
Factores de riesgo ambientales para desarrollar trastornos crónicos

Factores de riesgo ambientales en cachorros y gatitos para desarrollar trastornos crónicos en la edad adulta: un llamado a la investigación sobre la programación del desarrollo
 Virginie Gaillard1*
Virginie Gaillard1*  Sylvie Chastant2
Sylvie Chastant2  Gary Inglaterra3
Gary Inglaterra3  Oliver Forman4
Oliver Forman4  Alexander J. Alemán5
Alexander J. Alemán5  Jan S. Suchodolski6
Jan S. Suchodolski6  Cecilia Villaverde7
Cecilia Villaverde7  Pascale Chavatte-Palmer8,9
Pascale Chavatte-Palmer8,9  Franck Péron1
Franck Péron1- 1Centro de Investigación y Desarrollo, Royal Canin, Aimargues, Francia
- 2NeoCare, Université de Toulouse, Ecole Nationale Vétérinaire de Toulouse (ENVT), Toulouse, Francia
- 3Facultad de Medicina Veterinaria, Universidad de Nottingham, Nottingham, Reino Unido
- 4Wisdom Panel, Kinship, Waltham-on-the-Wolds, Leicestershire, Reino Unido
- 5Institute of Life Course and Medical Sciences of Small Animal Medicine, University of Liverpool, Liverpool, Reino Unido
- 6Laboratorio Gastrointestinal, Departamento de Ciencias Clínicas de Pequeños Animales, Texas A&M University, College Station, TX, Estados Unidos
- 7Expert Pet Nutrition, Fermoy, Condado de Cork, Irlanda
- 8Université Paris-Saclay, Université de Versailles Saint-Quentin-en-Yvelines (UVSQ), Institut National de Recherche Pour l’Agriculture, l’Alimentation et l’Environnement (INRAE), Biologie de la Reproduction, Environnement, Epigénétique et Développement (BREED), Jouy-en-Josas, Francia
- 9Ecole Nationale Vétérinaire d’Alfort, BREED, Maisons-Alfort, Francia
Muchos perros y gatos se ven afectados por enfermedades crónicas que afectan significativamente su salud y bienestar y las relaciones con los humanos. Algunas de estas enfermedades pueden ser difíciles de tratar, y una mejor comprensión de los factores de riesgo de la vida temprana para las enfermedades que ocurren en la edad adulta es clave para mejorar la atención veterinaria preventiva y las prácticas de cría. Este artículo revisa los factores de riesgo de la vida temprana para la obesidad y la enteropatía crónica, y para los problemas crónicos de comportamiento, que también pueden ser intratables con consecuencias que cambian la vida. Los aspectos de la vida temprana en cachorros y gatitos que pueden afectar el riesgo de trastornos adultos incluyen la nutrición materna, el establecimiento del microbioma intestinal, el comportamiento materno, el destete, la nutrición durante el crecimiento, la tasa de crecimiento, la socialización con congéneres y humanos, el realojamiento y la castración. A pesar de la evidencia en algunas especies de que los trastornos revisados aquí reflejan los orígenes del desarrollo de la salud y la enfermedad (DOHaD), la programación del desarrollo rara vez se ha estudiado en perros y gatos. Se discuten las prioridades y estrategias para aumentar el conocimiento de los factores de riesgo de la vida temprana y DOHaD en perros y gatos. Se proponen ventanas críticas de desarrollo: preconcepción, gestación, período de lactancia, precastración o prepubertad del crecimiento temprano, y crecimiento postcastración o postpubertad hasta el tamaño adulto, cuya duración depende de la especie y la raza. Los desafíos para la investigación de DOHaD en estas especies incluyen un gran número de razas con amplia variabilidad genética y fenotípica, y la existencia de muchos individuos de razas mixtas. Además, las dificultades para realizar estudios prospectivos de cohortes de por vida se ven exacerbadas por la discontinuidad en la cría de mascotas entre los criadores y los propietarios posteriores, y por la naturaleza dispersa de la propiedad de mascotas
Introducción
Existe una creciente conciencia de que los aspectos de la vida temprana en cachorros y gatitos, especialmente la nutrición durante la gestación y el crecimiento temprano, afectan el riesgo de mortalidad neonatal (1-3) y el desarrollo de enfermedades crónicas en la edad adulta (4, 5). En muchas especies de mamíferos, las experiencias tempranas de la vida y de los padres se han investigado como posibles contribuyentes a los orígenes del desarrollo de la salud y la enfermedad (DOHaD). El concepto de DOHaD abarca las observaciones de que las exposiciones ambientales durante el desarrollo pueden impulsar cambios epigenéticos que modifican o «programan» la expresión de genes, afectando el desarrollo estructural y funcional, con riesgos rápidos o tardíos para la salud. En los seres humanos, los primeros 1.000 días de vida, que se aproximan a la gestación más 2 años postnatales, se han identificado como un período crítico en el que la programación del desarrollo sienta las bases para un neurodesarrollo, crecimiento y salud óptimos (6). Gran parte de lo que se sabe sobre DOHaD y el epigenoma se deriva de modelos animales de laboratorio (7), pero el conocimiento experimental también se ha acumulado para rumiantes, cerdos e incluso caballos (7-9).
En perros y gatos, la mayoría de las investigaciones sobre la etiología de las afecciones crónicas de la edad adulta se han centrado en los predictores ambientales adultos y los factores de riesgo, sin investigar si estos tienen orígenes en el desarrollo. Las búsquedas bibliográficas en PubMED (31 de julio de 2022) con la amplia cadena de búsqueda (epigenética O «programación del desarrollo» O DOHaD O «orígenes del desarrollo de la salud y la enfermedad»), combinada con «Perros» o «Gatos» como encabezados de temas médicos, recuperaron 218 artículos. Después de seleccionar los títulos y resúmenes para determinar su relevancia, permanecieron 51 artículos relacionados con perros y 6 relacionados con gatos; Los temas de interés más evidentes fueron las modificaciones epigenéticas en células cancerosas [32 artículos (56%)] y los aspectos epigenéticos del fenotipo de la raza. En general, ha habido una consideración inadecuada en perros y gatos de la medida en que el medio ambiente, durante las diferentes etapas de crecimiento y maduración, puede influir en la aparición posterior de condiciones adultas y rasgos de comportamiento, incluso si estos factores ambientales son cronológicamente distantes. La investigación en carnívoros domésticos ha sido dirigida principalmente por expertos en campos especializados de la medicina veterinaria, incluyendo nutrición, reproducción, microbiología gastrointestinal y comportamiento, con una escasez de experiencia en DOHaD que cruza las disciplinas relevantes.®
Esta revisión proporciona una visión general de los principales factores de riesgo ambientales en cachorros y gatitos que pueden afectar la aparición de obesidad, enteropatía crónica (CE) y problemas de comportamiento en la edad adulta. Estos trastornos crónicos son comunes en los carnívoros domésticos, difíciles de tratar y tienen importantes efectos nocivos sobre la salud, la calidad de vida y potencialmente la longevidad (10-12). El comportamiento difícil puede conducir a una ruptura en el vínculo humano-animal, y puede resultar en abuso, renuncia o eutanasia de mascotas (13-15). Es posible que algunas de las variables modificables exploradas puedan representar riesgos continuos que comienzan o se hacen evidentes en la vida temprana, y algunas pueden ser manifestaciones de DOHaD, con cambios en el epigenoma en períodos de plasticidad del desarrollo. También existe la posibilidad de que las exposiciones desenmascaren los efectos de DOHaD. Las prioridades de investigación sugeridas se discuten para cada condición, basadas en la investigación existente en cachorros y gatitos, los factores en perros y gatos adultos que se sabe que están asociados con la condición y se hipotetiza que se establecen durante la vida temprana, y en el conocimiento de la programación del desarrollo en otras especies. Se proponen estrategias de investigación para aumentar nuestra comprensión del impacto a largo plazo del entorno temprano y los eventos de la vida de perros y gatos. Tales estrategias deben incluir estudios para determinar el papel de DOHaD como se ha hecho para otras especies. Estos estudios podrían permitir en última instancia la generación de directrices para informar la prevención de enfermedades desde la preconcepción. Esto no solo es importante para el bienestar animal, sino que debe considerarse en el contexto económico y social más amplio de la propiedad de perros y gatos.
Sobre la base de la evidencia en perros y gatos, humanos y animales de laboratorio, proponemos una línea de tiempo de exposiciones clave e hitos del desarrollo en cachorros y gatitos que dan forma y definen la «vida temprana». La vida temprana en esta revisión no pretende relacionarse con una edad cronológica fija o necesariamente con el mismo período de desarrollo considerado clásicamente en los estudios DOHaD en otras especies. Se utiliza para describir los períodos anteriores a la edad adulta en los que la maduración fisiológica y psicológica de los cachorros y gatitos puede verse afectada para bien o para mal, o surgen factores de riesgo modificables para enfermedades crónicas posteriores. Esto tiene como objetivo ayudar a enmarcar futuras investigaciones y alentar a los criadores, propietarios y veterinarios a adoptar un enfoque holístico, integrado y proactivo para promover la salud a largo plazo de las mascotas (10-12).
El contexto para la investigación sobre el desarrollo temprano de perros y gatos
Social
Los perros y gatos son apreciados como miembros de la familia en muchos hogares, lo que hace que su salud a largo plazo sea una alta prioridad para los propietarios. Los beneficios sociales de la propiedad de perros y gatos incluyen la promoción de la salud y el bienestar humanos (16, 17); Los perros también trabajan en una amplia variedad de roles de servicio. Si bien estos factores, combinados con una preocupación general por el bienestar animal, proporcionan una justificación para avanzar en nuestra comprensión de los factores de riesgo de enfermedades crónicas en los primeros años de vida, también significan que la aceptación de la investigación invasiva en estas especies es limitada; es probable que esta sea una de las razones del avance relativamente lento del conocimiento DOHaD en perros y gatos.
Con respecto a los estudios observacionales a gran escala, la dispersión de la población de mascotas hace que el estudio de las conexiones entre las exposiciones tempranas y tardías y los eventos sea particularmente difícil. No existe una red coherente de las partes relevantes a lo largo de la vida de una mascota. Por ejemplo, cada perro o gato individual puede tener un criador diferente que es responsable de su prenatal y los primeros 2-3 meses postnatales de vida, y cada mascota puede ser alojada posteriormente con un dueño diferente, que a su vez puede tener un veterinario diferente.
Económico
El tamaño de la población de mascotas y la importancia económica directa de las mascotas son medidas tangibles que ayudan a contextualizar la importancia de buscar vías para la medicina preventiva. Se estima que hay 92,9 millones de perros en Europa (25% de los hogares; datos de 2021) (18), y 83,7-88,9 millones en los Estados Unidos (45% de los hogares; datos de 2020) (19). La población total de gatos se estima en más de 113,6 millones en Europa (26 % de los hogares; datos de 2021) (18) y de 60,2 a 61,9 millones en los Estados Unidos (26 % de los hogares; datos de 2020) (19). Las ventas de productos alimenticios para mascotas ascendieron a 27 2 millones EUR en Europa en 2021 (18). En los Estados Unidos en 2021, las ventas de mercado fueron de $ 50.0 mil millones para alimentos y golosinas para mascotas, $ 34.3 mil millones para atención veterinaria y ventas de productos, y $ 9.5 mil millones para otros servicios fuera de la atención veterinaria, como alojamiento, aseo y seguro (20).
Biológico
Raza
La gran variabilidad fenotípica dentro de las especies caninas, y en menor medida las especies felinas, contribuye a la complejidad de la investigación en animales de compañía. La especie canina exhibe las más amplias diferencias morfológicas y de peso entre las razas de todas las especies de mamíferos terrestres. Más de 350 razas de perros están reconocidas por la Federación Cinológica Internacional (21). Los pesos adultos van desde 1 kg, para un Chihuahua, hasta más de 100 kg, para un Mastín Inglés. Además, muchos perros de compañía (hasta el 40% en el Reino Unido) son una mezcla de razas (22). La edad a la que se alcanza el peso corporal adulto se correlaciona con el tamaño de la raza del perro, que varía de ~ 9 a 10 meses para razas de juguete, pequeñas y medianas, a 11-15 meses para razas grandes y gigantes (23). La diversidad de tamaño es menos pronunciada en la población de gatos, en la que se reconocen 45 razas (24) y solo el 5-15% de los gatos tienen pedigrí (25). Los pesos de los gatos adultos varían de ~ 2 kg para un Munchkin a 10 kg para un Maine Coon (25).
Biología reproductiva
La comprensión de los factores de riesgo de la vida temprana para las enfermedades de los adultos y el potencial para la programación del desarrollo requiere un conocimiento de la biología específica de la especie de la concepción y el desarrollo fetal y neonatal (resumido para perros y gatos en la Figura Suplementaria 1, Tablas Suplementarias 1, 2). Esto permite que el momento de las exposiciones ambientales esté relacionado con la diferenciación de los tipos de células y el desarrollo de tejidos y órganos específicos. El desarrollo embrionario y fetal general es similar entre perros y gatos (26) con la excepción de la maduración de los ovocitos y la ovulación. La ovulación en los gatos es típicamente inducida por el coito (26), aunque la ovulación espontánea parece ser más que anecdótica (27). Los ovocitos se liberan en metafase II, por lo que la fertilización puede ocurrir tan pronto como llegan al oviducto (28). En perros, hay ovulación espontánea de ovocitos inmaduros en la profase I. La meiosis ovocitaria se reanuda después de ~ 48 h en el oviducto, y la fertilización ocurre a partir de 90 h después de la ovulación (28, 29). Otra diferencia en los perros es que los folículos se someten a luteinización preovulatoria, por lo que las concentraciones séricas de progesterona ya son altas en la ovulación (28, 29).
Hitos de los primeros años de vida
Los perros y los gatos comparten muchos hitos biológicos importantes con otras especies, pero el momento y los detalles biológicos difieren. Estos hitos incluyen eventos embrionarios y fetales (Figura suplementaria 1; Tablas suplementarias 1, 2), supervivencia neonatal, transición a alimentos sólidos y castración. Los períodos de desarrollo y maduración de órganos y órganos/sistemas en los que los factores externos pueden modificar su trayectoria de desarrollo son numerosos. Estos períodos críticos representan diferentes ventanas de oportunidad para promover un desarrollo beneficioso para la salud a largo plazo.
Exposiciones y eventos ambientales de la vida temprana como factores de riesgo para trastornos seleccionados en perros y gatos adultos
Obesidad en perros y gatos
Como afirma Kopelman (2000), «la obesidad puede definirse como una enfermedad en la que el exceso de grasa corporal se ha acumulado de tal manera que la salud puede verse afectada negativamente» (30). La obesidad se define como una enfermedad crónica recurrente, que a su vez puede predisponer a otras enfermedades no transmisibles, como la diabetes mellitus, las enfermedades cardiovasculares y el cáncer en perros y gatos (31). En el campo de la medicina veterinaria, más de 20 organizaciones veterinarias y asociadas nacionales e internacionales apoyan la clasificación de la obesidad como una enfermedad (32), que se considera el problema de salud número uno en los animales de compañía (33).
El sobrepeso y la obesidad tanto en perros como en gatos generalmente se miden determinando la puntuación de condición corporal (BCS), que se correlaciona bien con la masa de tejido adiposo (32-35). Sobre esta base, un estudio de perros en exposiciones familiares de mascotas en el Reino Unido informó que el 65% de los perros adultos tenían sobrepeso u obesidad, y el 9% tenía obesidad (10). En la encuesta de prevalencia de obesidad de 2018 en los Estados Unidos realizada por la Asociación para la Prevención de la Obesidad de las Mascotas (APOP), los veterinarios evaluaron que el 36.9% de los perros tenían sobrepeso y el 18.9% tenían obesidad (36). La obesidad y/o el sobrepeso en perros se asocian con muchas comorbilidades, deficiencias funcionales (37-43), una vida útil más corta (44) y una peor calidad de vida (45) (Figura 1). En la encuesta APOP 2018 para gatos, la prevalencia de sobrepeso y obesidad fue del 26 y 34%, respectivamente. La prevalencia de sobrepeso u obesidad en gatos adultos en las visitas de vacunación en Nueva Zelanda fue del 22 y 3%, respectivamente (50). En cuanto a los perros, el sobrepeso y/o la obesidad en los gatos se asocia con un mayor riesgo de una amplia gama de comorbilidades (46, 51, 52), una esperanza de vida reducida y un mayor riesgo de muerte (obesidad severa solamente) (53, 54), y algunos, pero no todos los datos, sugieren una calidad de vida reducida (55) (Figura 2).
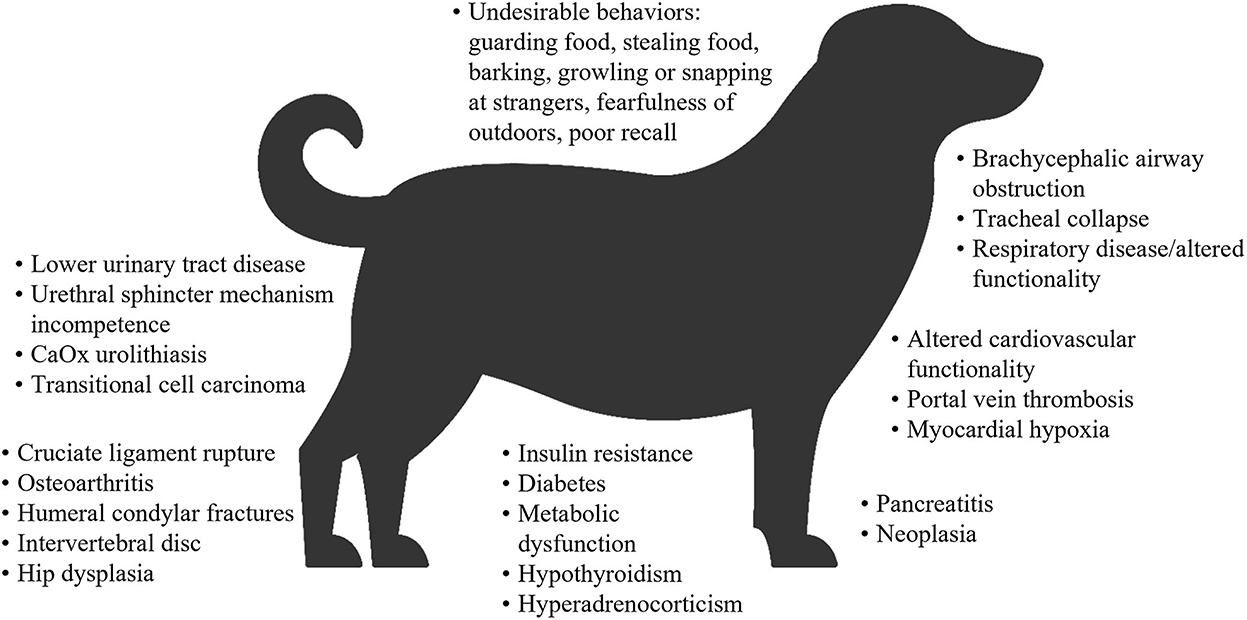 Figura 1. Resumen de las comorbilidades asociadas con la obesidad o el sobrepeso en perros (31, 37-40, 42, 43, 46-49). Algunas asociaciones fueron específicas para perros obesos o perros con sobrepeso. CaOx: oxalato de calcio.
Figura 1. Resumen de las comorbilidades asociadas con la obesidad o el sobrepeso en perros (31, 37-40, 42, 43, 46-49). Algunas asociaciones fueron específicas para perros obesos o perros con sobrepeso. CaOx: oxalato de calcio.
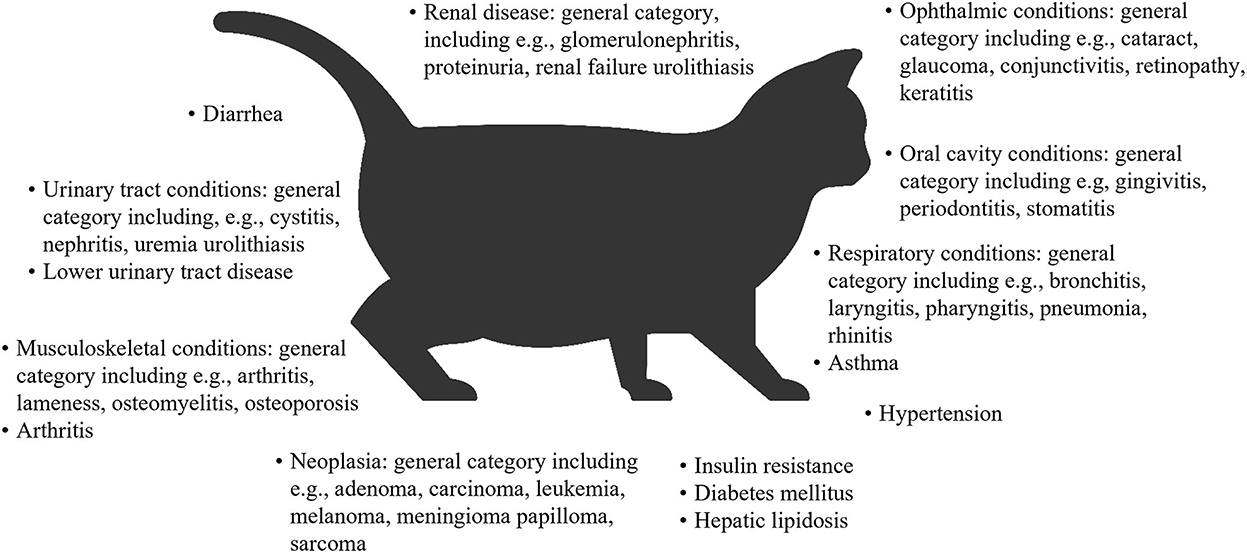 Figura 2. Resumen de las comorbilidades asociadas con la obesidad o el sobrepeso en gatos (31, 52, 53, 56-58). Algunas asociaciones se encontraron solo para ciertas puntuaciones de condición corporal dentro del rango de obesidad o sobrepeso. A menudo se encontraron asociaciones entre la obesidad o el sobrepeso y una categoría general de enfermedades o afecciones relacionadas con un órgano / sistema de órganos. En estos casos, se proporcionan ejemplos de algunas de las condiciones que el estudio incluyó en la definición de categoría.
Figura 2. Resumen de las comorbilidades asociadas con la obesidad o el sobrepeso en gatos (31, 52, 53, 56-58). Algunas asociaciones se encontraron solo para ciertas puntuaciones de condición corporal dentro del rango de obesidad o sobrepeso. A menudo se encontraron asociaciones entre la obesidad o el sobrepeso y una categoría general de enfermedades o afecciones relacionadas con un órgano / sistema de órganos. En estos casos, se proporcionan ejemplos de algunas de las condiciones que el estudio incluyó en la definición de categoría.
Factores de riesgo de obesidad en los primeros años de vida en perros adultos
Los factores de riesgo para la obesidad se pueden identificar en los primeros años de vida, ya en el período fetal. Por ejemplo, el bajo peso al nacer en Labrador Retrievers se ha asociado con sobrepeso en la edad adulta incluso después de ajustar por edad y estado de castración: el 70% de los perros con peso al nacer por debajo de la mediana tenían sobrepeso como adultos, en comparación con el 47% de los perros con pesos al nacer por encima de la mediana (5). No se encontró asociación entre la obesidad adulta y la tasa de crecimiento entre el nacimiento y el día 2 o entre el día 2 y el día 15. En contraste, en un estudio de hembras de perritos de colonia de Beagle criados en condiciones ambientales controladas, el peso al nacer no se correlacionó con el estado de sobrepeso adulto, pero la tasa de crecimiento rápido desde el nacimiento hasta las 2 semanas fue un predictor de sobrepeso adulto a los 2 años de edad (4). A la edad de 7 meses, BCS discriminó entre perros que tendrían sobrepeso como adultos y aquellos que tendrían un ligero sobrepeso o peso ideal (4). No se encontraron diferencias significativas entre los grupos de peso adulto en su ingesta de energía o gasto de energía en reposo corregido por el peso corporal metabólico a la edad de 4 meses. El equilibrio energético en reposo entre la edad de 7 y 10 meses fue significativamente mayor en los cachorros que tenían sobrepeso en comparación con el peso ideal en la edad adulta. Durante este estudio, los perros fueron alimentados ad libitum (tiempo restringido después del destete) con una dieta formulada para el crecimiento, o para adultos castrados, según corresponda. En un estudio diferente, un análisis retrospectivo de los registros de la práctica veterinaria encontró que los perros que eran obesos a los 3 años de edad (127 razas, 93% castrados) tenían un crecimiento más rápido en el peso corporal entre las 12 y 60 semanas de edad que el modelado en perros sanos en condición corporal ideal (59).
La castración es común para perros y gatos y a menudo se realiza antes de la pubertad mientras aún se están desarrollando. El momento exacto varía entre países, especies y razas, y sigue siendo un tema controvertido. La castración está bien establecida como uno de los factores de riesgo más importantes para la obesidad en adultos (46, 60-65). La castración de la prepubertad o la peripubertad en el contexto de esta revisión se considera una exposición ambiental temprana de relevancia para el riesgo de obesidad, aunque durante las etapas posteriores del desarrollo. El impacto potencial del sexo (60, 66, 67) y la edad de castración sobre los efectos de la castración en la obesidad adulta no están claros porque los hallazgos difieren según el estudio. Un estudio de cohorte prospectivo de Golden Retrievers identificó un riesgo ~ 42% mayor de obesidad en perros castrados entre 6 y 12 meses de edad en comparación con los castrados a los > 1 año de edad, pero no hay diferencia en el riesgo entre la castración a los <6 meses en comparación con 6-12 meses y >1 año (66). Por el contrario, en un estudio retrospectivo de registros veterinarios, la edad de castración (edades ≤ 6 meses, >6 meses a ≤ 1 año y >1 a ≤ 5 años de edad) no se asoció con el riesgo de obesidad (67). Cuando se examinaron los patrones de crecimiento de los perros de compañía de la misma fuente de datos patentada, la castración antes y después de las 37 semanas se asoció con ligeros cambios hacia arriba o hacia abajo en la trayectoria de crecimiento, respectivamente; Sin embargo, estos cambios fueron pequeños, lo que sugiere un impacto general limitado en el aumento de peso y, por lo tanto, en la obesidad futura (68). Las diferencias en el diseño del estudio y las razas de perros podrían explicar las aparentes inconsistencias entre estos estudios. De hecho, se informó que las interacciones entre el tamaño de la raza, la edad de castración y el número de visitas veterinarias por año afectan el riesgo de sobrepeso (67).
Los cambios hormonales resultantes de la castración podrían tener un efecto directo sobre el riesgo de obesidad. Los perros castrados tienen menores requerimientos de energía metabolizable que los perros sexualmente intactos (69, 70), y la castración puede aumentar el apetito indiscriminado (71). La evidencia sugiere que la castración también podría desenmascarar o aumentar los efectos de las exposiciones ambientales en el animal más joven. Los aumentos en el peso corporal de las perras Beagle después de la castración entre las edades de 7 y 10 meses fueron mayores en los perros identificados retrospectivamente como en riesgo de obesidad adulta al tener una tasa de crecimiento neonatal más alta que sus contemporáneos de un peso corporal adulto ideal (4). El crecimiento de Labrador Retrievers entre 2 y 21 días de vida se asoció con riesgo de obesidad en la edad adulta, pero solo en perros castrados (72).
Factores de riesgo de obesidad en los primeros años de vida en gatos adultos
En los seres humanos, la lactancia materna tiene un efecto protector contra la obesidad infantil en comparación con la alimentación con leche de fórmula (73), que podría estar asociada en parte con la presencia de leptina en la leche materna. La leptina es una hormona producida por el tejido adiposo que inhibe la ingesta de alimentos y modula el metabolismo de la glucosa; La principal fuente de neonatos puede ser la leche materna (74). En un pequeño estudio en gatos, las probabilidades de sobrepeso en la edad adulta fueron 3 veces menores en gatitos amamantados durante >6 semanas en comparación con <6 semanas, y hubo una predisposición al sobrepeso con una duración de lactancia de 11 semanas o menos (75). Los investigadores plantearon la hipótesis de que un corto período de succión podría conducir a perturbaciones en el desarrollo de mecanismos de control para la acumulación de grasa y la composición corporal a través de la reducción de la ingesta de leptina. Una tasa de crecimiento rápida en los gatos es un factor de riesgo clave para la obesidad. Una comparación de gatos de colonia alimentados ad libitum que tenían sobrepeso con aquellos de peso ideal en una mediana de 8,5 años de edad, mostró una asociación significativa entre la tasa de crecimiento entre 3 y 12 meses y el estado de sobrepeso posterior (76). En un estudio diferente que modeló el crecimiento de gatos de colonia alimentados ad libitum desde el destete, la tasa de crecimiento temprano indicada por el peso a las 15 semanas de edad fue un predictor significativo de sobrepeso a los 9 años (77). Las hipótesis para explicar estas asociaciones incluyen factores genéticos, epigenéticos e intrauterinos, además de la actividad física, la calidad de los alimentos, el comportamiento alimentario y el microbioma intestinal (76).
La tasa de crecimiento más rápida, el tamaño más pequeño de la camada, el menor peso al nacer y el sobrepeso materno antes del embarazo se asociaron con una predisposición de los gatitos a tener sobrepeso a los 8 meses de edad en un estudio centrado en factores genéticos y diseñado para reducir el potencial de factores de confusión no genéticos y diferencias epigenéticas (78). A pesar del diseño del estudio, algunos de los hallazgos sugirieron que la programación del desarrollo podría haber desempeñado un papel. Los autores especularon que la epigenética podría ser la base de la correlación negativa débil pero significativa del tamaño de la camada con el sobrepeso a los 8 meses. Además, las diferencias epigenéticas podrían haber contribuido a la observación de que, aunque tanto las madres con sobrepeso frente a las madres delgadas y de peso variable, como el sexo masculino frente al femenino se asociaron con un aumento de peso más rápido de los gatitos, esta relación se volvió estadísticamente significativa para el fenotipo materno más tarde que la diferencia sexual.
Al igual que en los perros, la castración de gatitos machos y hembras es un factor de riesgo para la obesidad adulta (65, 79, 80). Si bien la castración se asocia con un aumento del apetito y la ingesta de alimentos (81-83), también se asocia con menores requisitos de energía de mantenimiento (70, 81, 84). No hay pruebas suficientes en gatos para saber si la edad de castración se asocia con el riesgo de obesidad. Sin embargo, los cambios diferenciales en el apetito se han asociado con la edad de castración; Se observó hiperfagia aguda en gatas castradas a las 31 semanas de edad, pero no en las castradas a las 19 semanas de edad (85). Estos cambios de comportamiento pueden estar asociados con los efectos de la castración en las hormonas relacionadas con el apetito, como la grelina, la leptina, la adiponectina y el péptido-1 similar al glucagón (86). Por ejemplo, en un estudio de gatos machos adultos, las concentraciones séricas de adiponectina disminuyeron rápidamente después de la castración, y dentro de los 7 días, hubo un aumento significativo en la concentración sérica de grelina (83).
Posibles prioridades de investigación
Nutrición
El papel de la nutrición temprana en el desarrollo de la obesidad adulta exige una investigación más extensa y diversa. En cachorros y gatitos y sus padres, la nutrición es relativamente fácil de modificar tanto en la investigación como en entornos del «mundo real», y es probable que tenga múltiples impactos en los factores asociados con la obesidad (87-91). En su forma más simple, la ingesta excesiva crónica de calorías que comienza a una edad temprana da como resultado una acumulación progresiva de grasa corporal que finalmente se manifiesta como obesidad adulta. Sin embargo, una gran cantidad de evidencia en otras especies, incluidos los humanos, muestra que los insultos nutricionales tanto en el útero como en el postnatal pueden programar la obesidad posterior y otros trastornos metabólicos (92, 93). Los modelos de obesidad en especies politócicas demuestran que la mala nutrición materna (cuantitativa y/o cualitativa) puede modular aspectos de la deposición de grasa y la homeostasis energética en la descendencia a través de mecanismos epigenéticos (89, 94, 95). Se cree que las alteraciones en el desarrollo del eje hipotálamo-tejido adiposo de la descendencia son particularmente importantes para los rasgos obesogénicos, que se manifiestan como cambios estructurales, mala programación de la regulación del apetito que favorece las vías orexigénicas, leptina central y resistencia a la insulina, y alteraciones en la inervación noradrenérgica del tejido adiposo (96, 97). Se necesita investigación para determinar si el bajo peso al nacer en cachorros, como factor de riesgo para la obesidad adulta, es un ejemplo de programación fetal de un fenotipo «ahorrativo», por el cual un perfil metabólico establecido para hacer frente a una nutrición inadecuada durante el embarazo, más tarde se convierte en un factor de riesgo para la obesidad en el contexto de una nutrición postnatal abundante. La nutrición paterna en modelos animales de laboratorio también puede programar rasgos obesogénicos en la descendencia (98), pero esto no parece haber sido investigado aún en animales de compañía.
Los rasgos obesogénicos también pueden ser sensibles al entorno nutricional postnatal a medida que el desarrollo de órganos y vías hormonales continúa después del nacimiento en mamíferos (87). La literatura sobre la maduración postnatal de los carnívoros domésticos es limitada y, como en otras especies, el momento depende de los órganos involucrados (Tablas complementarias 1, 2). Por ejemplo, los cambios en la morfología de órganos como la glándula suprarrenal pueden ocurrir durante el primer año (99), la maduración funcional de los procesos digestivos puede no ocurrir hasta los 3 meses (100) y el sistema inmunológico puede no alcanzar todas las características adultas hasta los 12 meses (101). La mielinización del neocórtex continúa aumentando hasta ~ 9 meses después del nacimiento (102). Sin embargo, es razonable plantear la hipótesis de que la plasticidad del desarrollo se concentra en el período de lactancia. Se necesita investigación en perros y gatos para determinar los efectos de la dieta en la madre gestante y lactante sobre la cantidad y calidad del calostro y la leche, y si estos efectos tienen consecuencias para la composición corporal y el metabolismo de la descendencia adulta. La evaluación del impacto de la ingesta de alimentos y las intervenciones nutricionales de los primeros días de vida es particularmente relevante para los cachorros y gatitos de bajo peso al nacer al considerar las intervenciones nutricionales; El rápido crecimiento de recuperación se asocia con un mayor riesgo de obesidad adulta en otras especies (97).
Crecimiento
Tanto en cachorros como en gatitos, las tasas de crecimiento más altas se han asociado con la obesidad adulta (59, 77). No está claro si y cómo los aspectos de la regulación del equilibrio energético durante el crecimiento predisponen a los adultos a ser obesos o de peso ideal. Las disminuciones posprandiales de la grelina acilada, una hormona intestinal orexigénica, se retrasan en las hembras de Beagle de 7 meses de edad ya identificadas como en una trayectoria hacia el sobrepeso adulto, y esto puede promover la ingesta excesiva de alimentos (4). La concentración plasmática basal de leptina se asocia positivamente con la adiposidad, pero no parece ser un predictor temprano de aumento de peso. En humanos; La evidencia sugiere que el papel principal de la leptina es señalar las bajas reservas de grasa corporal en situaciones de balance energético negativo (4, 103). Se necesita investigación en poblaciones de estudio más grandes con diferentes razas y sexos para caracterizar aún más la dinámica del equilibrio energético durante el crecimiento asociado con la obesidad adulta, y para evaluar cualquier papel de la programación del desarrollo y los desencadenantes ambientales. Idealmente, los estudios de crecimiento y obesidad deberían evaluar la composición corporal. Sin embargo, mientras que por razones prácticas BCS se usa más comúnmente para evaluar la adiposidad, las escalas de puntuación solo se han validado adecuadamente en perros y gatos adultos. Los cachorros y gatitos tienen diferentes perfiles de composición corporal y morfologías en comparación con perros y gatos adultos (104-106), lo que hace que el diagnóstico del estado de sobrepeso con escalas BCS diseñadas para adultos sea insatisfactorio. Se podría lograr fácilmente una mayor objetividad y más uniformidad entre los estudios evaluando el crecimiento contra los estándares de tasa de crecimiento ahora disponibles para una amplia gama de diferentes tamaños de razas a partir de las 12 semanas de edad (68, 107). Estos estándares serán valiosos para facilitar la investigación del DOHaD en obesidad mediante la identificación de un crecimiento rápido o lento a una edad temprana, y para la determinación de casos en la composición corporal y los estudios metabólicos.
Microbiota intestinal
Se han observado diferencias en la microbiota intestinal y/o el microbioma de perros y gatos adultos obesos frente a los adultos delgados (108-112) y se han caracterizado cambios en perros y gatos obesos durante los estudios de pérdida de peso impulsados por la dieta (108, 113, 114). Sin embargo, las asociaciones entre la dieta, la microbiota intestinal, las hormonas enteroendocrinas y las alteraciones metabólicas son complejas, con estudios que informan hallazgos contrarios (113, 115, 116). Cuando se evaluaron los efectos de las proporciones de macronutrientes en las dietas alimentadas tanto a las madres como a sus gatitos, la composición de la dieta previa al destete no afectó el perfil de las poblaciones bacterianas en las heces de gatitos a las 8 semanas, pero sí moduló los niveles de expresión de genes en las vías metabólicas y de glucosa en muestras de sangre tomadas a las 18 semanas (117). Los hallazgos se invirtieron para una comparación entre dos dietas posteriores al destete. Lo que no se ha investigado directamente es ninguna asociación entre la microbiota intestinal tal como se está desarrollando en el cachorro y el gatito y la obesidad adulta, y el potencial de la nutrición temprana para influir en esto. La investigación en otras especies sobre programación del desarrollo sugiere que podría ser una vía fructífera de investigación (118, 119).
En ratones, la microbiota intestinal media cambios en la acetilación global de histonas y la metilación del ADN tanto localmente en las células del colon como distalmente en tejidos como el hígado y el tejido adiposo blanco (120). Estos cambios mediados por microbios se han demostrado en especies distintas de perros y gatos durante los primeros años de vida en un momento en que la microbiota intestinal se está desarrollando (121). Los metabolitos microbianos tienen un papel directo en las modificaciones epigenéticas, y la composición de la microbiota intestinal es relevante porque el perfil de los subproductos metabólicos de los componentes de la dieta, como los ácidos grasos de cadena corta (AGCC), puede diferir entre las especies bacterianas (120, 122). Factores como la lactancia materna frente a la alimentación con biberón, el estilo de vida, el medio ambiente y la exposición a los antimicrobianos también pueden afectar los rasgos obesogénicos a través de sus efectos sobre la microbiota emergente (123, 124).
Castración
La fuerza de la castración como un factor de riesgo modificable para la obesidad tanto en perros como en gatos exige una mayor comprensión de las interacciones entre las hormonas sexuales y la dieta sobre las hormonas relacionadas con el apetito y los metabolitos sanguíneos (86). El impacto de la castración en diferentes etapas de desarrollo (temprano vs. tardío) debe ser diseccionado para resolver las diferencias entre los estudios y explorar las diferencias de sexo, especie y raza. El impacto de las exposiciones ambientales, como la nutrición y la tasa de crecimiento durante los primeros días/semanas/meses, sobre los efectos de la castración posterior está poco investigado, pero los datos existentes justifican estudios prospectivos longitudinales adicionales (4). Una pregunta que debe abordarse es si la castración desenmascara o potencia los efectos de la programación del desarrollo de cachorros o gatitos.
Interacción de las exposiciones ambientales con susceptibilidades genéticas al sobrepeso y la obesidad
La interacción de los factores de riesgo genéticos con variables modificables en el desarrollo puede aumentar o disminuir la probabilidad de fenotipos particulares. En los seres humanos, los genes enriquecidos o sólo expresados dentro del sistema nervioso central tienen un papel central en la biología de la obesidad (125). El conocimiento de las susceptibilidades genéticas puede ayudar a los investigadores a diseñar estudios sobre programación del desarrollo e interpretar sus resultados.
Las razas de perros como Pug, Beagle, Golden Retriever, English Springer Spaniel, Border Terrier, Labrador Retriever y Cavalier King Charles Spaniel tienen un mayor riesgo de sobrepeso que los perros cruzados (126, 127), mientras que los gatos domésticos de pelo corto tienen un mayor riesgo de obesidad (127). Los genes candidatos para variantes genéticas sospechosas de aumentar el riesgo de obesidad en perros incluyen POMC, FTO, PPARG, MC4R y MC3R, INSIG2, GPR120 (127). Las variantes genéticas pueden estar restringidas a un pequeño número de razas, por ejemplo, una deleción de 14 pares de bases en POMC asociada con la obesidad y la motivación alimentaria encontrada en Labradores y Flat-coated Retrievers (128, 129). Los factores de riesgo genéticos deben ser una consideración en los estudios que investigan el impacto del entorno de la vida temprana en la obesidad. Los estudios de asociación de todo el genoma podrían ayudar a dilucidar los antecedentes genéticos de la obesidad en animales de compañía y existe un valor potencial tanto dentro de la raza como en los enfoques de raza y de raza cruzada a gran escala.
Enteropatía crónica en perros y gatos
La enteropatía crónica es un término general que abarca subgrupos de trastornos intestinales crónicos basados en la respuesta al tratamiento: enteropatía sensible a inmunosupresores [IRE, anteriormente conocida como enfermedad inflamatoria intestinal idiopática (EII)], enteropatía sensible a los alimentos y enteropatía sensible a antibióticos (12). La prevalencia de CE reportada en diferentes estudios varía de 1 a 18% (12). En gatos, la IRE coexiste frecuentemente con el linfoma de células pequeñas (130), que se considera que cae bajo el paraguas de CE en esta especie (131). Aunque la etiología subyacente de cada subtipo de CE no está clara, y puede no ser la misma, son afecciones inflamatorias crónicas, y la patogénesis refleja las interacciones entre la microbiota intestinal y los sistemas inmunes intestinales en el contexto de factores ambientales como la dieta y las susceptibilidades genéticas en algunas razas.
Factores de riesgo de la vida temprana para la enteropatía crónica en perros adultos
Hay muy pocos datos sobre los factores de riesgo de la AE en los primeros años de vida en perros; Sin embargo, estos datos limitados implican una amplia gama de variables que justifican una investigación completa. Los cachorros que históricamente se habían presentado en las etapas agudas de la infección por parvovirus canino a una mediana de 12 semanas de edad, tenían un mayor riesgo de signos gastrointestinales crónicos informados por el propietario en la vida posterior que los perros de control que se presentaron en la clínica veterinaria para un control de rutina o para signos no asociados con parvovirus [odds ratio 5.33 (IC 95%: 2.12–14.87)] (132). Un estudio similar también encontró que la enteritis previa por parvovirus era un factor de riesgo para los signos gastrointestinales persistentes, y entre los perros que se habían recuperado de la infección por parvovirus, los marcadores de gravedad de la enfermedad se asociaron con ese riesgo (133). En otro estudio, los factores de riesgo modificables tempranos para la CE en la edad adulta incluyeron la vacunación de la madre durante el embarazo, el tipo de alimento sólido alimentado a los cachorros durante los primeros 6 meses y la condición corporal del cachorro («delgado» en lugar de «peso normal») (134). Estos resultados deben interpretarse con precaución debido a las limitaciones metodológicas, como los cuestionarios retrospectivos de los propietarios, el sesgo de los participantes y los tipos de dieta amplios que no se controlaron nutricionalmente. En una revisión retrospectiva de los registros veterinarios de un hospital de enseñanza médica en los Estados Unidos, la castración se asoció con un aumento de las probabilidades de EII en machos y especialmente en perras (odds ratios para castrados vs. sexualmente intactos 1.43 y 2.0, respectivamente, p < 0.05 para ambos) (135). Los autores plantearon la hipótesis de que los mismos efectos antiinflamatorios y antioxidantes del estradiol demostrados en modelos murinos podrían proteger contra la EII en perros.
Factores de riesgo de la vida temprana para la enteropatía crónica en gatos adultos
Aunque la enteropatía crónica en gatos está bien descrita en la literatura (131, 136), no se encontraron estudios que hayan investigado factores de riesgo modificables en gatitos.
Posibles prioridades de investigación
La alteración de la microbiota intestinal madura, que podría deberse a la dieta o a los antimicrobianos, se asocia con un mayor riesgo de EII posterior en humanos o colitis experimental en modelos animales (137). La investigación sugiere que las modificaciones epigenéticas subyacen a las interacciones entre la dieta, el sistema inmunológico y la microbiota en el desarrollo de enfermedades crónicas, incluida la EII (121, 138).
Sugerimos que la investigación de cualquier asociación entre la microbiota intestinal en cachorros y gatitos y el desarrollo de CE en la edad adulta debe ser una prioridad de investigación (Figura 3). Existen interrelaciones complejas entre la microbiota intestinal, el metabolismo del huésped, el sistema inmunitario, la inflamación intestinal y la salud o disfunción gastrointestinal (139). ¿Existe una relación entre la microbiota intestinal que se desarrolla en cachorros y gatitos y la que se encuentra en perros y gatos adultos con CE, que difiere de la de adultos sanos? ¿Las perturbaciones en la microbiota intestinal en desarrollo afectan la maduración del sistema inmunológico y la adquisición de tolerancia de manera que predisponen a los cachorros y gatitos a la EC posterior? ¿Los efectos de la disbiosis de la microbiota en el metaboloma intestinal en estas mascotas programan epigenéticamente la susceptibilidad a la EC futura y/o disbiosis? Las piezas del rompecabezas se han caracterizado en cachorros y gatitos (140, 141), y por separado en adultos con CE (139, 142, 143), pero aún no se ha establecido la existencia de un vínculo entre estos.
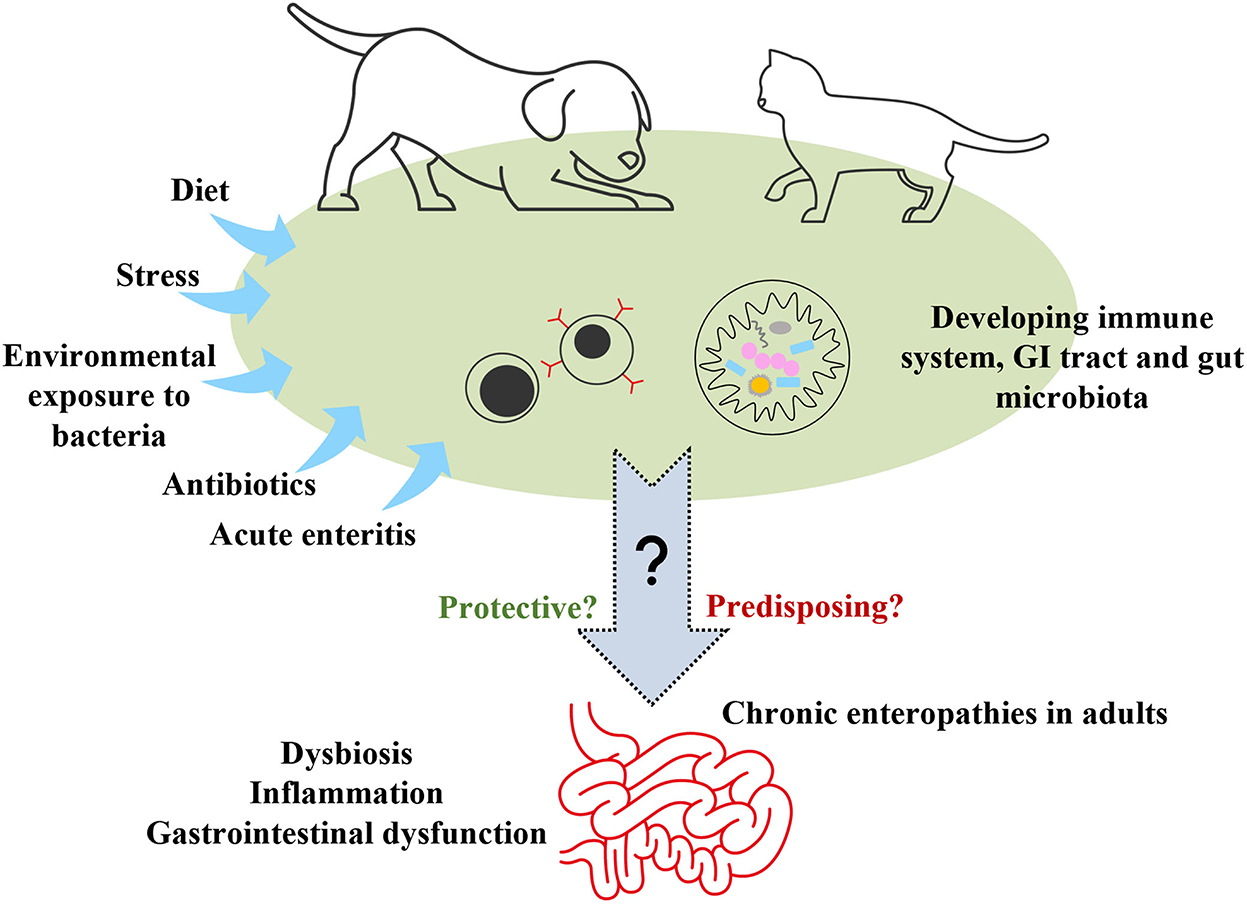 Figura 3. Exposiciones ambientales tempranas para investigar como posibles factores de riesgo para enteropatías crónicas en la edad adulta. GI gastrointestinal.
Figura 3. Exposiciones ambientales tempranas para investigar como posibles factores de riesgo para enteropatías crónicas en la edad adulta. GI gastrointestinal.
La disbiosis bacteriana se define como alteraciones en la composición de la microbiota intestinal bacteriana que conducen a cambios funcionales en el transcriptoma microbiano, proteoma o metaboloma, y / o disminución de la diversidad bacteriana (139, 144, 145). Se informa que el 72%-79% de los perros y el 76% de los gatos con CE tienen disbiosis según lo evaluado por los índices de disbiosis (142, 146, 147). La investigación en humanos y modelos animales sugiere que el papel de la disbiosis en la patogénesis de la EII podría ser causal (148, 149). Esto hace que el desarrollo de la microbiota intestinal en cachorros y gatitos, y las perturbaciones de esta, sean de particular interés como un factor de riesgo potencial para la CE.
La posibilidad en perros de transferencia bacteriana intrauterina de la madre al feto es controvertida, pero después del nacimiento, los datos sugieren que la madre siembra las bacterias iniciales y su perfil microbiano individual juega un papel fundamental en la configuración de la microbiota intestinal de su camada (150). La riqueza de especies bacterianas en el intestino neonatal aumenta a partir del día 2 después del nacimiento, y la microbiota intestinal cambia significativamente con la edad durante el período de lactancia y destete (151). Los mayores cambios en la microbiota de cachorros sanos se produjeron entre las 5 y 6 semanas de edad en una investigación (151), aunque las diferencias entre la microbiota de la descendencia y la madre todavía eran evidentes a las 8 semanas en otro estudio (152), y podrían ocurrir pequeños cambios hasta 1 año de edad (153). Al igual que con los cachorros, la microbiota intestinal de los gatitos sanos se desarrolla sustancialmente durante la lactancia y el destete, aunque el perfil adulto podría no alcanzarse completamente en esos períodos (154, 155). En un estudio en gatitos, los cambios en el microbioma todavía eran evidentes a las 18-30 semanas de edad, pero se habían estabilizado a las 30-42 semanas (156). En otro estudio, el microbioma fue relativamente estable en gatitos de 8 a 16 semanas (141, 157). Es evidente que se necesita más investigación longitudinal. Se afirma que es probable que la microbiota establecida en cachorros y gatitos sea generalmente estable durante la vida adulta sana como para los humanos, pero esto debe verificarse (139, 151, 152).
Es probable que la relevancia de las influencias dietéticas en la microbiota intestinal y el microbioma intestinal en cachorros y gatitos para el estado de salud gastrointestinal posterior sea multifactorial. La investigación existente debe ampliarse para investigar la dieta temprana como un riesgo potencial o factor protector para la CE. Por ejemplo, la suplementación pre y probiótica de Gran Danés en la última semana o las últimas 4 semanas de embarazo reduce el riesgo de gastroenteritis neonatal en su descendencia (158). Se plantea la hipótesis de que este efecto protector se transmite a través del vínculo entero-mamario, dado que en otros estudios alimentar a las madres con pro y / o prebióticos mejoró las propiedades inmunes de su calostro (159, 160). Otra hipótesis (no mutuamente excluyente) es que el efecto está mediado por la selección de bacterias promotoras de la salud en la madre que luego colonizan a los recién nacidos.
Las perturbaciones más profundas de la microbiota intestinal son las causadas por el uso de antibióticos. Se plantea la hipótesis de que el uso de antibióticos podría ser un factor de preparación importante en los cachorros para la EC posterior. En los seres humanos, el tratamiento con antibióticos en el primer año postnatal se asocia con un mayor riesgo de desarrollo posterior de EII (161-163). La diarrea aguda es común en los cachorros, y a menudo se trata con antibióticos. Los cambios en la microbiota fecal en perros con diarrea aguda y los grupos bacterianos involucrados no se informan consistentemente que sean los mismos que en la diarrea crónica, aunque la reducción en las concentraciones fecales de AGCC es un hallazgo compartido (139, 145, 164). En un estudio controlado prospectivo, el metronidazol (un tratamiento antibiótico común para la diarrea aguda) alteró significativamente el microbioma fecal y el metaboloma de perros sanos, incluida una disminución en la abundancia de Fusobacterias, que son bacterias productoras de AGCC clave, y la principal bacteria convertidora de ácidos biliares, Clostridium hiranonis, que se asoció con una reducción de los ácidos biliares secundarios (165). Los cambios persistieron en casi la mitad de los perros durante al menos 4 semanas. Los efectos a largo plazo de dicho tratamiento en cachorros que aún establecen una microbiota intestinal normal deben explorarse en estudios sobre programación del desarrollo. Se demostró que un tratamiento antibiótico (20 o 28 días) en gatos de 2 meses de edad con enfermedad del tracto respiratorio superior retrasa la maduración de su microbiota intestinal en comparación con los gatos sanos no tratados (166). La duración de los efectos difirió entre los antibióticos; El impacto de la amoxicilina-clavulanato en el microbioma ocurrió principalmente durante el tratamiento, mientras que el impacto de la doxiciclina se observó de 1 a 3 meses después de la retirada del antibiótico (166). La investigación debe extenderse al uso de antibióticos en perras preñadas; Los datos en humanos y ratones sugieren que este es un factor de riesgo para la enfermedad gastrointestinal en la descendencia (167, 168).
No se sabe si la microbiota intestinal influye en el desarrollo temprano del tracto gastrointestinal y la susceptibilidad a enfermedades crónicas a través de modificaciones epigenéticas en cachorros y gatitos. Sin embargo, los datos de estudios con ratones apuntan a la importancia de la microbiota intestinal en la modulación del desarrollo postnatal del intestino a través de la metilación del ADN de los genes en las células epiteliales intestinales asociadas con la inmunidad, el metabolismo y la regulación vascular (122, 162). Se sabe que los cambios en los metabolitos bacterianos asociados con la CE en perros influyen en otras especies en las modificaciones epigenéticas que afectan las vías inmunes e inflamatorias. Por ejemplo, la disminución de la abundancia fecal de Fusobacterium y Faecalibacterium en perros con CE se asocia con concentraciones fecales reducidas del propionato de AGCC (147, 169). Los ácidos grasos de cadena corta pueden regular las modificaciones epigenéticas al inhibir las histonas desacetilasas (HDAC) y contribuir con los donantes de acetilo para las modificaciones del ADN o las histonas.
Las anomalías o deficiencias en las respuestas inmunes a los antígenos ambientales, junto con las susceptibilidades genéticas, parecen desempeñar un papel central en el desarrollo de la CE en perros (170). La posibilidad de que parte de la inmunopatogénesis en perros con CE tenga orígenes en cambios epigenéticos fue planteada por una investigación de la expresión intestinal reducida de IgA mucosa encontrada en estos perros (171). La hipermetilación del gen para TACI se asoció negativamente con la expresión de IgA mucosa; Los autores plantearon la hipótesis de que tales cambios en el estado de metilación podrían haber sido inducidos por mediadores inflamatorios y la exposición del intestino a una microflora intestinal alterada (171). Por lo tanto, tales mecanismos podrían ser un vínculo entre las exposiciones ambientales durante el desarrollo y el riesgo de EC posterior.
En todas las vías de investigación sugeridas, los períodos de desarrollo de particular interés en cachorros y gatitos incluyen la colonización inicial del intestino neonatal, el destete y la transición de las instalaciones de cría a nuevos propietarios, cuando la diarrea es común, coincidiendo con cambios en la dieta, estrés y exposición a diferentes ambientes microbianos. Es necesario estudiar grandes poblaciones para comprender las variaciones interindividuales en la microbiota; puede que no haya una sola microbiota «normal» y «saludable». Se necesitan estudios sólidos que utilicen dietas nutricionalmente específicas y registren solo la CE diagnosticada por veterinarios. Aquellos que realizan investigaciones en programación del desarrollo deben, por supuesto, considerar las susceptibilidades de la raza y las asociaciones genéticas independientes de la raza con la enfermedad. Las razas de perros susceptibles a la CE incluyen Weimaraner, Rottweiler, Pastor Alemán, Border Collie y Boxer (12, 172).
Problemas de conducta en perros y gatos
Los problemas de comportamiento en perros y gatos son comunes y pueden afectar su bienestar y calidad de vida (173), su relación con los humanos y su idoneidad para el trabajo de asistencia (174). El comportamiento difícil es frecuentemente citado por los propietarios como al menos una de las razones por las que entregan a sus mascotas a los centros de rescate de animales, siendo la razón principal para el 10% de los perros en un estudio canadiense reciente (13), y la única razón para el 27% de los perros y el 19% de los gatos en un estudio estadounidense (14). También pueden llevar a algunos propietarios a buscar la eutanasia electiva para sus mascotas (15). Aunque la naturaleza de los problemas de comportamiento es amplia, como la agresión hacia los humanos y otros animales, la ansiedad por separación y la suciedad en la casa, al menos algunos rasgos de comportamiento adverso perjudiciales para el futuro a largo plazo de los perros y gatos pueden atribuirse a su entorno de vida temprana.
La mayor parte del desarrollo neurológico ocurre durante la vida fetal; Continúa rápidamente en el neonato, pero la formación y maduración de mielina continúa hasta al menos las 36 semanas de edad en perros (175). Las regiones del cerebro se desarrollan a diferentes ritmos a lo largo de la vida temprana, por lo que potencialmente permanecen susceptibles a las exposiciones ambientales (175-177). El desarrollo de rasgos conductuales y cognitivos se puede considerar en diferentes fases: gestación, el período neonatal que incluye alimentación, estimulación neurológica y maternidad en las primeras 3 semanas, socialización temprana de ~ 3 a 12 semanas de edad, socialización tardía de 12 semanas hasta 6 meses, y el período de enriquecimiento, que puede extenderse a 1 año de edad (177, 178). Se cree que las experiencias durante cada período tienen efectos acumulativos sobre la capacidad de entrenamiento, la salud y el rendimiento (177, 178).
Factores de riesgo de los primeros años de vida para problemas de comportamiento en perros adultos
No se identificó ninguna investigación en perros que investigara los efectos del estrés materno o la dieta durante el embarazo en el comportamiento de la descendencia, excepto una mención de que los cachorros de madres desnutridas estaban extremadamente nerviosos además de mostrar anomalías físicas (179).
La mala atención materna y la socialización antes de los 3 meses de edad se han asociado con el miedo en los perros, y la mala atención materna sola también se asoció con una combinación de miedo, sensibilidad al ruido y ansiedad por separación (180). Estos datos se derivaron de una encuesta a los propietarios, pero otros estudios con medidas más objetivas muestran que el nivel de maternidad puede afectar el rendimiento de los perros en las pruebas cognitivas, las respuestas al estrés y el temperamento en la vida posterior. Sin embargo, algunos resultados de la investigación parecen ser contradictorios en cuanto a si una exposición ambiental tiene un efecto positivo o negativo. Por ejemplo, en un estudio prospectivo, los perros guía que habían experimentado una maternidad más intensa tenían habilidades de resolución de problemas más pobres y mostraron niveles más altos de ansiedad a los 14-17 meses de edad, los cuales se asociaron con un riesgo significativamente mayor de fallar el programa de entrenamiento de perros guía (174). En contraste, se demostró un beneficio de mayor cuidado materno en cachorros Beagle machos y hembras; La duración media de la atención materna diaria en sus primeras 3 semanas se correlacionó positivamente con la exploración y la latencia del primer grito, y se correlacionó negativamente con el estrés en las pruebas de aislamiento a las 8 semanas de edad (181). Un estudio con seguimiento a largo plazo encontró que un mayor nivel de cuidado materno de los perros pastores alemanes machos y hembras se asoció con un mayor compromiso físico y social (por ejemplo, recuperación de balón, aceptación positiva del manejo), así como agresión en adultos jóvenes a los 18 meses de edad (182). En resumen, los estímulos en el período de lactancia parecen afectar algunos comportamientos de los perros adultos, pero la dirección de las asociaciones reportadas no siempre es intuitiva o consistente, tal vez reflejando la complejidad de la biología, así como las diferencias entre estudios en las pruebas de comportamiento, edades y razas (181).
En una revisión de siete estudios observacionales en perros originarios de establecimientos de cría comercial de alto volumen y vendidos en línea o a través de tiendas de mascotas, se destacaron los factores de riesgo para problemas conductuales y psicológicos posteriores (183). En el mayor de estos siete estudios, los perros del Reino Unido adquiridos de fuentes como tiendas de mascotas e Internet tenían 1,8 veces más probabilidades de mostrar agresión hacia los humanos que los perros adquiridos directamente de los criadores (183, 184). En todos los estudios, la agresión fue el comportamiento problemático más común asociado con los establecimientos de cría comercial o las granjas de cachorros y las tiendas de mascotas. Aunque no se investigaron los factores causales, las posibles causas discutidas incluyeron estrés en la madre, estimulación neonatal insuficiente o excesiva, destete temprano y separación materna, y aislamiento social entre las 3 y 12 semanas de edad.
Para el período de socialización temprana, el sistema nervioso central se ha desarrollado a una etapa que permite el condicionamiento y el aprendizaje asociado (177). La socialización de los cachorros con congéneres familiares es importante para el desarrollo de la competencia de comunicación, y las interacciones tempranas con congéneres no familiares pueden influir en el riesgo de agresividad en la vida adulta (177, 185). Por ejemplo, se encontró que la restricción del contacto de un cachorro con congéneres en las 8 semanas posteriores a su primera exposición a otros perros en un entorno público estaba asociada con la agresión hacia perros desconocidos cuando tenían entre 1 y 3 años de edad (185). La socialización temprana con humanos es importante para respuestas posteriores al manejo, entrenamiento con correa y pruebas de esfuerzo (186).
Los rasgos conductuales y las habilidades cognitivas no sociales continúan desarrollándose en los cachorros durante los últimos períodos de socialización y enriquecimiento (187-189). En perros de trabajo candidatos jóvenes, las medidas de control inhibitorio, atención y cognición espacial mejoraron entre los 3 y los 12 meses de edad (187). En un segundo estudio longitudinal, el rendimiento de las tareas cognitivas mejoró entre la edad de ~ 9 semanas y 21 meses, y el fenotipo adulto para algunos rasgos podría predecirse a partir de los resultados de las pruebas en la edad de cachorro (188). Sin embargo, se sabe poco sobre exposiciones específicas en estos períodos que podrían influir en el curso del desarrollo del cerebro, y es probable que el contexto ambiental general, la raza y el sexo también desempeñen un papel (189). Los cuestionarios completados por cuidadores adoptivos de cachorros desde ~ 2 meses de edad hasta el inicio del entrenamiento formal de perros guía, mostraron un efecto conductual positivo de crecer en un hogar con otro perro y con criadores de cachorros más experimentados (189). Los cachorros que habían sido atacados o amenazados por un perro desconocido mostraron significativamente más «miedo dirigido al perro» y «agresión dirigida a extraños» a la edad de 12 meses de edad en comparación con los cachorros que no habían experimentado ese trauma y tenían peores resultados de entrenamiento (189). Sin embargo, no se especificó la edad a la que se produjo el trauma.
Un vínculo entre los cambios epigenéticos y el comportamiento social dirigido por humanos en perros se encontró en un estudio (190). La metilación del ADN de la región promotora del gen del receptor de oxitocina (OXTR) se midió mediante pirosecuenciación de bisulfito seguida de PCR específica de metilación en células epiteliales de la boca obtenidas de varios cánidos. Se identificaron cuatro sitios metilados diferencialmente 5′-citosina-fosfato-guanine_3′ (CpG). Posteriormente se estudiaron en una gran población (n = 217) de Border Collies. El estado de metilación del ADN no solo difirió entre mujeres y hombres, sino que también se asoció con su respuesta en una prueba de «enfoque amenazante» de una manera dependiente del sexo. Por ejemplo, una mayor metilación en un sitio específico de CpG en perras tendió a corresponder con una mayor probabilidad de apaciguar el comportamiento en la prueba, mientras que la relación opuesta se encontró en perros machos. Además, los sitios de CpG difirieron en si la metilación del promotor aumenta o disminuye los niveles de expresión de OXTR (190), tanto el estado neutro en sí como la interacción del sexo con el estado neutro no predijeron los niveles de metilación en los tres sitios de CpG investigados. Este estudio destaca la complejidad de las relaciones entre las modificaciones epigenéticas y el comportamiento en perros, y la necesidad de investigar los factores ambientales que influyen en la epigenética del gen OXTR.
Factores de riesgo de los primeros años de vida para problemas de comportamiento en gatos adultos
Hay una escasez de conocimiento sobre el efecto del entorno materno durante el embarazo y los problemas de comportamiento posteriores en la descendencia felina adulta. Sin embargo, un estudio mostró que cuando los gatitos de las madres que habían estado desnutridas durante el embarazo fueron criados en gatos no privados de alimentos, tanto el desarrollo físico como el conductual se retrasaron (179, 191). Los comportamientos afectados incluyeron el tiempo dedicado a jugar y el uso de una bandeja de arena. Además, en la edad adulta los gatos mostraron un marcado comportamiento antisocial y alternancia entre comportamientos dominantes y sumisos. Estas observaciones podrían representar potencialmente el DOHaD clásico en gatos. En otra investigación, la restricción de proteínas de los gatos durante la gestación tardía y la lactancia afectó negativamente los procesos de unión tanto en madres como en gatitos en los primeros 12 días después del nacimiento (192).
Hay un desarrollo neurológico continuo de los gatitos durante los primeros 3 meses postnatales (193). Esto se observa, por ejemplo, en la capacidad cada vez más sofisticada para responder al sonido, el desarrollo de la colocación visual y la coordinación binocular, y los comportamientos macroscópicos durante las interacciones con los hermanos (193). Las experiencias tempranas de la vida de los gatos afectan el comportamiento en la edad adulta (194). Por ejemplo, el manejo humano de los gatitos durante el período sensible para la socialización se ha asociado con un comportamiento más amigable hacia los humanos a la edad de 1 año (195, 196). Se desconoce hasta qué punto la modulación epigenética y/o las diferencias genéticas contribuyen a estas observaciones.
Posibles prioridades de investigación
Sugerimos que la investigación sobre los factores de riesgo para los problemas de comportamiento en perros y gatos debe diversificarse para incluir evaluaciones en el útero y más estudios de influencias ambientales en animales de 6 a 12 meses. Esto no debe ser a expensas de un mayor trabajo en neonatos cuando el desarrollo del cerebro es particularmente plástico. Los estudios neonatales se ven facilitados por la relativa facilidad con la que se puede controlar el medio ambiente para individuos relacionados en la misma camada, aunque esto no permite la distinción entre efectos genéticos y ambientales. Es necesario explorar resultados aparentemente contradictorios para el efecto de la atención materna en las respuestas al estrés, tal vez comparando diferentes desafíos e intensidades de estrés a diferentes edades en animales mantenidos en ambientes estandarizados. Se requieren más estudios a largo plazo para determinar la durabilidad y reversibilidad de los efectos del entorno de la vida temprana en el comportamiento adulto.
Se desconoce hasta qué punto la modulación epigenética impulsa los factores de riesgo de trastornos del comportamiento en perros y gatos. Los pocos datos existentes que vinculan los cambios epigenéticos con el comportamiento del perro (190) resaltan la complejidad de las relaciones involucradas. Los factores ambientales que influyen en la variación epigenética conocida de OXTR son especialmente importantes para explorar. Los polimorfismos en el gen del receptor de dopamina 2 (DRD2) están asociados con el comportamiento temeroso en algunas razas de perros (197), y una variante del haplotipo en este gen se asocia con la separación de la ansiedad en Golden Retrievers (198). Por lo tanto, DRD2 podría ser otro gen de interés para estudiar los cambios epigenéticos que afectan el comportamiento.
Los estudios en otras especies proporcionan una rica fuente de datos de comportamiento del desarrollo y, por lo tanto, hipótesis para perros y gatos. Varios trastornos cognitivos, conductuales y emocionales en los niños se han asociado con el estrés durante el desarrollo (199, 200). El estrés prenatal puede provocar cambios estructurales y funcionales en múltiples regiones del cerebro fetal en desarrollo, incluido el eje hipotálamo-hipófisis (199, 201). La metilación del ADN del gen del receptor de glucocorticoides y OXTR son ejemplos de mecanismos que se cree que vinculan las experiencias infantiles con los trastornos psiquiátricos y el temperamento, respectivamente (202, 203). Las experiencias previas a la concepción de los padres también pueden ser relevantes y pueden afectar las respuestas ansiogénicas de sus hijos y las generaciones posteriores de descendientes (204, 205). Los estudios con roedores destacan la importancia del momento exacto de las exposiciones ambientales y el dimorfismo sexual en las secuelas del desarrollo (206). Las tensiones que pueden afectar el comportamiento de la descendencia incluyen, por ejemplo, el trauma psicológico previo a la concepción de cualquiera de los padres, la dieta materna durante el embarazo, la separación temprana de la descendencia de su madre y los comportamientos de maternidad (201, 205, 207, 208). La evidencia de varias especies también vincula la composición de la microbiota intestinal con el desarrollo neurocognitivo y conductual, construyendo el concepto de un eje microbioma-intestino-cerebro (138, 209, 210).
Estrategias de investigación para comprender los factores de riesgo de la vida temprana para enfermedades crónicas y problemas de comportamiento en perros y gatos, y el papel potencial de la programación del desarrollo
Los orígenes del desarrollo de la salud y la enfermedad aún no se han confirmado en las mascotas, pero los ejemplos discutidos sugieren que es probable que la programación del desarrollo sea tan importante como en otras especies. Recomendamos un enfoque multidisciplinario concertado para explorar la programación del desarrollo en perros y gatos y para cerrar las grandes brechas de conocimiento en comparación con otras especies.
La investigación humana y las campañas de información sobre DOHaD se llevan a cabo en el contexto de una ventana crítica de desarrollo específica de la especie. Creemos que la ventana de tiempo crítica general implicada en el desarrollo de perros y gatos se extiende desde la preconcepción hasta el final del crecimiento, comprendiendo cinco períodos: preconcepción, gestación, período de lactancia, precastración o prepubertad del crecimiento temprano, crecimiento posterior a la castración o post pubertad hasta el tamaño adulto (Figura 4). Dentro de esta ventana, habrá diferentes períodos críticos y, a veces, superpuestos para diferentes aspectos de la salud y la enfermedad de acuerdo con la plasticidad del desarrollo de los tejidos relevantes. El límite de edad superior de esta «ventana» dependerá de la especie y la raza. Las variables de particular interés para la investigación de DOHaD en perros y gatos, tanto individualmente como en combinación, incluyen el entorno materno y neonatal, la nutrición y el aumento de peso asociado y / o la tasa de crecimiento, la microbiota intestinal, el estrés al destete y la castración (Figura 4). El trabajo en otras especies destaca la necesidad de estudios para considerar el período periconcepcional (211), el momento exacto de la exposición ambiental, las diferencias en la programación entre los sexos (212-214), el papel de la placenta (215), las influencias paternas en el epigenoma de la descendencia, la raza y la variación genética (216-218).
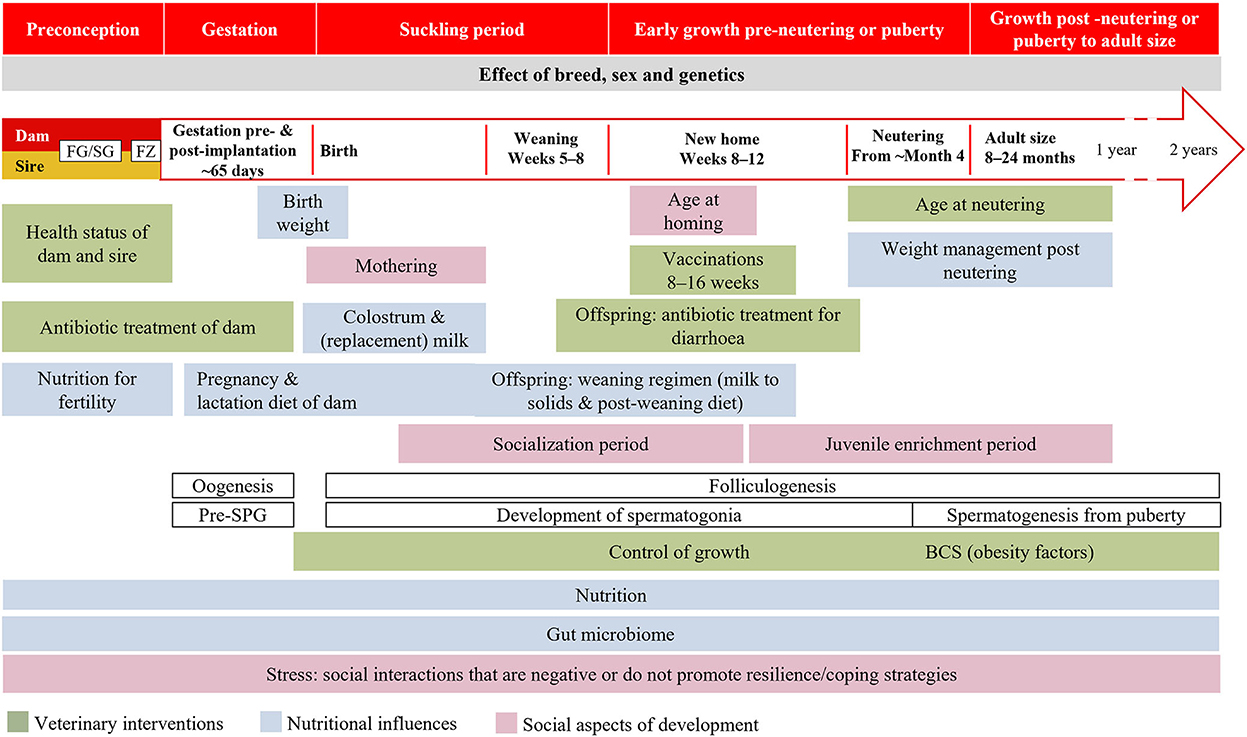 Figura 4. Ventanas de programación evolutiva propuestas en perros y gatos. BCS: puntuación de condición corporal; FG/SG: foliculogénesis/espermatogénesis; FZ: fertilización; Pre-SPG, prespermatogonia.
Figura 4. Ventanas de programación evolutiva propuestas en perros y gatos. BCS: puntuación de condición corporal; FG/SG: foliculogénesis/espermatogénesis; FZ: fertilización; Pre-SPG, prespermatogonia.
Es fundamental que toda la comunidad de investigación científica pueda acceder a grandes conjuntos de datos que abarquen datos de alta calidad de «toda la vida» y biobancos de muestras de tejido para explorar el impacto a largo plazo de las exposiciones en los primeros años de vida. Las colonias de investigación de gatos y perros con programas internos de cría son útiles para abordar el efecto de las intervenciones individuales, pero son poco comunes y no reflejan la situación de las mascotas domésticas, que están expuestas a muchos factores ambientales no controlados e interactivos. El análisis de grandes cohortes observacionales prospectivas de animales de propiedad privada, que tal vez abarquen hasta 15 años para abarcar toda la vida, puede identificar efectos de la programación que son pequeños en el individuo y muy variables entre individuos. Un desafío particular que debe abordarse es obtener datos longitudinales para un perro o gato que siga a ambos padres en el período previo a la concepción y se extienda a través de la gestación y la vida neonatal hasta la edad adulta y el final de la vida. Esto podría necesitar datos de dos criadores (uno para el padre y otro para la madre), al menos un propietario y probablemente al menos tres veterinarios (uno para el padre, un cuidador y otro para el cachorro o gatito). Estas partes interesadas deben comprometerse con las posibles implicaciones de amplio alcance y a largo plazo de DOHaD y las muchas oportunidades posibles de intervención. Las instituciones que crían y entrenan perros para funciones de servicio, y posteriormente monitorean su progreso, por ejemplo, perros guía, pueden proporcionar oportunidades de colaboración para los investigadores.
Aunque hay estudios longitudinales grandes, prospectivos y observacionales en curso, estos se dirigen principalmente a los perros después de que hayan abandonado el criador, por lo que carecerán de datos de los primeros 2 meses después del nacimiento. Por ejemplo, el proyecto Generation Pup operado por Dogs Trust en el Reino Unido se inició en 2016 para utilizar datos veterinarios y de propietarios de hasta 10,000 perros para identificar factores de riesgo modificables durante el desarrollo que afectan la salud y el bienestar de los adultos (219). La investigación investigará las relaciones entre el genotipo, el medio ambiente y los resultados de salud y comportamiento en diferentes etapas de la vida. También hay estudios de cohorte longitudinales específicos de la raza, como el Golden Retriever Lifetime Study dirigido por la Morris Animal Foundation en los Estados Unidos, que está recopilando datos sobre el estilo de vida, el medio ambiente, el comportamiento y la salud de 3,000 perros reclutados entre 2012 y 2015, incluidas muestras biológicas anuales (66, 220, 221). El proyecto epidemiológico Dogslife en curso (Universidad de Edimburgo, Universidad de Manchester, Universidad de Liverpool y el Kennel Club) recluta Labradores Retrievers de pedigrí del Reino Unido nacidos después de enero de 2010 (n = 6.084 perros en diciembre de 2015) (222, 223). Los propietarios completan cuestionarios cada mes durante el primer año de vida de su perro y cada 3 meses a partir de entonces (222, 223). En los Estados Unidos, el Proyecto de Envejecimiento Canino en curso (Universidad de Washington y Universidad de Texas A&M) ha reclutado a decenas de miles de perros de compañía para explorar aspectos de la «duración de la salud», es decir, el período de vida pasado libre de enfermedades (224). La cohorte de la Escuela Noruega de Ciencias Veterinarias establecida para investigar la enfermedad esquelética en cuatro razas de perros grandes (n = 700 cachorros reclutados 1998-2001), es notable como un ejemplo de una cohorte longitudinal en la que se reclutaron camadas de perros desde el momento del apareamiento de la madre, y se obtuvieron datos de criadores, propietarios y veterinarios (225, 226).
Los registros longitudinales de gatos y los estudios de cohortes parecen ser escasos. El histórico Estudio de Gatos de Bristol dirigido por la Universidad de Bristol es el primer estudio de cohorte de nacimiento reportado de gatitos (227). Los gatos se registraron entre las edades de 8 y 16 semanas, y los propietarios completan cuestionarios a intervalos establecidos. Los análisis reportados hasta la fecha incluyen la prevalencia y los factores de riesgo para la obesidad, y los signos del tracto urinario inferior informados por el propietario (228-230). El Registro de Fenotipo de Gato e Información de Salud en los Estados Unidos (UC Davis Veterinary Medicine) recolecta muestras de ADN de gatos sanos y enfermos con seguimiento a largo plazo cuando es posible (231).
Los datos sobre los mecanismos epigenéticos que vinculan las experiencias de la vida temprana con los trastornos de la edad adulta en perros y gatos son escasos; Son necesarios para ayudar a confirmar y comprender la programación del desarrollo, y desentrañar los efectos de los antecedentes genéticos. Los métodos para perfilar la metilación del ADN de todo el genoma están bien establecidos, y se puede lograr mucho antes de intentar identificar los genes específicos responsables de un fenotipo determinado epigenéticamente (232, 233). También deben estudiarse mecanismos epigenéticos distintos de la metilación del ADN, como las modificaciones de histonas, incluidas, entre otras, la metilación y la acetilación, y el ARN no codificante que puede regular la expresión génica durante la diferenciación y el desarrollo celular (234). Recientemente se ha establecido un repositorio disponible públicamente de datos epigenómicos caninos (BarkBase), que comprende los resultados de la secuenciación del ARN y los ensayos que determinan la accesibilidad de la cromatina en todo el genoma (235). La base de datos incluye 27 tejidos adultos diferentes y cinco tipos de tejidos fetales en cuatro puntos de tiempo embrionarios. El Royal Veterinary College ha instituido el Banco de Cerebros de Animales de Compañía, una colección estandarizada de tejido cerebral y otras muestras biológicas de perros y gatos sacrificados con afecciones neurológicas, junto con controles apropiados. Aunque tales bancos de tejidos podrían usarse para investigar cambios en el epigenoma, incluidos los asociados con la enfermedad, sin datos correspondientes sobre los aspectos ambientales del embarazo y la vida temprana, no proporcionarán evidencia de programación del desarrollo.
El efecto de las experiencias de la vida temprana y la programación del desarrollo en al menos algunas características fisiológicas se verá afectado por las diferencias genotípicas entre las muchas razas y razas mixtas. Dirigir la investigación a razas específicas sobre la base de su propensión a desarrollar la enfermedad o el comportamiento de interés puede ser ventajoso, por ejemplo, el Labrador Retriever para la obesidad.
Hay menos razas de gatos con las que lidiar en la investigación de DOHaD, pero en general las brechas de conocimiento son mayores que en los perros. Parece haber menos datos de campo longitudinales en gatos en comparación con perros, y probablemente haya menos conciencia pública sobre el impacto potencial de la programación del desarrollo en las enfermedades crónicas.
Observaciones finales
Existe evidencia directa de factores de riesgo en los primeros años de vida asociados con la obesidad y los problemas de comportamiento en perros y gatos, y en mucha menor medida la CE en perros. Sin embargo, se necesita una investigación prospectiva multidisciplinaria a largo plazo para confirmar DOHaD en estas especies. Los datos extensos de otras especies proporcionan una base científica para ayudar a priorizar los eventos y exposiciones de la vida temprana para la investigación. La diversidad de razas de perros y gatos, el manejo de la cría y los estilos de vida agrega complejidad a dicha investigación. Se cree que los criadores, propietarios y veterinarios tienen una ventana crítica de oportunidad en una o más de las etapas de la vida desde la preconcepción hasta el final de la fase de crecimiento del perro o gato en la que promover una programación beneficiosa para la salud a largo plazo. La apreciación por parte de cada uno de estos grupos de la ventana general de desarrollo también puede ayudar a fomentar la responsabilidad compartida, la transparencia y el intercambio de información.
Los perros y los gatos se consideran miembros de la familia y, sin embargo, la medicina veterinaria lucha para tratar afecciones comunes que afectan negativamente la calidad de vida de las mascotas, el vínculo especial propietario-mascota y los beneficios para la salud que las mascotas pueden aportar a las personas y a la sociedad. La medicina preventiva y las prácticas de cría desde la preconcepción en adelante deben tener una mayor prioridad y ser impulsadas por una mejor comprensión de la programación del desarrollo a nivel de la población.
Contribuciones del autor
VG, SC, GE, OF, AG, JS, CV, PC-P y FP contribuyeron sustancialmente a la concepción del artículo y a la interpretación de los datos presentados. Todos los autores críticos revisaron el manuscrito por su contenido intelectual importante. Los autores asumen toda la responsabilidad por el contenido científico del artículo y todos han aprobado la versión presentada.
Financiación
SC, GE, AG, JS, CV y PC-P recibieron estipendios de Royal Canin por su participación en una reunión de la Junta Asesora sobre Programación del Desarrollo (4 y 7 de diciembre de 2020). El apoyo editorial fue proporcionado por Susanna Ryan y financiado por Royal Canin SAS.
Conflicto de intereses
FP y VG son empleados de Royal Canin SAS. SC es un agente del gobierno francés y jefe de NeoCare ENVT, una unidad pública de investigación. La Unidad NeoCare fue parcialmente financiada por la División de Investigación Royal Canin de 2012 a 2021. Como tal, SC ha presentado resultados de investigación y recomendaciones clínicas en conferencias patrocinadas por Royal Canin. GE proporciona asesoramiento clínico veterinario al Waltham Petcare Science Institute, Waltham-on-the-Wolds, Leicestershire, Reino Unido. OF es un empleado de Wisdom Panel (Kinship), una compañía de cuidado de mascotas de Mars. AG es un empleado de la Universidad de Liverpool, pero su puesto cuenta con el apoyo financiero de Royal Canin, ha recibido una remuneración financiera por proporcionar material educativo, hablar en conferencias y trabajos de consultoría de Royal Canin, y también es miembro del Grupo Asesor Científico de Perros Guía. JS es un empleado del Laboratorio Gastrointestinal de la Universidad de Texas A&M, que ofrece pruebas de diagnóstico en base a una tarifa por servicio, y ha recibido honorarios de oradores y honorarios de consultoría de Royal Canin SAS, Nestlé Purina Petcare, ExeGI Pharma, Nutramax Laboratories y Hill’s Pet Nutrition. CV ha realizado trabajos de consultoría para una variedad de compañías de alimentos para mascotas (Nestlé Purina PetCare, Royal Canin, Mars Pet Care y Dechra Specific), ha participado como investigador en ensayos clínicos patrocinados por Royal Canin y Affinity Pet Care, desarrolla materiales educativos para Morris Institute, forma parte del Consejo Asesor Científico de FEDIAF y es miembro del Comité Global de Nutrición de WSAVA, y participa como orador o asistente en eventos de educación continua patrocinados u organizados por Royal Canin, Nestlé Purina PetCare y Hill’s Pet Nutrition.
El autor restante declara que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
El revisor AW declaró una afiliación compartida con el autor GE al editor de manejo en el momento de la revisión.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o reclamo que pueda ser hecho por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2022.944821/full#supplementary-material
Referencias
1. Mila H, Grellet A, Feugier A, Chastant-Maillard S. Impacto diferencial del peso al nacer y el crecimiento temprano en la mortalidad neonatal en cachorros. J Anim Sci. (2015) 93:4436–42. DOI: 10.2527/jas.2015-8971
Resumen de PubMed | Texto completo de CrossRef | Google Académico
2. Mugnier A, Chastant-Maillard S, Mila H, Lyazrhi F, Guiraud F, Adib-Lesaux A, et al. Bajo y muy bajo peso al nacer en cachorros: definiciones, factores de riesgo y supervivencia en una población a gran escala. BMC Vet Res. (2020) 16:354. DOI: 10.1186/S12917-020-02577-Z
Resumen de PubMed | Texto completo de CrossRef | Google Académico
3. Mugnier A, Mila H, Guiraud F, Brévaux J, Lecarpentier M, Martinez C, et al. El peso al nacer como factor de riesgo para la mortalidad neonatal: enfoque específico de la raza para identificar cachorros en riesgo. Prev Vet Med. (2019) 171:104746. doi: 10.1016/j.prevetmed.2019.104746
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Leclerc L, Thorin C, Flanagan J, Biourge V, Serisier S, Nguyen P. La tasa de crecimiento neonatal más alta y la puntuación de condición corporal a los 7 meses son factores predictivos de obesidad en perras Beagle adultas. BMC Vet Res. (2017) 13:104. DOI: 10.1186/S12917-017-0994-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
5. Mugnier A, Morin A, Cellard F, Devaux L, Delmas M, Adib-Lesaux A, et al. Asociación entre el peso al nacer y el riesgo de sobrepeso en la edad adulta en perros Labrador. PLoS ONE. (2020) 15:E0243820. doi: 10.1371/journal.pone.0243820
Resumen de PubMed | Texto completo de CrossRef | Google Académico
6. Linnér A, Almgren M. Programación epigenética: los primeros 1000 días importantes. Acta Paediatr. (2020) 109:443–52. doi: 10.1111/apa.15050
Resumen de PubMed | Texto completo de CrossRef | Google Académico
7. Bianco-Miotto T, Craig JM, Gasser YP, van Dijk SJ, Ozanne SE. Epigenética y DOHaD: desde lo básico hasta el nacimiento y más allá. J Dev Orig Health Dis. (2017) 8:513–9. doi: 10.1017/S2040174417000733
Resumen de PubMed | Texto completo de CrossRef | Google Académico
8. Chavatte-Palmer P, Tarrade A, Rousseau-Ralliard D. Dieta antes y durante el embarazo y la salud de la descendencia: la importancia de los modelos animales y lo que se puede aprender de ellos. Int J Medio Ambiente Salud Pública. (2016) 13:586. DOI: 10.3390/IJERPH13060586
Resumen de PubMed | Texto completo de CrossRef | Google Académico
9. Triantaphyllopoulos KA, Ikonomopoulos I, Bannister AJ. Epigenética y herencia de la variación fenotípica en el ganado. Cromatina epigenética. (2016) 9:31. DOI: 10.1186/S13072-016-0081-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
10. German AJ, Woods GRT, Holden SL, Brennan L, Burke C. Dangerous trends in pet obesity. Veterinario Rec. (2018) 182:25. DOI: 10.1136/vr.k2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Salonen M, Sulkama S, Mikkola S, Puurunen J, Hakanen E, Tiira K, et al. Prevalencia, comorbilidad y diferencias de raza en la ansiedad canina en 13.700 perros finlandeses. (2020) 10:2962. DOI: 10.1038/S41598-020-59837-Z
Resumen de PubMed | Texto completo de CrossRef | Google Académico
12. Dandrieux JRS, Mansfield CS. Enteropatía crónica en caninos: prevalencia, impacto y estrategias de manejo. Veterinario Med (Auckl). (2019) 10:203–14. doi: 10.2147/VMRR. S162774
Resumen de PubMed | Texto completo de CrossRef | Google Académico
13. Eagan BH, Gordon E, Protopopova A. Reasons for guardian-redonquishment of dogs to shelters: animal and regional predictors in British Columbia, Canada. Front Vet Sci. (2022) 9:857634. DOI: 10.3389/FVETS.2022.857634
Resumen de PubMed | Texto completo de CrossRef | Google Académico
14. Salman MD, Hutchison J, Ruch-Gallie R, Kogan L, New JC Jr, Kass PH, et al. Razones de comportamiento para la entrega de perros y gatos a 12 refugios. J Appl Anim Welf Sci. (2000) 3:93–106. doi: 10.1207/S15327604JAWS0302_2
15. Serpell J, Duffy DL, Jagoe A. Convertirse en un perro: experiencia temprana y el desarrollo del comportamiento. En:Serpell J, , editor. El perro doméstico: su comportamiento evolutivo e interacciones con las personas. 2ª edición. Cambridge: Cambridge University Press (2016). págs. 94–117. doi: 10.1017/9781139161800.006
Resumen de PubMed | Texto completo de CrossRef | Google Académico
16. Gee NR, Rodríguez KE, Fine AH, Trammell JP. Perros que apoyan la salud y el bienestar humano: un enfoque biopsicosocial. Front Vet Sci. (2021) 8:630465. DOI: 10.3389/FVETS.2021.630465
Resumen de PubMed | Texto completo de CrossRef | Google Académico
17. Qureshi AI, Memon MZ, Vazquez G, Suri MF. La propiedad de gatos y el riesgo de enfermedades cardiovasculares fatales. Resultados del segundo estudio nacional de seguimiento de mortalidad del estudio de examen de salud y nutrición. J Vasc Interv Neurol. (2009) 2:132–5.
18. FEDIAF. Facts and Figures 2021 European Overview (2022). Disponible en línea en: https://europeanpetfood.org/about/annual-report/ (consultado el 9 de mayo de 2022).
19. AVMA. Libro de consulta sobre propiedad de mascotas y demografía 2022. Washington, DC: Asociación Americana de Medicina Veterinaria (2022).
20. La Asociación Americana de Productos para Mascotas. Tamaño del mercado de la industria de mascotas, tendencias y estadísticas de propiedad 2021 (2022). Disponible en línea en: https://www.americanpetproducts.org/press_industrytrends.asp (consultado el 11 de julio de 2022).
21. Federación, Cynologique International. Presentación de nuestra organización. Disponible en línea en: http://www.fci.be/en/Presentation-of-our-organisation-4.html (consultado el 4 de agosto de 2021).
22. Grandjean D, Haymann F, Andre C, Bacque H, Bedossa T, Boogaets C, et al. The Dog Encyclopaedia. Tercera ed. Francia: Royal Canin SAS (2021).p. 930.
23. Hawthorne AJ, Booles D, Nugent PA, Gettinby G, Wilkinson J. Cambios en el peso corporal durante el crecimiento en cachorros de diferentes razas. J Nutr. (2004) 134:2027s−30s. doi: 10.1093/jn/134.8.2027S
Resumen de PubMed | Texto completo de CrossRef | Google Académico
24. The Cat Fanciers’ Association. CFA Breeds. Available online at: https://cfa.org/breeds/ (accessed August 4, 2021).
25. Caney SA, Fontbonne A, Deputte BL, German AJ, Dethioux F, German AJ, et al. Royal Canin. The Cat Encyclopedia. Second ed. Havasupai, Tewa: Aniwa Publishing (2017). p. 496.
26. Pieri N, Souza AF, Casals JB, Roballo K, Ambrósio CE, Martins DS. Comparative development of embryonic age by organogenesis in domestic dogs and cats. Reprod Domest Anim. (2015) 50:625–31. doi: 10.1111/rda.12539
27. Binder C, Aurich C, Reifinger M, Aurich J. Spontaneous ovulation in cats-uterine findings and correlations with animal weight and age. Anim Reprod Sci. (2019) 209:106167. doi: 10.1016/j.anireprosci.2019.106167
28. Reynaud K, Saint-Dizier M, Fontbonne A, Thoumire S, Chastant-Maillard S. Follicle growth, oocyte maturation, embryo development, and reproductive biotechnologies in dog and cat. Clin Theriogenology. (2020) 12:189–202.
29. Reynaud K, Fontbonne A, Marseloo N, Thoumire S, Chebrout M, de Lesegno CV, et al. In vivo meiotic resumption, fertilization and early embryonic development in the bitch. Reproduction. (2005) 130:193–201. doi: 10.1530/rep.1.00500
31. German AJ. The growing problem of obesity in dogs and cats. J Nutr. (2006) 136:1940s−6s. doi: 10.1093/jn/136.7.1940S
32. Ward E, German AJ, Churchhill JA. Asociación para la Prevención de la Obesidad en las Mascotas. Declaración de posición de la Iniciativa Global contra la Obesidad en las Mascotas (2019). Disponible en línea en: https://petobesityprevention.org/about/#GPOI (consultado el 9 de mayo de 2022).
33. Verbrugghe A. Epidemiología de la obesidad de los animales pequeños. En:Cline MG, Murphy M, , editores. Obesidad en el perro y el gato. Boca Raton, FL: CRC Press (2019). págs. 1–13. doi: 10.1201/9781315151625-1
34. Bjornvad CR, Nielsen DH, Armstrong PJ, McEvoy F, Hoelmkjaer KM, Jensen KS, et al. Evaluación de un sistema de puntuación de condición corporal de nueve puntos en gatos domésticos físicamente inactivos. Am J Vet Res. (2011) 72:433–7. doi: 10.2460/ajvr.72.4.433
Resumen de PubMed | Texto completo de CrossRef | Google Académico
35. Laflamme D. Desarrollo y validación de un sistema de puntuación de condición corporal para gatos: una herramienta clínica. Practicante Felino. (1997) 25:13–8.
36. Asociación para la Prevención de la Obesidad en las Mascotas. Resultados de la Encuesta de Obesidad de Mascotas 2018 (2019). Disponible en: https://petobesityprevention.org/2018 (consultado el 19 de octubre de 2021).
37. Tropf M, Nelson OL, Lee PM, Weng HY. Variables cardíacas y metabólicas en perros obesos. J Vet Pasante Med. (2017) 31:1000–7. DOI: 10.1111/JVIM.14775
Resumen de PubMed | Texto completo de CrossRef | Google Académico
38. Bach JF, Rozanski EA, Bedenice D, Chan DL, Freeman LM, Lofgren JL, et al. Asociación de la disfunción espiratoria de las vías respiratorias con una marcada obesidad en perros adultos sanos. Am J Vet Res. (2007) 68:670–5. doi: 10.2460/ajvr.68.6.670
Resumen de PubMed | Texto completo de CrossRef | Google Académico
39. Manens J, Ricci R, Damoiseaux C, Gault S, Contiero B, Diez M, et al. Efecto de la pérdida de peso corporal sobre la función cardiopulmonar evaluado por la prueba de caminata de 6 minutos y el análisis de gases en sangre arterial en perros obesos. J Vet Pasante Med. (2014) 28:371–8. doi: 10.1111/jvim.12260
Resumen de PubMed | Texto completo de CrossRef | Google Académico
40. Henegar JR, Bigler SA, Henegar LK, Tyagi SC, Hall JE. Cambios funcionales y estructurales en el riñón en las primeras etapas de la obesidad. J Am Soc Nefrol. (2001) 12:1211–7. doi: 10.1681/ASN. V1261211
Resumen de PubMed | Texto completo de CrossRef | Google Académico
41. Tvarijonaviciute A, Ceron JJ, Holden SL, Biourge V, Morris PJ, German AJ. Efecto de la pérdida de peso en perros obesos sobre los indicadores de función renal o enfermedad. J Vet Pasante Med. (2013) 27:31–8. DOI: 10.1111/JVIM.12029
Resumen de PubMed | Texto completo de CrossRef | Google Académico
42. German AJ, Hervera M, Hunter L, Holden SL, Morris PJ, Biourge V, et al. Mejora de la resistencia a la insulina y reducción de las adipocinas inflamatorias plasmáticas después de la pérdida de peso en perros obesos. Domest Anim Endocrinol. (2009) 37:214–26. doi: 10.1016/j.domaniend.2009.07.001
Resumen de PubMed | Texto completo de CrossRef | Google Académico
43. Tvarijonaviciute A, Ceron JJ, Holden SL, Cuthbertson DJ, Biourge V, Morris PJ, et al. Disfunción metabólica relacionada con la obesidad en perros: una comparación con el síndrome metabólico humano. BMC Vet Res. (2012) 8:147. DOI: 10.1186/1746-6148-8-147
Resumen de PubMed | Texto completo de CrossRef | Google Académico
44. Salt C, Morris PJ, Wilson D, Lund EM, German AJ. Asociación entre la esperanza de vida y la condición corporal en perros castrados propiedad del cliente. J Vet Pasante Med. (2019) 33:89–99. DOI: 10.1111/JVIM.15367
Resumen de PubMed | Texto completo de CrossRef | Google Académico
45. German AJ, Holden SL, Wiseman-Orr ML, Reid J, Nolan AM, Biourge V, et al. La calidad de vida se reduce en perros obesos, pero mejora después de una pérdida de peso exitosa. Veterinario J. (2012) 192:428–34. doi: 10.1016/j.tvjl.2011.09.015
Resumen de PubMed | Texto completo de CrossRef | Google Académico
46. Lund EM, Armstrong PJ, Kirk CA, Klausner JS. Prevalencia y factores de riesgo de obesidad en perros adultos de prácticas veterinarias privadas de los Estados Unidos. Pasante J Appl Res Vet Med. (2006) 4:177.
47. Pérez Alenza MD, Peña L, del Castillo N, Nieto AI. Factores que influyen en la incidencia y el pronóstico de los tumores mamarios caninos. J Small Anim Pract. (2000) 41:287–91. doi: 10.1111/j.1748-5827.2000.tb03203.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
48. Brown DC, Conzemius M, Shofer F. El peso corporal como factor predisponente para las fracturas condilares humerales, la ruptura cruzada craneal y la enfermedad del disco intervertebral en Cocker Spaniels. Vet Comp Orthop Traumatol. (1996) 9:75–8. DOI: 10.1055/S-0038-1632506
49. van Hagen MA, Ducro BJ, van den Broek J, Knol BW. Estimaciones de incidencia, factores de riesgo y heredabilidad de la cojera de las extremidades posteriores causada por la displasia de cadera en una cohorte de boxeadores al nacer. Am J Vet Res. (2005) 66:307–12. doi: 10.2460/ajvr.2005.66.307
Resumen de PubMed | Texto completo de CrossRef | Google Académico
50. Gates MC, Zito S, Harvey LC, Dale A, Walker JK. Evaluación de la obesidad en perros y gatos adultos que se presentan a citas de vacunación de rutina en la isla norte de Nueva Zelanda utilizando datos de registros médicos electrónicos. N Z Veterinario J. (2019) 67:126–33. doi: 10.1080/00480169.2019.1585990
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Parker VJ, Orcutt E, Love L. Fisiopatología de la obesidad: comorbilidades y consideraciones anestésicas. En:Cline MG, Murphy M, , editores. Obesidad en el perro y el gato. Primera ed. Boca Raton: CRC Press (2019). págs. 40–61. doi: 10.1201/9781315151625-3
Resumen de PubMed | Texto completo de CrossRef | Google Académico
52. Chiang CF, Villaverde C, Chang WC, Fascetti AJ, Larsen JA. Prevalencia, factores de riesgo y asociaciones de enfermedades de sobrepeso y obesidad en gatos que visitaron el hospital de enseñanza médica veterinaria de la Universidad de California, Davis desde enero de 2006 hasta diciembre de 2015. Top Companion Anim Med. (2022) 47:100620. doi: 10.1016/j.tcam.2021.100620
Resumen de PubMed | Texto completo de CrossRef | Google Académico
53. Teng KT, McGreevy PD, Toribio J, Raubenheimer D, Kendall K, Dhand NK. Asociaciones de la puntuación de la condición corporal con condiciones de salud relacionadas con el sobrepeso y la obesidad en gatos. J Small Anim Pract. (2018) 59:603–15. doi: 10.1111/jsap.12905
Resumen de PubMed | Texto completo de CrossRef | Google Académico
54. Teng KT, McGreevy PD, Toribio JL, Raubenheimer D, Kendall K, Dhand NK. Fuertes asociaciones de puntuación de condición corporal de nueve puntos con la supervivencia y la esperanza de vida en gatos. J Felino Med Surg. (2018) 20:1110–8. doi: 10.1177/1098612X17752198
Resumen de PubMed | Texto completo de CrossRef | Google Académico
55. Flanagan J, Bissot T, Hours MA, Moreno B, German AJ. Un estudio de cohorte multicéntrico internacional de pérdida de peso en gatos con sobrepeso: diferencias en el resultado en diferentes ubicaciones geográficas. PLoS ONE. (2018) 13:E0200414. doi: 10.1371/journal.pone.0200414
Resumen de PubMed | Texto completo de CrossRef | Google Académico
56. Feldhahn J, Rand J, Martin G. Sensibilidad a la insulina en gatos normales y diabéticos. J Felino Med Surg. (1999) 1:107–15. doi: 10.1016/S1098-612X(99)90067-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
57. Lund EM, Armstrong PJ, Kirk CA, Klausmer JS. Prevalencia y factores de riesgo de obesidad en gatos adultos de prácticas veterinarias privadas de los Estados Unidos. Pasante J Appl Res Vet Med. (2005) 3:88–96.
58. Valtolina C, Favier RP. Lipidosis hepática felina. Veterinario Clin North am Small Anim Pract. (2017) 47:683–702. doi: 10.1016/j.cvsm.2016.11.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
59. Salt C, Morris PJ, Butterwick RF, Lund EM, Cole TJ, German AJ. Comparación de patrones de crecimiento en perros sanos y perros en condición corporal anormal utilizando estándares de crecimiento. PLoS ONE. (2020) 15:E0238521. doi: 10.1371/journal.pone.0238521
Resumen de PubMed | Texto completo de CrossRef | Google Académico
60. Bjørnvad CR, Gloor S, Johansen SS, Sandøe P, Lund TB. La castración aumenta el riesgo de obesidad en perros machos, pero no en perras, un estudio transversal de los factores de riesgo relacionados con perros y dueños para la obesidad en perros de compañía daneses. Prev Vet Med. (2019) 170:104730. doi: 10.1016/j.prevetmed.2019.104730
Resumen de PubMed | Texto completo de CrossRef | Google Académico
61. Colliard L, Ancel J, Benet JJ, Paragon BM, Blanchard G. Risk factors for obesity in dogs in France. J Nutr. (2006) 136:1951s−4s. doi: 10.1093/jn/136.7.1951S
Resumen de PubMed | Texto completo de CrossRef | Google Académico
62. German AJ, Blackwell E, Evans M, Westgarth C. Overweight dogs exercise less frequently and for short periods: results of a large online survey of dog owners from the UK. J Nutr Sci. (2017) 6:e11. doi: 10.1017/jns.2017.6
Resumen de PubMed | Texto completo de CrossRef | Google Académico
63. McGreevy PD, Thomson PC, Pride C, Fawcett A, Grassi T, Jones B. Prevalencia de obesidad en perros examinados por las prácticas veterinarias australianas y los factores de riesgo involucrados. Veterinario Rec. (2005) 156:695–702. DOI: 10.1136/VR.156.22.695
Resumen de PubMed | Texto completo de CrossRef | Google Académico
64. Perry LM, Shmalberg J, Tanprasertsuk J, Massey D, Honaker RW, Jha AR. Factores de riesgo asociados con el sobrepeso y la obesidad canina en una encuesta informada por el propietario. bioRxiv. (2020). doi: 10.1101/2020.01.06.896399
65. Vendramini THA, Amaral AR, Pedrinelli V, Zafalon RVA, Rodrigues RBA, Brunetto MA. Castración en perros y gatos: evidencia científica actual e importancia de un manejo nutricional adecuado. (2020) 33:134–44. doi: 10.1017/S0954422419000271
Resumen de PubMed | Texto completo de CrossRef | Google Académico
66. Simpson M, Albright S, Wolfe B, Searfoss E, Street K, Diehl K, et al. Edad en la gonadectomía y riesgo de sobrepeso/obesidad y lesión ortopédica en una cohorte de Golden Retrievers. PLoS ONE. (2019) 14:E0209131. doi: 10.1371/journal.pone.0209131
Resumen de PubMed | Texto completo de CrossRef | Google Académico
67. Lefebvre SL, Yang M, Wang M, Elliott DA, Buff PR, Lund EM. Efecto de la edad en la gonadectomía sobre la probabilidad de que los perros tengan sobrepeso. J Am Vet Med Assoc. (2013) 243:236–43. doi: 10.2460/javma.243.2.236
Resumen de PubMed | Texto completo de CrossRef | Google Académico
68. Salt C, Morris PJ, German AJ, Wilson D, Lund EM, Cole TJ, et al. Tablas estándar de crecimiento para monitorear el peso corporal en perros de diferentes tamaños. PLoS ONE. (2017) 12:E0182064. doi: 10.1371/journal.pone.0182064
Resumen de PubMed | Texto completo de CrossRef | Google Académico
69. Bermingham EN, Thomas DG, Cave NJ, Morris PJ, Butterwick RF, German AJ. Requerimientos energéticos de perros adultos: un meta-análisis. PLoS ONE. (2014) 9:E109681. doi: 10.1371/journal.pone.0109681
Resumen de PubMed | Texto completo de CrossRef | Google Académico
70. Hoenig M, Ferguson DC. Efectos de la castración sobre las concentraciones hormonales y los requerimientos energéticos en gatos machos y hembras. Am J Vet Res. (2002) 63:634–9. doi: 10.2460/ajvr.2002.63.634
Resumen de PubMed | Texto completo de CrossRef | Google Académico
71. Kutzler MA. Posible relación entre los efectos adversos para la salud a largo plazo de la esterilización quirúrgica de eliminación de gónadas y la hormona luteinizante en perros. Animales (Basilea). (2020) 10:599. doi: 10.3390/ani10040599
Resumen de PubMed | Texto completo de CrossRef | Google Académico
72. Mugnier A, Cellard F, Morin A, Guiraud F, Mariani C, Adib-Lesaux A, et al. Impacto de factores neonatales y adultos en la condición corporal de los perros Labrador. Reprod Domest Anim. (2019) 54:36–7.
73. Dewey KG, Güngör D, Donovan SM, Madan EM, Venkatramanan S, Davis TA, et al. Lactancia materna y riesgo de sobrepeso en la infancia y más allá: una revisión sistemática con énfasis en el par de hermanos y estudios de intervención. Am J Clin Nutr. (2021) 114:1774–90. doi: 10.1093/ajcn/nqab206
Resumen de PubMed | Texto completo de CrossRef | Google Académico
74. Palou M, Picó C, Palou A. Leptin as a breast milk component for the prevention of obesity. (2018) 76:875–92. doi: 10.1093/nutrit/nuy046
Resumen de PubMed | Texto completo de CrossRef | Google Académico
75. van Lent D, Vernooij JCM, Corbee RJ. Los gatitos que amamantan 7 semanas o más tienen menos probabilidades de convertirse en gatos adultos con sobrepeso. Animales. (2021) 11:3434. DOI: 10.3390/ANI11123434
Resumen de PubMed | Texto completo de CrossRef | Google Académico
76. Serisier S, Feugier A, Venet C, Biourge V, German AJ. Tasa de crecimiento más rápida en gatos alimentados ad libitum: un factor de riesgo que predice la probabilidad de tener sobrepeso durante la edad adulta. J Nutr Sci. (2013) 2:e11. doi: 10.1017/jns.2013.10
Resumen de PubMed | Texto completo de CrossRef | Google Académico
77. Cave NJ, Bridges JP, Weidgraaf K, Thomas DG. Modelos mixtos no lineales de curvas de crecimiento de gatos domésticos de pelo corto en una colonia de cría, alojados en una instalación estacional para predecir la obesidad. J Anim Physiol Anim Nutr (Berl). (2018) 102:1390–400. doi: 10.1111/jpn.12930
Resumen de PubMed | Texto completo de CrossRef | Google Académico
78. Opsomer H, Liesegang A, Brugger D, Wichert B. Growth curves and body condition of young cats and their relation to maternal body condition. Animales. (2022) 12:1373. doi: 10.3390/ani12111373
Resumen de PubMed | Texto completo de CrossRef | Google Académico
79. Fettman MJ, Stanton CA, Banks LL, Hamar DW, Johnson DE, Hegstad RL, et al. Efectos de la castración sobre el peso corporal, la tasa metabólica y la tolerancia a la glucosa de gatos domésticos. Res Vet Sci. (1997) 62:131–6. doi: 10.1016/S0034-5288(97)90134-X
Resumen de PubMed | Texto completo de CrossRef | Google Académico
80. Wall M, Cave NJ, Vallee E. Owner and cat-related risk factors for feline overweight or obesity. Front Vet Sci. (2019) 6:266. DOI: 10.3389/fvets.2019.00266
Resumen de PubMed | Texto completo de CrossRef | Google Académico
81. Alexander LG, Salt C, Thomas G, Butterwick R. Efectos de la castración en la ingesta de alimentos, el peso corporal y la composición corporal en gatitas hembras en crecimiento. H. J Nutr. (2011) 106 Suppl 1:S19–23. doi: 10.1017/S0007114511001851
Resumen de PubMed | Texto completo de CrossRef | Google Académico
82. Kanchuk ML, Backus RC, Calvert CC, Morris JG, Rogers QR. El aumento de peso en gatos domésticos machos normales gonadectomizados y deficientes en lipoproteínas lipasa es el resultado de una mayor ingesta de alimentos y no de una disminución del gasto energético. J Nutr. (2003) 133:1866–74. doi: 10.1093/jn/133.6.1866
Resumen de PubMed | Texto completo de CrossRef | Google Académico
83. Wei A, Fascetti AJ, Kim K, Lee A, Graham JL, Havel PJ, et al. Efectos tempranos de la castración sobre el gasto energético en gatos machos adultos. PLoS ONE. (2014) 9:E89557. doi: 10.1371/journal.pone.0089557
Resumen de PubMed | Texto completo de CrossRef | Google Académico
84. Bermingham EN, Thomas DG, Morris PJ, Hawthorne AJ. Requerimientos energéticos de los gatos adultos. H. J Nutr. (2010) 103:1083–93. doi: 10.1017/S000711450999290X
Resumen de PubMed | Texto completo de CrossRef | Google Académico
85. Allaway D, Gilham M, Colyer A, Morris PJ. El impacto del tiempo de castración en el aumento de peso y la ingesta de energía en gatitas. J Nutr Sci. (2017) 6:e19. DOI: 10.1017/JNS.2017.20
Resumen de PubMed | Texto completo de CrossRef | Google Académico
86. de Godoy MRC and Pancosma Comparative Gut Physiology Symposium. Todo sobre la regulación del apetito: efectos de la dieta y los esteroides gonadales sobre la regulación del apetito y la ingesta de alimentos de los animales de compañía. J Anim Sci. (2018) 96:3526–36. doi: 10.1093/jas/sky146
Resumen de PubMed | Texto completo de CrossRef | Google Académico
87. Marousez L, Lesage J, Eberlé D. Epigenetics: linking early postnatal nutrition to obesity programming? Nutrientes. (2019) 11:2966. doi: 10.3390/nu11122966
Resumen de PubMed | Texto completo de CrossRef | Google Académico
88. Moullé VS, Parnet P. Efectos de la ingesta de nutrientes durante el embarazo y la lactancia en el páncreas endocrino de la descendencia. Nutrientes. (2019) 11:2708. doi: 10.3390/nu11112708
Resumen de PubMed | Texto completo de CrossRef | Google Académico
89. van Dijk SJ, Tellam RL, Morrison JL, Muhlhausler BS, Molloy PL. Desarrollos recientes sobre el papel de la epigenética en la obesidad y la enfermedad metabólica. Clin Epigenética. (2015) 7:66. doi: 10.1186/s13148-015-0101-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
90. Picó C, Reis F, Egas C, Mathias P, Matafome P. Lactation as a programming window for metabolic syndrome. Eur J Clin Invest. (2021) 51:E13482. DOI: 10.1111/ECI.13482
Resumen de PubMed | Texto completo de CrossRef | Google Académico
91. Ouni M, Schürmann A. Contribución epigenética a la obesidad. Genoma de mamíferos. (2020) 31:134–45. DOI: 10.1007/S00335-020-09835-3
Resumen de PubMed | Texto completo de CrossRef | Google Académico
92. Hoffman DJ, Powell TL, Barrett ES, Hardy DB. Orígenes del desarrollo de las enfermedades metabólicas. (2021) 101:739–95. doi: 10.1152/physrev.00002.2020
Resumen de PubMed | Texto completo de CrossRef | Google Académico
93. Hoffman DJ, Reynolds RM, Hardy DB. Orígenes del desarrollo de la salud y la enfermedad: conocimiento actual y mecanismos potenciales. (2017) 75:951–70. doi: 10.1093/nutrit/nux053
Resumen de PubMed | Texto completo de CrossRef | Google Académico
94. Kaspar D, Hastreiter S, Irmler M, Hrabé de Angelis M, Beckers J. Nutrition and its role in epigenetic inheritance of obesity and diabetes across generations. Genoma Mamm. (2020) 31:119–33. DOI: 10.1007/S00335-020-09839-Z
Resumen de PubMed | Texto completo de CrossRef | Google Académico
95. Obri A, Serra D, Herrero L, Mera P. El papel de la epigenética en el desarrollo de la obesidad. Biochem Pharmacol. (2020) 177:113973. doi: 10.1016/j.bcp.2020.113973
Resumen de PubMed | Texto completo de CrossRef | Google Académico
96. Bouret SG, Draper SJ, Simerly RB. Acción trófica de la leptina sobre las neuronas hipotalámicas que regulan la alimentación. Ciencia. (2004) 304:108–10. doi: 10.1126/science.1095004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
97. Bretón C. El eje hipotálamo-adiposo es un objetivo clave de la programación del desarrollo mediante la manipulación nutricional materna. J Endocrinol. (2013) 216:R19–31. doi: 10.1530/JOE-12-0157
Resumen de PubMed | Texto completo de CrossRef | Google Académico
98. Hur SSJ, Cropley JE, Suter CM. Paternal epigenetic programming: evolving metabolic disease risk. J Mol Endocrinol. (2017) 58:R159–r68. doi: 10.1530/JME-16-0236
Resumen de PubMed | Texto completo de CrossRef | Google Académico
99. Hullinger RL. Un estudio histomorfológico de los cambios de edad en la glándula suprarrenal canina. Tesis y disertaciones retrospectivas. Ames, IA: Universidad Estatal de Ciencia y Tecnología de Iowa (1966).
100. Buddington RK, Elnif J, Malo C, Donahoo JB. Actividades de las enzimas de membrana gástricas, pancreáticas e intestinales durante el desarrollo postnatal de los perros. Am J Vet Res. (2003) 64:627–34. DOI: 10.2460/AJVR.2003.64.627
Resumen de PubMed | Texto completo de CrossRef | Google Académico
101. Día MJ. Desarrollo del sistema inmunológico en el perro y el gato. J Comp Pathol. (2007) 137 Suppl 1:S10–5. doi: 10.1016/j.jcpa.2007.04.005
Resumen de PubMed | Texto completo de CrossRef | Google Académico
102. Fox MW. Visión general y crítica de etapas y períodos en el desarrollo canino. Dev Psychobiol. (1971) 4:37–54. doi: 10.1002/dev.420040104
Resumen de PubMed | Texto completo de CrossRef | Google Académico
103. Jeusette I, Daminet S, Nguyen P, Shibata H, Saito M, Honjoh T, et al. Efecto de la ovariectomía y la alimentación ad libitum sobre la composición corporal, el estado tiroideo, las concentraciones plasmáticas de grelina y leptina en perras. J Anim Physiol Anim Nutr. (2006) 90:12–8. doi: 10.1111/j.1439-0396.2005.00612.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
104. Spray CM, Widdowson EM. El efecto del crecimiento y desarrollo en la composición de los mamíferos. H. J Nutr. (1950) 4:332–53. doi: 10.1079/BJN19500058
Resumen de PubMed | Texto completo de CrossRef | Google Académico
105. Sheng HP, Huggins RA. Cambios en el agua, proteínas, sodio, potasio y cloruro en los tejidos con el crecimiento del Beagle. Crecimiento. (1975) 39:137–57.
106. Wang W, Brooks M, Gardner C, Milgram N. Efecto de la suplementación nutricional neuroactiva sobre el peso corporal y la composición en cachorros en crecimiento. J Nutr Sci. (2017) 6:e56. doi: 10.1017/jns.2017.57
Resumen de PubMed | Texto completo de CrossRef | Google Académico
107. Salt C, German AJ, Henzel KS, Butterwick RF. Tablas estándar de crecimiento para monitorear el peso corporal en gatitos domésticos de pelo corto intactos de los Estados Unidos. PLoS ONE. (2022) 17:E0277531. doi: 10.1371/journal.pone.0277531
Resumen de PubMed | Texto completo de CrossRef | Google Académico
108. Fischer MM, Kessler AM, Kieffer DA, Knotts TA, Kim K, Wei A, et al. Efectos de la obesidad, la restricción energética y la castración sobre la microbiota fecal de los gatos. H. J Nutr. (2017) 118:513–24. doi: 10.1017/S0007114517002379
Resumen de PubMed | Texto completo de CrossRef | Google Académico
109. Forster GM, Stockman J, Noyes N, Heuberger AL, Broeckling CD, Bantle CM, et al. Un estudio comparativo de la bioquímica sérica, el metaboloma y los parámetros del microbioma de perros de compañía clínicamente sanos, de peso normal, con sobrepeso y obesos. Top Companion Anim Med. (2018) 33:126–35. doi: 10.1053/j.tcam.2018.08.003
Resumen de PubMed | Texto completo de CrossRef | Google Académico
110. Handl S, German AJ, Holden SL, Dowd SE, Steiner JM, Heilmann RM, et al. Microbiota fecal en perros delgados y obesos. FEMS Microbiol Ecol. (2013) 84:332–43. doi: 10.1111/1574-6941.12067
Resumen de PubMed | Texto completo de CrossRef | Google Académico
111. Park HJ, Lee SE, Kim HB, Isaacson RE, Seo KW, Song KH. Asociación de la obesidad con leptina sérica, adiponectina y serotonina y microflora intestinal en perros Beagle. J Vet Pasante Med. (2015) 29:43–50. DOI: 10.1111/JVIM.12455
Resumen de PubMed | Texto completo de CrossRef | Google Académico
112. Kieler IN, Mølbak L, Hansen LL, Hermann-Bank ML, Bjornvad CR. Overweight and the feline gut microme—a pilot study. J Anim Physiol Anim Nutr. (2016) 100:478–84. doi: 10.1111/jpn.12409
Resumen de PubMed | Texto completo de CrossRef | Google Académico
113. Macedo HT, Rentas MF, Vendramini THA, Macegoza MV, Amaral AR, Jeremias JT, et al. La pérdida de peso en perros obesos promueve cambios importantes en el perfil de la microbiota fecal hasta el punto de parecerse a la microbiota de los perros delgados. Microbioma Anim. (2022) 4:6. DOI: 10.1186/s42523-021-00160-x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
114. Phungviwatnikul T, Lee AH, Belchik SE, Suchodolski JS, Swanson KS. La pérdida de peso y el consumo de dietas altas en proteínas y fibra afectan los perfiles de metabolitos sanguíneos, la composición corporal, la actividad física voluntaria, la microbiota fecal y los metabolitos fecales de los perros adultos. J Anim Sci. (2022) 100:skab379. doi: 10.1093/jas/skab379
Resumen de PubMed | Texto completo de CrossRef | Google Académico
115. Söder J, Wernersson S, Höglund K, Hagman R, Lindåse S, Dicksved J. Composition and short-term stability of gut microbiota in lean and spontaneously overweight healthy Labrador Retriever dogs. Acta Vet Scand. (2022) 64:8. DOI: 10.1186/S13028-022-00628-Z
Resumen de PubMed | Texto completo de CrossRef | Google Académico
116. Tal M, Weese JS, Gomez DE, Hesta M, Steiner JM, Verbrugghe A. La microbiota fecal bacteriana solo se ve mínimamente afectada por un plan estandarizado de pérdida de peso en gatos obesos. BMC Vet Res. (2020) 16:112. DOI: 10.1186/S12917-020-02318-2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
117. Bermingham EN, Kittelmann S, Young W, Kerr KR, Swanson KS, Roy NC, et al. La dieta post-destete afecta la composición microbiana fecal pero no la expresión del gen adiposo seleccionado en el gato (Felis catus). PLoS ONE. (2013) 8:E80992. doi: 10.1371/journal.pone.0080992
Resumen de PubMed | Texto completo de CrossRef | Google Académico
118. Pocheron AL, Le Dréan G, Billard H, Moyon T, Pagniez A, Heberden C, et al. La transferencia de microbiota materna programa el comportamiento alimentario de la descendencia. Microbiol frontal. (2021) 12:672224. DOI: 10.3389/fmicb.2021.672224
Resumen de PubMed | Texto completo de CrossRef | Google Académico
119. Cuevas-Sierra A, Ramos-Lopez O, Riezu-Boj JI, Milagro FI, Martinez JA. Dieta, microbiota intestinal y obesidad: vínculos con la genética y la epigenética del huésped y posibles aplicaciones. Adv Nutr. (2019) 10(suppl_1):S17–S30. doi: 10.1093/advances/nmy078
Resumen de PubMed | Texto completo de CrossRef | Google Académico
120. Krautkramer KA, Kreznar JH, Romano KA, Vivas EI, Barrett-Wilt GA, Rabaglia ME, et al. Las interacciones dieta-microbiota median la programación epigenética global en múltiples tejidos del huésped. Célula Mol. (2016) 64:982–92. doi: 10.1016/j.molcel.2016.10.025
Resumen de PubMed | Texto completo de CrossRef | Google Académico
121. Shock T, Badang L, Ferguson B, Martinez-Guryn K. La interacción entre la dieta, los microbios intestinales y la epigenética del huésped en la salud y la enfermedad. J Nutr Biochem. (2021) 95:108631. doi: 10.1016/j.jnutbio.2021.108631
Resumen de PubMed | Texto completo de CrossRef | Google Académico
122. Woo V, Alenghat T. Epigenetic regulation by gut microbiota. Microbios intestinales. (2022) 14:2022407. doi: 10.1080/19490976.2021.2022407
Resumen de PubMed | Texto completo de CrossRef | Google Académico
123. Sharma M, Li Y, Stoll ML, Tollefsbol TO. La conexión epigenética entre el microbioma intestinal en la obesidad y la diabetes. Genet frontal. (2019) 10:1329. doi: 10.3389/fgene.2019.01329
Resumen de PubMed | Texto completo de CrossRef | Google Académico
124. Tramper-Stranders G, Ambrozej D, Arcolaci A, Atanaskovic-Markovic M, Boccabella C, Bonini M, et al. Enlaces peligrosos: bacterias, terapias antimicrobianas y enfermedades alérgicas. Alergia. (2021) 76:3276–91. doi: 10.1111/all.15046
Resumen de PubMed | Texto completo de CrossRef | Google Académico
125. Loos RJF, Yeo GSH. La genética de la obesidad: del descubrimiento a la biología. Nat Rev Genet. (2022) 23:120–33. DOI: 10.1038/S41576-021-00414-Z
Resumen de PubMed | Texto completo de CrossRef | Google Académico
126. Pegram C, Raffan E, White E, Ashworth AH, Brodbelt DC, Church DB, et al. Frecuencia, predisposición de la raza y factores de riesgo demográficos para el estado de sobrepeso en perros en el Reino Unido. J Small Anim Pract. (2021) 62:521–30. DOI: 10.1111/JSAP.13325
Resumen de PubMed | Texto completo de CrossRef | Google Académico
127. Wallis N, Raffan E. La base genética de la obesidad y las enfermedades metabólicas relacionadas en humanos y animales de compañía. Genes. (2020) 11:1378. doi: 10.3390/genes11111378
Resumen de PubMed | Texto completo de CrossRef | Google Académico
128. Mankowska M, Krzeminska P, Graczyk M, Switonski M. Confirmación de que una deleción en el gen POMC está asociada con el peso corporal de los perros Labrador Retriever. Res Vet Sci. (2017) 112:116–8. doi: 10.1016/j.rvsc.2017.02.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
129. Raffan E, Dennis RJ, O’Donovan CJ, Becker JM, Scott RA, Smith SP, et al. Una deleción en el gen POMC canino se asocia con el peso y el apetito en perros Labrador Retriever propensos a la obesidad. Celda Metab. (2016) 23:893–900. doi: 10.1016/j.cmet.2016.04.012
Resumen de PubMed | Texto completo de CrossRef | Google Académico
130. Moore PF, Woo JC, Vernau W, Kosten S, Graham PS. Characterization of feline T cell receptor gamma (TCRG) variable region genes for the molecular diagnosis of feline intestinal T cell lymphoma. Veterinario Immunol Immunopathol. (2005) 106:167–78. doi: 10.1016/j.vetimm.2005.02.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
131. Marsilio S. Enteropatía crónica felina. J Small Anim Pract. (2021) 62:409–19. doi: 10.1111/jsap.13332
Resumen de PubMed | Texto completo de CrossRef | Google Académico
132. Kilian E, Suchodolski JS, Hartmann K, Mueller RS, Wess G, Unterer S. Long-term effects of canine parvovirus infection in dogs. PLoS ONE. (2018) 13:E0192198. doi: 10.1371/journal.pone.0192198
Resumen de PubMed | Texto completo de CrossRef | Google Académico
133. Sato-Takada K, Flemming AM, Voordouw MJ, Carr AP. Parvovirus enteritis y otros factores de riesgo asociados con signos gastrointestinales persistentes en perros más adelante en la vida: un estudio de cohorte retrospectivo. BMC Vet Res. (2022) 18:96. doi: 10.1186/s12917-022-03187-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
134. Hemida M, Vuori KA, Moore R, Anturaniemi J, Hielm-Björkman A. Las exposiciones modificables de la vida temprana y su asociación con el propietario informaron síntomas de enfermedad inflamatoria intestinal en perros adultos. Front Vet Sci. (2021) 8:552350. DOI: 10.3389/FVETS.2021.552350
Resumen de PubMed | Texto completo de CrossRef | Google Académico
135. Sundburg CR, Belanger JM, Bannasch DL, Famula TR, Oberbauer AM. Efectos de la gonadectomía sobre el riesgo de trastornos inmunes en el perro: un estudio retrospectivo. BMC Vet Res. (2016) 12:278. DOI: 10.1186/S12917-016-0911-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
136. Jergens AE. Enfermedad inflamatoria intestinal idiopática felina: lo que sabemos y lo que queda por desentrañar. J Felino Med Surg. (2012) 14:445–58. doi: 10.1177/1098612X12451548
Resumen de PubMed | Texto completo de CrossRef | Google Académico
137. Fofanova TY, Petrosino JF, Kellermayer R. Microbiome-epigenome interactions and the environmental origins of inflammatory bowel diseases. J Pediatr Gastroenterol Nutr. (2016) 62:208–19. doi: 10.1097/MPG.0000000000000950
Resumen de PubMed | Texto completo de CrossRef | Google Académico
138. Indrio F, Martini S, Francavilla R, Corvaglia L, Cristofori F, Mastrolia SA, et al. Asuntos epigenéticos: el vínculo entre la nutrición temprana, el microbioma y el desarrollo de la salud a largo plazo. Front-Pediatr. (2017) 5:178. DOI: 10.3389/FPED.2017.00178
Resumen de PubMed | Texto completo de CrossRef | Google Académico
139. Pilla R, Suchodolski JS. El papel del microbioma intestinal canino y el metaboloma en la salud y la enfermedad gastrointestinal. Front Vet Sci. (2019) 6:498. DOI: 10.3389/FVETS.2019.00498
Resumen de PubMed | Texto completo de CrossRef | Google Académico
140. Garrigues Q, Apper E, Chastant S, Mila H. Gut microbiota development in the growing dog: a dynamic process influence by maternal, environmental and host factors. Front Vet Sci. (2022) 9:964649. doi: 10.3389/fvets.2022.964649
Resumen de PubMed | Texto completo de CrossRef | Google Académico
141. Deusch O, O’Flynn C, Colyer A, Morris P, Allaway D, Jones PG, et al. La secuenciación profunda de escopeta basada en illumina revela efectos dietéticos sobre la estructura y función del microbioma fecal de gatitos en crecimiento. PLoS ONE. (2014) 9:E101021. doi: 10.1371/journal.pone.0101021
Resumen de PubMed | Texto completo de CrossRef | Google Académico
142. Sung C-H, Marsilio S, Chow B, Zornow KA, Slovak JE, Pilla R, et al. Un índice de disbiosis para evaluar la microbiota fecal en gatos sanos y gatos con enteropatías crónicas. J Felino Med Surg. (2022) 24:e1–12. doi: 10.1177/1098612X221077876
Resumen de PubMed | Texto completo de CrossRef | Google Académico
143. Marsilio S, Pilla R, Sarawichitr B, Chow B, Hill SL, Ackermann MR, et al. Caracterización del microbioma fecal en gatos con enfermedad inflamatoria intestinal o linfoma alimentario de células pequeñas. (2019) 9:19208. DOI: 10.1038/S41598-019-55691-W
Resumen de PubMed | Texto completo de CrossRef | Google Académico
144. Suchodolski JS, Dowd SE, Wilke V, Steiner JM, Jergens AE. La pirosecuenciación del gen 16s rRNA revela disbiosis bacteriana en el duodeno de perros con enfermedad inflamatoria intestinal idiopática. PLoS ONE. (2012) 7:E39333. doi: 10.1371/journal.pone.0039333
Resumen de PubMed | Texto completo de CrossRef | Google Académico
145. Suchodolski JS, Markel ME, Garcia-Mazcorro JF, Unterer S, Heilmann RM, Dowd SE, et al. El microbioma fecal en perros con diarrea aguda y enfermedad inflamatoria intestinal idiopática. PLoS ONE. (2012) 7:E51907. doi: 10.1371/journal.pone.0051907
Resumen de PubMed | Texto completo de CrossRef | Google Académico
146. Giaretta PR, Rech RR, Guard BC, Blake AB, Blick AK, Steiner JM, et al. Comparación de la expresión intestinal del transportador apical de ácidos biliares dependientes de sodio entre perros con y sin enteropatía inflamatoria crónica. J Vet Pasante Med. (2018) 32:1918–26. DOI: 10.1111/JVIM.15332
Resumen de PubMed | Texto completo de CrossRef | Google Académico
147. Minamoto Y, Minamoto T, Isaiah A, Sattasathuchana P, Buono A, Rangachari VR, et al. Concentraciones fecales de ácidos grasos de cadena corta y disbiosis en perros con enteropatía crónica. J Vet Pasante Med. (2019) 33:1608–18. DOI: 10.1111/JVIM.15520
Resumen de PubMed | Texto completo de CrossRef | Google Académico
148. Liu S, Zhao W, Lan P, Mou X. El microbioma en las enfermedades inflamatorias intestinales: de la patogénesis a la terapia. Célula proteica. (2021) 12:331–45. DOI: 10.1007/S13238-020-00745-3
Resumen de PubMed | Texto completo de CrossRef | Google Académico
149. DeGruttola AK, Low D, Mizoguchi A, Mizoguchi E. Current understanding of dysbiosis in disease in human and animal models. Inflamm Bowel Dis. (2016) 22:1137–50. doi: 10.1097/MIB.0000000000000750
Resumen de PubMed | Texto completo de CrossRef | Google Académico
150. Del Carro A, Corrò M, Bertero A, Colitti B, Banchi P, Bertolotti L, et al. La evolución de la flora microbiana de la basura madre desde el nacimiento hasta los 60 días de edad. BMC Vet Res. (2022) 18:95. DOI: 10.1186/S12917-022-03199-3
Resumen de PubMed | Texto completo de CrossRef | Google Académico
151. Blake AB, Cigarroa A, Klein HL, Khattab MR, Keating T, Van De Coevering P, et al. Etapas de desarrollo en microbiota, ácidos biliares y especies clostridiales en cachorros sanos. J Vet Pasante Med. (2020) 34:2345–56. DOI: 10.1111/JVIM.15928
Resumen de PubMed | Texto completo de CrossRef | Google Académico
152. Guard BC, Mila H, Steiner JM, Mariani C, Suchodolski JS, Chastant-Maillard S. Caracterización del microbioma fecal durante el desarrollo neonatal y pediátrico temprano en cachorros. PLoS ONE. (2017) 12:E0175718. doi: 10.1371/journal.pone.0175718
Resumen de PubMed | Texto completo de CrossRef | Google Académico
153. Tú yo, Kim MJ. Comparación de la microbiota intestinal de 96 perros sanos por rasgos individuales: raza, edad y puntuación de condición corporal. Animales. (2021) 11:2432. doi: 10.3390/ani11082432
Resumen de PubMed | Texto completo de CrossRef | Google Académico
154. Alessandri G, Argentini C, Milani C, Turroni F, Cristina Ossiprandi M, van Sinderen D, et al. Echar un vistazo a la comunidad intestinal bacteriana de animales de compañía: una perspectiva canina y felina. Microbio Biotechnol. (2020) 13:1708–32. doi: 10.1111/1751-7915.13656
Resumen de PubMed | Texto completo de CrossRef | Google Académico
155. Masuoka H, Shimada K, Kiyosue-Yasuda T, Kiyosue M, Oishi Y, Kimura S, et al. Transición de la microbiota intestinal de los gatos con la edad. PLoS ONE. (2017) 12:E0181739. doi: 10.1371/journal.pone.0181739
Resumen de PubMed | Texto completo de CrossRef | Google Académico
156. Deusch O, O’Flynn C, Colyer A, Swanson KS, Allaway D, Morris P, et al. El estudio longitudinal del microbioma fecal felino identifica cambios en la edad adulta temprana independientemente del desarrollo sexual. PLoS ONE. (2015) 10:E0144881. doi: 10.1371/journal.pone.0144881
Resumen de PubMed | Texto completo de CrossRef | Google Académico
157. Hooda S, Vester Boler BM, Kerr KR, Dowd SE, Swanson KS. El microbioma intestinal de los gatitos se ve afectado por la relación proteína: carbohidratos en la dieta y se asocia con las concentraciones de metabolitos y hormonas en la sangre. H. J Nutr. (2013) 109:1637–46. doi: 10.1017/S0007114512003479
Resumen de PubMed | Texto completo de CrossRef | Google Académico
158. Melandri M, Aiudi GG, Caira M, Alonge S. Un apoyo biótico durante el embarazo para fortalecer el rendimiento gastrointestinal en cachorros. Front Vet Sci. (2020) 7:417. DOI: 10.3389/FVETS.2020.00417
Resumen de PubMed | Texto completo de CrossRef | Google Académico
159. Alonge S, Aiudi GG, Lacalandra GM, Leoci R, Melandri M. Pre- y probióticos para aumentar el poder inmunológico del calostro en perros. Front Vet Sci. (2020) 7:570414. DOI: 10.3389/FVETS.2020.570414
Resumen de PubMed | Texto completo de CrossRef | Google Académico
160. Adogony V, Respondek F, Biourge V, Rudeaux F, Delaval J, Bind JL, et al. Efectos de scFOS dietético sobre inmunoglobulinas en calostros y leche de perras. J Anim Physiol Anim Nutr. (2007) 91:169–74. doi: 10.1111/j.1439-0396.2007.00688.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
161. Mark-Christensen A, Lange A, Erichsen R, Frøslev T, Esen B, Sørensen HT, et al. Exposición temprana a los antibióticos y riesgo de enfermedad de Crohn: un estudio nacional de cohorte de nacimiento danés. Inflamm Bowel Dis. (2022) 28:415–22. DOI: 10.1093/IBD/IZAB085
Resumen de PubMed | Texto completo de CrossRef | Google Académico
162. Pan WH, Sommer F, Falk-Paulsen M, Ulas T, Best P, Fazio A, et al. La exposición a la microbiota intestinal provoca distintos cambios en el metiloma y el transcriptoma en las células epiteliales intestinales durante el desarrollo postnatal. Genoma Med. (2018) 10:27. DOI: 10.1186/S13073-018-0534-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
163. Yu DH, Gadkari M, Zhou Q, Yu S, Gao N, Guan Y, et al. La regulación epigenética postnatal de las células madre intestinales requiere la metilación del ADN y está guiada por el microbioma. Genoma Biol. (2015) 16:211. DOI: 10.1186/S13059-015-0763-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
164. Guard BC, Barr JW, Reddivari L, Klemashevich C, Jayaraman A, Steiner JM, et al. Caracterización de la disbiosis microbiana y cambios metabolómicos en perros con diarrea aguda. PLoS ONE. (2015) 10:E0127259. doi: 10.1371/journal.pone.0127259
Resumen de PubMed | Texto completo de CrossRef | Google Académico
165. Pilla R, Gaschen FP, Barr JW, Olson E, Honneffer J, Guard BC, et al. Efectos del metronidazol sobre el microbioma fecal y el metaboloma en perros sanos. J Vet Pasante Med. (2020) 34:1853–66. DOI: 10.1111/JVIM.15871
Resumen de PubMed | Texto completo de CrossRef | Google Académico
166. Stavroulaki E, Suchodolski JS, Pilla R, Fosgate GT, Sung C-H, Lidbury JA, et al. Efectos a corto y largo plazo de la amoxicilina/ácido clavulánico o la doxiciclina en el microbioma gastrointestinal de gatos en crecimiento. PLoS ONE. (2021) 16:E0253031. doi: 10.1371/journal.pone.0253031
Resumen de PubMed | Texto completo de CrossRef | Google Académico
167. Schulfer AF, Battaglia T, Alvarez Y, Bijnens L, Ruiz VE, Ho M, et al. La transferencia intergeneracional de microbiota perturbada por antibióticos mejora la colitis en ratones susceptibles. Nat Microbiol. (2018) 3:234–42. DOI: 10.1038/S41564-017-0075-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
168. Agrawal M, Sabino J, Frias-Gomes C, Hillenbrand CM, Soudant C, Axelrad JE, et al. Exposiciones tempranas a la vida y el riesgo de enfermedad inflamatoria intestinal: revisión sistemática y metanálisis. EClinicalMedicine. (2021) 36:100884. doi: 10.1016/j.eclinm.2021.100884
Resumen de PubMed | Texto completo de CrossRef | Google Académico
169. Suchodolski JS. Consecuencias metabólicas de la disbiosis intestinal en perros con EII. Vancouver, BC: Cumbre de Nutrición de Animales de Compañía (CAN) de Nestlé Purina (2017).
170. Sacoor C, Barros LM, Montezinho L. ¿Cuáles son los biomarcadores potenciales que deben considerarse en el diagnóstico y manejo de las enteropatías inflamatorias crónicas caninas? Abierto Vet J. (2021) 10:412–30. doi: 10.4314/ovj.v10i4.9
Resumen de PubMed | Texto completo de CrossRef | Google Académico
171. Maeda S, Ohno K, Fujiwara-Igarashi A, Tomiyasu H, Fujino Y, Tsujimoto H. Methylation of TNFRSF13b and TNFRSF13c in duodenal mucosa in canine inflammatory bowel disease and its association with decreased mucosal IGA expression. Veterinario Immunol Immunopathol. (2014) 160:97–106. doi: 10.1016/j.vetimm.2014.04.005
Resumen de PubMed | Texto completo de CrossRef | Google Académico
172. Kathrani A, Werling D, Allenspach K. Canine breeds at high risk of developing inflammatory bowel disease in the South-Eastern UK. Veterinario Rec. (2011) 169:635. DOI: 10.1136/VR.D5380
Resumen de PubMed | Texto completo de CrossRef | Google Académico
173. Dreschel NA. Los efectos del miedo y la ansiedad en la salud y la esperanza de vida en los perros. Appl Anim Behav Sci. (2010) 125:157–62. doi: 10.1016/j.applanim.2010.04.003
174. Bray EE, Sammel MD, Cheney DL, Serpell JA, Seyfarth RM. Efectos de la inversión materna, el temperamento y la cognición en el éxito del perro guía. Proc Natl Acad Sci USA. (2017) 114:9128–33. doi: 10.1073/pnas.1704303114
Resumen de PubMed | Texto completo de CrossRef | Google Académico
175. Gross B, Garcia-Tapia D, Riedesel E, Ellinwood NM, Jens JK. Maduración normal del cerebro canino en imágenes de resonancia magnética. Ecografía Radiol Veterinaria. (2010) 51:361–73. doi: 10.1111/j.1740-8261.2010.01681.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
176. Brydges NM. Estrés prepuberal y desarrollo cerebral en roedores. Curr Opin Behav Sci. (2016) 7:8–14. doi: 10.1016/j.cobeha.2015.08.003
177. Dietz L, Arnold A-MK, Goerlich-Jansson VC, Vinke CM. La importancia de las experiencias de vida temprana para el desarrollo de trastornos del comportamiento en perros domésticos. Comportamiento. (2018) 155:83–114. doi: 10.1163/1568539X-00003486
178. Battaglia CL. Períodos de desarrollo temprano y los efectos de la estimulación y las experiencias sociales en el canino. J Vet Behav. (2009) 4:203–10. doi: 10.1016/j.jveb.2009.03.003
179. Houpt KA, Zicker S. Efectos dietéticos sobre el comportamiento canino y felino. Veterinario Clin North am Small Anim Pract. (2003) 33:405–16, VII–VIII. doi: 10.1016/S0195-5616(02)00115-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
180. Tiira K, Lohi H. Las experiencias tempranas de la vida y el ejercicio se asocian con ansiedades caninas. PLoS ONE. (2015) 10:E0141907. doi: 10.1371/journal.pone.0141907
Resumen de PubMed | Texto completo de CrossRef | Google Académico
181. Guardini G, Mariti C, Bowen J, Fatjó J, Ruzzante S, Martorell A, et al. Influencia del cuidado matutino materno en las respuestas conductuales de los cachorros de Beagle de 8 semanas de edad a nuevos estímulos ambientales y sociales. Appl Anim Behav Sci. (2016) 181:137–44. doi: 10.1016/j.applanim.2016.05.006
182. Foyer P, Wilsson E, Jensen P. Levels of maternal care in dogs affect adult offspring temperament. Sci Rep. (2016) 6:19253. doi: 10.1038/srep19253
183. McMillan FD. Resultados conductuales y psicológicos para perros vendidos como cachorros a través de tiendas de mascotas y / o nacidos en establecimientos de cría comercial: conocimiento actual y causas putativas. J Vet Behav. (2017) 19:14–26. doi: 10.1016/j.jveb.2017.01.001
184. Casey RA, Loftus B, Bolster C, Richards GJ, Blackwell EJ. Agresión dirigida humana en perros domésticos (Canis familiaris): ocurrencia en diferentes contextos y factores de riesgo. Appl Anim Behav Sci. (2014) 152:52–63. doi: 10.1016/j.applanim.2013.12.003
185. Wormald D, Lawrence AJ, Carter G, Fisher AD. Análisis de las correlaciones entre la exposición social temprana y la agresión reportada en el perro. J Vet Behav. (2016) 15:31–6. doi: 10.1016/j.jveb.2016.08.071
186. Freedman DG, King JA, Elliot O. Período crítico en el desarrollo social de los perros. Ciencia. (1961) 133:1016–7. doi: 10.1126/science.133.3457.1016
Resumen de PubMed | Texto completo de CrossRef | Google Académico
187. Lazarowski L, Krichbaum S, Waggoner LP, Katz JS. El desarrollo de habilidades de resolución de problemas en una población de perros candidatos a la detección (Canis familiaris). Anim Cogn. (2020) 23:755–68. DOI: 10.1007/S10071-020-01387-Y
Resumen de PubMed | Texto completo de CrossRef | Google Académico
188. Bray EE, Gruen ME, Gnanadesikan GE, Horschler DJ, Levy KM, Kennedy BS, et al. Desarrollo cognitivo del perro: un estudio longitudinal a lo largo de los primeros 2 años de vida. Anim Cogn. (2021) 24:311–28. DOI: 10.1007/S10071-020-01443-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
189. Serpell JA, Duffy DL. Los aspectos del entorno juvenil y adolescente predicen la agresión y el miedo en perros guía de 12 meses de edad. Front Vet Sci. (2016) 3:49. DOI: 10.3389/fvets.2016.00049
Resumen de PubMed | Texto completo de CrossRef | Google Académico
190. Cimarelli G, Virányi Z, Turcsán B, Rónai Z, Sasvári-Székely M, Bánlaki Z. El comportamiento social de los perros de compañía se asocia con la metilación periférica de OXTR. Psicópata frontal. (2017) 8:549. DOI: 10.3389/fpsyg.2017.00549
Resumen de PubMed | Texto completo de CrossRef | Google Académico
191. Simonson M. Effects of maternal malnourishment, development and behavior in successive generations in the rat and cat. En:Levitsky DA, , editor. Desnutrición, medio ambiente y comportamiento Nuevas perspectivas. Ithaca, NY: Cornell University Press (1979). págs. 133-48.
192. Gallo PV, Werboff J, Knox K. Restricción de proteínas durante la gestación y la lactancia: desarrollo del comportamiento de apego en gatos. Behav Neural Biol. (1980) 29:216–23. doi: 10.1016/S0163-1047(80)90547-6
Resumen de PubMed | Texto completo de CrossRef | Google Académico
193. Villablanca JR, Olmstead CE. Desarrollo neurológico de gatitos. Dev Psychobiol. (1979) 12:101–27. doi: 10.1002/dev.420120204
Resumen de PubMed | Texto completo de CrossRef | Google Académico
194. Turner DC. Una revisión de más de tres décadas de investigación sobre las interacciones y relaciones gato-humano y humano-gato. Procesos de comportamiento. (2017) 141(Pt 3):297–304. doi: 10.1016/j.beproc.2017.01.008
Resumen de PubMed | Texto completo de CrossRef | Google Académico
195. McCune S. El impacto de la paternidad y la socialización temprana en el desarrollo del comportamiento de los gatos hacia las personas y los objetos novedosos. Appl Anim Behav Sci. (1995) 45:109–24. doi: 10.1016/0168-1591(95)00603-P
196. Casey RA, Bradshaw JWS. Los efectos de la socialización adicional para gatitos en un centro de rescate sobre su comportamiento e idoneidad como mascota. Appl Anim Behav Sci. (2008) 114:196–205. doi: 10.1016/j.applanim.2008.01.003
197. Bellamy KK, Storengen LM, Handegård KW, Arnet EF, Prestrud KW, Overall KL, et al. DRD2 se asocia con el miedo en algunas razas de perros. J Vet Behav. (2018) 27:67–73. doi: 10.1016/j.jveb.2018.07.008
198. van Rooy D, Haase B, McGreevy PD, Thomson PC, Wade CM. Evaluación de genes candidatos OPRM1, DRD2, AVPR1a y OXTR en Golden retrievers con comportamientos relacionados con la separación. J Vet Behav. (2016) 16:22–7. doi: 10.1016/j.jveb.2016.03.001
199. van den Bergh BRH, Dahnke R, Mennes M. Prenatal stress and the developing brain: risks for neurodevelopmental disorders. Dev Psychopathol. (2018) 30:743–62. doi: 10.1017/S0954579418000342
Resumen de PubMed | Texto completo de CrossRef | Google Académico
200. Mourtzi N, Sertedaki A, Charmandari E. Señalización de glucocorticoides y alteraciones epigenéticas en trastornos relacionados con el estrés. Int J Mol Sci. (2021) 22:5964. doi: 10.3390/ijms22115964
Resumen de PubMed | Texto completo de CrossRef | Google Académico
201. van den Bergh BRH, van den Heuvel MI, Lahti M, Braeken M, de Rooij SR, Entringer S, et al. Orígenes del desarrollo prenatal del comportamiento y la salud mental: la influencia del estrés materno en el embarazo. Neurosci Biobehav Rev. (2020) 117: 26–64. doi: 10.1016/j.neubiorev.2017.07.003
Resumen de PubMed | Texto completo de CrossRef | Google Académico
202. Tyrka AR, Ridout KK, Parade SH. Adversidad infantil y regulación epigenética de los genes de señalización glucocorticoides: asociaciones en niños y adultos. Dev Psychopathol. (2016) 28(4pt2):1319–31. DOI: 10.1017/S0954579416000870
Resumen de PubMed | Texto completo de CrossRef | Google Académico
203. Krol KM, Moulder RG, Lillard TS, Grossmann T, Connelly JJ. Epigenetic dynamics in infancy and the impact of maternal engagement. Sci Adv. (2019) 5:eaay0680. doi: 10.1126/sciadv.aay0680
Resumen de PubMed | Texto completo de CrossRef | Google Académico
204. Franklin TB, Russig H, Weiss IC, Gräff J, Linder N, Michalon A, et al. Transmisión epigenética del impacto del estrés temprano a través de generaciones. Biol Psiquiatría. (2010) 68:408–15. doi: 10.1016/j.biopsych.2010.05.036
Resumen de PubMed | Texto completo de CrossRef | Google Académico
205. Jawaid A, Jehle KL, Mansuy IM. Impacto de la exposición de los padres en la salud de la descendencia en humanos. Tendencias Genet. (2021) 37:373–88. doi: 10.1016/j.tig.2020.10.006
Resumen de PubMed | Texto completo de CrossRef | Google Académico
206. Watkins AJ, Ursell E, Panton R, Papenbrock T, Hollis L, Cunningham C, et al. Las respuestas adaptativas de los embriones tempranos de ratón a la dieta materna protegen el crecimiento fetal pero predisponen a la enfermedad de inicio en adultos. Biol Reprod. (2008) 78:299–306. doi: 10.1095/biolreprod.107.064220
Resumen de PubMed | Texto completo de CrossRef | Google Académico
207. Louwies T, Johnson AC, Orock A, Yuan T, Greenwood-Van Meerveld B. El eje microbiota-intestino-cerebro: un papel emergente para el epigenoma. Exp Biol Med. (2020) 245:138–45. doi: 10.1177/1535370219891690
Resumen de PubMed | Texto completo de CrossRef | Google Académico
208. Szyf M. La epigenética del estrés perinatal. Diálogos Clin Neurosci. (2019) 21:369–78. doi: 10.31887/DCNS.2019.21.4/mszyf
209. Mohajeri MH, La Fata G, Steinert RE, Weber P. Relación entre el microbioma intestinal y la función cerebral. (2018) 76:481–96. doi: 10.1093/nutrit/nuy009
Resumen de PubMed | Texto completo de CrossRef | Google Académico
210. Zhu S, Jiang Y, Xu K, Cui M, Ye W, Zhao G, et al. El progreso de la investigación del microbioma intestinal relacionado con los trastornos cerebrales. J Neuroinflamación. (2020) 17:25. DOI: 10.1186/S12974-020-1705-Z
Resumen de PubMed | Texto completo de CrossRef | Google Académico
211. Sun C, Velazquez MA, Fleming TP. DOHaD y el período periconcepcional, una ventana crítica en el tiempo. En:Rosenfled CS, , editor. El epigenoma y los orígenes del desarrollo de la salud y la enfermedad. Nueva York, NY: Academic Press (2016). págs. 33–47. doi: 10.1016/B978-0-12-801383-0.00003-7
212. Aiken CE, Ozanne SE. Diferencias sexuales en los modelos de programación del desarrollo. Reproducción. (2013) 145:R1–13. doi: 10.1530/REP-11-0489
Resumen de PubMed | Texto completo de CrossRef | Google Académico
213. Bock J, Poeschel J, Schindler J, Börner F, Shachar-Dadon A, Ferdman N, et al. Impacto transgeneracional específico del sexo del estrés preconcepcional en el desarrollo de espinas dendríticas y longitud dendrítica en la corteza prefrontal medial. Función de estructura cerebral. (2016) 221:855–63. DOI: 10.1007/S00429-014-0940-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
214. Laguna-Barraza R, Bermejo-Álvarez P, Ramos-Ibeas P, de Frutos C, López-Cardona AP, Calle A, et al. Origen embrionario específico del sexo de la variabilidad fenotípica postnatal. Reprod Fertil Dev. (2012) 25:38–47. doi: 10.1071/RD12262
Resumen de PubMed | Texto completo de CrossRef | Google Académico
215. Tarrade A, Panchenko P, Junien C, Gabory A. Placental contribution to nutritional programming of health and diseases: epigenetics and sexual dimorphism. J Exp Biol. (2015) 218(Pt 1):50–8. doi: 10.1242/jeb.110320
Resumen de PubMed | Texto completo de CrossRef | Google Académico
216. Sinclair KD, Rutherford KM, Wallace JM, Brameld JM, Stöger R, Alberio R, et al. Epigenética y programación del desarrollo de los rasgos de bienestar y producción en animales de granja. Reprod Fertil Dev. (2016) 28:1443–78. DOI: 10.1071/RD16102
Resumen de PubMed | Texto completo de CrossRef | Google Académico
217. Torres-Rovira L, Tarrade A, Astiz S, Mourier E, Perez-Solana M, de la Cruz P, et al. Desarrollo de órganos dependientes del sexo y la raza y respuestas metabólicas en fetos de cerdos magros y obesos/resistentes a la leptina. PLoS ONE. (2013) 8:E66728. doi: 10.1371/journal.pone.0066728
Resumen de PubMed | Texto completo de CrossRef | Google Académico
218. Vonnahme KA, Hess BW, Nijland MJ, Nathanielsz PW, Ford SP. La diferenciación placentomal puede compensar la restricción de nutrientes maternos en ovejas adaptadas a condiciones de rango difíciles. J Anim Sci. (2006) 84:3451–9. DOI: 10.2527/JAS.2006-132
Resumen de PubMed | Texto completo de CrossRef | Google Académico
219. Murray JK, Kinsman RH, Lord MS, Da Costa REP, Woodward JL, Owczarczak-Garstecka SC, et al. ‘Generation pup’—protocol for a longitudinal study of dog behavior and health. BMC Vet Res. (2021) 17:1. DOI: 10.1186/S12917-020-02730-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
220. Chu ET, Simpson MJ, Diehl K, Page RL, Sams AJ, Boyko AR. La depresión endogámica causa una fecundidad reducida en los golden retrievers. Genoma Mamm. (2019) 30:166–72. doi: 10.1007/s00335-019-09805-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
221. Guy MK, Page RL, Jensen WA, Olson PN, Haworth JD, Searfoss EE, et al. El estudio de por vida del golden retriever: establecimiento de un estudio de cohorte observacional con relevancia traslacional para la salud humana. Philos Trans R Soc Lond B Biol Sci. (2015) 370:20140236. doi: 10.1098/rstb.2014.0230
Resumen de PubMed | Texto completo de CrossRef | Google Académico
222. Clements DN, Handel IG, Rose E, Querry D, Pugh CA, Ollier WE, et al. Dogslife: un estudio longitudinal basado en la web de la salud del Labrador Retriever en el Reino Unido. BMC Vet Res. (2013) 9:13. doi: 10.1186/1746-6148-9-13
Resumen de PubMed | Texto completo de CrossRef | Google Académico
223. Pugh CA, Bronsvoort BMC, Handel IG, Querry D, Rose E, Summers KM, et al. Las tasas de incidencia y los análisis de factores de riesgo para el propietario informaron vómitos y diarrea en Labrador Retrievers: hallazgos de la cohorte DogsLife. Prev Vet Med. (2017) 140:19–29. doi: 10.1016/j.prevetmed.2017.02.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
224. Creevy KE, Akey JM, Kaeberlein M, Promislow DEL. Un estudio científico abierto sobre el envejecimiento en perros de compañía. Naturaleza. (2022) 602:51–7. DOI: 10.1038/S41586-021-04282-9
Resumen de PubMed | Texto completo de CrossRef | Google Académico
225. Krontveit RI, Nødtvedt A, Sævik BK, Ropstad E, Skogmo HK, Trangerud C, et al. Estudio prospectivo sobre la displasia y el crecimiento de la cadera canina en una cohorte de cuatro razas grandes en Noruega (1998–2001). Prev Vet Med. (2010) 97:252–63. doi: 10.1016/j.prevetmed.2010.09.015
Resumen de PubMed | Texto completo de CrossRef | Google Académico
226. Sævik BK, Skancke EM, Trangerud C. Estudio longitudinal sobre diarrea y vómitos en perros jóvenes de cuatro razas grandes. Acta Vet Scand. (2012) 54:8. DOI: 10.1186/1751-0147-54-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
227. Murray JK, Casey RA, Gale E, Buffington CAT, Roberts C, Kinsman RH, et al. Perfil de cohorte: el ‘Bristol Cats Study’ (BCS), una cohorte de nacimiento de gatitos propiedad de hogares del Reino Unido. Int J Epidemiol. (2017) 46:1749–50E. DOI: 10.1093/IJE/DYX066
Resumen de PubMed | Texto completo de CrossRef | Google Académico
228. Longstaff L, Gruffydd-Jones TJ, Buffington CT, Casey RA, Murray JK. Signos del tracto urinario inferior informados por el propietario en una cohorte de gatos jóvenes. J Felino Med Surg. (2017) 19:609–18. doi: 10.1177/1098612X16643123
Resumen de PubMed | Texto completo de CrossRef | Google Académico
229. Rowe E, Browne W, Casey R, Gruffydd-Jones T, Murray J. Risk factors identified for owner-reported feline obesity at around one year of age: dry diet and indoor lifestyle. Prev Vet Med. (2015) 121:273–81. doi: 10.1016/j.prevetmed.2015.07.011
Resumen de PubMed | Texto completo de CrossRef | Google Académico
230. Rowe EC, Browne WJ, Casey RA, Gruffydd-Jones TJ, Murray JK. Factores de riesgo de la vida temprana identificados para el sobrepeso felino y la obesidad informados por el propietario alrededor de los dos años de edad. Prev Vet Med. (2017) 143:39–48. doi: 10.1016/j.prevetmed.2017.05.010
Resumen de PubMed | Texto completo de CrossRef | Google Académico
231. Cat Phenotype and Health Information Registry (Cat PHIR) (2021). Disponible en: https://vgl.ucdavis.edu/cat-phir (consultado el 22 de octubre de 2021).
232. Rauluseviciute I, Drabløs F, Rye MB. Datos de metilación del ADN por secuenciación: enfoques experimentales y recomendaciones para herramientas y tuberías para el análisis de datos. Clin Epigenética. (2019) 11:193. DOI: 10.1186/S13148-019-0795-X
Resumen de PubMed | Texto completo de CrossRef | Google Académico
233. Yong WS, Hsu FM, Chen PY. Perfilar la metilación del ADN de todo el genoma. Cromatina epigenética. (2016) 9:26. DOI: 10.1186/S13072-016-0075-3
Resumen de PubMed | Texto completo de CrossRef | Google Académico
234. Goyal D, Limesand SW, Goyal R. Epigenetic responses and the developmental origins of health and disease. J Endocrinol. (2019) 242:T105–T19. doi: 10.1530/JOE-19-0009
Resumen de PubMed | Texto completo de CrossRef | Google Académico
235. Megquier K, Genereux DP, Hekman J, Swofford R, Turner-Maier J, Johnson J, et al. Barkbase: anotación epigenómica de genomas caninos. Genes. (2019) 10:433. doi: 10.3390/genes10060433
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Palabras clave: comportamiento, epigenética, microbiota, nutrición, obesidad, enteropatía crónica, programación del desarrollo
Cita: Gaillard V, Chastant S, England G, Forman O, German AJ, Suchodolski JS, Villaverde C, Chavatte-Palmer P y Péron F (2022) Factores de riesgo ambientales en cachorros y gatitos para desarrollar trastornos crónicos en la edad adulta: un llamado a la investigación sobre la programación del desarrollo. Frente. Vet. Sci. 9:944821. DOI: 10.3389/FVETS.2022.944821
Recibido: 15 de mayo de 2022; Aprobado: 09 Diciembre 2022;
Publicado: 23 diciembre 2022.
Editado por:
Regiane R. Santos, Schothorst Feed Research, Países Bajos
Revisado por:
Ada Rota, Universidad de Pisa, Italia
Adam John Watkins, Universidad de Nottingham, Reino Unido
Sarah A. Reed, Universidad de Connecticut, Estados Unidos
Copyright © 2022 Gaillard, Chastant, Inglaterra, Forman, alemán, Suchodolski, Villaverde, Chavatte-Palmer y Péron. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Virginie Gaillard,  virginie.gaillard@royalcanin.com
virginie.gaillard@royalcanin.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía