
Investigación de genes de resistencia a los antimicrobianos en productos probióticos para animales de compañía

Investigación de genes de resistencia a los antimicrobianos en productos probióticos para animales de compañía
 Adam Kerek1,2*
Adam Kerek1,2* Emese Szabó1Ábel Szabó1Márton Papp2,3Krisztián Bányai1,2,4Gábor Kardos2,5,6,7Eszter Kaszab2,5,8
Emese Szabó1Ábel Szabó1Márton Papp2,3Krisztián Bányai1,2,4Gábor Kardos2,5,6,7Eszter Kaszab2,5,8 Krisztina Bali2,8
Krisztina Bali2,8 Ákos Jerzsele1,2
Ákos Jerzsele1,2- 1Departamento de Farmacología y Toxicología, Universidad de Medicina Veterinaria, Budapest (Hungría)
- número arábigoLaboratorio Nacional de Enfermedades Infecciosas de los Animales, Resistencia a los Antimicrobianos, Salud Pública Veterinaria y Seguridad de la Cadena Alimentaria, Universidad de Medicina Veterinaria, Budapest, Hungría
- 3Centro de Bioinformática, Universidad de Medicina Veterinaria, Budapest (Hungría)
- 4Instituto de Investigación Médica Veterinaria, Budapest, Hungría
- 5One Health Institute, Facultad de Ciencias de la Salud, Universidad de Debrecen, Debrecen, Hungría
- 6Centro Nacional de Salud Pública, Budapest, Hungría
- 7Departamento de Metagenómica, Universidad de Debrecen, Debrecen, Hungría
- 8Departamento de Microbiología y Enfermedades Infecciosas, Universidad de Medicina Veterinaria, Budapest, Hungría
Introducción: Uno de los mayores desafíos de nuestro tiempo es la resistencia a los antimicrobianos, que podría convertirse en la principal causa de muerte a nivel mundial dentro de unas pocas décadas. En el contexto de «Una sola salud», redunda en interés común mitigar la propagación mundial de la resistencia a los antimicrobianos mediante la búsqueda de soluciones alternativas, junto con la selección adecuada de medicamentos y su uso responsable. Los probióticos ofrecen una vía potencial para reducir el uso de antibióticos; sin embargo, hay una escasez de investigaciones que examinen los productos comerciales en términos de portadores de genes de resistencia a los antimicrobianos (ARG) involucrados en el desarrollo de resistencia a través de vectores microbianos.
Métodos: Nuestro estudio investigó 10 productos probióticos disponibles comercialmente para gatos y perros. Inicialmente, realizamos pruebas fenotípicas a través de la determinación de la concentración inhibitoria mínima (CMI) para antibióticos importantes en la salud animal y pública. Posteriormente, realizamos la secuenciación de nueva generación (NGS) de los productos para dilucidar el trasfondo genético detrás de la disminución de la sensibilidad fenotípica.
Resultados: En total, se identificaron 19 tipos de ARG, de los cuales el 57,9% se encontraron en plásmidos, y en dos casos se encontraron portadores como elementos genéticos móviles. Uno de los genes identificados fue el gen APH(3′)-Ia, capaz de inactivar antibióticos aminoglucósidos a través de la regulación de la producción de enzimas fosfotransferasas, mientras que el otro fue el gen tetS, capaz de conferir una sensibilidad reducida a los antibióticos de tetraciclina a través de la protección de la diana.
Discusión: Nuestros hallazgos subrayan la importancia de abordar las investigaciones sobre la resistencia a los antimicrobianos desde una perspectiva más amplia. Sugerimos que se justifiquen más estudios en esta área y planteamos preguntas sobre la necesidad de ampliar los estudios legalmente requeridos sobre productos probióticos de su uso en ganadería económica a su uso en animales de compañía.
1 Introducción
La resistencia a los antimicrobianos es la propiedad de las bacterias que les permite sobrevivir al tratamiento dirigido con antibióticos (1). En la actualidad, las infecciones causadas por cepas resistentes y multirresistentes se están extendiendo cada vez más (2) que, según la Organización Mundial de la Salud (OMS), se convertirán en una de las 10 principales amenazas para la salud mundial en el siglo XXI (3).
Los probióticos son microorganismos vivos que, al entrar en el organismo, ejercen efectos beneficiosos sobre todo el organismo huésped. La mayoría pertenecen a cepas bacterianas que se producen de forma natural en el intestino. Las especies pertenecientes a los géneros Enterococcus, Lactobacillus y Bifidobacterium se utilizan comúnmente para este propósito, que es el mantenimiento o la restauración del equilibrio del microbioma intestinal (4). Aunque se han realizado numerosos estudios sobre sus efectos en el cuerpo humano, se han realizado significativamente menos investigaciones sobre sus efectos en animales de compañía. Además de los efectos intestinales, es probable que los probióticos ejerzan efectos inmunomoduladores y reductores del estrés en los animales de compañía y ayuden a equilibrar los parámetros sanguíneos relacionados con la digestión (5). Ciertas cepas probióticas también exhiben efectos antimicrobianos contra bacterias gastrointestinales patógenas específicas (6). La investigación existente sugiere que los efectos de los probióticos son más favorables para las especies de bacterias específicas del huésped, especialmente en las enfermedades gastrointestinales (7). Para los organismos probióticos, puede ser deseable tener resistencia antibacteriana para sobrevivir en el tratamiento con antibióticos. Sin embargo, las bacterias pueden transferir genes de resistencia entre sí vertical u horizontalmente, como resultado, pueden ser capaces de transferir genes de resistencia a bacterias patógenas. El tracto gastrointestinal de los mamíferos proporciona condiciones favorables para la transferencia de genes. En el caso de los productos para el ganado, las regulaciones prohíben la inclusión de genes de resistencia relevantes desde una perspectiva de salud pública, pero dicha regulación no existe para los animales de compañía (8).
Durante el examen de la cepa de Bacillus amyloliquefaciens utilizada como probiótico, se encontró que, aunque era sensible a las fluoroquinolonas, exhibía una sensibilidad reducida a los grupos antibacterianos más antiguos (9). Sin embargo, otro estudio no detectó genes de resistencia en esta especie (10). La resistencia intrínseca es característica de las especies de Lactobacillus, que generalmente muestran resistencia a múltiples grupos de antibióticos, como aminoglucósidos, glicopéptidos, inhibidores de la síntesis de ácidos nucleicos e inhibidores de la síntesis de folato (11). En varios estudios se han detectado genes de resistencia a la penicilina en varios aislamientos (12-14). También se ha detectado resistencia a oxacilina y cefalosporina en Lactobacillus plantarum y Lactobacillus rhamnosus (15, 16). Mater et al. demostraron que las cepas de Lactobacillus acidophilus son capaces de adquirir genes de resistencia a la vancomicina de especies de Enterococcus a través de la transferencia horizontal de genes (17). Los genes de resistencia más comunes en las especies de Lactobacillus confieren resistencia a las tetraciclinas y macrólidos. Los genes tetM y tetS se pueden encontrar tanto en plásmidos como en cromosomas. Sin embargo, el gen tetL solo se describe en plásmidos. El gen ermB responsable de la resistencia a la eritromicina también se encuentra en los plásmidos (18). También se ha demostrado que las cepas con más de un gen de resistencia a la tetraciclina exhiben una mayor resistencia, ya que estos genes tienen un efecto sinérgico (19). Feld et al. aislaron una cepa de Lactobacillus plantarum con un transposón del gen tetM capaz de transferirlo a otras bacterias productoras de ácido láctico (20).
En un estudio posterior, en el que se examinaron cepas de Pediococcus acidilactici frente a 21 agentes antimicrobianos diferentes (penicilina, oxacilina, ampicilina, piperacilina, imipenem, vancomicina, estreptomicina, gentamicina, amikacina, kanamicina, tetraciclina, cloranfenicol, minociclina, doxiciclina, cotrimoxazol, azitromicina, eritromicina, clindamicina, norfloxacina, ciprofloxacina y levofloxacino), se encontró que solo cuatro cepas mostraron sensibilidad a la piperacilina, imipenem, cloranfenicol y eritromicina. Si bien mostraron solo una resistencia moderada a la clindamicina, la doxiciclina y la levofloxacina, fueron totalmente resistentes a los agentes restantes (21). En otra investigación, una cepa de Pediococcus pentosaceus probada contra 19 agentes solo fue resistente a ceftazidima y sulfametoxazol. Por el contrario, una cepa de Enterococcus faecalis fue casi totalmente resistente a todos los agentes. Con base en esto, las especies de Pediococcus parecen ser más seguras en términos de resistencia como probióticos en comparación con las especies de Enterococcus, lo que significa que es menos probable que contribuyan a la propagación de la resistencia a los antimicrobianos (22). Sin embargo, Enterococcus faecium es el componente más común de las preparaciones probióticas para animales de compañía. Se encontró que una cepa probiótica examinada por Bs et al. era resistente a la penicilina, ampicilina, eritromicina, kanamicina y estreptomicina. Sin embargo, fue sensible a la tetraciclina, el cloranfenicol y la rifampicina (23).
Por lo tanto, es particularmente importante regular adecuadamente los productos probióticos comercializados para animales de compañía en términos de porte de genes de resistencia a los antimicrobianos. En nuestro estudio, investigamos los productos probióticos más comunes para animales de compañía disponibles en Hungría a través de la secuenciación de próxima generación y comparamos los resultados con los perfiles de resistencia fenotípica de las cepas aisladas de estos productos.
2 Materiales y métodos
2.1 El origen de los productos y cepas
Compramos 10 productos probióticos disponibles para perros y gatos en minoristas que venden productos veterinarios en Hungría. El aislamiento de las cepas indicadas en los productos fue llevado a cabo por el Departamento de Microbiología y Enfermedades Infecciosas de la Universidad de Medicina Veterinaria de Budapest. El aislamiento de las cepas de Enterococcus faecium fue exitoso en los nueve productos que la incluyeron (9/9). Entre las otras cepas enumeradas en los productos, pudimos aislar Lactobacillus plantarum en un caso (1/4), Pediococcus pentasaceus en dos casos (2/2) y Pediococcus acidilactici en un caso (1/2). Además, ciertos productos indicaron la presencia de cepas de Pediococcus (una cepa) y cepas de Lactococcus (ocho cepas) en forma desactivada. La identidad de especie de las cepas se determinó mediante espectrometría de masas MALDI-TOF (Bruker, Mannheim, Alemania). Los aislados se utilizaron para las pruebas de concentración inhibitoria mínima (CMI). Las propiedades de cada producto se resumen en la Tabla 1.
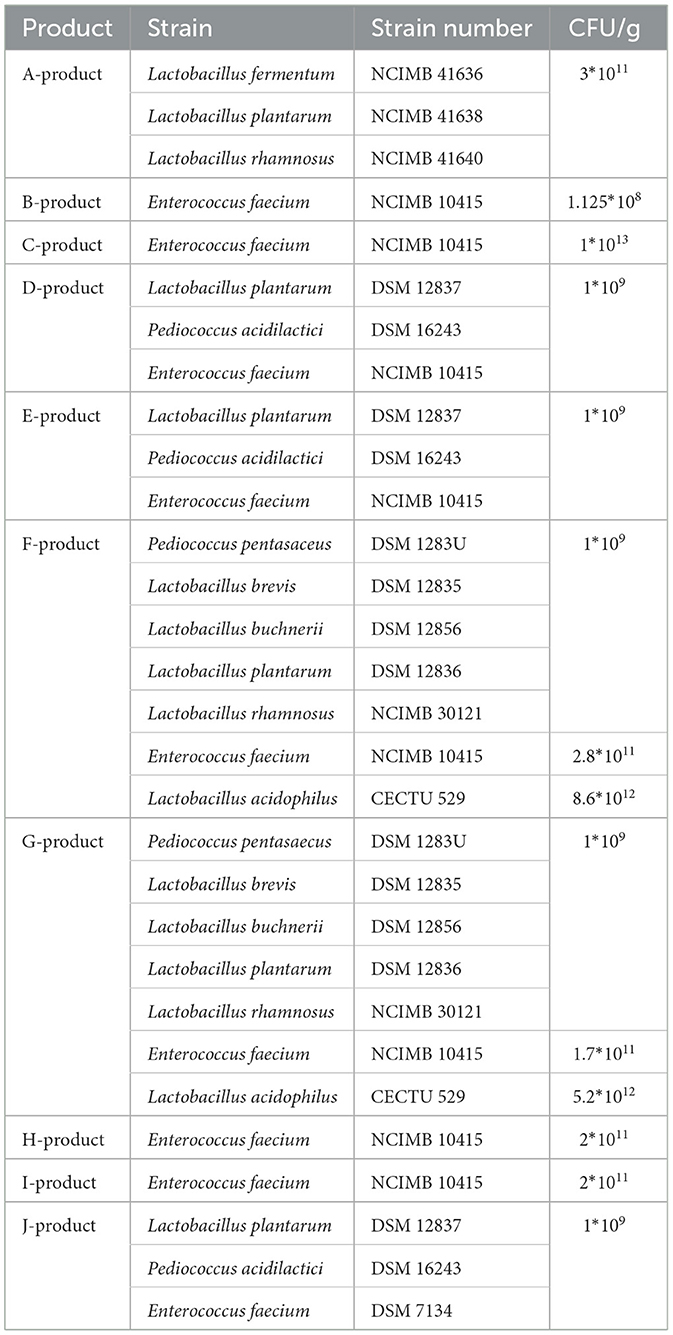
La clasificación de las cepas en categorías es supervisada por tres organizaciones internacionales principales. Según sus estándares, las cepas probióticas que se encuentran en los productos cuentan con identificadores únicos. La CECT (Colección Española de Cultivos Tipo) es una colección de cepas españolas acreditada con las normas ISO 9001 (24). La NCIMB (National Collection of Industrial, Food and Marine Bacteria) es una colección de cepas de propiedad privada ubicada en el Reino Unido (25). La DSM (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH) es una colección de cepas alemanas (26).
2.2 Preparación de la solución de antibióticos
Las soluciones de cepa se prepararon a partir de los ingredientes activos de acuerdo con las recomendaciones del Clinical Laboratory Standards Institute (CLSI), utilizando materiales de Merck KGaA, Darmstadt, Alemania (27). Para los experimentos, se prepararon soluciones de cepa a una concentración de 1.024 μg/mL, con ajustes para la pureza especificada por el fabricante de cada ingrediente activo. Los experimentos se llevaron a cabo dentro de un rango de dilución de 2 veces, que abarcó de 128 a 0,125 μg/mL.
2.3 Determinación de la concentración inhibitoria mínima
La expresión fenotípica de la resistencia se examinó mediante la determinación de los valores mínimos de concentración inhibitoria (CMI) de cada cepa bacteriana, lo que se llevó a cabo utilizando la metodología CLSI (27). Los puntos de corte se determinaron sobre la base de las directrices tanto del CLSI como del Comité Europeo de Pruebas de Susceptibilidad a los Antimicrobianos (EUCAST) (28). La selección de los agentes antimicrobianos se realizó con base en la literatura internacional, con el objetivo de cubrir los grupos de antimicrobianos más utilizados.
Las cepas bacterianas se almacenaron a -80 °C y se inocularon en 3 ml de caldo Mueller-Hinton (MHB) el día antes del experimento, seguido de una incubación a 37 °C durante 18-24 h. Los experimentos se llevaron a cabo utilizando una placa de microtitulación de 96 pocillos (VWR International, LLC., Debrecen, Hungría). La primera columna de las placas de trabajo, rellenada con 90 μl de MHB, recibió una dilución de 4 veces de las soluciones de tensión, estableciendo una concentración inicial de 128 μg/mL, a partir de la cual se preparó una serie de diluciones de 2 veces, excepto en las dos últimas columnas de la placa de microtitulación de 96 pocillos. La penúltima columna sirvió como control positivo (conteniendo solo bacterias), mientras que la última columna sirvió como control negativo (conteniendo solo el medicamento). La suspensión bacteriana ajustada a 0,5 de turbidez McFarland se inoculó en las placas de trabajo hasta la columna de control positivo utilizando un nefelómetro (CheBio fejleszto Kft., Budapest, Hungría) (27). La evaluación se llevó a cabo después de 24 h de incubación a 37 °C utilizando el lector automático MIC SWIN (CheBio fejleszto Kft., Budapest, Hungría) y el sistema VIZION (CheBio fejleszto Kft., Budapest, Hungría). El aislado de referencia utilizado fue Escherichia coli (ATCC 25922).
2.4 Secuenciación de nueva generación y análisis bioinformático
Las muestras de probióticos se prepararon utilizando 0,1 g de polvo probiótico disuelto en 1 ml de PBS (dilución 1:10). El ácido nucleico se extrajo de la mezcla utilizando el kit de minipreparación de microbios fecales/del suelo de ADN rápido (Zymo Research, CA, EE. UU.) de acuerdo con las instrucciones del fabricante. Las muestras se interrumpieron mediante el uso de un molino de bolas TissueLyzer LT (Qiagen, Alemania). La concentración de ADN purificado se midió con el equipo Qubit 2.0 utilizando el kit de ensayo Qubit dsDNA BR (Thermo Scientific, Waltham, MA, EE. UU.).
Las secuencias de nucleótidos se determinaron mediante secuenciación de próxima generación en un secuenciador Illumina® NextSeq 500 (Illumina, San Diego, CA, EE. UU.) siguiendo la guía de referencia proporcionada por Illumina. Para preparar las bibliotecas específicas de Illumina se utilizaron el kit de® preparación de bibliotecas de ADN Nextera XT (Illumina, San Diego, CA, EE. UU.) y el kit de índice Nextera XT v2 Set A (Illumina, San Diego, CA, EE. UU.) para preparar bibliotecas específicas de Illumina. Las muestras de ADN se diluyeron a 0,2 ng/μL en agua libre de nucleasas (Promega, Madison, WI, EE. UU.) en un volumen final de 2,5 μL. Los componentes de reacción se utilizaron a un volumen reducido. Para la reacción de etiquetado, se utilizaron 5 μL de tampón de ADN de Tagment con 2,5 μL de AmpliconTagment Mix. Durante el etiquetado, las muestras se incubaron a 55 °C durante 6 min, utilizando el sistema de PCR GeneAmp 9700 (Applied Biosystems/Thermo Fisher Scientific, Foster City, CA, EE. UU.). A continuación, se dejó enfriar las muestras a 10 °C antes de añadir 2,5 μL del tampón de etiquetado neutralizado. La neutralización se realizó durante 5 min a temperatura ambiente. A las muestras de ADN marcadas se les añadió un total de 7,5 μL de la mezcla maestra de PCR Nextera y 2,5 μL de cada uno de los cebadores índice i5 e i7. Los cebadores índice se adhirieron al ADN de la biblioteca a través de 12 ciclos de PCR (cada ciclo consistió en los siguientes pasos: 95 °C durante 10 s, 55 °C durante 30 s, seguidos de 72 °C durante 30 s). Después de los ciclos de PCR, las muestras se mantuvieron a 72 °C durante 5 min y luego a 10 °C. Las bibliotecas se purificaron utilizando el kit de extracción de fragmentos de ADN en gel/PCR (Geneaid Biotech Ltd., Taipei, Taiwán). La concentración de las bibliotecas purificadas se midió con el equipo Qubit 2.0 utilizando el kit de ensayo Qubit dsDNA HS (Thermo Fischer Scientific, Waltham, MA, EE. UU.). Los ADN de las bibliotecas se agruparon y se desnaturalizaron. El grupo de bibliotecas desnaturalizadas se cargó en una celda de flujo NextSeq 500/550 Mid Output a una concentración final de 1,5 pM. La secuenciación se realizó utilizando un secuenciador Illumina® NextSeq 500 (Illumina, San Diego, CA, EE. UU.).
Durante el procesamiento de los datos bioinformáticos, el control de calidad de las secuencias en bruto se realizó utilizando FastQC v0.11.9 (29), seguido de la eliminación de las secciones de baja calidad utilizando TrimGalore v0.6.6 (30). Las lecturas se ensamblaron en secuencias más largas (contigs) utilizando MEGAHIT v1.2.9 (31). A partir de estos contigs, se determinaron todos los posibles marcos de lectura abiertos (ORF) utilizando Prodigal v2.6.3 (32). Las secuencias de proteínas se derivaron de estos ORF en función de sus secuencias de nucleótidos. Posteriormente, las secuencias de proteínas se compararon con las secuencias de genes de resistencia a los antimicrobianos (ARG) en la Base de Datos Integral de Resistencia a los Antibióticos (CARD) utilizando el software Resistance Gene Identifier (RGI) v5.1.0 (33). Solo se conservaron los resultados que alcanzaron el umbral especificado (95%) en la base de datos CARD.
La movilidad potencial de los genes de resistencia identificados se evaluó utilizando el programa MobileElementFinder v1.0.3 (34), que predijo los genes que ocurren como elementos genéticos móviles (MGE) en los contigs. Solo se consideraron potencialmente móviles aquellos ARG encontrados dentro de la distancia de transposón compuesto más larga específica para la especie en la base de datos dentro del contig. Además, se examinó la codificación de plásmidos utilizando el software PlasFlow v1.1, y la presencia de genomas de fagos en contigs se determinó utilizando el software VirSorter v2.2.2 (35).
3 Resultados
3.1 Cepas de Enterococcus faecium
En la Tabla 2 se resumen los resultados de las pruebas de susceptibilidad de las cepas de Enterococcus faecium aisladas de los productos examinados. Todos los productos contenían la misma cepa (NCIMB10415) excepto el producto J (DSM7134). Se observaron valores de CMI similares para todas las cepas. Todas las cepas analizadas fueron sensibles a la penicilina, amoxicilina, amoxicilina-clavulanato, oxitetraciclina, doxiciclina, clindamicina, tilosina y vancomicina. Para gentamicina (100%; CMI >32 μg/mL) y sulfonamidas potenciadas (sulfametoxazol y trimetoprima en una proporción de 19:1; 100%; CMI 16 μg/mL), todas las cepas fueron resistentes. En cuanto a la gatifloxacina, fluoroquinolona de cuarta generación, seis cepas fueron susceptibles, mientras que tres fueron resistentes (66,7%; CMI >2 μg/mL).
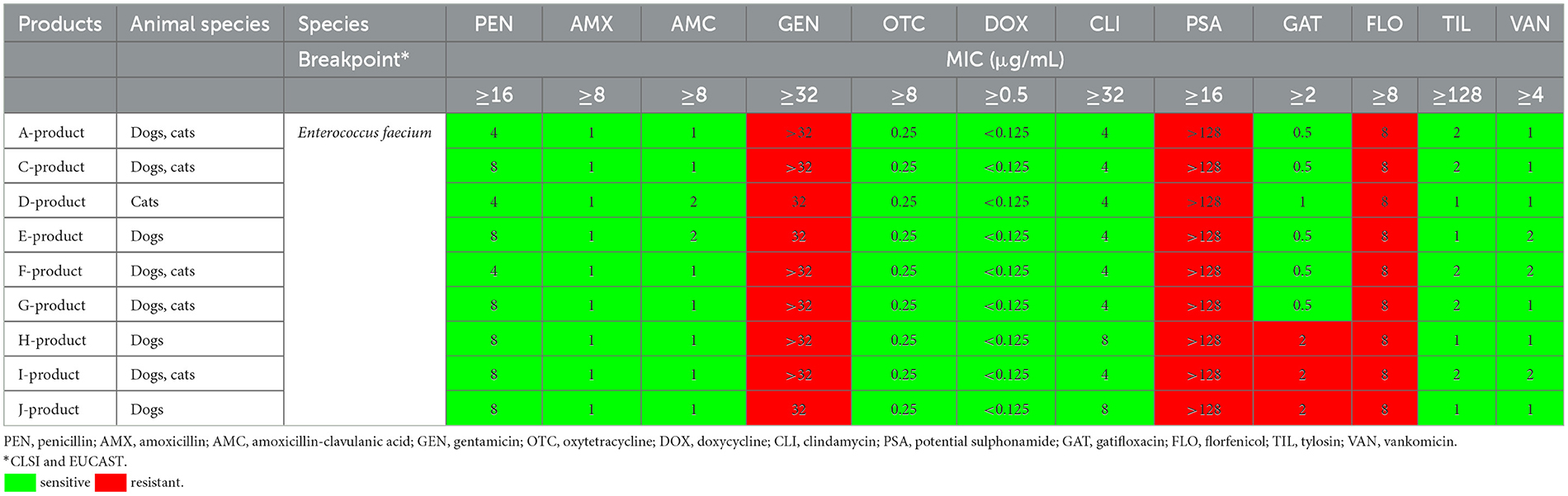
Tabla 2. Valores de concentración inhibitoria mínima (CMI) de cepas de Enterococcus faecium aisladas de los productos para antibióticos analizados.
3.2 Cepas de Lactobacillus y Pediococcus
En la Tabla 3 se resumen los perfiles de susceptibilidad a los antibióticos de las especies de Lactobacillus y Pediococcus. Entre las cepas de Lactobacillus, se aisló una cepa de Lactobacillus plantarum, que mostró sensibilidad a la penicilina, amoxicilina-clavulanato, gentamicina, oxitetraciclina, doxiciclina, clindamicina, tilosina y vancomicina. Fue resistente a la amoxicilina (>1 μg/mL de MIC), a la combinación de sulfonamidas (>16 μg/mL de MIC) y a gatifloxacino (>2 μg/mL de MIC). De las especies de Pediococcus, se aislaron dos cepas de Pediococcus pentasaceus y una de Pediococcus acidilactici, todas las cuales fueron susceptibles solo a clindamicina (< 8 μg/mL MIC), mostrando resistencia a todos los demás antibióticos probados.
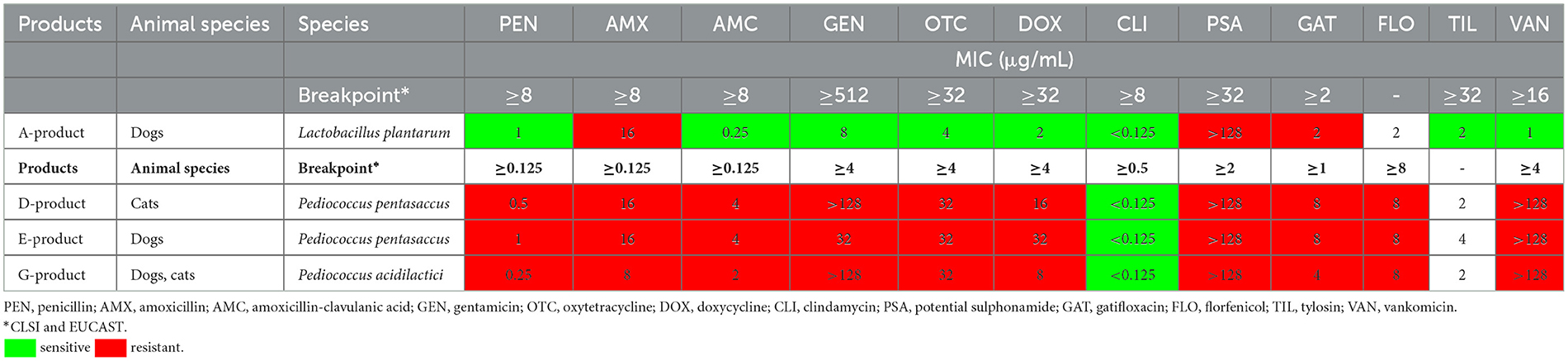
Tabla 3. Valores de concentración inhibitoria mínima (CMI) de las cepas de Lactobacillus y Pediococcus aisladas de los productos para los antibióticos probados.
3.3 Resultados del análisis metagenómico
En la Tabla 4 se resumen los resultados obtenidos de la secuenciación de nueva generación, incluyendo los genes de resistencia aislados de los productos, su origen taxonómico, indicando la familia ARG para cada gen, el mecanismo de resistencia determinado por el gen respectivo y los grupos antibióticos contra los que se confiere resistencia.
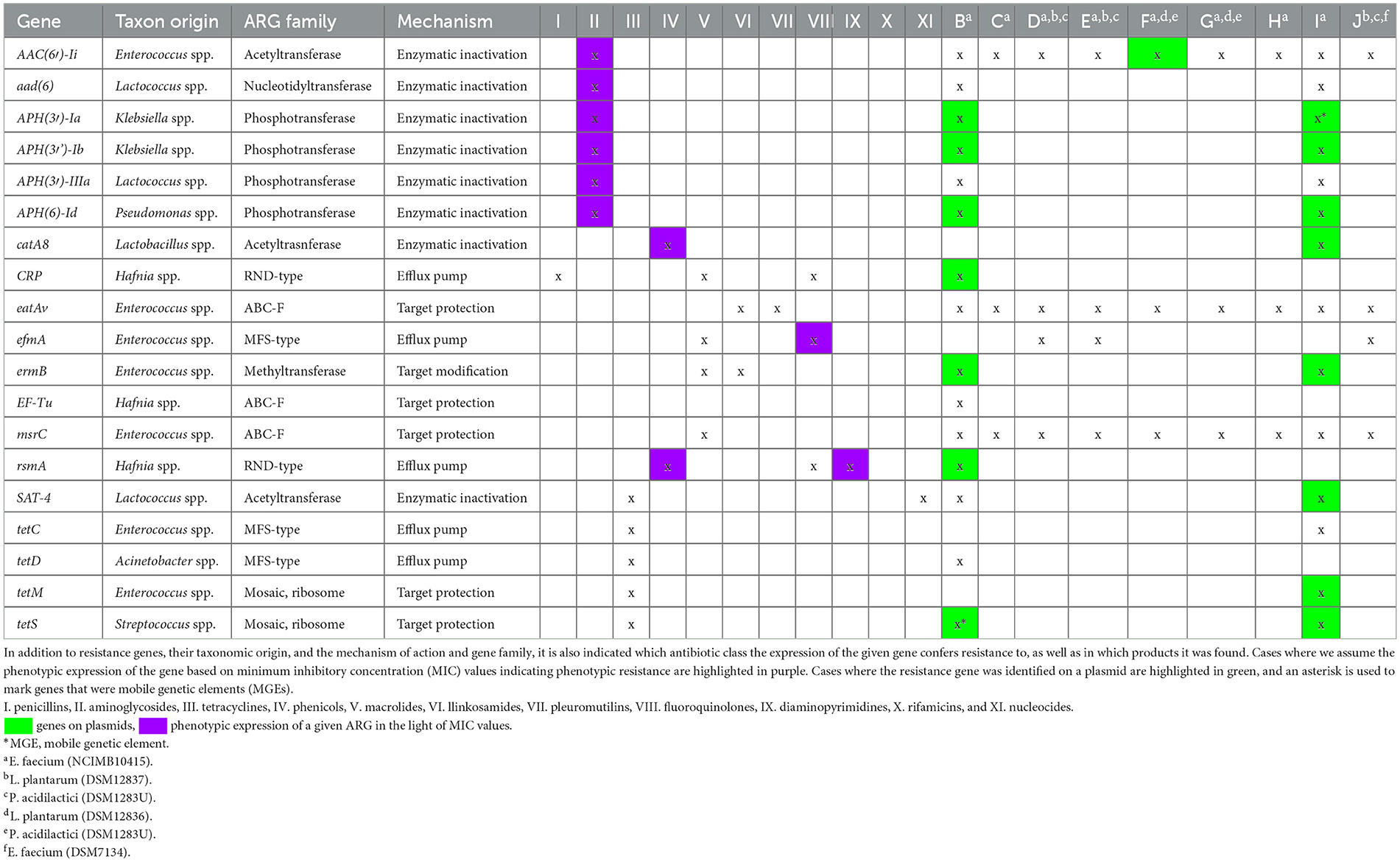
Se identificaron un total de 19 tipos de ARG, de los cuales 11 se encontraron en plásmidos, y no se identificaron genes en fagos. De particular preocupación es que en dos casos, un gen contenido en plásmidos también actuó como un elemento genético móvil (MGE). Uno de ellos fue el gen APH(3′)-Ia, que se encuentra en el producto I. La activación de este gen de resistencia por medios enzimáticos (fosfotransferasa) puede inactivar antibióticos aminoglucósidos (36). El otro gen era el gen tetS, que se encuentra en el producto A. Este gen es capaz de reducir la sensibilidad a los antibióticos tetraciclina a través de la protección diana (mosaico ribosómico), evitando así la unión del antibiótico (37).
Durante la determinación de los valores de MIC, se puede observar una correlación entre los valores de resistencia y los ARG identificados en las cepas de Enterococcus faecium de varios productos. La expresión enzimática del gen AAC(6′)-Ii (acetiltransferasa) conduce a la resistencia a los antibióticos aminoglucósidos. Este gen se encontró en todos los productos probados y podría ser uno de los genes responsables de la resistencia (≥32 μg/mL) observada en todas las cepas. Por lo general, se localiza cromosómicamente, pero en un caso (producto F), se encontró en un plásmido. El gen eatAv confiere resistencia a lincosamidas y pleuromutilinas a través de la protección de dianas (tipo ABC-F). Este gen estaba presente en todas las cepas de Enterococcus faecium de los productos analizados. El gen efmA determina una bomba de eflujo de tipo MFS para macrólidos y fluoroquinolonas, lo que podría contribuir a la resistencia a gatifloxacina (2 μg/mL MIC) observada en un caso (producto J). El gen rsmA determina una bomba de eflujo multifármaco de tipo RND, que puede conferir resistencia a fenicoles, fluoroquinolonas y diaminopirimidinas. Este gen también se encontró en todas las cepas de Enterococcus faecium y puede explicar la resistencia observada (>128 μg/mL de MIC) a las sulfamidas potenciadas, gatifloxacino (2 μg/mL de MIC) en los casos de productos H, I y J, así como la posible resistencia al florfenicol (8 μg/mL de MIC).
Todas las resistencias fenotípicas se atribuyeron con éxito a los ARG identificados. De particular preocupación es la alta proporción de genes encontrados en plásmidos (57,9%), con dos casos en los que estos también eran MGE. Por lo tanto, sobre la base de nuestros hallazgos, sería necesario realizar tales exámenes, similares a los requeridos para los animales destinados a la producción de alimentos, antes de introducir en la circulación preparaciones probióticas destinadas a animales de compañía. En el producto A, que contiene cepas de Lactobacillus plantarum (NCIMB41638), Lactobacillus fermentum (NCIMB41636) y Lactobacillus rhamnosus (NCIMB41640), no se encontraron ARG. Por lo tanto, la resistencia fenotípica observada en este caso no puede ser respaldada genéticamente por los genes detectados.
El producto B contenía Enterococcus faecium (NCIMB10415), de los cuales identificamos 15 ARG, de los cuales siete se localizaban en plásmidos y un gen (tetS) en un MGE. Cuatro genes se derivaron específicamente de especies de Enterococcus, mientras que el resto se originó de especies de Lactococcus, Klebsiella, Pseudomonas, Hafnia, Acinetobacter y Streptococcus. Del mismo modo, en el producto C y el producto H que contenían Enterococcus faecium (NCIMB10415), identificamos 3 ARGs, todos ellos genes cromosómicos originarios de Enterococcus spp. El producto D, el producto E y el producto J que contenían las especies Lactobacillus plantarum (DSM12837), Pediococcus acidilactici (DSM16243) y Enterococcus faecium (NCIMB10415), respectivamente, mostraron la presencia de los mismos cuatro ARG. Estos genes fueron localizados cromosómicamente, y todos se originaron a partir de Enterococcus spp.
El producto F contenía cepas de Pediococcus pentasaceus (DSM1283U), Lactobacillus brevis (DSM12835), Lactobacillus bucherii (DSM12856), Lactobacillus plantarum (DSM12836), Lactobacillus rhamnosus (NCIMB30121), Lactobacillus acidophilus (CECTU529) y Enterococcus faecium (NCIMB10415), con Pediococcus y Lactobacillus especies presentes en forma desactivada en el producto. Cuatro genes de resistencia coincidían con los encontrados en el producto E. Sin embargo, en el caso del producto F, el gen AAC(6′)-Ii se localizaba en un plásmido, mientras que el resto eran cromosómicos. Todos los genes identificados se atribuyeron a Enterococcus spp. La composición del producto G era idéntica a la del producto F, pero solo se pudieron identificar en los comprimidos dos ARG localizados cromosómicamente procedentes de Enterococcus spp.
El producto I, que contenía la cepa de Enterococcus faecium (NCIMB10415), contenía el mayor número de ARG identificados, con un total de 14. De estos, ocho se encontraron en plásmidos, y entre estos, el gen APH(3‘)-Ia también era un MGE. Cinco de los genes eran originalmente de origen Enterococcus spp., mientras que el resto puede haber sido adquirido de otras especies a través de la transferencia horizontal de genes, incluyendo las especies Staphylococcus, Klebsiella, Lactobacillales, Enterobacteriaceae, Enterobacterales y Streptococcus.
4 Discusión
Se investigó la susceptibilidad antibiótica in vitro de cepas probióticas aisladas de un total de 10 productos probióticos disponibles comercialmente para animales de compañía (perros, gatos), y se identificaron 19 ARG diferentes durante la exploración de los productos a través de la secuenciación de próxima generación.
Se observó resistencia a gentamicina en todas las cepas de Enterococcus faecium examinadas en nuestro estudio, con valores de CMI >32 μg/mL. Por el contrario, en el estudio realizado por Takeuchi et al., este valor superó los 500 μg/mL en el 22% de las cepas aisladas de Enterococcus faecium (38). Xu et al. detectaron valores extremadamente altos (9.000 μg/mL de MIC) en una cepa aislada de aguas residuales durante las pruebas de susceptibilidad antimicrobiana contra la sulfonamida (sulfametoxazol-trimetoprim) (39). En nuestra investigación se identificaron valores significativamente más bajos (16 μg/mL de MIC). De manera similar, Maasjost et al (40) obtuvieron resultados comparables para la susceptibilidad a la gentamicina y la sulfonamida en aislados de aves de corral (32 μg/mL de MIC, 16 μg/mL de MIC). También se observó resistencia a gatifloxacino en tres cepas, aunque a niveles relativamente bajos (>2 μg/mL de MIC). Este hallazgo está respaldado por varios estudios previos. Wenzler et al. trabajaron con aislados clínicos, donde el valor medio (2 μg/mL de MIC) coincidió con los descritos en nuestro estudio (41).
Nueve de las preparaciones que examinamos contenían Enterococcus faecium, en el que estaban presentes los 19 tipos de ARG. Xia et al. identificaron 18 ARGs en cepas de Enterococcus faecium, ninguna de las cuales era MGE (42), en contraste, describimos la probabilidad de movilidad para dos genes [APH(3′)-Ia, tetS]. Varios de los genes identificados confieren resistencia a los aminoglucósidos a través de diferentes vías enzimáticas. El gen AAC(6′)-Ii es una aminoglucósido transferasa que se encuentra en el cromosoma, descrita por primera vez en Enterococcus faecium (43). El gen AAD(6), que se encuentra en un plásmido, codifica la nucleótido transferasa aminoglucósido (44). Los miembros de la familia de genes APH (3′) confieren resistencia a los aminoglucósidos a través de la inactivación enzimática. El gen APH(3′)-Ia se encuentra en un transposón en las especies de Escherichia coli y Salmonella (36). El gen APH(3′)-IIIa está codificado por plásmidos, encontrándose en las especies de Staphylococcus aureus y Enterococcus (45). El gen APH(3′)-Ib se identificó en Escherichia coli, también encontrado en un plásmido (46). El APH(6)-Id opera a través de un mecanismo similar. Este gen se puede encontrar en un plásmido, transposón o cromosoma (47).
El gen catA8 codifica una enzima cloranfenicol acetiltransferasa. A través de la inactivación enzimática, neutraliza los fenicoles, principalmente el cloranfenicol (48). El análisis metagenómico del gen catA8 identificado por nosotros apoyó su origen en Lactobacillus y su presencia en un plásmido. El gen eatAv se identificó por primera vez en la bacteria Enterococcus faecium. Su mecanismo implica la protección de la diana, confiriendo resistencia a pleuromutilinas y lincosamidas (49). La expresión de ermB es inducida por el agente eritromicina. Como resultado, se desarrolla resistencia contra macrólidos y lincosamidas a través de la modificación de la diana (50).
El gen msrC, que se encuentra típicamente en las cepas de Enterococcus faecium, es un gen codificado cromosómicamente que confiere resistencia principalmente a los macrólidos, específicamente a la eritromicina. A través de la protección dirigida, previene la acción de los antibióticos (51). Observamos la expresión fenotípica de este gen en la mayoría de los casos en función de los valores de MIC, y en una preparación, lo identificamos en un plásmido. Urshev et al. detectaron el gen msrC en una cepa de Enterococcus faecium (52). Thumu et al. identificaron por primera vez el gen msrC en las especies de Pediococcus pentasaceus, pero también detectaron la presencia del gen ermB (53). En nuestras investigaciones, identificamos ambos genes en varias preparaciones, pero basándonos en el análisis metagenómico, encontramos que estos genes eran de origen Enterococcus spp. Sin embargo, fenotípicamente, observamos la expresión de resistencia definida por ellos. El gen rsmA codifica una pequeña proteína de unión al ARN que desempeña un papel regulador postranscripcional en la formación de los genes de virulencia de Pseudomonas aeruginosa. Confiere resistencia a diaminopirimidinas, fenicoles y fluoroquinolonas a través de un mecanismo de bomba de eflujo (54). El gen SAT-4, derivado de la bacteria Campylobacter coli, es una estreptotricina acetiltransferasa mediada por plásmidos (55).
El gen tetC es responsable de conferir resistencia a los antibióticos tetraciclina, principalmente en bacterias Gram-negativas, ya que regula la expresión de un gen de bomba de eflujo, que generalmente se encuentra en plásmidos. El gen tetD es similar, pero está presente exclusivamente en las especies Gram-negativas. La resistencia se desarrolla a través de mecanismos de protección del objetivo (56). Los genes tetM y tetS son responsables de las proteínas de protección de los ribosomas y se encuentran en elementos genéticos móviles, que se encuentran tanto en bacterias Gram-negativas como Gram-positivas. Al igual que Pan et al. (57), también identificamos el gen tetM en un plásmido, pero durante este estudio, se identificó que se originaba en Enterococcus. El gen tetS que detectamos resultó ser de origen Streptococcus spp., encontrado en un plásmido, y era móvil en el caso del producto B. Nawaz et al. identificaron estos genes durante su estudio de Lactobacillus plantarum, transfiriendo con éxito los genes a aislados de Enterococcus en condiciones experimentales (13). En nuestras investigaciones, el gen ermB identificado en los preparados que contenían Lactobacillus plantarum siempre resultó ser de origen Enterococcus spp.
Sobre la base de los resultados, se recomienda en la práctica considerar la introducción de regulaciones más estrictas durante la producción y la autorización de comercialización de probióticos, asegurando la reducción del grupo de ARG en los productos a un nivel mínimo, con especial atención a la exclusión de genes que pueden tener una importancia significativa para la salud pública. Las evaluaciones preliminares del perfil de resistencia durante la selección de cepas probióticas reducen la probabilidad de propagación de resistencias que pueden transferirse fácilmente, creando un puente entre la salud animal y la salud pública, y planteando un riesgo significativo a través de su uso. En todos los casos de probióticos, sería necesario compilar un panel de sustancias activas antibióticas basado en los resultados de las pruebas fenotípicas, sobre el cual se llevaría a cabo la exploración de los antecedentes genéticos mediante secuenciación de nueva generación, con especial atención al transporte de ARG en plásmidos o fagos, así como a su codificación como MGE. La exclusión de estos es esencial para minimizar las posibilidades de transferencia horizontal de genes. Las limitaciones de nuestros estudios incluyen que se llevaron a cabo con solo unos pocos productos involucrados. En el futuro, definitivamente vale la pena involucrar una gama más amplia de productos en los estudios y realizar más investigaciones paralelas para permitir análisis estadísticos.
5 Conclusión
Podemos concluir que, entre las cepas probióticas estudiadas, la bacteria Enterococcus faecium es la portadora de más genes de resistencia, de acuerdo con la literatura existente. Entre las especies de Lactobacillus y Pediococcus, identificamos genes de resistencia de cepas de Enterococcus en la mayoría de los casos, lo que también coincide con la literatura. Detectamos un gen derivado de Lactobacillus plantarum (efmA) responsable de la resistencia, que no había sido descrito previamente. La mayoría de los ARG identificados (57,9%) se encontraron en plásmidos; en dos preparaciones, el producto B (gen tetS) y el producto I [gen APH(3′)-Ia], identificamos ARG transmitidos por plásmidos, que también sirven como genes MGE. La literatura sobre este tema es relativamente escasa, particularmente en lo que respecta a los estudios que exploran el repertorio de ARG en preparaciones probióticas destinadas a animales de compañía. Nuestros resultados subrayan la importancia de llevar a cabo este tipo de estudios, y puede valer la pena considerar la posibilidad de imponer legalmente dichas investigaciones como condición para la distribución, de forma similar a la legislación aplicada a los productos destinados al ganado.
Declaración de disponibilidad de datos
Los conjuntos de datos presentados en este estudio se pueden encontrar en repositorios en línea. Los nombres del repositorio o repositorios y el número de acceso se pueden encontrar en el artículo/Material complementario.
Contribuciones de los autores
AK: Conceptualización, Metodología, Administración de proyectos, Supervisión, Visualización, Redacción – borrador original. ES: Curación de datos, Investigación, Redacción – borrador original. ÁS: Recursos, Redacción – borrador original. MP: Análisis formal, Software, Redacción – revisión y edición. KBán: Validación, Redacción – revisión y edición. GK: Validación, Redacción – revisión y edición. EK: Metodología, Redacción – revisión y edición. KBal: Metodología, Redacción – revisión y edición. ÁJ: Conceptualización, Obtención de fondos, Validación, Redacción – revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue apoyada por el Proyecto No. El MRR-2.3.1-21-2022-00001 se implementó con el apoyo proporcionado por el Mecanismo de Recuperación y Resiliencia (MRR), financiado en el marco de la estimación presupuestaria del Fondo Nacional de Recuperación, esquema de financiación RRF-2.3.1-21. Elaborado con el apoyo profesional del Programa de Becas para Doctorandos del Programa Cooperativo de Doctorado del Ministerio de Innovación y Tecnología financiado por el Fondo Nacional de Investigación, Desarrollo e Innovación.
Reconocimientos
Agradecemos al personal del Departamento de Microbiología y Enfermedades Infecciosas por su ayuda para aislar las cepas de los preparados. Además, extendemos nuestro agradecimiento a los asistentes del Laboratorio de Microbiología del Departamento de Farmacología y Toxicología por su apoyo en los trabajos preparatorios de nuestras investigaciones.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda hacer su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1464351/full#supplementary-material
Referencias
1. Alós J-I. Resistencia bacteriana a los antibióticos: una crisis global. Enfermedades Infecciosas y Microbiología Clínica. (2015) 33:692–9. doi: 10.1016/j.eimc.2014.10.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Munita JM, Arias CA. Mecanismos de resistencia a antibióticos. Microbiol Spectr. (2016) 4:15. DOI: 10.1128/microbiolspec. VMBF-0016-2015
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. QUIÉN. Se puso en marcha el Foro Mundial sobre la Resistencia a los Antimicrobianos para ayudar a hacer frente a las amenazas comunes para la salud del planeta. Disponible en: https://www.who.int/news-room/articles-detail/global-antimicrobial-resistance-forum-launched-to-help-tackle-common-threat-to-planetary-health (consultado el 14 de mayo de 2023).
4. Wilkins T, Sequoia J. Probióticos para afecciones gastrointestinales: un resumen de la evidencia. Am Médico de la Fam. (2017) 96:170–8.
5. Grześkowiak Ł, Endo A, Beasley S, Salminen S. Microbiota y probióticos en el bienestar canino y felino. Anaerobio. (2015) 34:14–23. doi: 10.1016/j.anaerobe.2015.04.002
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. He Y, Li F, Zhang W, An M, Li A, Wang Y, et al. Potencial probiótico de Bacillus amyloliquefaciens aislado de yaks tibetanos. Probiot Antimicr Prot. (2022) 22:5. doi: 10.1007/s12602-022-10027-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Kumar S, Pattanaik AK, Sharma S, Jadhav SE, Dutta N, Kumar A. Potencial probiótico de una bacteria Lactobacillus de origen fecal canino y su impacto en índices de salud intestinal seleccionados y respuesta inmune de perros. Probióticos: Proteínas antimicrobianas. (2017) 9:262–77. doi: 10.1007/s12602-017-9256-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Osmanagaoglu O, Kiran F, Ataoglu H. Evaluación del potencial probiótico in vitro de Pediococcus pentosaceus OZF aislado de leche materna humana. Probiot Antimicr Prot. (2010) 2:162–74. doi: 10.1007/s12602-010-9050-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Du H, Yao W, Kulyar MF-A, Ding Y, Zhu H, Pan H, et al. Efectos de Bacillus amyloliquefaciens TL106 aislado de cerdos tibetanos sobre el potencial probiótico y los microbios intestinales en lechones destetados. Microbiol Spectr. (2021) 10:E01205–21. doi: 10.1128/spectrum.01205-21
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Islam MI, Seo H, Redwan A, Kim S, Lee S, Siddiquee M, et al. Efecto anti-Clostridioides difficile in vitro e in vivo de una cepa probiótica de Bacillus amyloliquefaciens. J Microbiol Biotechnol. (2022) 32:46–55. doi: 10.4014/jmb.2107.07057
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Charteris WP, Kelly PM, Morelli L, Collins JK. Desarrollo y aplicación de una metodología in vitro para determinar la tolerancia al tránsito de especies potencialmente probióticas de Lactobacillus y Bifidobacterium en el tracto gastrointestinal humano superior. J Appl Microbiol. (1998) 84:759–68. doi: 10.1046/j.1365-2672.1998.00407.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Charteris WP, Kelly PM, Morelli L, Collins JK. Pruebas de sensibilidad a antibióticos por difusión en gradiente de lactobacilos potencialmente probióticos. J Food Prot. (2001) 64:2007–14. doi: 10.4315/0362-028X-64.12.2007
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Nawaz M, Wang J, Zhou A, Ma C, Wu X, Moore JE, et al. Caracterización y transferencia de resistencia a antibióticos en bacterias lácticas a partir de productos alimenticios fermentados. Curr Microbiol. (2011) 62:1081–9. doi: 10.1007/s00284-010-9856-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Belletti N, Gatti M, Bottari B, Neviani E, Tabanelli G, Gardini F. Resistencia a los antibióticos de lactobacilos aislados de dos quesos duros italianos. J Food Prot. (2009) 72:2162–9. doi: 10.4315/0362-028X-72.10.2162
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Coppola R, Succi M, Tremonte P, Reale A, Salzano G, Sorrentino E. Susceptibilidad a los antibióticos de las cepas de Lactobacillus rhamnosus aisladas del queso Parmigiano Reggiano. Lait. (2005) 85:193–204. doi: 10.1051/LAIT:2005007
16. Zdolec N, Filipović I, Fleck ZC, Marić A, Jankuloski D, Kozačinski L, et al. Susceptibilidad antimicrobiana de bacterias lácticas aisladas de embutidos fermentados y queso crudo. Veterinario Arh. (2011) 81:133–41.
17. Mater DDG, Langella P, Corthier G, Flores M-J. Una cepa probiótica de Lactobacillus puede adquirir resistencia a la vancomicina durante el tránsito digestivo en ratones. J Mol Microbiol Biotechnol. (2007) 14:123–7. doi: 10.1159/000106091
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Çataloluk O, Gogebakan B. Presencia de resistencia a los medicamentos en lactobacilos intestinales de origen lácteo y humano en Turquía. FEMS Microbiol Lett. (2004) 236:7–12. doi: 10.1111/j.1574-6968.2004.tb09620.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Ammor MS, Flórez AB, van Hoek AHAM, de los Reyes-Gavilán CG, Aarts HJM, Margolles A, et al. Caracterización molecular de la resistencia intrínseca y adquirida a antibióticos en bacterias lácticas y bifidobacterias. J Mol Microbiol Biotechnol. (2007) 14:6–15. doi: 10.1159/000106077
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Feld L, Bielak E, Hammer K, Wilcks A. Caracterización de un pequeño plásmido de resistencia a eritromicina pLFE1 a partir del aislado alimentario Lactobacillus plantarum M345. Plásmido. (2009) 61:159–70. doi: 10.1016/j.plasmid.2009.01.002
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Zhao M, Liu K, Zhang Y, Li Y, Zhou N, Li G. Características probióticas y análisis de la secuencia del genoma completo de Pediococcus acidilactici aislado de las heces de beagles adultos. Microbiol frontal. (2023) 14:1179953. doi: 10.3389/fmicb.2023.1179953
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Yang Q, Lü Y, Zhang M, Gong Y, Li Z, Tran NT, et al. Bacterias lácticas, Enterococcus faecalis Y17 y Pediococcus pentosaceus G11, mejora del rendimiento del crecimiento y de la inmunidad del cangrejo de fango (Scylla paramamosain). Pescados Mariscos Immunol. (2019) 93:135–43. doi: 10.1016/j.fsi.2019.07.050
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Bs S, Thankappan B, Mahendran R, Muthusamy G, Femil selta DR, Angayarkanni J. Evaluación de la producción de GABA y las actividades probióticas de Enterococcus faecium BS5. Probiot Antimicr Prot. (2021) 13:993–1004. doi: 10.1007/s12602-021-09759-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Rayos UV. Colección Cultura Tipo Española (2023). Disponible en: https://www.uv.es/uvweb/spanish-type-culture-collection/en/spanish-type-culture-collection-1285872233521.html (consultado el 1 de agosto de 2023).
25. NCIMB. Microbiología. Servicios analíticos. Almacenamiento de biomateriales (2023). Disponible en: https://www.ncimb.com/ (consultado el 1 de agosto de 2023).
26. Catálogo de la Colección Alemana de Microorganismos y Cultivos Celulares GmbH. (2023). Disponible en: https://www.dsmz.de/collection/catalogue/microorganisms/catalogue (consultado el 1 de agosto de 2023).
27. Instituto de Normas Clínicas y de Laboratorio. Métodos para la dilución de pruebas de susceptibilidad a los antimicrobianos para bacterias que crecen aeróbicamente. 11ª ed. Wayne, PA: Instituto de Estándares Clínicos y de Laboratorio (2018).
28. EUCAST. Tablas de puntos de interrupción para la interpretación de MIC y diámetros de zona. Comité Europeo de Pruebas de Susceptibilidad a los Antimicrobianos (2023).
29. Andrews S. FastQC: Una herramienta de control de calidad para datos de secuencia de alto rendimiento (2012). Disponible en: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (consultado el 25 de abril de 2022).
30. Krueger F, James F, Ewels P, Afyounian E, Schuster-Boeckler B. FelixKrueger/TrimGalore: v0.6.7—DOI a través de Zenodo. GitHub (2021).
31. Dinghua. MEGAHIT: una solución ultrarrápida de un solo nodo para el ensamblaje de metagenómica grande y compleja a través de un grafo sucinto de Bruijn. Bioinformática. (2022) 2022:BTV033. doi: 10.1093/bioinformática/BTV033
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Hyatt D, Chen G-L, Locascio PF, Land ML, Larimer FW, Hauser LJ. Pródigo: reconocimiento de genes procariotas e identificación del sitio de iniciación de la traducción. BMC Bioinformat. (2010) 11:119. doi: 10.1186/1471-2105-11-119
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Alcock BP, Raphenya AR, Lau TTY, Tsang KK, Bouchard M, Edalatmand A, et al. CARD 2020: vigilancia del resistoma antibiótico con la base de datos integral de resistencia a los antibióticos. Ácidos nucleicos Res. (2020) 48:D517-25. doi: 10.1093/nar/gkz935
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Johansson MHK, Bortolaia V, Tansirichaiya S, Aarestrup FM, Roberts AP, Petersen TN. Detección de elementos genéticos móviles asociados con la resistencia a los antibióticos en Salmonella enterica utilizando una herramienta web recientemente desarrollada: MobileElementFinder. J Quimiomadre antimicrobiana. (2021) 76:101–9. doi: 10.1093/jac/dkaa390
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Roux S, Enault F, Hurwitz BL, Sullivan MB. VirSorter: extracción de señales virales a partir de datos genómicos microbianos. ParesJ. (2015) 3:e985. doi: 10.7717/peerj.985
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Tauch A, Krieft S, Kalinowski J, Pühler A. El plásmido R pTP10 de 51.409 pb del aislado clínico multirresistente Corynebacterium striatum M82B está compuesto por segmentos de ADN identificados inicialmente en bacterias del suelo y en patógenos de plantas, animales y humanos. Mol Gen Genet. (2000) 263:1–11. doi: 10.1007/PL00008668
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Charpentier E, Gerbaud G, Courvalin P. Presencia del gen de resistencia a la tetraciclina de Listeria tet(S) en Enterococcus faecalis. Agentes antimicrobianos Quimiomadre. (1994) 38:2330–5. doi: 10.1128/AAC.38.10.2330
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Takeuchi K, Tomita H, Fujimoto S, Kudo M, Kuwano H, Ike Y. Resistencia a los medicamentos de los aislados clínicos de Enterococcus faecium y la transferencia conjugativa de rasgos de resistencia a la gentamicina y la eritromicina. FEMS Microbiol Lett. (2005) 243:347–54. doi: 10.1016/j.femsle.2004.12.022
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Xu J, Gallert C, Winter J. Múltiples resistencias a los antibióticos de los aislados de Enterococcus de aguas residuales crudas o filtradas con arena. Aplicación Microbiol Biotechnol. (2007) 74:493–500. doi: 10.1007/s00253-006-0668-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Maasjost J, Mühldorfer K, Jäckel SC de, Hafez HM. Patrones de susceptibilidad a los antimicrobianos de Enterococcus faecalis y Enterococcus faecium aislados de parvadas de aves de corral en Alemania. avdi. (2015) 59:143–8. doi: 10.1637/10928-090314-RegR
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Wenzler S, Schmidt-Eisenlohr E, Daschner F. Actividades comparativas in vitro de tres nuevas quinolonas y azitromicina contra patógenos aeróbicos causantes de infecciones del tracto respiratorio y heridas abdominales. Quimioterapia. (2004) 50:40–2. doi: 10.1159/000077284
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Xia M, Mu S, Fang Y, Zhang X, Yang G, Hou X, et al. Características genéticas y probióticas del Enterococcus faecium FUA027 productor de urolitina A. Alimentos. (2023) 12:1021. doi: 10.3390/alimentos12051021
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Costa Y, Galimand M, Leclercq R, Duval J, Courvalin P. Caracterización del gen cromosómico aac(6′)-Ii específico para Enterococcus faecium. Agentes antimicrobianos quimioterápicos. (1993) 37:1896–903. doi: 10.1128/AAC.37.9.1896
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Cerdá P, Goñi P, Millán L, Rubio C, Gómez-Lus R. Detección del grupo de genes de resistencia a aminoglucósidostreptotricina ant(6)-sat4-aph(3 ‘)-III en estreptococos del grupo viridans comensales. Int Microbiol. (2007) 10:57–60. doi: 10.2436/20.1501.01.8
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Trieu-Cuot P, Courvalin P. Secuencia de nucleótidos del gen del plásmido Streptococcus faecalis que codifica la fosfotransferasa 3′5′’-aminoglucósido fosfotransferasa tipo III. Gen. (1983) 23:331–41. doi: 10.1016/0378-1119(83)90022-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Pansegrau W, Miele L, Lurz R, Lanka E. Secuencia de nucleótidos del determinante de la resistencia a la kanamicina del plásmido RP4: homología con otros aminoglucósidos 3′-fosfotransferasas. Plásmido. (1987) 18:193–204. doi: 10.1016/0147-619X(87)90062-X
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Daly M, Villa L, Pezzella C, Fanning S, Carattoli A. Comparación de regiones génicas de resistencia a múltiples fármacos entre dos serotipos de Salmonella geográficamente no relacionados. J Quimiotera Antimicr. (2005) 55:558–61. doi: 10.1093/jac/dki015
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Perreten V, Schwarz FV, Teuber M, Levy SB. Mdt(A), una nueva proteína de eflujo que confiere resistencia múltiple a los antibióticos en Lactococcus lactis y Escherichia coli. Agentes antimicrobianos Quimiomadre. (2001) 45:1109–14. doi: 10.1128/AAC.45.4.1109-1114.2001
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Isnard C, Malbruny B, Leclercq R, Cattoir V. Bases genéticas para la resistencia in vitro e in vivo a Lincosamidas, Estreptograminas A y Pleuromutilinas (fenotipo LSAP) en Enterococcus faecium. Agentes antimicrobianos Quimiomadre. (2013) 57:4463–9. doi: 10.1128/AAC.01030-13
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Min Y-H, Kwon A-R, Yoon E-J, Shim M-J, Choi E-C. Atenuación traduccional y estabilización del ARNm como mecanismos de inducción de erm(B) por eritromicina. Agentes antimicrobianos Quimiomadre. (2008) 52:1782–9. doi: 10.1128/AAC.01376-07
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Singh KV, Malathum K, Murray BE. La alteración de un gen específico de la especie de Enterococcus faecium, un homólogo de los genes adquiridos de resistencia a macrólidos de los estafilococos, se asocia con un aumento en la susceptibilidad a los macrólidos. Agentes antimicrobianos Quimiomadre. (2001) 45:263–6. doi: 10.1128/AAC.45.1.263-266.2001
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Urshev Z, Yungareva T. Evaluación inicial de la seguridad de Enterococcus faecium LBBE81. Biotechnol Biotechnol Equip. (2021) 35:11–7. doi: 10.1080/13102818.2020.1840438
53. Thumu SCR, Halami PM. Presencia de genes de resistencia a eritromicina y tetraciclina en bacterias del ácido láctico procedentes de alimentos fermentados de origen indio. Antonie Van Leeuwenhoek. (2012) 102:541–51. doi: 10.1007/s10482-012-9749-4
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Mulcahy H, O’Callaghan J, O’Grady EP, Adams C, O’Gara F. El regulador postranscripcional RsmA desempeña un papel en la interacción entre Pseudomonas aeruginosa y las células epiteliales de las vías respiratorias humanas al regular positivamente el sistema de secreción de tipo III. Infectar a Immun. (2006) 74:3012–5. doi: 10.1128/IAI.74.5.3012-3015.2006
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Jacob J, Evers S, Bischoff K, Carlier C, Courvalin P. Caracterización del gen sat 4 que codifica una acetiltransferasa estreptotricina en Campylobacter coli BE/G4. FEMS Microbiol Lett. (1994) 120:13–7. doi: 10.1111/j.1574-6968.1994.tb07000.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Roberts MC. Actualización sobre los genes adquiridos de resistencia a la tetraciclina. FEMS Microbiol Lett. (2005) 245:195–203. doi: 10.1016/j.femsle.2005.02.034
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: probióticos, ARG, NGS, animales de compañía, resistencia a los antimicrobianos
Cita: Kerek A, Szabó E, Szabó Á, Papp M, Bányai K, Kardos G, Kaszab E, Bali K y Jerzsele Á (2024) Investigación de genes de resistencia a los antimicrobianos en productos probióticos para animales de compañía. Frente. Vet. Sci. 11:1464351. doi: 10.3389/fvets.2024.1464351
Recibido: 13 de julio de 2024; Aceptado: 07 de octubre de 2024;
Publicado: 22 de octubre de 2024.
Editado por:
Iram Liaqat, Universidad del Colegio Público, Lahore, Pakistán
Revisado por:
Riffat Iqbal, Colegio Público, Pakistán
Najma Arshad, Universidad del Punjab, Pakistán
Derechos de autor © 2024 Kerek, Szabó, Szabó, Papp, Bányai, Kardos, Kaszab, Bali y Jerzsele. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Adam Kerek, kerek.adam@univet.hu
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía