
Efectos del estrés por calor en los rasgos y metabolitos de la producción de leche

Efectos del estrés por calor en los rasgos y metabolitos de la producción de leche y estrategias de mitigación para razas de ganado lechero criadas en países tropicales y subtropicales
 Vicente Habimana1,2,3*,
Vicente Habimana1,2,3*,  Athumani Shabani Nguluma1,
Athumani Shabani Nguluma1,  Zabron Cuthibert Nziku4,
Zabron Cuthibert Nziku4,  Chinyere Charlotte Ekine-Dzivenu3,
Chinyere Charlotte Ekine-Dzivenu3,  Gota Morota5,
Gota Morota5,  Rafael Mrode3 y
Rafael Mrode3 y  Sebastian Wilson Chenyambuga1
Sebastian Wilson Chenyambuga1- 1Departamento de Ciencias Animales, Acuícolas y Pastizales, Universidad de Agricultura Sokoine, Morogoro, Tanzania
- 2SACIDS Africa Centre of Excellence for Infectious Diseases, SACIDS Foundation for One Health, Sokoine University of Agriculture, Morogoro, Tanzania
- 3Instituto Internacional de Investigaciones Pecuarias (ILRI), Nairobi, Kenya
- 4Instituto de Investigación Pecuaria de Tanzania (TALIRI) Zona Oriental, Tanga, Tanzania
- 5Escuela de Ciencias Animales, Instituto Politécnico y Universidad Estatal de Virginia, Blacksburg, VA, Estados Unidos
El estrés por calor es un problema importante para la industria láctea en muchas partes del mundo debido a sus efectos adversos sobre la productividad y la rentabilidad. El estrés por calor en el ganado lechero es causado por un aumento en la temperatura corporal central, lo que afecta la producción de grasa en la glándula mamaria. Reduce la producción de leche, la ingesta de materia seca y altera la composición de la leche, como los porcentajes de grasa, proteína, lactosa y sólidos no grasas, entre otros. Comprender los mecanismos biológicos de la adaptación climática, identificar y explorar firmas de selección, diversidad genómica e identificación de genes candidatos para la tolerancia al calor dentro de las razas lecheras indicina y taurina es una progresión importante hacia la cría de ganado lechero mejor adaptado a las condiciones climáticas cambiantes de los trópicos. Identificar las razas que son tolerantes al calor y su uso en programas de mejoramiento genético es crucial para mejorar la productividad y la rentabilidad del ganado lechero en los trópicos. El mejoramiento genético para la tolerancia al calor requiere la disponibilidad de parámetros genéticos, pero estos parámetros genéticos faltan actualmente en muchos países tropicales. En este artículo, revisamos los efectos de HS en el ganado lechero con respecto a (1) parámetros fisiológicos; (2) rendimiento de leche y rasgos de composición; y (3) metabolitos de leche y sangre para ganado lechero criado en países tropicales. Además, se discuten estrategias de mitigación como la modificación física del medio ambiente, el desarrollo nutricional y genético del ganado lechero tolerante al calor para prevenir los efectos adversos de HS en el ganado lechero. En climas tropicales, una estrategia más rentable para superar los efectos de HS es seleccionar genéticamente razas más adaptables y tolerantes al calor, el uso de animales cruzados para la producción de leche, es decir, cruces entre razas índicas como Gir, fulani blanco, N’Dama, Sahiwal o Boran a razas taurinas como Holstein-Friesian, Jersey o Brown Swiss. Los resultados de este examen contribuirán a la formulación de políticas con respecto a las estrategias para mitigar los efectos del SA en el ganado lechero de los países tropicales.
1. Introducción
El estrés por calor es una condición en la que los animales se vuelven incapaces de disipar la carga de calor generada por su metabolismo corporal y el medio ambiente, por lo que no pueden mantener su equilibrio térmico corporal (1-3). El estrés por calor conduce a un aumento de la temperatura rectal (RT), la frecuencia respiratoria (RR), la temperatura corporal central (TCC), la puntuación de jadeo (PS), la frecuencia del pulso (PR), la frecuencia de sudoración (SR) y la frecuencia cardíaca (FC) (4). El estrés por calor tiene efectos adversos en los rasgos de producción de leche, la fertilidad y la salud en el ganado lechero (5), especialmente en países tropicales donde el clima es siempre cálido y húmedo (6). El ganado lechero en estas regiones está expuesto a temperaturas ambiente (Ta) más altas, humedad relativa (HR), radiación solar (7) y variaciones del viento durante largos períodos de tiempo. En el ganado lechero, la temperatura y la humedad, generalmente combinadas como índice de temperatura-humedad, (THI) que superan su zona termoneutra causan una alta reducción en la ingesta de materia seca, la producción de leche y la fertilidad, que se asocian con cambios fisiológicos que incluyen CBT, RR, PR, RT y SR (8, 9). Estos efectos en la producción y composición de la leche, incluidos los porcentajes de grasa, proteína, lactosa y sólidos no grasos, pueden afectar directamente los ingresos generados por la producción lechera (10). Es crucial que el ganado lechero esté dentro de su zona termoneutral, donde no se requiere el gasto de energía para mantener la regulación interna de la temperatura corporal (3). La zona termoneutra para las razas de ganado lechero Bos taurus generalmente varía de 0.5 ° C a 20 ° C (8), aunque esto puede variar dependiendo de factores como el estado de producción, el nivel de aclimatación, el estado de embarazo, el tipo de alimento y las condiciones climáticas, incluida la velocidad del viento, la radiación solar y la humedad relativa (3, 8).
Incluso en países con temperaturas moderadas, como en la Unión Europea (UE) y los Estados Unidos de América (EE.UU.), el ganado lechero todavía puede experimentar estrés por calor (HS) debido al efecto del clima en el enfriamiento evaporativo (3). Se ha demostrado que el estrés por calor causa pérdidas económicas significativas en la industria láctea, con pérdidas anuales que van desde 1.69 a 2.26 mil millones de dólares en los Estados Unidos y una reducción de 0.73 millones de litros de leche en la India en 2020. En el Reino Unido, donde la mayoría de los rebaños lecheros están compuestos por razas Holstein-Friesian que son conocidas por su alto rendimiento de leche, la pérdida de producción de leche promedio anual proyectada es del 2,4% (~ 170 kg / vaca) en las regiones del suroeste (3). La selección genética para la tolerancia al calor en el ganado lechero sin comprometer su potencial de producción de leche se considera una estrategia sostenible para apoyar la producción de ganado lechero en ambientes tropicales. Las razas de ganado lechero en los países tropicales son diversas, incluido el ganado Bos indicus adaptado al calor y las razas Bos taurus menos resistentes, y pueden responder de manera diferente a los efectos de HS debido a su composición genética (11). Por lo tanto, la evaluación genética para la termotolerancia y la selección de animales tolerantes al calor basadas en regiones y climas específicos es posible y es una medida rentable para mejorar la producción de ganado lechero en condiciones de clima cálido. Cabe señalar que los rasgos de producción de leche a menudo son antagónicos a la tolerancia al calor, y la selección genética para una mayor producción de leche sin considerar la tolerancia al calor puede resultar en una mayor susceptibilidad a la HS (4, 12, 13). Por lo tanto, es necesario un enfoque equilibrado que considere tanto los rasgos de producción de leche como la tolerancia al calor para una selección genética efectiva en los programas de cría de ganado lechero. El estudio de Dikmen et al. (14) informó que las vacas Holstein tolerantes al calor tenían un mayor rendimiento de leche y un mejor rendimiento reproductivo en comparación con las vacas sensibles al calor en un ambiente tropical. Sigdel et al. (15) encontraron que el cruce de vacas Holstein con razas tolerantes al calor como Sahiwal y Gir mejoró el rendimiento de la leche y el rendimiento reproductivo en condiciones de HS. Además, Ekine-Dzivenu et al. (13) demostraron que la incorporación de la termotolerancia como criterio de selección en los programas de mejoramiento puede mejorar la producción de leche.
Además de la selección genética, también se pueden implementar modificaciones ambientales para mitigar el impacto de HS en el ganado lechero. Estos pueden incluir proporcionar sombra, mejorar la ventilación, usar sistemas de enfriamiento evaporativo y ajustar las prácticas de alimentación y riego. Sin embargo, estas medidas pueden no ser siempre rentables o factibles, especialmente en ambientes tropicales con recursos limitados (4, 6). Por lo tanto, la integración de la selección genética para la tolerancia al calor en los programas de mejoramiento puede ser una solución más sostenible y a largo plazo para mitigar los efectos adversos de HS en los rasgos de producción de leche, fertilidad y salud en el ganado lechero. Por lo tanto, el objetivo de esta revisión fue (1) caracterizar las respuestas genéticas y fenotípicas a HS, (2) identificar genes candidatos responsables de la tolerancia al calor; y (3) discutir estrategias de mitigación para razas de ganado lechero criadas en países tropicales. Las conclusiones de este examen contribuirán a la formulación de políticas con respecto a las estrategias para prevenir los efectos del SA en las razas de ganado lechero en el África subsahariana, donde el clima es muy cálido y húmedo casi todo el año.
2. Razas de ganado lechero criadas en países tropicales
Las razas de ganado utilizadas para la producción de leche en África se clasifican en cuatro grupos: el Bos taurus sin joroba, que se encuentra principalmente en África occidental; el Bos indicus jorobado (Cebú), taurina templada, que se ha diseminado ampliamente en África; Sanga que es un cruce de B. taurus x B. indicus, criado predominantemente en África oriental y meridional; y la cruz Sanga x Cebú conocida como Zenga (16, 17). La raza de ganado dominante criada en África es el ganado cebú, que está efectivamente adaptado a los climas tropicales y es tolerante a temperaturas extremas y HS. Además, la raza es altamente resistente a enfermedades y parásitos como garrapatas y tiene bajos requerimientos nutricionales (18). Sin embargo, la raza tiene un bajo potencial para la producción de leche.
En el África subsahariana (SSA), las razas comúnmente utilizadas para la producción lechera son principalmente cruces de razas locales y animales importados o semen de Estados Unidos, Canadá, Europa, Australia y Nueva Zelanda. Holstein es la raza exótica preferida para la producción de leche en SSA, seguida de Jersey, Ayrshire y Guernsey (19, 20). Entre las razas de ganado lechero introducidas en África, la raza de ganado lechero Holstein es la raza predominante en la mayoría de los países y es el genotipo más productivo en todo el mundo (21). Además, el genotipo Holstein es más favorecido por los agricultores debido a su capacidad para una alta producción de leche, lo que conduce a altas ganancias en la granja. Los genotipos de Jersey, Ayrshire y Guernsey se consideran productores de leche inferiores y, por lo tanto, son menos preferidos. Por otro lado, se prefieren los jerseys debido al alto contenido de grasa butírica de su leche y a los costos de cría relativamente más bajos debido a su pequeño tamaño (20). Es bien sabido que las razas Holstein y Jersey difieren en sus capacidades para responder a los efectos HS. Muchos estudios han indicado que la raza Jersey es más tolerante a la HS en comparación con el ganado lechero Holstein basado en la disminución de la producción de leche (19).
Debido a la falta de adaptabilidad de las razas de ganado lechero puro al ambiente tropical, varios países del África subsahariana han recurrido al uso de cruces de Holstein-Friesian, Ayrshire, Jersey, con Bos indicus para la producción de leche. El cruce de ganado autóctono con razas lecheras exóticas se ha utilizado para mejorar la producción de leche en regiones cálidas y lograr tasas de reproducción adecuadas. Actualmente, hay un gran número de cruces producidos cruzando los genotipos lecheros exóticos y el ganado cebú local que se utilizan como vacas lecheras (19) en los países subsaharianos. En la mayoría de los países tropicales, el cruzamiento se ha utilizado para combinar rasgos de alta producción de B. taurus (Holstein) con una mayor tolerancia al calor de B. indicus (Cebú). En algunos países, esta estrategia ha llevado al desarrollo de un genotipo de mayor producción y tolerante al calor, como el genotipo Girolando en Brasil (22). En Tailandia, el cruce entre Bos taurus (Holstein, Jersey y otras razas de ganado lechero) y Bos indicus (Sahiwal, Red Sindhi y Thai Native) es predominante y las principales vacas lecheras cruzadas (>87,5% de composición genética Holstein) son cruces de razas Holstein y Sahiwal o nativas tailandesas (23). Los estudios en países tropicales han indicado que los cruces Holstein-Friesian tienen un rendimiento de producción de leche relativamente mejor, mientras que los cruces de Jersey tienen una fertilidad más alta en comparación con el anterior (24). Además de los cruzamientos, para la producción de leche se utilizan genotipos de Bos indicus, por ejemplo, Sahiwal y Gir, que se consideran altamente resistentes al ambiente tropical y ligeramente sensibles a HS (25). La Tabla 1 muestra diferentes razas de ganado lechero criadas en países tropicales para la producción de leche.
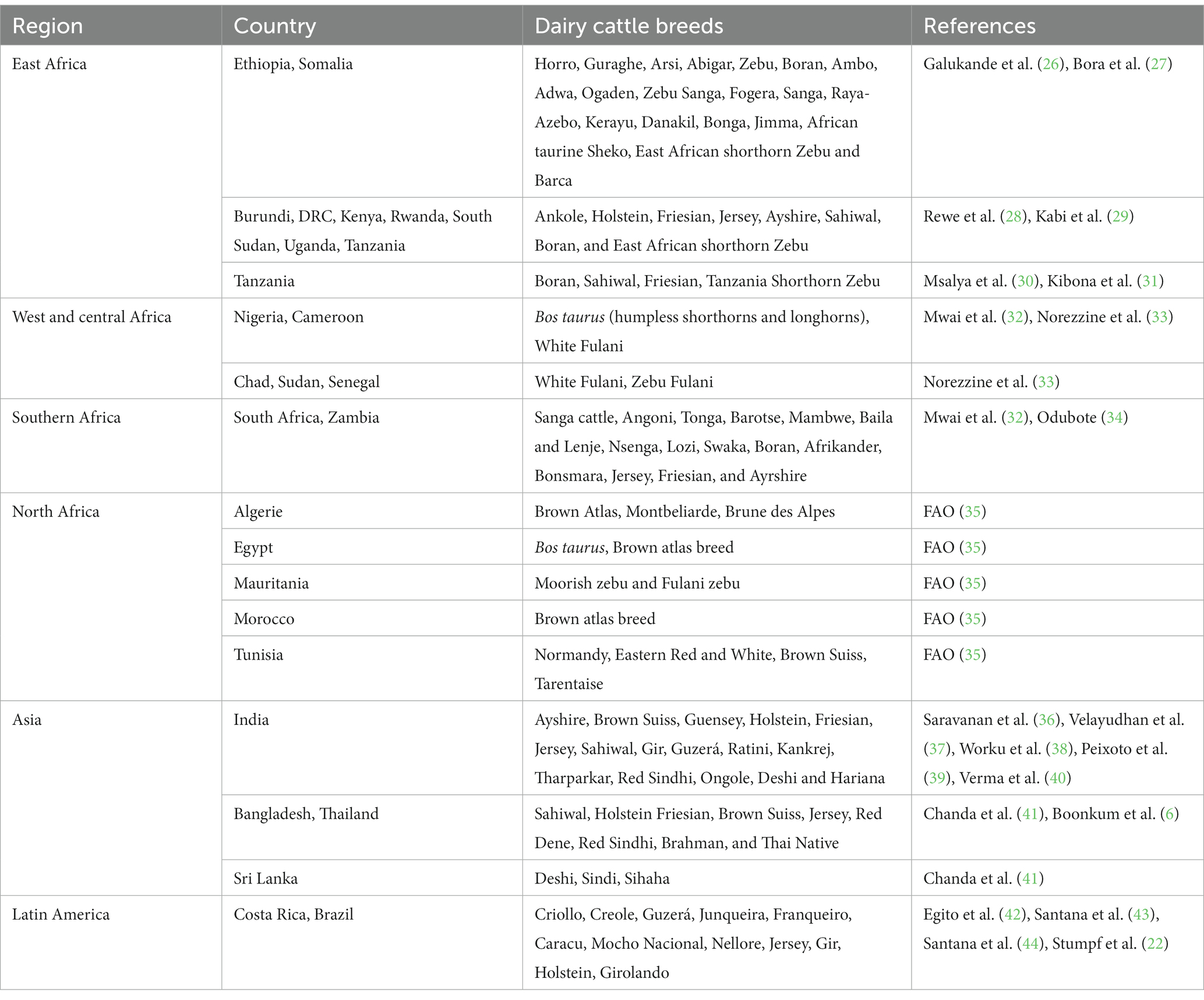 Tabla 1. Razas de ganado lechero criadas en países tropicales.
Tabla 1. Razas de ganado lechero criadas en países tropicales.
3. Indicadores y respuestas fenotípicas al estrés por calor para razas de ganado lechero criadas en los trópicos
Los indicadores del impacto de HS en animales incluyen: (1) parámetros fisiológicos (por ejemplo, frecuencia respiratoria, temperatura rectal, temperatura corporal-central, tasa de sudoración, puntaje de jadeo, entre otros); (2) rasgos de producción de leche; (3) biomarcadores de estrés oxidativo; y (4) genes que influyen en la tolerancia del HS. Además, el THI que combina los efectos de la temperatura ambiente y la humedad relativa, la velocidad del viento y la radiación solar también es un indicador de las condiciones de HS, y puede aproximarse al HS potencial en el ganado lechero. En la revisión anterior del Tribunal se presentó una revisión exhaustiva sobre esos biomarcadores de HS (45). La Tabla 2 muestra indicadores de SA en ganado lechero y los métodos utilizados para medirlos.
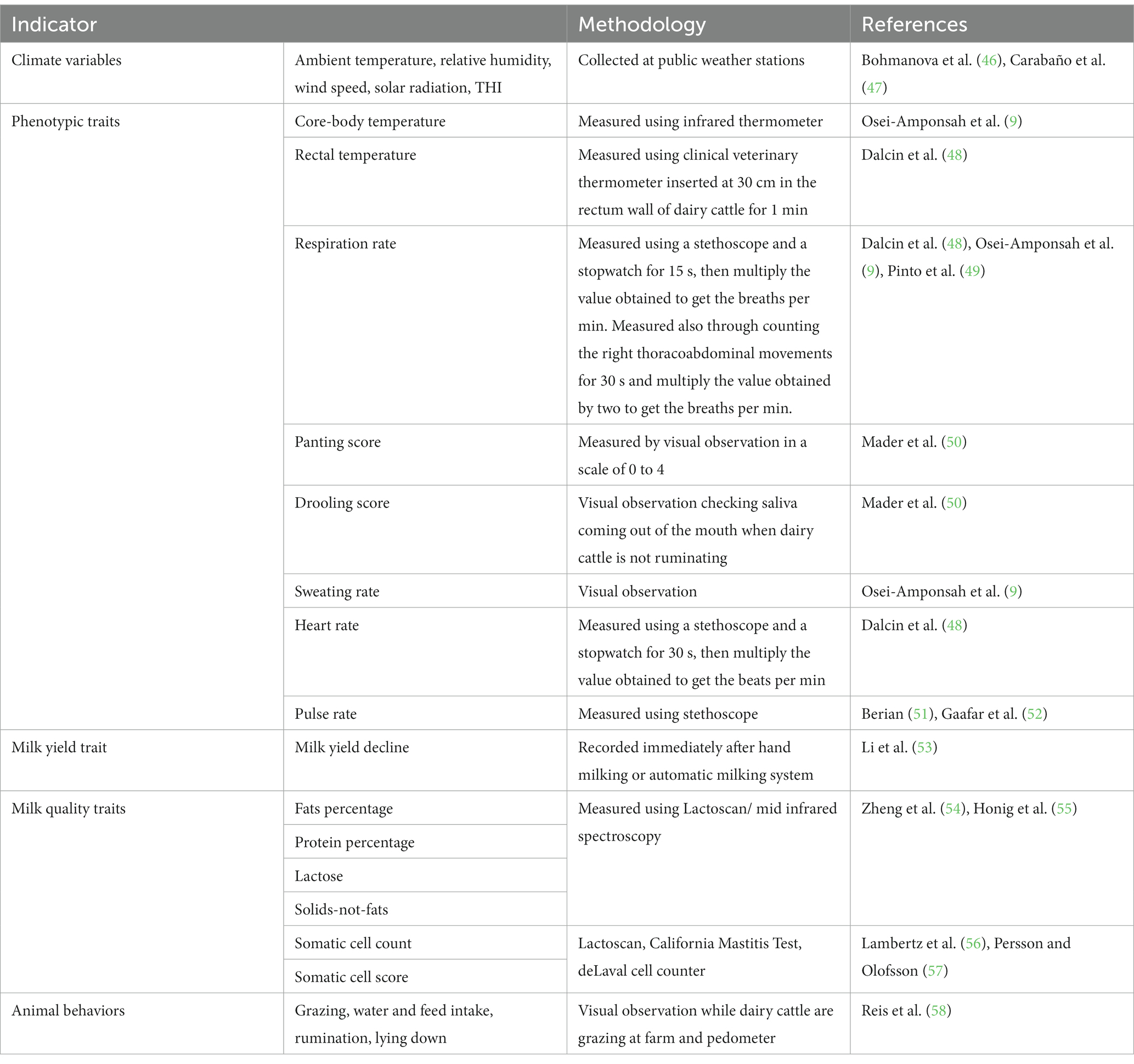 Tabla 2. Indicadores de estrés por calor en ganado lechero.
Tabla 2. Indicadores de estrés por calor en ganado lechero.
El ganado lechero ha desarrollado tres mecanismos de afrontamiento, es decir, aclimatación, aclimatación y adaptación para reducir los impactos de HS en sus sistemas biológicos (59, 60). La aclimatación es la respuesta fenotípica coordinada desarrollada por el ganado lechero a un factor estresante ambiental particular, mientras que la aclimatación es una respuesta coordinada a numerosos factores estresantes simultáneos como la temperatura, la humedad, la velocidad del viento y la radiación solar (59, 60). Tanto la aclimatación como la aclimatación actúan para mejorar la aptitud del ganado lechero a las condiciones climáticas cambiantes (59). Por otro lado, la adaptación implica cambios genéticos ya que los efectos negativos de las variables ambientales en un animal persisten durante mucho tiempo (59, 60). Para hacer frente a las condiciones de HS, el ganado lechero pasa mucho tiempo de pie y reduce la actividad para aumentar el área de superficie para la reducción de calor, la pérdida de agua sensible, el área de superficie radiante y el movimiento del aire a través de la convección, conducción, radiación y evaporación (61, 62). El estrés por calor impide que el ganado lechero exprese plenamente sus potenciales genéticos para la producción de leche y afecta la composición de la leche (63). El ganado lechero reacciona a la HS a través de: (1) disminución en la ingesta de alimento, (2) alto consumo de agua, (3) tasa metabólica basal alterada, (4) aumento de la evaporación del agua, (5) cambios fisiológicos, es decir, mayor temperatura corporal, mayor frecuencia respiratoria, frecuencia de jadeo, frecuencia del pulso, frecuencia cardíaca, temperatura rectal y (6) concentración alterada de hormonas en la sangre (45). Una vaca lechera que experimenta HS produce una pequeña cantidad de leche y tiene un alto riesgo de infección por enfermedad (61). En condiciones HS, la disminución en la ingesta de alimento ha sido reconocida como una causa importante de disminución en la producción de leche, ya que se asocia con un estado de balance energético negativo independientemente de la etapa de lactancia (41). La ingesta de alimento en vacas Holstein lactantes comienza a disminuir a temperaturas ambiente de 25-26 ° C y disminuye bruscamente cuando la temperatura es superior a 30 ° C. A 40°C, la ingesta de alimento puede disminuir en un 40% (41).
El estrés por calor se asocia con la reducción de los rendimientos de leche, grasa y proteínas (64-66). Por ejemplo, Negri et al. (67) reportaron pérdidas de producción de leche del 21% en un rebaño comercial de ganado lechero Holstein criado en la parte sur de Brasil debido a HS. En Bangladesh, Chanda et al. (41) indicaron que cuando una vaca Holstein lactante pasa de una temperatura ambiente de 18 a 30 °C, la producción de leche disminuye en alrededor del 15 %, seguida de una disminución del 35 % en el uso eficaz de energía por razones de producción.
El THI se utiliza para cuantificar la carga de calor ambiental en los animales. La producción de leche se asocia negativamente con THI. En Tailandia, Sungkhapreecha et al. (23) encontraron una disminución menor en la producción de leche de aproximadamente 70-80 g por punto de aumento en el valor de THI de 76 para vacas con >93.7% de composición genética Holstein, y una producción de leche 20-40% menor en comparación con otras poblaciones. En este estudio, el ganado lechero cruzado >87.5% de la genética Holstein fueron cruces entre Holstein y Sahiwal o razas nativas tailandesas (23). Además, la producción de leche, la grasa láctea, los sólidos no grasos y los porcentajes de proteínas se reducen en 15, 39.7, 18.9 y 16.9%, respectivamente, cuando una vaca Holstein lactante se transfiere de una temperatura ambiente de 18 a 30 ° C (68). La Tabla 3 presenta información sobre las respuestas fenotípicas del ganado lechero al SA para estudios realizados en países tropicales de Asia, África y América Latina.
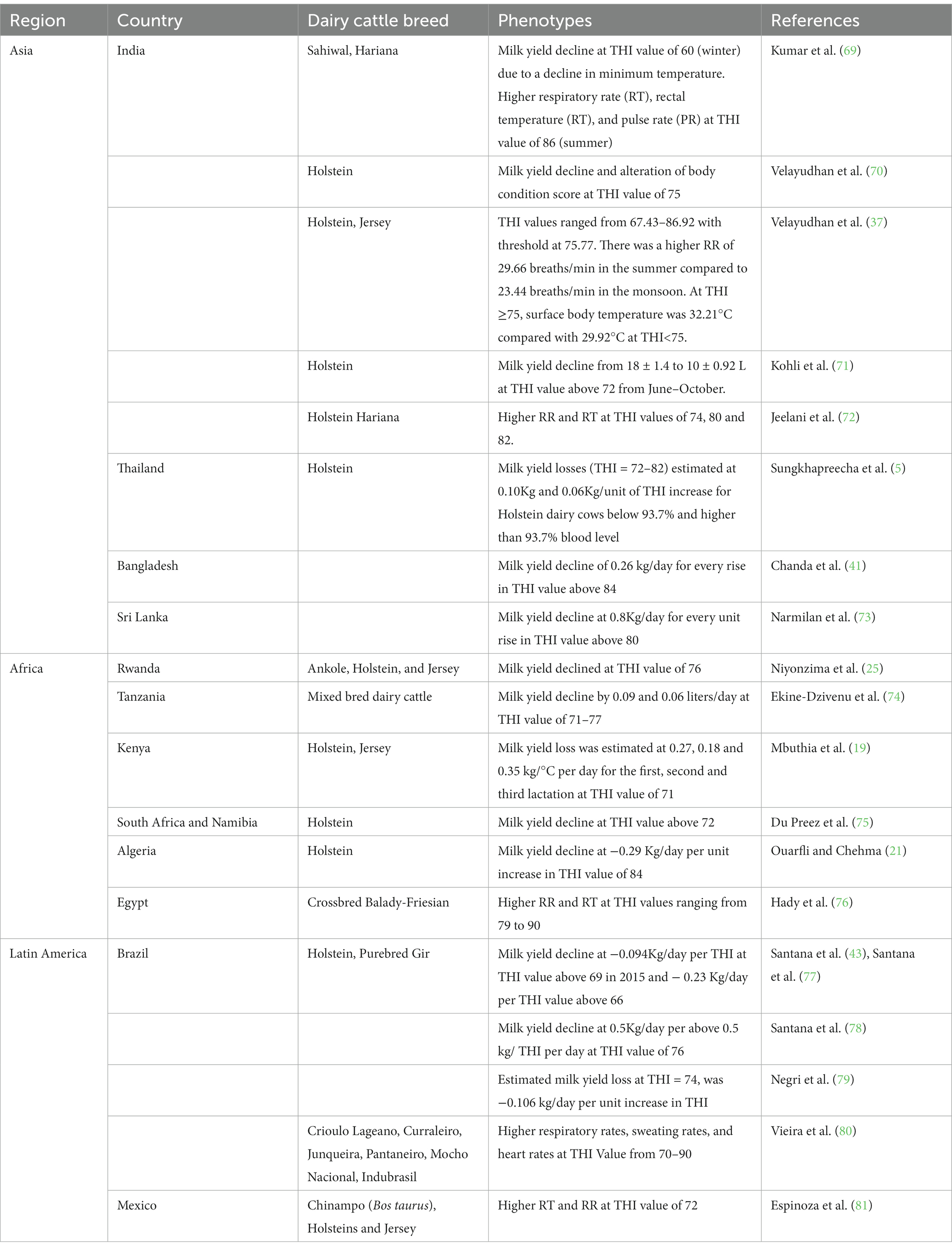 Tabla 3. Respuestas fenotípicas al estrés por calor para razas de ganado lechero criadas en países tropicales.
Tabla 3. Respuestas fenotípicas al estrés por calor para razas de ganado lechero criadas en países tropicales.
4. Efectos del estrés por calor en la leche y los metabolitos sanguíneos de las razas de ganado lechero criado en países tropicales y subtropicales
El estrés por calor induce la modificación de metabolitos en las glándulas mamarias del ganado lechero lactante, junto con la glucólisis, la lactosa, la cetona, el ciclo del ácido tricarboxílico (TCA), el metabolismo de los aminoácidos o nucleótidos, lo que impide el suministro de sustancias para la producción de leche en la glándula mamaria de las vacas Holstein lactantes (82). Por lo tanto, HS altera la síntesis y composición de la leche al alterar el metabolismo de sustancias en los tejidos de las glándulas mamarias de las vacas lecheras lactantes.
Los metabolitos de la leche y la sangre se utilizan para proporcionar información precisa sobre el grado de HS en las vacas lecheras a medida que cambian durante el HS (83). También ayudan en la selección genética de genotipos y animales individuales que son tolerantes al calor (82). Estos metabolitos se identifican en biofluidos animales (por ejemplo, suero, plasma, orina y leche) a través de enfoques metabolómicos. Un enfoque metabolómico se considera como un método potencial para el análisis extensivo de metabolitos de muestras de leche. Tres enfoques analíticos se utilizan ampliamente para estudiar el metaboloma de la leche: espectroscopia de resonancia magnética nuclear (RMN), cromatografía de gases y líquidos-espectrometría de masas (GC-MS / LC-MS). Entre estos enfoques, la RMN puede proporcionar información sobre la asociación directa entre la riqueza de metabolitos y las resonancias.
LC-MS tiene mayor sensibilidad y reproducibilidad, junto con una escala dinámica superior, volviéndose más digna para la investigación y determinación de metabolitos no volátiles y macromoleculares mayores (84). La metabolómica no dirigida que aplica LC-MS es un método confiable para la identificación de una miríada de cambios de metabolitos en una matriz biológica, como sangre, orina, células y tejidos (85). Por otro lado, la resonancia magnética nuclear de protones (RMN) combinada con el análisis estadístico multivariante son muy útiles para el perfil de metabolitos para investigar las variaciones metabólicas en sangre, leche e hígado del ganado lechero estresado por calor (86). Los metabolitos de la sangre y la leche (por ejemplo, glucosa, colesterol, ácidos grasos no esterificados, nitrógeno ureico en sangre, creatinina, entre otros) que se identifican a través de la metabolómica se utilizan generalmente para investigar el grado de HS en el ganado lechero (51, 87). En el estudio realizado por Zheng et al. (54) en la provincia de Heilongjiang en China en vacas lecheras Holstein, siete metabolitos sanguíneos incluyendo L-leucina, L-isoleucina, ácido butanoico y L-valina se asociaron significativamente negativamente con especies reactivas de oxígeno (ROS), mientras que cuatro metabolitos de la leche, es decir, galactinol, undecano, cis-aconitato y etanol se correlacionaron significativamente positivamente con ROS. Los metabolitos anteriores están involucrados en las vías metabólicas asociadas con aminoácidos, carbohidratos, metabolismo de traducción y metabolismo de cofactores y vitaminas. Tanto los metabolitos de la sangre como los de la leche se identificaron mediante GC-MS.
Uso de LC–MS y 1H-NMR, Tian et al. (88) en China, identificaron 53 biomarcadores de diagnóstico potenciales para HS a partir de muestras de leche de ganado lechero Holstein lactante estresado por calor. Entre ellos se incluyeron lactato, piruvato, citrato, creatina, prolina, β-hidroxibutirato, entre otros. Los metabolitos anteriores están involucrados en varias vías bioquímicas, incluyendo carbohidratos, aminoácidos, lípidos y metabolismo derivado de la microbiota intestinal. Yue et al. (83) investigaron los efectos de HS sobre la leche y los metabolitos sanguíneos utilizando resonancia magnética nuclear de protones (1H-RMN) entre los grupos HS y el ganado lechero Holstein lactante no HS en China e identificó ocho metabolitos de muestras de leche y doce metabolitos de muestras de plasma que no estaban significativamente relacionados entre los grupos experimentales estresados por calor y no estresados por calor. Los metabolitos identificados incluyen alanina, glicosa, glutamato, urea, histidina, leucina, lípidos, β-hidroxibutirato, citrato, colina, formiato, piridoxamina, entre otros. Los metabolitos anteriores están involucrados en diferentes vías metabólicas incluyendo proteólisis, glucólisis, gluconeogénesis, lipólisis, galactosa, purina y metabolismo de alanina, entre otros. Hu et al. (89) investigaron los efectos del HS sobre los metabolitos plasmáticos entre el ganado Sanhe y Holstein de Mongolia Interior en China utilizando 1H-RMN combinada con análisis estadísticos multivariantes. En su estudio, se identificaron 26 biomarcadores diagnósticos potenciales para HS (valina, leucina, isoleucina, glutamato, glutamina, creatinina, entre otros) involucrados en las vías metabólicas de aminoácidos, carbohidratos y lípidos. Fan et al. (82) identificaron 33 biomarcadores potenciales de metabolitos que pueden usarse para identificar el grado de HS en ganado lechero Holstein lactante en China. Entre ellos se detectaron glucosa, lactato, β-hidroxibutirato, creatinina, ácido fumárico y glicina como biomarcadores indicativos del grado de HS en ganado lechero lactante utilizando LC-MS/MS. Los metabolitos mencionados anteriormente están involucrados en la glucólisis, cetona, TCA, aminoácidos y nucleótidos de la vía metabólica. En este estudio, se detectaron cambios persuadidos por HS en los metabolitos asociados a la glucólisis para glucosa, lactato, piruvato, fructosa 1,6-bisfosfato, entre otros (82).
En el estudio posterior de Fan et al. (85), también realizado en China en ganado lechero Holstein, HS alteró el metabolismo energético, incluyendo la disminución de las concentraciones de glucosa, lactosa y galactosa-1-fosfato, el aumento de las concentraciones de acetoacetato y β-hidroxibutirato (metabolitos cetónicos), y las alteraciones de los metabolitos asociados a TCA (85). En su estudio, identificaron 34 metabolitos de la leche que están involucrados en las vías metabólicas de aminoácidos, glucólisis, lactosa, cetona, TCA y nucleótidos (85). Las alteraciones en la división de nutrientes pueden afectar el uso de nutrientes y la producción de leche en la glándula mamaria. Los resultados de estos estudios indican que la identificación de biomarcadores de diagnóstico para HS puede contribuir a una mejor comprensión de los mecanismos biológicos de las respuestas a HS (89). Esto es muy importante para la selección genómica del ganado lechero adaptable al calor y los programas de cría destinados a mejorar la tolerancia al calor en vacas lecheras lactantes (82). Además, el análisis de estos metabolitos puede ayudar a interpretar los mecanismos fisiológicos de las enfermedades metabólicas y la lactancia persuadidas por el HS (85).
Por otro lado, la HS altera parámetros sanguíneos como el volumen de células empaquetadas, glóbulos blancos, glóbulos rojos, hemoglobina, glucosa, concentraciones de electrolitos, leucocitos, tamaño de distribución de glóbulos rojos, volumen corpuscular medio y plaquetas, entre otros (22, 72). Por ejemplo, la neutrofilia ocurre en el ganado estresado por el calor debido a la acumulación de neutrófilos de la médula ósea hacia la sangre portal (22). El ancho de los glóbulos rojos (RDW) no se ve afectado por la elevación de THI y los parámetros fisiológicos, lo que implica que el calor no altera la configuración celular y el aumento de THI no influye en los niveles medios de concentración de hemoglobina celular (MCHC) (22). Se ha encontrado que el volumen plaquetario medio (MPV) aumenta con THI y rasgos fisiológicos (además de la frecuencia cardíaca y la puntuación de jadeo) (22). Kekana et al. (90) investigaron los efectos de HS en ganado lechero de Jersey preñado criado en la provincia de Limpopo de Sudáfrica y encontraron que a THI de 75-87, todo el ganado lechero lactante exhibió mayor proteína total, nitrógeno ureico en sangre y creatinina a los 21c y 75ésimo días de producción de leche. En el estudio de Jeelani et al. (72) realizado en la región subtropical de la India en 33 bovinos lecheros cruzados de Bos taurus (Holstein Friesian) x Bos indicus (Hariana / Tharparkar), tanto los recuentos de glóbulos rojos (RBC) como los de glóbulos blancos (WBC) no estaban significativamente relacionados con los determinados en THI de 72-74. Sin embargo, a THI de 76, los recuentos de glóbulos blancos fueron elevados mientras que los recuentos de glóbulos rojos se redujeron. La tendencia general mostró que el aumento en THI se correlaciona con la elevación en los recuentos de glóbulos blancos y la disminución en los recuentos de glóbulos rojos. Gaafar et al. (52) investigaron los efectos de HS en los parámetros sanguíneos de terneros lactantes frisones durante la temporada de invierno y verano en Egipto y encontraron efectos significativos de THI en diferentes parámetros sanguíneos. En general, las tendencias promedio de la concentración de hemoglobina (HGB) y el recuento de glóbulos blancos (WBC) y glóbulos rojos (RBC) disminuyeron significativamente en la estación cálida que en la estación fría como resultado de HS.
Además, el porcentaje de hematocrito (HCT) y el volumen celular medio (MCV), entre otros parámetros sanguíneos, aumentaron significativamente en la estación cálida que en la estación fría como resultado de la HS (22). La disminución en los recuentos de glóbulos blancos observada en la temporada de verano podría ser el resultado de la respuesta del sistema corporal a los estímulos de estrés después de venir de la larga temporada de invierno. En el estudio de Gao et al. (91) realizado en vacas Holstein lactantes en China, HS tendió a disminuir la glucosa plasmática (8%) y los ácidos grasos no esterificados (NEFA) (39,8%), y tendió a aumentar el nitrógeno ureico en sangre (BUN) (14,7%). El ganado lechero estresado por el calor mostró una reducción total de aminoácidos plasmáticos (17,1%), incluidos algunos aminoácidos esenciales (Ile 42,5%, Lys 18,2%) y aminoácidos no esenciales (Ser 10,7%, Arg 25,1%, Gly 20,8%, Cys 27%) (91). Rewe et al. (28) mostraron que en condiciones de HS, el ganado lechero lactante expuesto a una mayor temperatura muestra parámetros sanguíneos elevados que incluyen el recuento de eritrocitos, la concentración total de hemoglobina y el hematocrito, entre otros, junto con el pH y la densidad urinarios, y el contenido de materia seca fecal.
5. Modelos genéticos y parámetros para la tolerancia al calor en razas de ganado lechero criadas en países tropicales
La cuantificación del efecto genético se basa en las estimaciones de la varianza genética aditiva y las relaciones genéticas entre los rasgos en las poblaciones de ganado lechero (38). La estimación de los parámetros genéticos para la tolerancia al calor es crucial para diseñar esquemas de reproducción óptimos y para predecir la respuesta de selección. Para caracterizar esta relación, el umbral de confort y la disminución de la producción después de un umbral particular de THI se han utilizado a través de diferentes modelos genéticos (92). Varios estudios han determinado el efecto genético de HS en los rasgos de producción de leche y han estimado los componentes genéticos de HS en ganado lechero (47, 92–96). Los modelos estadísticos utilizados en los estudios de evaluación genética que aplican registros de producción de leche en el día de prueba incluyen (1) modelos de repetibilidad que consideran la misma relación genética entre los registros de leche de cada día de prueba, (2) modelos de rasgos únicos y rasgos múltiples que aplican cada registro de leche del día de prueba como parámetros diferentes, y (3) modelos de regresión aleatoria (RRM) que tratan una función covariable de los registros de leche repetidos en el día de prueba horas extras (97). En la revisión anterior del Tribunal se hizo una descripción más detallada de estos modelos (45). El RRM tiene el potencial de analizar los registros de leche de cada día de prueba asumiendo que las varianzas genéticas y no genéticas cambian con los días en la leche (DIM), la paridad, la etapa de lactancia al igual que las relaciones genéticas y no genéticas (97). El RRM también tiene en cuenta la variación genética aditiva y ambiental permanente dentro de la lactancia (93).
Los modelos de regresión aleatoria permiten la estimación de componentes de varianza genética y valores de reproducción a lo largo de toda la trayectoria de un descriptor dependiente del tiempo, como los días en la leche (DIM) o dependiente del medio ambiente, como la covariable THI (98). En RRM, cada animal tiene dos efectos genéticos, un efecto constante que corresponde al rendimiento animal en la zona termoneutra, y un efecto HS correspondiente a la tasa de descomposición de la producción de leche en condiciones HS (93). Modelar el efecto de una raza en función del tiempo y el entorno permite la identificación de interacciones GxE a través de diferencias en los componentes de covarianza genética en varias combinaciones de DIM sobre THI (46). Por otro lado, la optimización de los modelos genéticos que describen los efectos genéticos de HS en el ganado lechero en los países tropicales ha sido un desafío debido a la falta de un registro adecuado de los animales. Tradicionalmente, el modelo de línea discontinua (BL) se ha utilizado para describir la respuesta productiva del ganado lechero al aumento de las cargas de calor (47). Este modelo asume que la producción no cambia dentro de la zona termoneutra del ganado lechero donde no se observa respuesta al aumento de la temperatura y que después de un punto de ruptura particular que indica el comienzo del HS, la producción disminuye linealmente (47, 99). Dado que los patrones animales de respuesta a HS difieren según las condiciones climáticas, el tipo de raza (por ejemplo, Bos taurus versus Bos indicus), los sistemas de producción, la etapa de lactancia (lactante versus no lactante) y el estado fisiológico del animal, se considera una norma alternativa de reacción de ajuste RRM y funciones polinómicas de Legendre en lugar del modelo BL (45, 99).
Aunque interpretar biológicamente los parámetros de RRM puede ser difícil, son muy útiles por dos razones. En primer lugar, tienen una mayor flexibilidad para adaptarse a un patrón regular de disminución y, en segundo lugar, permiten la aplicación de la descomposición de la matriz de covarianza genética aditiva para detectar los criterios de selección (92). Tales criterios de selección no están asociados entre sí y podrían apoyar la mejora genética de la tolerancia al calor sin tener efecto sobre el grado de producción (92). Además, en los modelos de día de prueba utilizados para la selección de rasgos de producción de leche, una de las funciones de eigen se ha correlacionado con la persistencia de las lactancias. Esta función eigen también se ha recomendado como criterio de selección de este parámetro para evitar los problemas de correlaciones antagónicas entre el grado de producción y otras medidas de persistencia (92). Los dos enfoques anteriores se denominaron modelo splines (SP) y modelo de norma de reacción de regresión aleatoria (RN) basado en funciones polinómicas de Legendre (LP), y se han aplicado para estimar los parámetros genéticos de tolerancia al calor en razas de ganado lechero criadas en países tropicales. Negri et al. (79) aplicaron RRM con funciones LP de 3Rd e incluyó un modelo de THI en Brasil, y encontró una variación genética considerable para la tolerancia al calor. La Tabla 4 resume la información sobre los estudios que exploraron los modelos genéticos y los parámetros genéticos para la tolerancia al calor en razas de ganado lechero criado en países tropicales.
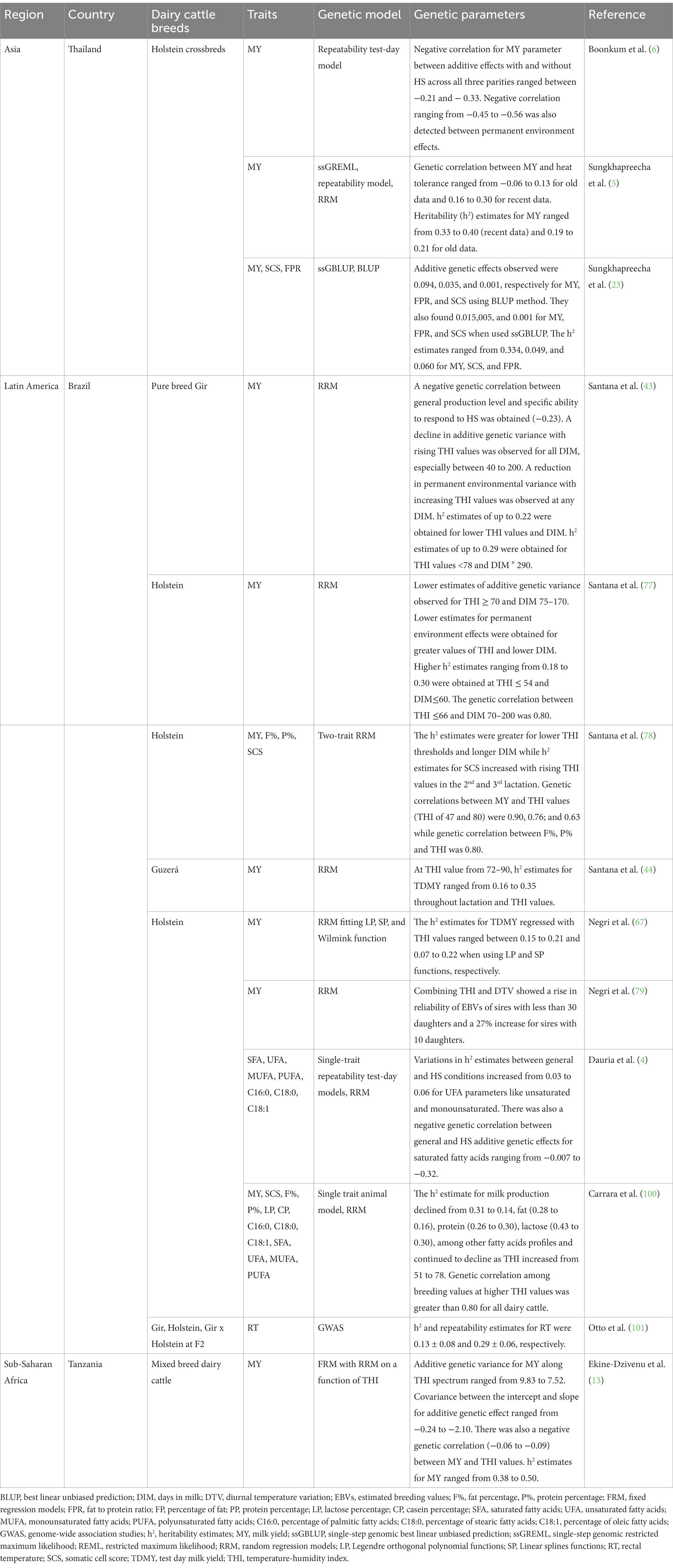 Tabla 4. Parámetros genéticos para la tolerancia al calor en razas de ganado lechero criadas en países tropicales.
Tabla 4. Parámetros genéticos para la tolerancia al calor en razas de ganado lechero criadas en países tropicales.
Boonkum et al. (6) estimaron los efectos genéticos de HS en los rasgos de producción de leche utilizando modelos de prueba de repetibilidad para cruces Holstein en Tailandia. En este estudio, observaron un umbral de THI que oscilaba entre 74 y 82. Sungkhapreecha et al. (5) utilizaron el modelo de máxima verosimilitud genómica restringida de un solo paso (ssGREML) y el modelo de repetibilidad y observaron umbrales THI de 72-82. En un estudio posterior, Sungkhapreecha et al. (23) utilizaron la mejor predicción genómica lineal imparcial genómica de un solo paso (ssGBLUP) y observaron un umbral THI de 76. En este estudio, las correlaciones genéticas entre los efectos genéticos aditivos fueron negativas para todos los rasgos utilizando ambos métodos. Sin embargo, se observaron los efectos genéticos aditivos positivos para SCS para ambos métodos.
En Brasil, Santana et al. (43) estimaron los parámetros genéticos para los rasgos de producción de leche utilizando RRM y observaron un valor umbral de HS de 69 con una pérdida de MY estimada en −0,019 kg/año. En este estudio, se observó que la población de ganado lechero Gir se estaba volviendo más susceptible a HS. También encontraron que se esperan respuestas más pequeñas a la selección para combinaciones de valores THI y DIM. En estudios posteriores, Santana et al. (77) informaron umbrales de THI que van de 54 a 79 y un antagonismo genético entre el rendimiento de la producción de leche y la resiliencia de HS. Este es un aspecto importante para los países tropicales con un clima cálido que tienen como objetivo seleccionar genéticamente una mayor producción de leche, pero también para los animales que son altamente resistentes a la temperatura y humedad extremas. Santana et al. (78) también utilizaron modelos animales de regresión aleatoria de dos rasgos para estimar los parámetros genéticos para MY, porcentaje de grasa (F%), porcentaje de proteína (P%) y SCS y observaron un valor THI de 76 como umbral de HS. En este estudio, el 53% de los registros de leche TD se obtuvieron a valores THI superiores a 66, y otro 14% a valores de THI superiores a 72. Hubo una disminución en la producción de leche estimada en 0,5 kg / día por vaca por THI. También observaron una reducción en MY, SCS, F % y P% en valores THI que oscilan entre 66 y 72. También señalaron que la respuesta a la selección puede variar en función de THI y DIM. En otro estudio, Santana et al. (44) estimaron los parámetros genéticos para los registros MY del ganado Guzera utilizando RRM. En este estudio, observaron umbrales THI que van de 72 a 90. Negri et al. (79) estimaron los componentes genéticos de MY para el ganado Holstein brasileño utilizando RRM que se ajustaba a los polinomios ortogonales de Legendre, splines lineales y función Wilmink. En este estudio, obtuvieron un umbral THI de 84 como umbral de HS con un rendimiento promedio de leche / día de 23.95 ± 1.9 kg / por THI por vaca para THI˃74. Observaron que la variación ambiental aumentaba a medida que disminuía la variación genética aditiva, esto interfería directamente en el2 estimaciones de MI rasgo.
En un estudio posterior, Negri et al. (67) obtuvieron un umbral de THI de 74, asociado con pérdidas de MY estimadas en −0.106 kg/día/THI por vaca. La variación de la temperatura diurna (DTV = 13) se asoció con −0,45 kg/día de pérdidas de MY. Recomendaron que es necesario incluir las variables climáticas en los modelos de evaluación genética para la selección genética de toros de alto rendimiento, ya que muchos toros podrían ser penalizados debido a que sus hijas se ven afectadas por HS. Dauria et al. (4) estimaron los parámetros genéticos de los ácidos grasos de la leche del ganado lechero brasileño Holstein utilizando modelos de día de prueba de repetibilidad de un solo rasgo y RRM. En este estudio, observaron un umbral THI de 68. Carrara et al. (100) estimaron los parámetros genéticos para los rasgos MY y ácidos grasos del ganado lechero Holstein brasileño utilizando un modelo animal de rasgo único y RRM. En este estudio, obtuvieron un umbral THI de 60. Además, observaron una pequeña variación en los componentes de la varianza sobre THI, lo que indica un HS moderado a bajo para el ganado lechero como resultado de las estrategias de mitigación que utilizan ventiladores, rociadores, árboles de sombra y otros dispositivos de enfriamiento aplicados en la granja. Otto et al. (101) estimaron los componentes genéticos de la RT para el ganado lechero cruzado Gir x Holstein utilizando GWAS. En este estudio, obtuvieron un umbral THI de 97 (42°C y 60% HR). En Tanzania, Ekine-Dzivenu et al. (13) estimaron los componentes genéticos de MY para ganado lechero de raza mixta utilizando un modelo de regresión fija con RRM en función de THI. En este estudio, identificaron valores de THI que oscilan entre 67-76. Encontraron que la selección genética para una mayor producción de leche sin tener en cuenta la tolerancia al calor conducirá a un deterioro de la tolerancia al calor en esta población de ganado lechero. En conclusión, las funciones de ajuste RN y LP de RRM son más adecuadas que otros modelos declarados durante la evaluación genética para la producción de leche y los rasgos de composición sobre diferentes umbrales de THI en estudios de tolerancia al calor. Las funciones RN son más convenientes para los descriptores ambientales continuos como THI ya que ayudan en la distinción de los individuos que están más o menos afectados por las variaciones ambientales (100). Además, a través de las funciones RN, es posible obtener dos componentes genéticos para cada individuo, es decir, capacidad de producción general y capacidad específica para responder a HS (77).
6. Genes responsables de la tolerancia al calor en razas de ganado lechero criadas en países tropicales
Comprender los mecanismos de adaptación climática y tolerancia al calor de las razas de ganado lechero criado en países tropicales es vital para la optimización de los sistemas de cría y la conservación de los recursos genéticos (102). La variación genética en la tolerancia al calor a nivel fisiológico y celular entre las razas Bos indicus y Bos taurus criadas en países tropicales se ha estimado en varios estudios (103). La tolerancia al calor se puede identificar en función de la expresión o activación de biomarcadores biológicos particulares, incluido el factor de transcripción de choque térmico (HSF), las proteínas de choque térmico (HSP70, HSP90 y HSP27) y el gen Slick Hair (36). HSF y HSP conocidos como chaperones moleculares, son genes cruciales asociados con la tolerancia al calor (37). Desempeñan un importante papel citoprotector durante el estrés celular al evitar la formación de proteínas no funcionales, mantener el plegamiento de proteínas y prevenir la agregación de proteínas que resultan en homeostasis de proteínas durante la respuesta celular a HS (37, 104). Las HSP también activan los mecanismos de respuesta inmune durante la HS (37). El estrés por calor causa un plegamiento incorrecto de las proteínas a nivel celular y una célula no realiza la replicación del ADN, la transcripción, la síntesis de polipéptidos y el transporte de materiales (105). Dados esos efectos HS, las HSP se activan y reducen la acumulación de las proteínas desnaturalizadas en la célula y mejoran su capacidad de supervivencia y capacidad para superar el estrés oxidativo y el estrés térmico (105). Además, HSP70 actúan como acompañantes moleculares y desempeñan un papel crucial en la termotolerancia celular, la apoptosis, la modulación inmune y la tolerancia al calor (37, 105). Las HSP70 funcionan como acompañantes inducibles y proporcionan una segunda ventana de protección al tiempo que estabilizan la conformación nativa de las proteínas y mantienen la capacidad de supervivencia celular durante las condiciones de HS (37, 106). Una inducción temprana de HSP70 se asocia con el desarrollo de tolerancia al calor en el ganado lechero (37). Además, HSP40, que alberga los genes DNAJA1, DNAJB1 y DNAJA2, regula la actividad ATPasa de HSP70 a través de la interacción con el dominio J de las proteínas HSP70 (104). Además, HSP40 y HSP90 funcionan como co-acompañantes de HSP70 y juegan un papel importante en la restauración de la conformación de proteínas después del choque térmico (106).
Los estudios han indicado que en todo el mundo existen cinco grupos continentales de ganado, a saber, taurina europea, taurina euroasiática, taurina asiática oriental, indicina china e indicina indicia india (107). La tolerancia al calor, la resistencia a los ectoparásitos, el bajo costo de mantenimiento y la baja susceptibilidad a los parásitos gastrointestinales son rasgos fisiológicos importantes de la indicina sobre la taurina (108, 109). Los rasgos fisiológicos anteriores ayudan a las razas Bos indicus a crecer en climas tropicales y subtropicales de África, el sudeste asiático, Brasil, el norte de Australia, el sur de China y partes de los Estados Unidos (107, 108). Las razas taurinas que han aumentado la tasa metabólica y los requisitos de alimentación se crían principalmente en Europa, América del Norte y Australia (108).
Para comprender mejor la variación en la respuesta de HS entre Holstein-Friesian (taurina) y Sahiwal (indicina) en la India, Kishore et al. (106) investigaron el perfil temporal de los genes candidatos HSP40, HSP60, HSP70 y HSP90 utilizando qRT-PCR a 42 ° C. En su estudio, observaron que Holstein-Friesian fue el más afectado por el choque térmico que Sahiwal, lo que indica que este último es altamente adaptable a las condiciones de HS. En otro estudio realizado en Brasil, Otto et al. (101) utilizaron GWAS a través de alelos de raza de origen (BOA) para comparar la respuesta HS entre las razas lecheras Holstein y Gir. En su estudio, se observó que la proporción de animales con alelos HH y GG fue de 2.9 y 49.9%, respectivamente, para una RT más alta en Holstein y Gir, lo que indica que el ganado lechero Holstein se vio más afectado por HS que los animales Gir (101).
En Nigeria, Morenikeji et al. (110) utilizaron el enfoque transcriptómico de ARN para comparar la respuesta de HS entre las razas de ganado lechero indicina (fulani blanco y N’Dama) y taurina (Angus y Holstein) e identificaron seis genes (ASIP, MC1R, TYRP1, TYRP2, TYR y ALOX12E) para ser responsables de la tolerancia al calor. Estos seis genes fueron regulados al alza en las razas indicinas que en las taurinas. En particular, se encontró que el gen ALOX12E estaba asociado con la retención de agua en indicine para evitar la deshidratación, un rasgo adaptativo importante para la supervivencia de fulani blanco y N’Dama en climas tropicales (110). Además, Onasanya et al. (111) indicaron que los genes HSP70 y HSP90 identificados en las razas de cebú de África occidental son genes candidatos termorreguladores que son menos sensibles a señales térmicas como las altas temperaturas. Estos genes de proteínas de choque térmico son predominantes en el cebú de África occidental y oriental, como el fulani blanco, N’Dama, Sahiwal, Boran y Ankole, que constituyen más del 90% de la población bovina en África (111).
Los análisis de firmas de selección tienen el potencial de identificar regiones genómicas específicas y genes candidatos asociados con rasgos adaptativos o productivos de diferentes razas de ganado lechero criadas en ambientes HS de países tropicales (40, 112, 113). Utilizando firmas de selección y variaciones en el número de copias para diferenciar las razas Bos taurus y Bos indicus, Jang et al. (114) identificaron 313 genes candidatos bajo selección y adaptación. Se identificó un mayor número de copias del gen HMGA2 en Bos indicus que en las razas Bos taurus. También identificaron el gen DNAJC18 en la deleción específica de indicina que en la taurina. Se ha demostrado que el gen candidato DNAJC18 desempeña un papel importante en la adaptación tropical y la tolerancia al calor del cebú de cuerno corto de África oriental (114). En la frontera entre China y Myanmar, Sun et al. (107) realizaron un estudio sobre el ganado jorobado Lincang e identificaron MED16, DNAJC8, HSPA4, FILIP1L, HELB, BCL2L1 y TPX2 como genes candidatos responsables de la tolerancia al calor y la resistencia al parásito. En la India, Saravanan et al. (36) estudiaron las firmas de selección entre razas taurinas y razas indicinas e identificaron genes candidatos responsables de la respuesta inmune y la tolerancia al calor. Entre los genes identificados se incluyen NCR3 (en Gir), ARID5A (en Ongole), HIST1H2BN (en Sahiwal), BEFB4 y DEFB7 (en Tharparkar) para la respuesta inmune. Los genes responsables de la tolerancia al calor incluyen HSPA1L y HSPA1B en Gir, DNAJB4 en Ongole y GRXCR1 en Tharparkar (36).
Genes como OLA1, SP9, HSPA12A se asociaron con tolerancia al calor en todas las razas de ganado Indicine (Gir, Hariana, Kankrej, Ongole, Red Sindhi, Sahiwal y Tharparkar), pero ninguno de estos genes se identificó en razas taurinas (36). La mayoría de los genes candidatos identificados en las razas taurinas estaban relacionados con los rasgos de producción de leche e incluían CACNA2D1, ADARB2, WDR37 y LARP4B (Ayshire), KCNIP4, SLIT2, ADAP2 y LTF (Brown Suiss), SYN3, TIMP3, RFK, ACACA, HNF1B y SCAP (Guernsey), ADARB2, WDR37, LARP4B, ZMYND11, PRNP y CDH4 en Holstein Friesian, y LPHN2, ADARB2, WDR37 y LARP4B en Jersey (36). Los genes que se encontraron asociados con la tolerancia al calor en las razas de ganado Indicine se pueden utilizar para la detección molecular de razas de ganado lechero con potencialidades de tolerancia al calor. Además, los genes candidatos identificados en diversos estudios pueden servir de base para explorar el uso de razas indicinas en esquemas de cruzamiento con el fin de transferir sus capacidades de adaptación y rusticidad a las razas taurinas (115). La Tabla 5 proporciona ejemplos de genes candidatos responsables de la tolerancia al calor que se han identificado en diferentes razas de ganado lechero criadas en países tropicales.
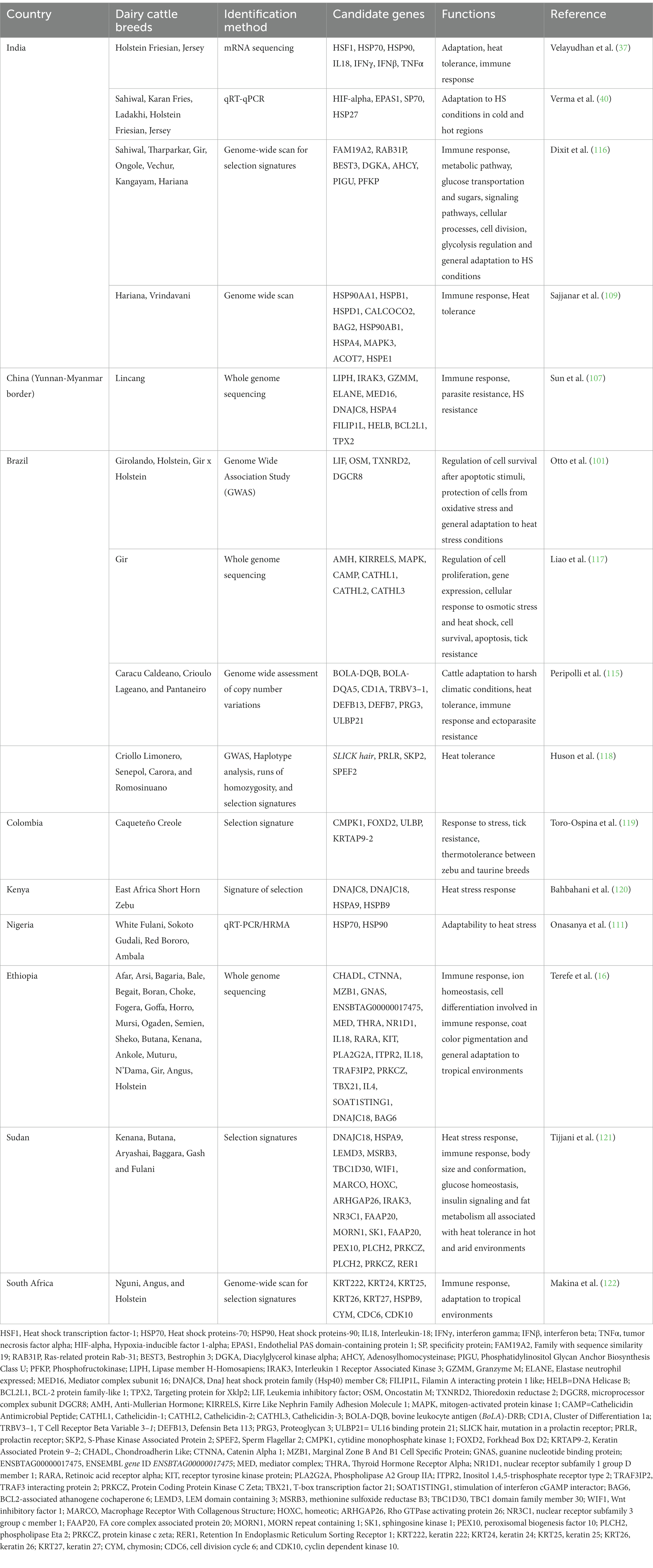 Tabla 5. Genes candidatos responsables de la tolerancia al calor en razas de ganado lechero criadas en países tropicales.
Tabla 5. Genes candidatos responsables de la tolerancia al calor en razas de ganado lechero criadas en países tropicales.7. Estrategias de mitigación del estrés por calor en ganado lechero en los trópicos
El estrés por calor causa un efecto perjudicial sustancial en la industria láctea, incluida la reducción del crecimiento, la eficiencia reproductiva y la producción y calidad de la leche. Por lo tanto, es necesario desarrollar estrategias para mitigar los efectos del SA. Sin embargo, no existe una estrategia única que se utilice para mitigar los impactos negativos del SA en la ganadería lechera. Las siguientes estrategias pueden reducir los efectos de HS en el ganado lechero: (1) modificación física del medio ambiente (sombreado, enfriamiento), (2) desarrollo genético de animales tolerantes al calor y (3) mejores estrategias de manejo de la alimentación (65, 123–126). Una combinación de las prácticas anteriores puede mejorar el rendimiento de producción del ganado lechero lactante en climas cálidos y húmedos, especialmente en países tropicales y subtropicales. Se ha indicado que las tecnologías de refrigeración tienen un alto impacto positivo en la eliminación de los efectos adversos del THI alto en vacas lecheras (1). Sin embargo, esas estrategias no alivian completamente los efectos adversos de la HS debido a factores relacionados con los animales, como la genética, el pelaje y el calor metabólico generado (127). Como tal, la identificación del ganado lechero resistente a una alta carga de calor y la utilización de estrategias de mitigación adecuadas pueden conducir a mejoras en el bienestar animal y la rentabilidad (128).
7.1. Modificación física del medio ambiente
Las estrategias comunes para mejorar la comodidad del ganado lechero lactante y mantener una producción óptima de leche durante los períodos de SA se ejecutan principalmente mediante la modificación física del entorno de producción. La modificación del medio ambiente implica tecnologías de reducción del calor que alteran las condiciones ambientales para evitar el SA, incluidos nebulizadores, almohadillas de enfriamiento y estrategias que mejoran el intercambio de calor entre el ganado lactante y el medio ambiente como los remojadores (129). Los sistemas de gestión como el riego, la sombra, los túneles, los rociadores y los graneros pueden mejorar la ventilación pasiva, mejorar la pérdida de calor corporal, disminuir la temperatura corporal y mejorar la ingesta de materia seca (DMI) (90, 126, 128). La ingesta de materia seca generalmente disminuye durante las condiciones de clima cálido, por lo que se debe aumentar la densidad de nutrientes de la dieta (126, 130, 131). Proporcionar sombra al ganado lechero cuando las temperaturas están por encima de su nivel de confort térmico puede ayudar a los productores a reducir el consumo de agua de las vacas. Esto es significativo porque la industria láctea tiene la segunda huella hídrica más alta en la producción animal (132). La eficacia de estas tecnologías para enfriar y alterar las condiciones térmicas en las vacas puede variar (46). Para mitigar el efecto adverso del SA en los sistemas de producción lechera de los pequeños agricultores, es necesaria una combinación de medidas de protección a corto y largo plazo. A corto plazo, el objetivo debe ser motivar a los pequeños agricultores a establecer estrategias de gestión del rebaño y cría de animales que protejan eficientemente a los animales del calor extremo. Estas estrategias de mitigación del calor podrían incorporar el uso de sombras de árboles, sombras de heno o paja o techos con láminas estriadas decoradas con revestimiento deliberado en la parte superior para devolver la luz solar y la utilización de techos elevados en los edificios para disminuir la temperatura asimilada en el interior. Otras medidas incluyen el suministro de suficiente alimento y agua de manera consistente y la mejora de la utilización de concentrado con un forraje mínimo dentro de los límites de mantener una función ruminal eficiente durante el clima cálido (74).
El enfriamiento evaporativo se ha utilizado con éxito como un sistema de reducción de calor durante un período prolongado del año en todo el suroeste de los Estados Unidos, pero se aplica menos en el sureste, donde hay bajas temperaturas y alta humedad (46). Por otro lado, los sistemas de refrigeración conductiva tienen el potencial de preservar el agua, se comparan mejor higiénicamente con las tecnologías de refrigeración basadas en la evaporación y proporcionan un alivio HS exitoso en comparación con las tecnologías basadas en ventiladores (133).
La provisión de sombra a los animales expuestos al sol es una forma exitosa de prevenir la radiación solar inmediata de onda corta (134) y minimiza los problemas de quemaduras solares, particularmente en animales con piel menos pigmentada (135). El ganado lechero lactante con acceso a la sombra tiene temperaturas rectales más bajas porque la carga de calor solar reducida les permite utilizar mejor las rutas sensibles de pérdida de calor (63). Además, la producción y composición de la leche mejoran cuando se proporciona sombra (134). Anderson et al. (63) informaron que el ganado lechero bajo sombra y enfriado por ventiladores y pulverización produjo 2 kg/día más de leche que el ganado lechero bajo sombra sola. La provisión de sombra y rociadores da como resultado un aumento del peso corporal, mayores rendimientos de leche, grasa láctea, proteínas, lactosa y menor temperatura rectal y frecuencia respiratoria (55, 132, 136, 137). Reis et al. (58) indicaron que la sombra de los árboles ayudó a reducir la HS en un 7,3%. Mojar el ganado lechero en ríos y estanques también es una forma exitosa de disipar el estrés térmico por evaporación y convección (135). La utilización de sombra natural o artificial dentro del sistema de producción de pastos salva al ganado de la radiación solar extrema y cambia su balance de radiación (58). Los estudios han demostrado que cuando un ganado lechero recibe sombra, aumenta el tiempo de rumia, aumenta la producción y la calidad de la leche y disminuye la TCC en comparación con un ganado lechero en un refugio sin sombra (61). En Brasil, Reis et al. (58) evaluaron las respuestas conductuales y fisiológicas de las razas de ganado lechero Gyr y Girolando criadas en ambientes sombreados y no sombreados. En este estudio, a THI de 79 a 83, el tiempo de rumia del ganado lechero Girolando y Gyr fue 1,7 veces mayor en el ganado lechero criado bajo sombra. El tiempo de reposo fue un 23% más largo en el ganado lechero expuesto al sol. El ganado lechero Girolando tenía un PS 35% más alto que el ganado lechero Gyr, independientemente del medio ambiente. El PS aumentó 2,5 veces durante la tarde en comparación con la mañana. Sin embargo, el efecto de este alto THI en el ganado lechero Girolando y Gyr se redujo por sombra de 2.5 a 5.2% (58).
7.2. Estrategias nutricionales y de alimentación para minimizar los efectos del estrés por calor en los animales
Mantener una ingesta suficiente de alimento es un método importante para reducir la disminución de la producción de leche. Entre las estrategias para aumentar la ingesta de nutrientes en la dieta se encuentra la provisión de alimentos de alta calidad, la alimentación de una gran cantidad de granos y la utilización de grasas suplementarias (138). El suministro de almidón degradable, como los granos de cereales (por ejemplo, grano de trigo y grano de maíz) reduce la cantidad de calor producido durante la digestión (138). En Australia, Garner et al. (138) proporcionaron dietas compuestas de grano de trigo, grano de maíz y heno alfa alfa a ganado lechero Holstein-Friesian expuesto a una cámara climática controlada de 33 ° C y 50% de humedad relativa. En este estudio, el ganado lechero al que se le ofrecieron granos de maíz tuvo un menor RR y una mayor ingesta de alimento. La disminución de la proporción de forraje a concentrado también puede aumentar la ración que se absorbe en mayor medida. Sin embargo, la alimentación excesiva de concentrado puede provocar trastornos del rumen, incluida la acidosis, y las vacas pueden dejar de alimentarse (130). La inclusión de bicarbonato de sodio en la dieta puede ayudar al rumen a acomodar una mayor cantidad de concentrado. Otros aditivos para piensos que se han utilizado en condiciones de HS para mantener la salud del rumen a partir de cambios en la dieta son las levaduras, que mejoran la degradación de la fibra, y los cultivos de hongos y la niacina, que mejora la utilización de energía (131). El suministro de cultivos de levadura como saccharomyces cerevisiae y enzimas fibrolíticas, por ejemplo, celulasas y hemicelulasas producidas por hongos y bacterias estabilizan el pH del rumen, mejoran la digestión de la fibra y aumentan el flujo de nutrientes postruminales, lo que conduce a una mejor tolerancia al calor en las vacas lecheras (130). Las modificaciones ambientales y de alimentación para aumentar la ingesta de alimento son específicas de cada sistema de producción y dependen de su valor adicional para mejorar el rendimiento, la aptitud y el bienestar de los animales en comparación con sus costos (65). Se recomienda elegir y alimentar alimentos frescos, sabrosos y de alta calidad en la mayor medida posible y proporcionar ingredientes que tengan una alta digestibilidad para el ganado para reducir el incremento de calor a través de la utilización de nutrientes en el animal (139).
7.3. Selección genética para el ganado lechero tolerante al calor
El desarrollo genético de un genotipo de ganado lechero tolerante al calor, el mantenimiento de una mayor producción de leche y el rendimiento reproductivo en entornos desafiantes ha sido un deseo de toda la vida de los investigadores y criadores de ganado (140, 141). La reducción de los efectos del SA en las razas de ganado lechero criadas en sistemas de producción de bajos insumos, especialmente en los países subsaharianos, debe basarse en la selección genética de animales que son termotolerantes y tienen capacidad genética para sobrevivir en condiciones estresantes. Esta estrategia es más adecuada y rentable para mejorar la producción de ganado lechero en ambientes tropicales (4). Además, los logros obtenidos a través de la selección genética son acumulativos y permanentes (96). Se consideran las siguientes estrategias al seleccionar genéticamente o criar una raza de ganado lechero termotolerante: (1) elección de raza o cruce para criar en la granja, ya que algunos genotipos son más tolerantes al calor que otros, (2) introgresión de genes que están asociados con la termotolerancia, y (3) uso de criterios de selección relacionados con la tolerancia al calor (99).
La identificación del ganado lechero resistente a la alta carga de calor y su uso en programas de mejora genética puede conducir a mejoras en el bienestar animal y la rentabilidad de las empresas lecheras (128). Un animal tolerante al calor es aquel que mantiene el equilibrio térmico durante las condiciones de SA (125). En los países tropicales, es mejor elegir genotipos de ganado lechero con composición genética de Bos indicus debido a sus capacidades de adaptación relativamente altas a HS. Se recomienda que la selección genética para la tolerancia al calor en los países tropicales se base no solo en la capacidad del ganado lechero para regular la temperatura corporal, sino también en la sostenibilidad de los rasgos de producción de leche durante las condiciones de SA (142). Una estrategia alternativa para la mejora genética de la tolerancia al calor con respecto a los rasgos de producción de leche es seleccionar una baja disminución de la producción de leche en lugar de la regulación de la temperatura corporal durante las condiciones de SA (142).
Los genotipos del ganado lechero tropical y subtropical tienen una mayor capacidad de adaptación a los factores estresantes climáticos que las razas templadas exóticas. Rasgos como la tolerancia al calor, la adaptabilidad a los cambios estacionales, la falta de alimento, la tolerancia a la infestación de garrapatas y los trastornos metabólicos, valen la pena en climas tropicales. Los rasgos mencionados anteriormente se observan generalmente en los genotipos del ganado lechero tropical, y deben introducirse en razas templadas mantenidas en los trópicos que tienen un rendimiento de producción más alto que los genotipos indígenas (143). El ganado cebú (Bos indicus), por ejemplo, posee genes de tolerancia al calor y el ganado derivado de genotipos cebú es altamente eficiente para controlar la temperatura corporal en respuesta al HS en comparación con el ganado de genotipos de Bos taurus de origen europeo (125). El ganado cebú (Bos indicus) es ampliamente reconocido como adaptado a condiciones climáticas tropicales y subtropicales que son prohibitivas para el ganado Bos taurus (144, 145).
Osei-Amponsah et al. (125) afirmaron que las modificaciones en la genética del rebaño y la selección de genotipos con composición genética de Bos indicus pueden ser adecuadas para desarrollar razas de ganado lechero termotolerantes. Por ejemplo, India, que es el mayor productor de leche del mundo, se ha embarcado en esquemas de cruzamiento para mejorar la producción de leche a partir de genotipos locales y el 90% de la población de ganado en el país es Bos indicus. Actualmente, el país tiene 22,1 millones de bovinos cruzados, compuesta por cruces de Holstein Friesian y Jersey (144). Otros estudios han demostrado que la temperatura rectal de un ganado lechero estresado por el calor es hereditaria, y la selección de este rasgo ofrece la posibilidad de mejorar la termotolerancia del ganado lechero (15). Las estimaciones de heredabilidad de la temperatura rectal en Holstein US varían de 0.13 a 0.17. Este nivel de estimación de heredabilidad implica que la selección genética para la respuesta del SA es factible (146).
Una estrategia genética alternativa para mitigar los efectos de HS sin cruzamiento es la tecnología de edición del genoma. Los polimorfismos asociados con la tolerancia al calor se pueden transferir de un genotipo a otro. El haplotipo SLICK detectado en genotipos tolerantes al calor criados en América Central y del Sur conduce a que los animales tengan el pelo corto y fino y con un mayor control de la temperatura corporal (147). La introgresión del gen del haplotipo SLICK a partir de genotipos de ganado que son genéticamente altamente resistentes a la HS puede conferir termotolerancia a genotipos lácteos templados que se adaptan a climas fríos (148). El haplotipo SLICK detectado inicialmente en el ganado Senepol ahora ha sido introducido en Holstein para mejorar su termotolerancia (149). Las vacas Holstein de haplotipo SLICK son altamente efectivas para controlar la temperatura corporal y no muestran una disminución notable en la producción de leche durante la HS (125, 148).
Los avances en el uso de datos de secuencia de tecnologías de fenómica, genómica y transcriptómica, junto con los hallazgos de los estudios de expresión génica, han llevado a la disponibilidad de valores genómicos precisos de reproducción que se pueden utilizar en la selección en todas las razas y generaciones. Los datos de la expresión génica o los estudios de asociación de genoma completo (GWAS) se pueden aplicar para mejorar aún más la precisión de la selección. Los GWAS se han aplicado con frecuencia para detectar regiones del genoma que tienen un impacto potencial en un rasgo de valor económico en la producción lechera (125). Utilizando GWAS a través de análisis condicionales para determinar con precisión la asociación subyacente a la tolerancia al calor y el grado de producción de leche en el ganado Holstein australiano, Cheruiyot et al. (11) identificaron posibles variantes candidatas y genes asociados con el sistema neuronal (ITPR1 e ITPR2) y la base de interacción ligando-receptor neuroactivo para la tolerancia al calor (NPFFR2 y CALCR). Esto ha dado un discernimiento innovador que puede ayudar a desarrollar programas de mejoramiento para mitigar HS. Se han realizado estudios GWAS para la temperatura rectal durante las condiciones de HS en vacas Holstein lactantes para detectar SNP conectados con genes termotolerantes y esto podría ayudar a la selección genética para la adaptación a HS (125). Además, utilizando la mejor metodología de predicción genómica lineal imparcial de un solo paso para detectar regiones genómicas y genes candidatos reconocidos involucrados en la producción de leche durante condiciones termoneutras y HS, Sigdel et al. (15) identificaron genes candidatos que aclaran las mayores variaciones genéticas aditivas para la producción de leche durante condiciones HS. En su estudio, se encontró que un sitio genómico en BTA15 estaba muy correlacionado con el grado de producción de leche durante HS para las tres paridades. Este sitio contiene genes candidatos incluyendo PEX16 y MAPK8IP1 que están involucrados en la respuesta celular a HS. El gen MAPK8IP1 está implicado en la regulación de la respuesta celular al choque térmico, lo que mejora secuencialmente la actividad de transcripción de diversos genes sensibles a HS que regulan múltiples funciones, como la supervivencia celular y la proliferación celular.
La selección genómica (GS) tiene una ventaja comparativa, ya que permite seleccionar toros jóvenes (y novillas) en función de sus valores genéticos estimados genómicos de termotolerancia (GEBV), además de otros parámetros (150). La selección genómica aplica marcadores de ADN de todo el genoma para detectar los efectos de varias mutaciones que causan alteración en un rasgo complejo como la tolerancia al calor, y permite seleccionar toros y novillas jóvenes en función de su GEBV y, por lo tanto, mejorar la ganancia genética (8). Esta estrategia debería promoverse en muchos países tropicales, ya que ha tenido éxito en los países desarrollados. Dado que los toros lecheros se seleccionan cada vez más para la cría sobre la base del GEBV (para una variedad de rasgos) en lugar de las pruebas de progenie, es necesario establecer el GEBV para la tolerancia al calor si este rasgo debe tenerse en cuenta en las decisiones de selección (150). La tolerancia al calor GEBV se ha estimado con una precisión que varía de 0,42 a 0,61 mediante la aplicación de marcadores SNP de alta densidad (125). En 2017, por primera vez, GEBV para tolerancia al calor se lanzó en Australia (125). Los valores genómicos estimados de mejoramiento para la tolerancia al calor son confiables y el uso de estos valores en el mejoramiento selectivo puede mejorar la ganancia genética y aumentar la rentabilidad en la granja. Sobre la base del GEBV, es posible identificar animales termotolerantes que son más adecuados para hacer frente a las condiciones climáticas actuales y futuras (150).
Ravagnolo y Misztal (96) recomendaron estrategias de selección genética para identificar animales tolerantes al calor utilizando los modelos de regresión aleatoria (RRM) que cuantifican el grado de HS basado en la información climática del día de la prueba. Este enfoque ofrece la posibilidad de identificar los efectos de las interacciones genotipo-por ambiente. Las firmas de selección, como los genes que regulan la anemia y el comportamiento alimentario en el ganado N’Dama tripanotolerante, el color del pelaje y el desarrollo del cuerno en el ganado Ankole, y la termotolerancia y la resistencia a las garrapatas observadas en el ganado cebú africano brindan oportunidades alternativas para la selección genética (125). Sobre la base de una pregunta que se plantea con frecuencia si es mejor seleccionar un ganado lechero para adaptarse al sistema, o alterar el sistema para adaptarse a un ganado lechero y como los productores lecheros en los países tropicales operan principalmente utilizando sistemas de pastoreo, es mejor identificar el genotipo del ganado lechero que los agricultores creen que se adaptará a su entorno de producción (99, 151 y 152).
8. Conclusión
Esta revisión proporciona una descripción completa de los genes y parámetros genéticos para la tolerancia al calor en razas de ganado lechero criadas en países tropicales. Destaca varios indicadores utilizados para medir la respuesta de HS en el ganado lechero, incluidas variables ambientales como la temperatura ambiente, la humedad relativa, la radiación solar y la velocidad del viento. También discutió la disminución en la producción de leche y las alteraciones en los rasgos de composición de la leche, como grasas, proteínas, lactosa, porcentajes de sólidos sin grasa y puntuación de células somáticas como indicadores de HS. Además, los parámetros fisiológicos como la temperatura corporal central, la frecuencia respiratoria, la temperatura rectal, la puntuación de jadeo, la puntuación de babeo, la frecuencia cardíaca y la frecuencia del pulso, así como los cambios de comportamiento, son indicadores potenciales de HS en el ganado lechero. Esta revisión describe varias razas lecheras de indicina y taurina criadas en países tropicales y los genes candidatos para la tolerancia al calor que se han identificado en razas de ganado lechero tropical. Discute tres enfoques, a saber, la espectrometría de masas en tándem de cromatografía de gases y líquidos, GC-LC/LC-MS y la resonancia magnética nuclear de protones (1H-RMN), utilizado para identificar los metabolitos de la leche y la sangre como biomarcadores vitales de HS en razas de ganado lechero, y los efectos de HS en esos metabolitos.
La respuesta fenotípica y los parámetros genéticos para la tolerancia al calor en razas de ganado lechero criados en Asia, América Latina y países subsaharianos también se presentan con varios modelos genéticos utilizados para estimar los componentes genéticos de la tolerancia al calor. La revisión enfatizó el antagonismo genético entre los rasgos de producción de leche y la tolerancia al calor, lo que indica la necesidad de considerar la sensibilidad genética de las posibles razas de ganado lechero criadas en ambientes tropicales para evitar los efectos adversos de HS. La revisión también destacó el uso potencial de las razas Bos indicus, que albergan un gran número de regiones genómicas y genes candidatos responsables de la adaptación climática, para cruzarse con razas Bos taurus para mejorar la producción de leche y la adaptación genética en países tropicales. Se discutieron las prácticas de manejo para mitigar la HS, como las modificaciones ambientales y las estrategias mejoradas de nutrición y alimentación, y se recomendó la selección genética para la tolerancia al calor como un enfoque rentable.
En conclusión, se recomienda la adopción de la selección genética para animales tolerantes al calor, junto con modificaciones ambientales, una mejor nutrición y el cruce de razas lecheras templadas con razas tropicales, para mitigar los impactos de HS en el ganado lechero, particularmente en los países tropicales. La investigación adicional sobre genes candidatos y parámetros genéticos asociados con la tolerancia al calor, así como el desarrollo de modelos de evaluación genética apropiados, pueden contribuir a la cría de razas de ganado lechero tolerantes al calor, mejorando en última instancia su rendimiento y resistencia frente al cambio climático. El cruce de razas templadas con razas locales que son resistentes a HS es una forma rápida de mejorar la termotolerancia de los animales y, por lo tanto, es muy recomendable en el África subsahariana.
Contribuciones del autor
VH realizó la investigación durante sus estudios de doctorado y preparó el borrador inicial del manuscrito. AN, ZN, CE-D, GM, RM y SC concibieron la idea del estudio, acordaron el contenido y editaron el manuscrito. Todos los autores contribuyeron al artículo y aprobaron la versión presentada.
Financiación
Este estudio fue financiado por la Asociación para las Habilidades en Ciencias Aplicadas, Ingeniería y Tecnología (PASET) a través del Fondo Regional de Becas e Innovación (RSIF) otorgado a VH para llevar a cabo estudios de doctorado en SACIDS Africa Centre of Excellence for Infectious Diseases, SACIDS Foundation for One Health, Sokoine University of Agriculture, Morogoro, Tanzania y el Instituto Internacional de Investigación Ganadera (ILRI, Nairobi, Kenia). Los financiadores no tuvieron ningún papel en el diseño del estudio, la decisión de publicar o la preparación del manuscrito. Los hallazgos y conclusiones de este estudio son los de los autores y no representan necesariamente los puntos de vista de los financiadores.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o reclamo que pueda ser hecho por su fabricante, no está garantizado ni respaldado por el editor.
Abreviaturas
BL: modelo de línea discontinua; BLUP, Mejor predicción lineal imparcial; BOA, Alelos de raza de origen; BUN: nitrógeno ureico en sangre; C16:0, Porcentaje de ácidos grasos palmíticos; C18:0, Porcentaje de ácidos grasos esteáricos; C18:1, Porcentaje de ácidos grasos oleicos; TCC: temperatura corporal-central; CP: porcentaje de caseína; DIM, Días en leche; República Democrática del Congo; DTV: variación diurna de la temperatura; EBVs, Valores genéticos estimados; F%, Porcentaje de grasa; FPR: proporción de grasa a proteína; PF: Porcentaje de grasa; FRM: modelos de regresión fija; GC–MS: Cromatografía de gases–espectrometría de masas; GEBV: Valores genéticos estimados genómicos; GxE: Genotipo por interacciones ambientales; GS: Selección genómica; GWAS, estudios de asociación de genoma completo; h2, estimaciones de heredabilidad; 1H-RMN: resonancia magnética nuclear de protones; HCT: Hematocrito; HGB: hemoglobina; FC: Tasa de calor; HS: Estrés por calor; HSP: proteínas de choque térmico; LC–MS/MS, espectrometría de masas en tándem de cromatografía líquida; LP: porcentaje de lactosa; LP: función polinómica de Legendre; MCV: volumen celular medio; MUFA: ácidos grasos monoinsaturados; MY, Producción de leche; NEFA: ácidos grasos no esterificados; P%, Porcentaje de proteína; PR: frecuencia del pulso; PP: porcentaje de proteína; PS: puntuación de Panting; PUFA: ácidos grasos poliinsaturados; qRT-PCR: reacción en cadena de la polimerasa cuantitativa en tiempo real; RBC: glóbulos rojos; REML: Máxima probabilidad restringida; HR: humedad relativa; RRM: modelos de regresión aleatoria; ROS: especies reactivas de oxígeno; RR: Frecuencia respiratoria; ARN: ácido ribonucleico; RN, Normas de reacción; RT: temperatura rectal; SCS: puntuación de células somáticas; SNPs: polimorfismo de nucleótido único; SFA: ácidos grasos saturados; SP: modelo Splines; RS: Tasas de sudoración; ssGBLUP, Mejor predicción lineal imparcial genómica de un solo paso; ssGREML: Probabilidad máxima restringida genómica de un solo paso; Ta: temperatura ambiente; TCA: ciclo del ácido tricarboxílico; TDMY: producción de leche el día de prueba; THI: Índice de temperatura-humedad; UFA: ácidos grasos insaturados; Reino Unido, Reino Unido; Estados Unidos, Estados Unidos de América; WBC: glóbulos blancos.
Referencias
1. Bernabucci, U, Biffani, S, Buggiotti, L, Vitali, A, Lacetera, N, y Nardone, A. Los efectos del estrés por calor en el ganado lechero italiano Holstein. J Dairy Sci. (2014) 97:471–86. DOI: 10.3168/JDS.2013-6611
2. Corazzin, M, Saccà, E, Lippe, G, Romanzin, A, Foletto, V, Da Borso, F, et al. Efecto del estrés por calor en el rendimiento de la vaca lechera y en la expresión de genes del metabolismo proteico en células mamarias. Animales. (2020) 10:1–13. DOI: 10.3390/ANI10112124
3. Marumo, JL, Lusseau, D, Speakman, JR, Mackie, M, and Hambly, C. Influence of environmental factors and parity on milk yield dynamics in barn-housed dairy cattle. J Dairy Sci. (2022) 105:1225–41. DOI: 10.3168/JDS.2021-20698
4. Dauria, BD, Sigdel, A, Petrini, J, Bóscollo, PP, Pilonetto, F, Salvian, M, et al. Efectos genéticos del estrés por calor en los ácidos grasos de la leche en el ganado Holstein brasileño. J Dairy Sci. (2022) 105:3296–305. DOI: 10.3168/JDS.2021-20914
5. Sungkhapreecha, P, Misztal, I, Hidalgo, J, Steyn, Y, Buaban, S, Duangjinda, M, et al. Cambios en los parámetros genéticos para la producción de leche y la tolerancia al calor en la población lechera cruzada tailandesa Holstein bajo diferentes niveles de estrés por calor y a lo largo del tiempo. J Dairy Sci. (2021) 104:12703–12. DOI: 10.3168/JDS.2021-20151
6. Boonkum, W, Misztal, I, Duangjinda, M, Pattarajinda, V, Tumwasorn, S, and Sanpote, J. Genetic effects of heat stress on milk yield of Thai Holstein crossbreeds. J Dairy Sci. (2011) 94:487–92. DOI: 10.3168/JDS.2010-3421
7. Cruz, GS, Urioste, JI y Saravia, CG. Efectos del estrés por calor en la producción de leche de vacas Holstein primíparas a escala regional utilizando grandes bases de datos. Archivos Latinoamericanos de Producción Animal. (2018) 26:67–76.
8. Garner, JB, Douglas, ML, Williams, SRO, Wales, WJ, Marett, LC, Nguyen, TTT, et al. La selección genómica mejora la tolerancia al calor en el ganado lechero. Informes científicos. (2016) 6:1–9. DOI: 10.1038/SEP34114
9. Osei-Amponsah, R, Dunshea, FR, Leury, BJ, Cheng, L, Cullen, B, Joy, A, et al. El estrés por calor afecta a las vacas lactantes que pastan en los pastos de verano australianos en una lechería robótica automática. Animales. (2020) 10:869. DOI: 10.3390/ANI10050869
10. Dunn, RJH, Mead, NE, Willett, KM, y Parker, DE. Análisis del estrés por calor en el ganado lechero del Reino Unido y el impacto en la producción de leche. Envir Res Letters. (2014) 9:4006. doi: 10.1088/1748-9326/9/6/064006
11. Cheruiyot, EK, Haile-Mariam, M, Cocks, BG, MacLeod, IM, Xiang, R, y Pryce, JE. Nuevos loci y vías neuronales para la resiliencia al estrés por calor en el ganado. Sci Rep. (2021) 11:1–16. DOI: 10.1038/S41598-021-95816-8
12. Carabaño, MJ, Pineda-Quiroga, C, Ugarte, E, Díaz, C, and Ramón, M. Genetic basis of thermotolerance in 2 local dairy sheep populations in the Iberian Peninsula. J Dairy Sci. (2021) 104:5755–67. DOI: 10.3168/JDS.2020-19503
13. Ekine-Dzivenu, CC, Mrode, R, Oloo, RD, Komwihangilo, D, Lyatuu, E, Msuta, G, et al. Análisis genómico de la producción de leche y la tolerancia al calor en el sistema lácteo de pequeños agricultores del África subsahariana. Proc 12th World Cong Genet App Lives Prod. (2022):1709–12. doi: 10.3920/978-90-8686-940-4_410
14. Dikmen, S, Cole, JB, Null, DJ, y Hansen, PJ. Mapeo de asociación de todo el genoma para la identificación de loci de rasgos cuantitativos para la temperatura rectal durante el estrés por calor en ganado Holstein. PLoS One. (2013) 8:1–7. doi: 10.1371/journal.pone.0069202
15. Sigdel, A, Abdollahi-Arpanahi, R, Aguilar, I, y Peñagaricano, F. El mapeo del genoma completo revela nuevos genes y vías involucradas en la producción de leche bajo estrés por calor en vacas Holstein estadounidenses. Genet frontal. (2019) 10:1–10. doi: 10.3389/fgene.2019.00928
16. Terefe, E, Belay, G, Tijjani, A, Han, J, y Hanotte, O. La resecuenciación del genoma completo revela diversidad genética y firmas de selección de ganado indígena etíope adaptado a entornos locales. Diversidad. (2023) 15:1–23. DOI: 10.3390/d15040540
17. Zulu, DN. Caracterización genética de razas bovinas nativas de Zambia. Tesis de maestría. Blacksburg: Instituto Politécnico de Virginia y Universidad Estatal (2008). 68 págs.
18. Richard, OA, Ebenezer, KA, y Frederick, YO. Cruce de ganado para la producción sostenible de leche en los trópicos. Int J Lives Prod. (2020) 11:108–13. DOI: 10.5897/IJLP2020.0717
19. Mbuthia, JM, Mayer, M, and Reinsch, N. Modeling heat stress effects on dairy cattle milk production in a tropical environment using test-day records and random regression models. Animal. (2021) 15:100222. doi: 10.1016/j.animal.2021.100222
20. Opoola, O, Banos, G, Ojango, JMK, Mrode, R, Simm, G, Banga, CB, et al. Análisis genético conjunto para el rendimiento del ganado lechero en países del África subsahariana. S Afric J Anim Sci. (2020) 50:507–20. doi: 10.4314/sajas.v50i4.3
21. Ouarfli, L, y Chehma, A. Efecto del índice de temperatura-humedad en el rendimiento de la leche de las vacas lecheras Holstein nacidas en el local bajo el clima sahariano. Arch Zoot. (2021) 24:24–36. DOI: 10.2478/Azibna-2021-0010
22. Stumpf, MT, Kolling, GJ, Fischer, V, dos Santos Daltro, D, Alfonzo, EPM, Dalcin, VC, et al. El índice elevado de temperatura-humedad induce alteraciones fisiológicas, sanguíneas y de leche en las vacas Holstein de una manera más pronunciada que en 2021/9 y 21040/10 Holstein × Gir. J Anim Behav Biomet. (31893) 21040:<>. doi: <>.<>/JABB.<>
23. Sungkhapreecha, P, Chankitisakul, V, Duangjinda, M, Buaban, S, and Boonkum, W. Determining heat stress effects of multiple genetic traits in tropical dairy cattle using single-step genomic BLUP. Veterinario Sci. (2022) 9:1–13. DOI: 10.3390/VETSci9020066
24. Chawala, AR, Sanchez-Molano, E, Dewhurst, RJ, Peters, A, Chagunda, MGG, and Banos, G. Breeding strategies for improving smallholder dairy cattle productivity in sub-Saharan Africa. J Anim Raza Genet. (2021) 138:668–87. DOI: 10.1111/JBG.12556
25. Niyonzima, YB, Strandberg, E, Hirwa, CD, Manzi, M, Ntawubizi, M, y Rydhmer, L. El efecto de la alta temperatura y la humedad en la producción de leche en Ankole y vacas cruzadas. Trop Anim Health Prod. (2022) 54:3092. DOI: 10.1007/S11250-022-03092-Z
26. Galukande, E, Mulindwa, H, Wurzinger, M, Roschinsky, R, Mwai, AO, y Sölkner, J. Cross-breeding cattle for milk production in the tropics: achievements, challenges and opportunities. Anim Genet Res. (2013):111–25. doi: 10.1017/S2078633612000471
27. Bora, SK, Tessema, TS, y Girmay, G. Genetic diversity and population structure of selected Ethiopian indigenous cattle breeds using microsatellite markers. Genet Res. (2023) 2023, 2023:1106755. DOI: 10.1155/2023/1106755
28. Rewe, TO, Peixoto, MGCD, Cardoso, VL, Vercesi Filho, AE, El Faro, L, y Strandberg, E. Gir for the giriama: the case for zebu dairying in the tropics – a review. Vive Res Rural Devel. (2015) 27.
29. Kabi, F, Muwanika, V, y Masembe, C. Indigenous cattle breeds and factors enhancing their variation, potential challenges of intensification and threats to genetic diversity in Uganda. Anim Genet Res. (2016) 58:1–12. doi: 10.1017/s2078633615000326
30. Msalya, G, Kim, ES, Laisser, ELK, Kipanyula, MJ, Karimuribo, ED, Kusiluka, LJM, et al. Determinación de la estructura genética y firmas de selección en tres cepas de cebú de cuernos cortos de Tanzania, ganado Boran y Frisón mediante análisis de SNP de todo el genoma. PLoS One. (2017) 12:1–18. doi: 10.1371/journal.pone.0171088
31. Kibona, CA, Yuejie, Z, y Tian, L. Towards developing a beef meat export policy in Tanzania: exploring the factors that influence beef meat exports. PLoS One. (2022) 17:E0270146. doi: 10.1371/journal.pone.0270146
32. Mwai, O, Hanotte, O, Kwon, YJ, y Cho, S. Revisión invitada – African indigenous cattle: unique genetic resources in a rapid changing world. Asiático Austr J Anim Sci. (2015) 28:911–21. doi: 10.5713/ajas.15.0002R
33. Norezzine, A, Duksi, F, Tsvetkova, AD, Ulybina, EA, Gins, MS, Rebouh, NY, et al. Caracterización genética del ganado blanco Fulani en Nigeria: un estudio comparativo. J Adv Vet Anim Res. (2019) 6:474–80. doi: 10.5455/javar.2019.f370
34. Odubote, IK. Caracterización de los sistemas de producción y prácticas de manejo de la cabaña bovina en Zambia. Trop Anim Health Prod. (2022) 54:1–11. DOI: 10.1007/S11250-022-03213-8
35. FAO. Caracterización y valor agregado a razas locales y sus productos en el cercano oriente y norte. África. (2012) 3:12–48.
36. Saravanan, KA, Panigrahi, M, Kumar, H, Parida, S, Bhushan, B, Gaur, GK, et al. Los escaneos genómicos para las firmas de selección revelaron genes candidatos para la adaptación y los rasgos de producción en una variedad de razas de ganado. Genómica. (2021) 113:955–63. doi: 10.1016/j.ygeno.2021.02.009
37. Velayudhan, SM, Brügemann, K, Alam, S, Yin, T, Devaraj, C, Sejian, V, et al. Respuestas moleculares, fisiológicas y hematológicas del ganado lechero cruzado en un clima de sabana tropical. Biología. (2023) 12:1–16. doi: 10.3390/biología12010026
38. Worku, D, Gowane, G, Alex, R, Joshi, P, and Verma, A. Inputs for optimization selection platform for milk production traits of dairy Sahiwal cattle. PLoS One. (2022) 17:1–19. doi: 10.1371/journal.pone.0267800
39. Peixoto, MGCD, Carvalho, MRS, Egito, AA, Steinberg, RS, Bruneli, FÂT, Machado, MA, et al. Diversidad genética y estructura genética poblacional de una metapoblación guzerá (Bos indicus). Animales. (2021) 11:1–14. DOI: 10.3390/ANI11041125
40. Verma, P, Sharma, A, Sodhi, M, Tiwari, M, Vivek, P, Kataria, RS, et al. Identificación de genes internos de referencia en células mononucleares de sangre periférica de poblaciones bovinas adaptadas a ambientes de Normoxia árida caliente e hipoxia árida fría. Genet frontal. (2022) 12:1–16. doi: 10.3389/fgene.2021.730599
41. Chanda, T, Debnath, G, Khan, K, Rahman, M, and Chanda, G. Impact of heat stress on milk yield and composition in early lactation of Holstein Friesian crossbred cattle. Banglad J Anim Sci. (2018) 46:192–7. doi: 10.3329/bjas.v46i3.36314
42. Egito, AA, Paiva, SR, Albuquerque, MDSM, Mariante, AS, Almeida, LD, Castro, SR, et al. Diversidad genética basada en microsatélites y relaciones entre diez razas de ganado criado criado en Brasil. BMC Genet. (2007) 8:1–14. DOI: 10.1186/1471-2156-8-83
43. Santana, ML, Pereira, RJ, Bignardi, AB, Vercesi Filho, AE, Menéndez-Buxadera, A, y El Faro, L. Detrimental effect of selection for milk yield on genetic tolerance to heat stress in purebred zebu cattle: genetic parameters and trends. J Dairy Sci. (2015) 98:9035–43. DOI: 10.3168/JDS.2015-9817
44. Santana, ML Jr, Pereira, RJ, Bignardi, AB, El Faro, L, Pires3, MFÁ, Andrade, RG, et al. El ganado Guzerá de doble propósito exhibe un alto rendimiento lechero bajo estrés por calor. J Anim Raza Genet. (2019):1–9. DOI: 10.1111/JBG.12450
45. Habimana, V, Ekine-Dzivenu, CC, Nguluma, AS, Nziku, ZC, Morota, G, Chenyambuga, SW, et al. Genes y modelos para estimar parámetros genéticos para la tolerancia al calor en ganado lechero. Genet frontal. (2023) 14:1–13. doi: 10.3389/fgene.2023.1127175
46. Bohmanova, J, Misztal, I, y Cole, JB. Índices de temperatura-humedad como indicadores de pérdidas de producción de leche por estrés térmico. J Dairy Sci. (2007) 90:1947–56. DOI: 10.3168/JDS.2006-513
47. Carabaño, MJ, Logar, B, Bormann, J, Minet, J, Vanrobays, ML, Díaz, C, et al. Modelado del estrés por calor bajo diferentes condiciones ambientales. J Dairy Sci. (2016) 99:3798–814. DOI: 10.3168/JDS.2015-10212
48. Dalcin, VC, Fischer, V, Daltro, D d S, Alfonzo, EPM, Stumpf, MT, Kolling, GJ, et al. Parámetros fisiológicos para el estrés térmico en ganado lechero. Revis Bras Zootec. (2016) 45:458–65. doi: 10.1590/S1806-92902016000800006
49. Pinto, S, Hoffmann, G, Ammon, C, and Amon, T. Critical THI thresholds based on the physiological parameters of lactating dairy cows. J Therm Biol. (2020) 88:102523. doi: 10.1016/j.jtherbio.2020.102523
50. Mader, TL, Davis, MS, y Brown-Brandl, T. Environmental factors influencing heat stress in feedlot cattle. J Anim Sci. (2006) 84:712–9. doi: 10.2527/2006.843712x
51. Berian, S. Effect of heat stress on physiological and Hemato-biochemical profile of cross baging boke. J Anim Res. (2019) 9:13. DOI: 10.30954/2277-940X.01.2019.13
52. Gaafar, H, El-Nahrawy, M, Mesbah, R, Shams, AS, Sayed, S, and Anas, AB. Impact of heat stress on growth performance and some blood and physiological parameters of suckling Friesian terners in Egypt. Inter J Plant, Anim Environ Sci. (2021) 11:545–65. doi: 10.26502/ijpaes.202121
53. Li, G, Chen, S, Chen, J, Peng, D, and Gu, X. Predicting rectal temperature and breathation rate responses in lactating dairy cows exposed to heat stress. J Dairy Sci. (2020) 103:5466–84. DOI: 10.3168/JDS.2019-16411
54. Zheng, S, Dong, J, Sun, Z, Qin, G, Li, C, Zhen, Y, et al. Correlación de los indicadores relacionados con el estrés oxidativo con la composición de la leche y los metabolitos en vacas lecheras de lactancia temprana. Veterinario Médico Sci. (2021):1–10. DOI: 10.1002/VMS3.615
55. Honig, H, Miron, J, Lehrer, H, Jackoby, S, Zachut, M, Zinou, A, et al. Rendimiento y bienestar de vacas lecheras de alto rendimiento sometidas a 5 u 8 sesiones de enfriamiento diarias en climas cálidos y húmedos. J Dairy Sci. (2012) 95:3736–42. DOI: 10.3168/JDS.2011-5054
56. Lambertz, C, Sanker, C, and Gauly, M. Climatic effects on milk production traits and somatic cell score in lactating Holstein-Friesian cows in different housing systems. J Dairy Sci. (2014) 97:319–29. DOI: 10.3168/JDS.2013-7217
57. Persson, Y, y Olofsson, I. Direct and indirect measurement of somatic cell count as indicator of intramammary infection in dairy winats. Acta Vet Scand. (2011) 53:1–5. DOI: 10.1186/1751-0147-53-15
58. Reis, NS, Ferreira, IC, Mazocco, LA, Souza, ACB, Pinho, GAS, Neto, ÁMDF, et al. La sombra modifica las respuestas conductuales y fisiológicas de vacas lecheras de producción baja a media en pastizales en un sistema integrado de cultivo-ganadería-bosque. Animales. (2021) 11:2411. doi: 10.3390/ani11082411
59. Bernabucci, U, Lacetera, N, Baumgard, LH, Rhoads, RP, Ronchi, B, and Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated rumiants. Animal. (2010) 4:1167–83. doi: 10.1017/S175173111000090X
60. Collier, RJ, Baumgard, LH, Zimbelman, RB, y Xiao, Y. Estrés por calor: fisiología de la aclimatación y adaptación. Frente Anim. (2019) 9:12–9. doi: 10.1093/af/vfy031
61. Becker, CA, Aghalari, A, Marufuzzaman, M, y Stone, AE. Predicción del estrés por calor del ganado lechero utilizando técnicas de aprendizaje automático. J Dairy Sci. (2021) 104:501–24. DOI: 10.3168/JDS.2020-18653
62. St-Pierre, NR, Cobanov, B, y Schnitkey, G. Economic losses from heat stress by US livestock industries1. J Dairy Sci. (2003) 86:E52–77. DOI: 10.3168/JDS. S0022-0302(03)74040-5
63. Anderson, SD, Bradford, BJ, Harner, JP, Tucker, CB, Choi, CY, Allen, JD, et al. Efectos de los ventiladores ajustables y estacionarios con nebulizadores sobre la temperatura corporal central y el comportamiento de mentira de las vacas lecheras lactantes en un clima semiárido. J Dairy Sci. (2013) 96:4738–50. DOI: 10.3168/JDS.2012-6401
64. Gálik, R, Lüttmerding, G, Boďo, Š, Knížková, I, y Kunc, P. Impact of heat stress on selected parameters of robotic milking. Animales. (2021) 11:1–10. doi: 10.3390/ani11113114
65. Hammami, H, Vandenplas, J, Vanrobays, M, Rekik, B, Bastin, C, and Gengler, N. Genetic analysis of heat stress effects on yield traits, ubre health, and fatty acids of Walloon Holstein cows. J Dairy Sci. (2015) 98:4956–68. DOI: 10.3168/JDS.2014-9148
66. Rong, Y, Zeng, M, Guan, X, Qu, K, Liu, J, Zhang, J, et al. Asociación de la variación genética de HSF1 con la tolerancia al calor en ganado chino. Animales. (2019) 9:3–9. DOI: 10.3390/ANI9121027
67. Negri, R, Aguilar, I, Feltes, GL y Cobuci, JA. Selección para el día de prueba Producción de leche y termotolerancia en ganado Holstein brasileño. Animales. (2021) 11:1–13. doi: 10.3390/ani11010128
68. Kadzere, CT, Murphy, MR, Silanikove, N, y Maltz, E. Estrés por calor en vacas lecheras lactantes: una revisión. Vidas Prod Sci. (2002) 77:59–91. doi: 10.1016/S0301-6226(01)00330-X
69. Kumar, J, Kumar, M, Madan, AK, Singh, Y, Yadav, B, y Anand, M. Effect of season on physiological parameters and production profile of Hariana and Sahiwal cattle. Haryana Vet. (2017) 56:69–71.
70. Velayudhan, SM, Brügemann, K, Sejian, V, Bhatta, R, Schlecht, E, Pinto, A, et al. Efectos del estrés por calor a través de la interfaz rural-urbana en las expresiones de rasgos fenotípicos del ganado lechero en una región de sabana tropical. Sostenibilidad. (2022) 14. DOI: 10.3390/SU14084590
71. Kohli, S, Atheya, Reino Unido, y Thapliyal, A. Evaluación del índice óptimo de humedad térmica para vacas lecheras cruzadas en el distrito de Dehradun, Uttarakhand. India Vet World. (2014) 7:916–21. doi: 10.14202/vetworld.2014.916-921
72. Jeelani, R, Konwar, D, Khan, A, Kumar, D, Chakraborty, D, and Brahma, B. Reassessment of temperature-moisture index for measuring heat stress in crossbred dairy botle of a sub-tropical region. J Therm Biol. (2019) 82:99–106. doi: 10.1016/j.jtherbio.2019.03.017
73. Narmilan, A, Puvanitha, N, Ahamed, AS, Santhirakumar, S, and Narmilan, A. Relationship between temperature-humidity index and Milk production of dairy cows in tropical climate. Asian J Dairy Food Res. (2021) 40:246–52. DOI: 10.18805/AJDFR. DR-213
74. Ekine-Dzivenu, CC, Mrode, R, Oyieng, E, Komwihangilo, D, Lyatuu, E, Msuta, G, et al. Evaluación del impacto del estrés por calor medido por el índice de temperatura-humedad (THI) en la producción de leche del día de prueba del ganado lechero de pequeños agricultores en un clima africano subsahariano. Vidas Sci. (2020) 242:4314. doi: 10.1016/j.livsci.2020.104314
75. Du Preez, JH, Giesecke, WH, Hattingh, PJ, y Eisenberg, BE. Estrés por calor en ganado lechero en condiciones del sur de África. Onderst J. (1990) 57:77–87.
76. Hady, MM, Melegy, TM, y Anwar, SR. Impact of the Egyptian summer season on oxidative stress biomarkers and some physiological parameters in crossbreeds and Egyptian buffaloes. Mundo veterinario. (2018) 11:771–7. doi: 10.14202/vetworld.2018.771-777
77. Santana, ML, Bignardi, AB, Pereira, RJ, Menéndez-Buxadera, A, and El Faro, L. Random regression models to account for the effect of genotype by environment interaction due to heat stress on the milk yield of Holstein cows under tropical conditions. J Appl Genet. (2016) 57:119–27. DOI: 10.1007/S13353-015-0301-X
78. Santana, ML, Bignardi, AB, Pereira, RJ, Stefani, G, y El Faro, L. Genetics of heat tolerance for milk yield and quality in Holsteins. Animal. (2017) 11:4–14. doi: 10.1017/S1751731116001725
79. Negri, R, Aguilar, I, Feltes, GL, Machado, JD, Neto, JB, Costa-Maia, FM, et al. Inclusión de variables bioclimáticas en evaluaciones genéticas de ganado lechero. Anim Biosci. (2021) 34:163–71. doi: 10.5713/ajas.19.0960
80. Vieira, R, Louvandini, H, Barcellos, J, Martins, CF, and McManus, C. Path and logistic analysis for heat tolerance in adapted breeds of bovinos in Brazil. Vidas Sci. (2022) 258:104888. doi: 10.1016/j.livsci.2022.104888
81. Espinoza, JL, Sánchez, J, Gracia, JA, Sánchez, JR, Ortega, R, y Palacios, A. La termorregulación difiere en Chinampo (Bos taurus) y en el ganado lechero nacido localmente. Turk J Vet Anim Sci. (2009) 33:175–80. DOI: 10.3906/VET-0704-24
82. Fan, C, Su, D, Tian, H, Li, X, Li, Y, Ran, L, et al. Perturbaciones metabólicas hepáticas de vacas lecheras lactantes con estrés por calor. Asian-Austr J Anim Sci. (2018) 31:1244–51. DOI: 10.5713/AJAS.17.0576
83. Yue, S, Ding, S, Zhou, J, Yang, C, Hu, X, Zhao, X, et al. El enfoque metabolómico explora los biomarcadores de diagnóstico y los cambios metabólicos en vacas lecheras con estrés por calor. Animal. (2020) 10:1741. doi: 10.3390/ani10101741
84. Lv, Z, Liu, H, Yang, Y, Bu, D, Zang, C, Yang, K, et al. Cambios en los metabolitos de la leche bovina con variantes de β-caseína revelados por la metabolómica. Animales. (2020) 10:1–11. DOI: 10.3390/ANI10060954
85. Fan, CY, Su, D, Tian, H, Hu, RT, Ran, L, Yang, Y, et al. Producción y composición de la leche y alteraciones metabólicas en la glándula mamaria de vacas lecheras lactantes con estrés por calor. J Integ Agric. (2019) 18:2844–53. doi: 10.1016/S2095-3119(19)62834-0
86. Contreras-Jodar, A, Nayan, NH, Hamzaoui, S, Caja, G, y Salama, AAK. El estrés por calor modifica el rendimiento de la lactancia y el perfil metabolómico urinario relacionado con la microbiota gastrointestinal de cabras lecheras. PLoS One. (2019) 14:1–14. doi: 10.1371/journal.pone.0202457
87. Hudaya, MF, Sitaresmi, PI, Noviandi, CT, Widyobroto, BP y Widayati, DT. Comportamiento y perfil sanguíneo en vacas lecheras Friesian-Holstein en la región especial de Yogyakarta. Indonesia J Anim Behav Biomet. (2020) 8:244–9. doi: 10.31893/JABB.20032
88. Tian, H, Zheng, N, Wang, W, Cheng, J, Li, S, Zhang, Y, et al. Estudio metabolómico integrado de la leche de vacas lecheras lactantes con estrés térmico. Sci Rep. (2016) 6:1–10. DOI: 10.1038/SEP24208
89. Hu, L, Brito, LF, Zhang, H, Zhao, M, Liu, H, Chai, H, et al. El perfil del metaboloma del plasma revela diferentes respuestas metabólicas al desafío agudo por frío entre el ganado Sanhe y Holstein del centro de Mongolia. J Dairy Sci. (2022) 105:9162–78. DOI: 10.3168/JDS.2022-21996
90. Kekana, TW, Nherera-Chokuda, FV, Muya, MC, Manyama, KM, y Lehloenya, KC. Producción de leche y metabolitos sanguíneos del ganado lechero influenciados por el índice de humedad térmica. Trop Anim Health Prod. (2018) 50:921–4. DOI: 10.1007/S11250-018-1513-Y
91. Gao, ST, Guo, J, Quan, SY, Nan, XM, Fernandez, MVS, Baumgard, LH, et al. Los efectos del estrés por calor en el metabolismo de las proteínas en vacas Holstein lactantes. J Dairy Sci. (2017) 100:5040–9. DOI: 10.3168/JDS.2016-11913
92. Carabaño, MJ, Bachagha, K, Ramón, M, and Díaz, C. Modeling heat stress effect on Holstein cows under hot and dry conditions: selection tools. J Dairy Sci. (2014) 97:7889–904. DOI: 10.3168/JDS.2014-8023
93. Bohlouli, M, Shodja, J, Alijani, S, y Eghbal, A. La relación entre el índice de temperatura-humedad y la producción de leche en el día de prueba del ganado lechero iraní Holstein utilizando el modelo de regresión aleatoria. Vidas Sci. (2013) 157:414–20. doi: 10.1016/j.livsci.2013.09.005
94. Hammami, H, Bormann, J, M’hamdi, N, Montaldo, HH, and Gengler, N. Evaluation of heat stress effects on production traits and somatic cell score of Holsteins in a temperate environment. J Dairy Sci. (2013) 96:1844–55. DOI: 10.3168/JDS.2012-5947
95. Luo, H, Brito, LF, Li, X, Su, G, Dou, J, Xu, W, et al. Parámetros genéticos para la temperatura rectal, la frecuencia respiratoria y la puntuación de babeo en el ganado Holstein y sus relaciones con diversos rasgos de fertilidad, producción, conformación corporal y salud. J Dairy Sci. (2021) 104:4390–403. DOI: 10.3168/JDS.2020-19192
96. Ravagnolo, O, y Misztal, I. Componente genético del estrés por calor en el ganado lechero, estimación de parámetros. J Dairy Sci. (2000) 83:2126–30. DOI: 10.3168/JDS. S0022-0302(00)75095-8
97. Cho, CI, Alam, M, Choi, TJ, Choy, YH, Choi, JG, Lee, SS, et al. Modelos para estimar parámetros genéticos de rasgos de producción de leche utilizando modelos de regresión aleatoria en ganado Holstein coreano. Asiático Austr J Anim Sci. (2016) 29:607–14. doi: 10.5713/ajas.15.0308
98. Bohlouli, M, Alijani, S, Naderi, S, Yin, T, and König, S. Prediction precisionies and genetic parameters for test-day traits from genomic and pedigree-based random regression models with or without heat stress interactions. J Dairy Sci. (2019) 102:488–502. DOI: 10.3168/JDS.2018-15329
99. Pryce, JE, Nguyen, TTT, Cheruiyot, EK, Marett, L, Garner, JB, and Haile-Mariam, M. Impact of hot weather on animal performance and genetic strategies to minimis the effect. Anim Prod Sci. (2022) 62:726–35. doi: 10.1071/AN21259
100. Carrara, ER, Petrini, J, Salvian, M, de Oliveira, HR, Rovadoscki, GA, Iung, LHDS, et al. Parámetros genéticos para la producción de leche y rasgos de calidad de vacas Holstein brasileñas en función del índice de temperatura y humedad. J Anim Raza Genet. (2021) 138:643–54. DOI: 10.1111/JBG.12636
101. Otto, PI, Guimarães, SEF, Verardo, LL, Azevedo, ALS, Vandenplas, J, Sevillano, CA, et al. Estudios de asociación de todo el genoma para la respuesta al estrés por calor en Bos taurus × Bos indicus ganado cruzado. J Dairy Sci. (2019) 102:8148–58. DOI: 10.3168/JDS.2018-15305
102. Freitas, PHF, Wang, Y, Yan, P, Oliveira, HR, Schenkel, FS, Zhang, Y, et al. Diversidad genética y firmas de selección para el estrés térmico en bovinos y otras dos especies de Bos adaptadas a condiciones climáticas divergentes. Genet frontal. (2021) 12:4823. doi: 10.3389/fgene.2021.604823
103. Kumar, A, Waiz, SA, Sridhar Goud, T, Tonk, RK, Grewal, A, Singh, SV, et al. Evaluación de la adaptabilidad de las razas de ganado cebú (Bos indicus) en dos condiciones climáticas diferentes: utilizando técnicas citogenéticas sobre la integridad del genoma. Int J Biometeorol. (2016) 60:873–82. DOI: 10.1007/S00484-015-1080-0
104. Srikanth, K, Kwon, A, Lee, E, and Chung, H. Characterization of genes and pathways that respond to heat stress in Holstein terneros through transcriptome analysis. Chaper de estrés celular. (2017) 22:29–42. DOI: 10.1007/S12192-016-0739-8
105. Hassan, FU, Nawaz, A, Rehman, MS, Ali, MA, Dilshad, SMR, and Yang, C. Prospects of HSP70 as a genetic marker for thermo-tolerance and immuno-modulation in animals under climate change scenario. Anim Nutrit. (2019) 5:340–50. doi: 10.1016/j.aninu.2019.06.005
106. Kishore, A, Sodhi, M, Kumari, P, Mohanty, AK, Sadana, DK, Kapila, N, et al. Células mononucleares de sangre periférica: un sistema celular potencial para comprender la respuesta diferencial de choque térmico en ganado nativo (Bos indicus), ganado exótico (Bos taurus) y búfalos ribereños (Bubalus bubalis) de la India. Chaper de estrés celular. (2014) 19:613–21. DOI: 10.1007/S12192-013-0486-Z
107. Sun, L, Qu, K, Ma, X, Hanif, Q, Zhang, J, Liu, J, et al. Los análisis del genoma completo revelan características genómicas y firmas de selección del ganado jorobado Lincang en la frontera entre China y Myanmar. Genet frontal. (2022) 13:1–11. doi: 10.3389/fgene.2022.833503
108. Iqbal, N, Liu, X, Yang, T, Huang, Z, Hanif, Q, Asif, M, et al. Variantes genómicas identificadas a partir de la resecuenciación del genoma completo de razas de ganado indicino de Pakistán. PLoS One. (2019) 14:1–21. doi: 10.1371/journal.pone.0215065
109. Sajjanar, B, Aalam, MT, Khan, O, Tanuj, GN, Sahoo, AP, Mishra, BP, et al. Genoma: el análisis de expresión amplia revela diferentes respuestas de choque térmico en ganado autóctono (Bos indicus) y cruzado (Bos indicus X Bos taurus). Genes Envir. (2023) 45:1–15. DOI: 10.1186/S41021-023-00271-8
110. Morenikeji, OB, Ajayi, OO, Peters, SO, Mujibi, FD, de Donato, M, Thomas, BN, et al. RNA-seq profiling of skin in temperate and tropical cattle. J Anim Sci Tech. (2020) 62:141–58. doi: 10.5187/jast.2020.62.2.141
111. Onasanya, GO, Msalya, GM, Thiruvenkadan, AK, Sreekumar, C, Tirumurugaan, GK, Fafiolu, AO, et al. Los genotipos heterocigotos de polimorfismo de nucleótido único en el gen de la proteína de choque térmico 70 influyen potencialmente en la tolerancia térmica entre cuatro razas de cebú de Nigeria. Genet frontal. (2021) 12:1–11. doi: 10.3389/fgene.2021.642213
112. Vanvanhossou, SFU, Yin, T, Scheper, C, Fries, R, Dossa, LH, and König, S. Unraveling admix, inbreeding, and recent selection signatures in West African indigenous cattle populations in Benin. Genet frontal. (2021) 12:1–19. DOI: 10.3389/fgene.2021.657282
113. Xu, L, Yang, L, Zhu, B, Zhang, W, Wang, Z, Chen, Y, et al. El escaneo de todo el genoma revela divergencia genética y diversa selección adaptativa en ganado local chino. BMC Genómica. (2019) 20:1–13. DOI: 10.1186/S12864-019-5822-Y
114. Jang, J, Terefe, E, Kim, K, Lee, YH, Belay, G, Tijjani, A, et al. Corrección a: variación del número de copias diferenciadas poblacionales de Bos taurus, Bos indicus y sus híbridos africanos. BMC Genómica. (2021) 22:531:1–12. DOI: 10.1186/S12864-021-07808-7
115. Peripolli, E, Stafuzza, NB, Machado, MA, do Carmo Panetto, JC, do Egito, AA, Baldi, F, et al. Evaluación de variantes del número de copias en tres razas bovinas brasileñas adaptadas localmente utilizando datos de resecuenciación del genoma completo. Anim Genet. (2023):1–17. doi: 10.1111/edad.13298
116. Dixit, SP, Bhatia, AK, Ganguly, I, Singh, S, Dash, S, Sharma, A, et al. Los análisis del genoma revelaron mezcla genética y firmas de selección en Bos indicus. Sci Rep. (2021) 11:1–11. DOI: 10.1038/S41598-021-01144-2
117. Liao, X, Peng, F, Forni, S, McLaren, D, Plastow, G, and Stothard, P. Whole genome sequencing of Gir cattle for identifying polymorphisms and loci under selection. Genoma. (2013) 56:592–8. DOI: 10.1139/GEN-2013-0082
118. Huson, HJ, Kim, ES, Godfrey, RW, Olson, TA, McClure, MC, Chase, CC, et al. Estudio de asociación de todo el genoma y orígenes ancestrales del pelaje de pelo liso en ganado adaptado tropicalmente. Genet frontal. (2014) 5:101. doi: 10.3389/fgene.2014.00101
119. Toro-Ospina, AM, Herrera Rios, AC, Bizarria Santos, W, Pimenta Schettini, G, Vallejo Aristizabal, VH, Tovar Claros, G, et al. Arquitectura genética y firmas de selección en el criollo caqueteño (ganado nativo colombiano). Diversidad. (2022) 14:828. DOI: 10.3390/d14100828
120. Bahbahani, H, Tijjani, A, Mukasa, C, Wragg, D, Almathen, F, Nash, O, et al. Firmas de selección para adaptación ambiental y cebú × aptitud híbrida taurina en cebú de cuernos cortos de África oriental. Genet frontal. (2017) 8:68. doi: 10.3389/fgene.2017.00068
121. Tijjani, A, Salim, B, da Silva, MVB, Eltahir, HA, Musa, TH, Marshall, K, et al. Firmas genómicas para la adaptación de tierras secas en regiones ricas en genes en ganado cebú africano. Genómica. (2022) 114:110423. doi: 10.1016/j.ygeno.2022.110423
122. Makina, SO, Muchadeyi, FC, Van Marle-Köster, E, Taylor, JF, Makgahlela, ML, y Maiwashe, A. Genome-wide scan for selection signatures in six cattle breeds in South Africa. Genet Selec Evol. (2015) 47:1–14. DOI: 10.1186/S12711-015-0173-X
123. Armstrong, DV. Interacción de estrés por calor con sombra y enfriamiento. J Dairy Sci. (1994) 77:2044–50. DOI: 10.3168/JDS. S0022-0302(94)77149-6
124. Joksimovic-Todorovic, M, Davidovic, V, Hristov, S, y Stankovic, B. Effect of heat stress on milk production in dairy cows. Biot Anim Husb. (2011) 27:1017–23. doi: 10.2298/bah1103017j
125. Osei-Amponsah, R, Chauhan, SS, Leury, BJ, Cheng, L, Cullen, B, Clarke, IJ, et al. Selección genética para la termotolerancia en rumiantes. Animales. (2019) 9:1–18. DOI: 10.3390/ANI9110948
126. Oeste, TJ. Efectos del estrés térmico en la producción en ganado lechero. J Dairy Sci. (2003) 86:2131–44. DOI: 10.3168/JDS. S0022-0302(03)73803-X
127. Wang, Z, Niu, K, Rushdi, HE, Zhang, M, Fu, T, Gao, T, et al. El estrés por calor induce cambios en las bacterias del rumen y el metaboloma de Buffalo. Animales. (2022) 12:1300. DOI: 10.3390/ANI12101300
128. Tresoldi, G, Schütz, KE, y Tucker, CB. Evaluación de la carga de calor en ganado lechero de lote seco: refinación de la metodología de muestreo en la granja. J Dairy Sci. (2016) 99:8970–80. DOI: 10.3168/JDS.2016-11353
129. Ouellet, V, Toledo, IM, Dado-Senn, B, Dahl, GE, and Laporta, J. Critical temperature-humidity index thresholds for dry cows in a subtropical climate. Frente Anim Sci. (2021) 2:6636. doi: 10.3389/fanim.2021.706636
130. Shwartz, G, Rhoads, ML, Vanbaale, MJ, Rhoads, RP y Baumgard, LH. Efectos de un cultivo de levadura suplementario en vacas Holstein lactantes estresadas por calor. J Dairy Sci. (2009) 92:935–42. DOI: 10.3168/JDS.2008-1496
131. Zimbelman, RB, Baumgard, LH, y Collier, RJ. Efectos de la niacina encapsulada sobre la pérdida de calor por evaporación y la temperatura corporal en vacas Holstein lactantes moderadamente estresadas por calor. J Dairy Sci. (2010) 93:2387–94. DOI: 10.3168/JDS.2009-2557
132. Palacio, S, Bergeron, R, Lachance, S, y Vasseur, E. Los efectos de proporcionar sombra portátil en el pasto en el comportamiento y la fisiología de las vacas lecheras. J Dairy Sci. (2015) 98:6085–93. DOI: 10.3168/JDS.2014-8932
133. Perano, KM, Usack, JG, Angenent, LT, y Gebremedhin, KG. Producción y respuestas fisiológicas del ganado lechero lactante estresado por calor al enfriamiento conductivo. J Dairy Sci. (2015) 98:5252–61. DOI: 10.3168/JDS.2014-8784
134. Van Laer, E, Tuyttens, FAM, Ampe, B, Sonck, B, Moons, CPH, and Vandaele, L. Effect of summer conditions and shade on the production and metabolism of Holstein dairy cows on pastture in temperate climate. Animal. (2015) 9:1547–58. doi: 10.1017/S1751731115000816
135. Domingos, HGT, Maia, ASC, Souza, JBF, Silva, RB, Vieira, FMC y Silva, RG. Efecto de la sombra y la aspersión de agua sobre las respuestas fisiológicas y los rendimientos de leche de las vacas Holstein en una región semiárida. Vidas Sci. (2013) 154:169–74. doi: 10.1016/j.livsci.2013.02.024
136. Ortiz, XA, Smith, JF, Villar, F, Hall, L, Allen, J, Oddy, A, et al. Una comparación de 2 sistemas de enfriamiento evaporativo en una granja lechera comercial en Arabia Saudita. J Dairy Sci. (2015) 98:8710–22. DOI: 10.3168/JDS.2015-9616
137. Tao, S, Thompson, IM, Monteiro, APA, Hayen, MJ, Young, LJ y Dahl, GE. Efecto del enfriamiento de vacas lecheras estresadas por calor durante el período seco sobre la respuesta a la insulina. J Dairy Sci. (2012) 95:5035–46. DOI: 10.3168/JDS.2012-5405
138. Garner, JB, Williams, SRO, Moate, PJ, Jacobs, JL, Hannah, MC, Morris, GL, et al. Los efectos del estrés por calor en las vacas lecheras ofrecieron dietas que contenían trigo o grano de maíz durante la lactancia tardía. Animales. (2022) 12:2031. doi: 10.3390/ani12162031
139. Godara, AS, Bhat, SA, Yogi, RK, Devi, S, and Sahoo, SP. Feeding strategies to amelioring the impact of heat stress in bovine. Inter J Sci Nature. (2016) 7:25–9.
140. Baena, MM, Tizioto, PC, Meirelles, SLC, and Regitano, LC d A. HSF1 and HSPA6 as functional candidate genes associated with heat tolerance in Angus bovinos. Revis Bras Zootec. (2018) 47:390. DOI: 10.1590/RBZ4720160390
141. Rolf, M. M. (2015). Base genética para la tolerancia al calor en el ganado. Oklahoma City: Taxi, 1–21.
142. Cuellar, CJ, Saleem, M, y Jensen, LM. Diferencias en la regulación de la temperatura corporal durante el estrés por calor y la depresión estacional en la producción de leche entre Holstein, Brown Swiss y vacas cruzadas. J Dairy Sci. (2023) 106:22725. DOI: 10.3168/JDS.2022-22725
143. Michael, P, De Cruz, CR, Nor, NM, Jamli, S, y Meng, GY. El potencial del uso de cruces templado-tropicales y subproductos agrícolas, asociado con el manejo del estrés por calor para la producción lechera en los trópicos: una revisión. Animales. (2022) 12:1–16. DOI: 10.3390/ANI12010001
144. Deb, R, Sajjanar, B, Singh, U, Kumar, S, Singh, R, Sengar, G, et al. Efecto del estrés por calor en el programa de expresión de Hsp90 entre las razas de ganado Sahiwal (Bos indicus) y Frieswal (Bos indicus × Bos taurus): un estudio comparativo. Genes. (2014) 536:435–40.
145. Maibam, U, Hooda, OK, Sharma, PS, Singh, SV, Mohanty, AK, Upadhyay, RC, et al. Variación estacional en la expresión de HSP70 y estrés oxidativo en la piel de ganado cebú (Tharparkar) y cruzado (Karan Fries) bajo clima tropical. Biolog Rhyt Res. (2017) 1016:1–15. doi: 10.1080/09291016.2017.1307905
146. Dikmen, S, Cole, JB, Null, DJ y Hansen, PJ. Heredabilidad de la temperatura rectal y correlaciones genéticas con rasgos de producción y reproducción en ganado lechero. J Dairy Sci. (2012) 95:3401–5. DOI: 10.3168/JDS.2011-4306
147. Camargo, DASL, y Pereira, FJ. Oportunidades de edición del genoma para mejorar la productividad del ganado en los trópicos. CABI Agric Biosci. (2022) 3:1–11. DOI: 10.1186/S43170-022-00075-W
148. Dikmen, S, Khan, FA, Huson, HJ, Sonstegard, TS, Moss, JI, Dahl, GE, et al. El locus de pelo SLICK derivado del ganado Senepol confiere termotolerancia a las vacas Holstein lactantes de manejo intensivo. J Dairy Sci. (2014) 97:5508–20. DOI: 10.3168/JDS.2014-8087
149. Dikmen, S, Alava, E, Pontes, E, Fear, JM, Dikmen, BY, Olson, TA, et al. Diferencias en la capacidad termorreguladora entre vacas Holstein lactantes de pelo liso y salvajes en respuesta al estrés por calor agudo. J Dairy Sci. (2008) 91:3395–402. DOI: 10.3168/JDS.2008-1072
150. Nguyen, TTT, Bowman, PJ, Haile-Mariam, M, Pryce, JE, y Hayes, BJ. Selección genómica para la tolerancia al estrés por calor en ganado lechero australiano. J Dairy Sci. (2016) 99:2849–62. DOI: 10.3168/JDS.2015-9685
151. Carabaño, MJ, Ramón, M, Menéndez-Buxadera, A, Molina, A, y Díaz, C. Selección de tolerancia al calor. Frente Anim. (2019) 9:62–8. doi: 10.1093/af/vfy033
Cita: Habimana V, Nguluma AS, Nziku ZC, Ekine-Dzivenu CC, Morota G, Mrode R y Chenyambuga SW (2023) Efectos del estrés por calor en los rasgos y metabolitos de la producción de leche y estrategias de mitigación para razas de ganado lechero criadas en países tropicales y subtropicales. Frente. Vet. Sci. 10:1121499. doi: 10.3389/fvets.2023.1121499
Editado por:
Johanna Ramírez-Díaz, Consejo Nacional de Investigación (CNR), Italia
Revisado por:
Eui-Soo Kim, Recombinetics, Estados Unidos
Angela Costa, Universidad de Bolonia, Italia
Copyright © 2023 Habimana, Nguluma, Nziku, Ekine-Dzivenu, Morota, Mrode y Chenyambuga. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Vincent Habimana, vincenthabimana5@gmail.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía