
Efectos de los prebióticos (lactoferrina) y diclazurilo en pollos infectados experimentalmente con Eimeria tenella

Efectos de los prebióticos (lactoferrina) y diclazurilo en pollos de engorde infectados experimentalmente con Eimeria tenella
 Asmaa G. Abd El Monsef1Nermin F. El Zohairy1
Asmaa G. Abd El Monsef1Nermin F. El Zohairy1 Marwa F. Hassan2Sanaa M. Salem3Asmaa Aboelabbas Gouda4Mogda K. Mansour2
Marwa F. Hassan2Sanaa M. Salem3Asmaa Aboelabbas Gouda4Mogda K. Mansour2 Abdulsalam A. M. Alkhaldi5Alzaylaee Hind6
Abdulsalam A. M. Alkhaldi5Alzaylaee Hind6 Ehab Kotb Elahallawy7,8*
Ehab Kotb Elahallawy7,8*- 1Departamento de Bioquímica, Toxicología y Deficiencia de Piensos, Instituto de Investigación de Salud Animal (AHRI), Centro de Investigación Agrícola (ARC), Sucursal de Zagazig, Zagazig, Egipto
- número arábigoDepartamento de Bioquímica, Toxicología y Carencia de Piensos, Instituto de Investigación de Sanidad Animal (AHRI), Centro de Investigación Agrícola (ARC), Dokki, Giza, Egipto
- 3Departamento de Patología, Instituto de Investigación de Salud Animal (AHRI), Centro de Investigación Agrícola (ARC), Sucursal de Zagazig, Zagazig, Egipto
- 4Departamento de Parasitología, Facultad de Medicina Veterinaria, Universidad de Zagazig, Zagazig, Egipto
- 5Departamento de Biología, Facultad de Ciencias, Universidad de Jouf, Sakaka, Arabia Saudita
- 6Departamento de Biología, Facultad de Ciencias, Universidad Princesa Nourah bint Abdulrahman, Riad, Arabia Saudita
- 7Departamento de Sanidad Animal, Grupo de Investigación en Sanidad Animal y Zoonosis (GISAZ), Universidad de Córdoba, Córdoba, España
- 8Departamento de Zoonosis, Facultad de Medicina Veterinaria, Universidad de Sohag, Sohag, Egipto
Introducción: La coccidiosis aviar representa un reto importante para la industria avícola en Egipto, lo que pone de relieve la necesidad urgente de validar nuevas dianas farmacológicas que ofrezcan perspectivas prometedoras para el desarrollo de anticoccidiales avanzados. Aunque numerosos informes destacan la actividad de la lactoferrina (LF) frente a diversos microorganismos, su potencial frente a Eimeria no ha sido explorado. El presente estudio evaluó el potencial efecto anticoccidial de LF y diclazurilo en pollos de engorde infectados experimentalmente con Eimeria tenella.
Métodos: Un total de 100 pollos de engorde de un día de edad se dividieron en cinco grupos iguales (20 cada uno) de la siguiente manera: el grupo 1 (G1) sirvió como grupo de control sano normal, el grupo 2 (G2) consistió en pollos infectados con 1 × 105 ooquistes esporulados de E. tenella a los 14 días de edad, el grupo 3 (G3) comprendió pollos infectados tratados con diclazurilo (0,5 mL/L en agua de bebida) durante 3 días sucesivamente, El grupo 4 (G4) incluyó pollos infectados tratados con LF (a una dosis de 250 mg/kg de dieta) desde el primer día de edad hasta el final del estudio, y el grupo 5 (G5) comprendió pollos infectados tratados con LF y diclazurilo.
Resultados: El grupo control positivo (G2) experimentó reducciones significativas en el peso corporal (PC), la ganancia de peso corporal, la glucosa sérica, la lipasa, la amilasa, la capacidad antioxidante total, varios índices hematológicos y proteínas totales, junto con alteraciones en varias enzimas antioxidantes. Por el contrario, los niveles séricos de aspartato aminotransferasa (AST), alanina aminotransferasa (ALT), fosfatasas alcalinas (ALP), urea, creatinina, óxido nítrico, volumen corpuscular medio (VCM), glóbulos blancos (GB), heterófilos, alfa 2, beta 1 y contenido hepático de malondialdehído estaban elevados en este grupo. Además, en G2 se observaron mayores recuentos de ooquistes y puntuaciones de lesiones, junto con alteraciones histopatológicas. Sorprendentemente, el tratamiento con diclazurilo y/o LF demostró potentes efectos antioxidantes y anticoccidiales, lo que resultó en una reducción del desprendimiento de ooquistes, puntuaciones de lesiones e infiltrados linfocitarios en el ciego. Además, estos tratamientos mejoraron los sistemas antioxidante e inmunológico en los pollos y restauraron todos los cambios histopatológicos reportados en el grupo infectado no tratado (G2).
Conclusión: Este estudio ofrece perspectivas novedosas sobre los posibles efectos anticoccidiales de la combinación de LF y diclazuril en pollos de engorde infectados con E. tenella, destacando las posibles acciones sinérgicas de LF en el tratamiento de la coccidiosis aviar.
1 Introducción
La coccidiosis aviar sigue siendo una de las principales enfermedades que influyen en la cría de aves de corral, causando una pérdida económica anual de más de 800 millones de dólares estadounidenses en todo el mundo (1). Esta enfermedad es causada por parásitos protozoarios intracelulares pertenecientes al género Eimeria (2). Se identificaron varias especies de Eimeria en aves de corral, a saber, Eimeria tenella, Eimeria mitis, Eimeria maxima, Eimeria acervulina, Eimeria brunetti, Eimeria necatrice y Eimeria praecox (3, 4). Sus efectos clínicos en las aves de corral van desde infecciones subclínicas hasta muertes de animales. Entre los otros, E. tenella y E. necatrix se clasifican como patógenos graves en aves de corral. La coccidiosis es la más común y difícil de manejar, particularmente en la cría de aves de corral, lo que provoca lesiones intestinales y reduce la conversión alimenticia, el ingreso de peso y la producción de huevos (5, 6). Considerada individualmente, cada especie de Eimeria puede afectar a una región específica del intestino; por ejemplo, E. tenella afecta principalmente al ciego, mientras que E. acervulina se dirige predominantemente al duodeno (7). Como se mencionó anteriormente, el parásito está ampliamente distribuido en el medio ambiente y se utilizan varias estrategias para su control, incluidos los agentes anticoccidiales. Sin embargo, los aislados resistentes a los medicamentos de Eimeria spp. emergen utilizando estos medicamentos (1). Entre otros, el diclazurilo (bencenoacetonitrilo) es un agente antiprotozoario a base de triazina, que contiene 1,2,4-triazina, y se ha utilizado durante mucho tiempo para controlar E. tenella (8). Este fármaco dificulta eficazmente las etapas sexual y asexual de los coccidios al impedir la excreción de ooquistes, lo que en última instancia detiene el ciclo de vida de estos parásitos (9). Además, se demostró que el diclazurilo es un agente anticoccidial análogo de nucleótidos altamente efectivo, lo que resultó en una reducción significativa de la eliminación media de ooquistes y destruyó todas las etapas de desarrollo intracelular de E. tenella (tanto esquizontes como gamontes) (10). El diclazurilo, ya sea solo o en comparación con otros agentes anticoccidiales en la proporción, tiene la actividad anticoccidial potencial (11). Sin embargo, ha habido un aumento preocupante en los informes de resistencia al diclazurilo y otros medicamentos anticoccidiales. Además, el uso excesivo de estos medicamentos puede dar lugar a residuos en los productos de origen animal, lo que supone un peligro potencial para la salud pública (8, 12). Por lo tanto, la búsqueda de nuevos agentes anticoccidiales eficaces sigue siendo una de las principales medidas para controlar la coccidiosis en las aves de corral. Varios informes anteriores exploraron los beneficios potenciales de los productos naturales y las plantas en el tratamiento de la coccidiosis (13, 14).
Curiosamente, los prebióticos y probióticos son cada vez más importantes en la prevención y el control de diversas enfermedades infecciosas, incluida la coccidiosis (15). Un estudio previo (16) ha señalado que diferentes suplementos probióticos dietéticos pueden afectar la inmunidad del huésped contra la coccidiosis. La lactoferrina (LF) es una glicoproteína de unión al hierro que tiene el potencial de mejorar el sistema de defensa preinmune del huésped. Se deriva de varias fuentes, incluidos los fluidos secretores como la leche materna o las lágrimas, las secreciones mucosas y los gránulos secundarios de neutrófilos y sangre (17). La LF tiene una estructura única y posee potentes actividades antimicrobianas e inmunomoduladoras relacionadas con la familia de la transferrina (18, 19). Además, se ha demostrado previamente que la FL mejora el rendimiento de las aves de corral (20). Los mecanismos celulares y moleculares responsables de los efectos inmunomoduladores de la FL no están completamente dilucidados. Sin embargo, los estudios in vitro e in vivo sugirieron la existencia de múltiples mecanismos, incluida la modulación de la producción de citoquinas/quimiocinas, la regulación de la producción de especies reactivas de oxígeno y el reclutamiento de células inmunitarias (21). Además de sus propiedades antioxidantes, la LF es un eliminador natural de hierro, activador/modulador de las vías de señalización y se dirige a la retroalimentación negativa de la respuesta inflamatoria (22-24). Además, la actividad antimicrobiana de la LF parece estar relacionada con la privación de hierro a través de la eliminación de un sustrato esencial que se requiere para el crecimiento bacteriano (25), mientras que su efecto bacteriostático se lleva a cabo principalmente mediante la degradación de los peptidoglicanos en la pared celular bacteriana afectando la permeabilidad de la membrana, lo que resulta en lisis celular (26). Si bien numerosos estudios han destacado las propiedades protectoras de LF como agente antibacteriano y antifúngico, la investigación limitada ha investigado su actividad potencial contra los parásitos, especialmente los coccidios.
Al revisar la literatura existente, es evidente que solo un estudio previo ha explorado la influencia de la LF bovina en Eimeria stiedae in vivo (ratones y conejos) e in vitro (células embrionarias de ratón y hepatobiliares de conejo) (27). Este estudio tuvo como objetivo evaluar los posibles efectos antiparasitarios e inmunomoduladores de la suplementación dietética con LF, ya sea sola o en combinación con el fármaco anticoccidial ampliamente utilizado diclazuril, en pollos de engorde infectados con Eimeria spp. La metodología consistió en evaluar signos clínicos, parámetros parasitológicos, marcadores de estrés oxidativo, cambios hematológicos, alteraciones en la expresión génica inmunológica y hallazgos histopatológicos.
2 Materiales y métodos
2.1 Consentimiento ético
El protocolo experimental fue aprobado por el Instituto de Investigación en Sanidad Animal (AHRI) de conformidad con el comité de la ARC y la ICUC (ARC, AHRI, IACUC, 85/23).
2.2 Drogas
El diclazurilo (Diclosol 1%), en una formulación soluble en agua, fue suministrado por la empresa Pharma Sweed-Egypt, ubicada en el distrito 10 de la ciudad de Ramadán, B3, Nº 2353/2004 y Código Nº 205.87.20.v3. El prebiótico LF se obtuvo de la empresa Dulex Lab Nutrition and Pharmaceutical, situada en New Cairo, lote n.º: B2117051.
2.3 Cepas de parásitos y preparación de parásitos
En este paso, se obtuvo ooquiste esporulado de E. tenella de la Unidad de Parasitología del Departamento de Enfermedades Avícolas e Investigación del Instituto de Investigación en Sanidad Animal (Dokki, Egipto). En este paso, se aislaron ooquistes de E. tenella del ciego de pollitos infectados naturalmente siguiendo los protocolos descritos en otros lugares (28, 29). Los ooquistes aislados se dejaron esporular a temperatura estándar en una solución de dicromato de potasio al 2,5%. Posteriormente, se empleó el método de flotación de azúcar concentrado para eliminar los restos de los ooquistes esporulados de Eimeria, seguido de tres enjuagues con agua destilada. A continuación, se utilizó la técnica Mc-Master para ajustar el recuento a 1 × 105 ooquistes esporulados por 1 ml (28, 29).
2.4 Aves y diseño experimental
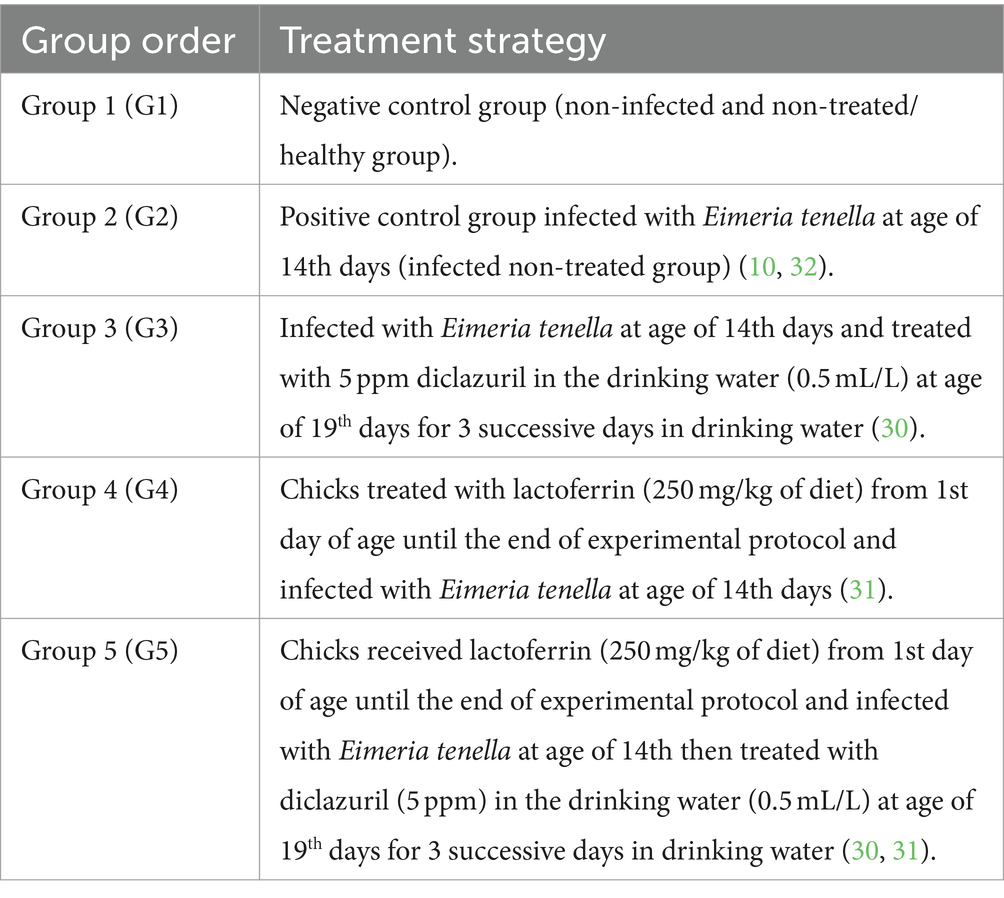
Se adquirieron cien pollos de engorde Cobb sanos de un día de edad (sin sexo) de una planta de incubación ubicada en El-Kahera Poultry Company, 10th de la ciudad de Ramadán, Egipto. A estos pollitos se les suministró una dieta comercial balanceada obtenida de la empresa Feed Mix hasta que cumplieron 7 días de edad. Posteriormente, fueron transferidos a jaulas con piso de alambre durante la duración del experimento, con estricto cumplimiento de las normas higiénicas. Los pollos tenían acceso sin restricciones a una dieta libre de antibióticos y anticoccidiales, junto con acceso ad libitum al agua del grifo. Las condiciones óptimas de temperatura se mantuvieron mediante radiadores eléctricos y ventiladores. Las muestras fecales se examinaron dos veces al día durante los primeros 14 días del experimento para confirmar la ausencia de ooquistes de Eimeria utilizando la técnica de flotación con solución saturada de NaCl (gravedad específica de 1,28). Además, todas las aves fueron vacunadas contra el virus de la enfermedad de Newcastle (VEN) utilizando las vacunas HitchnerB1 y LaSota a los 7 y 18 días de edad, respectivamente. También recibieron una vacuna contra la bursitis infecciosa a los 15 días de edad. Las vacunas contra el VEN se adquirieron a la empresa Intervet Boxmeer en Holanda, mientras que la vacuna Gumboro se obtuvo a la empresa Rhone-Merieau en Francia. De acuerdo con el protocolo experimental (30, 31) descrito en la Tabla 1, los pollitos se dividieron aleatoriamente en cinco grupos, cada uno compuesto por 20 individuos, y luego se dividieron en tres repeticiones biológicas, cada una de las cuales contenía de 6 a 7 pollos (con 3 jaulas por grupo). De estos, 80 animales estaban infectados, mientras que los 20 restantes no estaban infectados. Al día 14 de edad, a los pollitos de todos los grupos (G2-G5), excluyendo G1, se les administró 1 × 10 ooquistes esporulados de E. tenella 5/mL por pollito en 1 mL PBS utilizando una jeringa de insulina insertada directamente en el buche de cada ave.signos diariamente (10, 30, 32).
2.5 Evaluación de los signos clínicos y de la tasa de mortalidad
Los signos clínicos y el número de pollitos muertos en los grupos infectados y tratados se registraron diariamente durante todo el período experimental. La tasa de mortalidad se calcula como un porcentaje de acuerdo con la fórmula.
Tasa de mortalidad(%)=(número de pollitos muertos enunespecificadoperiodo×100)/Número total detodopollito
2.6 Evaluación de los resultados del crecimiento
Este paso implicó evaluar el peso corporal semanal (PC) y el aumento de peso corporal (BWG), que se registraron a lo largo del período de estudio de 22 días.
2.7 Muestreo
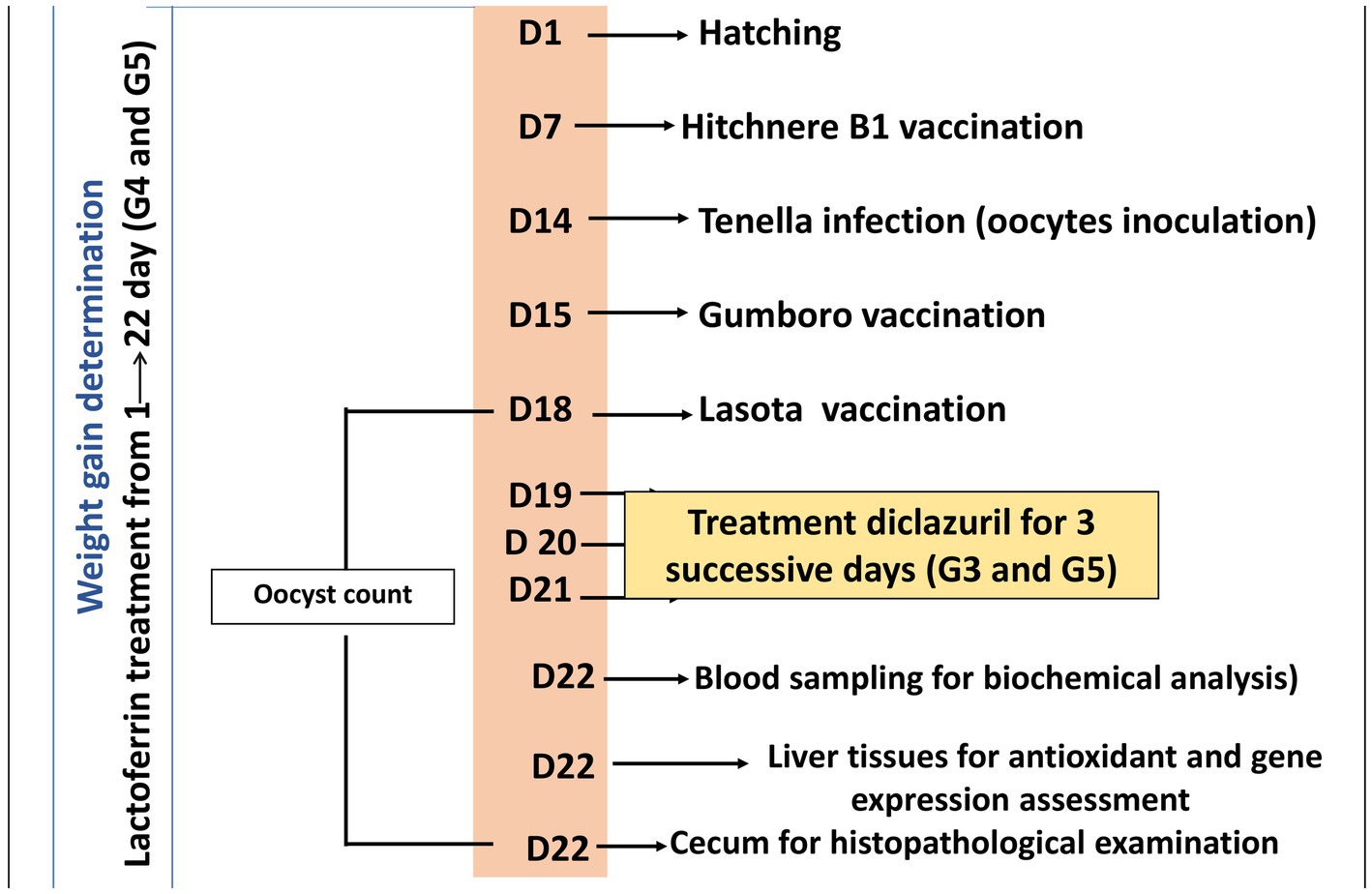
En el día 22 del experimento, los polluelos fueron sacrificados por dislocación del cuello y se recolectaron muestras de sangre, hígado y tejido de ciego. Se recogieron cinco muestras de sangre por vena alar de cada grupo. Cada muestra de sangre se dividió en dos tubos separados. El primer tubo contenía EDTA como anticoagulante para el hemograma completo, mientras que el segundo tubo no contenía ningún anticoagulante y se utilizó para la obtención de suero. Las muestras de suero se obtuvieron centrifugando las muestras de sangre a 3000 rpm durante 5 min. Los sueros se aislaron y almacenaron a -20 °C para su análisis bioquímico. En cuanto a las muestras de tejido, el tejido hepático se extirpó y se dividió en dos secciones: una para la evaluación antioxidante y otra para el análisis de la expresión génica. Además, se extirpó tejido de ciego y se preparó exclusivamente para el examen histopatológico. Los tratamientos experimentales y el cronograma de muestreo se dilucidan en la Figura 1.
2.8 Evaluación de la eficacia anticoccidial de prebióticos (lactoferrina) y diclazurilo
Este paso implicó la evaluación de la excreción de ooquistes fecales. La muestra fecal fresca de cada grupo experimental se recolectó diariamente en una bolsa de plástico limpia para su examen parasitológico. El recuento de ooquistes de Eimeria comenzó desde el cuarto día después de la infección hasta el final del experimento. Los ooquistes se contaron en 1 g de heces mediante el método de la cámara de McMaster (14, 33).
2.9 Examen histopatológico
Las muestras recolectadas de ciego se fijaron en formalina tamponada neutra al 10% durante 24 h, se deshidrataron en etanol graduado, se aclararon en xileno, se incluyeron en parafina y se seccionaron en secciones de tejido de 5 μm de espesor, luego se tiñeron con hematoxilina y eosina (H&E) y se examinaron microscópicamente para detectar cualquier alteración histopatológica (34). Todas las fotografías de la sección se tomaron con un microscopio Swift asociado a una cámara digital Swift. Las lesiones histopatológicas fueron puntuadas y estimadas por métodos semicuantitativos de la siguiente manera: «- = ausencia de lesión, + = 5-25%, ++ = 26-50% y +++ = > 50%» (35).
2.10 Análisis bioquímico
Se evaluó la cuantificación manual de eritrocitos (RBCs 106/mm3) y recuento de glóbulos blancos (WBCs) 103/mm3 según el método propuesto por Feldman et al. (36). También se estimó la concentración de hemoglobina (Hb) (g/dL) y el porcentaje de volumen de células empaquetadas (% de PCV) (37, 38). Se prepararon dos frotis de sangre inmediatamente después de recolectar cada muestra de sangre. Para el recuento diferencial de leucocitos se utilizaron frotis sanguíneos teñidos con tinción de Giemsa según Anderson y Latimer (39). Se estimaron las actividades de la alanina aminotransferasa (ALT) y la aspartato aminotransferasa (AST) según Reitman y Frankel (40). Además, la fosfatasa alcalina se ensayó mediante el método descrito por El-Aaser y El-Merzabani (41). El nivel de urea se midió siguiendo a Wybenga et al. (42), el nivel de creatinina según Henry (43) y el nivel de glucosa según Tietz (44). Además, se estimaron los niveles séricos de lipasa y amilasa según López (45), mientras que los niveles de óxido nítrico (NO) y capacidad antioxidante total (TAC) según Montgomery y Dymock y Koracevic et al. (46, 47), respectivamente. Se estimó el contenido hepático de actividades malonaldehído (MDA), superóxido dismutasa (SOD) y catalasa (CAT), y niveles reducidos de glutatión (GSH) de acuerdo con Okhawa et al. (48), Nishikimi et al. (49), Aebi (50) y Beutler et al. (51), respectivamente. Por su parte, la proteína total sérica y su patrón electroforético se estimaron de acuerdo con Sonnenwirth et al. (52) y Davis (53), respectivamente, y luego se calcularon de acuerdo con SynGene S. No. 17292*14518 sme*mpcs.
2.11 Análisis de parámetros antioxidantes
Los niveles hepáticos de malondialdehído (MDA), superóxido dismutasa (SOD), catalasa (CAT) y glutatión (GSH) se evaluaron mediante un método colorimétrico con el kit de biodiagnóstico (números de catálogo: MD 25 29, SD 25 21, CA 25 17 y GR 25 11, respectivamente). Se siguieron los protocolos para estas mediciones tal y como se describen en la literatura (48-51). Los tejidos hepáticos se extirparon, pesaron y cortaron en trozos pequeños, homogeneizados con un homogeneizador de vidrio en 9 volúmenes de tampón de fosfato de potasio 0,05 mM helado (pH 7,4) para preparar homogeneizados al 10%. Los tejidos homogeneizados se centrifugaron a 3000 rpm/15 min a 4°C y luego se utilizó el sobrenadante resultante para la determinación de los siguientes parámetros: SOD, CAT y MDA. Se picaron aproximadamente 0,1 g de tejidos hepáticos en pequeños trozos homogeneizados con un homogeneizador de vidrio en 0,2 mL de ácido metafosfórico (MPA) al 25% (ref. No.: 253-433-4, Sigma-Aldrich, Alemania), luego se agregaron 0,7 mL de agua destilada, se mezclaron e incubaron durante 1 h y se centrifugaron durante 10 min a 3,000 rpm, luego se retiró el sobrenadante limpio y se utilizó para la determinación de la concentración de GSH.
2.12 Transcripción de genes diana (IL-2 e IFN-γ) mediante qRT-PCR
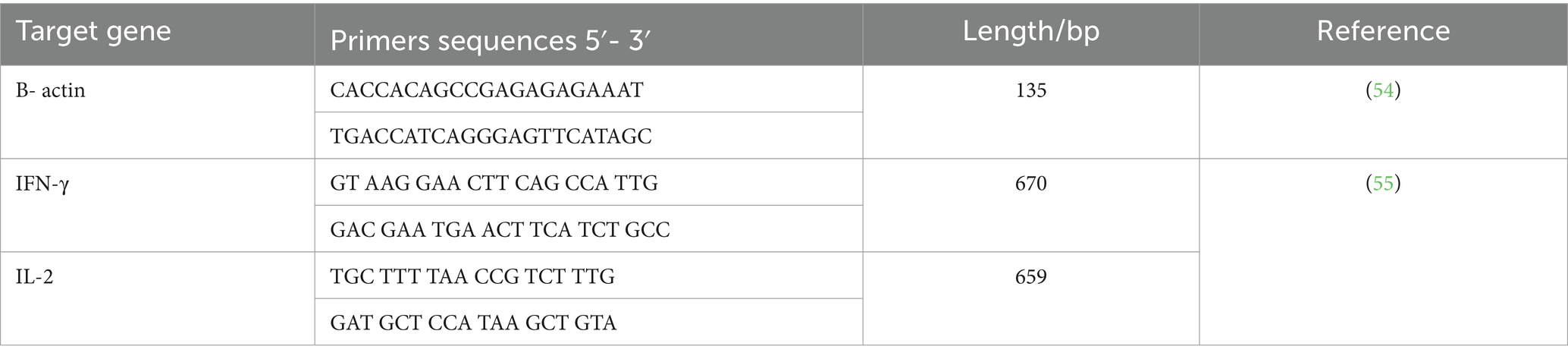
Este paso implicó la preparación de tejidos hepáticos de grupos experimentales que fueron recolectados, colocados en los tubos de Eppendorf, mantenidos inmediatamente en nitrógeno líquido y almacenados a -80 °C hasta la extracción del ARN para la determinación de la expresión génica de interleucina-2 (IL-2) e interferón-gamma (IFN-γ). A continuación, se estimó la expresión de ARNm de IL-2 e IFN-γ en el tejido hepático mediante PCR cuantitativa en tiempo real (qRT-PCR) que se realizó en la Unidad de Biotecnología del Instituto de Investigación en Sanidad Animal, sucursal de Zagazig (Egipto). La extracción de ARN de muestras de tejido hepático se realizó utilizando el kit QIAamp RNeasy Mini (Qiagen, Alemania, GmbH). Los cebadores utilizados se obtuvieron de Metabion (Alemania) y se muestran en la Tabla 2. El análisis de los resultados de la rt-PCR verde SYBR se determinó a través de las curvas de amplificación y los valores de los ciclos umbral (Ct), que se estimaron utilizando el software step one. Para evaluar el cambio de la expresión génica en el ARN de las muestras, se comparó la CT de cada muestra con la del grupo control en función del método «ΔΔCt» explicado por Yuan (56) en función de la siguiente relación: (2-ΔΔct).
2.13 Análisis estadístico
Los datos se analizaron estadísticamente mediante la prueba de análisis de varianza de una vía (ANOVA). Los resultados se dieron como media ± error estándar utilizando el programa SPSS 14.0 (2006) seguido de la prueba de Duncan. La significación estadística se estableció en p < 0,05.
3 Resultados
3.1 Efecto anticoccidial directo de la LF
3.1.1 Signos clínicos, puntuación de la lesión y tasa de mortalidad
En el presente trabajo, los pollos de G1 sirvieron como grupo de control normal negativo sano (no infectado) y, por lo tanto, no mostraron signos durante el período experimental. Mientras tanto, los pollos infectados con E. tenella y no tratados (G2; grupo control positivo) mostraron pérdida de apetito, embotamiento, depresión, pérdida de peso, plumas erizadas, deshidratación y diarrea sanguinolenta que apareció al quinto día después de la infección. En cuanto a la tasa de mortalidad registrada, el grupo infectado no tratado (G2) presentó una notable tasa de mortalidad del 30%, mientras que ninguno de los otros grupos reportó ninguna muerte. Por el contrario, los pollos infectados tratados con LF o diclazurilo (G3-G5) mostraron signos menos graves y los signos clínicos mejoraron gradualmente a partir del quinto día después de la infección y no mostraron mortalidad en todos estos grupos tratados.
3.1.2 Evaluación de los resultados del crecimiento
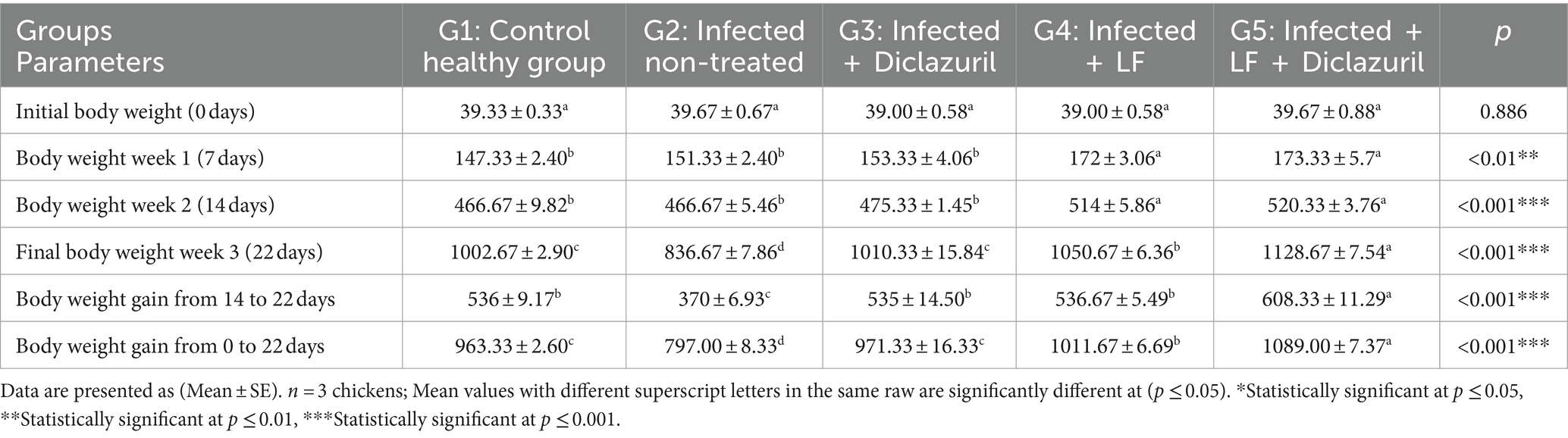
Los resultados obtenidos para la evaluación del rendimiento del crecimiento se muestran en la Tabla 3, revelando una disminución significativa (p < 0,001) en el peso corporal y el peso corporal (14 a 22 días o 0 a 22 días) en los pollos infectados no tratados (G2) en comparación con los pollos control normales (G1). Por el contrario, los pollos que recibieron diclazurilo y/o LF (G3, G4 y G5) mostraron un aumento significativo (p < 0,001) en el peso corporal y el peso corporal en comparación con los pollos infectados no tratados (G2). Cabe destacar que la suplementación con LF condujo a un aumento del peso corporal y del peso corporal a lo largo de toda la duración del estudio, como se observó tanto en los grupos G4 como en los G5.
3.1.3 Recuento de ooquistes
Los resultados de la Tabla 4 mostraron que los primeros ooquistes de E. tenella aparecieron en las caras al 4º día después de la infección, alcanzaron su punto máximo a los 4-5º días después de la infección (108 h después de la infección) y luego disminuyeron de nuevo gradualmente hasta el 8º día después de la infección. El uso de diclazurilo solo durante 3 días consecutivos inmediatamente después de la aparición de sangre en la gota (quinto día después de la infección) en pollos de engorde infectados experimentalmente mostró una disminución significativa en el número de ooquistes excretados en comparación con el grupo no tratado con E. tenella. Además, el uso de diclazurilo y/o LF tuvo un papel eficaz en la disminución del número de ooquistes. Se registró una reducción muy significativa en el número de excreción de ooquistes utilizando una combinación de LF y diclazurilo (Tabla 4).
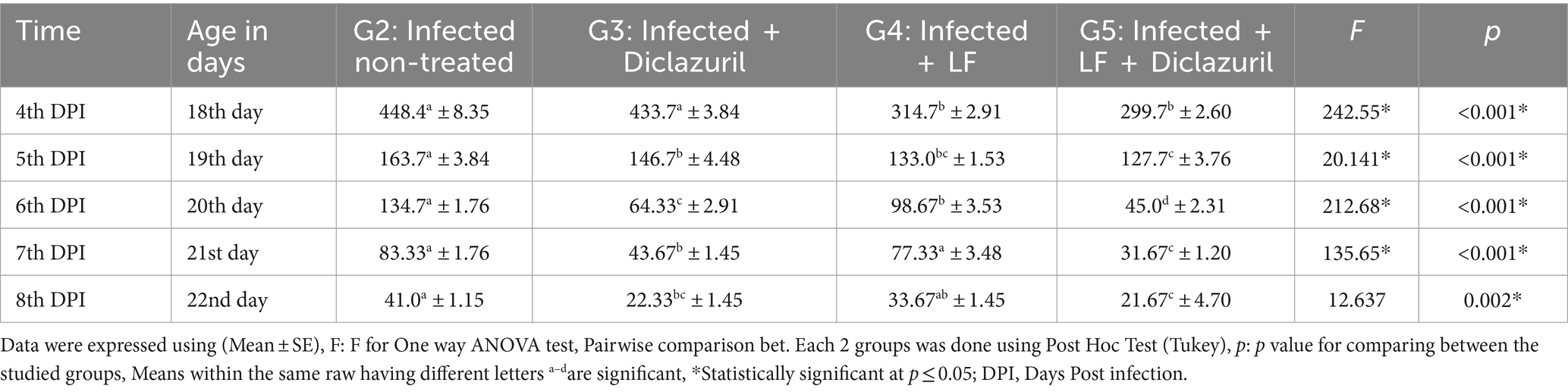
3.1.4 Histopatología
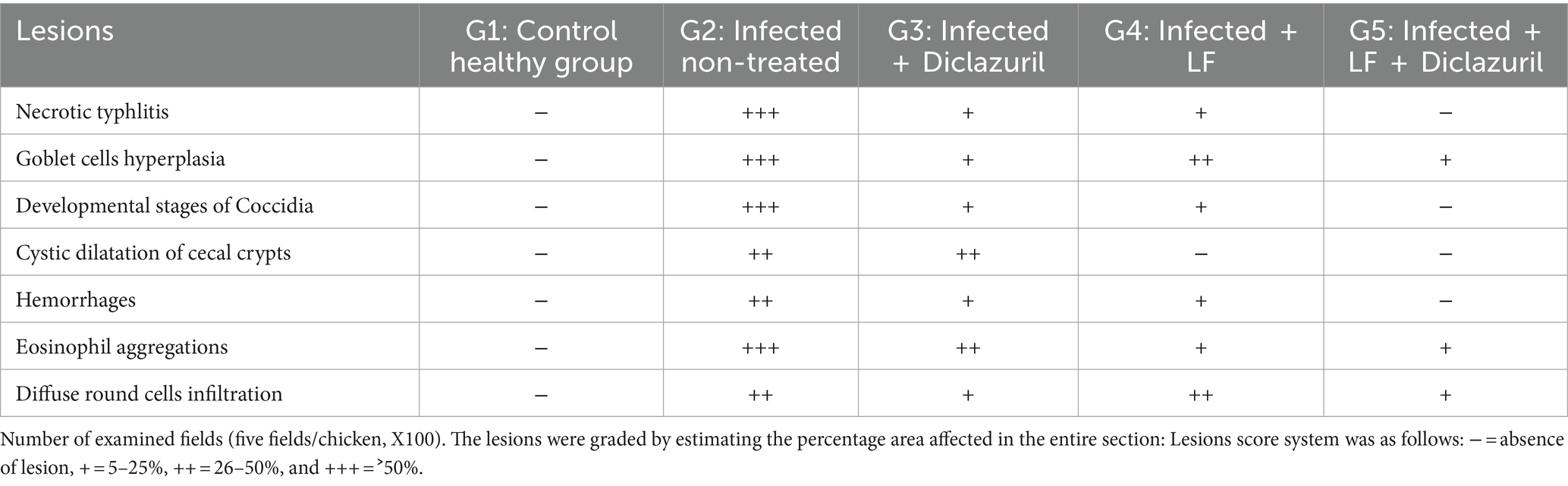
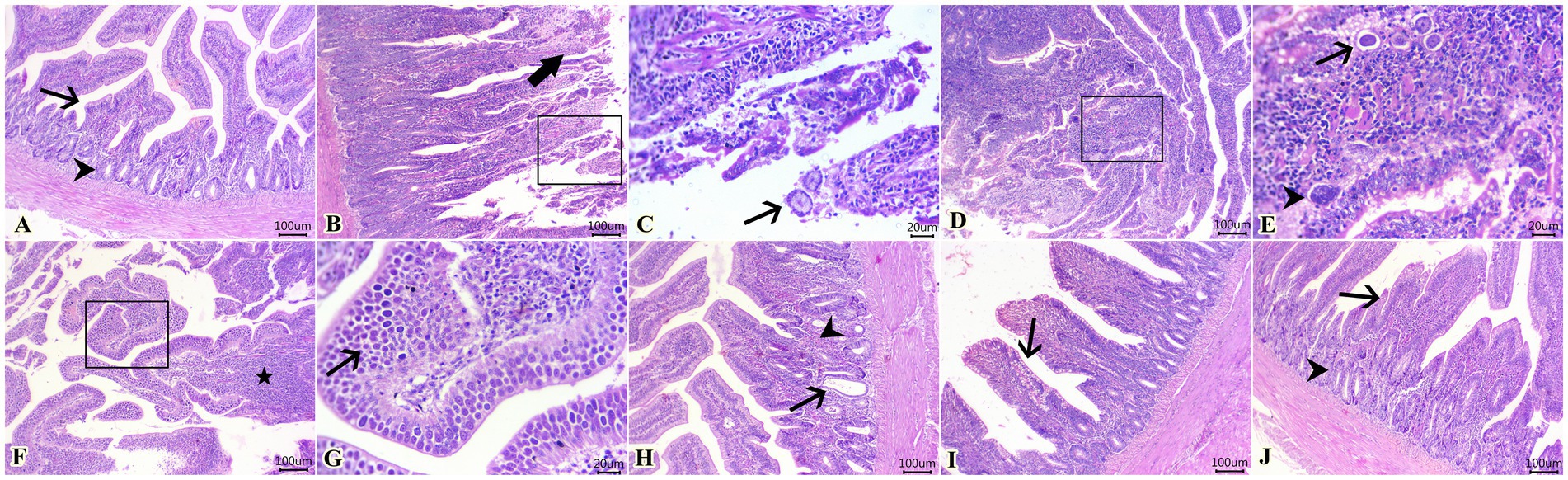
La puntuación de las lesiones en función de la extensión de la gravedad en los tejidos cecales entre los diferentes grupos experimentales se muestra en la Tabla 5. Como se ilustra, la infección experimental por E. tenella en el grupo control positivo (G2) indujo lesiones graves que recibieron la puntuación más alta en comparación con el grupo sano normal (G1). Por el contrario, como se muestra en la Tabla 5, el diclazuril y la FL causaron una mejoría significativa en la lesión histopatológica en comparación con el grupo infectado no tratado (G2). En cuanto a los hallazgos histopatológicos, G1 mostró arquitecturas histológicas normales de mucosa cecal, criptas, submucosa y capa muscular (Figura 2A). Por el contrario, G2 (Figuras 2B-G) mostró tiflitis necrótica representada por enterocitos necróticos, láminas descamadas de epitelio, infiltrados linfocíticos intensos dentro de lamia propia y capas submucosas, además de eritrocitos extravasados. También se encontró la presencia de varios estadios de desarrollo de coccidios dentro de enterocitos y láminas propias (macrogametos, microgametos y ooquistes) (Figuras 2B, C). En este grupo, los macrogametos se caracterizaron por gránulos eosinófilos periféricos y el microgameto contenía numerosos núcleos basófilos (Figuras 2D, E). Del mismo modo, algunas mucosas de revestimiento epitelial estaban fuertemente infestadas por un gran número de macrogametos con intensas infiltraciones de lámina propia con células redondas (Figuras 2F, G). Por el contrario, el grupo infectado tratado con diclazurilo (G3) mostró dilatación quística de algunas criptas cecales junto con hemorragia leve, agregación de eosinófilos e infiltración difusa de células mononucleares dentro de la lámina propia (Figura 2H). Además, los animales infectados tratados con el grupo tratado con LF (G4) exhibieron arquitecturas cecales normales con hiperplasia de células caliciformes, infiltración difusa de células mononucleares y hemorragias leves en la lámina propia (Figura 2I). Curiosamente, el último grupo (G5) tratado con una combinación de diclazurilo y LF G5 mostró una restauración de las estructuras histológicas normales del epitelio cecal y las criptas con una infiltración muy leve de la lámina propia y la submucosa con células redondas (Figura 2J).
3.2 Efecto protector indirecto de la LF
3.2.1 Evaluación del perfil sanguíneo y de los parámetros bioquímicos y antioxidantes en el suero
Los datos registrados en la Tabla 6 ilustraron que un aumento significativo (p < 0,001) en MCV, glóbulos blancos y % de heterófilos se asoció con un aumento no significativo en el % de basófilos en G2 en comparación con G1. En marcado contraste, todos los grupos tratados (G3-G5) mostraron una disminución significativa en todos los parámetros anteriores en comparación con los pollos infectados no tratados (G2). Por el contrario, se observó una disminución significativa (p < 0,05) de los glóbulos rojos, Hb, PCV, PLT, MCH, MCHC, linfocitos y monocitos, además de una disminución no significativa en el porcentaje de eosinófilos en el grupo no tratado con E. tenella (G2) en comparación con el grupo control sano. Todos los grupos tratados (G3-G5) mostraron un aumento significativo en todos los parámetros anteriores en comparación con G2 con pollos infectados no tratados (G2).
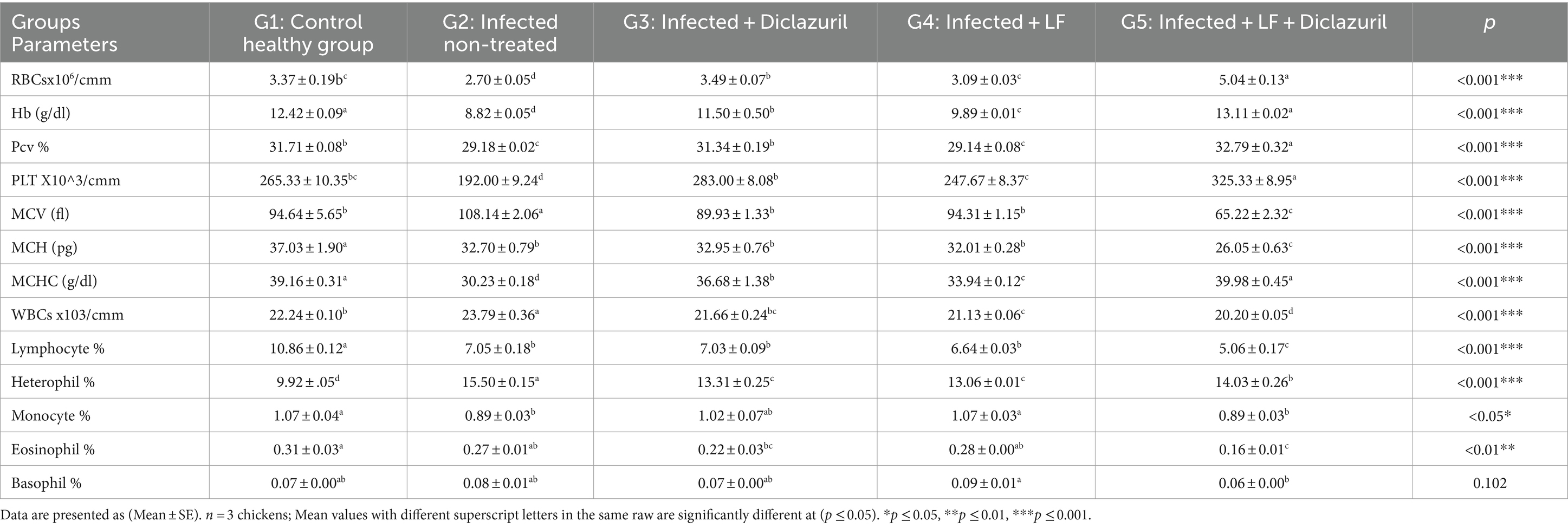
Tabla 6. Efecto del diclazuril y/o lactoferrina sobre el perfil sanguíneo durante la infección por E. tenella en pollos de engorde.
Como se muestra en la Tabla 7, el grupo infectado no tratado (G2) experimentó una disminución significativa (p < 0,05) de la proteína sérica total; albúmina, alfa 1, alfa total, beta 2, beta total, gamma 1, gamma 2, gamma total y globulina total y una disminución no significativa de alfa 2 y beta 1 en comparación con el grupo normal (G1). Por su parte, el tratamiento de los pollos infectados con diclazurilo y/o LF (G3, G4 y G5) mostró un aumento significativo (p < 0,05) en todos estos parámetros en comparación con G2. Se asocia con un aumento no significativo de la relación A:G en todos los grupos experimentales en comparación con el grupo normal.
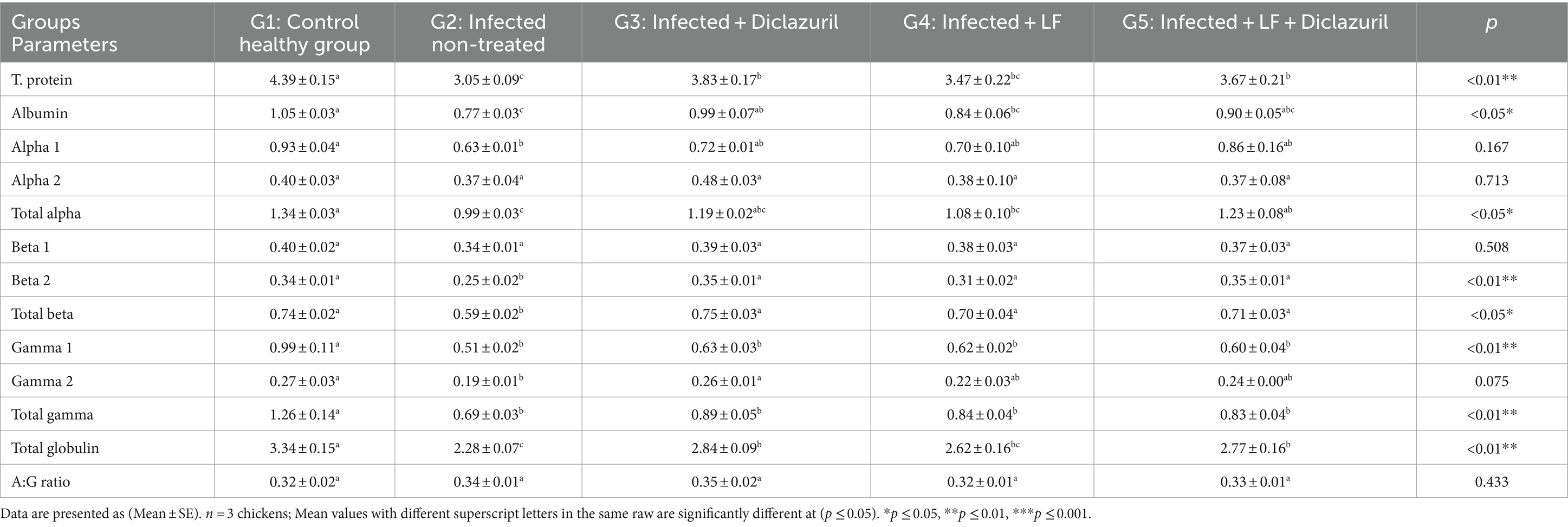
Además, los datos establecidos en la Tabla 8 revelaron que los niveles séricos de AST, ALT, ALP, urea, creatinina, NO y MDA en el hígado aumentaron significativamente (p < 0,001) en los pollos no tratados con E. tenella en comparación con el grupo normal. Mientras tanto, todos los grupos tratados mostraron una disminución significativa (p < 0,001) en todos los parámetros previos en comparación con los pollos no tratados con E. tenella. Por el contrario, se observó una disminución significativa (p < 0,01) de la glucosa sérica; lipasa (p < 0,01); amilasa; TAC; y se observaron niveles de SOD, CAT y GSH en el tejido hepático en el grupo no tratado con E. tenella en comparación con el grupo de control normal. Todos los grupos tratados mostraron un aumento significativo en todos los parámetros previos en comparación con los pollos no tratados con E. tenella con los mejores resultados en G5.
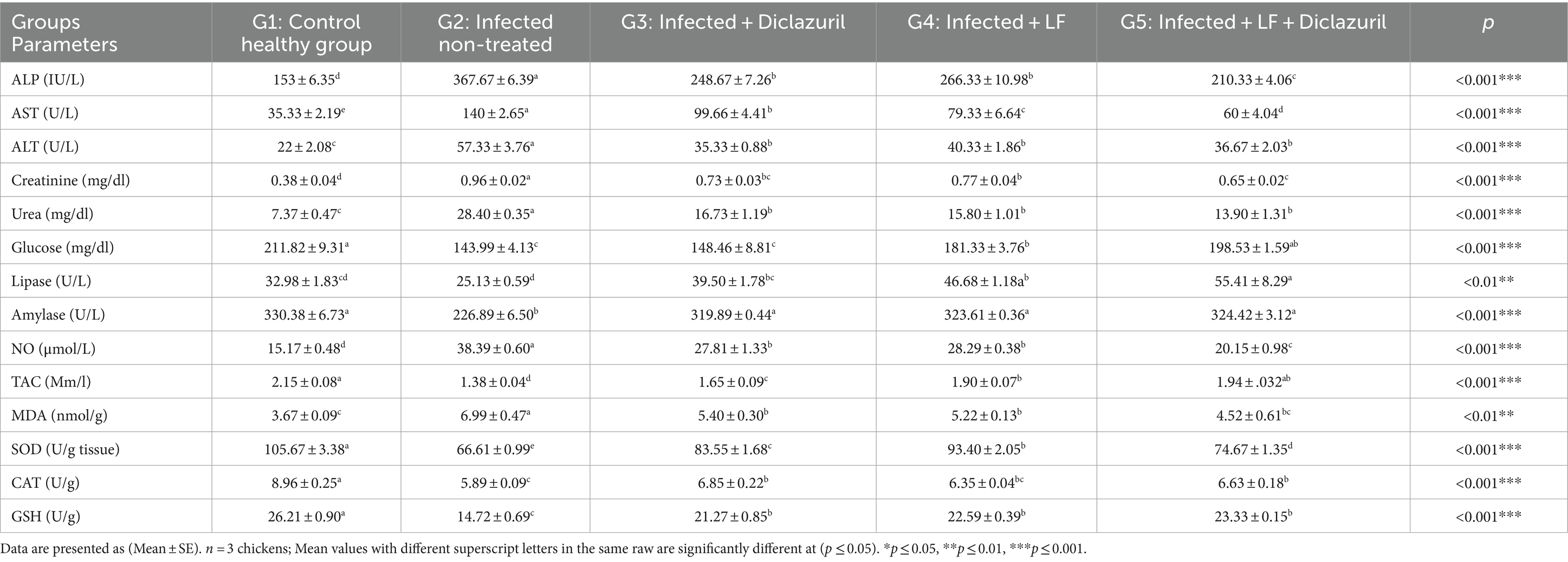
3.2.2 Evaluación de parámetros inflamatorios
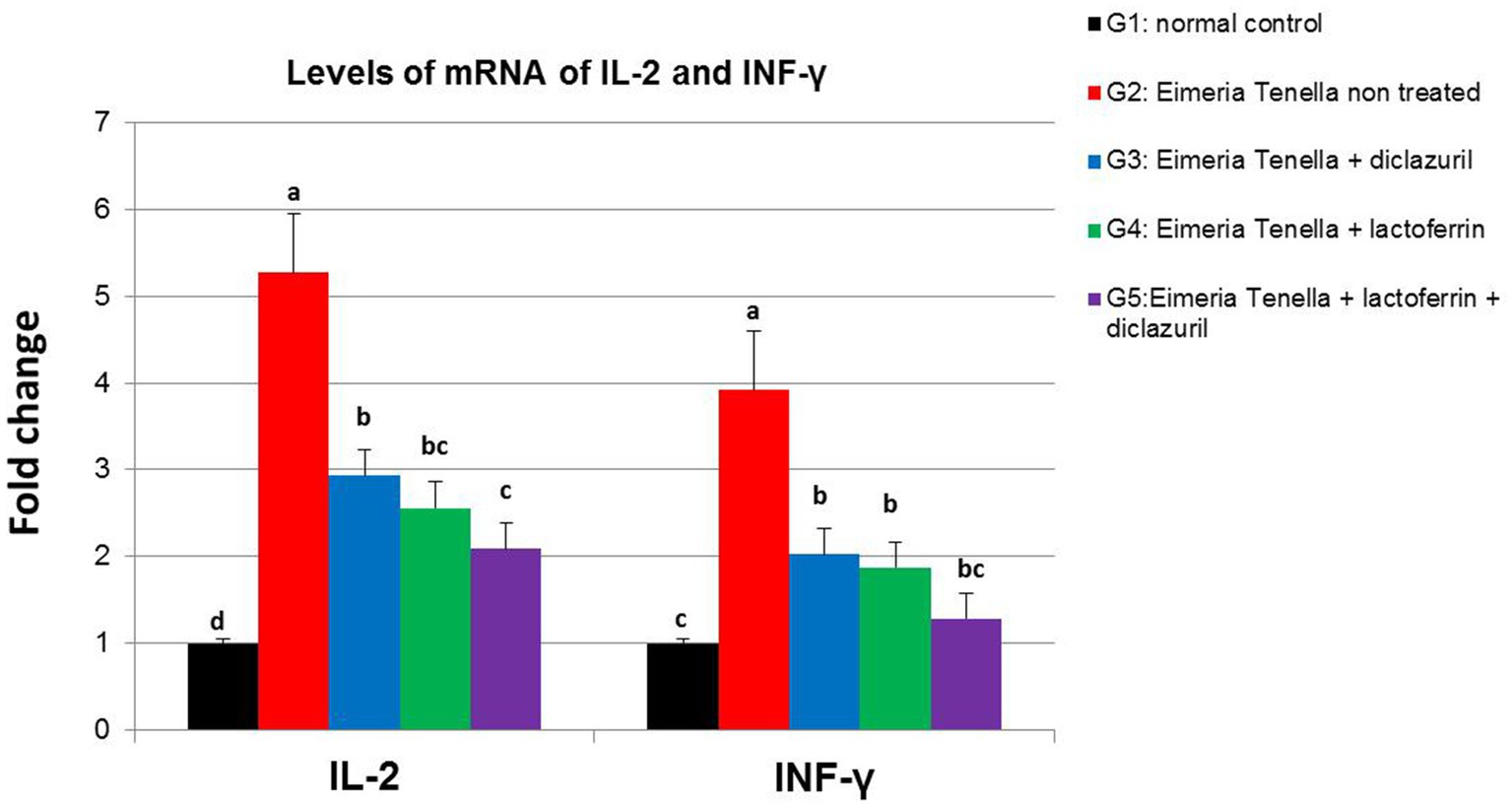
La figura 3 ilustra la alta regulación positiva (p < 0,001) de la expresión génica hepática de IL-2 e INF-γ en el grupo no tratado con E. tenella (G2) en comparación con el grupo sano (G1). Por su parte, los grupos tratados con diclazurilo y/o LF (G3-G5) mostraron una alta (p < 0,001) de regulación negativa de IL-2 e INF-γ en comparación con el grupo no tratado con E. tenella con efecto aditivo de ambos tratamientos LF y diclazurilo en G5.
4 Discusión
La coccidiosis aviar sigue siendo una de las principales enfermedades parasitarias que tienen un impacto económico negativo a nivel mundial en la industria avícola (57, 58). El control de la coccidiosis se basa principalmente en la aplicación de coccidiostáticos profilácticos en el alimento (59). Sin embargo, el desarrollo de resistencia anticoccidial a los aditivos químicos anticoccidiales para piensos y los posibles efectos nocivos sobre los peligros para la salud humana plantean una necesidad urgente de determinar las alternativas seguras para controlar la coccidiosis aviar. El estudio actual revela la eficacia potencial de LF y diclazuril en el tratamiento de la infección por E. tenella en pollos de engorde. Esto se establece a través de la evaluación integral de los hallazgos parasitológicos, bioquímicos, inmunológicos e histopatológicos, junto con evaluaciones del rendimiento del crecimiento, los signos clínicos, la puntuación de las lesiones y la tasa de mortalidad. Hasta donde sabemos, este es el primer informe que investiga los beneficios potenciales del uso de ambos materiales para tratar la coccidiosis.
En el presente estudio, el pico de excreción de ooquistes se produjo entre el 4º y el 5º día después de la infección (aproximadamente 108 h después de la infección). Este hallazgo se alinea estrechamente con Ahad et al. (60), quienes registraron el mayor recuento de ooquistes en el 4º día después de la infección. Sin embargo, otros estudios han documentado variaciones en el día pico de muda. En este sentido, Choi et al. (61) registraron el pico entre el 5º y el 6º o el 6º o 7º día después de la infección, dependiendo de la dosis de infección. Además, Gong et al. (62) observaron el pico en el 6º día post-infección, mientras que El-shazly et al. (33) lo observaron en el 9º día post-infección. Es probable que estas discrepancias en los tiempos máximos de excreción de ooquistes se deban a las variaciones en las dosis de inoculación de E. tenella (61), ya que las dosis más altas pueden acortar las generaciones asexuales y asexuales del parásito (63). En consecuencia, se han observado picos más tempranos en la excreción de ooquistes con las dosis de inoculación más altas (61). También es importante considerar que la patogenicidad de la coccidiosis está influenciada por una combinación de factores del huésped y del parásito (64). Estos factores abarcan la patogenicidad inherente de las especies de Eimeria, la capacidad reproductiva de las especies infectantes, la edad y cantidad de ooquistes ingeridos y la susceptibilidad de los pollos. Cada uno de estos elementos, ya sea individualmente o en combinación, puede influir en el momento de la excreción máxima de ooquistes (65-67).
mientras que, se produjo una disminución significativa en el número de ooquistes en el grupo tratado con LF (G4) debido a que la LF es una proteína multifuncional natural que inhibe el crecimiento de parásitos a través de la quelación del hierro y la capacidad de sus péptidos (Lactoferricinas) para atravesar la membrana celular protozoaria y la envoltura nuclear, por lo que los esporozoítos tratados con 1.000 μg/mL de bLfcin mostraron menos infectividad y menor penetración en las células huésped que los esporozoítos no tratados, como lo mencionan Omata et al. (27) y Reyes-López et al. (68). Curiosamente, la LF es una proteína inofensiva que podría usarse en combinación con dosis bajas de otros medicamentos. El tratamiento con LF y diclazurilo en el presente estudio mejoró la eficacia anticoccidial y resultó en una reducción significativa del recuento de ooquistes en comparación con G3.
Como se muestra en el presente estudio, el grupo no tratado (G2) mostró una serie de signos clínicos de infección en el quinto día después de la infección, como reducción del apetito, letargo, comportamiento depresivo, pérdida de peso y la presencia de diarrea sanguinolenta, que han sido reportados en la investigación previa (69) como signos clínicos comunes en la infección por coccidios. Estos signos podrían explicarse por la hipótesis de que el ambiente del tracto digestivo del huésped estimula la exciestación de ooquistes en las mollejas, lo que resulta en la liberación de esporozoítos que invaden y matan las células de la mucosa intestinal e inician el ciclo reproductivo del parásito. Por su parte, los grupos infectados tratados con diclazurilo y/o LF (G3-G5) mostraron grados más leves de síntomas, lo que podría atribuirse a la actividad anticoccidial del diclazurilo (70). En el presente estudio, G2 experimentó una disminución obvia en el peso corporal y el peso corporal, lo que podría atribuirse a un menor consumo de alimento y a la ruptura de la integridad intestinal como membrana absorbente, lo que daría lugar a una digestión de nutrientes y una utilización del alimento mucho menos efectivas (71). Estos resultados coincidieron con Choi et al. (61) quienes atribuyeron estos hallazgos a los efectos deletéreos del parásito Eimeria que invade el intestino del pollo y retarda su función intestinal. Por el contrario, todos los grupos tratados con diclazurilo y/o LF (G3-G5) mostraron un aumento significativo de la PCA y la PC. Resultados similares fueron reportados por El-Azm et al. (72) e Iraee et al. (73) quienes demostraron un aumento de peso corporal y peso corporal después del tratamiento con diclazurilo en el agua de bebida durante 3 días consecutivos en pollos infectados con especies de Eimeria. La posible explicación es la elevada actividad de las enzimas correspondientes, amilasa y lipasa, además de la actividad anticoccidial del diclazurilo, que aumenta la altura de las vellosidades, compensando la baja eficiencia alimenticia causada por la infección por Eimeria spp. (74). Como se muestra en el presente estudio, la alimentación con LF a partir del primer día de edad aumentó significativamente el peso corporal y el peso corporal en comparación con los observados en los grupos control (G1 y G2). Estos resultados fueron confirmados por un estudio previo (75), en el que se registró una mejora en el peso corporal y el peso corporal por la suplementación con LF, lo que puede estar relacionado con la mejora de la digestibilidad de los nutrientes, el aumento de la absorción de nutrientes (76) y el desencadenamiento de la proliferación de las células de la mucosa y su diferenciación (22). Los grupos tratados con LF (G4, G5) mostraron que la inhibición del crecimiento de patógenos invasores en el tracto gastrointestinal puede aumentar indirectamente el estado de salud y el rendimiento del crecimiento de las aves durante el período de cría (77), lo que refleja la actividad antimicrobiana de LF por la actividad secuestradora de hierro de los patógenos (19).
En el presente estudio, la infección por E. tenella causó daño agudo en el ciego de los pollos en pollos infectados no tratados (G2) combinado con la presencia de varios estadios de desarrollo de coccidios, enterocitos necróticos e infiltraciones leucocitarias en las células epiteliales cecales. De manera similar, investigaciones previas (30) reportaron la presencia de infiltración linfocítica y macrófagos con diferentes estadios del parásito coccidio en la infección cecal post-E. tenella. Por el contrario, el tratamiento de pollos infectados con diclazurilo y/o LF (G3-G5) mostró efectos protectores contra los efectos dañinos de E. tenella al atenuar el daño cecal. Este hallazgo es consistente con investigaciones previas (78) que reportan que el diclazurilo alivió el daño del ciego causado por E. tenella y mató todas las etapas de desarrollo de E. tenella, además de causar una actividad significativa en la detención del desarrollo del ciclo coccidial dentro de los pollos tratados (79), en conjunto consistentes sin hallazgos histopatológicos. Mientras tanto, los grupos infectados tratados (G3-G5) experimentaron la restauración de estos cambios histopatológicos reportados que podrían atribuirse al efecto anticoccidial del diclazurilo y la LF en todos los grupos tratados al mejorar los sistemas antioxidante e inmunológico en los pollos.
El presente estudio reveló que los parámetros hematológicos medidos (PCV, % Hb y recuento de glóbulos rojos) disminuyeron drásticamente en el grupo control positivo (G2), lo que puede ser el resultado de un daño esquizonte de segunda generación, lo que conduce a un daño sustancial de los vasos sanguíneos de la mucosa y pérdida de sangre (80). Del mismo modo, un estudio previo (81) registró una reducción significativa de los parámetros hematológicos en pollos infectados por E. tenella debido a la hemorragia aguda y a la rotura de tejido en la mucosa inducida por el ataque del parásito. Todos los grupos tratados (G3-G5) mostraron mejoría en los parámetros hematológicos, lo que está respaldado por un trabajo previo (82) que demostró que una dieta suplementaria de LF combinada con hierro era capaz de mejorar los índices hematológicos. Además, el diclazurilo reveló una actividad notable en la interrupción del ciclo de desarrollo de coccidiales dentro de las aves tratadas, especialmente cuando se aplica en el primer día de aparición de sangre en las heces (79). Mientras tanto, en el presente estudio, hubo un aumento significativo de leucocitos y heterófilos, además de una disminución del recuento de linfocitos en el grupo infectado no tratado (G2) en comparación con el grupo control sano (G1). Estos resultados podrían explicarse por la inducción de la expresión de la respuesta inflamatoria a la infección (83), que suele asociarse a neutrofilia y linfopenia (84). En marcado contraste, los pollos infectados suplementados con diclazurilo y/o LF (G3-G5) mostraron una reducción significativa en el total de leucocitos y heterófilos en comparación con el grupo infectado no tratado (G2), lo que ilustra las funciones inmunomoduladoras del diclazurilo y la LF. Del mismo modo, Ward y Conneely (85) registraron que la LF mostró un papel regulador en el sistema inmune in vivo.
La proteína total y sus componentes principales, la albúmina y las globulinas, desempeñan un papel clave en la actividad del sistema inmunológico en varias especies (86). Por lo tanto, la química sanguínea se vio muy afectada por el desafío de Eimeria en este estudio. Los grupos de control positivos experimentaron una disminución de la TP sérica, la albúmina, alfa 1 y las gammaglobulinas, mientras que hubo un aumento significativo de los niveles de AST, ALT, ALP, ácido úrico y creatinina, lo que podría deberse a la mala absorción de proteínas y otros nutrientes del intestino debido a la hemorragia y el daño de la mucosa en estas aves (2). La reducción en la ingesta de alimento y la absorción de proteínas eleva el catabolismo proteico en los tejidos musculares, lo que conduce a la degradación muscular y, finalmente, eleva los niveles séricos de ALT, AST, creatinina y ácido úrico (33). La actividad de la fosfatasa alcalina puede ser un signo sensible de la patogénesis en la infección por coccidiales del ciego (87). La hemorragia externa también puede causar hiperactividad de la médula ósea para liberar el exceso de glóbulos sanguíneos, lo que aumenta el nivel sérico de FA (88). Esto también podría estar relacionado con la destrucción celular debido a la infección parasitaria, que desencadena el escape de enzimas al torrente sanguíneo (89). La destrucción de las células induce la liberación de muchas enzimas como AST, ALT y ALP en el suero y, por lo tanto, sus niveles son elevados dependiendo del tipo y la extensión de la destrucción. Por el contrario, los grupos tratados con diclazurilo y/o LF (G3-G5) mostraron mejoría en todos estos parámetros en comparación con el grupo infectado no tratado (G2). Estos resultados son consistentes con trabajos previos (91) que informaron una mejoría de las funciones hepato-renales a través de una reducción significativa en la pérdida de proteínas, daño tisular y liberación de enzimas por LF, que se expresa por la elevación de la TP, la albúmina y la globulina séricas y la disminución de los niveles de ALP, ALT, AST y creatinina. La proteína sérica total expresada por electroforesis involucra albúmina y alfa, beta y gammaglobulinas. Las globulinas están implicadas en varias proteínas de fase aguda de importancia diagnóstica. En el presente estudio, los pollos tratados con LF registraron gammaglobulinas elevadas, lo que refleja sus efectos inmunomoduladores y preventivos en el hígado (92).
El presente estudio reportó una disminución altamente significativa en los niveles de glucosa sérica en G2 en comparación con el grupo control normal (G1). De manera similar, Abd El-Maksoud et al. (93) mencionaron que la glucosa sérica disminuyó en los pollos infectados con coccidios, lo que podría deberse a la anorexia y/o inflamación del tracto intestinal, lo que impidió la absorción de glucosa y la glucogenólisis hepática (94). Además, la reducción de los niveles séricos de glucosa puede estar relacionada con las actividades de prevención de la lipasa y la amilasa en los grupos infectados no tratados (95). Por el contrario, los grupos tratados con diclazurilo y LF (G3-G5) mostraron un aumento en los niveles séricos de glucosa, lipasa y amilasa. De manera similar, Ghasemi-Sadabadi et al. (96) observaron que el diclazurilo aumentó la glucosa sérica en pollos de engorde, lo que podría atribuirse a los roles de la LF en el refuerzo y aumento del tamaño y la función epitelial intestinal.
Nuestros resultados revelaron que G2 mostró un aumento significativo en la concentración de MDA y NO en comparación con el grupo control normal (G1). Estos resultados concuerdan con investigaciones previas (97) que informan de un aumento de la concentración de MDA y de una marcada reducción de la actividad de la SOD en aves infectadas, lo que indica la aparición de estrés oxidativo debido a la infección y la alteración del equilibrio antioxidante/prooxidante en favor de los prooxidantes. El estrés oxidativo relacionado con E. tenella en pollos de engorde da como resultado una reducción de los niveles de GSH y enzimas SOD que son importantes para inhibir la destrucción de radicales libres a lo largo de la infección por coccidios (98). Mientras tanto, los niveles de NO se elevaron en G2 en respuesta a las fases patógenas de esporozoítos que penetraron e inflamaron las células cecales (99). Por el contrario, el tratamiento de todos los animales infectados con LF y/o diclazurilo (G3-G5) mostró una mejoría de todos estos parámetros, ya que se redujo notablemente el MDA y se elevaron las enzimas antioxidantes como CAT y SOD, lo que refleja los efectos antioxidantes de la LF relacionados con sus propiedades de unión al hierro mediante el secuestro de hierro (100).
Se observó una alta regulación positiva en la expresión génica hepática de IL-2 e INF-γ en el grupo de control positivo (G2) en comparación con el grupo sano normal (G1). Las células T desempeñan un papel clave en la regulación de la inmunidad mediada por células mediante la secreción de citoquinas como INF-γ e IL2 y media la respuesta inmunitaria T en la coccidiosis aviar. Entre las citoquinas implicadas en la coccidiosis, el IFN-γ es un inmunomodulador representativo y el que más atención ha recibido debido a su influencia directa en el desarrollo intracelular de Eimeria. Resultados similares se observaron en pollos infectados con E. tenella (G2) en nuestro presente estudio. Se registró una expresión regulada al alza de IFN-γ en las amígdalas cecales, el bazo y las infecciones postprimarias y secundarias por E. tenella. Se encontró que el IFN-γ es liberado por las células T específicas mejoradas por mitógenos o antígenos que están presentes en la sangre de los pollos infectados con Eimeria (101). Por el contrario, los otros grupos tratados (G3-G5) podrían haber experimentado una mejora en la condición inmunológica de los pollos. Del mismo modo, un estudio previo (102) registró que la respuesta inmunitaria se modula utilizando aditivos alimentarios LF como prebióticos inmunoestimulantes. Esta mejora de la expresión génica inmune hepática en pollos alimentados con LF podría estar relacionada con la mejora de la inmunidad de las aves contra enfermedades infecciosas. Sin embargo, la información sobre la influencia de los aditivos LF en la respuesta inmunitaria y su impacto durante la infección de las aves de corral es limitada.
Nuestros resultados demostraron que el Grupo 5 (G5) mostró una mejora significativa en el estado oxidante/antioxidante, mejoró la salud del tejido cecal y redujo el desprendimiento de ooquistes. Estos hallazgos sugieren que el prebiótico potenció el efecto anticoccidial del diclazurilo, consistente con los resultados reportados por Farag et al. (103). De manera similar, Barberis et al. (104) destacaron la eficacia de la combinación de prebióticos y fármacos anticoccidiales en el control de las infecciones por coccidios en pollos, observando mejoras en las respuestas inmunes y el rendimiento del crecimiento. Dada la creciente resistencia a los fármacos en las cepas de Eimeria, la combinación de LF prebióticos con fármacos anticoccidiales presenta una estrategia prometedora para mejorar la eficacia de los fármacos y controlar las infecciones por coccidios, lo que podría servir como una alternativa eficaz en el tratamiento de estas infecciones. Sin embargo, es importante considerar que el presente estudio tiene algunas limitaciones, como que no se midió el consumo de alimento y la tasa de conversión alimenticia, lo que podría haber fortalecido aún más los hallazgos. Además, aunque el estudio se centró en medir los parámetros antioxidantes en los tejidos hepáticos, al igual que numerosos estudios anteriores, habría sido beneficioso evaluar también estos parámetros en los tejidos cecales para un análisis más completo.
5 Conclusión
El estudio actual concluyó que la LF podría poseer una potente actividad anticoccidial y antiinflamatoria, ya que redujo la gravedad de la lesión y el desprendimiento de los ooquistes. Sin embargo, esto debería ir precedido de una futura ampliación de su evaluación sobre el terreno. Curiosamente, el uso combinado de diclazuril con LF dio lugar a una mejoría superior del estado oxidante/antioxidante y de los tejidos cecales, así como a puntuaciones más bajas de las lesiones y al desprendimiento de ooquistes, lo que se refleja en los hallazgos histopatológicos de los tejidos cecales. Claramente, se puede recomendar el uso de LF como un potencial anticoccidial natural en pollos de engorde para combatir los efectos secundarios reportados de los agentes anticoccidiales comunes. Además, se podría concluir que la LF se puede utilizar como una alternativa natural promotora del crecimiento en pollos de engorde, ya que tiene efectos positivos sobre el rendimiento del crecimiento. Se recomienda realizar más investigaciones para explorar las principales vías mecanicistas implicadas en las acciones anticoccidiales de la FL y su posible aplicación en enfermedades similares de importancia veterinaria y médica.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio se incluyen en el artículo/material complementario, las consultas adicionales pueden dirigirse al autor correspondiente.
Declaración ética
El estudio en animales fue aprobado por AHRI de conformidad con el comité de la ARC y la ICUC (ARC, AHRI, IACUC, 85/23). El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
AGA: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Administración de proyectos, Recursos, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. NE: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Administración de proyectos, Recursos, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. MH: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Administración de proyectos, Recursos, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. SS: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Administración de proyectos, Recursos, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. AG: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Administración de proyectos, Recursos, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. MM: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Administración de proyectos, Recursos, Software, Supervisión, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. AAMA: Curación de datos, análisis formal, adquisición de fondos, software, validación, redacción, revisión y edición. HA: Curación de datos, análisis formal, adquisición de fondos, investigación, recursos, software, validación, redacción, revisión y edición. EE: Conceptualización, Curación de datos, Análisis formal, Obtención de fondos, Investigación, Metodología, Recursos, Software, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición.
Financiación
El/los autor/es declara(n) que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio fue apoyado por los investigadores de la Universidad Princesa Nourah bint Abdulrahman que apoyan el Proyecto No. (PNURSP2024R401), Universidad Princesa Nourah bint Abdulrahman, Riad, Arabia Saudita.
Reconocimientos
Nos gustaría agradecer a los investigadores de la Universidad Princesa Nourah bint Abdulrahman que apoyan el Proyecto No. (PNURSP2024R401), Universidad Princesa Nourah bint Abdulrahman, Riad, Arabia Saudita. Se agradece enormemente a los autores a la empresa farmacéutica y de nutrición de laboratorio Dulex, New Cairo, Egipto, por proporcionarnos la materia prima de lactoferrina.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Fatoba, AJ, y Adeleke, MA. Diagnóstico y control de la coccidiosis del pollo: una actualización reciente. J Parásito Dis. (2018) 42:483–93. doi: 10.1007/s12639-018-1048-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Williams, RB . Coccidiosis intercurrente y enteritis necrótica de pollos: manejo racional e integrado de la enfermedad mediante el mantenimiento de la integridad intestinal. Pathol aviar. (2005) 34:159–80. doi: 10.1080/03079450500112195
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Lee, Y, Park, I, Wickramasuriya, SS, Ben, AJ, Koziol, ME, y Lillehoj, HS. La administración concomitante de IL-7 de pollo o péptido 2 de lisina NK mejora la eficacia de la vacunación con el factor de elongación de Eimeria-1α contra la infección por Eimeria maxima en pollos de engorde. Poult Sci. (2022) 101:102013. doi: 10.1016/j.psj.2022.102013
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. López-Osorio, S, Chaparro-Gutiérrez, JJ, y Gómez-Osorio, LM. Visión general del ciclo de vida de la Eimeria avícola y de las interacciones huésped-parásito. Front Vet Sci. (2020) 7:384. doi: 10.3389/fvets.2020.00384
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Blake, DP, Clark, EL, Macdonald, SE, Thenmozhi, V, Kundu, K, Garg, R, et al. Diversidad poblacional, genética y antigénica del apicomplejo Eimeria tenella y su relevancia para el desarrollo de vacunas. Proc Natl Acad Sci USA. (2015) 112:E5343–50. doi: 10.1073/pnas.1506468112
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Noack, S, Chapman, HD y Selzer, PM. Medicamentos anticoccidiales de la industria ganadera. Parasitol Res. (2019) 118:2009–26. doi: 10.1007/s00436-019-06343-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Williams, RB . Aspectos epidemiológicos del uso de vacunas anticoccidiales vivas para pollos. Int J Parasitol. (1998) 28:1089–98. doi: 10.1016/S0020-7519(98)00066-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Zhang, M, Qiu, J, Shu, X, Tang, X, Sha, X, Wu, L, et al. Farmacocinética, actividad y eliminación de residuos de R- y S-Diclazurilo en pollos de engorde. J Agric Food Chem. (2020) 68:8987–95. doi: 10.1021/acs.jafc.0c03091
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Elokil, AA, Abouelezz, KFM, Ahmad, HI, Pan, Y y Li, S. Investigación de los impactos de la exposición a antibióticos en la diversidad de la microbiota intestinal en pollitos. Animales (Basilea). (2020) 10:896. doi: 10.3390/ani10050896
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Elkomy, A, Aboubakr, M y Medhat, Y. Eficacia anticoccidial del diclazurilo en pollos de engorde infectados experimentalmente con Eimeria tenella. Benha Vet Med J. (2015) 29:23–8. doi: 10.21608/bvmj.2015.31541
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Awaad, MH, Afify, MA, Zouelfakar, SA, e Hilali, MA. Eficacia anticoccidial de sapogeninas esteroideas (coccidiostato orgánico) en pollos de engorde (ensayos de semicampo y de campo). Egypt Vet Med Soci Parasitol J. (2003) 1:123–36.
12. Allen, PC, y Fetterer, RH. Avances recientes en biología e inmunobiología de especies de Eimeria y en el diagnóstico y control de la infección por estos parásitos coccidios de las aves de corral. Clin Microbiol Rev. (2002) 15:58–65. doi: 10.1128/CMR.15.1.58-65.2002
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Abd-ELrahmanSM, MSA, Mohamed, SE, El-Khadragy, MF, Dyab, AK, Hamad, N, Safwat, MM, et al. Efecto comparativo de la alicina y el extracto de ajo alcohólico sobre la morfología e infectividad de ooquistes de Eimeria tenella en pollos. Animales (Basilea). (2022) 12:3185. doi: 10.3390/ani12223185
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Elmahallawy, EK, Fehaid, A, El-Shewehy, DMM, Ramez, AM, Alkhaldi, AAM, Mady, R, et al. La S-metilcisteína mejora el daño intestinal inducido por la infección por Eimeria tenella al atacar el estrés oxidativo y los moduladores inflamatorios. Front Vet Sci. (2022) 8:754991. doi: 10.3389/fvets.2021.754991
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Cai, H, Luo, S, Liu, Q, Zhou, Q, Yan, Z, Kang, Z, et al. Efectos de una preparación probiótica compleja, Fengqiang Shengtai y vacuna contra la coccidiosis sobre el rendimiento y la microbiota intestinal de pollos de engorde desafiados con vectores parásitos de Eimeria spp. (2023) 16:253. doi: 10.1186/s13071-023-05855-5
16. Taherpour, K, Moravej, H, Taheri, HR y Shivazad, M. Efecto de la inclusión dietética de glicéridos probióticos, prebióticos y de ácido butírico sobre la resistencia contra la coccidiosis en pollos de engorde. J Poult Sci. (2012) 49:57–61. doi: 10.2141/jpsa.011031
17. Kell, DB, Heyden, EL, y Pretorius, E. La biología de la lactoferrina, una proteína de unión al hierro que puede ayudar a defenderse contra virus y bacterias. Frente Immunol. (2020) 11:1221. doi: 10.3389/fimmu.2020.01221
18. Elazab, MFA, Elbaiomy, AEA, Ahmed, MS, Alsharif, KF, Dahran, N, Elmahallawy, EK, et al. Efectos mejoradores de la lactoferrina bovina sobre la hematotoxicidad inducida por benceno en ratas albinas. Front Vet Sci. (2022) 9:907580. doi: 10.3389/fvets.2022.907580
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Vega-Bautista, A, de la Garza, M, Carrero, JC, Campos-Rodríguez, R, Godínez-Victoria, M, y Drago-Serrano, ME. El impacto de la lactoferrina en el crecimiento de las bacterias intestinales. Int J Mol Sci. (2019) 20:4707. doi: 10.3390/ijms20194707
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Humphrey, BD, Huang, N y Klasing, KC. El arroz que expresa lactoferrina y lisozima tiene propiedades similares a los antibióticos cuando se alimenta a los pollitos. J Nutr. (2002) 132:1214–8. doi: 10.1093/jn/132.6.1214
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. de la Rosa, G, Yang, D, Tewary, P, Varadhachary, A, y Oppenheim, JJ. La lactoferrina actúa como una alarmina para promover el reclutamiento y la activación de APC y respuestas inmunes específicas de antígeno. J Immunol. (2008) 180:6868–76. doi: 10.4049/jimmunol.180.10.6868
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Legrand, D . La lactoferrina, una molécula clave en los procesos inmunitarios e inflamatorios. Biochem Cell Biol. (2012) 90:252–68. doi: 10.1139/o11-056
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Actor, JK, Hwang, SA, Olsen, M, Zimecki, M, Hunter, RL Jr y Kruzel, ML. Inmunomodulación de lactoferrina de la respuesta a DTH en ratones. Int Immunopharmacol. (2002) 2:475–86. doi: 10.1016/S1567-5769(01)00189-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Baveye, S, Elass, E, Mazurier, J, Spik, G y Legrand, D. Lactoferrina: una glicoproteína multifuncional involucrada en la modulación del proceso inflamatorio. Clin Chem Lab Med. (1999) 37:281–6. doi: 10.1515/cclm.1999.049
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Giansanti, F, Panella, G, Leboffe, L, y Antonini, G. Lactoferrina de la leche: propiedades nutracéuticas y farmacológicas. Productos farmacéuticos (Basilea). (2016) 9:61. doi: 10.3390/ph9040061
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Sahin, O, Ziaei, A, Karaismailoğlu, E, y Taheri, N. Niveles séricos de enzima convertidora de angiotensina y lisozima en pacientes con afectación ocular de enfermedades autoinmunes e infecciosas. BMC Ophthalmol. (2016) 16:19. doi: 10.1186/s12886-016-0194-4
27. Omata, Y, Satake, M, Maeda, R, Saito, A, Shimazaki, K, Yamauchi, K, et al. Reducción de la infectividad de los esporozoítos de toxoplasma gondii y Eimeria stiedai mediante tratamiento con lactoferricina bovina. J Vet Med Sci. (2001) 63:187–90. doi: 10.1292/jvms.63.187
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Soulsby, EJL . (1982). Helmintos, artrópodos y protozoos de animales domésticos. 7ª Edn. Bailliere Tindall: Londres. 573–574
29. Conway, DP, y Mackenzie, ME. Procedimientos de diagnóstico y pruebas de coccidiosis aviar. Tercera Edición. Editorial Blackwell. (2007). 13.
30. Abd El Ghany, AM, Ibrahimb, HA, Hassanb, SWM, y Galalb, AAA. Eficacia del aceite de alicina y diclazurilo en pollos de engorde: efecto sobre la coccidiosis cecal y antioxidante del tejido cecal. EVMPS. (2021) 17:146–68. doi: 10.21608/evmspj.2021.297930
31. Enany, M, El Gammal, AEA, Solimane, R, El Sissi, A y Hebashy, A. Evaluación del efecto inmunomodulador de la lactoferrina sobre la respuesta inmunitaria de pollos de engorde. Veterinario del Canal de Suez Med J. (2017) 22:135–46. doi: 10.21608/scvmj.2017.62452
32. Dalloul, RA, Lillehoj, HS, Shellem, TA, y Doerr, JA. Mejora de la inmunidad de la mucosa contra Eimeria acervulina en pollos de engorde alimentados con un probiótico a base de lactobacilos. Poult Sci. (2003) 82:62–6. doi: 10.1093/ps/82.1.62
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. El-Shazly, KA, El-Latif, AA, Abdo, W, El-Morsey, A, El-Aziz, MIA, y El-Mogazy, H. Actividad anticoccidial de la fluoroquinolona lomefloxacino contra la infección experimental por Eimeria tenella en pollos de engorde. Parasitol Res. (2020) 119:1955–68. doi: 10.1007/s00436-020-06692-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Suvarna, KS, Layton, C, y Bancroft, JD. Libro electrónico de Teoría y práctica de las técnicas histológicas de Bancroft. Ámsterdam: Elsevier Health Sciences (2018).
35. Gibson-Corley, KN, Olivier, AK y Meyerholz, DK. Principios para una puntuación histopatológica válida en la investigación. Veterinario Pathol. (2013) 50:1007–15. doi: 10.1177/0300985813485099
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Feldman Zinkl, JG, y Jain, NCBF. Hematología veterinaria de Schalm. 5ª ed. Filadelfia, Pensilvania: Lippincott Williams y Wilkins (2000).
37. Dein, FJ . Hematología En: Harrison GJ, Harrison LR (eds): Medicina y Cirugía Aviar Clínica.Filadelfia, WB Saunders Co. (1986). págs. 174-191.
38. Coles, EH . Patología clínica veterinaria. 4ª ed. Filadelfia, Pensilvania: WB Saunders Company (1986).
39. Andreasen, CB, y Latimer, KS. Características de tinción citoquímica de heterófilos y eosinófilos de pollo. Veterinario Clin Pathol. (1990) 19:51–4. doi: 10.1111/j.1939-165X.1990.tb00543.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Reitman, S, y Frankel, S. Método colorimétrico para la determinación de transaminasas glutámicas oxalacéticas y glutámicas pirúvicas séricas. Am J Clin Pathol. (1957) 28:56–63. doi: 10.1093/ajcp/28.1.56
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. El-Aaser, AA, y El-Merzabani, MM. Determinación simultánea de las actividades de la 5′-nucleotidasa y la fosfatasa alcalina en suero. Z Klin Chem Klin Biochem. (1975) 13:453–9. doi: 10.1515/cclm.1975.13.10.453
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Wybenga, DR, Di Giorgio, J, y Pileggi, VJ. Métodos manuales y automatizados para la medición de nitrógeno ureico en suero entero. Clin Chem. (1971) 17:891–5. doi: 10.1093/clinchem/17.9.891
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Henry, RJ, Cannon, CD, y Winkelman, JW. «Química clínica, principios y técnicas». 2ª Ed., W.B. Saunders, Co. Harport y Rowhogerstown, Departamento Médico. (1974). 1629.
44. Tietz, noroeste . Guía clínica de pruebas de laboratorio. 3ª Edn Filadelfia, Pensilvania: WB Saunders; (1995)268–273.
45. López, J, Carl, A, Burtis Ashwood, Edward, R, Bruns y David, E. Tietz libro de texto de química clínica y diagnóstico molecular (5ª edición): Elsevier, St. Louis, EE. UU., 2012, 2238 pp, 909 ilustraciones. (2013) Indian J Clin Biochem 28: 104-105. doi: 10.1007/s12291-012-0287-7
46. Montgomery, HAC, y Dymock, JF. La determinación de nitrito en agua. Analista. (1961) 1:414–6. doi: 10.1039/AN9618600411
47. Koracevic, D, Koracevic, G, Djordjevic, V, Andrejevic, S, y Cosic, V. Método para la medición de la actividad antioxidante en fluidos humanos. J Clin Pathol. (2001) 54:356–61. doi: 10.1136/jcp.54.5.356
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Ohkawa, H, Ohishi, N y Yagi, K. Ensayo de peróxidos lipídicos en tejidos animales mediante reacción de ácido tiobarbitúrico. Bioquímica anal. (1979) 95:351–8. doi: 10.1016/0003-2697(79)90738-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Nishikimi, M, Appaji, N, y Yagi, K. La ocurrencia del anión superóxido en la reacción de metosulfato de fenazina reducido y oxígeno molecular. Biochem Biophys Res Commun. (1972) 46:849–54. doi: 10.1016/S0006-291X(72)80218-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Aebi, H . Catalasa in vitro. Métodos: Enzimol. (1974) 105:121–6. doi: 10.1016/s0076-6879(84)05016-3
51. Beutler, E, Duron, O, y Kelly, BM. Método mejorado para la determinación del glutatión en sangre. J Lab Clin Med. (1963) 61:882–8.
52. Sonnenwirth, AC, Gradwohl, RBH, y Jaret, L. Métodos de laboratorio clínico y diagnóstico de Gradwohl. 8ª Ed., 258-259. Londres: C.V. Mosby Co. (1980)
53. Davis, BJ . Electroforesis discal. II método y aplicación a la proteína sérica humana. Ann N Y Acad Sci. (1964) 121:404–27. doi: 10.1111/j.1749-6632.1964.tb14213.x
54. Wang, YC, Deng, JL, Xu, SW, Peng, X, Zuo, ZC, Cui, HM, et al. Efectos de la zearalenona sobre los niveles de ARNm de IL-2, IL-6 e IFN-γ en los linfocitos esplénicos de pollos. Sci World J. (2012) 2012:567327. doi: 10.1100/2012/567327
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Zhou, H, Buitenhuis, AJ, Weigend, S, y Lamont, SJ. Polimorfismos promotores de genes candidatos y cinética de respuesta de anticuerpos en pollos: interferón-γ, interleucina-2 e inmunoglobulina de cadena ligera. Poult Sci. (2001) 80:1679–89. doi: 10.1093/ps/80.12.1679
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Yuan, JS, Reed, A, Chen, F y Stewart, CN. Análisis estadístico de datos de PCR en tiempo real. BMC Bioinformática. (2006) 7:1–12. doi: 10.1186/1471-2105-7-85
57. Youssefi, MR, Alipour, R, Fakouri, Z, Shahavi, MH, Nasrabadi, NT, Tabari, MA, et al. La suplementación dietética con nanoemulsión de eugenol alivia los efectos negativos de la coccidiosis experimental en la salud y el rendimiento del crecimiento de los pollos de engorde. Moléculas. (2023) 28:2200. doi: 10.3390/moléculas28052200
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Liu, Y, Zheng, J, Li, J, Gong, P y Zhang, X. Inmunidad protectora inducida por una vacuna de ADN que codifica el romboide de Eimeria tenella contra el desafío homólogo. Parasitol Res. (2013) 112:251–7. doi: 10.1007/s00436-012-3132-2
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Daofeng, Q, Wenxiu, M, Yongmeng, Y y Jianzhong, H. Efecto de la dinitolmida intercalada en montmorillonita en la infección por E. tenella en pollos. Parasitol Res. (2014) 113:1233–8. doi: 10.1007/s00436-014-3762-7
60. Ahad, S, Tanveer, S, y Malik, TA. Actividad anticoccidial del extracto acuoso de un hongo silvestre (Ganoderma applanatum) durante la infección por coccidial inducida experimentalmente en pollos de engorde. J Parásito Dis. (2016) 40:408–14. doi: 10.1007/s12639-014-0518-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Choi, J, Ko, H, Tompkins, YH, Teng, PY, Lourenco, JM, Callaway, TR, et al. Efectos de la infección por Eimeria tenella en parámetros clave para la eficiencia alimenticia en pollos de engorde. Animales. (2021) 11:3428. doi: 10.3390/ANI11123428
Resumen de PubMed | Texto completo de Crossref | Google Académico
62. Gong, Z, Qu, Z, Wei, H, Chang, F y Cai, J. Características biológicas de una línea precoz de Eimeria tenella. Parasitol Res. (2024) 123:167. doi: 10.1007/s00436-024-08190-5
63. Walker, RA, Sharman, PA, Miller, CM, Lippuner, C, Okoniewski, M, Eichenberger, RM, et al. El análisis de secuenciación de ARN del transcriptoma de gametocitos de Eimeria tenella revela pistas sobre la base molecular de la reproducción sexual y la biogénesis de ooquistes. Genómica. (2015) 16:94. doi: 10.1186/s12864-015-1298-6
Resumen de PubMed | Texto completo de Crossref | Google Académico
64. Hungerford, TG . Enfermedades de las aves de corral, incluidas las aves de jaula y las palomas, 4ª ed. Sídney: Angus and Robertson Ltd. (1969) 335-340.
65. Fernando, MA . Eimeria: Infecciones del intestino En: PL Long editor. Coccidiosis del hombre y de los animales. Boca Ratón, FL: CRC Press (1990). 63–75.
66. Calnek, BW, Barnes, HJ, Beard, CW, McDougald, LR, y Saif, YM. Enfermedades de las aves de corral. 10ª ed. Ames, IA: Iowa State University Press (1997).
67. Souza, GC, Esteves, GF, Volpato, FA, Miotto, R, Mores, MAZ, Ibelli, AMG, et al. Efectos de las concentraciones variables de Eimeria challenge en la integridad intestinal de pollos de engorde. Aves de corral. (2024) 3:1–14. doi: 10.3390/poultry3010001
68. Reyes-López, M, Ramírez-Rico, G, Serrano-Luna, J, y de la Garza, M. Actividad de la Apo-Lactoferrina sobre protozoos patógenos. Farmacia. (2022) 14:1702. doi: 10.3390/pharmaceutics14081702
Resumen de PubMed | Texto completo de Crossref | Google Académico
69. Iacob, OC y Duma, V. Aspectos clínicos, paraclínicos y morfopatológicos en la eimeriosis cecal de pollos de engorde. Rev Sci Parasitol. (2009) 10:43–50.
70. Sivaseelan, S, Malmarugan, S, Balachandran, P, Vijayakumar, S, Poomathi, K, y Balasubramania, GA. Brote de enteritis necrótica en pollo asociada con ascaridiasis. VetJ de la India. (2012):90.
71. Walk, CL, Cowieson, AJ, Remus, JC, Novak, CL y McElroy, AP. Efectos de las enzimas dietéticas sobre el rendimiento y el número de células caliciformes intestinales de pollos de engorde expuestos a una vacuna viva contra ooquistes de coccidios. Poult Sci. (2011) 90:91–8. doi: 10.3382/ps.2010-00760
Resumen de PubMed | Texto completo de Crossref | Google Académico
72. El-Azm, IMA, El-Hamid, HSA, Ellakany, HF y El-Shall, NA. Sensibilidad de dos aislados de campo local de Eimeria tenella a maduramicina y diclazurilo. Zagazig Vet J. (2010) 38:8–18.
73. Iraee, HA, Iraee, MA, Youssefi, MR, y Tabari, MA. Parámetros de comportamiento de crecimiento en coccidiosis experimental de pollos tratados con Diclazuril y Clopidol: la necesidad de evaluar nuevos recursos anticoccidiales. Irán J Vet Med. (2015) 9:189–94.
74. Nabian, S, Arabkhazaeli, F, Seifouri, P y Farahani, A. Análisis morfométrico del intestino en coccidiosis experimental en pollos de engorde tratados con fármacos anticoccidiales. Irán J Parasitol. (2018) 13:493.
75. Siqueiros-Cendón, T, Arévalo-Gallegos, S, Iglesias-Figueroa, BF, García-Montoya, IA, Salazar-Martínez, J, y Rascón-Cruz, Q. Efectos inmunomoduladores de la lactoferrina. Acta Pharmacol Sin. (2014) 35:557–66. doi: 10.1038/aps.2013.200
Resumen de PubMed | Texto completo de Crossref | Google Académico
76. Geier, MS, Torok, VA, Guo, P, Allison, GE, Boulianne, M, Janardhana, V, et al. Los efectos de la lactoferrina en el entorno intestinal de los pollos de engorde. Hno. Poult Sci. (2011) 52:564–72. doi: 10.1080/00071668.2011.607429
Resumen de PubMed | Texto completo de Crossref | Google Académico
77. Weinberg, ED . Propiedades antibióticas y aplicaciones de la lactoferrina. Curr Pharm Des. (2007) 13:801–11. doi: 10.2174/138161207780363095
Resumen de PubMed | Texto completo de Crossref | Google Académico
78. Tian, E, Zhou, BH, Wang, XY, Zhao, J, Deng, W y Wang, HW. Efecto del diclazurilo sobre la morfología intestinal y la expresión de SIgA en pollos infectados con Eimeria tenella. Parasitol Res. (2014) 113:4057–64. doi: 10.1007/s00436-014-4074-7
Resumen de PubMed | Texto completo de Crossref | Google Académico
79. El-Dakhly, KHM, El-Sawah, AA, Shalaby, AA, y El-Nesr, KHA. Eficacia de lactobacillus acidophilus y/o diclazuril para la inhibición y control de la infección por Eimeria tenella en polluelos balady. Kafrelsheikh Vet Med J. (2006) 4:1–19. doi: 10.21608/kvmj.2006.109296
Resumen de PubMed | Texto completo de Crossref | Google Académico
80. Ellakany, HF, Abuakkada, SS, Oda, SS y El-Sayed, YS. Influencia de los bajos niveles de aflatoxinas dietéticas en las infecciones por Eimeria tenella en pollos de engorde. Trop Anim Salud Prod. (2011) 43:249–57. doi: 10.1007/s11250-010-9685-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
81. Adulugba, I, Goselle, O, Ajayi, O, Pam, K y Friday, S. Combinación fitosintética como grandes potenciadores de los parámetros hematológicos: un estudio de caso en aves de corral. Am J Fitomedicina Clin Ther. (2017) 5:1. doi: 10.21767/2321-2748.100322
82. Kawakami, H, Hiratsuka, M y Dosako, S. Efectos de la lactoferrina saturada de hierro en la absorción de hierro. Agric Biol Chem. (1988) 52:903–8. doi: 10.1080/00021369.1988.10868784
83. Çam, Y, Atasever, A, Eraslan, G, Kibar, M, Atalay, Ö, Beyaz, L, et al. Eimeria stiedae: infección experimental en conejos y efecto del tratamiento con toltrazuril e ivermectina. Exp parasitol. (2008) 119:164–72. doi: 10.1016/j.exppara.2008.01.005
Resumen de PubMed | Texto completo de Crossref | Google Académico
84. Anwar, AH, Kazmi, SIH y Khan, MN. Efecto de la coccidiosis inducida experimentalmente sobre algunos parámetros sanguíneos de terneros de búfalo. Pak J Biol Sci. (1999) 2:1024–6. doi: 10.3923/pjbs.1999.1024.1026
85. Ward, PP y Conneely, OM. Lactoferrina: papel en la homeostasis del hierro y la defensa del huésped contra la infección microbiana. Biometales. (2004) 17:203–8. doi: 10.1023/B:BIOM.0000027693.60932.26
Resumen de PubMed | Texto completo de Crossref | Google Académico
86. Moradian, AM, Dorafshan, S, Paykan Heyrati, F y Ebrahimi, E. Efectos de la lactoferrina bovina en la dieta sobre el crecimiento, los parámetros hematobioquímicos, las funciones inmunitarias y la tolerancia al estrés por exposición al aire en el cíclido africano Sciaenochromis fryeri. Aquac Nutr. (2018) 24:392–9. doi: 10.1111/anu.12570
87. Kogut, MH y Powell, KC. Hallazgos preliminares de alteraciones en la actividad de la fosfatasa alcalina sérica en pollos durante infecciones coccidiales. J Comp Pathol. (1993) 108:113–9. doi: 10.1016/S0021-9975(08)80215-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
88. Adamu, M, Boonkaewwan, C, Gongruttananun, N, y Vongpakorn, M. Cambios hematológicos, bioquímicos e histopatológicos causados por la coccidiosis en pollos. Agric Natural Resour. (2013) 47:238–46.
89. Hanada, K, Kumagai, K, Yasuda, S, Miura, Y, Kawano, M, Fukasawa, M, et al. Maquinaria molecular para el tráfico no vesicular de ceramidas. Naturaleza. (2003) 426:803–9. doi: 10.1038/nature02188
Resumen de PubMed | Texto completo de Crossref | Google Académico
90. Newsome, PN, Cramb, R, Davison, SM, Dillon, JF, Foulerton, M, Godfrey, EM, et al. Pautas sobre el manejo de los análisis de sangre hepáticos anormales. Tripa. (2018) 67:6–19. doi: 10.1136/gutjnl-2017-314924
91. El-Sherbeny, EM, y El-Shenawy, FA. Efecto de la lactoferrina sola o en combinación con bacitracina en la infección por Clostridium perfringens en conejos. Egipto J Chem. (2023) 66:13–30. doi: 10.21608/EJCHEM.2022.137842.6068
92. Assaraj, QSH, Alattar, HA, Farid, AS, y Fararah, KM. Influencia de la lactoferrina en la respuesta inmune en ratas intoxicadas por diazinón. Benha Vet Med J. (2018) 34:169–81. doi: 10.21608/BVMJ.2018.29426
93. Abd El-Maksoud, HA, Afaf, D, Abdel-Magid, M, y El-Badry, MA. Efecto bioquímico de la infestación por coccidios en gallinas ponedoras. Benha Vet Med J. (2014) 26:127–33.
94. Melkamu, S, y Asrat, M. Revista internacional de investigación avanzada en ciencias biológicas cambios bioquímicos causados por Eimeria spp en pollos de engorde. Int J Adv Res Biol Sci. (2019) 6:77–83. doi: 10.22192/ijarbs
95. Lee, KW, Ho Hong, Y, Lee, SH, Jang, SI, Park, MS, Bautista, DA, et al. Efectos de los programas de antibióticos y anticoccidiales promotores del crecimiento sobre el rendimiento y el estado inmunológico de los pollos de engorde. Res Vet Sci. (2012) 93:721–8. doi: 10.1016/j.rvsc.2012.01.001
Resumen de PubMed | Texto completo de Crossref | Google Académico
96. Ghasemi-Sadabadi, M, Ebrahimnezhad, Y, Shaddel-Tili, A, Bannapour-Ghaffari, V, y Saemi, P-BS. Comparación del uso de coccidiostáticos ionóforos y no ionóforos sobre el rendimiento, las características de la canal, los parámetros bioquímicos de la sangre y la flora microbiana intestinal en pollos de engorde. Irán J Appl Anim Sci. (2020) 10:693–704.
97. El-Maddawy, ZK, El-Sawy, AESF, Ashoura, NR, Aboelenin, SM, Soliman, MM, Ellakany, HF, et al. Uso de nanopartículas de óxido de zinc como agentes anticoccidiales en pollos de engorde junto con su impacto en el rendimiento del crecimiento, el estado antioxidante y el perfil hematobioquímico. Vida (Basilea). (2022) 12:74. doi: 10.3390/life12010074
Resumen de PubMed | Texto completo de Crossref | Google Académico
98. Alsulami, MN, y El-Saadony, MT. Suplementación de las dietas de pollos de engorde con nanopartículas bacterianas de selenio que mejoran el rendimiento, las características de la canal, los índices sanguíneos, el estado antioxidante y la microbiota cecal de pollos de engorde infectados con Eimeria tenella. Poult Sci. (2023) 102:103111. doi: 10.1016/j.psj.2023.103111
Resumen de PubMed | Texto completo de Crossref | Google Académico
99. Domínguez, PA, Pro-Martínez, A, Narciso-Gaytán, C, Hernández-Cázares, A, Sosa-Montes, E, Pérez-Hernández, P, et al. La suplementación simultánea de arginina y vitaminas antioxidantes E y C reduce el estrés oxidativo en pollos de engorde después de un desafío con Eimeria spp. Can J Anim Sci. (2015) 95:143–53. doi: 10.4141/cjas-2014-083
100. Raghuveer, TS, McGuire, EM, Martin, SM, Wagner, BA, Rebouché, CJ, Buettner, GR, et al. La lactoferrina en la dieta de los recién nacidos prematuros atenúa los productos de oxidación inducidos por el hierro. Pediatr Res. (2002) 52:964–72. doi: 10.1203/00006450-200212000-00024
Resumen de PubMed | Texto completo de Crossref | Google Académico
101. Breed, DGJ, Schetters, TPM, Verhoeven, NAP y Vermeulen, AN. Caracterización de la capacidad de respuesta relacionada con el fenotipo de los linfocitos de sangre periférica de pollos infectados con Eimeria tenella. Parásitos Inmunológicos. (1997) 19:563–9. doi: 10.1046/j.1365-3024.1997.d01-174.x
Resumen de PubMed | Texto completo de Crossref | Google Académico
102. Hashem, NMA, El-Son, MAM, Ateya, AI y Saleh, RM. Impacto de la suplementación con lactoferrina en el estrés oxidativo, la expresión génica y la disfunción inmunitaria inducida por Aeromonas veronii en tilapia del Nilo (Oreochromis niloticus). Aquac Res. (2022) 53:2392–407. doi: 10.1111/are.15757
103. Farag, EA, Ali, MA, y Seddiek, SA. Eficacia comparativa de los nanooligosacáridos prebióticos y diclazurilo en pollos infectados experimentalmente con Eimeria tenella. Zag Vet J. (2009) 37:102–15.
Palabras clave: anticoccidial, lactoferrina, diclazurilo, Eimeria tenella, pollos de engorde
Cita: Abd El Monsef AG, El Zohairy NF, Hassan MF, Salem SM, Gouda AA, Mansour MK, Alkhaldi AAM, Alzaylaee H y Elmahallawy EK (2024) Efectos del prebiótico (lactoferrina) y diclazurilo en pollos de engorde infectados experimentalmente con Eimeria tenella. Frente. Vet. Sci. 11:1416459. doi: 10.3389/fvets.2024.1416459
Editado por:
Vikrant Sudan, Universidad de Ciencias Veterinarias y Animales Guru Angad Dev, India
Revisado por:
Mohamed S. Ahmed, Universidad de Kafrelsheikh, Egipto
Sara Taha Elazab, Universidad de Mansoura, Egipto
Shawky Mohamed Aboelhadid, Universidad de Beni-Suef, Egipto
Derechos de autor © 2024 Abd El Monsef, El Zohairy, Hassan, Salem, Gouda, Mansour, Alkhaldi, Alzaylaee y Elmahallawy. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Ehab Kotb Elmahallawy, sa2elele@uco.es
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.