
Efectos del sistema de cría mixto en jaula vs. suelo de red sobre la histomorfología y los perfiles de expresión génica del bazo de ganso
Efectos del sistema de cría mixto en jaula vs. suelo de red sobre la histomorfología y los perfiles de expresión génica del bazo de ganso
 Qingliang Chen1,2,3†
Qingliang Chen1,2,3†  Canción Yang1,2,3†
Canción Yang1,2,3†  Zhiyu He1,2,3 Guang Yang1,2 Junqi Wang1,2,3
Zhiyu He1,2,3 Guang Yang1,2 Junqi Wang1,2,3  Xiaopeng Li1,2,3 Wanxia Wang4
Xiaopeng Li1,2,3 Wanxia Wang4  Xin Yuan1,2 Jiwei Hu1,2,3 Hua He1,2,3
Xin Yuan1,2 Jiwei Hu1,2,3 Hua He1,2,3  Liang Li1,2,3
Liang Li1,2,3  Jiwen Wang1,2,3
Jiwen Wang1,2,3  Shenqiang Hu1,2,3*
Shenqiang Hu1,2,3*- 1Exploración e Innovación de Recursos Genéticos de Animales de Granja Laboratorio Clave de la Provincia de Sichuan, Universidad Agrícola de Sichuan, Chengdu, China
- 2Laboratorio Clave de Multiómica Ganadera y Avícola, Ministerio de Agricultura y Asuntos Rurales, Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Sichuan, Chengdu, China
- 3Laboratorio Estatal Clave de Ganadería y Cría de Aves de Corral, Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Sichuan, Chengdu, China
- 4Departamento de Producción Animal, Estación General de Cría de Animales de la Provincia de Sichuan, Chengdu, Sichuan, China
Debido a las demandas tanto de protección ambiental como de modernización de la industria del ganso en China, los sistemas tradicionales de cría de gansos en la ribera han ido pasando gradualmente a los modernos sistemas de cría intensiva en tierras secas, como el sistema de cría mixta de suelo neto (MRS) y el sistema de cría en jaulas (CRS). Sin embargo, las respuestas inmunitarias de los gansos a los diferentes sistemas de cría en tierras secas siguen siendo poco conocidas. Este estudio tuvo como objetivo investigar y comparar los efectos dependientes de la edad de MRS y CRS en las características histomorfológicas esplénicas y los perfiles de expresión de genes relacionados con el sistema inmunológico entre tres razas de ganso económicamente importantes, que incluyen el ganso blanco de Sichuan (SW), el ganso de Gang (GE) y el ganso de las Landas (LD). El análisis morfológico reveló que el peso esplénico y el índice orgánico del SW fueron mayores en el CRS que en el MRS (p < 0,05). Las observaciones histológicas mostraron que para el SW y el LD, el diámetro y el área del corpúsculo esplénico y el diámetro de la arteria trabecular fueron mayores bajo MRS que bajo CRS a las 30 o 43 semanas de edad (p < 0,05), mientras que el área de pulpa roja esplénica de GE fue mayor bajo CRS que bajo MRS a las 43 semanas de edad (p < 0,05). Además, a las 43 semanas de edad, se observaron niveles más altos de expresión de ARNm de NGF, SPI1 y VEGFA en el bazo de SW bajo MRS que bajo CRS (p < 0,05), mientras que se observaron niveles más altos de HSPA2 y NGF en bazos de LD bajo MRS que bajo CRS (p < 0,05). En el caso de la GE, hubo niveles más altos de expresión de ARNm de MYD88 en el bazo bajo CRS a las 30 semanas de edad (p < 0,05). Además, nuestro análisis de correlación mostró que parecía haber asociaciones positivas más pronunciadas entre los parámetros histológicos esplénicos y los niveles de expresión de varios genes clave relacionados con el sistema inmunitario bajo MRS que bajo CRS. Por lo tanto, se especula que los gansos criados bajo MRS podrían exhibir funciones inmunes mejoradas que los que se crían bajo CRS, particularmente para SW y LD. Aunque se supone que estas diferencias fenotípicas están asociadas con los perfiles de expresión diferencial dependientes de la edad de HSPA2, MYD88, NGF, SPI1 y VEGFA en el bazo de ganso, los mecanismos reguladores subyacentes esperan más investigaciones.
1 Introducción
China es el mayor productor y consumidor mundial de gansos de engorde, y el consumo de productos de ganso ha aumentado año tras año (1). Además, China es rica en recursos genéticos de gansos, ya que tiene 31 razas autóctonas de gansos y varias razas de gansos europeos importadas. Todas las razas de gansos autóctonos chinos, excepto el ganso Yili, se derivan del ganso cisne (Anser cygnoides), mientras que el ganso Yili y las razas domésticas europeas se derivan del ganso común (Anser anser) (2). En los últimos años, debido a las demandas tanto de protección ambiental como de modernización de la industria del ganso en China, los sistemas de cría de gansos han ido pasando gradualmente de los tradicionales de cría a los modernos de cría intensiva en tierras secas, como el sistema de cría en el suelo (FRS), el sistema de cría en red (NRS), el sistema de cría mixta en el suelo (MRS), y el sistema de cría en jaulas (SRI) (3).
Cada vez hay más pruebas de que la producción y la salud de las aves de corral se vieron significativamente afectadas por el sistema de cría. Se ha demostrado que las aves criadas en sistemas de interior y exterior mostraron diferentes respuestas inmunitarias. El ambiente al aire libre expuso a las aves a una gama más amplia de microorganismos y otros antígenos, lo que podría estimular su sistema inmunológico para que se desarrollara rápidamente y, por lo tanto, mostró respuestas inmunes más fuertes (4, 5). Los sistemas de cría en secano en interiores, como MRS y CRS, que son más propicios para controlar las condiciones de alojamiento al realizar la separación del estiércol, pueden mejorar en gran medida la eficiencia y la salud de la producción avícola. Un estudio anterior informó que los pollos de engorde Desi criados bajo CRS exhibieron un mejor rendimiento de crecimiento (como la tasa de conversión alimenticia, el peso corporal y la tasa muscular del pecho) que los que estaban bajo FRS (6). De manera similar, en comparación con FRS, los patos Chaohu criados bajo NRS mostraron un mejor rendimiento de crecimiento, como el peso corporal y la ganancia diaria promedio (7). En comparación, se sabe menos sobre los efectos de los diferentes sistemas de cría en la producción y la salud de los gansos. Se demostró que los gansos criados en corrales equipados con piscina mostraron un menor peso corporal, ingesta de alimento y tasa de conversión alimenticia en comparación con los que estaban bajo FRS (8). Otro estudio informó que los gansos de Yangzhou criados bajo CRS mostraron una ganancia diaria promedio y un peso corporal más altos que los bajo FRS (3). Además, se informó que el sistema de cría altera significativamente las características histomorfológicas ileales del ganso y la composición microbiana cecal, lo que podría afectar sus funciones fisiológicas y su rendimiento productivo (9). Sin embargo, hasta la fecha, las respuestas inmunitarias de los gansos a los diferentes sistemas de cría en tierras secas siguen siendo poco conocidas.
El bazo, que sirve como un órgano inmunológico importante en las aves de corral, desempeña un papel crucial en las respuestas inmunes humorales y celulares al apoyar la generación, maduración y almacenamiento de linfocitos (10). Los niveles de expresión de genes clave relacionados con el sistema inmunitario en el bazo de las aves de corral se utilizan comúnmente como indicadores clave de sus respuestas inmunitarias (11). Se ha demostrado que el sistema de cría está implicado en la regulación de la expresión de genes relacionados con el sistema inmunitario para modular las funciones inmunitarias de las aves de corral. Por ejemplo, los pollos de engorde criados bajo FRS exhibieron niveles más altos de expresión de ARNm de interleucina-1β e interferón-γ en el íleon que aquellos bajo CRS (12). En comparación con FRS, el NRS aumentó los niveles séricos de interleucina-1β, interferón-γ, interleucina-4 e inmunoglobulinas en patos de carne, lo que mejoró las funciones inmunológicas de los patos (13). En comparación con el sistema de cría en interiores, los niveles de expresión de ARNm del receptor 7 (TLR7) fueron significativamente más altos en la bursa, el pulmón, el duodeno, el íleon y el ciego de los patos criados en libertad (14). Además, la expresión de varios genes clave relacionados con el sistema inmunitario, incluido el receptor toll like 2 (TLR2), el receptor toll like 4 (TLR4), la subunidad 1 del factor nuclear kappa B (NF-κB1) y la interleucina 6 (IL-6), aumentó significativamente en el bazo de los pollos de engorde criados bajo FRS que en los de CRS (15). En conjunto, estos resultados sugirieron que el sistema de cría puede afectar el desarrollo esplénico y las respuestas inmunitarias de las aves de corral mediante la modulación de la actividad de las vías de señalización TLR2/4, NF-κB y MAPK. Cabe destacar que se ha demostrado que la vía de señalización única NF-κB desempeña un papel central en la mediación de las respuestas inmunitarias celulares a los estímulos externos (16, 17). Por lo tanto, también es importante explorar el mecanismo molecular por el cual el sistema de cría ejerce los efectos dependientes de la edad sobre el desarrollo esplénico y las funciones inmunitarias del ganso mediante el examen de los perfiles de expresión de estos genes clave relacionados con el sistema inmunitario.
Teniendo en cuenta que tanto las dos razas de ganso autóctonas chinas (ganso blanco de Sichuan, SW y ganso de Gang, GE) como una raza de ganso europeo importada (ganso de las Landas, LD) se han criado ampliamente en China debido a sus notables rasgos y valores económicos significativos, es importante comparar los impactos de diferentes sistemas de cría en tierras secas en las respuestas inmunitarias de estas tres razas de ganso. El presente estudio tuvo como objetivo, en primer lugar, comparar los efectos dependientes de la edad de MRS y CRS en las características histomorfológicas esplénicas entre tres razas de gansos y, posteriormente, identificar los genes clave responsables de las variaciones dependientes de la edad en las respuestas inmunes esplénicas bajo diferentes sistemas de cría entre tres razas de gansos. Se espera que estos resultados proporcionen una base teórica para mejorar las funciones inmunitarias de los gansos en los sistemas de cría en tierras secas.
2 Materiales y métodos
2.1 Animales de experimentación
En este estudio se utilizaron tres razas de gansos de tamaño mediano, incluyendo SW, GE y LD, que fueron proporcionadas por la Granja Experimental de Cría de Aves Acuáticas de la Universidad Agrícola de Sichuan (Ya’an, Sichuan, China). Un total de 100 gansos machos de un día de edad de cada una de estas tres razas de gansos nacieron del mismo lote y fueron alimentados en las mismas condiciones ambientales durante el período de 0 a 10 semanas de edad. Posteriormente, los gansos sanos de cada raza se dividieron en partes iguales en dos grupos (MRS y CRS), y se les proporcionó acceso gratuito a alimento y agua hasta la recolección de la muestra. Los gansos bajo SRI se criaron en jaulas individuales con las dimensiones de largo × ancho ×altura: 0,55 m × 0,37 m × 0,7 m, a una altura de 1,5 m sobre el nivel del suelo. Los gansos bajo MRS se criaron en un área interior con las dimensiones de largo × ancho: 6 m × 13 m, que consistió en un área de 60 m2 redes de plástico a una altura de 1 m sobre el nivel del suelo y una altura de 18 m2 lecho de fermentación. El horario de iluminación diario para los gansos bajo ambos sistemas de cría es de 16 h encendido y 8 h apagado, con las luces encendidas a las 08:00 a.m.
2.2 Recogida de muestras
Tanto a las 30 (alcanzar la madurez sexual) como a las 43 (alcanzar la madurez corporal) semanas de edad, se midieron los pesos corporales después de 12 h de ayuno, y se seleccionaron aleatoriamente 8 gansos machos con pesos corporales similares de cada raza de ganso de CRS y MRS, respectivamente. Todos los gansos experimentales seleccionados (n = 8 por semana por raza y por sistema de cría) fueron sacrificados mediante la inhalación de dióxido de carbono, seguida de una luxación cervical. Después del sacrificio, el bazo y el timo se recogieron y pesaron rápidamente. El índice de spleen/órgano timo se calculó mediante la siguiente fórmula: índice de órgano bazo/timo (%) = (peso del spleen/órgano timo (g)/peso corporal (kg)) × 100%. Para cada grupo, la mitad de los bazos de cada raza en cada punto de muestreo (n = 4) se utilizaron para el examen histológico, mientras que el resto (n = 4) se congeló rápidamente en nitrógeno líquido y finalmente se almacenó a -80 °C hasta la extracción del ARN.
2.3 Observación histológica
Los bazos de ganso recién recolectados se fijaron primero con formaldehído al 4% a temperatura ambiente durante 72 h, luego se deshidrataron a través de una serie de diferentes concentraciones de etanol y, finalmente, se transfirieron a xileno y se incrustaron en cera de parafina (18). Finalmente, cada muestra se cortó en rodajas de 5 μm de grosor, que se tiñeron con hematoxilina y eosina (H&E). Estas rebanadas teñidas fueron observadas y fotografiadas utilizando un microscopio de cámara trinocular digital BA410-Digital (Motic China Group Co. Ltd., Xiamen, China). De acuerdo con los métodos descritos en varios estudios previos (19-21), los parámetros histológicos esplénicos del ganso se analizaron utilizando el software Image-Pro Plus 6.0 (National Institutes of Health, Bethesda, MD).
En detalle, se observó el área de pulpa roja esplénica de ganso (RPA) midiendo el área de interés (AOI) del área cubierta por pulpa roja. El área del corpúsculo esplénico (ALA) se observó midiendo el AOI del corpúsculo esplénico. El diámetro del corpúsculo esplénico (ALD) se calculó midiendo el diámetro promedio del AOI del corpúsculo esplénico. El área de la trabécula esplénica (TLA) se observó midiendo el AOI de la trabécula esplénica. El diámetro de la arteria trabecular (TAD) se calculó midiendo el diámetro promedio del AOI de la arteria trabecular. El diámetro de la arteria central (CAD) se calculó midiendo el diámetro medio de la AOI de la arteria central. El diámetro o área media de cada parámetro histológico esplénico se calculó como la media de las observaciones de todos los individuos examinados por grupo experimental.
2.4 Extracción de ARN y PCR cuantitativa con transcripción inversa en tiempo real (RT-qPCR)
El ARN total se extrajo de los tejidos del bazo mediante el reactivo Trizol (Invitrogen, Massachusetts, CA, Estados Unidos) siguiendo las instrucciones del fabricante. La calidad del ARN se evaluó mediante un espectrofotómetro NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA) y electroforesis en un gel de agarosa al 1,5%. Aproximadamente 1 μg de ARN total de cada muestra se transcribió inversamente en ADNc mediante HiScript RT SuperMix para qPCR (+limpiador de ADNg; Vazyme, Nanjing, China) de acuerdo con las instrucciones del fabricante. Las reacciones de RT-qPCR se realizaron en el sistema de detección de PCR en tiempo real Bio-Rad CFX96 (Bio-Rad, Hercules, CA, Estados Unidos) utilizando 2 × UDG Fast SYBR Green qPCR Master Mix (Cofitt, Chengdu, China). El sistema de reacción se realizó en un volumen total de 20 μL, incluyendo 10 μL de 2 × UDG Fast SYBR Green qPCR Master Mix (Cofitt, Chengdu, China), 0,4 μL de cada cebador, 0,4 μL de ADNc, 8,8 μL de ddH®20. Las condiciones de amplificación de RT-qPCR se enumeraron de la siguiente manera: preprocesamiento de UDG a 50 °C durante 2 min; activación enzimática a 94°C durante 10 min; predesnaturalización a 95°C durante 3 min; seguido de 40 ciclos de desnaturalización a 95 °C durante 5 s, y recocido/extensión a la temperatura correspondiente de cada juego de imprimación durante 30 s. La especificidad objetivo de cada conjunto de cebadores se evaluó mediante el análisis de la curva de fusión, y la identidad de todos los amplicones se verificó mediante secuenciación. Los controles sin plantilla y los controles negativos sin transcriptasa inversa también se incluyeron en todas las series de RT-qPCR. Cada muestra se ejecutó por triplicado. Los niveles relativos de expresión de ARNm de los genes diana se normalizaron a los dos genes de referencia GAPDH y β-ACTINA utilizando los 2-ΔΔCT método (22). Los pares de cebadores utilizados en nuestro análisis de RT-qPCR se enumeran en la Tabla 1.
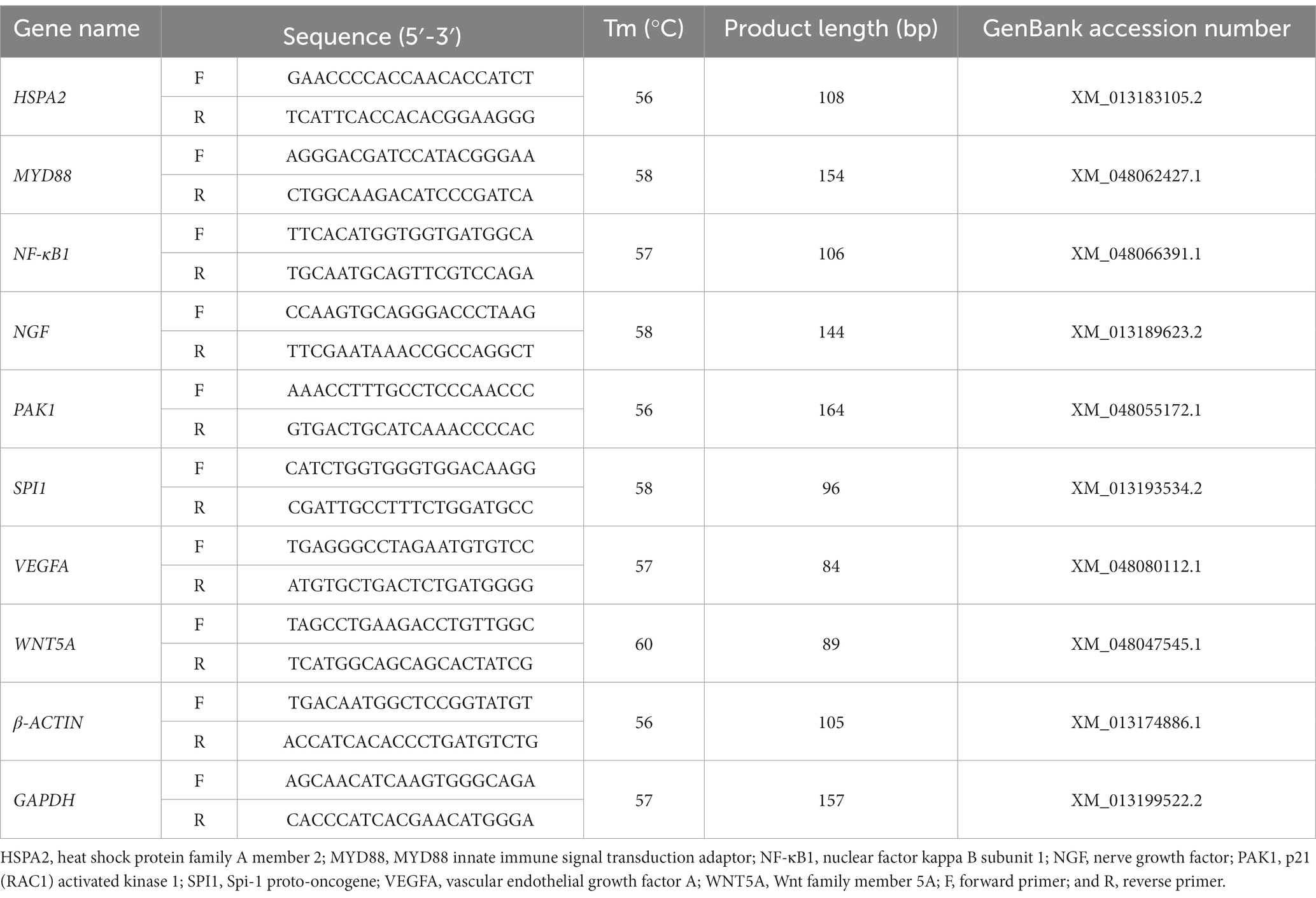 Tabla 1. Secuencias de cebadores utilizadas para la PCR cuantitativa de transcripción inversa en tiempo real.
Tabla 1. Secuencias de cebadores utilizadas para la PCR cuantitativa de transcripción inversa en tiempo real.
2.5 Análisis estadístico
Todos los datos se expresaron como la media ± SEM. Para el análisis estadístico se utilizó el software SAS 9.4 (SAS Institute Inc., Carolina del Norte, Estados Unidos). Los datos se analizaron mediante análisis de varianza de dos vías (ANOVA) utilizando el procedimiento de modelo lineal general (GLM) en el software SAS 9.4, con el sistema de recría (MRS o CRS) y la edad (30 o 43 semanas de edad) como factores fijos. Se realizó el procedimiento de la prueba t de Student para evaluar la significancia entre dos grupos diferentes. Se utilizó el análisis de correlación de Spearman para analizar las correlaciones entre las características histomorfológicas esplénicas y los niveles de expresión génica relacionados con el sistema inmunitario. Un valor de probabilidad (P) menor que 0,01 se consideró estadísticamente extremadamente significativo diferente, mientras que un valor de p menor que 0,05 se consideró significativamente diferente. Para dibujar las figuras se utilizaron tanto GraphPad Prism 8.0.2 (GraphPad Software, San Diego, CA, Estados Unidos) como el software R 3.14 (Lucent Technologies Inc., Milpitas, Estados Unidos). Los resultados con p < 0,05 se consideraron estadísticamente significativos.
3 Resultados
3.1 Efectos dependientes de la edad de MRS y CRS sobre el peso y los índices de órganos del bazo y el timo entre tres razas de gansos
Como se presenta en la Figura 1 y en la Tabla Suplementaria 1, para SW, los pesos e índices de órganos de bazo y timo fueron afectados estadísticamente significativamente por el sistema de cría (p < 0.05). En detalle, el peso esplénico y el índice orgánico de SW fueron menores bajo MRS que bajo CRS a las 30 o 43 semanas de edad (p < 0,05). Del mismo modo, el peso del timo y el índice de órganos del SW fueron menores tanto en la TMS como en el CRS a las 30 y 43 semanas de edad (p < 0,05), y los de la LD fueron menores en la MRS que en el CRS a las 30 semanas de edad (p < 0,05). No hubo diferencia significativa en los parámetros morfológicos esplénicos de la GE entre MRS y CRS a las 30 y 43 semanas de edad. En particular, el peso del timo y el índice de órganos disminuyeron generalmente durante el período de 30 a 40 semanas de edad entre tres razas de gansos, y se vieron afectados significativamente por el factor edad en GE (p < 0,05).
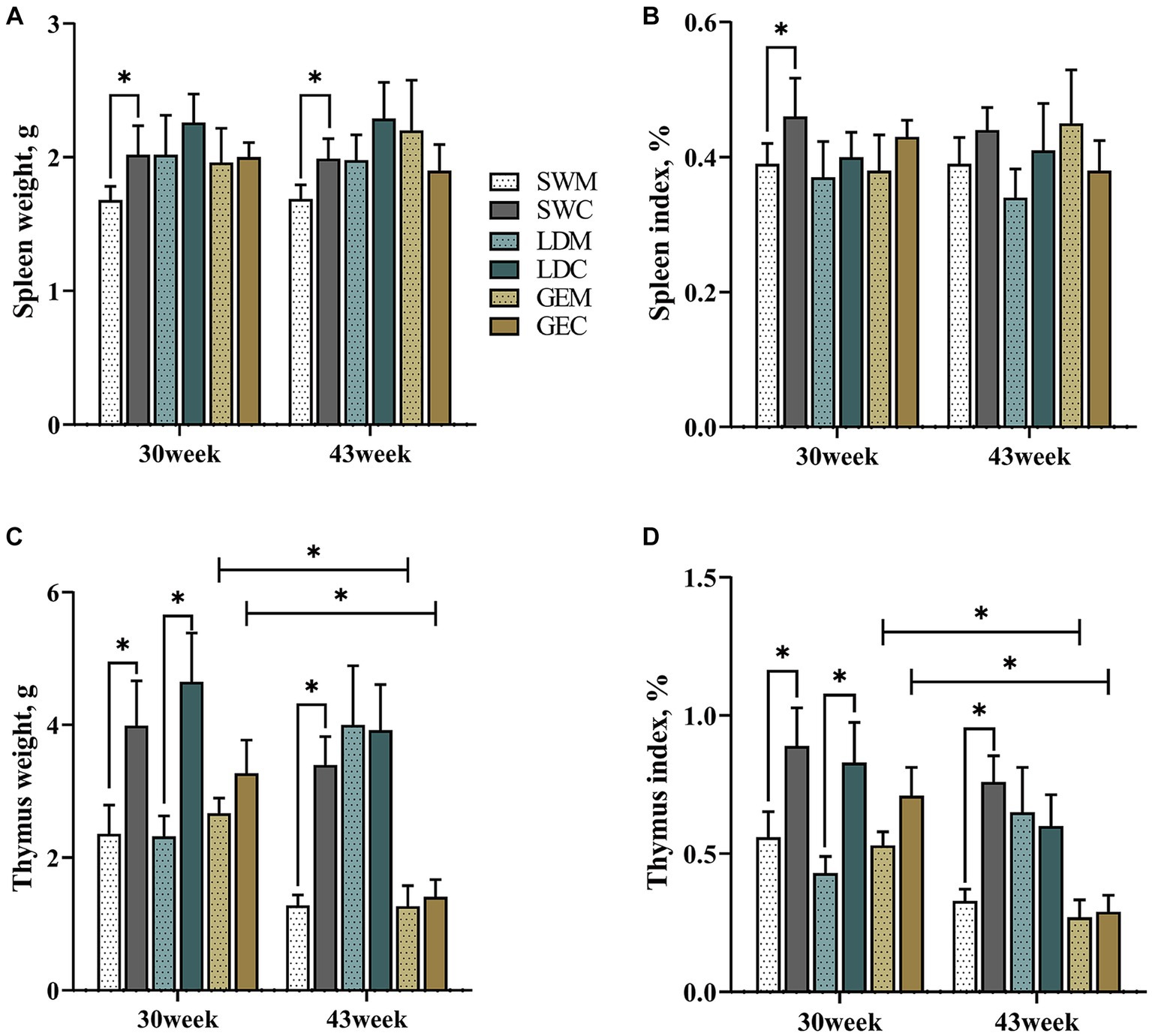 Figura 1. Comparación de los efectos de MRS y CRS en el desarrollo de órganos inmunes entre tres razas de gansos. (A) Peso del bazo. (B) Índice de órganos del bazo. (C) Peso del timo. (D) Índice de órganos del timo. «*» indica una diferencia significativa entre los dos grupos designados al nivel de p < 0,05. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; SWM; Ganso blanco de Sichuan bajo MRS; SWC, Ganso blanco de Sichuan bajo CRS; LDM, ganso de las Landas bajo MRS; LDC, ganso de las Landas en el CRS; GEM, Ganso de la pandilla bajo MRS; y GEC, Ganso de la pandilla bajo CRS.
Figura 1. Comparación de los efectos de MRS y CRS en el desarrollo de órganos inmunes entre tres razas de gansos. (A) Peso del bazo. (B) Índice de órganos del bazo. (C) Peso del timo. (D) Índice de órganos del timo. «*» indica una diferencia significativa entre los dos grupos designados al nivel de p < 0,05. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; SWM; Ganso blanco de Sichuan bajo MRS; SWC, Ganso blanco de Sichuan bajo CRS; LDM, ganso de las Landas bajo MRS; LDC, ganso de las Landas en el CRS; GEM, Ganso de la pandilla bajo MRS; y GEC, Ganso de la pandilla bajo CRS.
3.2 Efectos dependientes de la edad de MRS y CRS en los parámetros histológicos esplénicos en tres razas de gansos
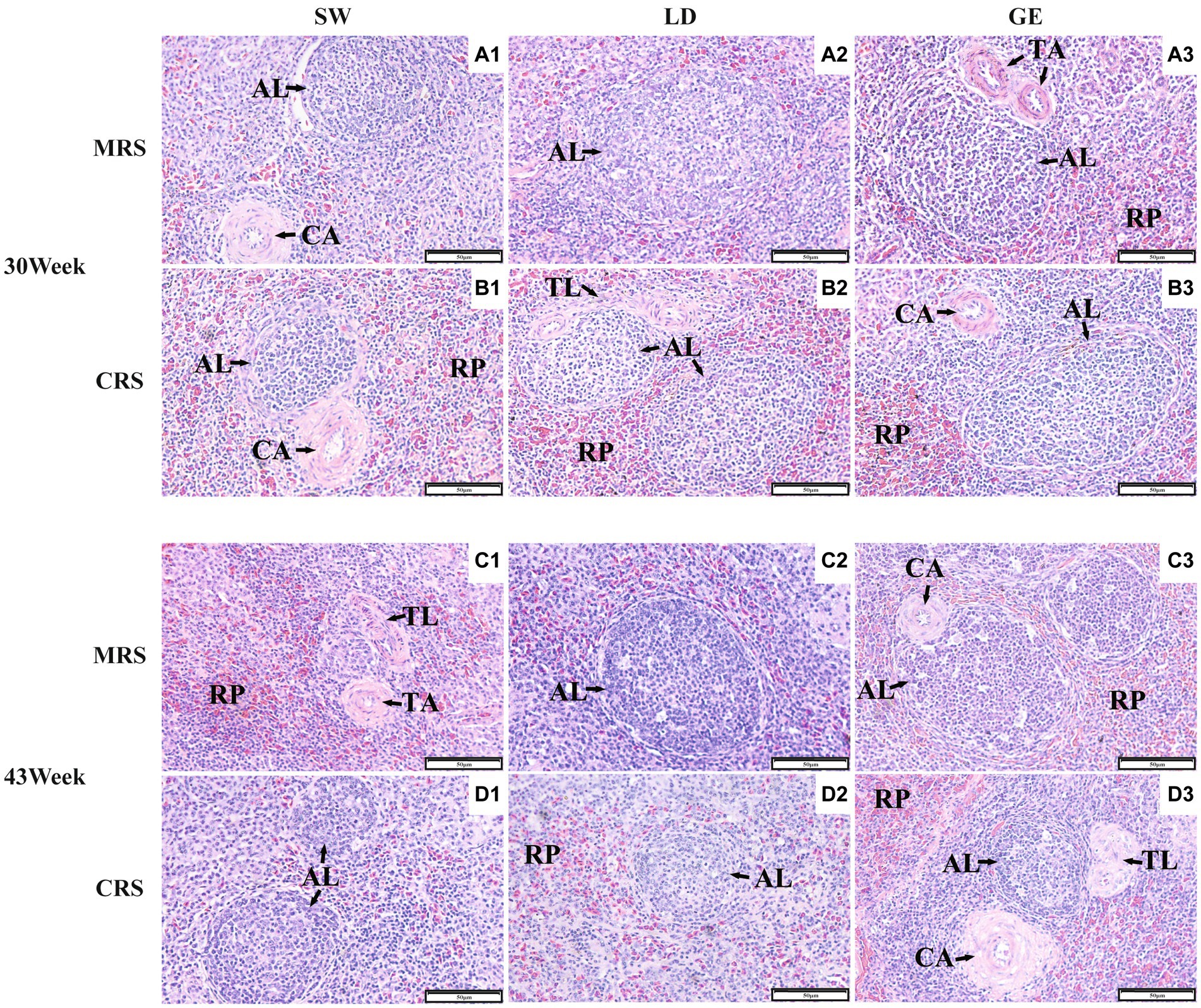
A continuación, investigamos los efectos de MRS y CRS sobre los parámetros histológicos esplénicos en tres razas de gansos. Como se muestra en la Figura 2, el bazo de ganso estaba compuesto histológicamente principalmente por estroma y parénquima. El estroma del bazo de ganso estaba compuesto principalmente por una red de tejido conectivo reticular que proporciona soporte y contención a las células sanguíneas y las células inmunitarias, como los linfocitos, los macrófagos y las células dendríticas. El parénquima del bazo de ganso estaba formado por pulpa roja y blanca, sin ninguna demarcación definida de límites. La pulpa roja estaba compuesta principalmente por sinusoides llenos de sangre, mientras que la pulpa blanca está poblada principalmente por tejido linfoide como el corpúsculo esplénico.
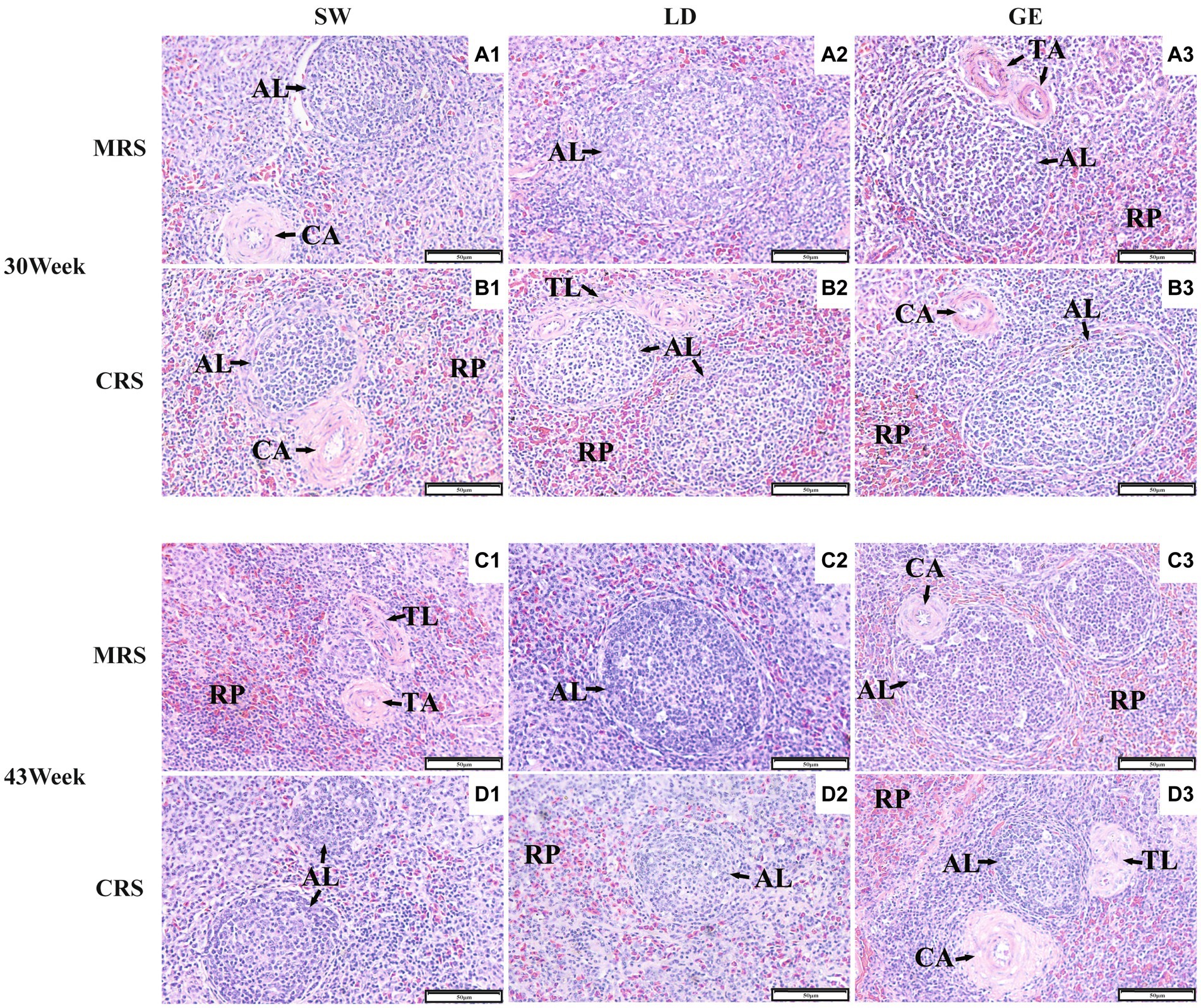 Figura 2. Observaciones histológicas del bazo de tres razas de gansos bajo dos sistemas de cría diferentes. Imágenes histológicas representativas (200 ×) del bazo de gansos bajo MRS a las 30 semanas de edad (A1-A3), bajo CRS a las 30 semanas de edad (B1-B3), bajo MRS a las 43 semanas de edad (C1-C3) y bajo CRS a las 43 semanas de edad (D1-D3). Los números 1, 2 y 3 representan las diferentes razas SW, LD y GE. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; AL: corpúsculo esplénico; CA: arteria central; TA: arteria trabecular; TL: trabécula esplénica; y RP, pulpa roja.
Figura 2. Observaciones histológicas del bazo de tres razas de gansos bajo dos sistemas de cría diferentes. Imágenes histológicas representativas (200 ×) del bazo de gansos bajo MRS a las 30 semanas de edad (A1-A3), bajo CRS a las 30 semanas de edad (B1-B3), bajo MRS a las 43 semanas de edad (C1-C3) y bajo CRS a las 43 semanas de edad (D1-D3). Los números 1, 2 y 3 representan las diferentes razas SW, LD y GE. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; AL: corpúsculo esplénico; CA: arteria central; TA: arteria trabecular; TL: trabécula esplénica; y RP, pulpa roja.
Además, se analizaron las diferencias en los parámetros histológicos esplénicos medidos entre la EMR y la SRC. Como se muestra en la Figura 3 y en la Tabla Suplementaria 2, el ALA, el ALD y el TAD de SW se vieron afectados significativamente por el sistema de cría (p < 0,001) y la edad (p < 0,05). Para LD, el TLA y el TAD se vieron afectados significativamente por el sistema de cría (p < 0,05). Para GE, el RPA se vio afectado significativamente por el sistema de cría (p < 0,001), mientras que el TAD se vio afectado significativamente por la edad (p < 0,05). Para el SW y el LD, algunos parámetros histológicos esplénicos (incluyendo el ALA, el ALD y el TAD) fueron mayores bajo MRS que bajo CRS a las 30 o 43 semanas de edad (p < 0,05). Para GE, el RPA fue mayor bajo CRS que bajo MRS (p < 0,05).
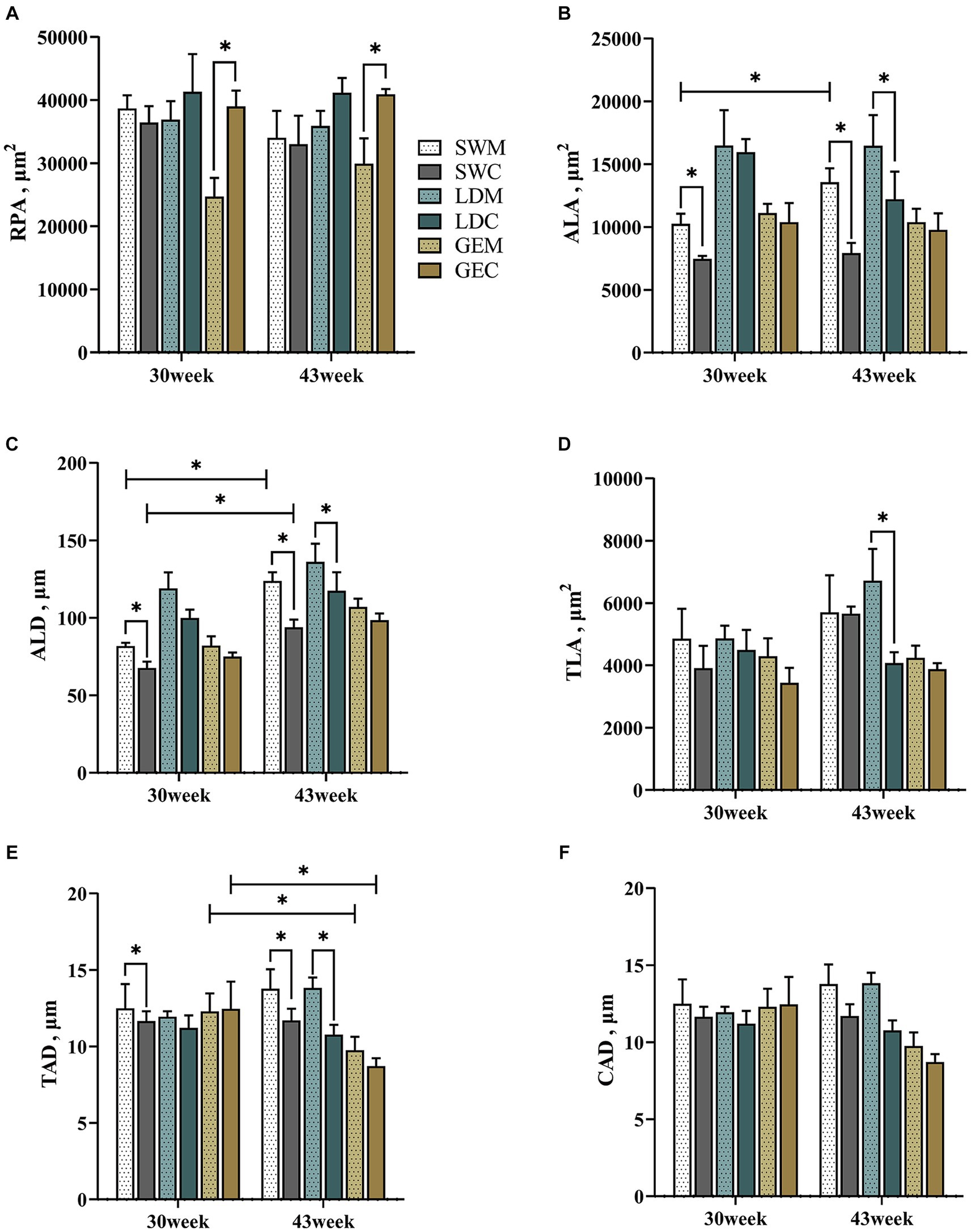 Figura 3. Comparación de los efectos de la MRS frente a la CRS sobre los parámetros histológicos del bazo entre tres razas de gansos. (A) Área de pulpa roja. (B) Área del corpúsculo esplénico. (C) Diámetro del corpúsculo esplénico. (D) Área de la trabécula esplénica. (E) Diámetro de la arteria trabecular. (F) Diámetro de la arteria central. «*» indica una diferencia significativa entre los dos grupos designados al nivel de p < 0,05. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; SWM; Ganso blanco de Sichuan bajo MRS; SWC, Ganso blanco de Sichuan bajo CRS; LDM, ganso de las Landas bajo MRS; LDC, ganso de las Landas en el CRS; GEM, Ganso de la pandilla bajo MRS; y GEC, Ganso de la pandilla bajo CRS.
Figura 3. Comparación de los efectos de la MRS frente a la CRS sobre los parámetros histológicos del bazo entre tres razas de gansos. (A) Área de pulpa roja. (B) Área del corpúsculo esplénico. (C) Diámetro del corpúsculo esplénico. (D) Área de la trabécula esplénica. (E) Diámetro de la arteria trabecular. (F) Diámetro de la arteria central. «*» indica una diferencia significativa entre los dos grupos designados al nivel de p < 0,05. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; SWM; Ganso blanco de Sichuan bajo MRS; SWC, Ganso blanco de Sichuan bajo CRS; LDM, ganso de las Landas bajo MRS; LDC, ganso de las Landas en el CRS; GEM, Ganso de la pandilla bajo MRS; y GEC, Ganso de la pandilla bajo CRS.
3.3 Efectos dependientes de la edad de MRS y CRS en los perfiles de expresión génica relacionados con el sistema inmunitario en el bazo entre tres razas de gansos
Además, exploramos las diferencias en los niveles de expresión de ARNm de 8 genes relacionados con el sistema inmunitario, incluidos HSPA2, MYD88, NF-κB1, NGF, PAK1, SPI1, VEGFA y WNT5A, durante el desarrollo del bazo de gansos criados bajo MRS y CRS. Como se muestra en la Figura 4, para SW, los niveles de ARNm de NGF, SPI1 y VEGFA fueron significativamente más altos bajo MRS que bajo CRS a las 43 semanas de edad (p < 0,05). Del mismo modo, se observaron niveles de expresión más altos de HSPA2 y NGF en los bazos de LD bajo MRS que en los de CRS (p < 0,05). Para GE, los niveles de expresión de VEGFA fueron significativamente más altos bajo MRS que bajo CRS a las 43 semanas de edad (p < 0,05), mientras que los niveles de expresión de MYD88 fueron significativamente más bajos bajo MRS que bajo CRS a las 30 semanas de edad (p < 0,05). Además, entre tres razas de gansos, la expresión de NF-κB1, PAK1 y WNT5A tendió a ser mayor bajo MRS en comparación con CRS (0,05 < p < 0,1), con una diferencia más pronunciada a las 43 semanas de edad.
 Figura 4. Comparación de los efectos de MRS versus CRS en los perfiles de expresión génica relacionados con el sistema inmunitario del bazo entre tres razas de gansos. Los perfiles de expresión de ARNm de (A) HSPA2, (B) MYD88, (C) NF-κB1, (D) NGF, (E) PAK1, (F) SPI1, (G) VEGFA y (H ) WNT5A en bazos de tres razas de gansos bajo dos sistemas de cría diferentes. «*» indica una diferencia significativa entre los dos grupos designados al nivel de p < 0,05. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; SWM; Ganso blanco de Sichuan bajo MRS; SWC, Ganso blanco de Sichuan bajo CRS; LDM, ganso de las Landas bajo MRS; LDC, ganso de las Landas en el CRS; GEM, Ganso de la pandilla bajo MRS; y GEC, Ganso de la pandilla bajo CRS.
Figura 4. Comparación de los efectos de MRS versus CRS en los perfiles de expresión génica relacionados con el sistema inmunitario del bazo entre tres razas de gansos. Los perfiles de expresión de ARNm de (A) HSPA2, (B) MYD88, (C) NF-κB1, (D) NGF, (E) PAK1, (F) SPI1, (G) VEGFA y (H ) WNT5A en bazos de tres razas de gansos bajo dos sistemas de cría diferentes. «*» indica una diferencia significativa entre los dos grupos designados al nivel de p < 0,05. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas; SWM; Ganso blanco de Sichuan bajo MRS; SWC, Ganso blanco de Sichuan bajo CRS; LDM, ganso de las Landas bajo MRS; LDC, ganso de las Landas en el CRS; GEM, Ganso de la pandilla bajo MRS; y GEC, Ganso de la pandilla bajo CRS.
3.4 Análisis de correlación entre los parámetros histológicos esplénicos del ganso y los perfiles de expresión génica en respuesta a diferentes sistemas de cría
Finalmente, se analizaron las correlaciones entre los parámetros histológicos esplénicos del ganso y los perfiles de expresión de genes relacionados con el sistema inmunológico bajo diferentes sistemas de cría. Como se muestra en la Figura 5 y en la Figura 1 suplementaria, en CRS, algunos parámetros histológicos esplénicos (incluidos el RPA, ALA, ALD y CAD) mostraron correlaciones negativas con el ARNm (perfiles de expresión de HSPA2, MYD88, WNT5A y PAK1) mientras que el TLA mostró correlaciones positivas con los perfiles de expresión de NF-κB1, HSPA2, MYD88 y PAK1 . Por el contrario, bajo MRS, parecía haber asociaciones positivas más pronunciadas entre los parámetros histológicos esplénicos (incluidos ALA, ALD, TLA y TAD) y los niveles de expresión de ARNm de HSPA2, MYD88, NGF, SPI1 y VEGFA.
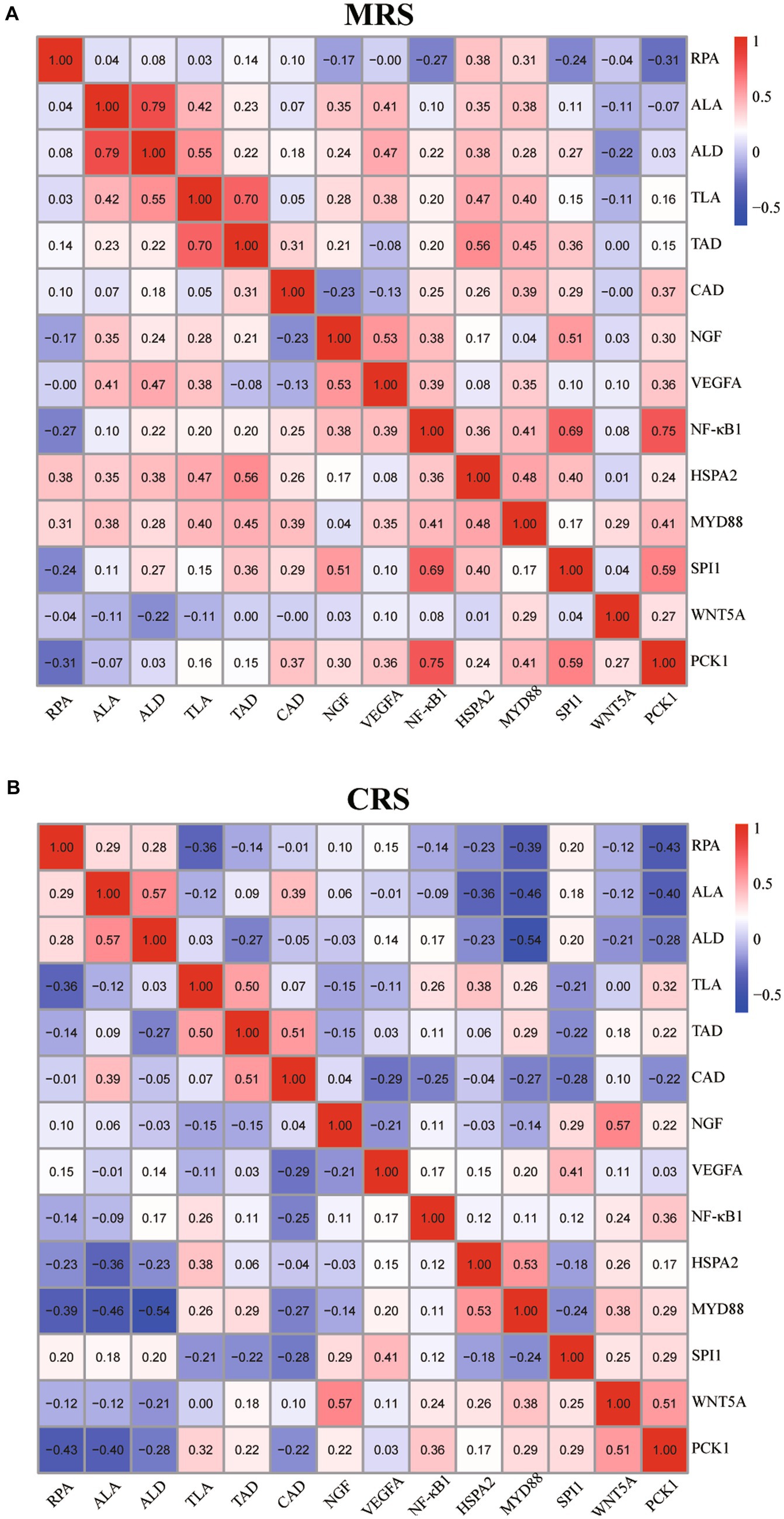
Figura 5. Análisis de correlación de los parámetros histológicos del bazo de ganso y de los perfiles de expresión génica relacionados con el sistema inmunitario bajo MRS (A) y CRS (B). Tanto para la MRS como para la CRS, n = 24. El color representa el coeficiente de correlación de Spearman. Abreviaturas: MRS, sistema de cría mixto de piso neto; CRS, sistema de cría en jaulas
4 Discusión
Se ha informado ampliamente que el sistema de cría tiene efectos significativos en la producción y la salud de las aves de corral. Sin embargo, en comparación con los pollos y los patos, se sabe poco sobre los efectos de los sistemas de cría en tierras secas en las respuestas inmunitarias de los gansos. En este trabajo, analizamos exhaustivamente las respuestas morfológicas, histológicas y moleculares esplénicas de tres razas de gansos a los dos sistemas de cría en tierras secas (MRS y CRS). Nuestro análisis morfológico reveló que el peso esplénico y el índice orgánico de SW fueron mayores en el CRS que en el MRS, mientras que no hubo diferencias significativas en el LD y el GE. El peso del timo y el índice de órganos de tres razas de gansos, especialmente para GE, mostraron una tendencia decreciente durante el desarrollo. En las aves de corral, tanto el bazo como el timo son vitales para mantener las funciones inmunitarias normales del cuerpo (23). Se informó que las gallinas criadas al aire libre generalmente tenían un peso del bazo y un índice de órganos más altos que las criadas en el interior (24). El peso del bazo y el timo de los patos criados bajo NRS fue mayor que el de los criados bajo FRS (13). Por lo tanto, se podría inferir que el sistema de cría en tierras secas tiene efectos significativos en el desarrollo esplénico del ganso, y la magnitud de estos efectos puede depender de la raza, la edad y su interacción.
Las observaciones histológicas mostraron que el bazo de ganso se puede dividir en dos compartimentos principales, que incluyen la pulpa roja que contiene sangre y la pulpa blanca que está llena de células linfoides. Se informó que la pulpa roja actúa como un filtro de sangre al extraer células envejecidas, muertas u opsonizadas de la circulación (25). Un estudio previo sobre el desarrollo postembrionario de la barrera sangre-bazo del pato mostró que la RPA aumentaba gradualmente a medida que el bazo se desarrollaba, al igual que las funciones de barrera biológica (19). En el presente estudio, se observó un mayor ARP en los bazos de GE criados bajo CRS que bajo MRS, lo que puede conducir a diferencias en sus funciones inmunes. El corpúsculo esplénico es donde se reúnen y maduran las células B, y la vaina linfática periarterial alrededor de la arteria central está formada por células T maduras. A través de estas estructuras histológicas únicas, el bazo puede montar respuestas inmunitarias complejas y eliminar patógenos de la sangre de manera efectiva (26). Un estudio reciente sobre patos ponedoras mostró que la inflamación resultaba en la proliferación de los corpúsculos esplénicos y linfocitos (27). Nuestros resultados mostraron que para SW y LD, el ALA, el ALD y el TAD fueron mayores bajo MRS que bajo CRS a las 30 o 43 semanas de edad, lo que implica que los gansos criados bajo MRS podrían exhibir funciones inmunes mejoradas que aquellos bajo CRS, particularmente para SW y LD. Se observó que, aunque los parámetros morfológicos e histológicos esplénicos del ganso se vieron afectados de manera diferente por los dos sistemas de cría en tierras secas, los parámetros histológicos podrían ser mejores indicadores de sus funciones inmunológicas.
Para revelar los mecanismos moleculares por los cuales el sistema de cría regula el desarrollo del bazo del ganso, examinamos más a fondo la expresión de varios genes clave relacionados con el sistema inmunológico. Entre ellas, HSPA2, un miembro de la familia de proteínas de choque térmico, puede proteger a las células del estrés adverso como el estrés oxidativo, la inflamación y la apoptosis. Mientras tanto, HSPA2 está implicado en la activación de las células presentadoras de antígenos, la presentación indirecta (o cebado cruzado) y la translocación de NF-κB a los núcleos (28). El NGF puede potenciar la actividad de las células inmunitarias mediante la activación de la señalización TLR4-NF-κB y otras vías antiinflamatorias en humanos, ratones y aves (29). Se informó que los niveles regulados al alza de SPI1 también podrían activar la señalización TLR4/NFκB al unirse al promotor de TLR4, lo que en consecuencia agravó la apoptosis celular, la inflamación y el reclutamiento de células inmunitarias (30). Además, existe evidencia de que el VEGFA estuvo implicado en la regulación de las respuestas inflamatorias crónicas a través de la vía TLR4-NF-κB (31). En particular, la vía NF-κB no solo es crucial para activar varias citoquinas proinflamatorias en varios tipos de células (32), sino que también sirve como un regulador clave de la inmunidad innata y adaptativa, la respuesta al estrés, la proliferación celular y la apoptosis (33). En este estudio, observamos que los niveles de expresión de ARNm de HSPA2, NGF, SPI1 y VEGFA se expresaron significativamente de manera diferencial en el bazo de gansos criados bajo MRS y CRS, lo que sugiere que el sistema de cría puede afectar el desarrollo del bazo de ganso y las funciones inmunes al regular la expresión de estos genes clave relacionados con el sistema inmunológico.
Además, MYD88 es una proteína adaptadora clave que media las vías de señalización iniciadas por los receptores tipo Toll (TLR) y la familia de receptores de interleucina-1 (IL-1R). (34). Tras la activación por componentes microbianos o citoquinas, MYD88 puede reclutar quinasas de la familia IRAK, lo que conduce a la activación de las vías de señalización posteriores, incluidas las vías de señalización MAPK y NF-κB (35). A través de estas vías, MYD88 está implicado en la activación de células inmunitarias del bazo, como los macrófagos, lo que favorece la producción de citoquinas inflamatorias y el inicio de la respuesta inmunitaria (36, 37). Nuestros resultados mostraron que, para la GE, los niveles de expresión del ARNm de MYD88 fueron más altos en el bazo de los gansos criados bajo CRS que en MRS, lo que sugiere un impacto potencial del sistema de cría en la respuesta inmune esplénica. Además, un estudio anterior informó que la sobreexpresión de PAK1 reguló las respuestas inmunes del ratón contra los patógenos al promover la polarización inflamatoria de los macrófagos a través de la activación de la señalización NF-κB (38). Se ha demostrado que el WNT5A, que se reconoce como un factor proinflamatorio, estimula la liberación de citoquinas proinflamatorias, la angiogénesis y la linfangiogénesis a través de las vías de señalización NF-κB y MAPK (39, 40). En este estudio, no hubo diferencias significativas en la expresión de PAK1 y WNT5A en el bazo de los gansos bajo MRS y CRS, lo que implica que podrían no mediar directamente los efectos del sistema de cría en el desarrollo del bazo del ganso.
Las estrechas relaciones entre las características histomorfológicas esplénicas, los perfiles de expresión de genes relacionados con el sistema inmunitario y las funciones inmunitarias del cuerpo se han descrito ampliamente en varios animales domésticos (13, 15). Del mismo modo, nuestros resultados mostraron que parecía haber asociaciones positivas más pronunciadas entre los parámetros histológicos esplénicos (incluidos el ALA, ALD, TLA y TAD) y los niveles de expresión de ARNm de varios genes clave relacionados con el sistema inmunitario (HSPA2, MYD88, NGF, SPI1 y VEGFA) en gansos criados bajo MRS que bajo CRS. Además, se encontró que los efectos dependientes de la edad del sistema de cría en los parámetros histológicos esplénicos del ganso y los perfiles de expresión de genes relacionados con el sistema inmunológico estaban fuertemente sincronizados, lo que sugiere que estos genes podrían desempeñar un papel crucial en la regulación de las funciones inmunes del ganso. Sus perfiles diferenciales de expresión del desarrollo pueden ser responsables de las diferencias en las respuestas inmunológicas de los gansos a MRS y CRS.
5 Conclusión
En conclusión, demostramos los efectos diferenciales dependientes de la edad del SRC y el SRM sobre las características histomorfológicas esplénicas y los perfiles de expresión de genes relacionados con el sistema inmunitario entre tres razas de gansos. Los resultados mostraron que los gansos criados bajo MRS podrían exhibir funciones inmunes mejoradas que los bajo CRS, particularmente para SW y LD. Aunque se supone que estas diferencias fenotípicas están asociadas con los perfiles de expresión diferencial dependientes de la edad de HSPA2, MYD88, NGF, SPI1 y VEGFA en el bazo de ganso, los mecanismos reguladores subyacentes esperan más investigaciones. Cabe señalar que la diferente densidad de población entre CRS y MRS, que sirve como una de las propiedades inherentes de los sistemas de cría en tierras secas, podría, al menos en parte, afectar la respuesta inmune del ganso. Sin embargo, este estudio proporciona información novedosa sobre la selección de sistemas adecuados de cría en tierras secas para mejorar el estado inmunológico del ganso y los mecanismos reguladores del sistema de cría en el desarrollo del bazo del ganso.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio están incluidas en el artículo/Material complementario, las consultas posteriores pueden dirigirse al autor correspondiente.
Declaración ética
Los estudios en animales fueron aprobados por el Comité de Ética y Bienestar Animal de la Universidad Agrícola de Sichuan. Los estudios se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
Control de calidad: Conceptualización, Curación de datos, Análisis formal, Administración de proyectos, Redacción – borrador original. YS: Curación de datos, Metodología, Redacción – borrador original. ZH: Conceptualización, Análisis formal, Escritura – borrador original. GY: Investigación, Metodología, Escritura – borrador original. JuW: Recursos, Software, Escritura – borrador original. XL: Metodología, Recursos, Software, Redacción – borrador original. WW: Adquisición de fondos, Recursos, Redacción – revisión y edición. XY: Investigación, Escritura – revisión y edición. JH: Recursos, Redacción – revisión y edición. HH: Administración de proyectos, redacción, revisión y edición. LL: Metodología, Redacción – revisión y edición. JiW: Adquisición de fondos, administración de proyectos, validación, redacción, revisión y edición. SH: Obtención de fondos, administración de proyectos, validación, visualización, redacción, revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Este estudio contó con el apoyo financiero del Programa de Ciencia y Tecnología de Sichuan (2023NSFSC0227), el Programa de Apoyo a la Tecnología Clave de la Provincia de Sichuan (2021YFYZ0014) y el Sistema de Investigación Agrícola de China del Ministerio de Ciencia y Pesca y el MARA (CARS-42-4).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1335152/full#supplementary-material
Referencias
1. Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO). Disponible en: https://www.fao.org/faostat/en/#data/QCL. (2022).
2. Shi, XW, Wang, JW, Zeng, FT y Qiu, XP. Los patrones de escisión del ADN mitocondrial distinguen el origen independiente de los gansos domésticos chinos y los gansos domésticos occidentales. Biochem Genet. (2006) 44:237–45. DOI: 10.1007/S10528-006-9028-Z
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Liu, BY, Wang, ZY, Yang, HM, Wang, JM, Xu, D, Zhang, R, et al. Influencia del sistema de cría en el rendimiento del crecimiento, las características de la canal y la calidad de la carne de los gansos de Yangzhou. Poult Sci. (2011) 90:653–9. doi: 10.3382/ps.2009-00591
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Chen, S, Xiang, H, Zhang, H, Zhu, X, Wang, D, Wang, J, et al. El sistema de cría provoca cambios en el comportamiento, el microbioma y la expresión génica de los pollos. Poult Sci. (2019) 98:3365–76. doi: 10.3382/ps/pez140
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Ocejo, M, Oporto, B, y Hurtado, A. Caracterización de la secuenciación del amplicón de 16s Rrna de la composición del microbioma cecal de pollos de engorde y pollos de crecimiento lento de corral a lo largo de su vida productiva. Sci Rep. (2019) 9:2506. doi: 10.1038/s41598-019-39323-x
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Bhimraj, AM, Popat, DS, Dinani, O, Babu, M, Rajani, A, y Valli, P. Efecto de los sistemas de cría sobre el rendimiento del crecimiento y las características de la canal del pollo desi. Int J Curr Microbiol App Sci. (2018) 7:3517–24. doi: 10.20546/ijcmas.2018.707.408
7. Zhang, C, Ah Kan Razafindrabe, RH, Chen, K, Zhao, X, Yang, L, Wang, L, et al. Efectos de diferentes sistemas de cría sobre el rendimiento del crecimiento, las características de la canal, la calidad de la carne y los parámetros bioquímicos séricos de los patos Chaohu. Anim Sci J. (2018) 89:672–8. doi: 10.1111/asj.12976
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Liao, SC, Lu, PX, Shen, SY, Hsiao, CC, Lien, CY, Wang, SD, et al. Efectos de diferentes condiciones de piscina y tipos de suelo sobre el rendimiento del crecimiento y la dermatitis de las almohadillas de los pies en gansos romanos blancos criados en interiores. Animales (Basilea). (2021) 11:1705. doi: 10.3390/ani11061705
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Li, X, Hu, S, Wang, W, Tang, B, Zheng, C, Hu, J, et al. Efectos del sistema de cría en jaulas frente al sistema de cría en el suelo sobre la histomorfología intestinal del ganso y la composición microbiana cecal. Poult Sci. (2022) 101:101931. doi: 10.1016/j.psj.2022.101931
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Smith, KG, y Hunt, JL. Sobre el uso de la masa del bazo como medida de la fortaleza del sistema inmunológico aviar. Ecología. (2004) 138:28–31. doi: 10.1007/s00442-003-1409-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Redmond, SB, Tell, RM, Coble, D, Mueller, C, Palic, D, Andreasen, CB, et al. Respuestas diferenciales de citoquinas esplénicas a la modulación inmunitaria de la dieta por diversas líneas de pollos. Poult Sci. (2010) 89:1635–41. doi: 10.3382/ps.2010-00846
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Yan, L, Lv, ZZ, An, S, Xing, K, Wang, ZG, Lv, MB, et al. Efectos del sistema de cría y Narasin en el rendimiento del crecimiento, el desarrollo gastrointestinal y la microbiota intestinal de pollos de engorde. Poult Sci. (2021) 100:100840. doi: 10.1016/j.psj.2020.10.073
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Guo, Y, Wang, Y, Liu, Z, Guo, X, Deng, Y, Ouyang, Q, et al. Efectos de los sistemas de cría sobre el rendimiento productivo, la capacidad antioxidante y el estado inmunológico de patos de carne a diferentes edades. Animal. (2021) 15:100199. doi: 10.1016/j.animal.2021.100199
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Kolluri, G, Ramamurthy, N, Churchil, RR, Dhinakar Raj, G, y Kannaki, TR. Influencia de la edad, el sexo y los sistemas de cría en el patrón de expresión del receptor tipo toll 7 (Tlr7) en el intestino, los pulmones y los tejidos linfoides de patos autóctonos. Br Poultry Sci. (2014) 55:59–67. doi: 10.1080/00071668.2013.867926
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Song, B, Li, P, Xu, H, Wang, Z, Yuan, J, Zhang, B, et al. Efectos del sistema de cría y el tratamiento con antibióticos sobre la función inmunitaria, la microbiota intestinal y los metabolitos de los pollos de engorde. J Anim Sci Biotechnol. (2022) 13:144. doi: 10.1186/s40104-022-00788-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. Byun, EB, Sung, NY, Byun, EH, Song, DS, Kim, JK, Park, JH, et al. El trímero C1 de procianidina inhibe la señalización de Mapk y Nf-Κb inducida por Lps a través de Tlr4 en macrófagos. Int Immunopharmacol. (2013) 15:450–6. doi: 10.1016/j.intimp.2012.11.021
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Sasaki, Y, e Iwai, K. Roles de la vía Nf-Κb en la biología de los linfocitos B. Señalización del receptor de células B. (2016) 393:177209. doi: 10.1007/82_2015_479
18. Feldman, AT, y Wolfe, D. Procesamiento de tejidos y tinción de hematoxilina y eosina. Métodos Mol Biol. (2014) 1180:31–43. doi: 10.1007/978-1-4939-1050-2_3
19. Xu, M, Li, W, Yang, S, Sun, X, Tarique, I, Yang, P, et al. Caracterización morfológica del desarrollo postembrionario de la barrera hemato-bazo en pato. Poult Sci. (2020) 99:3823–30. doi: 10.1016/j.psj.2020.05.012
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Juan, JL. El bazo aviar: un órgano descuidado. Q Rev Biol. (1994) 69:327–51. doi: 10.1086/418649
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Khodanovich, MY, Anan’ina, TV, Krutenkova, EP, Akulov, AE, Kudabaeva, MS, Svetlik, MV, et al. Desafíos y soluciones prácticas para la resonancia magnética y la comparación y las mediciones histológicas utilizando las herramientas de software disponibles de Imagej. Biomedicina. (2022) 10:1556. doi: 10.3390/biomedicines10071556
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Livak, KJ, y Schmittgen, TD. Análisis de datos relativos de expresión génica mediante Pcr cuantitativa en tiempo real y el método 2− Δδct. Métodos. (2001) 25:402–8. doi: 10.1006/meth.2001.1262
23. Li, JP, Xiong, X, Gan, XM, Pu, FJ, Ma, SC, Bai, LL, et al. El análisis del transcriptoma de la Bursa de Fabricius y el Thymus de patos ponedoras revela cambios en la expresión génica inmune que subyacen a los impactos de las densidades de población. Br Poultry Sci. (2021) 62:820–6. doi: 10.1080/00071668.2021.1943309
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. Shini, S, Shini, A y Huff, GR. Efectos de la administración crónica y repetida de corticosterona en la cría de pollos sobre la fisiología, el inicio de la puesta y la producción de huevos de las gallinas. Physiol Behave. (2009) 98:73–7. doi: 10.1016/j.physbeh.2009.04.012
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Lewis, SM, Williams, A, y Eisenbarth, SC. Estructura y función del sistema inmune en el bazo. Sci Immunol. (2019) 4:eaau6085. doi: 10.1126/sciimmunol.aau6085
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Cesta, MF. Estructura, función e histología normales del bazo. Toxicol Pathol. (2006) 34:455–65. doi: 10.1080/01926230600867743
27. Sun, X, Liu, E, Wang, T, Zhang, Q, Yang, P, Ahmed, N, et al. La nueva evidencia histológica de la barrera hemato-bazo en pato (Anas Platyrhynchos). Histol Histopathol. (2019) 34:33–45. doi: 10.14670/hh-18-019
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Li, Z, Menoret, A y Srivastava, P. Roles de las proteínas de choque térmico en la presentación de antígenos y la presentación cruzada. Curr Opin Immunol. (2002) 14:45–51. doi: 10.1016/s0952-7915(01)00297-7
29. Minnone, G, De Benedetti, F, y Bracci-Laudiero, L. Ngf y sus receptores en la regulación de la respuesta inflamatoria. Int J Mol Sci. (2017) 18:1028. doi: 10.3390/ijms18051028
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Liu, Z y Huang, S. La regulación positiva de Spi1 durante el infarto de miocardio agrava la lesión del tejido cardíaco y la progresión de la enfermedad a través de la activación del eje Tlr4/Nfκb. Am J Transl Res. (2022) 14:2709–27.
31. Wang, L, Zhao, Y, Qian, J, Sun, L, Lu, Y, Li, H, et al. Señalización del receptor tipo Toll-4 en el linfoma de células del manto: efectos sobre el crecimiento tumoral y la evasión inmunitaria. Cáncer. (2013) 119:782–91. doi: 10.1002/cncr.27792
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Bova, CA, Olsen, JC y Swanstrom, R. La familia de genes Env de retrovirus aviares: análisis molecular del rango de huéspedes y variantes antigénicas. J Virol. (1988) 62:75–83. doi: 10.1128/jvi.62.1.75-83.1988
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Kuwata, H, Matsumoto, M, Atarashi, K, Morishita, H, Hirotani, T, Koga, R, et al. Ikappabns inhibe la inducción de un subconjunto de genes dependientes de receptores tipo Toll y limita la inflamación. Inmunidad. (2006) 24:41–51. doi: 10.1016/j.immuni.2005.11.004
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Deguine, J, y Barton, GM. Myd88: un actor central en la señalización inmune innata. F1000Prime Rep. (2014) 6:97. doi: 10.12703/p6-97
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Zhou, M, Xu, W, Wang, J, Yan, J, Shi, Y, Zhang, C, et al. Aumento de la autofagia dependiente de Mtor, a través de la señalización ascendente de Tlr4-Myd88-Mapk y la vía descendente de Nf-Κb apaga la inflamación intestinal y la lesión por estrés oxidativo. EBioMedicina. (2018) 35:345–60. doi: 10.1016/j.ebiom.2018.08.035
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Bayer, AL, y Alcaide, P. Myd88: en el corazón de la señalización inflamatoria y la enfermedad cardiovascular. J Mol Cell Cardiol. (2021) 161:75–85. doi: 10.1016/j.yjmcc.2021.08.001
Resumen de PubMed | Texto completo de Crossref | Google Académico
37. Muzio, M, Ni, J, Feng, P y Dixit, VM. Irak-2 y Myd88, miembros de la familia Irak-2 y Myd88 como mediadores proximales de la señalización de Il-1. Ciencia. (1997) 278:1612–5. doi: 10.1126/science.278.5343.1612
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Zhang, W, Liu, H, Liu, W, Liu, Y y Xu, J. La pérdida mediada por Polycomb de MicroRNA let-7c determina la polarización inflamatoria de los macrófagos a través de la vía Nf-Κb dependiente de Pak1. La muerte celular difiere. (2015) 22:287–97. doi: 10.1038/cdd.2014.142
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Pereira, C, Schaer, DJ, Bachli, EB, Kurrer, MO y Schoedon, G. La señalización Wnt5a/Camkii contribuye a la respuesta inflamatoria de los macrófagos y es una diana para la acción antiinflamatoria de la proteína C activada y la interleucina-10. Arterioscler Tromb Vasc Biol. (2008) 28:504–10. doi: 10.1161/atvbaha.107.157438
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Shao, Y, Zheng, Q, Wang, W, Xin, N, Song, X y Zhao, C. Funciones biológicas de Wnt5a derivado de macrófagos y su papel en las enfermedades humanas. Oncotarget. (2016) 7:67674–84. doi: 10.18632/oncotarget.11874
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: ganso, sistema de cría de secano, bazo, histomorfología, expresión génica
Cita: Chen Q, Song Y, He Z, Yang G, Wang J, Li X, Wang W, Yuan X, Hu J, He H, Li L, Wang J y Hu S (2024) Efectos del sistema de cría mixto en jaula vs. suelo de red en la histomorfología del bazo de ganso y los perfiles de expresión génica. Frente. Vet. Sci. 11:1335152. doi: 10.3389/fvets.2024.1335152
Editado por:
Izhar Hyder Qazi, Universidad Shaheed Benazir Bhutto de Ciencias Veterinarias y Animales, Pakistán
Revisado por:
Servet Yalcin, Universidad Ege, Türkiye
Xiaoyu Huang, Universidad Agrícola de Gansu, China
Derechos de autor © 2024 Chen, Song, He, Yang, Wang, Li, Wang, Yuan, Hu, He, Li, Wang y Hu. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Shenqiang Hu, shenqiang.hu@sicau.edu.cn
†Estos autores han contribuido igualmente a este trabajo
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía