
Impacto de la dieta alta en espirulina, extruida o suplementada con enzimas en pollos

Impacto de la dieta alta en espirulina, extruida o suplementada con enzimas, sobre las células sanguíneas, los metabolitos sistémicos y los perfiles hepáticos de lípidos y minerales de pollos de engorde
 María P. Spínola1 ,2 †
María P. Spínola1 ,2 †  Cristina M. Alfaia1 ,2 †
Cristina M. Alfaia1 ,2 †  Mónica M. Costa1 ,2 †
Mónica M. Costa1 ,2 †  Rui M. A. Pinto3 ,4
Rui M. A. Pinto3 ,4  Paula A. Lopes1 ,2
Paula A. Lopes1 ,2  José M. Pestana1 ,2 João C. Tavares5 Ana R. Mendes1 ,2 ,5 ,6
José M. Pestana1 ,2 João C. Tavares5 Ana R. Mendes1 ,2 ,5 ,6  Miguel P. Mourato5 ,6
Miguel P. Mourato5 ,6  Beatriz Tavares5 Daniela F. P. Carvalho5 ,6 Cátia F. Martins1 ,2 ,5 ,6 Joana I. Ferreira1 ,2 Madalena M. Lordelo5 ,6
Beatriz Tavares5 Daniela F. P. Carvalho5 ,6 Cátia F. Martins1 ,2 ,5 ,6 Joana I. Ferreira1 ,2 Madalena M. Lordelo5 ,6  José A. M. Pates1 ,2 *
José A. M. Pates1 ,2 *- 1CIISA – Centro de Investigação Interdisciplinar em Sanidade Animal, Faculdade de Medicina Veterinária, Universidade de Lisboa, Lisbon, Portugal
- 2Laboratório Associado para Ciência Animal e Veterinária (AL4AnimalS), Lisboa, Portugal
- 3JCS, Laboratório de Análises Clínicas Dr. Joaquim Chaves, Avenida General Norton de Matos, Algés, Portugal
- 4iMED.UL, Faculdade de Farmácia, Universidade de Lisboa, Avenida Professor Gama Pinto, Lisboa, Portugal
- 5Instituto Superior de Agronomía, Universidade de Lisboa, Lisboa, Portugal
- 6LEAF – Vinculando Paisaje, Medio Ambiente, Agricultura y Alimentación, Instituto Superior de Agronomía, Universidade de Lisboa, Laboratorio Asociado TERRA, Lisboa, Portugal
Se exploró el impacto de la inclusión dietética del 15% de Spirulina (Arthrospira platensis) en pollos de engorde, centrándose en los componentes celulares de la sangre, los metabolitos sistémicos y la composición hepática de lípidos y minerales. De los 14 a los 35 días de edad, se dividieron 120 pollos de engorde y se distribuyeron en cuatro tratamientos dietéticos: una dieta estándar a base de harina de maíz y soja (control), una dieta de espirulina al 15%, una dieta de espirulina extruida al 15% y una dieta de espirulina al 15% superdosificada con una mezcla de enzimas (0,20% de pancreatina porcina más 0,01% de lisozima). El análisis hematológico no reveló desviaciones significativas (p > 0,05) en los recuentos de células sanguíneas entre los tratamientos, lo que sugiere que la inclusión alta de Spirulina mantiene el equilibrio hematológico. La evaluación metabólica sistémica indicó una mayor capacidad antioxidante en aves con dietas de Spirulina (p < 0,001), lo que apunta a una posible reducción del estrés oxidativo. Sin embargo, el estudio observó un impacto perjudicial en las métricas de rendimiento del crecimiento, como el peso corporal final y el índice de conversión alimenticia (ambos p < 0,001), en los tratamientos alimentados con espirulina, ya que la suplementación con mezcla de enzimas superdosificadas no alivió estos efectos, pero la extrusión los mitigó. En cuanto a la composición hepática, las aves con dietas extruidas con espirulina y suplementadas con enzimas mostraron un aumento notable de los ácidos grasos n-3 (EPA, DPA, DHA) (p < 0,001), lo que llevó a una mejora de la relación AGPI n-6/n-3 (p < 0,001). A pesar de este cambio positivo, se observó una reducción de los lípidos hepáticos totales (p = 0,003) sin un cambio significativo en los niveles de colesterol. Nuestros hallazgos subrayan la necesidad de una mayor exploración de los niveles óptimos de inclusión, los métodos de procesamiento y las posibles mejoras enzimáticas de la espirulina en las dietas de los pollos de engorde. En última instancia, esta investigación tiene como objetivo lograr un equilibrio entre la promoción de beneficios para la salud y el mantenimiento de un rendimiento óptimo del crecimiento en la nutrición de las aves de corral.
1 Introducción
La creciente búsqueda mundial de fuentes de proteínas sostenibles y respetuosas con el medio ambiente, especialmente en la avicultura, ha impulsado la exploración de componentes de piensos no convencionales. Entre estos, las microalgas, particularmente las del género Spirulina, han emergido como contendientes notables (1-3). Arthrospira platensis, conocida ubicuamente como Spirulina, es una cianobacteria reconocida por su contenido superior de proteínas, carbohidratos, ácidos grasos esenciales, vitaminas, pigmentos y diverso perfil mineral. Sin embargo, estos contenidos están sujetos a modulaciones en función de las condiciones de cultivo y las variaciones estacionales (4, 5).
Además de su sólida cartera nutricional, la espirulina presenta una serie de beneficios funcionales que incluyen propiedades antimicrobianas, antioxidantes, antiinflamatorias e inmunomoduladoras (5-7). Sin embargo, la pared celular predominantemente indigerible del alga se asemeja a la de las bacterias gramnegativas con una composición principalmente de peptidoglicano y lipopolisacáridos, lo que presenta desafíos de digestibilidad para los animales monogástricos (8). Además, la bioaccesibilidad de las proteínas de las algas puede verse frustrada debido a los complejos proteína-pigmento, como las ficocianinas interconectadas con las membranas tilacoides (9).
Aunque las carbohidrasas, como la lisozima, se han mostrado prometedoras para desmantelar parcialmente la pared celular de la espirulina (10), un proceso de gelificación parece atrapar nutrientes, impidiendo su digestión (3, 11). Se han realizado intentos preliminares de hidrolizar proteínas de Spirulina a través de pretratamientos físicos junto con pancreatina o tripsina (12, 13), pero el ámbito de la implementación de dichas enzimas para degradar proteínas en la alimentación de A. platensis para animales monogástricos sigue siendo en gran medida inexplorado. Sin embargo, los estudios emergentes acentúan la eficacia potencial de enzimas exógenas específicas, como las enzimas activas en carbohidratos, para romper la pared celular de la espirulina y amplificar la biodisponibilidad de sus beneficios nutricionales (14). La incorporación de estas enzimas exógenas como aditivos alimentarios en las dietas de las aves de corral es una estrategia muy utilizada para mitigar la escasez de enzimas endógenas y neutralizar los factores antinutricionales, mejorando la digestibilidad de los componentes de la dieta (15, 16). Además, estas enzimas favorecen la generación de oligosacáridos prebióticos, lo que favorece la salud intestinal y los parámetros de crecimiento (17).
Tradicionalmente, las microalgas se han utilizado más como suplementos que como ingredientes principales en las dietas monogástricas (18). Las investigaciones anteriores de este equipo evaluaron las ramificaciones de la integración de microalgas como ingrediente alimentario en el rendimiento del crecimiento, el estado de salud y la composición de lípidos hepáticos en pollos de engorde (19, 20), y se extendieron a los cerdos (19, 20). Esta investigación es pionera en el examen de los efectos sinérgicos de la alimentación prolongada con un alto nivel de A. platensis, ya sea sola o suplementada con diferentes enzimas (predominantemente peptidasas, EC 3.4), sobre la salud y el metabolismo lipídico hepático en pollos de engorde. Si bien los impactos positivos de la suplementación con espirulina en la dieta sobre el rendimiento y la salud de los pollos de engorde se han investigado tentativamente [por ejemplo, Pestana et al. (3)], persiste un ámbito inexplorado con respecto a los efectos interactivos de las multienzimas en superdosis junto con niveles más altos de inclusión de espirulina. De hecho, informes anteriores mostraron el potencial de las multienzimas superdosificadas (más altas de lo recomendado) para mejorar el rendimiento del crecimiento, la salud intestinal y la absorción de nutrientes en las aves de corral (21-23). La aplicación de espirulina extruida en uno de los tratamientos dietéticos parece ser una combinación prometedora, como se ha demostrado en estudios in vitro, para los pretratamientos enzimáticos (por ejemplo, pancreatina porcina), ya que la alta presión y temperatura pueden causar una alteración en la pared celular de las microalgas, contribuyendo a mejorar la biodisponibilidad de proteínas relevantes, como la ficocianina (12, 13).
El propósito de este trabajo fue evaluar el impacto de la incorporación de un 15% de biomasa de A. platensis en dietas de pollos de engorde, ya sea de forma exclusiva o en combinación con una mezcla de enzimas superdosificantes (pancreatina y lisozima), o cuando se extruye espirulina, desde el día 14 hasta el 35, sobre las células sanguíneas de los pollos de engorde, los metabolitos plasmáticos y la composición hepática, que comprende ácidos grasos, diterpenos antioxidantes, pigmentos y perfil mineral.
2 Materiales y métodos
2.1 Alojamiento de animales y tratamientos dietéticos
La investigación incluyó un grupo de ciento veinte pollos de engorde Ross 308 machos de un día de edad adquiridos en Pinto Valouro (Bombarral, Portugal), cada uno con un peso inicial promedio de 39,3 ± 2,30 g. Desde el día 1, se alojaron aleatoriamente en 40 recintos con malla metálica (56 cm de alto x 56 cm de largo x 50 cm de ancho) durante 35 días en condiciones ambientales reguladas. La temperatura ambiente y el flujo de aire se supervisaron de forma persistente según los protocolos previamente establecidos (2, 19). Adhiriéndose a la doctrina de las 3R (Reducir, Refinar y Reemplazar) para reducir el número de animales empleados, cada recinto albergaba tres polluelos, con 10 recintos idénticos designados para cada experimento dietético. El ensayo experimental se llevó a cabo en la Escuela de Agricultura de la Universidad de Lisboa (Lat: 38°42′27.5″N; GNL: 9°10′56.3″W). El Comité de Bienestar Animal de la Escuela de Agricultura de la Universidad de Lisboa (ORBEA/ISA) aprobó todas las metodologías experimentales con animales, asignando al estudio un número de código de protocolo 0421/000/000/2022.
En la quincena inicial, a los pollitos se les administró una dieta básica formulada a partir de harina de maíz y soja procedente de Rações Veríssimo S. A. (Leiria, Portugal). Después de esta fase, desde el día 14 hasta el día 35, hicieron la transición a una de las cuatro dietas experimentales distintas que se suministran ad libitum diariamente. Estas dietas abarcaron: (1) una dieta estándar basada en harina de maíz y soja (CTR); (2) una dieta fortificada con un 15% de polvo de espirulina (obtenida de Allmicroalgae, Pataias, Portugal) (SP); (3) una dieta que contenga un 15% de polvo de espirulina extruida (SPE); y (4) una dieta suplementada con un 15% de espirulina en polvo junto con una mezcla de enzimas a medida, constituida por un 0,20% de extracto de pancreatina porcina (procedente de Merck, Darmstadt, Alemania) y un 0,01% de lisozima extraída de la clara de huevo de gallina (procedente de Sigma-Aldrich, Missouri, Estados Unidos) (SPM). La extrusión de microalgas fue realizada por la empresa Sparos (Olhão, Portugal), siguiendo unas condiciones detalladas: 340 mL de adición de agua por minuto, a 34 bares y 118°C para el último barril de extrusión. Este procedimiento se realizó de 3 a 7 s. Luego, los gránulos de algas se secaron durante 8 y 10 min a 120 °C (12, 13). El extracto encapsulado de pancreatina porcina comprendía 350 FIP-U/g de proteasa, 6.000 FIP-U/g de lipasa y 7.500 FIP-U/g de amilasa. El polvo de lisozima contenía 70.000 U/mg de proteína. En el cuadro 1 se proporciona una composición detallada de los componentes y aditivos de los piensos para cada dieta experimental. Durante el ensayo, se observó una mortalidad del 2,5% (tres animales de un total de 120).
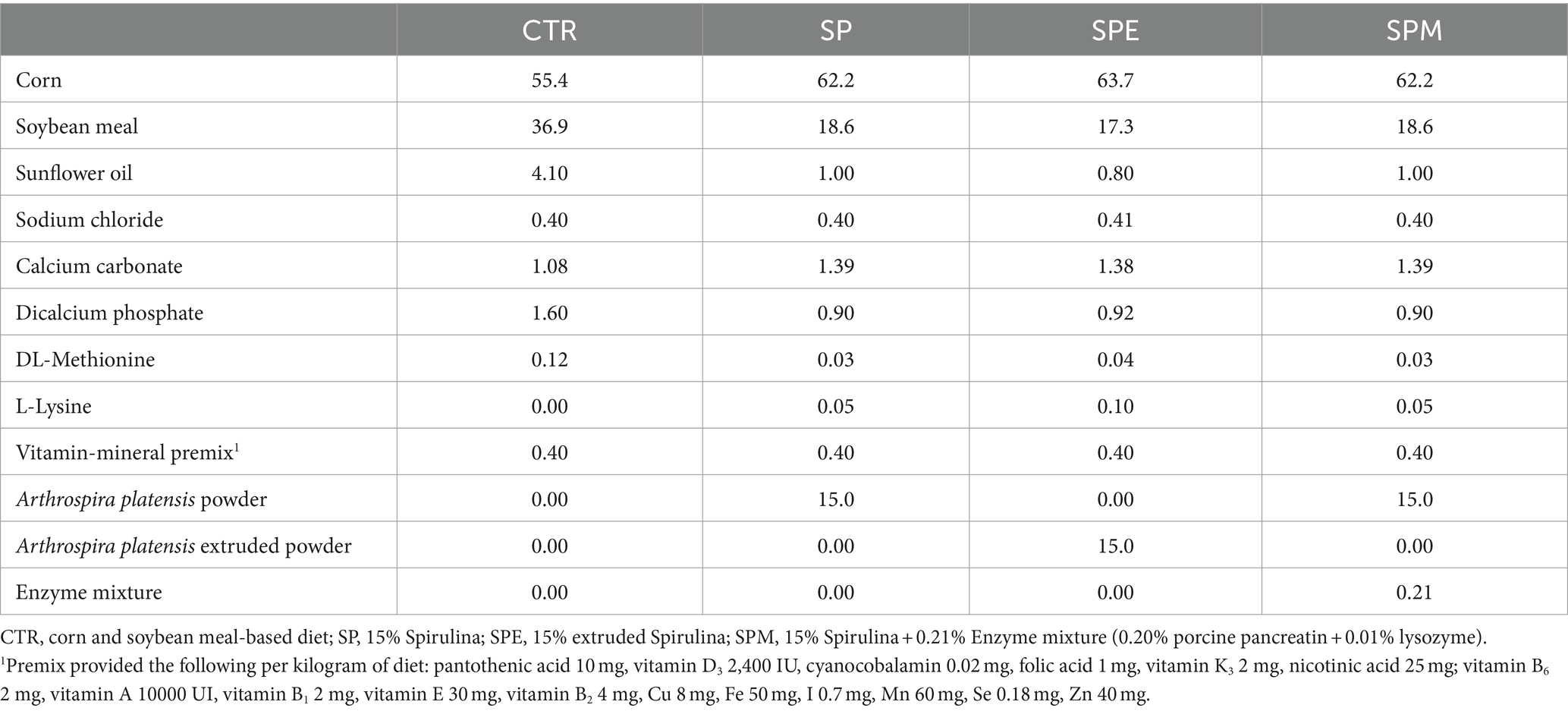 Tabla 1. Composición de ingredientes de las dietas experimentales de pollos de engorde desde el día 14 hasta el día 35 (%, según la base de la alimentación).
Tabla 1. Composición de ingredientes de las dietas experimentales de pollos de engorde desde el día 14 hasta el día 35 (%, según la base de la alimentación).
Se realizaron evaluaciones semanales para registrar los pesos de los pollos de engorde y los comederos, con el aprovisionamiento diario de alimento para calcular los parámetros de ganancia de peso corporal (BWG) (determinado por el diferencial de peso semanal dividido por 7), la ingesta diaria promedio de alimento (ADFI) (calculada como el consumo semanal por jaula, dividido por 7) y el índice de conversión alimenticia (FCR) (el cociente de consumo semanal dividido por 3 y ganancia semanal de peso corporal).
2.2 Análisis de microalgas y dietas experimentales
En la Tabla 2 se muestra la composición proximal de las microalgas y las dietas experimentales ofrecidas a los pollos de engorde desde el día 14 hasta el 35. La evaluación del contenido de materia seca, proteína bruta, grasa bruta y cenizas se realizó de acuerdo con las metodologías estipuladas por la AOAC (24). La cuantificación del contenido energético bruto tanto en la microalga como en las dietas se realizó empleando una calorimetría de bomba adiabática (Parr 1.261, Parr Instrument Company, Moline, IL).
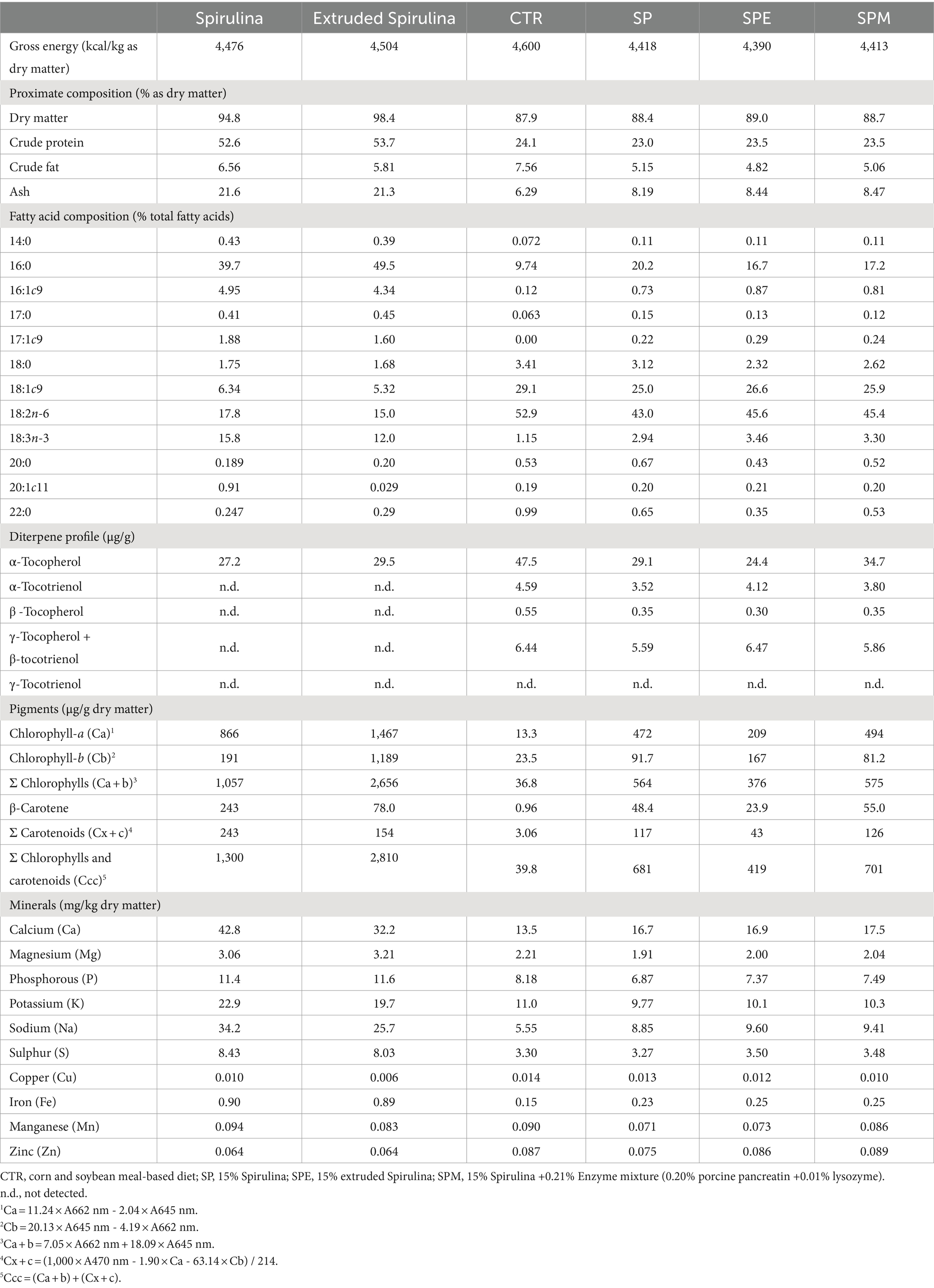 Tabla 2. Composiciones proximales y nutricionales de Arthrospira platensis y dietas experimentales ofrecidas a pollos de engorde desde el día 14 hasta el 35.
Tabla 2. Composiciones proximales y nutricionales de Arthrospira platensis y dietas experimentales ofrecidas a pollos de engorde desde el día 14 hasta el 35.
La composición de ácidos grasos en A. platensis y las dietas experimentales se realizó después de una única etapa de extracción y metilación ácida, alineándose con el método delineado por Sukhija y Palmquist (25). Los ésteres metílicos de ácidos grasos resultantes (FAME) se analizaron mediante cromatografía de gases interconectada con un detector de ionización de llama (HP7890A Hewlett-Packard, Avondale, PA), desplegando una columna capilar Supelcowax® 10 (30 m × 0,20 mm de diámetro interior, 0,20 μm de espesor de película, Supelco, Bellefonte, PA, Estados Unidos), mediante el protocolo propuesto por Alfaia et al. (26). La identificación de FAME dentro de las muestras se logró mediante la yuxtaposición con una mezcla estándar de FAME (mezcla de FAME de 37 componentes C4-C24, Supelco, Bellefonte, PA, EE. UU.). El éster metílico del ácido heneicosanoico (21,0) se alistó como el estándar interno para la cuantificación de ácidos grasos individuales, con el contenido de ácidos grasos articulado como un porcentaje de los ácidos grasos totales.
Los diterpenos y β-caroteno dentro de las dietas experimentales se evaluaron siguiendo la metodología delineada por Prates et al. (27). Las muestras duplicadas, cada una con un peso de 0,1 g, se enriquecieron con ácido ascórbico (vitamina C) y se sometieron a una reacción de saponificación en un baño de agua mantenido a 80 °C durante 15 min. Después de la extracción y centrifugación (2.500 rpm, 10 min), las fases de n-hexano se examinaron utilizando un sistema de HPLC (Agilent 1.100 Series, Agilent Technologies Inc., Palo Alto, CA, Estados Unidos) equipado con una columna de sílice de fase normal (Zorbax RX-Sil, tamaño de partícula de 5 μm, 250 mm × 4,6 mm de diámetro interior). El β-caroteno se detectó empleando un detector de matriz de fotodiodos UV-visible (λ = 450 nm), mientras que los tocoferoles y los tocotrienoles se identificaron con un detector de fluorescencia (excitación λ = 295 nm y emisión λ = 325 nm). La cuantificación del β-caroteno y los análogos de la vitamina E se facilitó mediante la utilización de curvas estándar que correlacionan el área del pico con la concentración.
El perfil pigmentario de las dietas experimentales se determinó con alteraciones menores a los protocolos expuestos por Hynstova et al. (28) y Pestana et al. (3). Concisamente, se amalgamaron 0,1 g de muestras con 5 mL de acetona y se homogeneizaron en la oscuridad, seguido de centrifugación (2.500 rpm, 10 min) se aisló y analizó el sobrenadante. Las clorofilas a y b se detectaron en longitudes de onda de 662 nm y 645 nm, respectivamente, y carotenoides totales a 470 nm mediante espectrofotometría UV-Vis (Genesys 150, Thermo Scientific, Waltham, Massachusetts, Estados Unidos). El contenido de pigmento se dedujo empleando ecuaciones proporcionadas por Hynstova et al. (28).
La composición mineral de A. platensis y las dietas experimentales se descifraron asignando 0,3 g de muestras en un tubo de digestión, introduciendo ácido nítrico (65%) y ácido clorhídrico (37%), y digiriendo a 95°C dentro de una cámara ventilada durante 16 h. Posteriormente, se adicionó peróxido de hidrógeno (30%) según lo descrito por Ribeiro et al. (29). La solución resultante se filtró y analizó empleando Espectrometría de Emisión Óptica de Plasma Acoplado Inductivamente (ICP-OES, iCAP 7.200 duo Thermo Scientific, Waltham, MA, Estados Unidos), con cuantificación de minerales lograda a través de patrones y curvas de calibración.
2.3 Sacrificio y recogida de muestras
Al final del ensayo, en una sala con condiciones específicas para el sacrificio, se seleccionó un pollo de engorde de 35 días, peso medio, de cada corral, para su sacrificio, mediante aturdimiento eléctrico seguido de exanguinación manual. Se recogieron muestras de sangre (10 mL), el mismo día, directamente en los tubos de Sarstedt (Numbrecht, Alemania), y posteriormente se centrifugaron para obtener plasma. Se recogieron muestras de hígado, se picaron, se sellaron al vacío y se almacenaron a -20 °C para su posterior análisis. El resto de las aves fueron sacrificadas como se describió anteriormente.
2.4 Análisis de células sanguíneas y metabolitos plasmáticos
Según lo articulado por Madeira et al. (19), los glóbulos rojos, los glóbulos blancos y los trombocitos se enumeraron empleando analizadores Sysmex XN-10 (Sysmex Corporation, Kobe, Japón). El recuento de glóbulos rojos se determinó mediante el método de variación de impedancia, después del enfoque hidrodinámico. Se realizó la enumeración diferencial de glóbulos blancos (%) en frotis de sangre, que se decoloraron utilizando la técnica de May-Grünwald-Giemsa. La concentración de hemoglobina se midió fotométricamente, sirviendo como reactivo el lauril sulfato de sodio.
Metabolitos plasmáticos seleccionados, que abarcan glucosa, urea, creatinina, proteínas totales y diversas formas lipídicas (triacilgliceroles, TAG; colesterol total; HDL-colesterol; y colesterol LDL), además de las actividades enzimáticas de la alanina aminotransferasa (ALT, EC 2.6.1.2), la aspartato aminotransferasa (AST, EC 2.6.1.1), la fosfatasa alcalina (ALP, EC 3.1.3.1) y la γ-glutamiltransferasa (GGT, EC 2.3.2.13) se determinaron utilizando kits de diagnóstico del Sistema Analítico Modular Hitachi de Roche Diagnostics (Mannheim, Alemania). Se aplicaron fórmulas delineadas por Friedewald et al. (30) y Covaci et al. (31) para el cálculo del colesterol VLDL y de los lípidos totales, respectivamente. Los niveles de proteína C reactiva se evaluaron mediante inmunoturbidimetría (Roche Diagnostics, Meylan, Francia). Electrolitos primarios, incluidos Na, K y Cl++− fueron evaluados a través de potenciometría indirecta.
2.5 Análisis de la composición de lípidos totales y ácidos grasos en el hígado
Los lípidos totales se aislaron de las muestras de hígado liofilizadas, por duplicado, empleando una mezcla solvente de diclorometano y metanol (2:1 v/v), y se cuantificaron mediante análisis gravimétrico según el protocolo delineado por Folch et al. (32). La composición de ácidos grasos dentro de las muestras de hígado se descifró de manera similar a la metodología aplicada para A. platensis y las dietas experimentales. No obstante, se aprovechó un proceso secuencial de transesterificación alcalina seguido de transesterificación ácida para convertir los ácidos grasos en FAME, divergiendo del único procedimiento de metilación ácida. Las condiciones cromatográficas fueron congruentes con el protocolo previamente articulado por Alfaia et al. (26).
2.6 Determinación de colesterol total, diterpenos y β-caroteno en el hígado
Para la extracción de colesterol total, diterpenos y β-caroteno, se emplearon muestras de hígado duplicadas, con un peso de 0,75 g cada una, siguiendo el mismo procedimiento delineado para las dietas experimentales. Esto abarcó la saponificación directa, la extracción singular de n-hexano y el posterior análisis por HPLC según el protocolo detallado en Prates et al. (27).
Para la extracción de pigmentos (que abarca clorofila-a, clorofila-b y carotenoides totales) de muestras de hígado, se utilizó una versión ligeramente adaptada del método utilizado para las dietas experimentales. En resumen, se amalgamaron 1,5 g de tejido hepático con 3 ml de acetona, seguido de una homogeneización de 1 min en un ambiente oscuro. Después de la centrifugación (3.000 rpm durante 5 min a 4°C), el sobrenadante fue secuestrado y examinado bajo las condiciones estipuladas por Coelho et al. (33). Las cantidades de pigmento se determinaron utilizando las ecuaciones presentadas por Hynstova et al. (28).
2.7 Determinación del perfil mineral en el hígado
El perfil mineral hepático se midió siguiendo el procedimiento establecido para las microalgas y dietas, excepto para el bromuro, cuya determinación se realizó según el método expuesto por Delgado et al. (34).
2.8 Análisis estadístico
El conjunto de datos se sometió a análisis como un diseño completamente aleatorizado empleando el modelo Mixto Lineal Generalizado (GLM) en el marco del software del Sistema de Análisis Estadístico (SAS) (SAS Institute Inc., Cary, NC) (35). La dieta experimental fue el único factor fijo y el efecto primario dentro del modelo estadístico. La unidad experimental fue el recinto o el animal, dependiendo de los parámetros evaluados. El recinto se designó para evaluar el peso corporal, la ganancia de peso corporal, la ingesta media diaria de alimento y el índice de conversión alimenticia, mientras que el ave individual fue la unidad de análisis para el resto de los parámetros (metabolitos plasmáticos y parámetros hepáticos completos). La homogeneidad de las varianzas de los datos se comprobó mediante la prueba de Levene y la normalidad mediante la prueba de Shapiro-Wilk. Para las comparaciones múltiples entre medias de mínimos cuadrados, se invocó el método ajustado de Tukey-Kramer (opción PDIFF). Todos los valores se articularon como la media junto con el SEM (error estándar de la media). Se estableció un umbral de p-valores menores de 0,05 para la determinación de la significación estadística.
Para explorar las interrelaciones entre los parámetros sanguíneos y las variables hepáticas, se realizó un Análisis de Componentes Principales (ACP) utilizando el paquete de software SPSS Statistics for Windows (IBM Corp., versión 2020, versión 27.0, Armonk, NY, Estados Unidos).
3 Resultados
3.1 Efecto de las dietas experimentales sobre el rendimiento y la ingesta de alimento de las aves
La investigación sobre el rendimiento del crecimiento de los pollos de engorde bajo diferentes tratamientos se presenta en la Figura 1 con fines contextuales. Esta evaluación abarcó el período comprendido entre los días 14 y 35 del desarrollo de los pollos. Los resultados mostraron que las dietas que contenían únicamente espirulina (SP) y espirulina combinadas con la mezcla de enzimas (SPM) redujeron significativamente el peso corporal final (PC) de las aves, en comparación con la dieta de control. Sin embargo, no se observaron diferencias significativas en el peso corporal final entre el grupo alimentado con espirulina extruida (SPE) y el grupo control. Un patrón similar surgió en las métricas de aumento de peso corporal (BWG), con la dieta SPE mostrando resultados comparables al grupo de control, claramente diferentes de las dietas SP y SPM. En cuanto a la ingesta media diaria de alimento (ADFI), se observó un aumento interesante en el grupo SPE en comparación con la dieta SP. En cuanto al índice de conversión alimenticia (FCR), todos los grupos que incorporaron Spirulina condujeron a un aumento del FCR en relación con el testigo, lo que significó una disminución en la eficiencia alimenticia.
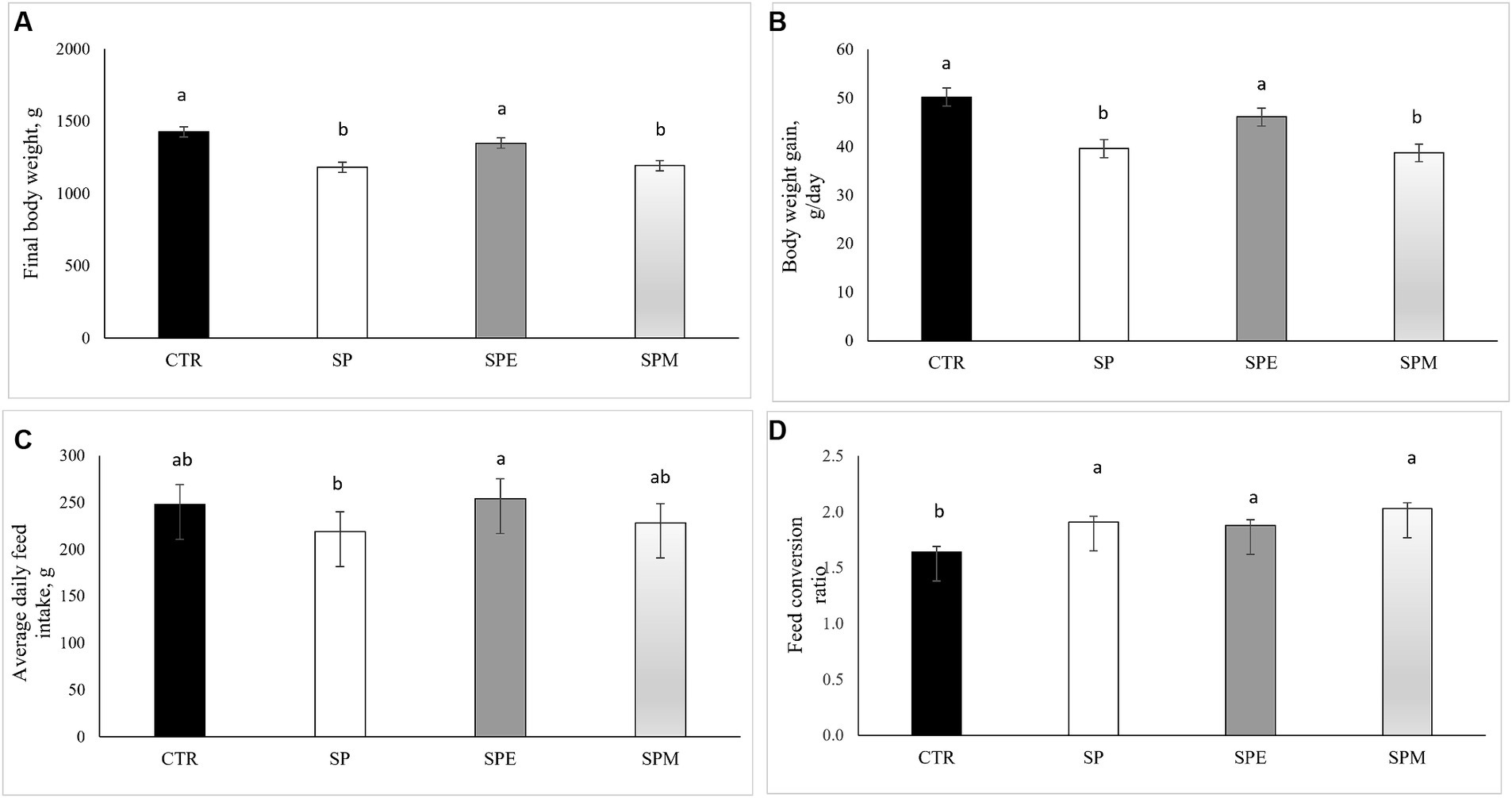 Figura 1. Impacto de los tratamientos dietéticos en el rendimiento del crecimiento de los pollos de engorde: (A) Peso corporal final (g); (B) Aumento de peso corporal (g/día); C) Ingesta media diaria de alimento (g); (D) Índice de conversión alimenticia. CTR, dieta a base de harina de maíz y soja; SP, 15% Espirulina; SPE, 15% de Espirulina extruida; SPM, 15% Espirulina + 0,21% mezcla de enzimas (0,20% pancreatina porcina +0,01% lisozima). Los valores se muestran como media ± SEM (error estándar de la media). a,b,cDiferentes superíndices dentro de una fila indican una diferencia significativa (p < 0,05).
Figura 1. Impacto de los tratamientos dietéticos en el rendimiento del crecimiento de los pollos de engorde: (A) Peso corporal final (g); (B) Aumento de peso corporal (g/día); C) Ingesta media diaria de alimento (g); (D) Índice de conversión alimenticia. CTR, dieta a base de harina de maíz y soja; SP, 15% Espirulina; SPE, 15% de Espirulina extruida; SPM, 15% Espirulina + 0,21% mezcla de enzimas (0,20% pancreatina porcina +0,01% lisozima). Los valores se muestran como media ± SEM (error estándar de la media). a,b,cDiferentes superíndices dentro de una fila indican una diferencia significativa (p < 0,05).
3.2 Efecto de las dietas experimentales sobre el perfil hematológico y bioquímico
En la Tabla 3 se muestra el recuento de células sanguíneas y los metabolitos plasmáticos de pollos de engorde alimentados con un 15% de A. platensis, ya sea exclusivamente, o cuando la espirulina se extruye o se combina con una mezcla de enzimas (pancreatina y lisozima). Los recuentos de glóbulos blancos (p = 0,702), granulocitos (p = 0,769), linfocitos (p = 0,741), monocitos (p = 0,473), glóbulos rojos (p = 0,377), trombocitos (p = 0,708) y hemoglobina (p = 0,062) no se vieron afectados por las dietas.
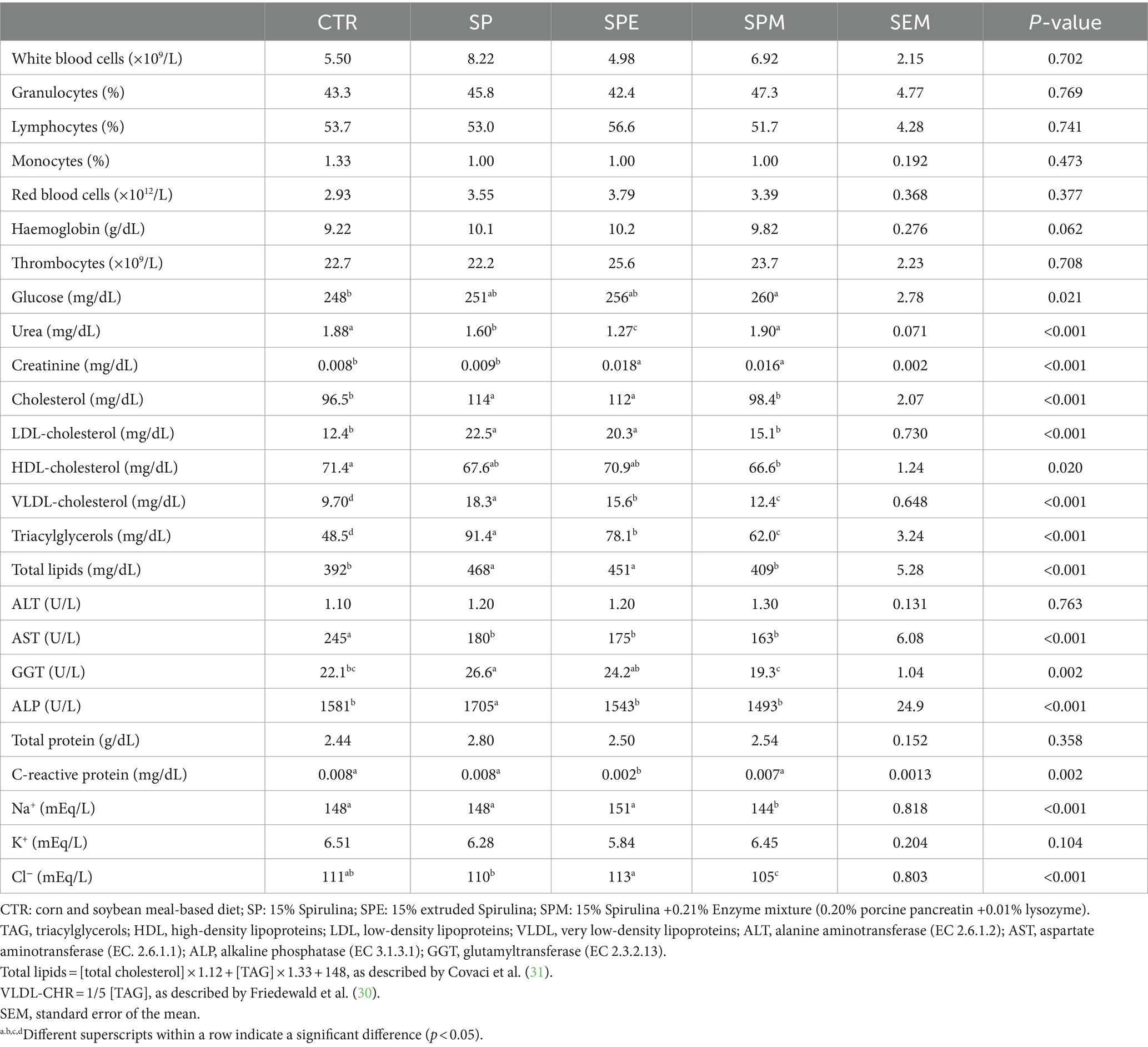 Tabla 3. Perfiles hematológicos y bioquímicos de pollos de engorde alimentados con las dietas experimentales.
Tabla 3. Perfiles hematológicos y bioquímicos de pollos de engorde alimentados con las dietas experimentales.
En cuanto a los metabolitos plasmáticos, los niveles de glucosa alcanzaron los niveles más altos en pollos de engorde alimentados con SPM, intermedios en SP o SPE, y más bajos en pollos de engorde alimentados con la dieta control (p = 0,021). La urea se redujo en pollos de engorde alimentados con SP y SPE en relación con el control y SPM (p < 0.001). La función renal se evaluó además mediante creatinina, que mostró los valores más altos en pollos de engorde alimentados con SPE y SPM (p < 0,001). Consistentemente, la dieta SP aumentó los parámetros lipídicos que se redujeron con la suplementación con mezcla de enzimas para colesterol (p < 0,001), colesterol LDL (p < 0,001), colesterol VLDL (p < 0,001), triacilgliceroles (p < 0,001) y lípidos totales (p < 0,001). En cuanto a la función hepática, la AST se redujo con las dietas basadas en Spirulina (p < 0,001) incluso si la ALT se mantuvo sin cambios (p = 0,753) en todos los tratamientos dietéticos. La FA (p < 0,001) y la GGT (p = 0,002) fueron aumentadas por la dieta SP en comparación con los otros tratamientos dietéticos. Las proteínas totales se mantuvieron sin cambios a lo largo de las dietas (p = 0,358). La dieta SPE redujo la proteína C reactiva en fase aguda sobre los otros tres tratamientos dietéticos (p = 0,002). En la evaluación de los principales electrolitos, el sodio (p < 0,001) y el cloruro (p < 0,001) se redujeron con la dieta SPM en relación con los otros tratamientos dietéticos. Por el contrario, no se observaron cambios significativos para el potasio (p = 0,104).
3.3 Efecto de las dietas experimentales sobre la composición hepática de lípidos totales, colesterol y ácidos grasos
El perfil lipídico hepático y la composición de ácidos grasos de pollos de engorde alimentados con 15% de A. platensis, ya sea exclusivamente, o cuando la espirulina se extruye o se combina con una mezcla de enzimas (lisozima y pancreatina porcina) se muestran en la Tabla 4. Se observó una disminución en los lípidos totales (p = 0,003) entre los pollos de engorde a los que se les administró Spirulina sola o en su estado extruido, en contraste con una disminución en los niveles de colesterol dentro del grupo control (p < 0,001). Aunque el agregado de ácidos grasos saturados (AGS) permaneció invariable a lo largo de los regímenes dietéticos (p > 0,05), ciertos ácidos grasos específicos como el ácido láurico (12:0) experimentaron una elevación en los niveles entre los pollos de engorde sometidos a la dieta de espirulina extruida (p = 0,015), mientras que los niveles de ácido palmítico (16,0) y ácido araquídico (20,0) aumentaron en escenarios en los que se administró espirulina extruida o espirulina acoplada con enzimas (p = 0,009 y p = 0,005, respectivamente).
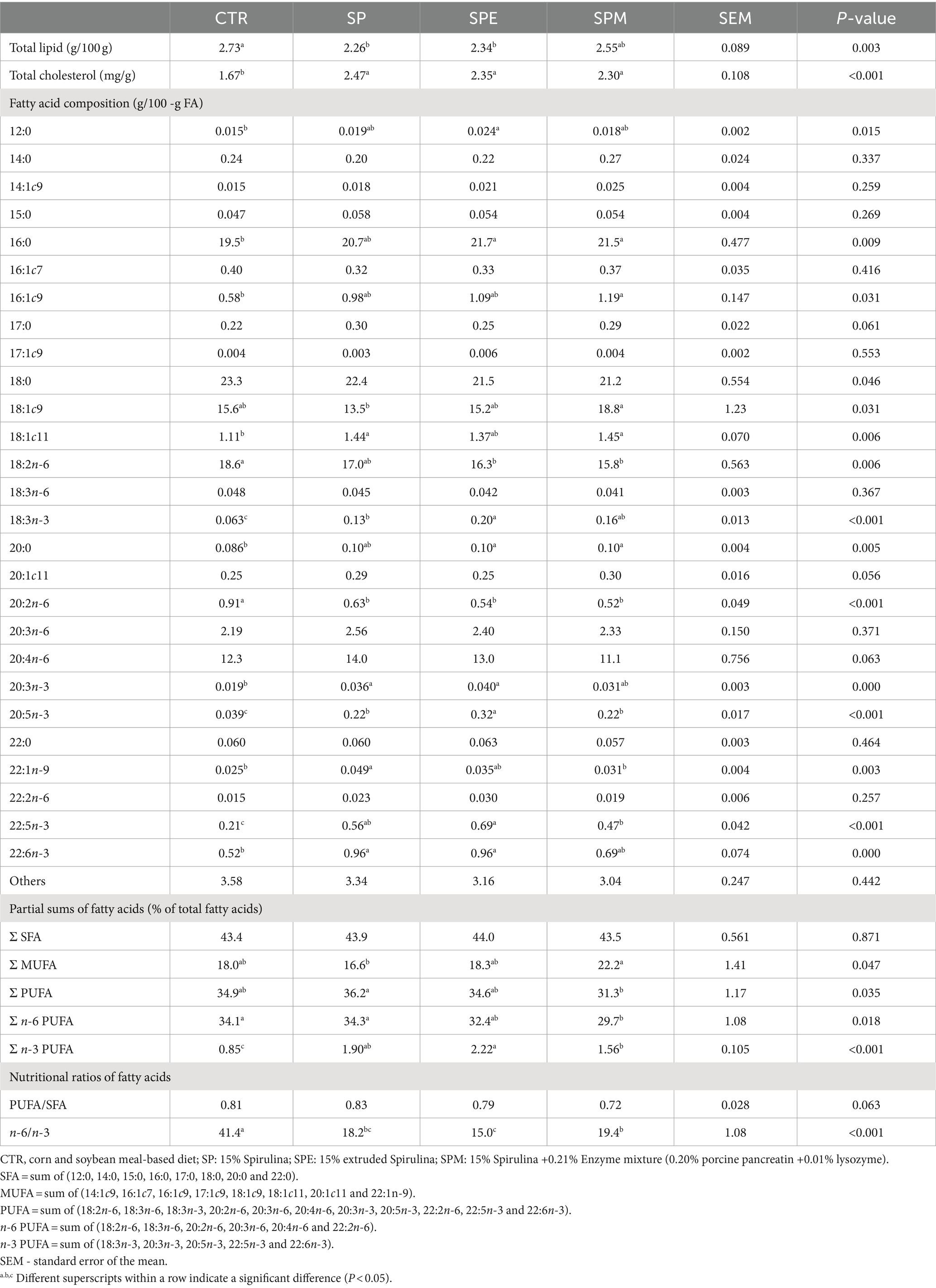 Tabla 4. Composición hepática de lípidos totales, colesterol total y ácidos grasos de pollos de engorde alimentados con dietas experimentales.
Tabla 4. Composición hepática de lípidos totales, colesterol total y ácidos grasos de pollos de engorde alimentados con dietas experimentales.
La agregación de ácidos grasos monoinsaturados (MUFA) alcanzó su punto máximo en los pollos de engorde alimentados con Spirulina junto con la mezcla de enzimas, exhibió valores intermedios en aquellos alimentados únicamente con Spirulina o la dieta de control, y se desplomó a los más bajos en aquellos a los que se les administró Spirulina extruida (p = 0.047). Este patrón fue impulsado predominantemente por fluctuaciones en los niveles de ácido oleico predominante (18:1c9) (p = 0,031) y ácidos grasos menores ácido palmitoleico (16:1 c9) y ácido vaccénico (18:1c11) (p = 0,006). Por el contrario, se observó una tendencia inversamente proporcional en el contenido de ácidos grasos poliinsaturados totales (AGPI) (p = 0,035), y específicamente en la subcategoría de AGPI n-6 (p = 0,018), atribuible en gran medida a las proporciones de ácido linoleico (18:2n-6) (p = 0,006) y ácido eicosadienoico (20:2n-6) (p < 0,001).
Además, se observó una elevación de los AGPI n-3 totales en los pollos de engorde alimentados con espirulina extruida, seguidos de los suplementados con espirulina y enzimas (p < 0,001), en comparación con el grupo control. Este ascenso culminó con una notable reducción de la relación n-6/n-3 (p < 0,001), particularmente pronunciada en la dieta SPE. Todos los ácidos grasos n-3 individuales registraron un repunte bajo las dietas a base de espirulina (p < 0,001) en relación con el control.
3.4 Efecto de las dietas experimentales sobre el perfil diterpénico hepático y los pigmentos
La Tabla 5 muestra el perfil de diterpenos hepáticos y la composición de pigmentos de pollos de engorde alimentados con 15% de A. platensis, ya sea exclusivamente, o cuando la espirulina se extruye o se combina con una mezcla de enzimas (lisozima y pancreatina porcina). Al examinar el perfil de diterpenos, se detectó β-caroteno, un precursor de la vitamina A, en el hígado de los pollos de engorde en todas las dietas experimentales, aunque sin cambios significativos entre los tratamientos. Por el contrario, se observó una reducción notable en los niveles de α-tocoferol en las aves que recibieron dietas suplementadas con Spirulina (p < 0,001). Esta disminución fue pronunciada independientemente de si la espirulina se extruyó o se complementó con la mezcla de enzimas. De manera similar, los niveles agregados de γ-tocoferol y β-tocotrienol exhibieron una disminución significativa en los tratamientos alimentados con Spirulina en comparación con el control (p < 0.001). Los niveles de α-tocotrienol presentaron una varianza marginal entre los tratamientos, acercándose al umbral de significación (p = 0,052).
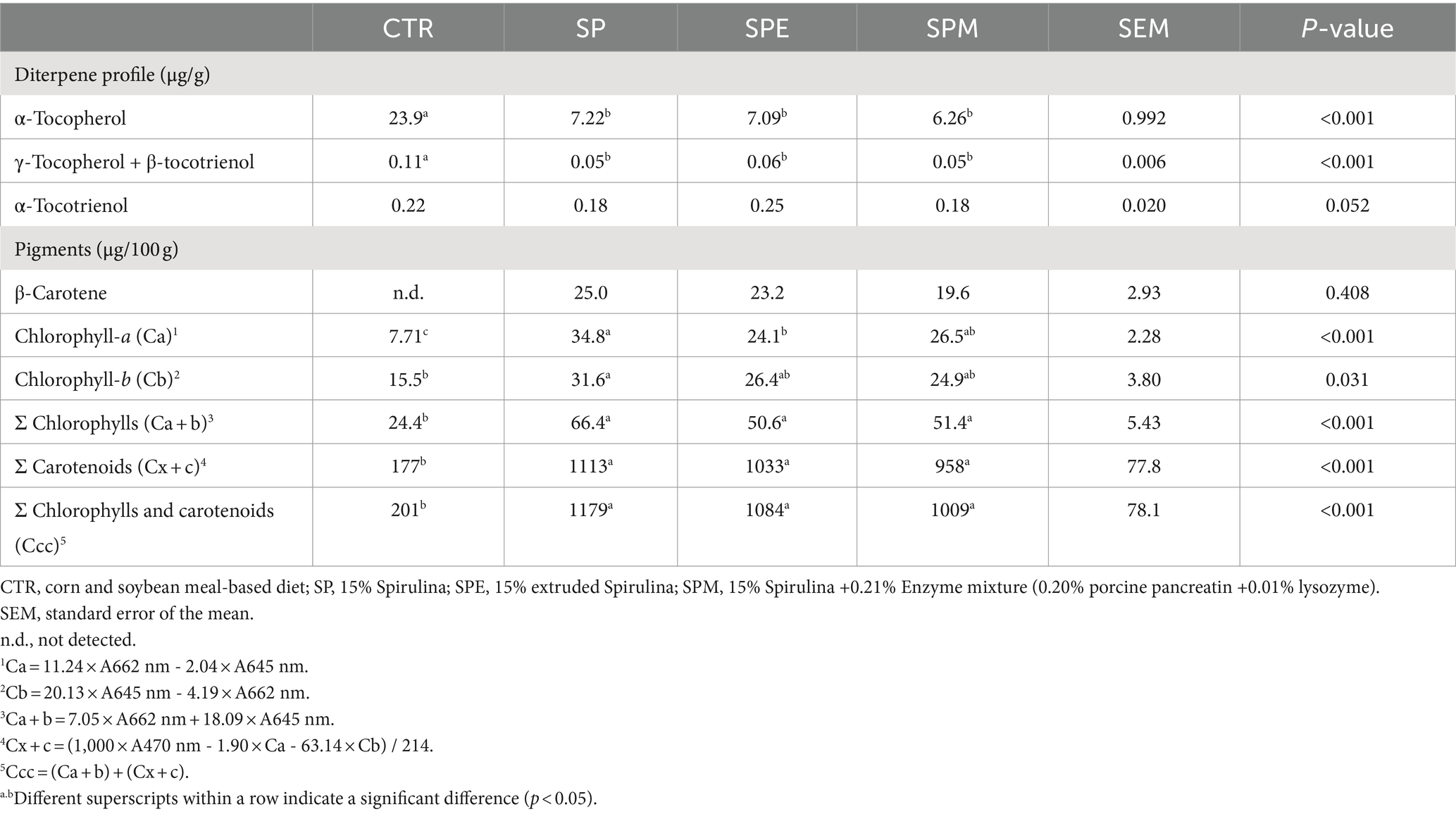 Tabla 5. Perfil diterpénico hepático y pigmentos de pollos de engorde alimentados con dietas experimentales.
Tabla 5. Perfil diterpénico hepático y pigmentos de pollos de engorde alimentados con dietas experimentales.
Con respecto a los pigmentos, los datos manifestaron una marcada elevación de la clorofila-a, las clorofilas totales, los carotenoides totales y la suma de las clorofilas totales más los carotenoides en el hígado de los pollos de engorde bajo todas las dietas inclusivas de Spirulina (p < 0,001). Estas mejoras se vieron afectadas de manera consistente, sin la extrusión de Spirulina o la suplementación de la mezcla de enzimas. Sorprendentemente, los niveles de clorofila-b se duplicaron en las aves alimentadas únicamente con Spirulina en comparación con las de la dieta control (p = 0,031).
3.5 Efecto de las dietas experimentales sobre el perfil mineral hepático
En la Tabla 6 se muestra el perfil mineral en el hígado de pollos de engorde alimentados con 15% de A. platensis, ya sea exclusivamente, o cuando la espirulina se extruye o se combina con una mezcla de enzimas (pancreatina y lisozima). Entre los minerales observados, el manganeso (Mn) emergió como un elemento distinto que muestra un aumento significativo en los niveles en pollos de engorde alimentados con espirulina extruida o espirulina suplementada con el brebaje enzimático, en comparación con el grupo de control (p = 0,008).
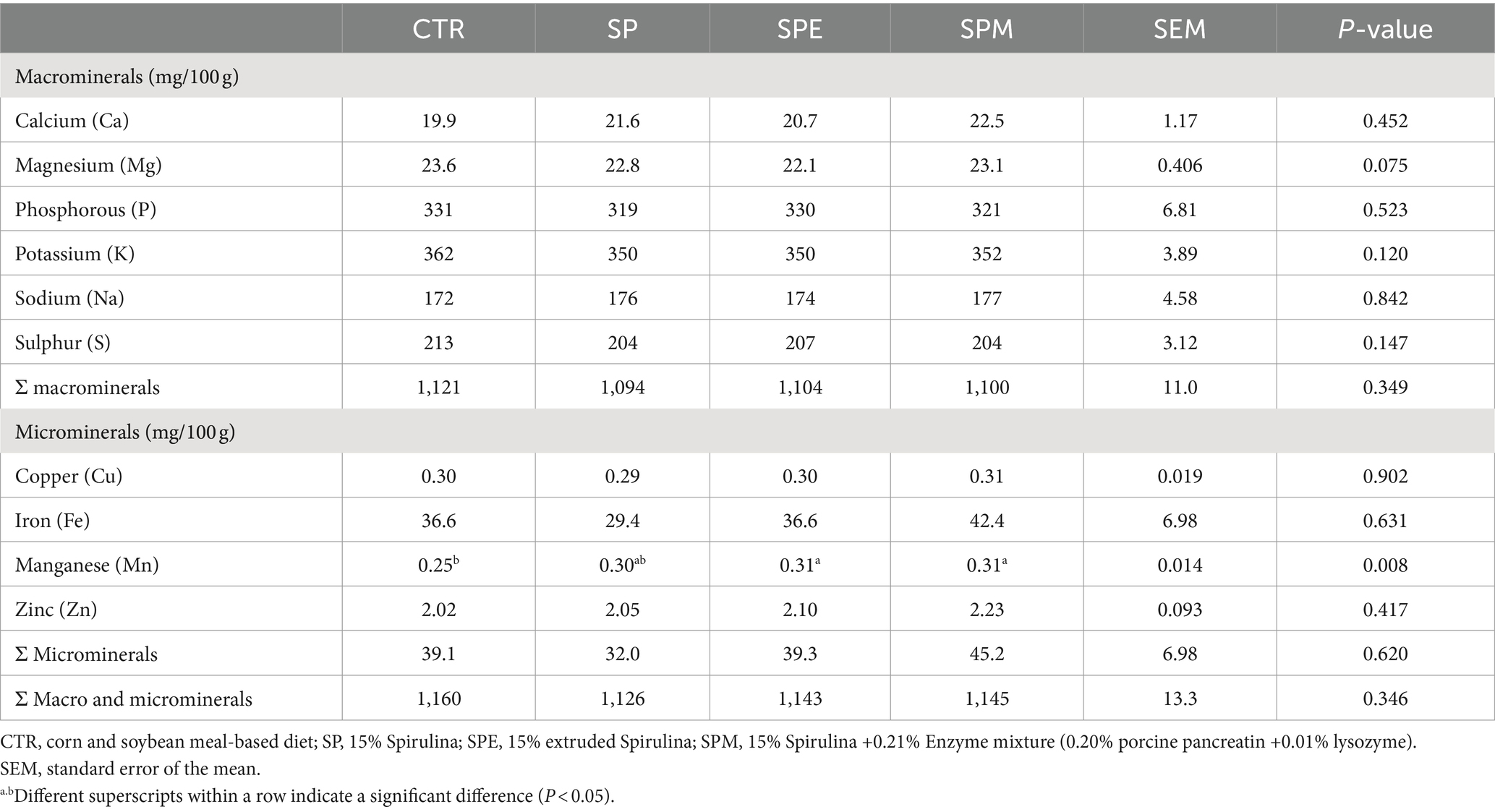 Tabla 6. Perfil mineral hepático de pollos de engorde alimentados con dietas experimentales.
Tabla 6. Perfil mineral hepático de pollos de engorde alimentados con dietas experimentales.
Contrariamente a la dinámica del manganeso, el resto de los micro y macrominerales, tanto de forma individual como acumulativa, mantuvieron un estado estacionario en todas las facciones experimentales (p > 0,05). El cuadro de macrominerales, que abarca el calcio (Ca), el magnesio (Mg), el fósforo (P), el potasio (K), el sodio (Na) y el azufre (S), junto con el consorcio de microminerales de cobre (Cu), hierro (Fe) y zinc (Zn), mostró una notable estabilidad.
3.6 Análisis de componentes principales de la composición plasmática y hepática
El Análisis de Componentes Principales (ACP) se realizó utilizando variables de metabolitos plasmáticos de pollos de engorde y los lípidos hepáticos mostraron distintos patrones de variabilidad entre los tratamientos dietéticos (Figura 2).
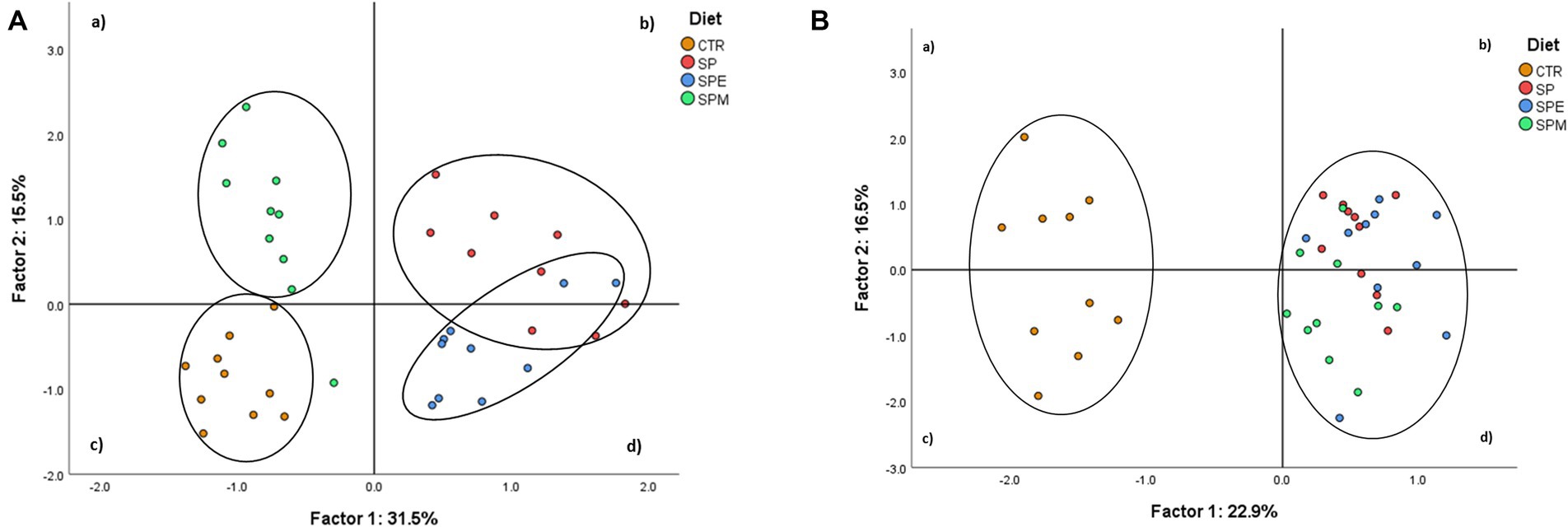 Figura 2. Análisis de componentes principales del perfil de metabolitos plasmáticos (A) y de la composición bioquímica hepática (B) de los tratamientos dietéticos. CTR, dieta a base de harina de maíz y soja; SP, 15% Espirulina; SPE, 15% de Espirulina extruida; SPM, 15% Espirulina + 0,21% mezcla de enzimas (0,20% pancreatina porcina +0,01% lisozima).
Figura 2. Análisis de componentes principales del perfil de metabolitos plasmáticos (A) y de la composición bioquímica hepática (B) de los tratamientos dietéticos. CTR, dieta a base de harina de maíz y soja; SP, 15% Espirulina; SPE, 15% de Espirulina extruida; SPM, 15% Espirulina + 0,21% mezcla de enzimas (0,20% pancreatina porcina +0,01% lisozima).
La Figura 2A representa los tratamientos dietéticos distribuidos según la variabilidad de los datos de metabolitos plasmáticos. Los dos primeros factores cubrieron el 47,0% de la varianza total (31,5% para el Factor 1 y 15,5% para el Factor 2), lo que llevó a una clara separación de tratamientos. El grupo SPM estuvo representado mayoritariamente en el cuadrante (a), el control en el cuadrante (c) y los tratamientos SP y SPE en los cuadrantes (b) y (d). Además, en la Figura 2B se representan los tratamientos en función de la composición química hepática. Los dos primeros factores cubrieron el 39,4% de la varianza total (22,9% para el Factor 1 y 16,5% para el Factor 2) formando dos conglomerados. Estos correspondieron al control ubicado en los cuadrantes (a) y (c), y a los tratamientos que contienen espirulina ubicados en los cuadrantes (b) y (d). Las variables de metabolitos plasmáticos con mayor poder de discriminación fueron lípidos totales (0,96), colesterol LDL (0,92), colesterol (0,91), colesterol VLDL (0,83) y triacilgliceroles (0,83), para el Factor 1; y colesterol HDL (−0,76), AST (−0,59), colesterol VLDL (0,45), triacilgliceroles (0,45), urea (0,43), K+ (0,38) y proteína C reactiva (0,33), para el Factor 2 (Tabla 7). Las variables hepáticas con mayor poder discriminante fueron α-tocoferol (−0,93), 20:5n-3 (0,83), carotenoides totales (0,81), γ-tocoferol+ β-tocotrienol (−0,78), β-caroteno (0,77), clorofila-a (0,77), 22:5n-3 (0,75) y 18:3n-3 (0,74) para el factor 1; y 18:1c9 (−0,94), 20:4n-6 (0,87), 16:1c9 (−0,75), 14:1c9 (−0,74) y 18:0 (0,71) para el factor 2.
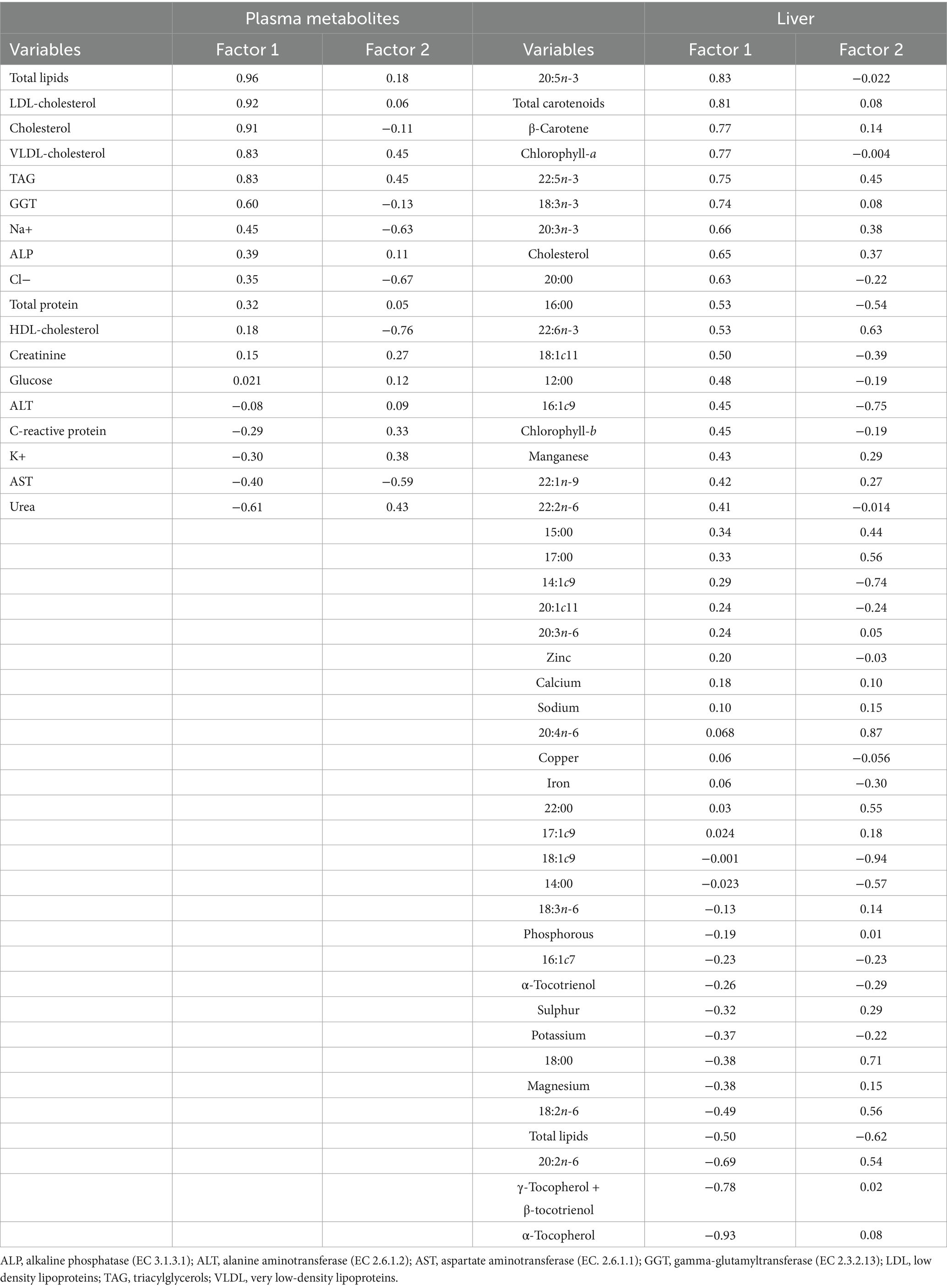 Tabla 7. Cargas de los dos primeros componentes principales de los metabolitos plasmáticos y composición química hepática.
Tabla 7. Cargas de los dos primeros componentes principales de los metabolitos plasmáticos y composición química hepática.
4 Discusión
Los datos zootécnicos de las aves aquí presentados resuenan con la literatura existente, ya que el rendimiento de los pollos de engorde está influenciado por múltiples factores que abarcan el nivel de microalgas en la dieta, la duración del período de alimentación y la eficacia de los pollos de engorde en la digestión de las paredes celulares de las microalgas. Park et al. (2) en un estudio anterior ilustraron que una modesta inclusión dietética de A. platensis (que oscila entre 0,25 y 1,0%) durante 35 días mejoró la eficiencia alimenticia en pollos de engorde. Por el contrario, nuestra investigación anterior reveló una disminución en el peso corporal y el peso corporal final, junto con un aumento en la FCR, en aves alimentadas con una dieta de A. platensis al 15%, ya sea sola o combinada con enzimas exógenas, durante 21 días (3). El esfuerzo actual, empleando el mismo nivel dietético de 15% de A. platensis, con o sin mezclas de enzimas comerciales enriquecidas con peptidasas (VemoZyme P y pancreatina porcina), subrayó un impacto perjudicial en el rendimiento del crecimiento de los pollos de engorde después de 4 semanas de alimentación, a través de la disminución del peso corporal, el peso corporal final y el ADFI, junto con el aumento de la FCR. Esta tendencia fue constante a lo largo del período experimental, destacando un impacto notable de estas dietas en el rendimiento del crecimiento de los pollos de engorde. Sin embargo, el grupo alimentado con Espirulina extruida cambió el deterioro del rendimiento del crecimiento asociado con los tratamientos alimentados con Espirulina, lo que condujo a valores superiores para el peso corporal final y el peso corporal (14 a 35 días), que fueron equivalentes a los encontrados con la dieta control. El proceso de extrusión no se comprende claramente, pero la alteración de las paredes celulares de las microalgas puede, por lo tanto, mejorar la biodisponibilidad, con la liberación de proteínas importantes, como las ficocianinas. Una mejora en la biodisponibilidad podría facilitar una hidrólisis más eficiente por parte de las enzimas digestivas endógenas de los pollos de engorde, mejorando así la absorción y la consiguiente utilización de los nutrientes, especialmente las proteínas, contribuyendo a una mejora en el rendimiento de crecimiento de los pollos de engorde. Becker (36) propuso un límite del 10% de inclusión de microalgas en las dietas de las aves de corral para evitar posibles resultados adversos, como una FCR elevada durante períodos prolongados. El rendimiento adverso de las aves con niveles más altos de espirulina podría atribuirse a la gelificación de proteínas, lo que podría impedir la absorción de aminoácidos y aumentar la viscosidad (11). Además, el robusto contenido de proteínas en la espirulina (50 a 70%) podría presentar resistencia a la acción proteolítica de las peptidasas endógenas de las aves (37). El intento de mitigar esta disminución en el rendimiento del crecimiento animal a través de la suplementación con peptidasa exógena no arrojó resultados positivos. Un estudio previo en gallos (21) también describió la ausencia de efectos sobre el rendimiento del crecimiento animal de una pancreatina comercial superdosificada (250 a 1.000 mg/kg) incorporada en una dieta estándar, aunque se detectó una mejora en la morfología intestinal. Por el contrario, los estudios en pollos de engorde mostraron una disminución en la tasa de conversión alimenticia (22) y un aumento en la digestibilidad de los nutrientes (23) cuando se alimentó con niveles similares (350 a 1,000 mg / kg) de una mezcla de enzimas súper dosificada agregada a la dieta básica. Sin embargo, ninguno de los estudios probó el efecto de la pancreatina en las microalgas dietéticas. En el presente estudio, se utilizó la doble cantidad de pancreatina (2000 mg/kg) en un intento de maximizar la hidrólisis de los compuestos de Spirulina, pero no se evaluó el impacto de dicha mezcla enzimática en otras microalgas adecuadas para pollos de engorde, lo que sería interesante para la pursue. Aunque A. platensis, al ser rica en oligosacáridos, se promociona por su potencial prebiótico que podría modular la fermentación intestinal, este efecto no compensó el impacto adverso sobre el peso corporal y la GMD en este estudio; por lo tanto, no se puede prevenir el deterioro del crecimiento (38). Esto arroja luz sobre el imperativo de una comprensión más profunda de las repercusiones dietéticas y la dinámica de utilización de nutrientes en pollos de engorde, para aprovechar mejor la destreza nutricional de A. platensis y evitar impactos perjudiciales en el crecimiento.
A. platensis a un alto nivel de incorporación dietética no promovió cambios significativos en los recuentos de células sanguíneas, lo que sugiere el mantenimiento de la homeostasis hematológica. Este resultado no está alineado con los hallazgos de Sugiharto et al. (39) que reportaron una reducción significativa en los valores de hemoglobina, eritrocitos y hematocrito en aves alimentadas con 1% de A. platensis durante 35 días. Varios estudios reportaron un equilibrio de la respuesta inmune inducida por A. platensis (39-41). En un informe anterior de este equipo de investigación (Lopes et al., datos no publicados) utilizando el 15% de A. platensis en la dieta de pollos de engorde, observamos que, si bien el recuento de glóbulos blancos se mantuvo sin cambios en las dietas, se representaron distintas variaciones en los recuentos de granulocitos y linfocitos. El cambio en el recuento de monocitos reflejó uno de los linfocitos. Los glóbulos blancos son una parte del sistema inmunitario que protege al cuerpo de las infecciones. Estas células circulan a través del torrente sanguíneo y los tejidos para responder a lesiones o enfermedades atacando a cualquier organismo desconocido que ingrese al cuerpo. Las anomalías en su recuento apuntan hacia la aparición de enfermedades infecciosas o inflamatorias, leucemia, linfoma y trastornos de la médula ósea (42). Más recientemente, se ha prestado mucha atención a los efectos inmunomoduladores de la espirulina cuando se aplica como aditivo para piensos avícolas, destacando su potencial para aumentar la resistencia a las enfermedades y mejorar las tasas de crecimiento, especialmente en condiciones de estrés (40).
El estado clínico y la condición física del animal pueden ser fácilmente percibidos por parámetros bioquímicos sistémicos (43). En este estudio, observamos que la inclusión de A. platensis en la dieta a un nivel alto aumentó los niveles de glucosa en aves alimentadas con la mezcla de enzimas en relación con la dieta de referencia. Lo más probable es que este hallazgo esté asociado con la composición rica en carbohidratos de A. platensis (44, 45). Curiosamente, mientras que los niveles de creatinina mejoraron con la espirulina extruida o combinada con enzimas, se observó una reducción significativa de la urea en las aves alimentadas con A. platensis, individualmente o extruida en relación con los otros tratamientos. Esto puede sugerir que A. platensis no puso en peligro la función renal. Además, la reducción de los niveles de urea en las aves alimentadas con la microalga podría ser consecuencia de su menor digestibilidad proteica. A menudo, los electrolitos comunes como el sodio y el potasio se asocian con trastornos de insuficiencia renal (46). El riñón desempeña un papel fundamental en el mantenimiento del equilibrio y el transporte de cloruro del cuerpo, que está estrechamente asociado con el transporte de sodio (47). A pesar de ser estadísticamente significativas, las variaciones observadas en los valores plasmáticos de sodio y cloruro, que fueron consistentemente reducidos por A. platensis en combinación con una mezcla de enzimas, no son relevantes desde un punto de vista fisiológico. De acuerdo con estos hallazgos, el elemento potasio no varió entre los tratamientos dietéticos. En cuanto a la evaluación de la función hepática a excepción de la ALT, que no mostró cambios entre los tratamientos experimentales, las variaciones observadas para las actividades enzimáticas de AST, ALP y GGT se mantuvieron dentro de las cifras de referencia para las aves (33, 48, 49). Este hecho nos permite concluir que la función hepática no se altera aunque se detecten pequeños cambios en ciertas aminotransferasas. Por lo tanto, al combinar datos hepáticos y renales, la incorporación de A. platensis en las dietas no impuso ningún peligro toxicológico para las aves, lo que subraya el potencial de esta microalga como ingrediente dietético para la producción de pollos de engorde.
El perfil lipídico plasmático estuvo influenciado en gran medida por las dietas. Nuestro estudio no apoya las propiedades hipocolesterolistas y lipídicas de A. platensis cuando se administran individualmente, como se informó previamente para otras especies de microalgas (50). Se encontró un patrón consistente de aumento del colesterol total, colesterol LDL, colesterol VLDL, triacilgliceroles y lípidos totales cuando se administró espirulina como suplemento individual. Hasta donde sabemos, este aumento lipémico se debe a la adición de espirulina y puede ser responsable de una mayor absorción de grasa en el tracto intestinal (50). No obstante, para la mayoría de estos parámetros lipídicos se detectó un efecto hipolipidémico contrapesado para la combinación de pancreatina con lisozima alcanzando niveles cercanos a los observados en las aves control. Las proteínas totales en plasma se mantuvieron sin cambios en las dietas experimentales, incluso si se esperaba un aumento en los pollos de engorde alimentados con dietas de A. platensis. La espirulina es una rica fuente de proteínas con niveles de proteínas que varían entre el 50 y el 70% (37), lo que convierte a esta microalga en una alternativa putativa a las fuentes de proteínas convencionales, como la harina de soja tradicional (51-53). Además, las diferencias estadísticamente significativas observadas para la proteína de fase aguda, proteína C reactiva, carecen de significado fisiológico porque los valores medios encontrados entre los tratamientos dietéticos fueron similares y seguidos de un error estándar residual. Los impactos de la aplicación de A. platensis dietética a un nivel de incorporación superior al 10%, como el 15% aquí utilizado, son escasos. El origen de las discrepancias encontradas entre nuestro estudio y la literatura son factores clave determinantes, incluyendo la dosis y la fuente de la microalga, la extensión del ensayo experimental y las condiciones específicas. Además, los principales constituyentes de la biomasa de microalgas dependen en gran medida de la cepa de microalgas, la ubicación geográfica, la época del año de cosecha y las condiciones de cultivo (5, 28).
El hígado, al ser el principal tejido lipogénico de las aves de corral, es esencial para dar forma al perfil de ácidos grasos de los pollos de engorde (54). Nuestros datos experimentales delinean varias interacciones matizadas entre la inclusión dietética de A. platensis y los perfiles lipídicos hepáticos en pollos de engorde. La alteración en los lípidos totales y el colesterol observada en este estudio se alinea con la literatura existente en la que se ha demostrado que la suplementación dietética con microalgas como la espirulina modula el metabolismo de los lípidos en pollos de engorde, lo que a menudo conduce a una reducción de los lípidos totales (40). El aumento del colesterol en los tratamientos alimentados con dietas a base de espirulina corrobora los hallazgos de otros estudios (3, 55). La suma sin cambios de AGS en todos los tratamientos dietéticos, a pesar del aumento en ciertos AGS individuales, es un resultado intrigante. Este hallazgo es algo divergente de la literatura en la que la suplementación con microalgas se ha asociado con una disminución de los AGS debido a su perfil rico en AGPI (56). La elevación de MUFA en los tratamientos alimentados con espirulina con una mezcla de enzimas se alinea con los hallazgos en los que la suplementación con microalgas mejoró la composición de MUFA, particularmente ácido oleico (18:1 c9) (57). Por otro lado, la disminución de los AGPI y de los AGPI n-6 en algunos tratamientos es contraria a nuestras expectativas, dado el rico perfil de AGPI de la Espirulina. El incremento de los AGPI n-3 y la reducción de la relación n-6/n-3, particularmente en la dieta SPE, es un hallazgo prometedor, ya que una relación n-6/n-3 más baja a menudo se considera beneficiosa para la salud (58). Este enriquecimiento en AGPI n-3, junto con una relación equilibrada n-6/n-3, aumenta potencialmente el mérito nutricional del hígado de pollo de engorde, lo que puede traducirse en un perfil de carne más saludable y propicio para el consumo humano. Estos hallazgos se hacen eco de la compleja interacción entre los componentes de la dieta y el metabolismo de los lípidos hepáticos, lo que subraya la necesidad de un equilibrio meticuloso para optimizar la salud de los animales y la carne resultante.
El aumento de todos los ácidos grasos n-3 individuales por las dietas basadas en espirulina es un hallazgo sustancial y está respaldado por la literatura (56). Los impactos diferenciales observados entre la espirulina extruida y las dietas suplementadas con enzimas resaltan la complejidad de las interacciones entre el procesamiento de microalgas, la suplementación enzimática y la biodisponibilidad de nutrientes. Se justifica una mayor exploración en este campo para dilucidar los mecanismos que sustentan estas observaciones.
Nuestro examen abarcó además la evaluación del impacto de las dietas de A. platensis, con o sin suplementación enzimática, sobre los perfiles hepáticos de tocoferol y pigmentos. Una gran cantidad de componentes bioactivos que se originan en las algas, que abarcan antioxidantes, pigmentos, vitaminas y polisacáridos, son elogiados por sus atributos beneficiosos para la salud tanto en animales como en humanos. La espirulina, en particular, se erige como una fuente auspiciosa de vitaminas con potentes capacidades antioxidantes, conocidas por mitigar la inflamación. La vitamina E, un nutriente liposoluble presente en varios alimentos, funciona como un antioxidante celular, fortaleciendo las células contra el ataque de los radicales libres. Si bien el régimen dietético no moduló significativamente los niveles de compuestos de vitamina E, el α-tocoferol emergió como el homólogo predominante de la vitamina E en todos los tratamientos experimentales, un hallazgo que está en armonía con la composición de la dieta. Por el contrario, el γ-tocoferol junto con el β-tocotrienol se discernieron como constituyentes menores. En marcado contraste, el β-caroteno, un precursor de la vitamina A, junto con la clorofila-a y las clorofilas totales, fue testigo de una marcada mejora en las dietas de A. platensis, alineándose perfectamente con su composición nutricional innata (2, 60). Esto revela la biodisponibilidad de estos elementos dietéticos y atestigua el aumento nutricional otorgado por la inclusión de Spirulina. Las clorofilas y los carotenoides, pigmentos lipofílicos naturales, son fundamentales para mantener la homeostasis antioxidante (61), desempeñando así un papel fundamental tanto en la salud animal como en la humana (62). Las elevaciones en los niveles de clorofila y carotenoides en el hígado, como se observa en este estudio, reflejan el robusto potencial antioxidante promovido por la espirulina, capaz de fomentar un entorno antioxidante propicio en los pollos de engorde. Esta potencialidad antioxidante protege el tejido hepático del estrés oxidativo, que es indispensable para mantener la salud y la función hepática. Además, los niveles inalterados de tocoferol en todos los tratamientos subrayan la estabilidad y biodisponibilidad de este antioxidante vital, incluso frente a modificaciones dietéticas. Estos hallazgos acentúan colectivamente la destreza nutricional y de promoción de la salud de la espirulina y, por extensión, subrayan su potencial como ingrediente alimenticio viable y rico en nutrientes en las dietas de las aves de corral.
La espirulina, al igual que las microalgas en general, incorpora una rica cantidad de minerales fundamentales (63), lo que las convierte en complementos dietéticos adecuados para los animales (64). En particular, los minerales como el cobre, el yodo, el hierro, el potasio y el zinc, que orquestan actividades fisiológicas clave como el metabolismo celular (por ejemplo, el yodo) y la regulación osmótica (por ejemplo, el sodio), son abundantes en las microalgas (65). A la luz de esto, nuestro estudio se aventuró a discernir el impacto de las dietas de A. platensis en el terreno mineral hepático de pollos de engorde. Como se muestra en este estudio, la concurrencia general de macrominerales mantuvo un nivel de constancia en todo el espectro dietético de la espirulina, aunque el sodio y el magnesio manifestaron trayectorias divergentes. El reservorio hepático de sodio experimentó un repunte en las aves alimentadas con dietas de espirulina, lo que marca un avance positivo. El sodio, un nutriente indispensable, desempeña un papel importante en la homeostasis celular, el equilibrio de líquidos y electrolitos y la modulación de la presión arterial. Además, es fundamental para orquestar la excitabilidad de los músculos y las células nerviosas, además de facilitar el tránsito de nutrientes y sustratos a través de las membranas plasmáticas (66). Por el contrario, a pesar de una disminución en los niveles de magnesio en aves alimentadas con dietas de A. platensis, la disminución fue marginal. El magnesio, un mineral versátil, sirve como cofactor para una extensión de más de 300 enzimas, modulando así actividades fisiológicas cruciales como la contracción muscular, la conducción neuromuscular, el control glucémico, la contracción miocárdica y la regulación de la presión arterial (67). Por otro lado, la infusión de A. platensis con suplementación enzimática no provocó ninguna alteración perceptible en los niveles de microminerales.
Los microminerales, como el zinc, el manganeso y el cobre, se estiman como cofactores por excelencia de las enzimas antioxidantes como la superóxido dismutasa (68) y constituyen la vanguardia de la defensa antioxidante. El hierro, una entidad metálica esencial para la vida, está implicado en la generación de especies de oxígeno deletéreas a través de la reacción de Fenton, que culmina en la génesis del potente radical hidroxilo. El transporte, la utilización y el almacenamiento del hierro se llevan a cabo meticulosamente mediante proteínas especializadas como la transferrina, la ferritina y las hemoproteínas (69, 70). El aumento del manganeso hepático acentúa el papel potencial de la espirulina en la dieta para mejorar la biodisponibilidad o la retención de ciertos oligoelementos, lo que posiblemente fortalezca el estado nutricional mineral de los pollos de engorde. En conjunto, los hallazgos de microminerales se hacen eco del mantenimiento del equilibrio redox dentro de las aves, lo que podría respaldar un estado de homeostasis antioxidante. Esta narrativa encaja con la narrativa general de que las microalgas, especialmente la espirulina, son una fuente nutritiva de minerales esenciales, lo que potencialmente fortalece el estado nutricional mineral de los pollos de engorde y, por extensión, muestra su promesa como un suplemento dietético sólido en la nutrición de las aves de corral.
Los resultados del ACP se alinean con estudios previos (33, 71, 72), demostrando efectivamente los distintos impactos de las dietas experimentales sobre los metabolitos plasmáticos y las variables hepáticas en pollos de engorde. El PCA reveló cuatro grupos distintos correspondientes a cada grupo de tratamiento, con una superposición notable entre los tratamientos SP y SPE, pero una clara demarcación de los tratamientos SPM y control. Este patrón sugiere una influencia significativa de la mezcla de enzimas en variables discriminantes clave como los lípidos totales, los triacilgliceroles y el colesterol. Sin embargo, estos hallazgos de ACP no reflejan completamente los datos presentados en la Tabla 3, lo que indica una interacción compleja entre la composición de la dieta y los resultados metabólicos. El impacto diferencial de la suplementación enzimática en dietas que contienen microalgas, observado en este estudio, se hace eco de hallazgos previos en los que la inclusión de un 0,001% de carbohidrasas, ya sea una humedad de cuatro enzimas (exo-β-glucosaminidasa, alginato liasa, peptidoglicano ácido N-acetilmurámico desacetilasa y lisozima) (NzyTech, Lisboa, Portugal) o un alimento PL25 ulvan liasa (NzyTech, Lisboa, Portugal) combinado con un 10% de Chlorella vulgaris (33) y un 15% de Ulva lactuca ( 71), respectivamente, alteraron la dinámica nutricional en las dietas de los pollos de engorde. Además, el análisis de variables hepáticas del PCA distinguió los tratamientos de control de los alimentados con microalgas, un hallazgo consistente con Costa et al. (72), quienes reportaron una diferenciación similar en dietas suplementadas con 15% de Laminaria digitata y 0.001% de alginato liasa. El poder distintivo de la espirulina se atribuye a los cambios en variables discriminantes clave como ciertos AGPI n-3 (18:3 n-3, 20:5 n-3 y 22:5 n-3), α y γ-tocoferoles, β-caroteno, clorofila-a y carotenoides totales. Estas observaciones corroboran los hallazgos presentados en las Tablas 4 y 5, que mostraron un aumento de estos ácidos grasos y pigmentos y una disminución de los niveles de vitamina E con la inclusión de microalgas. Aunque Altmann et al. (73) también mostraron un claro impacto de la Espirulina para los AGPI n-3, los resultados obtenidos fueron opuestos en relación con los presentados en este estudio, lo que podría atribuirse a la diferente composición nutricional de las microalgas. En conjunto, estos resultados subrayan la capacidad de la espirulina, particularmente cuando se combina con suplementos enzimáticos, para modular los componentes lipídicos del plasma. Además, los datos demuestran el efecto de la microalga por sí sola en la mejora del metabolismo y la composición de los lípidos hepáticos, principalmente a través de la acumulación de AGPI n-3 beneficiosos para la salud y pigmentos antioxidantes. Una mayor difusión de los resultados actuales a través de las redes sociales, evidenciando los beneficios y las limitaciones de la incorporación de Spirulina en la dieta de los pollos de engorde al público en general, representa una perspectiva prometedora para la difusión del conocimiento científico (74).
5 Conclusión
Este estudio dilucida el impacto multifacético de la inclusión dietética del 15% de Spirulina en pollos de engorde, destacando las alteraciones en la capacidad antioxidante sistémica, la composición de ácidos grasos hepáticos y el rendimiento del crecimiento. Específicamente, las dietas extruidas y suplementadas con enzimas de Spirulina elevaron significativamente los niveles hepáticos de ácidos grasos n-3, lo que fomentó un cambio favorable en la proporción de AGPI n-6 / n-3, lo que es indicativo de posibles beneficios para la salud. Este nivel de inclusión de Spirulina también condujo a una reducción de los lípidos hepáticos totales y a un aumento de los carotenoides antioxidantes en el hígado del pollo de engorde. Sin embargo, el colesterol hepático se incrementó y el α-tocoferol se redujo con la adición de Espirulina, que no se revirtió ni con el pretratamiento de extrusión ni con la suplementación enzimática. La incorporación de microalgas también fue responsable de un aumento del colesterol total, LDL-colesterol y lípidos totales en sangre, aunque la mezcla enzimática contrarrestó este efecto. La microalga tuvo efectos adversos sobre el rendimiento del crecimiento, que no fueron mitigados por la suplementación enzimática, sino parcialmente revertidos por la extrusión de microalgas. La extrusión de la espirulina pareció mejorar la biodisponibilidad de algunos nutrientes, lo que significa una posible vía de procesamiento para maximizar los beneficios nutricionales de la espirulina. Estos hallazgos subrayan una compleja interacción entre la inclusión de espirulina en la dieta, su procesamiento, la suplementación enzimática y los efectos resultantes sobre la salud de los pollos de engorde y el rendimiento del crecimiento.
Los estudios futuros deben intentar delinear los niveles óptimos de inclusión y los métodos de procesamiento de la espirulina para mitigar los efectos adversos sobre el colesterol y el α-tocoferol y, por lo tanto, maximizar los beneficios para la salud. Además, los mecanismos precisos que subyacen a los efectos observados, en particular la interacción entre el procesamiento de la espirulina, la suplementación enzimática y la biodisponibilidad de nutrientes, justifican una investigación en profundidad. El pretratamiento por extrusión de microalgas muestra potencial para su aplicación a escala industrial. Por lo tanto, ampliar la investigación para incluir evaluaciones de los efectos a largo plazo de la inclusión de espirulina en la dieta y explorar diferentes niveles de suplementación enzimática combinada con algas preextruidas puede proporcionar información más matizada sobre el aprovechamiento de los beneficios potenciales de la espirulina en las dietas de las aves de corral.
Declaración de disponibilidad de datos
Los datos brutos que respaldan las conclusiones de este artículo serán puestos a disposición por los autores, sin reservas indebidas.
Declaración ética
La investigación cumplió estrictamente con el marco normativo estipulado por la Unión Europea (Directiva 2010/63/UE) y obtuvo el respaldo necesario del Comité de Revisión Ética de CIISA/FMV, el Comité de Bienestar Animal de la Autoridad Veterinaria Nacional (Direção Geral de Alimentação e Veterinária, Portugal) y ORBEA/ISA (código de aprobación: 0421/000/000/2022).
Contribuciones de los autores
MS: Redacción – revisión y edición, Investigación, Análisis formal, Metodología. CA: Redacción – revisión y edición, Redacción – borrador original. MC: Redacción – revisión y edición, Redacción – borrador original, Validación, Investigación, Análisis formal, Metodología. RP: Redacción – revisión y edición, Investigación, Metodología. PL: Redacción – revisión y edición, Redacción – borrador original. JPe: Redacción – revisión y edición, Investigación, Metodología. JT: Escritura – revisión y edición, Investigación. AM: Escritura – revisión y edición, Investigación. MM: Redacción – revisión y edición, Investigación, Metodología. BT: Escritura – revisión y edición, Investigación. DC: Escritura – revisión y edición, Investigación, Metodología. CM: Redacción – revisión y edición, Investigación, Metodología. JF: Escritura – revisión y edición, Investigación. ML: Escritura – revisión y edición, Recursos. JPr: Redacción – revisión y edición, Redacción – borrador original, Validación, Supervisión, Recursos, Administración de proyectos, Adquisición de fondos, Conceptualización.
Financiación
El/los autor/es declaran/n que se recibió apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue financiada por subvenciones otorgadas por la Fundación para la Ciencia y la Tecnología (FCT, Lisboa, Portugal) a CIISA (UIDB/00276/2020), AL4AnimalS (LA/P/0059/2020) y LEAF (UIDB/04129/2020). El Programa de Estímulo de Empleo Científico de la FCT proporcionó asistencia financiera adicional a PL (DL57/2016/CP1438/CT0007), una beca postdoctoral a JP (SFRH/BPD/116816/2016), una beca de doctorado a MS (UI/BD/153071/2022) y una beca de doctorado a AM (2022.11690.BD). El proyecto también recibió financiación del proyecto Portugal 2020 (P2020/17/SI/70114/2019) y un contrato de investigación asociado para MC.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Abd El-Hady, A, y El-Ghalid, O. Spirulina platensis algae (Spa): un nuevo aditivo para piensos avícolas. Efecto de la suplementación con Spa en dietas de pollos de engorde sobre el rendimiento productivo, el perfil lipídico y el metabolismo calcio-fósforo. Worlds Poult Sci J. (2018) 7498:1–7.
2. Park, J, Lee, S, y Kim, I-S. Efecto de la espirulina (Arthrospira) platensis en la dieta sobre el rendimiento del crecimiento, la actividad enzimática antioxidante, la digestibilidad de los nutrientes, la microflora cecal, la emisión de gases nocivos de excretas y la calidad de la carne de pechuga de pollos de engorde. Poult Sci. (2018) 97:2451–9. doi: 10.3382/ps/pey093
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Pestana, JM, Puerta, B, Santos, H, Madeira, MS, Alfaia, CM, Lopes, PA, et al. Impacto de la incorporación dietética de Spirulina (Arthrospira platensis) y enzimas exógenas en el rendimiento de los pollos de engorde, las características de la canal y la calidad de la carne. Poult Sci. (2020) 99:2519–32. doi: 10.1016/j.psj.2019.11.069
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. Bortolini, DG, Maciel, GM, Fernandes, IAA, Pedro, AC, Rubio, FTV, Branco, IG, et al. Propiedades funcionales de los compuestos bioactivos de Spirulina Spp.: Estado actual y tendencias futuras. Químico de los alimentos. (2022) 5:100134. doi: 10.1016/j.fochms.2022.100134
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Tounsi, L, Hentati, F, Ben Hlima, H, Barkallah, M, Smaoui, S, Fendri, I, et al. Microalgas como materia prima para polisacáridos bioactivos. Int J Biol Macromol. (2022) 221:1238–50. doi: 10.1016/j.ijbiomac.2022.08.206
Resumen de PubMed | Texto completo de Crossref | Google Académico
6. Grover, P, Bhatnagar, A, Kumari, N, Narayan Bhatt, A, Kumar Nishad, D, y Purkayastha, J. C-Ficocianina, una nueva proteína de Spirulina platensis, estudios in vivo de toxicidad, antioxidantes e inmunomoduladores. Saudi J Biol Sci. (2021) 28:1853–9. doi: 10.1016/j.sjbs.2020.12.037
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Pilotto, MR, Milanez, S, Moreira, RT, Rosa, RD, y Perazzolo, LM. Posibles efectos inmunomoduladores y protectores del suplemento dietético a base de Arthrospira sobre las defensas inmunitarias intestinales de los camarones. Pescados y mariscos Immunol. (2019) 88:47–52. doi: 10.1016/j.fsi.2019.02.062
Resumen de PubMed | Texto completo de Crossref | Google Académico
8. Cohen, Z . Los productos químicos de la espirulina en: A Vonshak editor. Spirulina platensis Arthrospira: Fisiología, Biología Celular y Biotecnología. Londres: Taylor & Francis (1997). 175–204.
9. Böcker, L, Hostettler, T, Diener, M, Eder, S, Demuth, T, Adamcik, J, et al. Cambios funcionales y estructurales resueltos en el tiempo y la temperatura de la ficocianina extraída de Arthrospira platensis/Spirulina. Químico de los alimentos. (2020) 316:126374. doi: 10.1016/j.foodchem.2020.126374
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Coelho, D, Lopes, PA, Cardoso, V, Ponte, P, Brás, J, Madeira, MS, et al. Una mezcla constituida por dos enzimas para mejorar la degradación de la pared celular de la microalga Arthrospira platensis para dietas monogástricas. J Anim Physiol Anim Nutr. (2020) 104:310–21. doi: 10.1111/jpn.13239
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. Evans, AM, Smith, DL y Moritz, JS. Efectos de la incorporación de algas en las formulaciones de la dieta iniciadora de pollos de engorde sobre la digestibilidad de los nutrientes y el rendimiento de las aves de 3 a 21 D. J Appl Poult Res. (2015) 24:206–14. doi: 10.3382/japr/pfv027
12. Costa, MM, Spínola, MP, y Prates, JAM. Combinación de pretratamientos mecánicos/físicos con tripsina o pancreatina sobre la degradación de la proteína Arthrospira platensis. Agricultura. (2023) 13:198. doi: 10.3390/agriculture13010198
13. Spínola, MP, Costa, MM, y Prates, JA. Estudios sobre el impacto de pretratamientos seleccionados en la solubilidad proteica de la microalga Arthrospira platensis. Agricultura. (2023) 13:221. doi: 10.3390/agriculture13010221
14. Shirazi, HM, Karimi-Sabet, J y Ghotbi, C. Producción de biodiésel a partir de materia prima de microalgas Spirulina mediante transesterificación directa cerca de la condición de metanol supercrítico. Bioresour Technol. (2017) 239:378–86. doi: 10.1016/j.biortech.2017.04.073
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Alagawany, M, Elnesr, SS, y Farag, MR. El papel de las enzimas exógenas en la promoción del crecimiento y la mejora de la digestibilidad de los nutrientes en las aves de corral. Irán J Vet Res. (2018) 19:157–64.
16. Singh, AK, Tiwari, UP, Berrocoso, JD, Dersjant-Li, Y, Awati, A y Jha, R. Efectos de una combinación de xilanasa, amilasa y proteasa, y probióticos sobre los principales nutrientes, incluidos los aminoácidos y la utilización de polisacáridos sin almidón en pollos de engorde alimentados con diferentes niveles de fibras. Poult Sci. (2019) 98:5571–81. doi: 10.3382/ps/pez310
Resumen de PubMed | Texto completo de Crossref | Google Académico
17. Bedford, MR, y Cowieson, AJ. Enzimas exógenas y sus efectos sobre la microbiología intestinal. Anim Feed Sci Technol. (2012) 173:76–85. doi: 10.1016/j.anifeedsci.2011.12.018
18. El-Hady, AMA, Elghalid, OA, Elnaggar, AS, y El-khalek, EA. Evaluación del rendimiento del crecimiento y del estado fisiológico de la suplementación con algas Spirulina platensis en la dieta de pollos de engorde. Livest Sci. (2022) 263:105009. doi: 10.1016/j.livsci.2022.105009.19
19. Madeira, MSMS, Lopes, PAAB, Martins, CF, Assunção, JMP, Alfaia, CMRPM, Pinto, RMA, et al. La dieta Arthrospira platensis mejora el potencial antioxidante sistémico y cambia los lípidos plasmáticos sin afectar las vías metabólicas hepáticas relacionadas en lechones posdestetados. BMC Vet Res. (2021) 17:158. doi: 10.1186/s12917-021-02869-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
20. Martins, CF, Pestana, JM, Alfaia, CM, Costa, M, Ribeiro, DM, Coelho, D, et al. Efectos de Chlorella vulgaris como ingrediente alimentario sobre la calidad y el valor nutricional de la carne de lechones destetados. Seguridad de alimentos. (2021) 10:1155. doi: 10.3390/foods10061155
Resumen de PubMed | Texto completo de Crossref | Google Académico
21. Asare, E, Yang, Z, Yang, H y Wang, Z. Evaluación de la pancreatina dietética como enzima exógena sobre el rendimiento del crecimiento, la expresión génica, las respuestas inmunológicas, las inmunoglobinas séricas y la morfología intestinal en gallos. J Appl Anim Res. (2022) 50:61–8. doi: 10.1080/09712119.2021.2019044
22. Bromfield, JI, Hoffman, LC, Horyanto, D, y Soumeh, EA. Mejora del rendimiento del crecimiento, el desarrollo de órganos, la calidad de la carne y la mineralización ósea de pollos de engorde a través de la superdosificación multienzimática en dietas de energía reducida. Animales. (2021) 11:2791. doi: 10.3390/ani11102791
Resumen de PubMed | Texto completo de Crossref | Google Académico
23. Madigan-Stretton, J, Mikkelsen, D, y Soumeh, EA. Superdosis multienzimática en dietas de pollos de engorde: las implicaciones para la morfología intestinal, el perfil microbiano, la digestibilidad de los nutrientes y la mineralización ósea. Animales. (2020) 11:1. doi: 10.3390/ani11010001
Resumen de PubMed | Texto completo de Crossref | Google Académico
24. AOAC . Métodos oficiales de análisis. 17ª ed. Washington D.C., EE.UU.: Asociación de Químicos Analíticos Oficiales (2000).
25. Sukhija, P, y Palmquist, D. Método rápido para la determinación de la composición total de ácidos grasos de piensos y heces. J Agric Food Chem. (1987) 36:1202–6. doi: 10.1021/jf00084a019
26. Alfaia, CM, Pestana, JM, Rodrigues, M, Coelho, D, Aires, MJ, Ribeiro, DM, et al. Influencia de la Chlorella vulgaris en la dieta y las enzimas activas en carbohidratos en el rendimiento del crecimiento, la calidad de la carne y la composición lipídica de pollos de engorde. Poult Sci. (2021) 100:926–37. doi: 10.1016/j.psj.2020.11.034
27. Prates, JA, Quaresma, MA, Bessa, RJ, Fontes, CMG, y Alfaia, CMP. Cuantificación simultánea por Hplc de colesterol total, tocoferoles y β-caroteno en ternera Barrosã-Pdo. Químico de los alimentos. (2006) 94:469–77. doi: 10.1016/j.foodchem.2005.01.021
28. Hynstova, V, Sterbova, D, Klejdus, B, Hedbavny, J, Huska, D y Adam, V. Separación, identificación y cuantificación de carotenoides y clorofilas en suplementos dietéticos que contienen Chlorella vulgaris y Spirulina platensis mediante cromatografía en capa fina de alta resolución. J Pharm Biomed Anal. (2018) 148:108–18. doi: 10.1016/j.jpba.2017.09.018
Resumen de PubMed | Texto completo de Crossref | Google Académico
29. Ribeiro, DM, Scanlon, T, Kilminster, T, Martins, CF, Greeff, J, Milton, J, et al. Perfil mineral de tejidos musculares y hepáticos de corderos merino australiano, Damara y Dorper: efecto de la pérdida de peso. J Anim Physiol Anim Nutr. (2020) 104:823–30. doi: 10.1111/jpn.13339
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Friedewald, WT, Levy, RI, y Fredrickson, DS. Estimación de la concentración de colesterol de lipoproteínas de baja densidad en plasma, sin el uso de la ultracentrífuga preparativa. Clin Chem. (1972) 18:499–502. doi: 10.1093/clinchem/18.6.499
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Covaci, A, Voorspoels, S, Thomsen, C, van Bavel, B, y Neels, H. Evaluación de lípidos totales utilizando métodos enzimáticos para la normalización de los niveles de contaminantes orgánicos persistentes en suero. Sci Total Environ. (2006) 366:361–6. doi: 10.1016/j.scitotenv.2006.03.006
Resumen de PubMed | Texto completo de Crossref | Google Académico
32. Folch, J, Lees, M y Sloane Stanley, GH. Un método sencillo para el aislamiento y purificación de lípidos totales a partir de tejidos animales. J Biol Chem. (1957) 226:497–509. doi: 10.1016/S0021-9258(18)64849-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Coelho, DFM, Alfaia, CMRPM, Assunção, JMP, Costa, M, Pinto, RMA, de Andrade Fontes, CMG, et al. Impacto de la incorporación de Chlorella vulgaris en la dieta y enzimas activas de carbohidratos sobre los metabolitos plasmáticos y la composición lipídica hepática de pollos de engorde. BMC Vet Res. (2021) 17:229. doi: 10.1186/s12917-021-02932-8
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Delgado, I, Ventura, M, Gueifão, S, Coelho, I, Nascimento, AC, Silva, JAL, et al. 12ª edición especial del IFDC 2017 – Contenido de yodo, selenio y hierro en los alimentos clave portugueses tal como se consumen. J Compost de alimentos anal. (2019) 79:39–46. doi: 10.1016/j.jfca.2019.03.004
35. Instituto SAS . Guía del usuario de Sas/stat 9.1, vol. 1–7. Cary, Carolina del Norte: Instituto SAS (2004).
36. Becker, EW . Microalgas para la nutrición humana y animal. Manual Microal Cul. (2013):461–503. doi: 10.1002/9781118567166.ch25
37. Soni, RA, Sudhakar, K, y Rana, RS. Espirulina: desde el crecimiento hasta el producto nutricional: una revisión. Tendencias Food Sci Technol. (2017) 69:157–71. doi: 10.1016/j.tifs.2017.09.010
38. Cai, B, Yi, X, Han, Q, Pan, J, Chen, H, Sun, H, et al. Caracterización estructural de oligosacáridos de Spirulina platensis y su efecto sobre la microbiota fecal in vitro. Ciencia Alimentaria Bienestar Humano. (2022) 11:109–18. doi: 10.1016/j.fshw.2021.07.012
39. Sugiharto, S, Yudiarti, T, Isroli, I y Widiastuti, E. Efecto de la duración de la alimentación de Spirulina platensis sobre el rendimiento del crecimiento, los parámetros hematológicos, la población microbiana intestinal y las características de la canal de los pollos de engorde. S Afr J Anim Sci. (2018) 48:98–107. doi: 10.4314/sajas.v48i1.12
40. El-Shall, NA, Jiang, S, Farag, MR, Azzam, M, Al-Abdullatif, AA, Alhotan, R, et al. Potencial de Spirulina platensis como suplemento alimenticio para aves de corral para mejorar el rendimiento del crecimiento y la modulación inmunológica. Frente Immunol. (2023) 14:1072787. doi: 10.3389/fimmu.2023.1072787
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Mirzaie, S, Zirak-Khattab, F, Hosseini, SA y Donyaei-Darian, H. Efectos de la espirulina dietética sobre el estado antioxidante, el perfil lipídico, la respuesta inmunitaria y las características de rendimiento de pollos de engorde criados a alta temperatura ambiente. Australas Asiáticas J Anim Sci. (2018) 31:556–63. doi: 10.5713/ajas.17.0483
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. King, W, Toler, K y Woodell-May, J. Papel de los glóbulos blancos en las terapias autólogas basadas en sangre y médula ósea. Biomed Res Int. (2018) 2018:6510842–8. doi: 10.1155/2018/6510842
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Muriel, R, Schmidt, D, Calabuig, CP, Patino-Martinez, J, y Ferrer, M. Factores que afectan los parámetros bioquímicos del plasma y la condición física de los polluelos de águila pescadora (Pandion Haliaetus). J Ornithol. (2013) 154:619–32. doi: 10.1007/s10336-012-0925-0
44. Martins, CF, Ribeiro, DM, Costa, M, Coelho, D, Alfaia, CM, Lordelo, M, et al. Utilizar microalgas como recurso alimenticio sostenible para mejorar la calidad y el valor nutricional de la carne de cerdo y aves de corral. Seguridad de alimentos. (2021) 10:2933. doi: 10.3390/foods10122933
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Wu, H, Li, T, Lv, J, Chen, Z, Wu, J, Wang, N, et al. Características de crecimiento y composición bioquímica de Arthrospira platensis inducidas por deficiencia simultánea de nitrógeno y medio suplementado con agua de mar en un estanque de canalización al aire libre en invierno. Seguridad de alimentos. (2021) 10:2974. doi: 10.3390/foods10122974
Resumen de PubMed | Texto completo de Crossref | Google Académico
46. Molla, MD, Degef, M, Bekele, A, Geto, Z, Challa, F, Lejisa, T, et al. Evaluación de electrolitos séricos y prueba de función renal para la detección de la enfermedad renal crónica entre los miembros del personal del Instituto de Salud Pública de Etiopía, Addis Abeba, Etiopía. BMC Nefrol. (2020) 21:494. doi: 10.1186/s12882-020-02166-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Nagami, GT . Hipercloremia: por qué y cómo. Nefrología. (2016) 36:347–53. doi: 10.1016/j.nefro.2016.04.001
Resumen de PubMed | Texto completo de Crossref | Google Académico
48. Kaneko, JJ, Harvey, JW y Bruss, M. Bioquímica clínica de animales domésticos. 5ª ed. Cambridge, MA, EE.UU.: Academic Press (1997).
49. Kuttappan, VA, Huff, GR, Huff, WE, Hargis, BM, Apple, JK, Coon, C, et al. Comparación de los perfiles hematológicos y serológicos de pollos de engorde con grados normales y severos de rayas blancas en filetes de pechuga. Poult Sci. (2013) 92:339–45. doi: 10.3382/ps.2012-02647
Resumen de PubMed | Texto completo de Crossref | Google Académico
50. Sano, T, Kumamoto, Y, Kamiya, N, Okuda, M y Tanaka, Y. Efecto del extracto lipofílico de Chlorella vulgaris sobre la hiperlipidemia alimentaria en ratas alimentadas con colesterol. Arteria. (1988) 15:217–24.
51. Austic, RE, Mustafa, A, Jung, B, Gatrell, S y Lei, XG. Potencial y limitación de una nueva biomasa de microalgas de diatomeas desgrasadas en sustitución de la harina de soja y el maíz en dietas para pollos de engorde. J Agric Food Chem. (2013) 61:7341–8. DOI: 10.1021/JF401957Z
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Spolaore, P, Joannis-Cassan, C, Duran, E e Isambert, A. Aplicaciones comerciales de las microalgas. J Biosci Bioeng. (2006) 101:87–96. doi: 10.1263/jbb.101.87
53. Swiatkiewicz, S, Arczewska-Wlosek, A, y Józefiak, D. Aplicación de la biomasa de microalgas en la nutrición avícola. Worlds Poult Sci J. (2015) 71:663–72. doi: 10.1017/S0043933915002457
54. Hermier, D . Metabolismo y engorde de lipoproteínas en aves. J Nutr. (1997) 127:805S–8S. doi: 10.1093/jn/127.5.805S
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Abdel-Aty, E, El-Sheekh, M, Abdel Daim, M y Saber, H. Efectos de Spirulina platensis y Amphora Coffeaeformis como suplementos dietéticos sobre los parámetros bioquímicos sanguíneos, la población microbiana intestinal y el rendimiento productivo en pollos de engorde. Environ Sci Pollut Res. (2020) 28:1801–11. doi: 10.1007/s11356-020-10597-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Bonos, E, Kasapidou, E, Kargopoulos, A, Karampampas, A, Nikolakakis, I, Christaki, E, et al. Espirulina como ingrediente funcional en las dietas de pollos de engorde. S Afr J Anim Sci. (2016) 46:94–102. doi: 10.4314/sajas.v46i1.12
57. Gatrell, SK, Kim, J, Derksen, TJ, O’Neil, EV, y Lei, XG. Creación de pollo enriquecido con ácidos grasos Ω-3 utilizando biomasa de microalgas verdes desgrasadas. J Agric Food Chem. (2015) 63:9315–22. doi: 10.1021/acs.jafc.5b03137
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Simopoulos, AP . Un aumento en la proporción de ácidos grasos Omega-6/Omega-3 aumenta el riesgo de obesidad. Nutrientes. (2016) 8:128. doi: 10.3390/nu8030128
Resumen de PubMed | Texto completo de Crossref | Google Académico
59. Brigelius-Flohé, R, Kelly, FJ, Salonen, JT, Neuzil, J, Zingg, JM y Azzi, A. La perspectiva europea sobre la vitamina E: conocimientos actuales e investigaciones futuras. Am J Clin Nutr. (2002) 76:703–16. doi: 10.1093/ajcn/76.4.703
Resumen de PubMed | Texto completo de Crossref | Google Académico
60. Holman, BWB, y Malau-Aduli, OEA. Espirulina como suplemento ganadero y alimento para animales. J Anim Physiol Anim Nutr. (2013) 97:615–23. doi: 10.1111/j.1439-0396.2012.01328.x
61. Pérez-Gálvez, A, Viera, I, y Roca, M. Carotenoides y clorofilas como antioxidantes. Antioxidantes. (2020) 9:505. doi: 10.3390/antiox9060505
Resumen de PubMed | Texto completo de Crossref | Google Académico
62. Nabi, F, Arain, MA, Rajput, N, Alagawany, M, Soomro, J, Umer, M, et al. Beneficios para la salud de los carotenoides y su posible aplicación en la industria avícola: una revisión. J Anim Physiol Anim Nutr. (2020) 104:1809–18. doi: 10.1111/jpn.13375
Resumen de PubMed | Texto completo de Crossref | Google Académico
63. Christaki, E, Florou-Paneri, P, y Bonos, E. Microalgas: un ingrediente novedoso en nutrición. Int J Food Sci Nutr. (2011) 62:794–9. doi: 10.3109/09637486.2011.582460
64. Morais, T, Inácio, A, Coutinho, T, Ministro, M, Cotas, J, Pereira, L, et al. Potencial de las algas marinas en la alimentación animal: una revisión. J Mar Sci Ing. (2020) 8:559. doi: 10.3390/jmse8080559
65. Priyadarshani, I, y Rath, B. Aplicaciones comerciales e industriales de las microalgas: una revisión. J Biomasa de algas util. (2012) 3:89–100.
66. Strazzullo, P, y Leclercq, C. Sodio. Adv Nutr. (2014) 5:188–90. doi: 10.3945/an.113.005215
Resumen de PubMed | Texto completo de Crossref | Google Académico
67. Al Alawi, AM, Majoni, SW, y Falhammar, H. Magnesio y salud humana: perspectivas y direcciones de investigación. Int J Endocrinol. (2018) 2018:9041694–17. doi: 10.1155/2018/9041694
Resumen de PubMed | Texto completo de Crossref | Google Académico
68. Fukai, T, Folz, RJ, Landmesser, U, y Harrison, DG. Superóxido dismutasa extracelular y enfermedad cardiovascular. Cardiovasc Res. (2002) 55:239–49. doi: 10.1016/j.yjmcc.2009.08.010
69. Beaumont, C, Laverdant, C, Ardaillou, R, Nordmann, R, David, G, y Sureau, C. Le Métabolisme Intracellulaire du Fer. Bull Acad Natl Med. (2000) 184:313–24.
70. Comporti, M, Signorini, C, Buonocore, G, y Ciccoli, L. Liberación de hierro, estrés oxidativo y envejecimiento eritrocitario. Gratis Radic Biol Med. (2002) 32:568–76. doi: 10.1016/s0891-5849(02)00759-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
71. Alfaia, CM, Costa, MM, Pinto, RMA, Pestana, JM, Mourato, M, Carvalho, P, et al. Metabolitos plasmáticos y composición hepática de pollos de engorde en respuesta a Ulva lactuca dietética con Ulvan Liasa o una mezcla comercial de enzimas. Moléculas. (2022) 27:7425. doi: 10.3390/moléculas27217425
72. Costa, MM, Lopes, PA, Assunção, JMP, Alfaia, CMRPM, Coelho, DFM, Mourato, MP, et al. Efectos combinados de la dieta de Laminaria digitata con alginato liasa sobre los metabolitos plasmáticos y la composición hepática de lípidos, pigmentos y minerales de pollos de engorde. BMC Vet Res. (2022) 18:153. doi: 10.1186/s12917-022-03250-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
73. Altmann, BA, Wigger, R, Ciulu, M, y Mörlein, D. El efecto de los piensos proteicos alternativos de insectos o microalgas en la calidad de la carne de los pollos de engorde. J Sci Food Agric. (2020) 100:4292–302. doi: 10.1002/jsfa.10473
74. Muca, E, Buonaiuto, G, Lamanna, M, Silvestrelli, S, Ghiaccio, F, Federiconi, A, et al. Llegar a un público más amplio: el papel de Instagram en la educación y el compromiso con la nutrición de las vacas lecheras. Animales. (2023) 13:3503. doi: 10.3390/ani13223503
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: enzimas alimenticias, compuestos hepáticos, extrusión de microalgas, metabolitos plasmáticos, nutrición avícola
Cita: Spínola MP, Alfaia CM, Costa MM, Pinto RMA, Lopes PA, Pestana JM, Tavares JC, Mendes AR, Mourato MP, Tavares B, Carvalho DFP, Martins CF, Ferreira JI, Lordelo MM y Prates JAM (2024) Impacto de la dieta alta en espirulina, extruida o suplementada con enzimas, en las células sanguíneas, los metabolitos sistémicos y los perfiles lipídicos y minerales hepáticos de pollos de engorde. Frente. Vet. Sci. 11:1342310. doi: 10.3389/fvets.2024.1342310
Editado por:
Damiano Cavallini, Universidad de Bolonia, Italia
Revisado por:
Tumisang Ben Matshogo, Universidad del Noroeste, Sudáfrica
Giovanni Buonaiuto, Universidad de Bolonia, Italia
Godfrey Mhlongo, Universidad del Noroeste, Potchefstroom, Sudáfrica, en colaboración con el revisor TM
Copyright © 2024 Spínola, Alfaia, Costa, Pinto, Lopes, Pestana, Tavares, Mendes, Mourato, Tavares, Carvalho, Martins, Ferreira, Lordelo y Prates. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY). S
*Correspondencia: José A. M. Prates, japrates@fmv.ulisboa.pt
†Estos autores han contribuido igualmente a este trabajo y comparten la primera autoría
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía