registradores ingeribles. Reprod Domest Anim. (2018) 53 Supl. 3:63-9. doi: 10.1111/rda.13330
Resumen de PubMed | Texto completo de CrossRef | Google Académico
68. Zhang M, Sun X, Cheng J, Guo M. Análisis y comparación de los perfiles nutricionales de la leche canina con la leche bovina y caprina. Alimentos (Basilea, Suiza). (2022) 11:30472. doi: 10.3390/alimentos11030472
Resumen de PubMed | Texto completo de CrossRef | Google Académico
69. Heinze CR, Freeman LM, Martin CR, Power ML, Fascetti AJ. Comparación de la composición de nutrientes de los sustitutos comerciales de la leche para perros con la de la leche para perros. J am vet med assoc. (2014) 244:1413–22. doi: 10.2460/javma.244.12.1413
Resumen de PubMed | Texto completo de CrossRef | Google Académico
70. Adkins Y, Lepine AJ, Lönnerdal B. Cambios en la composición de proteínas y nutrientes de la leche a lo largo de la lactancia en perros. Am J Vet Res. (2001) 62:1266–72. doi: 10.2460/ajvr.2001.62.1266
Resumen de PubMed | Texto completo de CrossRef | Google Académico
71. Gunderson EP. Impacto de la lactancia materna en el metabolismo materno: implicaciones para las mujeres con diabetes gestacional. Curr Diab Rep. (2014) 14:1–16. doi: 10.1007/s11892-013-0460-2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
72. Butte NF, Hopkinson JM, Mehta N, Moon JK, Smith EO. Ajustes en el gasto energético y la utilización del sustrato durante el final del embarazo y la lactancia. Am J Clin Nutr. (1999) 69:299–307. doi: 10.1093/ajcn/69.2.299
Resumen de PubMed | Texto completo de CrossRef | Google Académico
73. Motil KJ, Montandon CM, Garza C. Tasas metabólicas basales y postprandiales en mujeres lactantes y no lactantes. Am J Clin Nutr. (1990) 52:610–5. doi: 10.1093/ajcn/52.4.610
Resumen de PubMed | Texto completo de CrossRef | Google Académico
74. Scantlebury M, Butterwick R, Speakman JR. Energética de la lactancia en razas de perros domésticos (Canis familiaris) de dos tamaños. Comp Biochem Physiol A Mol Integr Physiol. (2000) 125:197–210. doi: 10.1016/s1095-6433(99)00175-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
75. Scantlebury M, Butterwick R, Speakman JR. Energética y variación del tamaño de la camada en razas de perros domésticos Canis familiaris de dos tamaños. Comp Biochem Physiol A Mol Integr Physiol. (2001) 129:919–31. doi: 10.1016/s1095-6433(01)00359-2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
76. Sadurskis A, Kabir N, Wager J, Forsum E. Metabolismo energético, composición corporal y producción de leche en mujeres suecas sanas durante la lactancia. Am J Clin Nutr. (1988) 48:44–9. doi: 10.1093/ajcn/48.1.44
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Palabras clave: perro, metabolómica, metabolismo, embarazo, lactancia, reproducción
Cita: Ottka C, Vapalahti K, Arlt SP, Bartel A y Lohi H (2023) Las diferencias metabólicas del anestro, el celo, el embarazo, el pseudoembarazo y la lactancia en 800 perras. Frente. Vet. Sci. 10:1105113. doi: 10.3389/fvets.2023.1105113
Recibido: 22 de noviembre de 2022; Aceptado: 16 de enero de 2023;
Publicado: 02 de febrero de 2023.
Editado por:
Josipa Kuleš, Universidad de Zagreb, Croacia
Revisado por:
Branimira Špoljarić, Universidad de Zagreb, Croacia
Alessandra Rota, Universidad de Pisa, Italia
Derechos de autor © 2023 Ottka, Vapalahti, Arlt, Bartel y Lohi. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Claudia Ottka,  claudia.ottka@petbiomics.com
claudia.ottka@petbiomics.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía