
Influencia de las vacunas heterologas y homólogas, y sus componentes, en la respuesta inmune del huésped y la protección contra la paratuberculosis de la cabra experimental
Influencia de las vacunas heterologas y homólogas, y sus componentes, en la respuesta inmune del huésped y la protección contra la paratuberculosis de la cabra experimental
 Natalia Elguezabal3,
Natalia Elguezabal3,  Iker A. Sevilla3, Raquel Vallejo1,2, José Espinosa1,2, María del Carmen Ferreras1,2, Julio Benavides2 y
Iker A. Sevilla3, Raquel Vallejo1,2, José Espinosa1,2, María del Carmen Ferreras1,2, Julio Benavides2 y  Valentín Pérez1,2
Valentín Pérez1,2- 1Departamento de Sanidad Animal, Facultad de Veterinaria, Universidad de León, León, España
- 2Departamento de Sanidad Animal, Instituto de Ganadería de Montaña (CSIC-ULE), León, España
- 3Departamento de Sanidad Animal, NEIKER-Instituto Vasco de Investigación y Desarrollo Agrario, Derio, España
La vacunación contra la paratuberculosis, una enfermedad crónica de los rumiantes causada por Mycobacterium avium subsp. paratuberculosis (Mapa), se ha considerado como el método de control más eficaz. Sin embargo, la protección está incompleta y los mecanismos que operan en la respuesta de los animales a la vacunación no se entienden completamente. Por lo tanto, este estudio analizó la respuesta inmunitaria y los efectos sobre la protección contra la infección por Map, provocadas por las vacunas paratuberculosis (Silirum®) y tuberculosis (M. bovis [HIMB]) inactivadas por calor y sus componentes en un modelo experimental caprino. Cincuenta niños de cabra se dividieron en 10 grupos (n = 5) según su vacunación (Silirum®, HIMB y no vacunados), inmunización (bacterias inactivadas o adyuvantes) y/o infección. El desafío oral con Map se realizó 45 días después de la vacunación/inmunización (dpv), y los animales fueron sacrificados a 190 dpv. La respuesta inmune periférica y la proporción de subpoblaciones de linfocitos se evaluaron mensualmente mediante el ensayo de inmunoabsorción ligada a enzimas y el análisis de citometría de flujo, respectivamente. Se realizaron la respuesta inmune local, la proporción de subpoblaciones de linfocitos tisulares, la detección de mapas (reacción en cadena de la polimerasa) y el examen histológico en tejidos linfoides asociados al intestino. Todos los grupos infectados desarrollaron lesiones granulomatosas paratuberculosis a pesar de la vacunación o la inmunización. Los grupos vacunados por Silirum® y HIMB mostraron una reducción considerable de la lesión consistente con una importante respuesta inmune celular y humoral periférica. Además, se observó un menor número de granulomas en grupos inmunizados con bacterias y adyuvantes inactivados en comparación con el grupo no vacunado e infectado. Sin embargo, a pesar de no ser significativa, esta reducción fue aún mayor en los grupos inmunizados adyuvantes, que desarrollaron una lesión granulomatosa más leve sin respuestas inmunes periféricas detectables asociadas con la inmunización. No se detectaron cambios en la proporción periférica y local de los subconjuntos de linfocitos o en la respuesta inmune local en relación con la vacunación/inmunización o la infección. A pesar de que la vacunación contra la paratuberculosis y la tuberculosis mostraron una protección parcial y cruzada contra la infección por mapas, respectivamente, solo el examen histológico podría evaluar la progresión de la infección en estos animales. Además, el patrón observado en la reducción de las lesiones en grupos inmunizados adyuvantes sugiere la posible implicación de una respuesta inmune ine inespecífica que reduce el desarrollo de lesiones granulomatosas.
Introducción
La paratuberculosis es una enfermedad crónica y debilitante de los rumiantes causada por Mycobacterium avium subsp.paratuberculosis (Mapa) (1) y responsable de importantes pérdidas económicas globales estimadas en aproximadamente 12,61 millones de dólares y 364,31 millones de dólares en ganado lechero español y de la Unión Europea, respectivamente (2, 3). Map es una bacteria ubicua transmitida entre el ganado a una edad temprana a través de la vía fecal-oral, aunque las manifestaciones clínicas pueden aparecer varios años después de la infección (4). La respuesta inmune protectora inicial contra Map se ha relacionado con una fuerte respuesta inmune mediada por células caracterizada por la liberación de la citocina proinflamatoria interferón γ (IFN-γ), la formación de granulomas y la eliminación de las micobacterias (5, 6). En este sentido, los animales infectados por mapas pueden mostrar una variedad de lesiones que han demostrado estar estrechamente relacionadas con las diferentes fases de la enfermedad. Las lesiones tempranas de paratuberculosis se caracterizan por granulomas pequeños y bien delimitados, llamados formas focales o multifocales, ubicados dentro del tejido linfoide intestinal que también se pueden ver en animales adultos, donde se consideran como formas de latencia o resistencia; la interrupción de la respuesta inmune mediada por células protectoras conduce al desarrollo de lesiones difusas y a Los métodos patológicos para la caracterización de la lesión paratuberculosis se han aplicado con éxito en trabajos anteriores como un indicador fiable de la presencia de la infección por mapa y la forma de la enfermedad mostrada por los animales infectados (9-11).
El largo período de incubación y la capacidad de Map para persistir en los programas de control de obstaculizan el medio ambiente basados en medidas de gestión de la higiene o en el diagnóstico precoz y la eliminación de animales positivos (12, 13). Por esta razón, la vacunación con vacunas de muerte por calor disponibles en el mercado se ha considerado como la medida más rentable para el control de la paratuberculosis, ya que su uso reduce la incidencia de la enfermedad dentro de la manada (14,15). La vacunación conduce a una reducción tanto de la colonización de los tejidos intestinales como del número de animales clínicamente afectados, logrando una disminución de aproximadamente un 90 % en la aparición de animales que se derra el mapa (16-18).
Sin embargo, los mecanismos que podrían estar involucrados en la protección asociada con la vacunación aún no se entienden completamente (12, 13). La protección conferida por las vacunas contra la infección por mapas se ha correlacionado con una respuesta inmune temprana y fuerte mediada por células (19-21), a pesar del hecho de que hay un bajo porcentaje de fracaso de la vacunación, donde algunos animales vacunados podrían permanecer altamente infecciosos y desarrollar lesiones graves (16, 22, 23).
La mayoría de las vacunas paratuberculosis que matan por calor se basan en la cepa Map 316F, que ha demostrado una reducción de la virulencia (a Carga bacteriana de los tejidos) durante la infección experimental de los terneros con este aislado vivo, probablemente debido a los pasajes continuos in vitro (24). En este sentido, la inmunización subcutánea de ovejas con cepa Map 316F matada por el calor por sí sola no mostró una respuesta inmune mediada por células y humoral (25). La suplementación con adyuvantes de aceite mineral es un principio central para mejorar la inmunogenicidad de los antígenos y su liberación continua con el fin de promover una respuesta inmune robusta a través del aumento de una fuerte inflamación local en el lugar de la inyección (26). En un experimento anterior, se encontraron variaciones significativas en la respuesta inmune cuando se administraron por vía subcutánea vacunas para la paratuberculosa hechas con diferentes adyuvantes a ovejas (25). Por otro lado, se ha demostrado que la única administración de adyuvantes provoca una respuesta inmune inespecífica cuyo efecto protector no se ha evaluado en detalle (27-29). En el caso de las vacunas contra la paratuberculosis, el efecto que cada componente podría tener en una respuesta protectora contra la infección por Map no se ha aclarado completamente.
Además, Map comparte un alto número de antígenos comunes con micobacterias relacionadas como Mycobacterium bovis (Mbv) o Mycobacterium caprae, responsables de la tuberculosis en rumiantes, un hecho que va más allá de las reacciones cruzadas que aparecen cuando se utilizan pruebas inmunológicas para el diagnóstico de tuberculosis en animales infectados por paratuberculosis o vacunados (30 Esto también podría estar asociado con la reducción significativa de las lesiones relacionadas con la tuberculosis lograda después de la vacunación contra la paratuberculosis en cabras y terneros (32, 33). A pesar de este hecho, aún se desconoce el efecto de la inmunización con micobacterias relacionadas con la tuberculosis en el desarrollo de la paratuberculosis.
Por lo tanto, el objetivo de este estudio fue analizar el efecto de la vacunación homóloga o heteróloga de las cabras, y un desafío posterior con Map, en la respuesta inmune y la protección, y evaluar si los diferentes componentes de estas vacunas (bacterias inactivadas o adyuvantes) podrían participar en la respuesta del huésped vacunado contra Map.
Materiales y métodos
Declaración ética
Todos los procedimientos experimentales se llevaron a cabo de acuerdo con las leyes europeas (86/609) y españolas (R.D. 223/1988, R.D. 1021/2005, R.D. 53/2013) y aprobado por el gobierno local siguiendo el informe y las sugerencias del Subcomité de Experimentos y Bienestar Animal de la Universidad de León (ULE) (OEBA-ULE-016-2017).
Vacunas, productos de inmunización e inóculo de desafío
La vacuna comercial Silirum® contra la paratuberculosis y sus componentes (el adyuvante MontanideTM y la cepa Map 316F incluida en la vacuna) fueron preparadas por separado y enviadas por el fabricante, CZ Vaccines (Porriño, España), mientras que la vacuna Mbv inactivada por calor (HIMB) y sus componentes (un adyuvante En resumen, cada dosis de la vacuna HIMB (1 ml) consistió en una suspensión acuosa de 107 unidades formadoras de colonias (CFU) inactivadas por calor de la cepa de Mbv 1403 NEIKER (84-85 °C durante 45 min) emulsionada en el adyuvante MontanideTM ISA 50 V 2 (Seppic, Francia). Además, cada bacteria inactivada y los productos de inmunización adyuvante se ajustaron a la misma dosis administrada en las vacunas Silirum® y HIMB. Por lo tanto, cada dosis (1 ml) de bacterias inactivadas (109 UFC de Map 316F y 107 UFC de cepas Mbv 1403) se diluía en solución salina con fosfato (PBS), mientras que cada dosis de adyuvante MontanideTM (1 ml) (utilizada en las vacunas Silirum® y HIMB) se emulsionó en PBS
Además, la cepa bovina Map 764 se preparó para el desafío como lo describió anteriormente Fernández et al.(11). En resumen, la cepa Map 764 se cultivó en caldo Middlebrook 7H9 enriquecido con un 10% de ácido oleico-albúmina-dextrosa-catalasa (OADC) y 2 mg · L−1 de micobatina J (7H9 OADC MJ) durante 3 semanas a 37 ± 1°C. Luego, los cultivos se cosecharon por centrifugación a 3.000 g durante 10 minutos, y los gránulos bacterianos se lavaron dos veces y se reuspendieron en PBS. Con el fin de interrumpir los grupos bacterianos, la suspensión resultante se pasó hacia arriba y hacia abajo a través de una aguja de calibre 27 varias veces y se hizo vórtice. La concentración bacteriana se estimó mediante la densidad óptica (O.D.) y la estimación de la UFC de diluciones en serie de 10 veces chapadas en 7H9 OADC MJ consolidado con agar. Finalmente, las suspensiones se ajustaron a 1,2 × 1010 UFC · mL-1 y se mantuvieron a 4 °C durante todo el período de desafío (2 semanas), y los grupos bacterianos se interrumpieron de nuevo antes de la inoculación oral, como se mencionó anteriormente.
Diseño experimental
En este estudio se utilizaron un total de 50 hembras de cabras de raza Murciano-Granadina de 1 mes de edad. Los animales fueron seleccionados de un rebaño sin signos clínicos y dieron negativo para la tuberculosis y la tuberculosis en los últimos 10 años. Además, no se identificaron reactores positivos en las campañas oficiales anuales de erradicación de la tuberculosis, basadas en pruebas cutáneas intradérmicos, realizadas por las autoridades regionales de salud animal durante los últimos 5 años. El estado libre de paratuberculosis y tuberculosis de los animales experimentales se confirmó utilizando el ensayo de inmunoabsorbente ligado a enzimas de anticuerpos (ELISA) contra Map (ID Screen® Paratuberculosis indirect, IDVet, Gabrels, Francia) y Mbv (INgezim Tuberculosis DR, Eurofins Technology, Madrid, España) y la prueba de Después de un período de adaptación de 15 días en las instalaciones del Instituto de Ganadería de Montaña (IGM-ULE) en León (España), los niños cabras fueron asignados al azar en corrales separados, sometidos a prácticas de gestión estándar, y su estado de salud y bienestar se revisaba diariamente.
Al comienzo del estudio, y de acuerdo con el protocolo de vacunación y/o infección a seguir, las cabras se clasificaron en 10 grupos (n = 5) y se distribuyeron en diferentes corrales para evitar el contacto directo entre los grupos de acuerdo con el siguiente esquema (Figura 1):
 FIGURA 1. Esquema de diseño experimental. Las cabras se dividieron en siete grupos y se vacunaron con diferentes componentes 0 días después de la vacunación (dpv). Desde entonces hasta el sacrificio (190 dpv), se tomaron muestras de sangre (B) a intervalos de 30 días. Cinco cabras de cada grupo fueron desafiadas por vía oral con la cepa Map 764 a 45 dpv y divididas en 10 grupos: NV, no vacunadas y no infectadas; NVI, no vacunadas e infectadas; VS, Silirum® vacunadas y no infectadas; VSI, Silirum® vacunadas e infectadas; VH, HIMB vacunadas y no Finalmente, a 190 dpv, se recogieron heces (F), y se llevaron a cabo la tala y necropsias completas para la recolección de muestras de tejido (T).
FIGURA 1. Esquema de diseño experimental. Las cabras se dividieron en siete grupos y se vacunaron con diferentes componentes 0 días después de la vacunación (dpv). Desde entonces hasta el sacrificio (190 dpv), se tomaron muestras de sangre (B) a intervalos de 30 días. Cinco cabras de cada grupo fueron desafiadas por vía oral con la cepa Map 764 a 45 dpv y divididas en 10 grupos: NV, no vacunadas y no infectadas; NVI, no vacunadas e infectadas; VS, Silirum® vacunadas y no infectadas; VSI, Silirum® vacunadas e infectadas; VH, HIMB vacunadas y no Finalmente, a 190 dpv, se recogieron heces (F), y se llevaron a cabo la tala y necropsias completas para la recolección de muestras de tejido (T).
VS: Silirum® vacunado y no infectado
VSI: Silirum® vacunado e infectado
VH: HIMB vacunado y no infectado
VHI: HIMB vacunado e infectado
StrSI: inmunizado con la cepa Map 316F Silirum® e infectado
StrHI: inmunizado con la cepa Mbv 1403 HIMB e infectado
AdjSI: inmunizado con el adyuvante MontanideTM Silirum® e infectado
AdjHI: inmunizado con la vacuna MontanideTM HIMB adyuvante e infectado
NV: no vacunado y no infectado
NVI: no vacunado e infectado
No se incluyó animales no infectados en grupos inmunizados con bacterias o adyuvantes inactivados, debido al hecho de que el estudio se centró en la evaluación del efecto de los componentes de las vacunas en la protección, después de la infección por Map, y esto implicaría la inclusión de un gran número de animales y grupos que obstaculizarían el estudio.
La vacunación se realizó al comienzo del experimento, el día 0, mediante una inyección subcutánea en la pechuga con 1 ml de la vacuna Silirum®, 109 Map 316F Silirum® cepas de la vacuna CFU, 1 ml de adyuvante MontanideTM Silirum® (CZ Vaccines, Porriño, España), 1 ml de vacuna HIM
Cuarenta y cinco días después de la vacunación (dpv), los animales fueron desafiados por vía oral (Figura 1) utilizando una jeringa automática con una cantidad total de 1,2 × 1010 Mapa 764-CFUs diluidos en 40 ml de PBS como se describió previamente por Fernández et al. (11), mientras que 40 ml de PBS se administraron por vía oral A lo largo del ensayo experimental, todas las cabras fueron monitoreadas diariamente en busca de signos clínicos y muestreadas cada 30 días hasta el sacrificio (Figura 1). A 190 dpv, se realizaron necropsias completas y muestreo post mortem en todas las cabras después de ser sacrificadas humanamente por sedación profunda con xilazina (XILAGESIC®, Laboratorios Calier, Barcelona, España) y una posterior inyección intravenosa de T61® (MSD Animal Health, Salamanca, España) seguida de exhalación (Figura 1).
Colección de muestras
Las muestras de sangre se recogieron mensualmente de la vena yugular en tubos de Vacutainer con heparina de litio (Becton Dickinson and Company, Reino Unido) y sin anticoagulante (Becton Dickinson and Company, Reino Unido). Luego, las muestras heparinizadas se procesaron inmediatamente para la prueba de liberación de IFN-γ en respuesta al derivado proteico de los antígenos aviarios (PPDa) y bovinos (PPDb) a 0, 30, 60, 90, 120, 150 y 190 dpv (Figura 1). Al mismo tiempo, también se recogió sangre heparinizada para el aislamiento de células mononucleares de sangre periférica (PBMC) (35) y su posterior caracterización por citometría de flujo. Además, se procesaron muestras no heparinizadas para pruebas de determinación de anticuerpos Map (ID Screen® Paratuberculosis indirect, IDVet, Gabrels, Francia) y específicas de Mbv (INgezim Tuberculosis DR, Eurofins Technology, Madrid, España) a 0, 30, 60, 90, 120, 150 y 190 dpv (Figura 1).
Las muestras fecales de cada cabra se recogieron por separado en guantes de plástico desechables directamente del recto a 190 dpv, antes de la eutanasia del animal, y se congelaron a -20 °C hasta el procesamiento para la detección del mapa a través del cultivo bacteriológico.
Los animales fueron sacrificados en el día 190 dpv y se realizó una necropsia regulada, ordenada y completa. Después del examen general de las vísceras, se tomaron muestras de íleno (zonas proximales, media y distales), jejuno (zonas proximales, medias y distales), válvula ileocecal y parches de Peyer jejunal (al menos tres parches de cada zona: proximal, media y distal), junto con ganglios Además, también se recogieron y almacenaron muestras de íleo distal, parches jejunales de Peyer y ganglio linfático mesentérica a -20 °C para el aislamiento del mapa por cultivo y la detección mediante reacción en cadena de la polimerasa cuantitativa en tiempo real (qPCR), así como procesadas inmediatamente para el aislamiento de leucocitos con el
Aislamiento de PBMC y leucocitos tisulares
Los PBMC se aislaron como se describió anteriormente (35). En resumen, se centrifugaron 30 ml de sangre periférica heparinizada, y los PBMC se aislaron por centrifugación de gradiente utilizando LymphoprepTM (STEMCELL Technologies®, Colonia, Alemania). Los PBMC resultantes se lavaron tres veces con PBS, y las suspensiones celulares se reuspendieron en medio RPMI1640 suplementado + GlutaMaxTM (Gibco, Paisley, Reino Unido) y se contaron en una cámara Neubauer y se ajustaron a una concentración final de 106 células · mL-1. Las muestras intestinales y los ganglios linfáticos mesentéricos se recogieron y lavaron en PBS durante la necropsia, se pusieron en tubos de halcón individuales que contenían 30 ml de medio estéril suplementado RPMI1640 y se procesaron en el laboratorio dentro de los 30 minutos posteriores a la recolección. Para el aislamiento de linfocitos tisulares, se abrieron 5 cm de parches de íleon y jejunal de Peyer longitudinalmente mostrando la mucosa y se lavó con PBS hasta que se eliminaron los restos fecales, mientras que se extirpó la grasa pericapsular de los ganglios linfáticos. Se extirpó el exceso de tejido alrededor de cada parche de Peyer, y la mucosa de los parches de íleon y Peyer se raspó y picó. Además, 50 mg de tejido linfático mesentérico se cortaron en trozos pequeños y se cortaron con una hoja de bisturí. El tejido picado se suspendió en 11 ml de PBS con EDTA (2 mM) y se procesó con una licuadora (Masticador, IUL) durante 2,5 minutos. Luego, 10 ml de la porción homogeneizada superior se pasaron a través del filtro de 40 μm (Thermo Fisher Scientific, Madrid, España), y la suspensión resultante se superponó en un volumen igual de LymphoprepTM y se centrífugó a 800 g durante 30 minutos sin parada ni aceleración. Las células de la capa de interfaz se lavaron tres veces con PBS/EDTA, se contaron en una cámara Neubauer y se resuspenderon en RPMI1640 suplementado a una concentración final de 106 células · mL-1.
La viabilidad celular determinada por la exclusión del tinte azul trypan solía ser >90 % tanto para los PBMC como para los linfocitos tisulares (datos no mostrados).
Respuesta inmune humoral periférica y mediada por células locales y periférica
Las muestras enteras de sangre periférica heparinizada tomadas a 0, 30, 60, 90, 120, 150 y 190 dpv se procesaron dentro de las 3 horas posteriores a la recogida. Para cada animal, tres pozos de una placa de tejido de 24 pozos (Thermo Fischer Scientific, Rochester, NY) se llenaron con 1,5 ml de sangre cada uno y se mezclaron con 100 μL de PBS estéril (control negativo) o se estimularon con 100 μL de PPDa o PPDb (CZ Vaccines, Porriño Después de 22 horas de estimulación, las placas se centrifugaron a 750 g durante 15 minutos, y el plasma se recogió y almacenó a -20 °C hasta que se hizo la prueba.
Para el análisis de la respuesta inmune local mediada por células, un total de 2 × 106 leucocitos mononucleares aislados del íleon, los parches de Peyer y el ganglio linfático mesentérico se semlinó por pozo en tres pozos cada uno de una placa de tejido de 24 pozos y se estimularon como se describe para la sangre entera. Después de 22 horas de estimulación, las placas se centrifugaron a 750 g durante 15 minutos, y los supernatantes se recogieron y almacenaron a -20 °C hasta que se probaron.
La producción de IFN-γ en estos ensayos se evaluó por duplicado utilizando ELISA comercial para IFN-γ bovino siguiendo las instrucciones del fabricante (prueba Bovigam® Mbv IFN-γ para ganado, Thermo Fisher Scientific, Waltham, EE. UU.), y los valores de absorbancia se midieron espectrofotométricamente utilizando un lector ELISA ELX800 (Bio Los valores de O.D. se ajustaron dividiendo el valor de O.D. de plasma o sobrenadante por el valor de O.D. de control negativo de cada placa para evitar variaciones entreplaca. Los resultados se expresaron como valores de índice aveaviano y bovino por medio del cociente entre la O.D. media de PPDa u O.D. del plasma estimulado por PPDb, respectivamente, y la O.D. media del plasma de control negativo (11, 36).
Se permitió que las muestras de sangre no heparinas coagularan y se retractaban, y el suero se almacenó a -20 °C hasta que se utilizó. Cada suero se probó para detectar anticuerpos específicos contra Map (37) y Mbv (38) utilizando pruebas ELISA comerciales (ID Screen® Paratuberculosis indirect, IDVet, Gabrels, Francia; e INgezim Tuberculosis DR Eurofins Technology, Madrid, España, respectivamente) siguiendo las instrucciones del fabricante. Los resultados se expresaron como la relación S/P de los anticuerpos Map y Mbv calculada dividiendo el O.D. corregido de la muestra por el O.D. corregido del control positivo y multiplicándolo por 100 (39).
Detección cuantitativa del mapa en tiempo real
El ADN se extrajo de 50 mg de íleon distal, parches Peyer jejunales y ganglio linfático mesentérica utilizando el kit de purificación de ADN de tejido Maxwell® 16 (Promega, WI, EE. UU.) con el instrumento Maxwell 16 (Promega) siguiendo las instrucciones del fabricante. A continuación, el ADN se cuantificó utilizando el kit del sistema QuantiFluorTM ONEdsDNA (Promega, WI, EE. UU.) y el fluorímetro QuantusTM (Promega, WI, EE. UU.). El ADN extraído se diluió a 50 ng · μL-1 y se almacenó a -20°C hasta que se realizó la qPCR. El ADN genómico de 2 × 108 UFC de mapa y el de 50 mg de tejidos de un animal no infectado se extrajeron y cuantificaron para generar una curva estándar.
La detección de la secuencia del mapa IS900se realizó como lo describió anteriormente Arteche-Villasol et al.(40). Además, la cuantificación de ADN de mapa y la sensibilidad analítica de qPCR se evaluaron mediante la construcción de una curva estándar diluida de 10 veces utilizando ADNgenomico de mapa que oscilaba entre 1.000 pg y 0,001 pg/reacción mezclada con 100 ng/reacción de ADN tisular de un animal no infectado. Las muestras se consideraron positivas cuando el pico de disociación (Tm) fue de 89,1 ± 1,5 °C y los ciclos umbral (Ct) fueron ≤ 37 (41, 42). Los resultados de la qPCR se analizaron utilizando 7500 Software v2.0.6 (Applied BiosystemsTM). La cantidad de ADN del mapa (pg) de cada pozo se calculó mediante la interpolación de sus valores de Ct con la curva estándar como se describió anteriormente (43), y la cantidad media se calculó a partir de ambos duplicados.
Tejido y cultura fecal del mapa
El íleon distal, los parches de Peyer jejunal y el ganglio linfático mesentérérico de cada animal se probaron individualmente, mientras que las muestras fecales se agruparon en grupos de tres cabras cada una. Los segmentos de parche de íleon y Peyer (12 cm) y 2 gramos de ganglio linfático mesentérica se procesaron como se describe para el aislamiento de leucocitos tisulares y siguiendo los métodos descritos anteriormente (44). En resumen, 2 g de cada grupo de muestras de tejido y fecales se descontaminaron con 38 ml de cloruro de hexadecilpiridinio y se homogeneizaron en una licuadora (Masticador, IUL) durante aproximadamente 15 s. Después de 18 horas de descontaminación, se utilizaron 200 μL de la suspensión para inocular dos tubos que contienen el medio de yema de huevo de Herrold suplementado con piruvato de sodio y micobacitina J (MJ) y dos tubos que contenían 7H9 OADC suplementado con MJ, penicilina, anfoteric Los cultivos se incubaron a 37 °C ± 1°C, y el crecimiento se comprobó mediante un examen bajo un microscopio estereoscópico después de 8, 12, 16 y 20 semanas de postnoculación. Los cultivos se consideraron positivos si se observaban una o más colonias cartográficas características en cualquier tubo. Las colonias aisladas en ambos medios fueron confirmadas por una PCR múltiplex en tiempo real que detecta las secuencias de mapas IS900e ISMap02 (45).
Análisis citométrico de flujo de PBMC y leucocitos tisulares
Se llevó a cabo un análisis de citometría de flujo de un solo color para la caracterización fenotípica de PBMC aislados a 0, 30, 60, 90, 120, 150 y 190 dpv y leucocitos mononucleares aislados de íleo distal, parches de Peyer jejunal y ganglio linfático mesentero. Un número total de 2 × 105 células por pozo se sembró en una placa de 96 pozos (Thermo Fisher Scientific, Roskilde, Dinamarca) e incubaron con anticuerpos primarios contra los marcadores de superficie de linfocitos detallados en la Tabla 1 durante 1 hora a 4 °C. Posteriormente, las células se lavaron dos veces con PBS e incubaron con anticuerpos secundarios conjugados apropiados durante 1 hora a 4 °C (Tabla 1). Finalmente, las células se fijaron con el 1% de CellFIXTM (Becton Dickinson and Company, Erembodegem, Bélgica) hasta que se analizaron. La adquisición de muestras de 10.000 eventos se realizó utilizando un citometro de flujo (MACSQuant, Miltenyi Biotec®), donde los eventos se cerraron durante el análisis de adquisición como se describió anteriormente en otros lugares (35) para descartar la presencia de aire y dobles. Luego, se llevó a cabo el análisis de los datos utilizando el MACSQuantify10 SoftwareTM (Miltenyi Biotec®), y los resultados se expresaron como porcentaje de células positivas.
 TABLA 1. Anticuerpos primarios y secundarios utilizados en el análisis de citometría de flujo de PBMC y subpoblaciones de linfocitos tisulares.
TABLA 1. Anticuerpos primarios y secundarios utilizados en el análisis de citometría de flujo de PBMC y subpoblaciones de linfocitos tisulares.
Examen histopatológico
Los tejidos fijos para el examen histopatológico se procesaron convencionalmente para la incrustación de parafina y se tiñeron con la técnica hematoxilina-eosina y Ziehl-Neelsen para la detección de bacilos ácidos rápidos (46).
Las lesiones consistentes con la infección por Map se clasificaron como focales, multifocales a y multifocales b, o formas difusas de acuerdo con la presencia y ubicación de los granulomas y siguiendo las pautas descritas anteriormente para pequeños rumiantes (8, 46). En resumen, las lesiones se caracterizaron como formas focales cuando los granulomas se limitaban al tejido linfoide de los parches de Peyer; formas multifocales cuando los granulomas se encontraban en la lámina propia adyacente al tejido linfoide (multifocal a) o no (multifocal b) y formas difusas cuando los granulomas se propagaban a amplias áreas La clasificación de cada animal se basó en su lesión granulomatosa más grave. Después del examen histopatológico, se cuantificó el número de granulomas por sección de tejido en todas las muestras de tejido como se describe en otros lugares (11, 46). Se seleccionaron tres secciones de tejido de cada sitio intestinal (es decir, ileum (zonas proximal, media y distal; jejuno – zonas proximal, media y distal; valva ileocecal y parches de Peyer jejunal, zonas proximales, media y distales) y dos secciones de cada ganglio linfático (es decir, ganglios
Análisis estadístico
La distribución normal de los resultados de los anticuerpos específicos de Map y Mbv, la producción local y periférica de IFN-γ y la proporción de subpoblaciones de linfocitos periféricos y tisulares se evaluaron para la normalidad utilizando la prueba de Shapiro-Wilk. Los datos de la producción de IFN-γ y las pruebas ELISA de anticuerpos se transformaron logarítmicamente. Luego, la citometría de flujo y los resultados de ELISA de la producción periférica de IFN-γ y anticuerpos se analizaron utilizando un procedimiento de modelo lineal generalizado (GLM) para evaluar los principales efectos de la vacunación, el desafío, el tiempo y sus interacciones. Posteriormente, se estimaron las diferencias entre los grupos de vacunación para cada vez que se muestreó utilizando la corrección de Tukey-Kramer para múltiples comparaciones. Del mismo modo, los principales efectos de la vacunación, los tejidos y sus interacciones se estimaron en los resultados de la producción local de IFN-γ, seguido de la evaluación de las diferencias entre los grupos de vacunación y los tejidos utilizando la prueba de comparaciones múltiples Tukey-Kramer. Además, las diferencias entre los grupos en el número de cabras con lesiones se evaluaron utilizando las pruebas exactas de χ2 y Fisher. Además, después de la transformación logarítmica, las diferencias en el recuento de granuloma entre los grupos de vacunación se calcularon utilizando la prueba t de Student. Todos los análisis estadísticos se llevaron a cabo utilizando el software GraphPad Prism 6.0 (San Diego, CA, EE. UU.), excluyendo los GLM que se realizaron con el software R 3.5.3 (R Development Core Team, 2019). P < 0,05 se consideró estadísticamente significativo.
Resultados
Respuesta inmunitaria mediada por células periféricas y locales (IFN-γ)
Los resultados de GLM mostraron que los valores del índice de aves y bovino alcanzaron niveles significativos de 60 a 190 dpv en los grupos VS, VSI, VH y VHI (p < 0,001 y p < 0,01 respectivamente) (Figuras 2A, B). Además, la inmunización de la vacuna Silirum® (VS y VSI) y la vacuna HIMB (VH y VHI) ejerció un efecto considerable en los valores del índice de aves, independientemente del estado de infección (p < 0,01 y p < 0,001, respectivamente) (Figura 2A), pero solo los grupos VHI, VH y StrHI mostraron un efecto significativo Además, el desafío oral afectó significativamente a los niveles del índice aviar de los grupos VSI, VHI (p < 0,001) y NVI (p < 0,05) y en los valores del índice bovino de los grupos VSI, VHI (p < 0,01) y StrHI (p < 0,05).
 FIGURA 2. Cinética de la producción de IFN-γ por sangre entera estimulada con antígenos de aves (PPDa) y bovinos (PPDb). Los resultados se expresan como valores de índice de A.D. de aves (A) y bovinos (B) de cada grupo de vacunación (n = 5) (NV, no vacunados y no infectados; NVI, no vacunados e infectados; VS, Silirum® vacunado y no infectados; VSI, Silirum® vacunado e infectado; VH, La línea roja punteada vertical representa el tiempo del desafío oral del mapa (45 dpv). Las diferencias significativas determinadas por múltiples comparaciones se representaron como *p < 0,05, **p < 0,01, ***p < 0,001, ****p < 0,0001.
FIGURA 2. Cinética de la producción de IFN-γ por sangre entera estimulada con antígenos de aves (PPDa) y bovinos (PPDb). Los resultados se expresan como valores de índice de A.D. de aves (A) y bovinos (B) de cada grupo de vacunación (n = 5) (NV, no vacunados y no infectados; NVI, no vacunados e infectados; VS, Silirum® vacunado y no infectados; VSI, Silirum® vacunado e infectado; VH, La línea roja punteada vertical representa el tiempo del desafío oral del mapa (45 dpv). Las diferencias significativas determinadas por múltiples comparaciones se representaron como *p < 0,05, **p < 0,01, ***p < 0,001, ****p < 0,0001.
El análisis de comparaciones múltiples mostró que el grupo VSI tenía valores de índice avia más altos que NV, NVI, StrSI, AdjSI y AdjHI a 60 y 120 dpv, mientras que el grupo VS mostró estas diferencias solo a 120 dpv (Figura 2A). Por el contrario, estos grupos no mostraron ninguna diferencia significativa en los niveles del índice bovino (Figura 2B). Además, no se observaron diferencias entre VS y VSI ni en los valores del índice aviado o bovino en ningún momento. Por otro lado, el grupo VHI mostró valores de índice ave más grandes que NVI, NV, StrSI, AdjSI y AdjHI a 120 dpv (Figura 2A). Además, los valores del índice bovino de este grupo fueron significativamente más altos que NV, NVI, StrSI, AdjSI y AdjHI a 90, 120, 150 y 190 dpv y mayores que VH a 120 dpv (Figura 2B).
La producción local de IFN-γ en leucocitos mononucleares purificados de los tejidos y estimulados con PPDa y PPDb fue menor que la observada en los PBMC. Además, los valores del índice aviar fueron más heterogéneos que los niveles del índice bovinos por la mayor variabilidad individual observada en el primero (Figuras 3A, B), específicamente en los grupos NVI, VH, VHI, StrHI y AdjSI (Figura 3A). Los parches de Peyer y el ganglio linfático mesentérica del grupo NVI mostraron los valores más altos del índice aviar, seguidos del íleon, los parches de Peyer y el ganglio linfático mesentérica del grupo StrHI (Figura 3A). Sin embargo, el análisis estadístico no mostró ningún efecto significativo de la vacunación o la ubicación del tejido en los valores del índice aviar o bovino (p > 0,05). Además, tampoco se observaron diferencias significativas entre los grupos vacunados en el análisis de comparaciones múltiples (p < 0,05). Mientras tanto, se observaron diferencias significativas en los valores del índice aviar dentro del grupo NVI, donde la producción de IFN-γ fue mayor en los parches de Peyer y el ganglio linfático mesentérico que en el íleon (p < 0,05) (Figura 3A). Además, a pesar de la alta variabilidad dentro de los grupos observados en los valores del índice bovino, los valores del ganglio linfático mesentérico del grupo VH fueron considerablemente más altos que los de los parches de íleon y Peyer (p < 0,01) (Figura 3B).
 FIGURA 3. Producción de IFN-γ por leucocitos a partir de íleon (IL), parches Peyer jejunales (JPP) y ganglios linfáticos mesenteros (MLN) estimulados con antígenos aviarios (PPDa) y bovinos (PPDb). Los resultados se expresan como valores de índice de O.D. de aves (A) y bovinos de cada grupo de vacunación (n = 5) (NV, no vacunados y no infectados; NVI, no vacunados e infectados; VS, Silirum® vacunado y no infectados; VSI, Silirum® vacunado e infectado; VH, HIMB vacuna Las barras y las líneas verticales representan valores medios y desviaciones estándar, respectivamente. Las diferencias significativas determinadas por múltiples comparaciones se representaron como *p < 0,05, **p < 0,01, ***p < 0,001, ****p < 0,0001.
FIGURA 3. Producción de IFN-γ por leucocitos a partir de íleon (IL), parches Peyer jejunales (JPP) y ganglios linfáticos mesenteros (MLN) estimulados con antígenos aviarios (PPDa) y bovinos (PPDb). Los resultados se expresan como valores de índice de O.D. de aves (A) y bovinos de cada grupo de vacunación (n = 5) (NV, no vacunados y no infectados; NVI, no vacunados e infectados; VS, Silirum® vacunado y no infectados; VSI, Silirum® vacunado e infectado; VH, HIMB vacuna Las barras y las líneas verticales representan valores medios y desviaciones estándar, respectivamente. Las diferencias significativas determinadas por múltiples comparaciones se representaron como *p < 0,05, **p < 0,01, ***p < 0,001, ****p < 0,0001.
Respuesta humorística periférica
Los resultados de GLM estimaron que la producción de anticuerpos específicos tanto de Map como de Mbv alcanzó niveles significativos de 60 a 190 dpv (p < 0,001 y p < 0,0001, respectivamente) en los grupos vacunados por Silirum® y HIMB (Figuras 4A, B). Además, el desafío oral impulsó la producción de anticuerpos específicos de Map en los grupos vacunados VSI, VHI (p < 0,001) y NVI (p < 0,05) (Figura 4A) y la producción de anticuerpos específicos de Mbv en VHI (p < 0,001) (Figura 4B). Además, el análisis de comparaciones múltiples mostró que solo los VSI tenían valores S/P específicos del mapa significativamente más altos en comparación con los grupos inmunizados con adyuvantes, bacterias inactivadas y no vacunados a 90, 120, 150 y 190 dpv, mientras que no se observaron diferencias significativas en VHI (Figura 4A). Además, no se observaron diferencias significativas en los niveles de anticuerpos Map ni entre los grupos de vacunación Silirum® y HIMB ni entre los grupos vacunados infectados y no infectados.
 FIGURA 4. Cinética de los niveles de anticuerpos séricas en la sangre periférica. Los resultados se expresan como la relación S/P de los anticuerpos específicos de Map (A) y Mbv (B) de cada grupo de vacunación (n = 5) (NV, no vacunados y no infectados; NVI, no vacunados e infectados; VS, Silirum® vacunado y no infectados; VSI, Silirum® vacunado e infectado; VH La línea roja punteada vertical representa el tiempo del desafío oral del mapa (45 dpv). Las diferencias significativas determinadas por múltiples comparaciones se representaron como *p < 0,05, **p < 0,01, ***p < 0,001, ****p < 0,0001.
FIGURA 4. Cinética de los niveles de anticuerpos séricas en la sangre periférica. Los resultados se expresan como la relación S/P de los anticuerpos específicos de Map (A) y Mbv (B) de cada grupo de vacunación (n = 5) (NV, no vacunados y no infectados; NVI, no vacunados e infectados; VS, Silirum® vacunado y no infectados; VSI, Silirum® vacunado e infectado; VH La línea roja punteada vertical representa el tiempo del desafío oral del mapa (45 dpv). Las diferencias significativas determinadas por múltiples comparaciones se representaron como *p < 0,05, **p < 0,01, ***p < 0,001, ****p < 0,0001.
Los valores S/P específicos de Mbv fueron significativamente más altos cuando se comparaba el grupo VHI con los grupos VS, VSI, StrSI, StrHI, AdjSI y AdjHI en 60, 90, 120, 150 y 190 dpv (Figura 4B). Sin embargo, se observaron diferencias significativas entre VHI y VH solo en 120, 150 y 190 dpv (Figura 4B).
PCR cuantitativa y bacteriología de tejidos y heces
La detección de mapas fue confirmada por qPCR (n = 4) y cultivo bacteriológico (n = 3) dentro de cabras infectadas (n = 35). Los animales positivos a qPCR correspondieron a (i) dos cabras de StrHI (linfondulomesenérico y los parches de Peyer, respectivamente), (ii) una de VHI (latos de pererior) y (iii) uno de AdjSI (pernos de oveja). Además, los animales positivos al cultivo bacteriológico correspondían a una cabra cada uno de NVI, VHI (ileum, parches de Peyer y ganglios linfáticos mesentéricos positivos) y VSI (ileum y linfáticos positivos mesentéricos). Solo los parches de Peyer de la cabra de VHI mostraron resultados positivos para ambas técnicas de detección. Todas las muestras de tejido de grupos no infectados fueron negativas a la amplificación de la secuencia IS900por qPCR o cultivo bacteriológico. La cultura bacteriológica de Map fue negativa en todas las muestras fecales analizadas.
Subconjuntos relativos de linfocitos sanguíneos y tejidos periféricos
Las desviaciones medias y estándar de las proporciones relativas de tres linfocitos T (CD4+, CD8+ y CD3+), un linfocito T γδ y dos linfocitos B (CD21+ y CD20+) se estimaron mediante citometría de flujo.
A lo largo de todos los grupos, la proporción relativa más alta de células positivas en PBMC aisladas de sangre entera correspondió a los linfocitos T γδ (30,46 % ± 3,55%), mientras que la proporción más baja se observó en los linfocitos B CD20+ (5,96 ± 1,48%) a lo largo del estudio ( Sin embargo, a pesar de que el análisis estadístico muestra oscilaciones significativas (p < 0,001) de las proporciones relativas de los linfocitos CD4+, CD8+, WC1+, CD21+, CD3+ y CD20+ en diferentes muestreos, no se encontraron diferencias significativas entre los grupos (p > 0,05).
La citometría de flujo realizada en células purificadas a partir de muestras de tejido mostró que los niveles de proporción relativa más altos correspondían a los linfocitos CD21+ B en el íleon (39,47 ± 4,71%), los parches de Peyer (26,81 ± 3,59%) y el ganglio linfático mesentérico (44.90 ± El análisis estadístico mostró una influencia significativa de la vacunación e inmunización con bacterias y adyuvantes inactivados en la proporción relativa de subconjuntos de linfocitos tisulares (p < 0,05). Sin embargo, los resultados de las comparaciones entre grupos no mostraron un patrón claro entre la proporción relativa de subpoblaciones de linfocitos y los diferentes grupos probablemente por la variabilidad individual observada dentro de los grupos. La media, la desviación estándar y los resultados de las comparaciones múltiples de subpoblaciones relativas de linfocitos en el íleon, los parches de Peyer y los ganglios linfáticos mesenteros se resumen en las Tablas Suplementarias 2-4.
Hallazgos patológicos
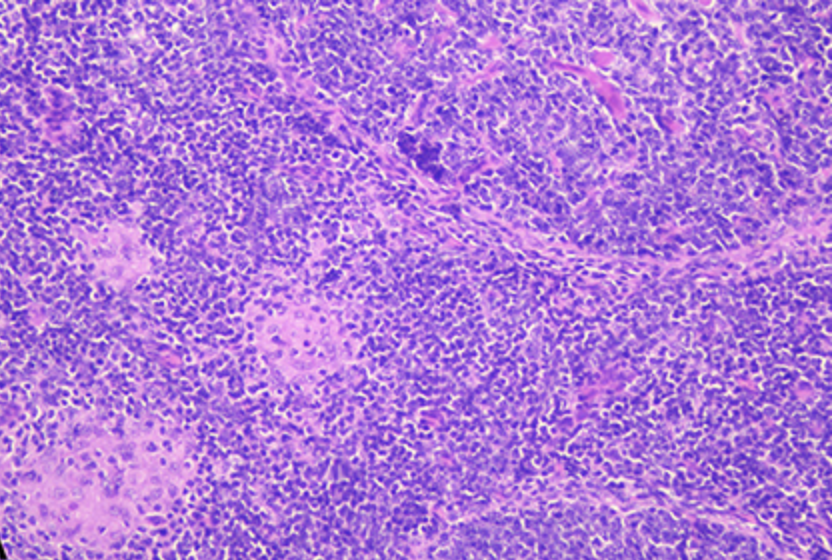
No se encontraron lesiones gruesas compatibles con la paratuberculosis en ningún animal. Se detectaron lesiones granulomatosas microscópicas características de la infección por mapa en todos los grupos infectados, con diferencias en la gravedad y la distribución. El número de cabras por grupo con lesiones según el tejido y el número de cabras por grupo clasificados en términos de gravedad de las lesiones se muestran en la Tabla 2. Las cabras con granulomas pequeños y bien definidos compuestos de macrófagos, con un citoplasma pálido abundante y un núcleo grande, escoltados por unos pocos linfocitos ubicados exclusivamente en el área interfollicular del tejido linfoide intestinal, se clasificaron como focales (n = 5) (Figura 5A). Además, las cabras con granulomas bien definidos en el área interfollicular de los parches de Peyer y también en la lámina propia estrechamente asociada con el tejido linfoide intestinal se clasificaron como a multifocal (n = 5) (Figura 5B). Finalmente, la presencia de lesiones granulomatosas no solo en los parches de Peyer y lamina propia relacionada, sino también en áreas de la mucosa sin ninguna asociación con el tejido linfoide, se consideró como multifocal b (n = 13) (Figura 5C). No se notaron lesiones difusas en los tejidos de ninguna cabra (n = 0). Los grupos VSI y VHI fueron los menos afectados con solo un animal de cada grupo desarrollando lesiones granulomatosas, mostrando diferencias significativas en comparación con el NVI en el que las lesiones estaban presentes en todos los animales (p < 0,05). Además, las lesiones de la cabra VSI se clasificaron como focales, mientras que la cabra del grupo VHI mostró lesiones más graves, categorizadas como b multifocales que todas las cabras del NVI. Además, la mayoría de las cabras inmunizadas con cepas (StrSI y StrHI) desarrollaron formas b multifocales, a excepción de un animal de StrHI con una lesión focal. Por el contrario, entre los animales inmunizados con adyuvantes (AdjSI y AdjHI), solo un animal del AdjSI mostró lesiones b multifocales, mientras que se encontraron lesiones a focales y multifocales en las cabras restantes. No se detectaron diferencias significativas en el número de cabras con lesiones granulomatosas en grupos inmunizados con bacterias o adyuvantes inactivados (p > 0,05). Las lesiones granulomatosas se localizaron principalmente en los parches jejunales de Peyer, seguidos de íleon distal, ganglios linfáticos, válvula ileocecal y jejuno, aunque las lesiones observadas en los ganglios linfáticos siempre se asociaron con la presencia de lesiones granulomatosas en el intestino (Tabla 3). No se observaron lesiones microscópicas en los grupos que permanecieron como no infectados. No se detectaron bacilos ácidos en las lesiones de ninguna de las cabras infectadas.
 TABLA 2. Número de animales de cada grupo con lesiones microscópicas consistentes con la paratuberculosis de acuerdo con el tejido examinado y su gravedad (focal, multifocal a, multifocal b y difuso).
TABLA 2. Número de animales de cada grupo con lesiones microscópicas consistentes con la paratuberculosis de acuerdo con el tejido examinado y su gravedad (focal, multifocal a, multifocal b y difuso).
 FIGURA 5. Tipos de lesiones granulomatosas paratuberculosis encontradas en el estudio. (A) La lesión focal: granulomas bien delimitados compuestos por pequeños grupos de macrófagos ubicados en el área interfollicular de los parches de Peyer. (B) Lesión multifocal: pequeños granulomas ubicados en el área interfollicular del tejido linfoide (asterisco) y en la lámina propia asociada (cabezas de flecha). (C) Lesión multifocal b: granulomas ubicados en la lámina propia en un área desprovista de tejido linfoide.
FIGURA 5. Tipos de lesiones granulomatosas paratuberculosis encontradas en el estudio. (A) La lesión focal: granulomas bien delimitados compuestos por pequeños grupos de macrófagos ubicados en el área interfollicular de los parches de Peyer. (B) Lesión multifocal: pequeños granulomas ubicados en el área interfollicular del tejido linfoide (asterisco) y en la lámina propia asociada (cabezas de flecha). (C) Lesión multifocal b: granulomas ubicados en la lámina propia en un área desprovista de tejido linfoide.
 TABLA 3. Resultados del recuento de granulomas. Número de granulomas según su ubicación para cada grupo y número total y porcentaje (%) de granulomas por grupo.
TABLA 3. Resultados del recuento de granulomas. Número de granulomas según su ubicación para cada grupo y número total y porcentaje (%) de granulomas por grupo.
El número total de granulomas por grupo se resumió en la Tabla 3. El grupo NVI mostró el número más alto, mientras que el más bajo correspondía a los grupos VSI y VHI. Además, el número de granulomas observados en los grupos StrSI y StrHI fue mayor que los observados en AdjSI y AdjHI. Se observaron diferencias significativas en el número medio de granulomas por grupo (Figura 6) entre los grupos NVI (159,60 ± 271,90) y VSI (4,60 ± 10,29) y VHI (6,20 ± 13,86) (p < 0,05). Sin embargo, no se encontraron diferencias significativas en los grupos StrSI (57,60 ± 61,54), StrHI (61,40 ± 83,22), AdjSI (20,00 ± 26,82) y AdjHI (29,20 ± 33,86) (p > 0,05).
 FIGURA 6. Cuenta media de granulomas de los tejidos de los grupos vacunados e infectados. Los datos se expresan como número medio de granulomas por grupo, y las desviaciones estándar en cada grupo (n = 5) y las diferencias significativas se representaron como *p < 0,05, **p <0,01, ***p < 0,001, ****p < 0,0001.
FIGURA 6. Cuenta media de granulomas de los tejidos de los grupos vacunados e infectados. Los datos se expresan como número medio de granulomas por grupo, y las desviaciones estándar en cada grupo (n = 5) y las diferencias significativas se representaron como *p < 0,05, **p <0,01, ***p < 0,001, ****p < 0,0001.
Discusión
Los estudios de vacunación contra la paratuberculosis realizados en rumiantes han demostrado resultados valiosos en la reducción del número de animales clínicamente afectados y la prevalencia de la enfermedad (16, 23). Sin embargo, la incapacidad de la respuesta inmune protectora provocada por la vacunación para prevenir completamente la infección y su progresión y propagación se aleja del término «vacuna ideal» (47, 48). Esta desventaja se debe en parte a la falta de conocimiento sobre los mecanismos periféricos y locales relacionados con el éxito o el fracaso de la protección contra las micobacterias. Aquí, este trabajo analiza la respuesta inmune local y periférica provocada por la vacunación homóloga (Silirum®) y heteróloga (vacunaHIMB) y la inmunización con sus componentes por separado y su relación con el grado de protección estimado por la evaluación de las lesiones granulomatoscópicos y la presencia del mapa en los tejidos.
A pesar de la identificación de lesiones granulomatosas distintivas de paratuberculosis en todos los grupos de cabras infectadas, independientemente de su estado de vacunación o inmunización, la variación en su gravedad refleja las observaciones de los estudios de campo y experimentales, ya que las lesiones observadas en las cabras vacunadas por paratuberculosis fueron leves (focales) y escasas, No se observaron lesiones difusas, posible debido al corto período de incubación de este trabajo (4-5 meses), en línea con otros estudios en los que se observaron lesiones b multifocales hasta 7 meses después de la infección, mientras que no se notificaron lesiones de tipo difuso hasta 12 meses (46, 49). A pesar de este hecho, los resultados obtenidos aquí coinciden con otros estudios experimentales en los que las lesiones observadas en animales vacunados se limitaron a formas regresivas (focales), contenidas en el área interfollicular del tejido linfoide intestinal, evitando su diseminación a través del intestino pero sin prevenir la infección, como se confirmó mediante la detección de Map en tejidos de
Hasta la fecha, los estudios de eficacia de la vacunación in vivo se han centrado principalmente en la inducción temprana y sostenida de IFN-γ periférico (20, 21, 52), ya que la liberación de esta citocina se ha asociado con la activación de mecanismos antimicobacterianos y la prevención del crecimiento bacteriano intracelular (53, 54). Ciertamente, la dinámica de producción de IFN-γ observada en los grupos vacunados por Silirum® (es decir, VS y VSI) fue consistente con otros estudios realizados con niños caprinos vacunados con vacunas cales que matan (52, 55). Además de la respuesta inmune mediada por células, los animales vacunados por Silirum® también mostraron un aumento de los niveles de anticuerpos de Map. El papel de la respuesta humoral se ha subestimado, ya que se ha considerado ineficaz contra los patógenos intracelulares (56, 57). Sin embargo, un número emergente de estudios realizados en condiciones experimentales han sugerido el posible efecto positivo de los anticuerpos en el control de la paratuberculosis, incluso en relación con la vacunación (58-60). Esto se apoya en la fuerte respuesta humoral inicial observada en ovejas vacunadas por la paratuberculosis y la capacidad de estos animales para modular esa respuesta en etapas posteriores para prevenir la progresión de la infección (60). Tanto las respuestas celulares como las humorales se mejoraron significativamente en este estudio después del desafío experimental con Map. Estos resultados imitan el aumento de IFN-γ observado en cabras vacunadas por paratuberculosis y expuestas naturalmente en condiciones de campo (61), aunque el aumento significativo en la producción de anticuerpos después del desafío oral de Map aquí observó contrastes con estudios anteriores donde la infección oral experimental en ovejas vacunadas no mostró diferencias con animales no vacunados o incluso condujo a una Sin embargo, el aumento de las respuestas celulares y humorales aquí observadas no impidieron la aparición de lesiones granulomatosas en el grupo VSI. Por lo tanto, a pesar de que la vacunación podría estar limitando la progresión y la gravedad de las lesiones, ya que solo una cabra VSI desarrolló un bajo número de granulomas focales, la respuesta celular y humoral periférica no permitió predecir la presencia de lesiones o infecciones en animales vacunados.
Además, los cambios en la proporción relativa de subpoblaciones de linfocitos también se han relacionado con infecciones micobacterianas (62–64). En este sentido, las vacas con enfermedad clínica han mostrado proporciones más bajas de linfocitos CD4+, CD8+ y γδ T que las vacas subclínicas en PBMC recién aislados (65). Además, en otro estudio, se detectaron porcentajes más altos de linfocitos γδ T y B y una disminución de CD4+ en los tejidos de ovejas infectadas durante la infección experimental temprana de Map (10). En cuanto al efecto de la vacunación, se ha informado que la infección de las ovejas vacunadas por Gudair® con Map conduce a una disminución en las proporciones de linfocitos periféricos CD4+ y B a los 13 días posteriores al desafío evaluados en PBMC estimulados in vitro por Map, pero esto no se evaluó en células no estimuladas y recién aisladas (22).Por el contrario, no se detectaron cambios en la proporción relativa de linfocitos T periféricos (CD4+, CD8+, γδ y CD3+) o linfocitos B (CD20+ y CD21+) en PBMC recién aislados entre cualquier grupo de este estudio. Además, se estimaron diferencias en la proporción relativa del tejido de subpoblaciones de linfocitos, pero no se observó una relación clara entre estos resultados y la vacunación/inmunización y/o infección, probablemente debido a la alta variabilidad individual dentro de los grupos probados. Del mismo modo, no se observó ninguna relación entre la producción local de IFN-γ y el desarrollo de lesiones, ya sea en VSI u otros grupos infectados. En este sentido, solo los parches de Peyer y el ganglio linfático mesentérico del grupo NVI produjeron una producción de IFN-γ ligeramente más alta. Este resultado es consistente con el mayor número de granulomas detectados en este grupo, ya que un mayor número de lesiones granulomatosas se han relacionado con un mayor número de células inmunomarcadas con IFN-γ (6) y es probable que las lesiones presentes en los grupos infectados restantes no sean suficientes para generar una respuesta inmune mediada por células locales sustanciales en respuesta a los Por lo tanto, ni las respuestas inmunitarias celulares y humorales periféricas ni la evaluación de la proporción relativa de subpoblaciones de linfocitos en sangre y tejidos enteros ni la producción local de IFN-γ pudieron predecir aquellos animales que permanecen desprotegidos temprano después de la infección. Además, se ha demostrado que la vacunación reduce el número de bacterias excretadas tanto en condiciones de campo como experimentales (17, 21, 37, 66). En este estudio, Map no se aisló en las heces de ninguna cabra. Se ha detectado una mayor excreción de mapas en relación con el desarrollo de lesiones graves y signos clínicos en animales infectados de forma natural y experimental (67, 68), aunque podría detectarse intermitente en animales con paratuberculosis subclínica (53, 69). Sin embargo, las heces se recogieron solo en el sacrificio (145 días después de la infección), por lo que la presencia de Map podría haberse subestimado debido al corto tiempo posterior al desafío y al tiempo limitado muestreado. Además, se ha observado que la excreción fecal se asocia con frecuencia con animales con un alto número de tejidos positivos para el cultivo y lesiones graves (70). Además, el bajo número de tejidos Map-positivos detectados por cultivo bacteriológico o qPCR (7 de 35 cabras infectadas) no fue sorprendente, ya que en varios trabajos anteriores, ya sea en casos naturales o experimentales (46, 71), se ha demostrado que la carga bacteriana en animales con lesiones similares a las encontradas en este estudio estaba ausente o muy baja.
Curiosamente, solo una cabra del grupo VHI desarrolló lesiones granulomatosas paratuberculosis, aunque clasificadas como multifocales b, mostrando la eficacia de la protección heteróloga que la vacunación contra la tuberculosis confiere contra la infección de Map. Al igual que la vacunación homóloga, los grupos vacunados con HIMB (es decir, VH y VHI) mostraron una producción significativa de IFN-γ y anticuerpos como se describió anteriormente en cabras (66, 72), incluso cuando la sangre se estimulaba con PPDa. Además, también se observó un aumento en la producción de IFN-γ en cabras vacunadas por vía subcutánea con la vacuna HIMB y desafiadas endobronquialmente con una micobacteria estrechamente relacionada (M. caprae) en condiciones experimentales (66), demostrando una vez más que la elevación de estas respuestas no garantiza el éxito de la vacunación, ya que Se ha informado previamente de un efecto de protección cruzada relacionado con la vacunación, ya que se observó una reducción de las lesiones pulmonares en cabras vacunadas por la paratuberculosis (32) o en terneros (33) infectados por endobronquialmente con M. caprae o Mbv, respectivamente. Además, a pesar de este efecto beneficioso, se notó una reacción cruzada contra los antígenos micobacterianos estándar en el ensayo de liberación de IFN-γ y en la determinación de anticuerpos específicos de Map e implicó una desventaja relevante para la diferenciación entre animales infectados y vacunados (DIVA) (31). Esta reacción cruzada se ha descrito previamente en respuesta a los antígenos micobacterianos PPDa y PPDb, aunque por lo general esta producción está sesgada hacia el antígeno relacionado con la estimulación homóloga (73-75). Es notable que se detectaron anticuerpos específicos contra Mbv solo en el grupo VHI y, por un corto tiempo, en el grupo VH, mientras que los anticuerpos específicos contra Map se encontraron fácilmente no solo en el grupo VSI, sino también en los grupos VS, VH y VHI. Los avances recientes en la evaluación de las proteínas recombinantes para las pruebas serológicas de Mbv, como la utilizada aquí (MBP83), han hecho posible minimizar la reacción cruzada inducida por la vacunación paratuberculosis en el diagnóstico de tuberculosis (38, 76). Por lo tanto, el desarrollo y el uso/implementación de antígenos más específicos (reactivos DIVA) tanto para el ensayo IFN-γ como para la detección de anticuerpos son necesarios para mejorar el diagnóstico inmunológico de la paratuberculosis y la tuberculosis y para superar las interferencias producidas por la vacunación (31).
Abordando el efecto de las cepas inactivadas Silirum® y HIMB individualmente, se observó que las cabras de StrSI y StrHI mostraban un mayor número de lesiones granulomatosas que afectaban a un mayor número de animales que las de los grupos VSI o VHI. Además, a pesar de que el número de granulomas fue menor que el de las cabras de la NVI, estos también se clasificaron como b multifocales, lo que demuestra la progresión de la infección en estos grupos. En cuanto a las respuestas inmunitarias periféricas, solo el grupo StrHI mostró altos niveles periféricos de IFN-γ en respuesta a ambos antígenos micobacterianos, pero no a la producción de anticuerpos periféricos, lo que podría sugerir la capacidad de esta cepa para estimular solo la respuesta inmune celular periférica. En este sentido, en un estudio realizado en ganado, la administración oral de la cepa inactivada de Mbv 1403 suspendida en PBS no mostró un aumento de la producción de IFN-γ en respuesta al antígeno bovino, mientras que la inmunización parenteral con esta cepa homogeneizada en el adyuvante MontanideTM ISA 50 V 2 (vacuna IBR) mostró Por el contrario, no se detectaron respuestas inmunes celulares o humorales periféricas evidentes en el grupo StrSI, aunque IFN-γ aumentó a 150 dpv. En este sentido, es posible que este aumento tardío fuera el efecto de la infección por Map en lugar de la inmunización en sí, ya que los animales con la mayor respuesta inmune periférica mediada por células correspondieron a aquellos con las lesiones granulomatosas más numerosas (datos no mostrados). Por lo tanto, esta respuesta inmune periférica limitada provocada por las bacterias inactivadas podría ser una consecuencia de su débil inmunogenicidad en ausencia de adyuvantes (78, 79).
Además, la inmunización con adyuvantes (es decir, AdjSI y AdjHI) en ausencia de antígenos bacterianos dio lugar a un menor número de lesiones granulomatosas que en el grupo StrSI, StrHI o incluso NVI. Además, estas lesiones se clasificaron principalmente como a focal y multifocal, y solo un animal del AdjSI desarrollaba lesiones b multifocales. En este sentido, no se detectaron respuestas inmunes celulares o humorales periféricas significativas en grupos inmunizados adyuvantes. Esto está de acuerdo con un estudio anterior en el que no se registró una respuesta inmune periférica específica en los terneros inmunizados adyuvantes MontanideTM ISA 50 V 2 (80). Sin embargo, solo AdjSI mostró un aumento posterior en la respuesta periférica IFN-γ (150 dpv) y los anticuerpos específicos de Map (190 dpv) posiblemente asociados con la presencia de lesiones multifocales a y b en contraste con AdjHI que mostraron principalmente lesiones focales similares al grupo VSI, aunque el efecto principal de los adyuvantes no se pudo evaluar Esto es similar al aumento posterior observado en StrSI, StrHI y NVI, en los que también se detectaron lesiones b multifocales. Por lo tanto, es posible que el mayor número y extensión de las lesiones en estos grupos se correlacionaran con una mayor respuesta de los linfocitos sensibilizados que podrían haber migrado del intestino a la sangre, como se había planteado anteriormente (6).
El papel principal de los adyuvantes de aceite mineral en la vacunación es aumentar la respuesta inmune a través de la liberación sostenida del antígeno en el lugar de la inyección, asegurando la estimulación constante de la respuesta inmunitaria (78). Los adyuvantes MontanideTM, tal como se utilizan aquí, provocan un liposoma que protege el antígeno inactivado de la degradación, llevándolo a drenar los ganglios linfáticos a través del reclutamiento de células y linfocitos que presentan antígenos (79). Sin embargo, se han reportado diferencias en los perfiles de respuesta inmune dependiendo del tipo de adyuvante MontanideTM (25, 81, 82). Por ejemplo, las respuestas de anticuerpos específicas del antígeno IFN-γ y Map específicas provocadas por la vacunación de ovejas con la cepa Map 316F que mata por calor combinada con MontanideTM ISA 50 V 2 fueron más bajas que las provocadas por la vacunación con la vacuna comercial Gudair® (MontanideTM ISA 103 y 80) (25). Sin embargo, estas diferencias podrían estar relacionadas con la composición de los adyuvantes (tipo de ácidos grasos o emulsionante) y la intensidad de la respuesta inflamatoria en el lugar de la inyección, y a pesar de su importancia en la formulación de la vacuna, su interacción primaria con la respuesta inmune del huésped aún no se entiende completamente, ya que su efecto a menudo se estudia en combinación con Por esa razón, a pesar de la ausencia de diferencias estadísticas por la alta variabilidad individual, es sorprendente que el número limitado y la gravedad de las lesiones granulomatosas en los grupos AdjSI y AdjHI sin la liberación de un antígeno fueran bastante similares a los observados en los grupos VSI y VHI. En un estudio realizado en terneros, solo dos de los seis terneros inmunizados con MontanideTM ISA 50 V 2 mostraron un cultivo positivo de Mbv en los ganglios prescapulares derecho después del desafío de Mbv intranodular (80), lo que sugiere que el adyuvante podría limitar, en cierta medida, la propagación de la infección. Se sabe que la aplicación de adyuvantes de agua en aceite mostró un claro efecto estimulante sobre la respuesta inmune del huésped antes de la inmunización con un antígeno (27). Sin embargo, en ausencia de una clara respuesta inmune periférica o local específica, se trata de plantear la hipótesis de que esta respuesta inmune protectora podría ser mediada por mecanismos no específicos que podrían ayudar al huésped a controlar la infección micobacterial. Este mecanismo es desconocido, pero otras células inmunes, como las células que presentan antígenos (es decir, macrófagos, células dendríticas) podrían estar mediando en esta respuesta a medida que se reclutan en el sitio de inoculación (26) y posiblemente se están activando debido a la respuesta inflamatoria. De hecho, se ha planteado la hipótesis de que estas células pueden estar involucradas en el desarrollo de una respuesta inmune «entrenada» caracterizada por su respuesta aumentada y no específica después de la reinfección independientemente de la respuesta inmune adaptativa de la memoria (83).
En conjunto, este estudio ha evaluado los perfiles inmunológicos de dos vacunas inactivadas por calor contra la paratuberculosis y la tuberculosis antes y después del desafío de Map. Nuestros resultados destacaron el hecho de que los predictores como la respuesta periférica y local IFN-γ, la producción de anticuerpos periféricos y la evaluación periférica y local de las subpoblaciones de linfocitos no pueden evaluar el resultado de la vacunación solo mediante la evaluación efectiva de las lesiones microscópicas que ha demostrado ser un método eficaz para estimar el estado de infección (9, 11, Sin embargo, tanto la vacunación homóloga como la vacunación heteróloga pudieron conferir una alta respuesta inmune protectora contra la infección de Map, limitando la progresión de las lesiones granulomatosas y demostrando adicionalmente el beneficio potencial de la vacunación contra la tuberculosis en el control de la paratuberculosis. En este sentido, se necesitan más estudios centrados en la reducción de las interferencias en las pruebas serológicas utilizadas para el diagnóstico de la paratuberculosis y la tuberculosis, posiblemente dirigidos a rutas de vacunación alternativas o diferentes adyuvantes utilizados. Además, se observó un efecto interesante en la reducción del número y la gravedad de las lesiones granulomatosas paratuberculosis en grupos inmunizados con adyuvantes de aceite mineral refinado. Por lo tanto, se necesitan más estudios para evaluar el efecto de cada componente de la vacuna que podría estar involucrado en la protección contra las micobacterias.
Declaración de disponibilidad de datos
Los autores pondrán a disposición los datos sin perjuicios que respalden las conclusiones de este artículo.
Declaración ética
El estudio sobre animales fue revisado y aprobado por el Subcomité de Experimentos y Bienestar Animal de la Universidad de León (ULE) (OEBA-ULE-016-2017).
Contribuciones del autor
El vicepresidente, JB, DG-E y NA-V diseñaron y llevaron a cabo el experimento. JE ayudó en el análisis estadístico. VP, JB, DG-E, NA-V, RV y MF participaron en las necropsias, así como en la recolección y análisis de muestras de sangre y tejido. El mapa del inóculo y la preparación de la vacuna HIMB fueron llevados a cabo por NE e IS que, junto con NA-V, participaron en el análisis bacteriológico. El manuscrito fue escrito por VP, JB, DG-E y NA-V. La versión final enviada fue leída y aprobada por todos los autores.
Financiación
Este trabajo fue apoyado financieramente por el Ministerio de Ciencia e Innovación de España (proyectos AGL2015-66540-C2-1-R y RTI2018-099496-B-I00), la Junta de Castilla y León (proyecto LE259P18) y el Instituto Nacional de Investigación Agronómica (proyecto RTA 2017-00089-00-00). NA-V recibió un contrato predoctoral (BES-2016-076513) del Ministerio de Ciencia e Innovación español y DG-E y JE de un contrato postdoctoral del Ministerio de Ciencia e Innovación (subvenciones no. FJCI-2017-32020 y FJC2019-042422-I respectivamente).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o reclamación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Agradecimientos
Nos gustaría dar las gracias al personal del Instituto de Ganadería de Montaña (CSIC-ULE) por la entrega de los animales de experimentación. También agradecemos a CZ Vaccines por proporcionar los productos de inmunización utilizados en el estudio. Además, queremos dar las gracias al personal del laboratorio de técnicas instrumentales de la Universidad de León y a la ayuda técnica de Carmen Agudín, Marta Silva, María Teresa Carro, Elena Molina y Ainara Badiola.
Material complementario
El material complementario de este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2021.744568/full#supplementary-material
Referencias
1. Rathnaiah G, Zinniel DK, Bannantine JP, Stabel JR, Gröhn YT, Collins MT, et al. Patogénesis, genética molecular y genómica de Mycobacterium avium subsp. paratuberculosis, el agente etiológico de la enfermedad de Johne. Ciencia veterinaria frontal (2017) 4:187. doi: 10.3389/fvets.2017.00187
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
2. García AB, Shalloo L. Revisión invitada: el impacto económico y el control de la paratuberculosis en el ganado. J Dairy Sci.(2015) 98:5019–39. doi: 10.3168/jds.2014-9241
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
3. Rasmussen P, Barkema HW, Mason S, Beaulieu E, Hall DC. Pérdidas económicas debido a la enfermedad de Johne (paratuberculosis) en el ganado lechero. J Dairy Sci. (2021) 104:3123–43. doi: 10.3168/jds.2020-19381
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
4. Clarke CJ. La patología y la patogénesis de la paratuberculosis en rumiantes y otras especies. Camino de J Comp. (1997) 116:217–61. doi: 10.1016/S0021-9975(97)80001-1
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
5. Stabel JR. Transiciones en las respuestas inmunitarias a Mycobacterium paratuberculosis. Microbiol veterinario. (2000) 77:465–73. doi: 10.1016/S0378-1135(00)00331-X
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
6. Fernández M, Fuertes M, Elguezabal N, Castaño P, Royo M, Ferreras MC, et al. Expresión inmunohistoquímica del interferón-γ en diferentes tipos de lesiones granulomatosas asociadas con la paratuberculosis bovina. Comp Immunol Microbiol Infecta Dis. (2017) 51:1–8. doi: 10.1016/j.cimid.2017.01.002
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
7. Pérez V, García Marín JF, Badiola JJ. Descripción y clasificación de los diferentes tipos de lesiones asociadas con la infección natural por paratuberculosis en ovejas. Camino de J Comp. (1996) 114:107-22. doi: 10.1016/S0021-9975(96)80001-6
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
8. Corpa JM, Garrido J, García Marín JF, Pérez V. Clasificación de las lesiones observadas en casos naturales de paratuberculosis en cabras. Camino de J Comp. (2000) 122:255–65. doi: 10.1053/jcpa.1999.0368
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
9. Begg DJ. O’Brien R, Mackintosh, CG, Griffin JFT. Modelo experimental de infección para la enfermedad de Johne en ovejas.Infecta el sistema inmunológico. (2005) 73:5603-11. doi: 10.1128/IAI.73.9.5603-5611.2005
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
10. de Silva K, Begg D, Carter N, Taylor D, Di Fiore L, Whittington R. La respuesta temprana a la proliferación de linfocitos en ovejas expuestas a Mycobacterium avium subsp. paratuberculosis se compró al estado de infección. Inmunobiología.(2010) 215:12–25. doi: 10.1016/j.imbio.2009.01.014
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
11. Fernández M, Benavides J, Sevilla IA, Fuertes M, Castaño P, Delgado L, et al. Infección experimental de corderos con cepas de tipo C y S de la subespecie de Mycobacterium avium paratuberculosis: hallazgos inmunológicos y patológicos.Veterinario Res. (2014) 45:5. doi: 10.1186/1297-9716-45-5
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
12. Bastida F, Juste RA. Control de la paratuberculosis: una revisión centrada en la vacunación. J Vacunas Basadas En El Inmunológico. (2011) 9:8. doi: 10.1186/1476-8518-9-8
13. Juste R, Pérez V. Control de la paratuberculosis en ovejas y cabras. Vet Clin Food Anim. (2011) 27:127–38. doi: 10.1016/j.cvfa.2010.10.020
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
14. Eppleston J, Reddacliff L, Windsor P, Links I, Whittington R. Observaciones preliminares sobre la prevalencia de la excreción de ovejasMycobacterium avium subsp paratuberculosis después de 3 años de un programa de vacunación para la enfermedad de las ovejas Johne. Aust Vet J. (2005) 83:637–8. doi: 10.1111/j.1751-0813.2005.tb13279.x
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
15. Juste RA, Alonso-Hearn M, Molina E, Geijo M, Vazquez P, Sevilla IA, et al. Reducción significativa de la eliminación bacteriana y mejora en la producción de leche en las granjas lecheras después del uso de una nueva vacuna paratuberculosa inactivada en un ensayo de campo. Notas de la Reca de BMC. (2009) 2:233. doi: 10.1186/1756-0500-2-233
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
16. Reddacliff L, Eppleston J, Windsor P, Whittington R, Jones S. Eficacia de una vacuna muerta para el control de la paratuberculosis en rebaños de ovejas australianas. Microbiol veterinario. (2006) 115:77-90. doi: 10.1016/j.vetmic.2005.12.021
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
17. Sweeney RW, Whitlock RH, Bowersock TL, Cleary DL, Meinert TR, Habecker PL, et al. Efecto de la administración subcutánea de una vacuna paratuberculosis Mycobacterium avium subsp muerta en la colonización de tejidos después de la exposición oral al organismo en terneros. Am J Vet Res. (2009) 70:493–7. doi: 10.2460/ajvr.70.4.493
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
18. Alonso-Hearn M, Molina E, Geijo M, Vazquez P, Sevilla IA, Garrido JM, et al. La inmunización del ganado lechero adulto con una nueva vacuna asesinada por calor se asocia con una vida productiva más larga antes de que las vacas sean enviadas a la matanza con sospecha de paratuberculosis. J Dairy Sci. (2012) 95:618–29. doi: 10.3168/jds.2009-2860
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
19. Gwozdz JM, Thompson KG, Manktelow BW, Murray A, West DM. Vacunación contra la paratuberculosis de corderos ya infectados experimentalmente con la subespecie de Mycobacterium avium paratuberculosis. Aust Vet J. (2000) 78:560–6. doi: 10.1111/j.1751-0813.2000.tb11902.x
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
20. Begg DJ, Griffin JFT. Vacunación de ovejas contra M. paratuberculosis: parámetros inmunológicos y eficacia protectora. Vacuna. (2005) 23:4999–5008. doi: 10.1016/j.vaccine.2005.05.031
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
21. Mercier P, Brémaud I, Gautier MP. Vacunación de niños menores de un mes con una vacuna muerta y reducción de la frecuencia de excreción fecal de la subespecie de Mycobacterium avium paratuberculosis. Rumino pequeño Res. (2014) 121:425–33. doi: 10.1016/j.smallrumres.2014.09.007
22. de Silva K, Plain KM, Begg DJ, Purdie AC, Whittington RJ. Las células T CD4+, las células T γδ y las células B están asociadas con la falta de protección contra la vacuna en la infección por la subcuaspecie de Mycobacterium avium paratuberculosis.Vacuna. (2015) 33:149-55. doi: 10.1016/j.vaccine.2014.10.082
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
23. Dhand NK, Eppleston J, Whittington RJ, Windsor PA. Cambios en la prevalencia de la paratuberculosis ovina después de la vactura con Gudair®: resultados de un estudio longitudinal realizado durante una década. Vacuna. (2016) 34:5107-13. doi: 10.1016/j.vaccine.2016.08.064
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
24. Watkins C, Schock A, May L, Denham S, Sales J, Welch L, et al. Evaluación de la virulencia de las cepas de la vacuna de la subespecie de Mycobacterium avium paratuberculosis en un modelo de ternero. Microbiol veterinario. (2010) 146:63–9. doi: 10.1016/j.vetmic.2010.04.017
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
25. Begg DJ, Dhungyel O, Naddi A, Dhand NK, Plain KM, de Silva K, et al. La inmunogenicidad y la reactividad tisular de la vacuna de células enteras inactivada por Mycobacterium avium subsp paratuberculosis dependen del adyuvante utilizado.Heliyon. (2019) 5:e01911. doi: 10.1016/j.heliyon.2019.e01911
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
26. Aucouturier J, Dupuis L, Ganne V. Adyuvantes diseñados para vacunas veterinarias y humanas. Vacuna. (2001) 19:2666–72. doi: 10.1016/S0264-410X(00)00498-9
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
27. van der Heijden PHJ, Bokhout BA, Bianchi ATJ, Scholten JW, Stok W. Aplicación separada de adyuvante y antígeno: el efecto de una emulsión de agua en aceite en la respuesta celular formadora de placa esplénica a los glóbulos rojos de las ovejas en ratones. Inmunobiol. (1986) 171:143–54. doi: 10.1016/S0171-2985(86)80023-7
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
28. Danielsson R, Eriksson H. Adyuvantes de aluminio en las vacunas, una forma de modular la respuesta inmunitaria. Semin Cell Dev Biol. (2021) 115:3–9. doi: 10.1016/j.semcdb.2020.12.008
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
29. Li X, Xing R, Xu C, Liu S, Qin Y, Li K, et al. Efecto inmunoestimulador del quitosano y el quitosano cuaternario: una revisión de los posibles adyuvantes de la vacuna. Carbohydr Polym. (2021) 264:118050. doi: 10.1016/j.carbpol.2021.118050
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
30. Garrido JM, Vazquez P, Molina E, Plazaola JM, Sevilla IA, Geijo MV, et al. La vacunación contra la paratuberculosis solo causa una reactividad cruzada limitada en la prueba cutánea para el diagnóstico de la tuberculosis bovina. PLoS ONE. (2013) 8:e80985. doi: 10.1371/journal.pone.0080985
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
31. Serrano M, Elguezabal N, Sevilla IA, Geijo MV, Molina E, Arrazuria R, et al. Detección de tuberculosis en terneros vacunados por paratuberculosis: nuevas alternativas contra la interferencia. PLoS ONE. (2017) 12:e0169735. doi: 10.1371/journal.pone.0169735
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
32. Pérez de, Val B, Nofrarías M, López-Soria S, Garrido JM, Vordermeier HM, Villareal-Ramos B, Martín M, Puentes E, Juste RA, Domingo M. Efectos de la vacunación contra la paratuberculosis en la tuberculosis en cabras: interferencias diagnósticas y protección cruzada. BMC Vet Res. (2012) 8:191. doi: 10.1186/1746-6148-8-191
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
33. Serrano M, Elguezabal N, Sevilla IA, Geijo MV, Molina E, Juste RA, et al. Los resultados preliminares indican que la vacuna inactivada contra la paratuberulosis podría modificar el curso de la infección experimental por Mycobacterium bovis en los terneros. Ciencia veterinaria frontal (2017) 4:175. doi: 10.3389/fvets.2017.00175
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
34. Garrido JM, Sevilla IA, Beltrán-Beck B, Minguijón E, Ballesteros C, Galindo RC, et al. Protección contra la tuberculosis en el jabalí euroasiático vacunado con Mycobacterium bovis inactivado por el calor. PloS ONE. (2011) 6:e24905. doi: 10.1371/journal.pone.0024905
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
35. Arteche-Villasol N, Benavides J, Espinosa J, Vallejo R, Royo M, Ferreras MC, et al. Aislamiento optimizado in vitro de diferentes subpoblaciones de células inmunes de la sangre periférica y técnicas comparativas para la generación de macrófagos derivados de monocitos en pequeños rumiantes. Inmunopático inmune veterinario. (2020) 230:110131. doi: 10.1016/j.vetimm.2020.110131
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
36. Delgado L, Juste RA, Muñoz M, Morales S, Benavides J, Ferreras MC, et al. Diferencias en la respuesta inmune periférica entre corderos y ovejas adultas infectadas experimentalmente con Mycobacterium avium suspecies paratuberculosis.Inmunopático inmune veterinario. (2012) 145:23-31. doi: 10.1016/j.vetimm.2011.10.005
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
37. Vidal E, Arrieta-Villegas C, Grasa M, Mercader I, Domingo M, Pérez de Val B. Evaluación de campo de la eficacia de la vacuna Mycobacterium bovis BCG contra la tuberculosis en cabras. BMC Vet Res. (2017) 13:252. doi: 10.1186/s12917-017-1182-5
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
38. Roy Á, Infantes-Lorenzo JA, Blázquez JC, Venteo A, Mayoral FJ, Domínguez M, et al. Análisis temporal de la interferencia causada por la vacunación paratuberculosis en las pruebas de diagnóstico de tuberculosis en cabras. Prev Vet Med. (2018) 156:68-75. doi: 10.1016/j.prevetmed.2018.05.010
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
39. Lybeck KR, Storset AK, Djønne B, Valheim M, Olsen I. La excreción fecal se detectó antes de las respuestas inmunitarias en cabras infectadas naturalmente con Mycobacterium avium subsp. paratuberculosis. Res Vet Sci. (2011) 91:32–9. doi: 10.1016/j.rvsc.2010.08.012
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
40. Arteche-Villasol N, Gutiérrez-Expósito D, Vallejo R, Espinosa J, Elguezabal N, Ladero-Auñon I, et al. Respuesta temprana de los macrófagos derivados de monocitos de cabras vacunadas y no vacunadas contra la infección in vitro con Mycobacterium avium subsp. paratuberculosis. Veterinario Res. (2021) 52:69. doi: 10.1186/s13567-021-00940-y
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
41. Hines II ME, Turnquist SE, Ilha MRS, Rajeev S, Jones AL, Whittington L, et al. Evaluación de nuevos candidatos a vacunas orales y validación de un modelo caprino de la enfermedad de Johne. La Célula Frontal Infecta El Microbiol. (2014) 4:26. doi: 10.3389/fcimb.2014.00026
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
42. Plain KM, Marsh IB, Waldron AM, Galea F, Whittington AM, Saunders VF, et al. Ensayo de PCR fecal directa de alto rendimiento para la detección de Mycobacterium avium subsp. paratuberculosis en ovejas y ganado. J Clin Microbiol.(2014) 52:745–57. doi: 10.1128/JCM.03233-13
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
43. Rodríguez-Lázaro D, D’Agostino M, Herrewegh A, Pla M, Cook N, Ikonomopoulos J. Métodos basados en PCR en tiempo real para la detección de Mycobacterium avium subsp. paratuberculosis en agua y leche. Int J Food Microbiol. (2005) 101:93–104. doi: 10.1016/j.ijfoodmicro.2004.09.005
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
44. Adúriz JJ, Juste RA, Cortabarria N. Falta de dependencia de micobactina de las micobacteriáceas aisladas en Middlebrook 7H11 de casos clínicos de paratuberculosis ovina. Microbiol veterinario. (1995) 45:211–7. doi: 10.1016/0378-1135(95)00037-B
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
45. Sevilla IA, Garrido JM, Molina E, Geijo MV, Elguezabal N, Vázquez P, et al. Desarrollo y evaluación de una nueva PCR triplex dirigida a elementos multicopias para la detección de Mycobacterium avium subsp. paratuberculosis en las heces. Appl Environ Microbiol. (2014) 80:3757–68. doi: 10.1128/AEM.01026-14
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
46. Delgado L, García Marín JF, Muñoz M, Benavides J, Juste RA, García-Pariente C, et al. Hallazgos patológicos en ovejas jóvenes y adultas después de una infección experimental con 2 dosis diferentes de la subespecie de Mycobacterium avium paratuberculosis.Vet Pathol. (2013) 50:857–66. doi: 10.1177/0300985813476066
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
47. Rosseels V, Huygen K. Vacunación contra la paratuberculosis. Vacunas de expertos en Rev. (2008) 7:817–32. doi: 10.1586/14760584.7.6.817
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
48. Gupta S, Singh SV, Singh M, Chaubey KK, Karthik K, Bhatia AK, et al. Enfoques de vacunación para el «gestión terapéutica» de la infección por paratuberculosis de la subespecie de Mycobacterium avium en el ganado doméstico. Veterinario P. (2019) 39:143–52. doi: 10.1080/01652176.2019.1667042
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
49. Krüger C, Köhler H, Liebler-Tenorio EM. Desarrollo secuencial de lesiones 3, 6, 9 y 12 meses después de la infección experimental de niños caprinos con Mycobacterium avium subsp paratuberculosis. Vet Pathol. (2015) 52:276-90. doi: 10.1177/0300985814533804
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
50. Juste RA, García Marín JF, Peris B, Sáez de, Ocáriz C, Badiola JJ. Infección experimental de corderos vacunados y no vacunados con Mycobacterium paratuberculosis. Camino de J Comp. (1994) 110:185–94. doi: 10.1016/S0021-9975(08)80189-2
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
51. Faisal, SM. Chen JW, Yan F, Chen TT, Useh NM, Yan W, et al. Evaluación de un mutante de Mycobacterium avium subsp paratuberculosis leuD como candidato a vacuna contra el desafío en un modelo caprino. Inmunidad de la vacuna Clin.(2013) 20:572–81. doi: 10.1128/CVI.00653-12
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
52. Corpa JM, Pérez V, García Marín JF. Diferencias en las respuestas inmunitarias en corderos y niños vacunados contra la paratuberculosis, según la edad de vacunación. Microbiol veterinario. (2000) 77:475–85. doi: 10.1016/S0378-1135(00)00332-1
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
53. Storset AK, Hasvold HJ, Valheim M, Brun-Hansen H, Berntsen G, Whist SK, et al. Prensa CMcL, Holstad G, Larsen HJS. Paratuberculosis subclínica en cabras después de una infección experimental: un estudio inmunológico y microbiológico.Inmunopático inmune veterinario. (2001) 80:271–87. doi: 10.1016/S0165-2427(01)00294-X
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
54. Vazquez P, Garrido JM, Juste RA. Respuesta específica de antiodiódica e interferón-gamma asociada con formas inmunobiológicas de paratuberculosis bovina en ganado frisón sacrificado. PLoS ONE. (2013) 8:e64568. doi: 10.1371/journal.pone.0064568
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
55. Koets A, Ravesloot L, Ruuls R, Dinkla A, Eisenberg S, Lievaart-Peterson K. Efectos de la edad y el medio ambiente en las respuestas inmunitarias adaptativas a la vacunación contra la paratuberculosis de Mycobacterium avium subsp. (MAP) en cabras lecheras en relación con las estrategias de control de la paratuberculosis. Veterinario de ciencia. (2019) 6:62. doi: 10.3390/vetsci6030062
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
56. Pérez V, Tellechea J, Badiola JJ, Gutiérrez M, García Marín JF. Relación entre la respuesta serológica y los hallazgos patológicos en ovejas con paratuberculosis adquirida de forma natural. Am J Vet Res. (1997) 58:799–803. Disponible en línea en: https://www.avma.org/Pages/home.aspx
57. Pollock JM, McNair J, Welsh MD, Girvin RM, Kennedy HE, Mackie DP, et al. Respuesta inmune en la tuberculosis bovina. Tuberculosis. (2001) 81:103–7. doi: 10.1054/tube.2000.0258
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
58. Waters WR, Miller JM, Palmer MV, Stabel JR, Jones DE, Koistinen KA, et al. Inducción temprana de las respuestas inmunitarias humorales y celulares durante la infección experimental por micobacterium avium subsp. paratuberculosis de los terneros. Infecta el sistema inmunológico. (2003) 71:5130–8. doi: 10.1128/IAI.71.9.5130-5138.2003
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
59. Begg DJ, de Silva K, Carter N, Plain KM, Purdie A. Whittington RJ. ¿Existe realmente un dominio Th1 sobre Th2 en las primeras etapas de las infecciones por paratuberculosis de la subespecie de Mycobacterium avium? Inmunobiología. (2011) 216:840–6. doi: 10.1016/j.imbio.2010.12.004
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
60. Pooley HB, Begg DJ, Plain KM, Whittington RJ, Purdie AC, de Silva K. La respuesta inmune humoral es esencial para una protección vacunal exitosa contra la paratuberculosis en ovejas. BMC Vet Res. (2019) 15:223. doi: 10.1186/s12917-019-1972-z
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
61. Storset AK, Berg I, Djønne B. Evaluación de la prueba de interferón gamma para el diagnóstico de la paratuberculosis en cabras. Inmunopático inmune veterinario. (2005) 107:87–94. doi: 10.1016/j.vetimm.2005.03.015
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
62. Denis M, Wedlock DN, Buddle BM. Capacidad de los subconjuntos de células T y sus mediadores solubles para modular la replicación de Mycobacterium bovis en macrófagos bovinos. Inmunol celular. (2004) 232:1–8. doi: 10.1016/j.cellimm.2005.01.003
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
63. Gillan S, O’Brien R, Hughes AD, Griffin JFT. Identificación de parámetros inmunes para diferenciar los estados de enfermedad entre las ovejas infectadas con Mycobacterium avium subsp. paratuberculosis. Inmunidad de la vacuna Clin.(2010) 17:108-17. doi: 10.1128/CVI.00359-09
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
64. Frie MC, Sporer KRB, Kirkpatrick BW, Coussens PM. Perfiles de activación de células T y B de vacas con y sin enfermedad de Johne en respuesta a la estimulación in vitro con la subespecie de Mycobacterium avium paratuberculosis. Inmunopático inmune veterinario. (2017) 193–4:50–6. doi: 10.1016/j.vetimm.2017.10.005
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
65. Stabel JR, Bannantine JP. Respuestas inmunitarias celulares divergentes específicas del antígeno durante los estados subclínicos y clincal asintomáticos de la enfermedad en vacas infectadas naturalmente con Mycobacterium avium subsp. paratuberculosis. Infec Immun. (2020) 88:e00650-19. doi: 10.1128/IAI.00650-19
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
66. Arrieta-Villegas C, Perálvarez T, Vidal E, Puighibet Z, Moll X, Canturri A, et al. Eficacia de la vacunación parenteral contra la tuberculosis con Mycobacterium bovis inactivado por el calor en cabras de desafío experimental. PLoS ONE. (2018) 13:e0196948. doi: 10.1371/journal.pone.0196948
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
67. Kurade NP, Tripathi BN, Rajukumar K, Parihar NS. Desarrollo secuencial de lesiones histológicas y su relación con el aislamiento bacteriano, la desprendimiento fecal y las respuestas inmunitarias durante las etapas progresivas de la infección experimental de corderos con Mycobacterium avium subsp. Paratuberculosis. Vet Pathol. (2004) 41:378–87. doi: 10.1354/vp.41-4-378
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
68. Taniguchi Y, Sakakibara S, Fujihara M, Yagi A, Fujiyoshi S. La asociación entre la detección de Mycobacterium avium subsp.paratuberculosis DNA en las heces y la clasificación histopatológica. J Vet Med Sci. (2020) 82:541–5. doi: 10.1292/jvms.18-0724
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
69. Whittington RJ, Fell S, Walker D, McAllister S, Marsh I, Sergeant E, et al. Uso de cultivos fecales agrupados para la detección sensible y económica de la infección por paratuberculosis de Mycobacterium avium subsp. en rebaños de ovejas. J Clin Microbiol. (2000) 38:2550–6. doi: 10.1128/JCM.38.7.2550-2556.2000
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
70. Mortier RA, Barkema HW, Orsel K, Wolf R, De Buck J. Patrones de desprendimiento de terneros lácteos infectados experimentalmente con la subespecie de Mycobacterium avium paratuberculosis. Veterinario Res. (2014) 45:71. doi: 10.1186/s13567-014-0071-1
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
71. González J, Geijo MV, García-Pariente C, Verna A, Corpa JM, Reyes LE, et al. Clasificación histopatológica de las lesiones asociadas con la infección natural por paratuberculosis en el ganado. Camino de J Comp. (2005) 133:184–96. doi: 10.1016/j.jcpa.2005.04.007
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
72. Roy Á, Risalde MA, Bezos J, Casal C, Romero B, Sevilla I, et al. Respuesta de las cabras a la vacunación intramuscular con Mycobacterium bovis muerto por el calor y desafío natural. Comp Immunol Microbiol Infecta Dis. (2018) 60:28-34. doi: 10.1016/j.cimid.2018.09.006
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
73. Muskens J, van Zijderveld F, Eger A, Bakker D. Evaluación de la respuesta inmunitaria a largo plazo en el ganado después de la vacunación contra la paratuberculosis en dos rebaños lácteos holandeses. Microbiol veterinario. (2002) 86:269–78. doi: 10.1016/S0378-1135(02)00006-8
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
74. Thom M, Howard C, Villarreal-Ramos B, Mead E, Vordermeier M, Hope J. Consecuencia de la exposición previa a las micobacterias ambientales en la vacunación contra el BCG y el diagnóstico de la infección por tuberculosis. Tuberculosis.(2008) 88:324-34. doi: 10.1016/j.tube.2007.12.002
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
75. Álvarez J, de Juan L, Bezos J, Romero B, Sáez JL, Marqués S, et al. Efecto de la paratuberculosis en el diagnóstico de tuberculosis bovina en un rebaño de ganado con una infección mixta utilizando el ensayo de detección de interferón-gamma. Microbiol veterinario. (2009) 135:389–93. doi: 10.1016/j.vetmic.2008.09.060
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
76. Tewari D, Hoving E, Linscott R, Martel E, Lawrence J, Wolfgang D, et al. Respuesta de anticuerpos paratuberculosis de Mycobacterium avium subsp., desprendimiento fecal y reactividad cruzada de anticuerpos a Mycobacterium bovis en manadas infectadas por M avium subspparatuberculosisvacunadas contra la enfermedad de Johne. Inmunidad de la vacuna Clin. (2014) 21:698–703. doi: 10.1128/CVI.00032-14
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
77. Jones GJ, Steinbach S, Sevilla IA, Garrido JM, Juste R, Vordermeier HM. La vacunación oral de ganado con inactivado por calor Mycobacterium bovis no compromete las pruebas de diagnóstico de tuberculosis bovina. Inmunopático inmune veterinario. (2016) 182:85–8. doi: 10.1016/j.vetimm.2016.10.010
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
78. Coffman RL, Sher A, Seder RA. Adyuvantes de la vacuna: poner en trabajo la inmunidad innata. Inmunidad. (2010) 33:492–503. doi: 10.1016/j.immuni.2010.10.002
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
79. Awate S, Babiuk LA, Mutwiri G. Mecanismo de acción de los adyuvantes. Inmunidad frontal. (2013) 4:114. doi: 10.3389/fimmu.2013.00114
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
80. van der Heijden EMDL, Chileshe J, Vernooij JCM, Gortazar C, Juste RA, Sevilla I, et al. Perfiles de respuesta inmune de los terneros después de la vacunación con BCG vivo y candidatos a la vacuna Mycobacterium bovis inactivados. PLoS ONE. (2017) 12:e0188448. doi: 10.1371/journal.pone.0188448
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
81. Vordermeier HM, Dean GS, Rosenkrands I, Agger EM, Andersen P, Kaveh DA, et al. Los adyuvantes inducen fenotipos inmunológicos distintos en un modelo de vacuna contra la tuberculosis bovina. Inmunidad de la vacuna Clin. (2009)16:1443–8. doi: 10.1128/CVI.00229-09
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
82. Klimka A, Michels L, Glowalla E, Tosetti B, Krönke M, Krut O. La montanida ISA 71 VG es ventajosa para el adyuvante de Freund en la inmunización contra la infección por S. aureus de ratones. Scand J Immunol. (2015) 81:291–7. doi: 10.1111/sji.12279
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
83. Netea MG. Entrenamiento de la inmunidad innata: el concepto cambiante de la memoria inmunológica en la defensa innata del huésped. Eur J Clin Invest. (2013) 43:881–4. doi: 10.1111/eci.12132
Resumen de PubMed | Texto completo de CrossRef | Google Scholar
Palabras clave: vacunación, paratuberculosis, tuberculosis, HIMB, adyuvante, protección cruzada
Cita: Arteche-Villasol N, Gutiérrez-Expósito D, Elguezabal N, Sevilla IA, Vallejo R, Espinosa J, Ferreras MC, Benavides J y Pérez V (2022) Influencia de las vacunas heterogéneas y homólogas, y sus componentes, en la respuesta inmune del huésped y la protección contra la paratuberculosis de cabra experimental Delante. Veterinario. Ciencia. 8:744568. doi: 10.3389/fvets.2021.744568
Recibido: 20 de julio de 2021; aceptado: 02 de diciembre de 2021;
Publicado: 05 de enero de 2022.
Editado por:
Francisco Javier Salguero, Salud Pública de Inglaterra, Reino Unido
Revisado por:
Beatriz Vidana, Universidad de Bristol, Reino Unido
Mourad Tayebi, Universidad del Oeste de Sydney, Australia
Copyright © 2022 Arteche-Villasol, Gutiérrez-Expósito, Elguezabal, Sevilla, Vallejo, Espinosa, Ferreras, Benavides y Pérez.
*Correspondencia: Noive Arteche-Villasol, nartv@unileon.es
†Estos autores han contribuido por igual a este trabajo
Descargo de responsabilidad: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamación que pueda ser hecha por su fabricante no está garantizado o respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía