
Explorando el efecto aditivo de los flavonoides Ampelopsis grossedentata y Tween 80 en la alimentación de cabras nubias
Explorando el efecto aditivo de los flavonoides Ampelopsis grossedentata y Tween 80 en la alimentación de cabras nubias
 Junhong Zhu†
Junhong Zhu† Ying Lu†
Ying Lu† Zhendong GaoYuqing ChongMengfei Li
Zhendong GaoYuqing ChongMengfei Li Weidong Deng
Weidong Deng Dongmei Xi*
Dongmei Xi*- Laboratorio Clave Provincial de Nutrición y Piensos Animales de Yunnan, Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Yunnan, Kunming, China
Introducción: La prohibición de los antibióticos en la cría de animales subraya la necesidad crucial de contar con aditivos alimentarios seguros y naturales. Este estudio investiga los efectos de los flavonoides (AGF) de Ampelopsis grossedentata (AGF) y Tween 80 sobre el rendimiento del crecimiento, los índices sanguíneos y la microbiota ruminal de cabras nubias, evaluando su potencial como aditivos alimentarios alternativos en el manejo del ganado.
Métodos: Treinta y dos cabras fueron divididas aleatoriamente en cuatro grupos. Al grupo control (grupo CON) se le proporcionó una dieta basal, mientras que los grupos experimentales recibieron dietas suplementadas con diversos aditivos dietéticos durante 100 días: una dieta basal suplementada con 25 mg/kg de monensina (grupo MN), una dieta basal que contenía 2,0 g/kg de flavonoides Ampelopsis grossedentata (grupo AGF) o una dieta basal que contenía 7,5 ml/kg de Tween 80 (grupo TW). Se recolectaron muestras de sangre y líquido ruminal para su análisis al final del período de alimentación. El rendimiento del crecimiento se monitoreó a través de pesajes regulares y mediciones de consumo de alimento. Los índices sanguíneos se analizaron utilizando técnicas bioquímicas estándar, mientras que la composición microbiana del líquido ruminal se determinó mediante la secuenciación del gen 16S rRNA de alto rendimiento para evaluar la diversidad y función microbiana. A continuación, se evaluaron los efectos de los tratamientos dietéticos sobre el rendimiento del crecimiento, los índices sanguíneos y la composición microbiana del rumen.
Resultados: El grupo AGF exhibió un aumento significativo de la ganancia diaria promedio y una disminución de la relación alimento-ganancia (p < 0.05). El análisis de los índices sanguíneos no reveló diferencias entre los grupos CON y AGF, ya que ambos mostraron mayores concentraciones de triglicéridos, colesterol de lipoproteínas de baja densidad, transaminasa glutámico-pirúvica, fosfatasa alcalina y lactato deshidrogenasa en comparación con el grupo de monensina (p < 0,05). El grupo TW tuvo niveles significativamente más altos de glucosa, transaminasa glutámico-oxaloacética y transaminasas glutámico-pirúvicas que el grupo MN (p < 0,05). El análisis de la diversidad microbiana reveló que el grupo TW tenía una diversidad alfa significativamente mayor que otros grupos, mientras que el análisis de la diversidad beta mostró una mayor similitud entre la microbiota ruminal de los grupos AGF y CON. El análisis de LEfSe identificó a Proteobacteria, Deferribacteres, Ehryarchaeo y Elusimicrobia como biomarcadores que distinguen la microbiota ruminal entre los grupos. En conclusión, la suplementación con AGF aumentó la abundancia relativa de bacterias beneficiosas en el rumen de las cabras de Nubia y, por lo tanto, mejoró el rendimiento del crecimiento. La suplementación con TW aumentó significativamente la diversidad y abundancia microbiana del rumen, lo que sugiere beneficios para la salud del rumen a pesar de la mala palatabilidad. Estos hallazgos ponen de relieve el potencial del AGF como un nuevo aditivo ecológico con importantes implicaciones para la eficiencia y el desarrollo de la ganadería.
1 Introducción
Los aditivos antibióticos se han utilizado ampliamente en la cría de animales (1). Sin embargo, debido a problemas como los residuos de medicamentos, su uso ha sido prohibido en muchos países (2). Esta prohibición ha planteado importantes desafíos para la ganadería en su conjunto, lo que subraya la necesidad crítica de explorar aditivos alimentarios naturales, ecológicos y seguros. El objetivo de este estudio fue evaluar los efectos de dos posibles alternativas, Ampelopsis grossedentata flavone (AGF), derivada de extractos naturales de plantas, y Tween 80, ampliamente utilizada como emulsionante, como aditivos alimentarios sobre el rendimiento del crecimiento, los índices sanguíneos y la microbiota ruminal en cabras nubias.
AGF, acrónimo de flavonoides Ampelopsis grossedentata, representa un grupo de extractos de plantas naturales derivadas de la vid Ampelopsis grossedentata, conocida por sus importantes propiedades antioxidantes. Su componente principal es la dihidromiricetina (3), que tiene diversas funciones fisiológicas como antiinflamatoria, antiestrés oxidativo (4), antibacteriana (5), antiapoptosis (6), regulación inmunitaria (7), y desempeña un papel importante en la mejora del rendimiento del crecimiento animal y la mejora de la inmunidad animal (8). Informes recientes indican que el AGF no solo muestra una fuerte actividad antioxidante, sino que también tiene excelentes propiedades protectoras del hígado que pueden convertirse en productos destinados a la antidiabetes, antioxidantes y protección hepática (9-11). El estudio encontró que el té de ratán previno los trastornos metabólicos causados por una dieta alta en grasas al mejorar la homeostasis de la glucosa en ratas. La investigación de Wang y sus colegas encontró que las cualidades antioxidantes del AGF juegan un papel protector en el mantenimiento de la integridad de la barrera intestinal dentro de la línea celular epitelial porcina (12). Zhu et al. encontraron que la incorporación de AGF en la dieta no solo mejora el ecosistema microbiano, sino que también tiene un potencial significativo para promover el crecimiento del ganado juvenil, contribuyendo así a las ganancias económicas (13). Sin embargo, el papel específico del AGF como aditivo alimentario en rumiantes aún requiere más investigación.
El nombre químico de Tween 80 es monooleato de sorbido de polioxietileno. Puede aumentar la cantidad de liberación de enzimas y mejorar la actividad enzimática, promover la interacción entre la enzima y el sustrato (14). Es ampliamente utilizado como emulsionante en el procesamiento de alimentos (15). El Tween 80 de grado alimenticio generalmente se considera seguro como aditivo para piensos (16). Los experimentos han demostrado que Tween 80 tiene un cierto efecto regulador sobre los microorganismos del rumen, lo que puede mejorar la actividad de la celulasa y promover el crecimiento de bacterias no celulolíticas (17, 18). El estudio de Martha encontró que Tween 80 y su derivado ácido oleico promovían el crecimiento de corynebacterium accolens (19). La investigación ha demostrado que la adición del surfactante Tween-80 aumenta eficazmente la capacidad natural de las bacterias para biodegradar el naftaleno (20). Sin embargo, solo unos pocos estudios han verificado el efecto de Tween 80 en rumiantes, con particularmente pocos informes sobre su aplicación en la producción de ovejas de carne.
Por lo tanto, el objetivo de este estudio fue investigar exhaustivamente los efectos de la suplementación dietética con AGF y Tween 80 sobre el rendimiento del crecimiento, los índices sanguíneos y la microbiota ruminal en cabras nubias, así como evaluar la viabilidad de estos dos aditivos como alternativas a la monensina. A través de este estudio, esperamos proporcionar una base teórica para la promoción y aplicación de estos aditivos alimentarios naturales y seguros en la industria ganadera.
2 Materiales y métodos
El estudio se llevó a cabo en la granja del condado de Zhaotong, provincia de Yunnan, China. Enclavada en el terreno montañoso medio de la montaña Houlong en las coordenadas N27°31′13″, E103°54′51″, la granja se alza orgullosa a una altitud de 1.614 m. Esta localidad experimenta un clima climático caracterizado por altos niveles de humedad que oscilan entre el 20% y el 60% de humedad relativa, y una inclinación por las precipitaciones. A lo largo de los meses de verano, de junio a septiembre, las temperaturas diarias alcanzan un máximo de 26 °C. Sin embargo, un delta de temperatura diurna conspicuo. La propuesta de investigación y los procedimientos experimentales pertinentes fueron aprobados por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Agrícola de Yunnan (Número de caso: 202200301).
2.1 Animales y diseño de experimentos
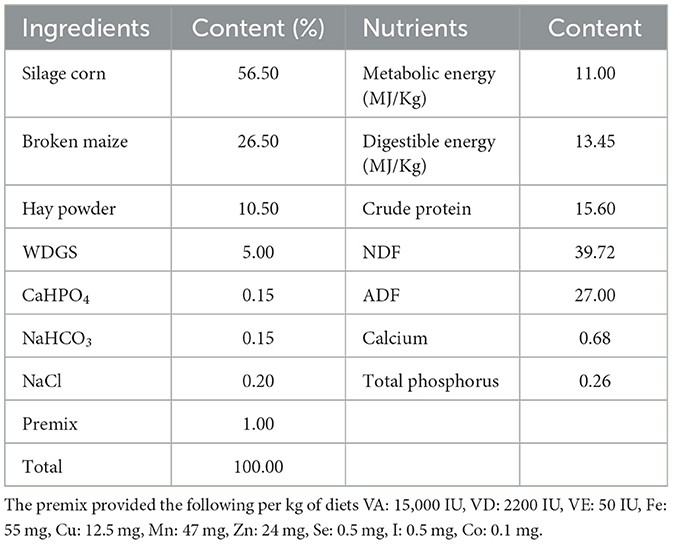
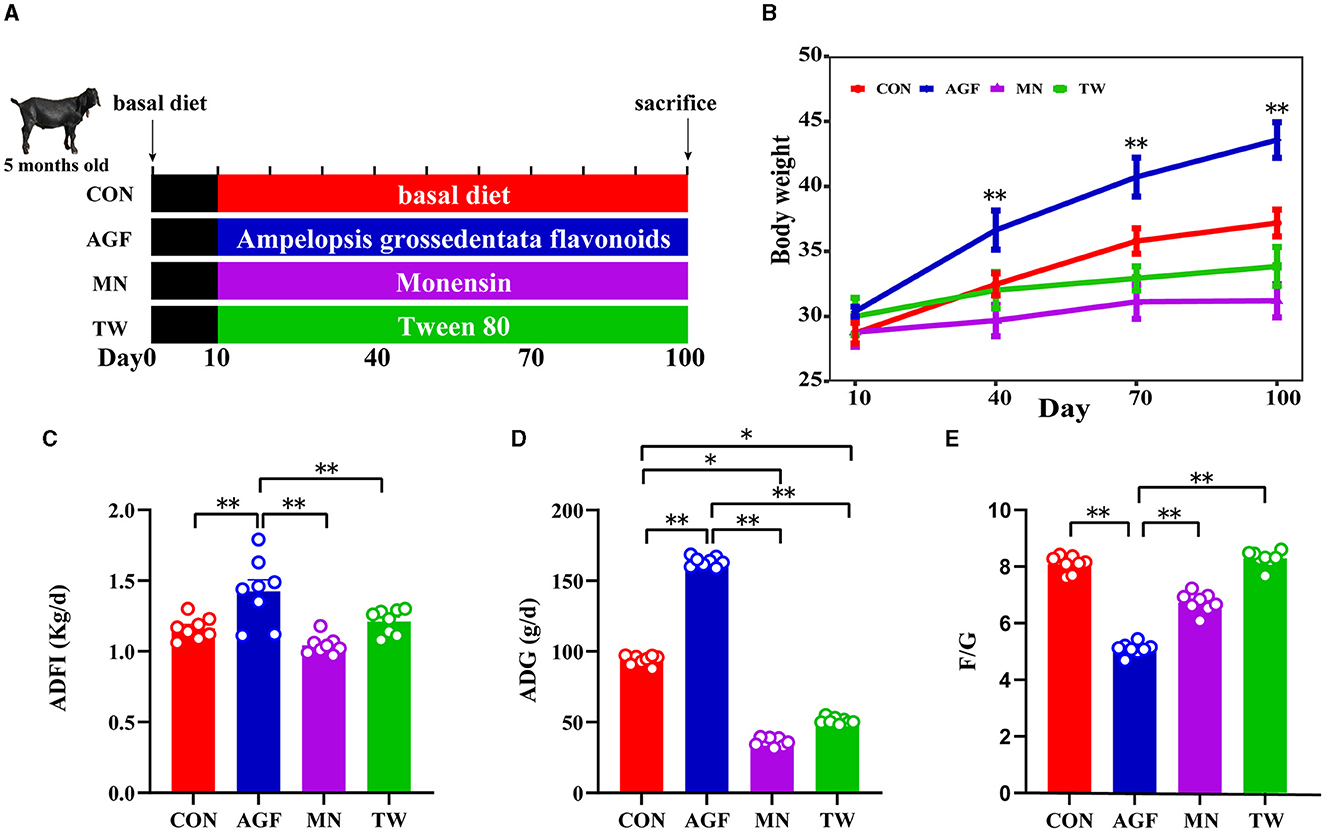
El estudio se llevó a cabo utilizando un enfoque de diseño completamente aleatorizado. Se formaron cuatro grupos a partir de un total de treinta y dos cabras nubias (media inicial ± EE; 27 ± 1,39 kg de peso corporal y 5 ± 0,5 meses de edad) designadas para el engorde, cada grupo compuesto por ocho cabras (mitad macho y mitad hembra). Las formulaciones de la dieta basal seguidas en este estudio se basaron en las directrices proporcionadas por el NRC (21) y NY/T816-2004 (Tabla 1). El grupo control (grupo CON) fue alimentado con una dieta basal estándar. Por el contrario, los grupos experimentales recibieron variaciones de esta dieta: uno con una adición de 2,0 g/kg de flavonoides Ampelopsis grossedentata (grupo AGF), otro con 25 mg/kg de monensina (grupo MN) y el último mejorado con 7,5 mL/kg de Tween 80 (grupo TW). Alimentamos y manejamos uniformemente a todas las cabras experimentales. Con una duración de 100 días, el experimento se inició con una fase preparatoria de 10 días dedicada a la vacunación, la desparasitación, los ajustes de manejo y las transiciones dietéticas. La disposición específica se muestra en la Figura 1A. Las cabras fueron alimentadas dos veces al día, a las 8:00 y a las 16:00 h con libre acceso a agua dulce.
2.2 Recogida de muestras
Eldía 101, a las 07:00 horas, se recogieron muestras de sangre de las venas yugulares de ocho cabras de cada grupo, utilizando tubos vacutainer. Estas muestras se centrifugaron inmediatamente a 3.000 rpm y a una temperatura de 4 °C. Después de la centrifugación, separamos el plasma y lo almacenamos a -20 °C para futuros análisis. Además, el mismo día, antes de la alimentación, recogimos muestras de fluido ruminal de seis cabras de cada grupo, seleccionadas por su peso más cercano a la media del grupo, utilizando una bomba de vacío. Para el análisis microbiano, empleamos secuenciación de alto rendimiento para identificar secuencias microbianas únicas en la región 16S (22).
2.3 Evolución del crecimiento
Se llevaban registros diarios del consumo de alimento. En los puntos horarios programados (07:00)los días 11, 41, 71y 101, las cabras se someten a una evaluación de peso después del ayuno. El índice de conversión alimenticia se determinó aplicando la fórmula estándar.
2.4 Fisiología y bioquímica del plasma
Se utilizó un analizador bioquímico automatizado (Roche, Basilea, Suiza) para medir la fisiología del plasma y los índices bioquímicos. Estos incluyeron triglicéridos (TG), proteínas totales (TP), glucosa (GLU), UREA, colesterol total (CT), colesterol de lipoproteínas de alta densidad (HDL-CH), colesterol de lipoproteínas de baja densidad (LDL-CH), globulina (GLOB), albúmina (ALB), alanina aminotransferasa (ALT), aspartato aminotransferasa (AST), fosfatasa alcalina (ALP) y lactato deshidrogenasa (LDH).
2.5 Secuenciación del gen ARNr 16S
El ADN microbiano se extrajo utilizando los kits de ADN de suelo HiPure (Magen, Guangzhou, China) de acuerdo con los protocolos del fabricante. Se empleó el espectrofotómetro NanoDrop 2000 (Thermo Fisher Scientific, Madison, WI, EE.UU.) para la cuantificación del ADN, asegurando la obtención de ADN genómico de alta calidad en cantidades adecuadas (23). A continuación, las regiones V3-V4 de los genes 16S de ADNr se amplificaron mediante PCR, utilizando cebadores específicos directos (5′-CCTACGGGNGGCWGCAG-3′) e inversos (5′-GGACTACHVGGGTATCTAAT-3′) (24). Se utilizó la PCR para amplificar el gen del ARN ribosómico, con un protocolo que comenzó a 94 °C durante 2 min, seguido de 30 ciclos de 98 °C durante 10 s, 62 °C durante 30 s y 68 °C durante 30 s, culminando en una extensión final a 68 °C durante 5 min. Los amplicones resultantes se extrajeron de geles de agarosa al 2% y se sometieron a purificación a través del kit de extracción de gel de ADN AxyPrep (Axygen Biosciences, Union City, CA), según las pautas proporcionadas por el fabricante. La cuantificación se llevó a cabo mediante el sistema de PCR en tiempo real ABI StepOnePlus (Life Technologies, Foster City, CA). Después de la purificación, los amplicones se sometieron a una secuenciación de extremos pareados en la plataforma Illumina (PE250) siguiendo procedimientos estándar.
Se empleó FASTP (versión 0.18.0) (25) para filtrar los datos brutos de la plataforma Illumina, lo que dio como resultado lecturas limpias para el posterior análisis de ensamblaje. Se utilizó el software FLASH (versión 1.2.11) (26) para fusionar lecturas limpias en etiquetas basadas en una superposición mínima de 10 pb y una tasa máxima de discordancia del 2%. Siguiendo los criterios de filtrado descritos en la literatura (27), se empleó QIIME (versión 1.9.1) (28) para filtrar aún más las etiquetas de baja calidad, lo que produjo un conjunto de etiquetas limpias de alta calidad. Utilizando el algoritmo UCHIME (29) con una base de datos de referencia (versión r20110519), se realizó la comprobación de quimeras en las etiquetas limpias. Las etiquetas limpias resultantes, el filtrado posterior a la quimera, se sometieron a un análisis posterior. Utilizando el software UPARSE (versión 7.1) (24), las unidades taxonómicas operativas (OTU) se sometieron a un reagrupamiento con un umbral de similitud de secuencia del 97% (30). Las secuencias quiméricas fueron identificadas y eliminadas empleando el algoritmo de Uchime (31, 32).
El Análisis de Componentes Principales (ACP) basado en las tablas de abundancia de OTU se llevó a cabo utilizando el paquete Vegan en lenguaje R (versión 2.5.3) (33). El mapa de calor de abundancia de especies se generó utilizando el paquete de mapas de calor del lenguaje R (versión 1.0.12) (34). El análisis de correlación de especies de Pearson se calculó utilizando el paquete psych (versión 1.8.4) (35). El grafo de la red de correlación de especies se construyó utilizando el paquete de grafos del lenguaje R (versión 1.1.2) (36). Los índices de diversidad alfa, incluidos Chao1 y Simpson, se calcularon utilizando QIIME versión 1.9.1 (28). Los índices de diversidad, incluidas las curvas de dilución y las curvas de rango-abundancia, se visualizaron a través del paquete ggplot2 en R (versión 2.2.1) (37). Los análisis de escalamiento multidimensional no métrico (NMDS) se llevaron a cabo utilizando el paquete vegan (versión 2.5.3) en R, y los gráficos resultantes se generaron a través del paquete ggplot2 (versión 2.2.1) (38, 39). El análisis de Upset Intergrupal, dirigido a identificar especies únicas y compartidas, se realizó utilizando el paquete UpSetR en R (40). Se utilizó el software LEfSe (versión 1.0) para identificar las características de los biomarcadores dentro de cada grupo (41). Se empleó PICRUSt2 (versión 1.0) para inferir el análisis de la vía KEGG de las OTU (35). Las diferencias entre los grupos se evaluaron a través de la prueba t de Welch y la prueba de suma de rangos de Wilcoxon para comparaciones por pares, junto con la prueba de diferencia honestamente significativa (HSD) de Tukey y la prueba H de Kruskal-Wallis para análisis de grupos múltiples, todas realizadas utilizando el paquete vegano (versión 2.5.3) en R (33).
2.6 Análisis de datos
Se analizó el impacto de cuatro dietas sobre el rendimiento del crecimiento, los metabolitos plasmáticos, el pH ruminal y la microbiota ruminal de las cabras mediante un ANOVA de una vía en el programa SPSS 25.0 (IBM Corp, Armonk, NY, EE.UU.), y los resultados se presentaron como medias ± EE agrupados (42). Las pruebas de rangos múltiples de LSD y Duncan facilitaron las comparaciones múltiples de medias, categorizando las diferencias como extremadamente significativas (p < 0,01), significativas (p < 0,05) o no significativas (p > 0,05) en función de los valores de p.
3 Resultados
3.1 Evolución del crecimiento
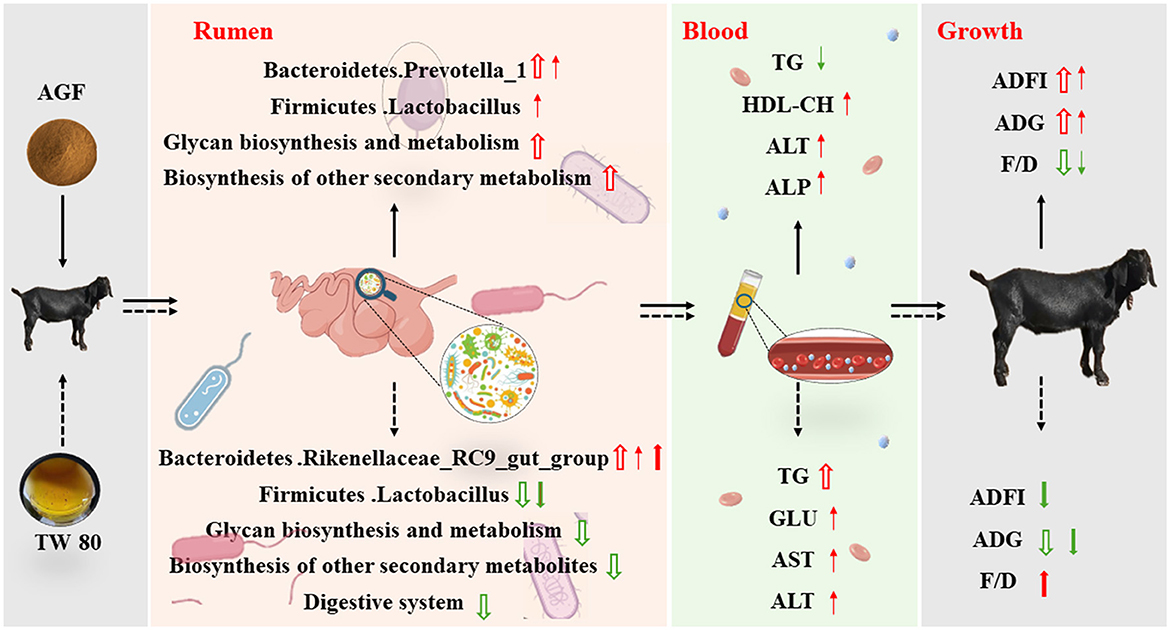
La comparación inicial del peso corporal no indicó diferencias significativas entre los grupos CON, AGF, MN y TW (p > 0,05). Sin embargo, una ingesta de 90 días de flavonoides Ampelopsis grossedentata condujo a un aumento significativo en el peso corporal en cabras en comparación con el grupo CON (Figura 1B). La ingesta media diaria de alimento (ADFI) y la ganancia media diaria (GMD) en el grupo AGF fueron significativamente superiores a las del grupo CON, así como a las de los grupos MN y TW (p < 0,01; Figuras 1C y D). Además, la relación alimentación-ganancia (F/G) en el grupo AGF fue significativamente menor que la del grupo CON, y también significativamente menor que la de los grupos MN y TW (p < 0,01; Figura 1E).
3.2 Índices sanguíneos
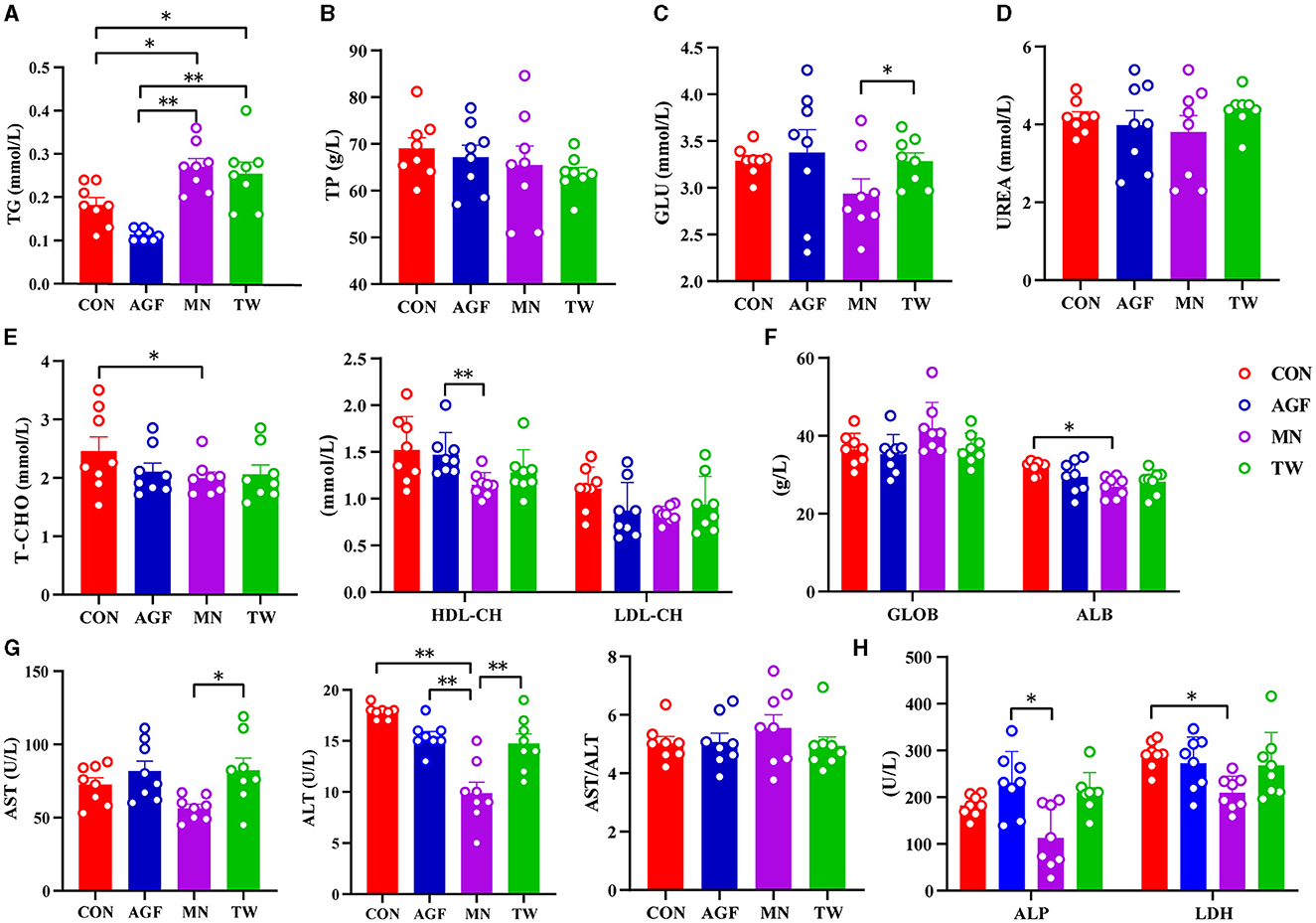
No hubo diferencias significativas en las concentraciones plasmáticas de TP y UREA entre los cuatro grupos (p> 0,05) (Figuras 2B, D). Al final del experimento, no hubo diferencias significativas en la concentración plasmática de TG entre los grupos AGF y CON (p > 0,05). Sin embargo, la concentración de TG en ambos grupos fue significativamente mayor que en los grupos MN y TW (p < 0,05; Figura 2A). No hubo diferencias significativas en las concentraciones plasmáticas de T-CHO y ALB entre los grupos AGF, TW y CON (p > 0,05). Sin embargo, el grupo CON tuvo concentraciones significativamente más altas de T-CHO y ALB que el grupo MN (p < 0,05). Las diferencias en las concentraciones plasmáticas de LDL-CH entre los grupos AGF, TW y CON no fueron significativas (p > 0,05). Sin embargo, la concentración de LDL-CH en el grupo AGF fue significativamente mayor que en el grupo MN (p < 0,05; Figuras 2E, F). No hubo diferencias significativas (p > 0,05) en las concentraciones plasmáticas de GLU, AST y ALT entre los grupos AGF, TW y CON. Las concentraciones de GLU, AST y ALT en el grupo MN fueron significativamente menores que las del grupo TW (p < 0,05), pero no hubo diferencias significativas (p > 0,05) entre los grupos MN y AGF. En particular, la concentración plasmática de ALT en el grupo MN fue significativamente menor que en los grupos CON y AGF (Figuras 2C, G). Las concentraciones de ALP y LDH en el grupo MN fueron significativamente menores que las del grupo AGF y en el grupo CON, respectivamente (p < 0,05) (Figura 2H).
3.3 Composición y diversidad de la microbiota ruminal
3.3.1 pH del rumen
Se midió el pH ruminal en diferentes grupos, arrojando valores de 6,71 ± 0,03 (CON), 6,62 ± 0,02 (AGF), 6,42 ± 0,05 (MN) y 6,61 ± 0,06 (TW), respectivamente. El análisis indicó que los diversos aditivos no afectaron significativamente el pH ruminal en cabras (p > 0,05).
3.3.2 Análisis de la diversidad microbiana en el rumen
La cobertura de The Good de esta secuenciación superó el 99%, oscilando entre el 99,70% y el 99,84% (Figura 3A). Se realizó un análisis de diversidad en las OTU para comprender mejor la riqueza y diversidad de la composición microbiana entre los grupos CON, AGF, MN y TW. En comparación con el grupo CON, los cinco índices de diversidad α en el grupo AGF fueron significativamente más bajos (p < 0,05). Por el contrario, no hubo diferencias significativas en los cinco índices de α diversidad entre los grupos TW y CON (p > 0,05). La α diversidad en el grupo AGF fue significativamente menor que en los grupos MN y TW (p < 0,05). La riqueza de especies microbianas en el grupo AGF disminuyó significativamente (p < 0.05; Figura 3B), mientras que no hubo diferencia significativa en la uniformidad (p > 0,05; Figura 3C). El análisis de coordenadas principales (PCoA) es un método de reducción de dimensionalidad que emplea un plano bidimensional para visualizar las distancias entre muestras. Las coordenadas 1 representan el 14,44% de la variación y las coordenadas 2 representan el 11,11% de la varianza (Figura 3D). La composición microbiana ruminal mostró mayor similitud entre los grupos TW y MN.
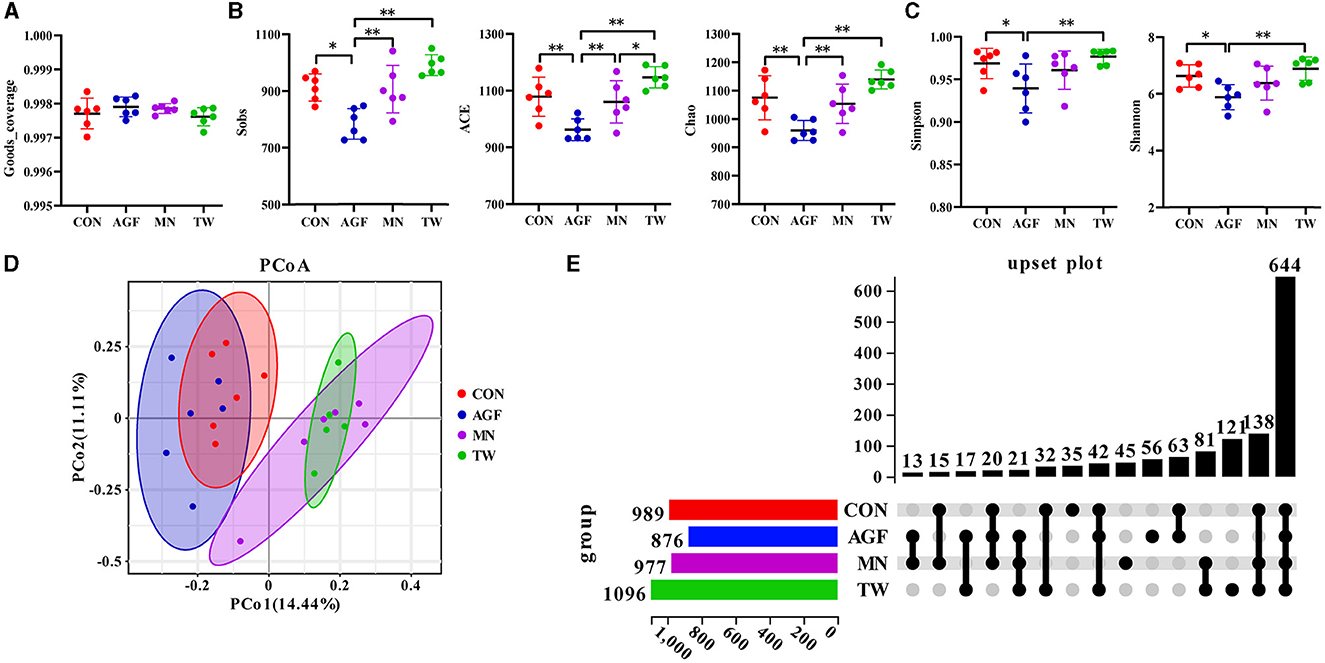
3.3.3 Clúster de OTU
Tras la exclusión de las etiquetas consideradas de baja calidad o carentes de importancia biológica, el análisis de las 32 muestras arrojó un total de 2.879.868 etiquetas efectivas. Cada muestra generó entre 110.629 y 128.204 etiquetas efectivas, con una tasa efectiva superior al 92% (Tabla Suplementaria S1). Se observaron un total de 989, 876, 977 y 1.096 unidades taxonómicas operativas (OTU) en los grupos CON, AGF, MN y TW, siendo 35, 56, 45 y 121 OTU exclusivas de los grupos CON, AGF, MN y TW, respectivamente (Figura 3E). Hubo 644 OTU comunes a los cuatro grupos, y las intersecciones de los grupos CON y AGF, MN, TW contenían 63, 15 y 32 OTU, respectivamente (Figura 3E).
3.3.4 Composición microbiana
Dentro del nivel de filo, Bacteroidetes y Firmicutes constituyeron los filos predominantes, representando colectivamente más del 90% de los microorganismos en los cuatro grupos (Figura 4A), y sus abundancias relativas oscilaron entre el 33,38% y el 76,59% y el 18,07% y el 56,22%, respectivamente (Tabla Suplementaria S2). En comparación con el grupo CON, no hubo diferencias significativas en la abundancia relativa de Bacteroides y Firmicutes en los grupos AGF y TW (p > 0,05). Además, las abundancias relativas de Proteobacteria y Kiritimatiellaeota en el grupo AGF no mostraron diferencias significativas (p > 0.05). Sin embargo, en el grupo TW, la abundancia relativa de Proteobacteria disminuyó significativamente, mientras que la abundancia relativa de Kiritimatiellaeota aumentó significativamente (p < 0.05). En comparación con el grupo MN, las abundancias relativas de Bacteroides y Proteobacteria fueron significativamente mayores en el grupo AGF (p < 0,01), mientras que las abundancias relativas de Firmicutes y Kiritimatiellaeota fueron significativamente menores (p < 0,05). No hubo diferencias significativas en las abundancias relativas de estos taxones en el grupo TW (p > 0.05) (Figura 4B).
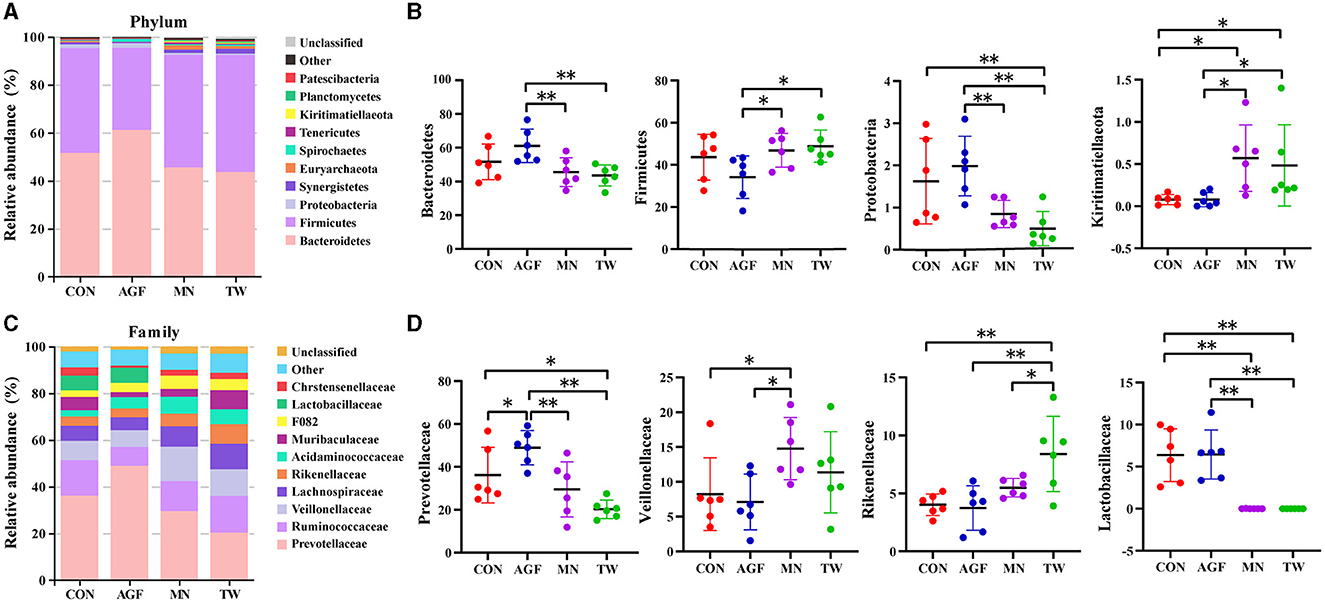
A nivel familiar, Prevotellaceae emergió como la microbiota dominante (Figura 4C). En comparación con el grupo CON, la abundancia relativa de Prevotellaceae en el grupo AGF aumentó significativamente, mientras que la del grupo TW disminuyó significativamente (p < 0,05). En comparación con el grupo MN, la abundancia relativa de Prevotellaceae en el grupo AGF aumentó significativamente (p < 0,05), pero no en el grupo TW (p > 0,05). En comparación con el grupo CON, la abundancia relativa de Veillonellaceae, Rikenellaceae y Lactobacillaceae en el grupo AGF no fue significativamente diferente (p > 0,05), mientras que la abundancia relativa de Rikenellaceae en el grupo TW aumentó significativamente (p < 0,05). La abundancia relativa de Lactobacillaceae disminuyó significativamente (p < 0,01). En comparación con el grupo MN, la abundancia relativa de Veillonellaceae en el grupo AGF disminuyó significativamente, y la abundancia relativa de Lactobacillaceae aumentó significativamente (p < 0,05). La abundancia relativa de Rikenellaceae en el grupo TW se incrementó significativamente, y la abundancia relativa de Lactobacillaceae disminuyó significativamente (p < 0.05) (Figura 4D).
Dentro del nivel de género, en comparación con el grupo CON, la abundancia relativa de Prevotella_1 en el grupo AGF es significativamente mayor (p < 0.05), la abundancia relativa de Rikenellaceae_RC9_gut_group y Lactobacillus en el grupo TW no es significativamente diferente, y la abundancia relativa de Prevotella_1 en el grupo TW no es significativamente diferente (p > 0.05) (Figuras 5A, B). La abundancia relativa de Rikenellaceae_RC9_gut_group se incrementó significativamente, y la abundancia relativa de Lactobacillus disminuyó significativamente (p < 0.05). En comparación con el grupo MN, la abundancia relativa de Prevotella_1 y Lactobacillus en el grupo AGF aumentó significativamente (p < 0,05), mientras que la abundancia relativa de Rikenellaceae_RC9_gut_group no fue significativamente diferente (p > 0,05) (Figura 5B).

Se realizó un análisis de correlación entre los géneros microbianos ruminales dominantes en el grupo AGF y varios parámetros fisiológicos y bioquímicos en plasma, y los resultados se mostraron en la Figura 5C. Veillonellaceae_UCG-001 se correlaciona significativamente positivamente con GLU (p < 0,05). Succiniclasticum se correlaciona significativamente negativamente con ALT (p < 0,05). El Ruminococcaceae_NK4A214_group y Prevotellaceae_UCG-001 mostraron una correlación positiva significativa con el HDL-CH (p < 0,01). Por el contrario, el Eubacterium_coprostanoligenes_group mostró una correlación negativa significativa con la UREA y la AST (p < 0,01).
3.3.5 Análisis de la microbiota diferencial ruminal
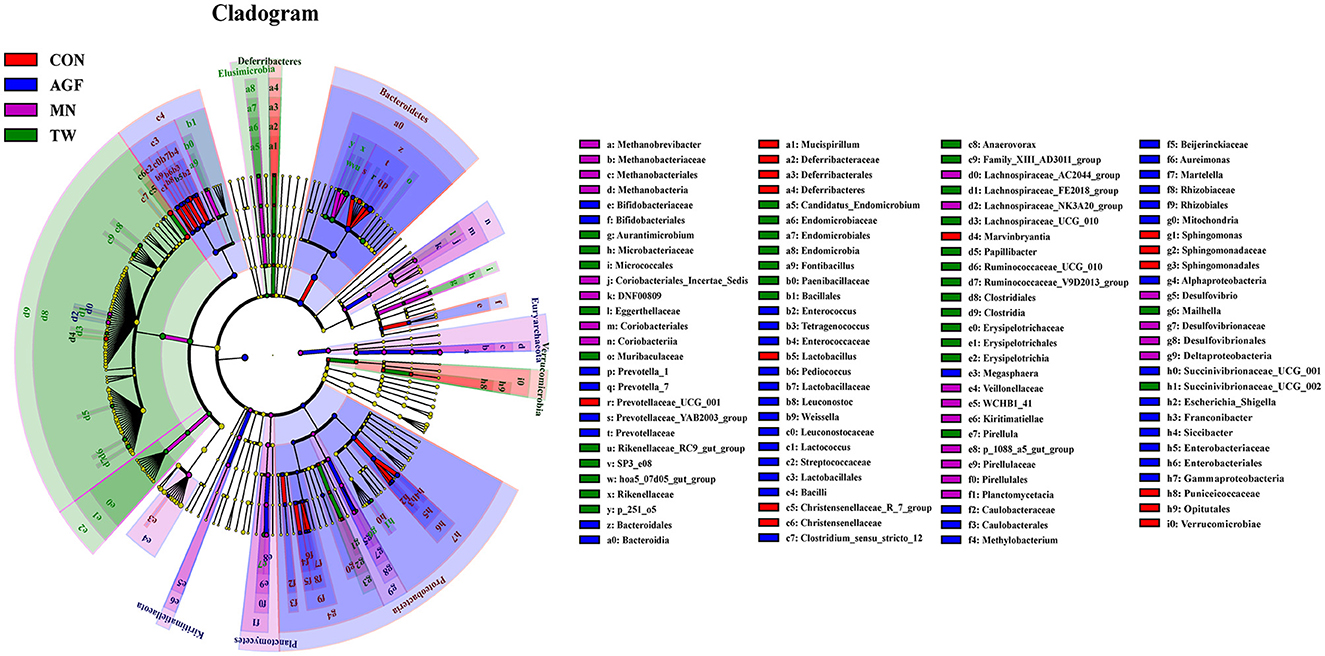
En los grupos AGF, CON, MN y TW, se identificaron un total de 117 biomarcadores en varios niveles taxonómicos. En particular, Proteobacteria, Deferribacteres, Euryarchaeota y Elusimicrobia emergieron como taxones clave que contribuyeron a las diferencias observadas en la microbiota ruminal entre estos grupos (Figura 6). En los grupos AGF y MN se identificaron un total de 112 biomarcadores en varios niveles taxonómicos. En particular, Prevotellaceae y Clostridia emergieron como taxones clave que contribuyeron a las diferencias observadas en la microbiota ruminal entre estos grupos (Tabla suplementaria S3). En los grupos AGF y CON, se identificaron un total de 24 biomarcadores en varios niveles taxonómicos. En particular, Clostridia y Pseudomonadales emergieron como taxones clave que contribuyeron a las diferencias observadas en la microbiota ruminal entre estos grupos (Tabla suplementaria S4). En los grupos TW y MN se identificaron un total de 12 biomarcadores en varios niveles taxonómicos. En particular, Erysipelotrichia y Bacterium_VCD2007 emergieron como taxones clave que contribuyeron a las diferencias observadas en la microbiota ruminal entre estos grupos (Tabla suplementaria S5). En los grupos TW y CON, se identificaron un total de 92 biomarcadores en varios niveles taxonómicos. En particular, Rikenellaceae y Prevotellaceae emergieron como taxones clave que contribuyeron a las diferencias observadas en la microbiota ruminal entre estos grupos (Tabla suplementaria S6).
3.3.6 Predicción del perfil funcional de la microbiota
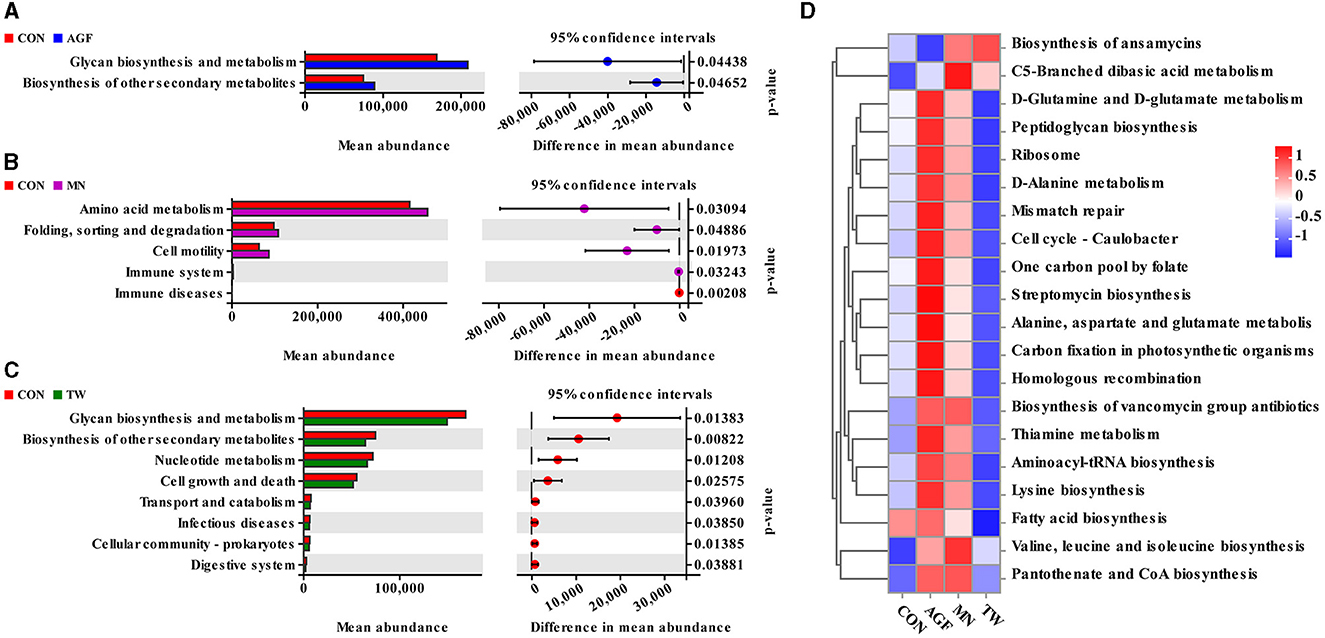
La predicción funcional de la microbiota ruminal se realizó en la plataforma PICRUSt2. Se aplicó la prueba t de Welch para identificar diferencias significativas en las vías funcionales entre los cuatro grupos en el nivel 2. Dos categorías funcionales mostraron diferencias significativas entre los grupos AGF y CON (p < 0,05). En el grupo AGF, tanto la biosíntesis como el metabolismo de los glicanos, y la biosíntesis de otros metabolitos secundarios, se elevaron significativamente en comparación con el grupo CON (p < 0,05) (Figura 7A). Se observaron disparidades significativas en cinco categorías funcionales entre los grupos MN y CON (p < 0,05). En el grupo MN, los niveles de metabolismo, plegamiento, clasificación y degradación de aminoácidos y motilidad celular fueron significativamente elevados en comparación con el grupo CON (p < 0,05) (Figura 7B). Ocho clasificaciones funcionales en los grupos TW y CON mostraron diferencias estadísticamente significativas, siendo el grupo TW significativamente menor que el grupo CON (p < 0,05) (Figura 7C). En el nivel 3, el estudio predijo un total de veinte categorías funcionales dentro de la microbiota ruminal de las cabras. Los grupos AGF y MN mostraron un enriquecimiento similar en dieciocho categorías funcionales (Figura 7D).
4 Discusión
La «prohibición» total puede tener un cierto impacto en la capacidad de producción de la ganadería a corto plazo. En la actualidad, se ha encontrado que los extractos de plantas (43), los probióticos (44), las preparaciones enzimáticas (45), el quitosano (46) y los péptidos antimicrobianos (47) tienen las características de alta eficiencia y sin efectos secundarios. Este grupo de investigación ha llevado a cabo experimentos sobre aditivos vegetales naturales y aditivos alimentarios, con el fin de superar los defectos de los antibióticos en diferentes grados. Entre estos, se espera que los flavonoides Ampelopsis grossedentata, un extracto de planta (48), y Tween 80, un tensioactivo no iónico (49), los cuales no son tóxicos e inofensivos, se conviertan en nuevos aditivos verdes para piensos libres de contaminación. Sin embargo, la información sobre el uso de AGF en rumiantes sigue siendo escasa. La información relativa a la aplicación de AGF en animales rumiantes sigue siendo limitada. Por lo tanto, el objetivo de este estudio es determinar la eficacia de AGF y TW mediante el análisis de su influencia en el rendimiento del crecimiento de las cabras, los metabolitos plasmáticos y la microbiota ruminal, todos ellos indicadores clave del crecimiento y el estado de salud de los animales.
La mejora del rendimiento del crecimiento de las cabras está estrechamente relacionada con el ADFI y la utilización del alimento. Los resultados mostraron que tanto el ADFI como el ADG fueron más altos en el grupo AGF que en otros grupos, con la menor relación F/G, lo que indica una alta utilización del alimento y un efecto de engorde efectivo. Esta eficacia se atribuye a la capacidad de los flavonoides de Ampelopsis grossedentata para aliviar las respuestas al estrés oxidativo, reducir el daño gastrointestinal y promover el crecimiento y desarrollo gastrointestinal de las cabras (4). Contrariamente a lo esperado, el grupo TW demostró un efecto adverso, diferente de estudios previos. Por ejemplo, McAllister et al. (16) observaron que el tratamiento con Tween 80 en cabras redujo significativamente el IMD y la ganancia diaria en comparación con los controles no tratados. Lee et al. (17) reportaron un aumento en el IMD y en la producción de leche en vacas tratadas con Tween 80. Las diferentes respuestas pueden atribuirse a la atractiva fragancia de los flavonoides Ampelopsis grossedentata que mejoran la ingesta de alimento, mientras que el olor ligeramente desagradable y el sabor amargo de Tween 80 podrían disuadir a las cabras de comer. Teniendo en cuenta estos factores, los flavonoides de Ampelopsis grossedentata se muestran prometedores como sustituto de la monensina para mejorar el rendimiento del crecimiento, pero se requieren pruebas más rigurosas.
Los cambios en la composición plasmática pueden reflejar el nivel nutricional y la función orgánica de los rumiantes (50). La ALT y la AST son dos tipos de aminotransferasas, desempeñan un papel crucial en la evaluación de la función hepática, los aminoácidos y las proteínas, el metabolismo e influyen indirectamente en la descomposición y síntesis de grasas y azúcares (51). Normalmente, los niveles de AST en cabras oscilan entre 40.0 y 123.0 U/L, pero pueden aumentar cuando el hígado está dañado (52). En este experimento, la actividad plasmática de AST en todos los grupos de cabras estuvo dentro del rango normal. La fosfatasa alcalina (ALP), una enzima que se encuentra ampliamente en varios tejidos como el hígado, los huesos, los intestinos, los riñones y la placenta, se secreta desde el hígado a la bilis. Se asocia con el crecimiento óseo en animales, así como con el transporte y utilización de azúcares, lípidos, proteínas y ácido fosfórico. Los niveles de FA reflejan el crecimiento y desarrollo del ganado y las aves de corral (53). La actividad de ALP en el plasma de cabra del grupo AGF fue significativamente mayor que en el grupo MN, lo que justifica una mayor investigación. El HDL-CH, considerado colesterol bueno, transporta los lípidos de los tejidos al hígado. En este experimento, los niveles de HDL-CH en el grupo AGF fueron significativamente más altos que en el grupo MN. El AGF es útil para la absorción y el transporte de lípidos en rumiantes. En resumen, se ha demostrado que el AGF mejora el crecimiento y el desarrollo y los niveles de lípidos y los perfiles lipídicos en cabras.
El pH del líquido ruminal refleja el estado de fermentación ruminal principalmente al afectar la actividad microbiana del rumen. Los estudios han demostrado que la composición de la dieta, la secreción de saliva, la utilización del producto de fermentación ruminal, la eficiencia de absorción y antes y después de la alimentación pueden causar cambios en el pH del líquido ruminal, y la composición dietética es el factor clave que causa cambios en el pH (54). Además, cuando el pH del rumen es de 6,4 a 6,8, las bacterias que descomponen la celulosa pueden lograr un ambiente adecuado (55). Cuando el pH del rumen era de 6,2 a 7,0, el ecosistema microbiano del rumen era relativamente estable, lo que podía asegurar la fermentación normal del rumen (56). En este experimento, la suplementación de la dieta con varios aditivos alimentarios no influyó notablemente en el pH del líquido ruminal, que se mantuvo dentro del rango de 6,4 a 6,8, lo que indica que el entorno ruminal era relativamente estable y el número medido de microorganismos ruminales reflejaba con precisión la microflora.
Los microorganismos ruminales y sus huéspedes constituyen un microecosistema complejo, que desempeña un papel importante en la nutrición de los rumiantes. El estudio en profundidad del microecosistema ruminal es propicio para comprender mejor el mecanismo de la nutrición de los rumiantes (57). La diversidad alfa se refiere a la diversidad dentro de un hábitat o ecosistema en particular. La cobertura de Good refleja la saturación de secuenciación de la muestra. Los índices Sobs, Chao1 y ACE se refieren principalmente a la riqueza de especies de las muestras. Simpson y Shannon reflejan de manera integral la riqueza y la uniformidad de las especies, por lo que el nivel del índice también se verá afectado por la uniformidad. Cuanto más uniformemente se distribuyan las especies en la muestra, mayor será la diversidad. En este experimento, la saturación secuencial de cada grupo fue suficiente. Los índices Sobs, ACE, Chao1, Shannon y Simpson del grupo TW fueron significativamente más altos que los de los otros tres grupos. En este experimento, el grupo TW tuvo la mayor cantidad de OTU y el grupo AGF tuvo menos OTU, lo que también se correspondió con la diversidad de resultados. En pocas palabras, los hallazgos sugieren que el grupo TW exhibió la mayor riqueza de especies y uniformidad en el fluido ruminal de cabra, por lo que también posee el mayor nivel de diversidad de especies.
Firmicutes y Bacteroidetes emergieron como los dos filos bacterianos predominantes en este estudio, corroborando los hallazgos de investigaciones anteriores (58). Entre los 10 principales filos, la abundancia relativa de Bacteroides y Proteobacteria en el grupo TW es significativamente menor que la del grupo AGF, mientras que la abundancia relativa de Firmicutes y Kiritimatiellaeota muestra la tendencia opuesta. Los bacteroides desempeñan un papel clave en el metabolismo de polisacáridos y oligosacáridos, proporcionando así nutrientes y vitaminas tanto al huésped como a otros microbios que residen en los intestinos (59). Los intestinos firmes, estimulados por la fibra dietética, y sus metabolitos muestran funciones significativas que promueven la salud (60). Prevotella_1 es el género dominante, lo cual es consistente con los resultados de estudios previos, y su abundancia relativa en el grupo AGF es significativamente mayor que en otros grupos. Además, Prevotella es capaz de utilizar almidón, monosacáridos y varios polisacáridos no celulósicos (60). Estos resultados indican que la síntesis y el metabolismo de los azúcares en el grupo AGF son más fuertes, lo que es consistente con la predicción funcional en este experimento.
El Rikenellaceae_RC9_gut_group, que forma parte de la familia Rikenellaceae, fermenta principalmente polisacáridos no absorbidos en el intestino del huésped, lo que conduce a la producción de ácidos grasos de cadena corta como acetato, propionato y butirato (61). En este experimento, la abundancia relativa de Rikenellaceae_RC9_gut_group en el grupo TW fue significativamente mayor que en otros grupos. Sin embargo, la función del metabolismo de la glucosa del grupo TW fue significativamente menor que la de los grupos CON y AGF, lo que se especula que se debe a la menor abundancia de Rikenellaceae_RC9_gut_group en comparación con Prevotella_1 entre el total de géneros bacterianos. Lactobacillus se considera una bacteria beneficiosa, crucial para preservar el equilibrio de la microbiota intestinal y frustrar la invasión de patógenos (62). La abundancia relativa de Lactobacillus en los grupos AGF y CON fue significativamente mayor que en los grupos TW y MN. En conclusión, el equilibrio microbiano del rumen y la capacidad anabólica de la glucosa del grupo AGF fueron relativamente fuertes, y la diversidad del grupo AGF fue más cercana a la del grupo CON, lo que lo convierte en una buena opción para los aditivos verdes.
5 Conclusiones
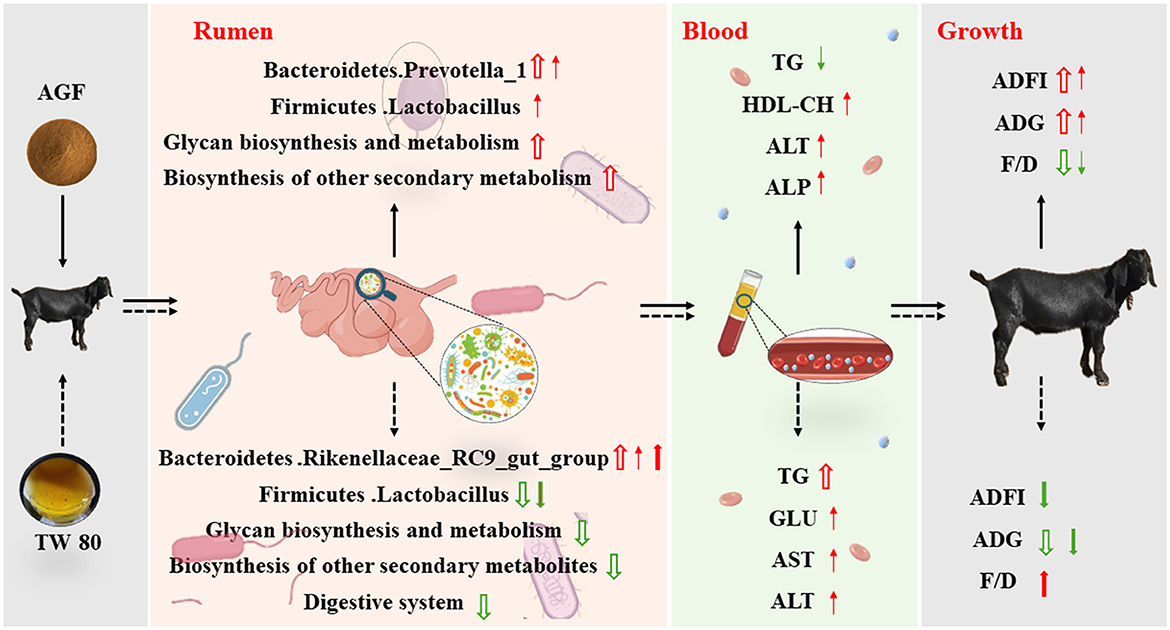
El diagrama de ajuste de este estudio se muestra en la Figura 8, que se dibuja de acuerdo con los resultados experimentales. En conclusión, la adición dietética de flavonoides de Ampelopsis grossedentata puede aumentar significativamente la abundancia relativa de bacterias beneficiosas y afectar los índices fisiológicos y bioquímicos del plasma de las cabras. Mejorar aún más la ingesta media diaria de alimento y la ganancia diaria de las cabras y reducir la relación entre el alimento y la ganancia. La suplementación dietética de Tween 80 aumenta significativamente la diversidad y abundancia de microorganismos ruminales, optimizando así la comunidad microbiana y mejorando la salud ruminal. Sin embargo, a pesar de estos beneficios, la abundancia de bacterias beneficiosas es menor que en el grupo de control, lo que sugiere que las mejoras en la diversidad microbiana del rumen por sí solas pueden no compensar completamente la menor palatabilidad. En consecuencia, el efecto de la alimentación es más pobre en comparación con el grupo AGF, lo que indica que el método de alimentación necesita una mayor mejora. Con todo, los flavonoides de Ampelopsis grossedentata muestran potencial como un nuevo aditivo verde para reemplazar la monensina y reducir el estrés en la cría de animales.
Declaración de disponibilidad de datos
Los datos presentados en el estudio están depositados en el repositorio del NCBI, número de acceso PRJna956984.
Declaración ética
Los estudios en animales fueron aprobados por el Comité de Cuidado y Uso de Animales de la Universidad Agrícola de Yunnan. Los estudios se llevaron a cabo de acuerdo con la legislación local y los requisitos institucionales. Se obtuvo el consentimiento informado por escrito de los propietarios para la participación de sus animales en este estudio.
Contribuciones de los autores
JZ: Conceptualización, Curación de datos, Análisis formal, Investigación, Metodología, Validación, Visualización, Redacción – borrador original, Redacción – revisión y edición. YL: Conceptualización, Investigación, Validación, Redacción – borrador original, Escritura – revisión y edición. ZG: Investigación, Software, Escritura – Revisión y Edición. YC: Investigación, Software, Escritura – Revisión y Edición. ML: Análisis formal, Investigación, Redacción – revisión y edición. WD: Investigación, Recursos, Escritura – revisión y edición. DX: Curación de datos, Obtención de fondos, Metodología, Supervisión, Redacción, revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue financiada por el Proyecto Nacional de Investigación y Desarrollo (2022YFD1100408).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2024.1411071/full#supplementary-material
Referencias
1. Wen R, Li C, Zhao M, Wang H, Tang Y. Retirada de antibióticos promotores del crecimiento en China y su impacto en el patógeno transmitido por los alimentos Campylobacter coli de origen porcino. Microbiol frontal. (2022) 13:1004725. doi: 10.3389/fmicb.2022.1004725
Resumen de PubMed | Texto completo de Crossref | Google Académico
2. Toutain PL, Ferran AA, Bousquet-Melou A, Pelligand L, Lees P. La medicina veterinaria se refiere a los nuevos medicamentos antimicrobianos verdes. Microbiol frontal. (2016) 7:1196. doi: 10.3389/fmicb.2016.01196
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Zhang S, Gao S, Chen Y, Xu S, Yu S, Zhou J. Identificación de enzimas de hidroxilación y análisis metabólico de la síntesis de dihidromiricetina en Ampelopsis Grossedentata. Genes. (2022) 13:2318. doi: 10.3390/genes13122318
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Hou X, Tong Q, Wang W, Xiong W, Shi C, Fang J. La dihidromiricetina protege las células endoteliales del daño por estrés oxidativo inducido por el peróxido de hidrógeno mediante la regulación de las vías mitocondriales. Ciencia de la vida. (2015) 130:38–46. doi: 10.1016/j.lfs.2015.03.007
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Wu Y, Bai J, Zhong K, Huang Y, Gao H. Un mecanismo antibacteriano dual implicado en la disrupción de la membrana y la unión al ADN de la 2r,3r-dihidromiricetina de las agujas de pino de cedrus deodara contra Staphylococcus aureus. Químico de los alimentos. (2017) 218:463–70. doi: 10.1016/j.foodchem.2016.07.090
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Zhang Y, Yang H, Li S, Li WD, Wang Y. Consumo de café y té y riesgo de desarrollar accidente cerebrovascular, demencia y demencia posterior al accidente cerebrovascular: un estudio de cohorte en el biobanco del Reino Unido. PLoS Med. (2021) 18:E1003830. doi: 10.1371/journal.pmed.1003830
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Zhang X, Wang L, Peng L, Tian X, Qiu X, Cao H, et al. La dihidromiricetina protege a los huvecs del daño oxidativo inducido por el nitroprusiato de sodio mediante la activación de la vía de señalización Pi3k/Akt/Foxo3a. J Cell Mol Med. (2019) 23:4829–38. doi: 10.1111/jcmm.14406
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Wu RR, Li X, Cao YH, Peng X, Liu GF, Liu ZK, et al. Plantas medicinales chinas de Ampelopsis Grossedentata: una revisión de sus características botánicas, uso, fitoquímica, componentes farmacológicos activos y toxicología. Moléculas. (2023) 28:7145. doi: 10.3390/moléculas28207145
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Xiao XN, Wang F, Yuan YT, Liu J, Liu YZ Yi X. Actividad antibacteriana y modo de acción de la dihidromiricetina de las hojas de Ampelopsis Grossedentata contra las bacterias transmitidas por los alimentos. Moléculas. (2019) 24:2831. doi: 10.3390/moléculas24152831
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Liang H, He K, Li T, Cui S, Tang M, Kang S, et al. Mecanismo y actividad antibacteriana del extracto de té de vid y dihidromiricetina frente a Staphylococcus aureus. Sci Rep. (2020) 10:21416. doi: 10.1038/s41598-020-78379-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Luo QJ, Zhou WC, Liu XY, Li YJ, Xie QL, Wang B, et al. Constituyentes químicos y actividades inhibidoras, antioxidantes y hepatoprotectoras de la a-glucosidasa de Ampelopsis Grossedentata. Moléculas. (2023) 28:7956. doi: 10.3390/moléculas28247956
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Wang Z, Jiang Q, Li P, Shi P, Liu C, Wang W, et al. El extracto acuoso de Ampelopsis Grossedentata alivia el estrés oxidativo y la inflamación intestinal. Antioxidantes. (2023) 12:0547. doi: 10.3390/antiox12030547
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Zhu J, Liu X, Lu Y, Yue D, He X, Deng W, et al. Explorar el impacto de los flavonoides de Ampelopsis Grossedentata en el rendimiento del crecimiento, la microbiota ruminal y la fisiología y bioquímica del plasma de los niños. Animales. (2023) 13:2454. doi: 10.3390/ani13152454
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. Akin DE. Evaluación por microscopía electrónica y cultivo anaeróbico de tipos de bacterias ruminales asociadas a la digestión de paredes celulares forrajeras. Appl Environ Microbiol. (1980) 39:242–52. doi: 10.1128/aem.39.1.242-252.1980
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Lu Y, Wang YY, Yang N, Zhang D, Zhang FY, Gao HT, et al. El emulsionante alimentario polisorbato 80 aumenta la absorción intestinal de ftalato de Di-(2-etilhexilo) en ratas. Toxicol Sci. (2014) 139:317–27. doi: 10.1093/toxsci/kfu055
Resumen de PubMed | Texto completo de Crossref | Google Académico
16. McAllister TA, Stanford K, Bae HD, Treacher RJ, Hristov AN, Baah J, et al. Efecto de un tensioactivo y enzimas exógenas sobre la digestibilidad del alimento y sobre el rendimiento del crecimiento. Canadiense J Animal Sci. (2000) 80:35–44. doi: 10.4141/A99-053
17. Lee SS, Ahn BH, Kim HS, Kim CH, Cheng KJ, Ha JK. Efectos de los tensioactivos no iónicos en las distribuciones enzimáticas del contenido ruminal, el crecimiento anaeróbico de los microbios ruminales, las características de fermentación ruminal y el rendimiento de las vacas lactantes. Asiático Austral J Animal Sci. (2003) 16:104–15. doi: 10.5713/ajas.2003.104
18. Okino S, Ikeo M, Ueno Y, Taneda D. Efectos de Tween 80 en la estabilidad de la celulasa en condiciones agitadas. Bioresour Technol. (2013) 142:535–9. doi: 10.1016/j.biortech.2013.05.078
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Menberu MA, Hayes AJ, Liu S, Psaltis AJ, Wormald PJ, Vreugde S. Tween 80 y su derivado ácido oleico promueven el crecimiento de Corynebacterium accolens e inhiben los aislados clínicos de Staphylococcus aureus. Int Forum Alergia Rhinol. (2021) 11:810–3. doi: 10.1002/alr.22730
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Olukanni OD, Albert AA, Farinto M, Awotula AO, Osuntoki AA. Tween-80 mejoró la biodegradación de naftaleno por klebsiella quasipneumoniae. Antonie Van Leeuwenhoek. (2023) 116:697–709. doi: 10.1007/s10482-023-01839-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Consejo Nacional de Investigación (NRC) (2007). Requerimientos de nutrientes de los pequeños rumiantes: ovejas, cabras, cérvidos y camélidos del Nuevo Mundo. Washington, DC: The National Academies Press.
22. Lodge-Ivey SL, Browne-Silva J, Horvath MB. Nota técnica: diversidad bacteriana y productos finales de fermentación en muestras de fluido ruminal recogidas mediante lavado oral o cánula ruminal. J Anim Sci. (2009) 87:2333–7. doi: 10.2527/jas.2008-1472
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Thermo Fisher Scientific (2009). Espectrofotómetros NanoDrop 2000/2000c. [Manual de usuario]. Disponible en línea en: https://www.thermofisher.com/nanodrop
24. Guo M, Wu F, Hao G, Qi Q, Li R, Li N, et al. Bacillus subtilis mejora la inmunidad y la resistencia a enfermedades en conejos. Frente Immunol. (2017) 8:354. doi: 10.3389/fimmu.2017.00354
Resumen de PubMed | Texto completo de Crossref | Google Académico
25. Chen S, Zhou Y, Chen Y, Gu J. Fastp: un preprocesador fastq todo en uno ultrarrápido. Bioinformática. (2018) 34:i884-i90. doi: 10.1093/bioinformatics/bty560
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. Magoč T, Salzberg SL. Flash: ajuste rápido de la longitud de lecturas cortas para mejorar los ensamblajes del genoma. Bioinformática. (2011) 27:2957–63. doi: 10.1093/bioinformatics/btr507
Resumen de PubMed | Texto completo de Crossref | Google Académico
27. Bokulich NA, Subramanian S, Faith JJ, Gevers D, Gordon JI, Knight R, et al. El filtrado de calidad mejora enormemente las estimaciones de diversidad de la secuenciación de amplicones de illumina. Métodos Nat. (2013) 10:57–9. doi: 10.1038/nmeth.2276
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. Qiime permite el análisis de datos de secuenciación comunitaria de alto rendimiento. Métodos Nat. (2010) 7:335–6. doi: 10.1038/nmeth.f.303
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R. Uchime mejora la sensibilidad y la velocidad de detección de quimeras. Bioinformática. (2011) 27:2194–200. doi: 10.1093/bioinformatics/btr381
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Edgar RC. Músculo: alineación de secuencias múltiples con alta precisión y alto rendimiento. Ácidos nucleicos Res. (2004) 32:1792–7. doi: 10.1093/nar/gkh340
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Price MN, Dehal PS, Arkin AP. Fasttree 2: árboles de máxima probabilidad aproximada para alineaciones grandes. PLoS UNO. (2010) 5:e9490. doi: 10.1371/journal.pone.0009490
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Lozupone C, Knight R. Unifrac: un nuevo método filogenético para comparar comunidades microbianas. Appl Environ Microbiol. (2005) 71:8228–35. doi: 10.1128/AEM.71.12.8228-8235.2005
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Oksanen JB, Kindt FG. R Vegano: paquete de ecología comunitaria. Paquete R. (2010). Disponible en línea en: https://cran.r-project.org (consultado el 10 de febrero de 2024).
34. Kolde RK. M R. Paquete ‘Pheatmap’. Paquete R (2015). Disponible en línea en: https://CRAN.R-project.org/package=pheatmap (consultado el 14 de febrero de 2024).
35. Revelle WR MW. Paquete ‘Psych’. La amplia red de archivos de R. (2015). Disponible en línea en: https://CRAN.R-project.org/package=psychTools (consultado el 13 de febrero de 2024).
36. Csardi MG. Paquete«Igraph». (2013) 3:2013. Disponible en línea en: https://CRAN.R-project.org/package=igraph (consultado el 10 de febrero de 2024).
37. Wickham HC. W Ggplot2: una implementación de la gramática de los gráficos. Versión 07 del paquete R. (2008). Disponible en línea en: http://CRANR-projectorg/package=~ggplot2 (consultado el 10 de febrero de 2024).
38. Langille MG, Zaneveld J, Caporaso JG, McDonald D, Knights D, Reyes JA, et al. Perfil funcional predictivo de comunidades microbianas utilizando secuencias de genes marcadores de ARNr 16s. Nat Biotechnol. (2013) 31:814–21. doi: 10.1038/nbt.2676
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Ward TL, J. Meulemans, J. Bugbase predice los fenotipos del microbioma a nivel de organismo. BioRxiv. (2017). Pág. 133462. doi: 10.1101/133462
40. Conway JR, Lex A, Gehlenborg N. Upsetr: un paquete de R para la visualización de conjuntos que se intersecan y sus propiedades. Bioinformática. (2017) 33:2938–40. doi: 10.1093/bioinformatics/btx364
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Descubrimiento y explicación de biomarcadores metagenómicos. Genoma Biol. (2011) 12:R60. doi: 10.1186/gb-2011-12-6-r60
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Pollesel M, Tassinari M, Frabetti A, Fornasini D, Cavallini D. Efecto del orden de paridad de las hembras sobre los parámetros de homogeneidad de la camada. Ital J Anim Sci. (2020) 19:1188–94. doi: 10.1080/1828051X.2020.1827990
43. Jayanegara A, Yogianto Y, Wina E, Sudarman A, Kondo M, Obitsu T, et al. Efectos combinados de extractos de plantas ricos en taninos y saponinas como aditivos alimentarios para mitigar la formación ruminal in vitro de metano y amoníaco. Animales. (2020) 10:1531. doi: 10.3390/ani10091531
Resumen de PubMed | Texto completo de Crossref | Google Académico
44. Markowiak P, Sliżewska K. Efectos de los probióticos, prebióticos y simbióticos en la salud humana. Nutrientes. (2017) 9:1021. doi: 10.3390/nu9091021
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Peters A, Lebzien P, Meyer U, Borchert U, Bulang M, Flachowsky G. Efecto de las enzimas fibrolíticas exógenas en la fermentación ruminal y la digestión de nutrientes en vacas lecheras. Arch Anim Nutr. (2010) 64:221–37. doi: 10.1080/17450391003693142
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Swiatkiewicz S, Swiatkiewicz M, Arczewska-Wlosek A, Jozefiak D. Chitosan y sus derivados de oligosacáridos (chito-oligosacáridos) como suplementos alimenticios en la nutrición de aves de corral y cerdos. J Anim Physiol Anim Nutr. (2015) 99:1–12. doi: 10.1111/jpn.12222
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Liu Q, Yao S, Chen Y, Gao S, Yang Y, Deng J, et al. Uso de péptidos antimicrobianos como aditivo alimentario para cabras juveniles. Sci Rep. (2017) 7:12254. doi: 10.1038/s41598-017-12394-4
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Chen J, Wu Y, Zou J, Gao K. Inhibición de la A-glucosidasa y actividad antihiperglucémica de los flavonoides de ampelopsis grossedentata y los derivados flavonoides. Bioorg Med Chem. (2016) 24:1488–94. doi: 10.1016/j.bmc.2016.02.018
Resumen de PubMed | Texto completo de Crossref | Google Académico
49. Kaur G, Mehta SK. Desarrollos de microemulsiones basadas en polisorbato (Tween): administración preclínica de fármacos, toxicidad y aplicaciones antimicrobianas. Int J Pharm. (2017) 529:134–60. doi: 10.1016/j.ijpharm.2017.06.059
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Nozad S, Ramin AG, Moghadam G, Asri-Rezaei S, Babapour A, Ramin S. Relación entre las concentraciones de urea, proteína, creatinina, triglicéridos y macrominerales en sangre con la calidad y cantidad de leche en vacas Holstein lecheras. Foro Veter Res. (2012) 3:55–9. doi: 10.2298/AVB1101003N
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Mohammadi V, Anassori E, Jafari S. Medición de los cambios en los metabolitos bioquímicos relacionados con la energía durante el período periparto en ovejas de raza makouei. Foro Veter Res. (2016) 7:35–9.
52. Russell KE, Roussel AJ. Evaluación del perfil químico sérico de rumiantes. Veter Clinics North Am Food Animal Pract. (2007) 23:403–26. doi: 10.1016/j.cvfa.2007.07.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Poupon R. Fosfatasa alcalina hepática: un eslabón perdido entre la coléresis y la inflamación biliar. Hepatología. (2015) 61:2080–90. doi: 10.1002/hep.27715
Resumen de PubMed | Texto completo de Crossref | Google Académico
54. Jiang FG, Lin XY, Yan ZG, Hu ZY, Wang Y, Wang ZH. Efectos de la fuente de forraje y el tamaño de partícula sobre la actividad masticatoria, el pH ruminal y la secreción de saliva en vacas Holstein lactantes. Anim Sci J. (2019) 90:382–92. doi: 10.1111/asj.13153
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Liang J, Zhang P, Zhang R, Chang J, Chen L, Wang G, et al. Respuesta de los microorganismos ruminales al Ph durante la hidrólisis anaerobia y la acidogénesis de la biomasa lignocelulósica. Gestión de residuos. (2024) 174:476–86. doi: 10.1016/j.wasman.2023.12.035
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Mackie RI, Gilchrist FM. Cambios en las bacterias productoras y utilizadas de lactato en relación con el pH en el rumen de las ovejas durante la adaptación gradual a una dieta alta en concentrados. Appl Environ Microbiol. (1979) 38:422–30. doi: 10.1128/aem.38.3.422-430.1979
Resumen de PubMed | Texto completo de Crossref | Google Académico
57. Pan X, Li Z, Li B, Zhao C, Wang Y, Chen Y, et al. Dinámica de la expresión génica del rumen, colonización del microbioma y su interacción en cabras. BMC Genómica. (2021) 22:288. doi: 10.1186/s12864-021-07595-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Cui X, Wang Z, Yan T, Chang S, Wang H, Hou F. Diversidad bacteriana ruminal de ovejas tibetanas (ovis aries) asociada con diferentes tipos de forraje en la meseta qinghai-tibetana. Can J Microbiol. (2019) 65:859–69. doi: 10.1139/cjm-2019-0154
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Zafar H, Saier MH. Especies de bacteroides intestinales en la salud y la enfermedad. Microbios intestinales. (2021) 13:1–20. doi: 10.1080/19490976.2020.1848158
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Sun Y, Zhang S, Nie Q, He H, Tan H, Geng F, et al. Intestino firme: relación con la fibra dietética y papel en la homeostasis del huésped. Crit Rev Food Sci Nutr. (2023) 63:12073–88. doi: 10.1080/10408398.2022.2098249
Resumen de PubMed | Texto completo de Crossref | Google Académico
61. Su XL, Tian Q, Zhang J, Yuan XZ, Shi XS, Guo RB, et al. Acetobacteroides hydrogenigenes gen. Nov, SP nov, una bacteria anaerobia productora de hidrógeno de la familia Rikenellaceae aislada de un carrizal. Int J System Evolut Microbiol. (2014) 64:2986–91. doi: 10.1099/ijs.0.063917-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
62. Wang Y, Li A, Jiang X, Zhang H, Mehmood K, Zhang L, et al. Potencial probiótico de cepas de leuconostoc pseudomesenteroides y lactobacillus aisladas de yaks. Microbiol frontal. (2018) 9:2987. doi: 10.3389/fmicb.2018.02987
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: Flavonoides Ampelopsis grossedentata, Tween 80, cabras nubias, rendimiento del crecimiento, índices sanguíneos, microbiota ruminal
Cita: Zhu J, Lu Y, Gao Z, Chong Y, Li M, Deng W y Xi D (2024) Explorando el efecto aditivo de los flavonoides Ampelopsis grossedentata y Tween 80 en la alimentación de cabras nubias. Frente. Vet. Sci. 11:1411071. doi: 10.3389/fvets.2024.1411071
Recibido: 02 de abril de 2024; Aceptado: 25 de junio de 2024;
Publicado: 12 julio 2024.
Editado por:
Haoyu Liu, Universidad de Yangzhou, China
Revisado por:
Rongfang Li, Universidad Agrícola de Hunan, China
Houqiang Luo, Facultad Vocacional de Ciencia y Tecnología de Wenzhou, China
Derechos de autor © 2024 Zhu, Lu, Gao, Chong, Li, Deng y Xi. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY). S
*Correspondencia: Dongmei Xi, dmxiynau@163.com
†Estos autores han contribuido igualmente a este trabajo
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente a las de sus organizaciones afiliadas, o las del editor, de los editores y de los revisores. Cualquier producto que puede ser evaluada en este artículo o afirmación que puede ser hecha por su El fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía