
La suplementación con ácido benzoico mejora el rendimiento del crecimiento, la digestibilidad y el metabolismo del nitrógeno de los corderos

La suplementación con ácido benzoico mejora el rendimiento del crecimiento, la digestibilidad de los nutrientes y el metabolismo del nitrógeno de los corderos destetados
 Wenjie Zhang1
Wenjie Zhang1  Shuo Sol1 Yaqian Zhang1 Yanan Zhang1 Jianguo Wang2 Zhiqiang Liu2
Shuo Sol1 Yaqian Zhang1 Yanan Zhang1 Jianguo Wang2 Zhiqiang Liu2  Kailun Yang1*
Kailun Yang1*- 1Laboratorio Clave de Nutrición de Herbívoros para la Producción de Carne y Leche de Xinjiang, Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Xinjiang, Ürümqi, China
- 2Xinjiang Shangpin Meiyang Technology Co., Ltd., Changji, China
El nitrógeno es uno de los componentes esenciales de las proteínas y los ácidos nucleicos y desempeña un papel crucial en el crecimiento y desarrollo de los rumiantes. Sin embargo, la tasa de utilización de nitrógeno de los rumiantes es menor que la de los animales monogástricos, lo que no solo reduce la conversión y utilización de proteínas, sino que también aumenta la descarga de nitrógeno del estiércol y causa contaminación ambiental. La etapa de cordero es un período importante en la vida de las ovejas, que puede afectar el rendimiento de la producción y la calidad de la carne de las ovejas de engorde. El propósito de este experimento fue explorar los efectos de la suplementación con ácido benzoico sobre el rendimiento del crecimiento, la digestibilidad de los nutrientes, el metabolismo del nitrógeno y los parámetros plasmáticos de los corderos destetados. Un total de 40 corderos machos destetados de oveja Hu con peso corporal similar se dividieron aleatoriamente en 4 grupos: control sin ácido benzoico (0 BA) y los corderos de otros 3 grupos fueron alimentados con 0,5, 1 y 1,5% de ácido benzoico sobre la base de una dieta experimental (0,5, 1 y 1,5 BA, respectivamente). El experimento duró 60 días. Los resultados mostraron que la ganancia media diaria del grupo de 1 BA se incrementó significativamente (p < 0,05) en comparación con los grupos de 0 y 1,5 BA, mientras que se observó una tendencia opuesta de la relación entre la ingesta de materia seca y la ganancia media diaria. La digestibilidad de la materia seca, la materia orgánica, la fibra detergente neutra y la fibra detergente ácida del grupo 1 BA se incrementó significativamente (p < 0,05) en comparación con los grupos 0 y 1,5 BA, así como el contenido plasmático de albúmina. Además, las concentraciones urinarias de ácido hipúrico y nitrógeno hipurato en los grupos de 1 y 1,5 BA fueron mayores (p < 0,05) que en los grupos de 0 y 0,5 BA. Además, la ingesta de nitrógeno en los grupos 0,5 y 1 BA aumentó significativamente (p < 0,05) en comparación con otros grupos. A la 1 h después de la alimentación matutina, la concentración plasmática de ácido benzoico del grupo 1 BA alcanzó el valor máximo y fue mayor (p < 0,05) que otros grupos, y luego comenzó a disminuir. Del mismo modo, la concentración de ácido hipúrico en plasma de los grupos 1 y 1,5 BA fue mayor (p < 0,05) que la del grupo 0 BA de 1 a 4 h después de la alimentación matutina. A las 3 h después de la alimentación, la concentración de nitrógeno ureico en el plasma del grupo 0 BA fue mayor (p < 0,05) que la del grupo 1,5 BA. En general, la suplementación adecuada de ácido benzoico (1%) en la dieta puede mejorar el rendimiento del crecimiento y el metabolismo del nitrógeno de los corderos destetados.
1 Introducción
Con la escasez de recursos alimenticios y el agravamiento adicional de la contaminación ambiental, es difícil aumentar la producción de alimentos de los animales simplemente ampliando el número de cabezas reproductoras e ignorando la contaminación ambiental. La innovación tecnológica es una de las principales formas de transformar el modo de producción de los animales y mejorar la tasa de producción y la tasa de utilización de los recursos de los animales, lo que es beneficioso para el establecimiento de una sociedad que ahorre recursos y sea respetuosa con el medio ambiente en el futuro (1). Una gran cantidad de sustancias nitrogenadas excretadas por el ganado es una causa importante de contaminación ambiental en la ganadería (2, 3). Estas sustancias nitrogenadas se derivan principalmente de la proteína bruta no digerida y de la degradación de los aminoácidos en los piensos (4, 5). El nitrógeno es uno de los componentes esenciales de las proteínas y los ácidos nucleicos y desempeña un papel crucial en el crecimiento y desarrollo de los rumiantes. Sin embargo, con el sistema de alimentación dietética actual, la tasa de utilización de nitrógeno de los rumiantes es menor que la de los animales monogástricos (6). La tasa de utilización de nitrógeno de los rumiantes es de aproximadamente 20 a 36%, y el 64 a 80% restante de nitrógeno se excreta en forma de heces y orina (7), lo que no solo reduce la conversión y utilización de proteínas, sino que también aumenta la descarga de nitrógeno del estiércol (8-10). La menor utilización de nitrógeno por parte de los rumiantes restringe gravemente los beneficios económicos de las explotaciones ganaderas. Por lo tanto, el aumento de la eficiencia de la conversión y utilización de nitrógeno en rumiantes mediante una estrategia nutricional es de gran importancia para la industria de los rumiantes, así como para la reducción de la contaminación ambiental (11).
El ácido hipúrico (AH), también conocido como benzoilglicina, es el conjugado de glicina del ácido benzoico (BA), y se encuentra en altas concentraciones en la orina de los herbívoros (12). En el caso de los herbívoros, incluidos los rumiantes, los polifenoles de las dietas basadas en plantas pueden formar BA bajo la acción conjunta de microorganismos intestinales, que posteriormente se absorben a través del intestino y se transportan al hígado para su metabolismo (13). En las mitocondrias hepáticas de los animales, el BA y la glicina son catalizados por enzimas para formar HA, que luego se excreta en la orina (14). En las primeras investigaciones, Doak (15) encontró que el nitrógeno ureico representaba el 76,4% del nitrógeno urinario en el oeste, y las proporciones de nitrógeno alantoína y nitrógeno HA eran de 4,1 y 2,6%, respectivamente. El estudio de Bristow et al. analizó el contenido de nitrógeno en la orina de bovinos y ovinos, y encontró que el 69% del nitrógeno total en la orina bovina estaba en forma de urea, el 7,3% en forma de alantoína y el 5,8% en forma de HA. En la orina de oveja, el nitrógeno en forma de urea representó el 83% del contenido total de nitrógeno, y tanto el nitrógeno HA como el nitrógeno alantoína representaron el 4,3% del nitrógeno total (16). Por lo tanto, el nitrógeno excretado por HA se puede utilizar como otra forma de excreción de nitrógeno en rumiantes.
El BA es un precursor importante en el proceso de formación de AH, y su contenido afecta directamente a la excreción de HA. Como compuesto orgánico de ácido aromático débilmente ácido, el BA se usa ampliamente como acidificante orgánico y conservante en la industria de alimentos, medicamentos y piensos debido a su amplio efecto antibacteriano y su fuerte capacidad antibacteriana (17, 18). El BA puede mejorar el rendimiento del crecimiento y la conversión alimenticia de los animales monogástricos al promover la producción y activación de enzimas digestivas gastrointestinales (19), mejorar la capacidad de absorción intestinal (20), mejorar la barrera intestinal (21) y regular la microbiota intestinal (22). Sin embargo, se ha prestado poca atención a los efectos del BA sobre el metabolismo de la urea, el metabolismo del nitrógeno y el rendimiento del crecimiento de los rumiantes. Cabe destacar que tanto la síntesis de AH como el ciclo de la urea se producen en las mitocondrias hepáticas de los rumiantes. Entre ellos, el ciclo de la urea consta principalmente de 1 molécula de NH3 y 1 molécula de CO2 catalizado por la carbamilo fosfato sintetasa I para producir carbamilo fosfato (23). El NH +4 4+ produced by glutamine in mitochondria can be used to synthesize carbamyl phosphate as well as glycine. When the entry of BA into liver cells increases, HA synthesis is elevated, and the consumption of glycine correspondingly increases, thus up-regulating the synthesis of glycine in liver mitochondria. If the amount of NH +4 4+ in the mitochondria used to synthesize glycine increases, does the amount used to synthesize carbamyl phosphate decrease, then reducing urea production? However, there is a lack of relevant research at present. Therefore, this study was carried out to evaluate the effects of different levels of BA supplementation on nutrient digestibility, urea metabolism, nitrogen metabolism and growth performance of weaned lambs.
2 Materiales y métodos
2.1 Declaración ética
Todos los procedimientos de cuidado y manejo de animales en este estudio se llevaron a cabo bajo la guía del Cuidado y Uso de Animales de Laboratorio en China y fueron aprobados por (número de protocolo: 2020022) el Comité de Cuidado de Animales de la Universidad Agrícola de Xinjiang (Urumqi, Xinjiang, China).
2.2 Animales de experimentación y gestión de la alimentación
El experimento con animales se llevó a cabo en una granja comercial de ovejas ubicada en el Parque de Ciencia y Tecnología Agrícola, Changji, China. Se utilizaron un total de 40 corderos machos sanos de oveja Hu con edad y peso corporal similares (peso corporal, 17,27 ± 1,52 kg) después del destete. Después de marcar con marcas en las orejas, los corderos se asignaron aleatoriamente a 4 grupos, cada uno con 10 animales. Todos los corderos fueron alimentados con la misma dieta basal que fue formulada de acuerdo con el NRC (24). Las composiciones de los piensos y los niveles de nutrientes de la dieta experimental se presentan en la Tabla 1. Los corderos de cada grupo se suplementaron con 0, 0,5, 1 y 1,5% de BA (Comprado a Henan Xizheng Industry Co., China; Pureza ≥ 99,5%) en la dieta basal, y los tratamientos fueron etiquetados como grupos 0, 0,5, 1 y 1,5 BA, respectivamente. La cantidad aditiva de BA se basó en los estudios previos en ganado vacuno (25) y cerdos de engorde (26).
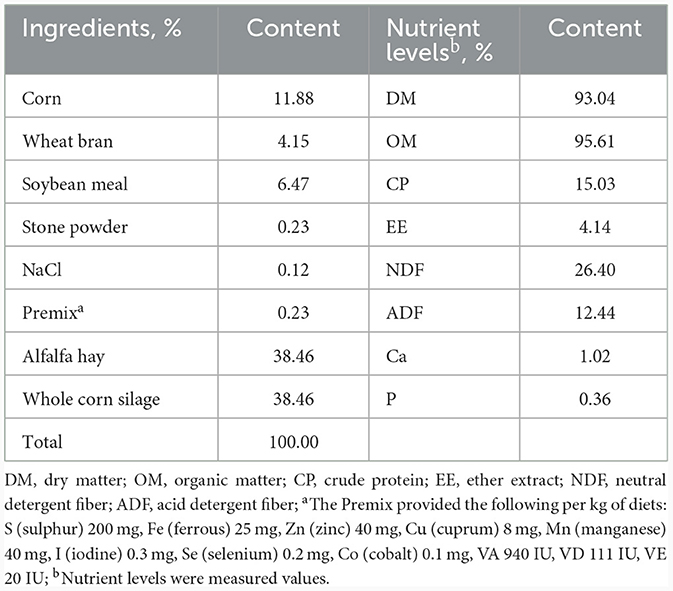 Tabla 1. Ingredientes de los piensos y niveles de nutrientes de la dieta (base MS).
Tabla 1. Ingredientes de los piensos y niveles de nutrientes de la dieta (base MS).
El presente estudio se realizó de febrero a abril de 2023. Todos los animales de 4 tratamientos fueron criados en 40 corrales con 1 cordero en cada corral (1 × 1,2 m). Los 40 corrales se ubicaron dentro de un establo abierto por dos lados y dispuestos en dos filas de 20, separadas por el carril central de alimentación. Los corrales están rodeados por barras metálicas horizontales, que también delimitan los corrales en el carril de alimentación. El piso tenía una base de concreto cubierta con lecho de paja de cebada, del cual se añadía una escama fresca (alrededor de 1,5 kg) por corral sobre el lecho permanente una vez al día. Los corderos estaban desatados y no tenían acceso a una zona de potrero. El BA se mezcló completamente con la dieta basal. Los corderos fueron alimentados dos veces al día a las 10:00 y a las 17:00, respectivamente, permitiendo entre un 5 y un 10% de orts, y se les dio libre acceso a agua potable. Antes de la estela de alimentación, se limpió y esterilizó el cobertizo experimental y se eliminaron los parásitos. A una fase adaptativa de 5 días le siguieron 60 días de período experimental.
2.3 Medición del rendimiento del crecimiento
Antes de la alimentación matutina, el peso corporal de todos los corderos se midió en d 0, 30 y 60, y la ganancia media diaria (GMD) se calculó por peso corporal inicial y final. El consumo de alimento se registró de acuerdo con la diferencia de alimento ofrecido y rechazado y se convirtió en consumo de materia seca (MSI). La relación de conversión alimenticia (F:G) se determinó dividiendo el IMD por la GMD.
2.4 Recogida de muestras de orina, heces y sangre
Del día 50 al 55 del experimento, se seleccionaron aleatoriamente 6 corderos de cada grupo para recolectar muestras de orina y heces. Se utilizó un dispositivo de recolección de orina de fabricación propia para recolectar muestras de orina de cordero, y la orina se recolectó cada 4 horas durante todo el día para registrar la producción diaria de orina de los corderos en detalle. Toda la orina de cada cordero durante el experimento de digestión y metabolismo se agitó por completo. El peso total de la orina se determinó mediante una balanza electrónica (Deante Sensor Technology Co., Ltd., Tianjin, China), y el 10% del total de la orina se submuestreó y se almacenó en frascos de muestras de orina. Inmediatamente, el pH de la orina se determinó con un medidor de pH portátil (Ruibin Technology Co., Ltd., Guangzhou, China). A continuación, se mezclaron muestras de orina por 100 mL con 10 mL de ácido sulfúrico al 10% para la acidificación (27) y se conservaron a −20°C para el análisis del contenido de BA y el metabolismo del nitrógeno.
Además, se recogieron muestras fecales en placas de tamiz de nylon colocadas bajo el suelo del establo individual de corderos. Las heces se recogieron cada 4 h durante todo el día. Después de que las muestras fecales de cordero se mezclaron completamente durante 6 días consecutivos, se pesó aleatoriamente el 10% de la cantidad total. Mientras tanto, el alimento fresco y los orts se muestrearon diariamente. Las muestras fecales se mezclaron por cordero y se submuestrearon. Todos los piensos, orts y muestras fecales (los 100 g de heces se mezclaron con 10 mL de ácido sulfúrico al 10%) se secaron a 65 °C en un horno de aire forzado (Hengmai drying equipment Co., Ltd., Changzhou, China) durante 48 h a un peso constante. A continuación, las muestras secadas al aire se molieron para pasar a través de un tamiz de 1 mm (Xulang machinery Equipment Co., Ltd., Guangzhou, China) para medir el metabolismo del nitrógeno y la digestibilidad aparente.
Antes de la alimentación matutina (0 h) y 1, 2, 3 y 4 h después de la alimentación matutina en d 40, se seleccionaron aleatoriamente 6 corderos de cada tratamiento para recolectar muestras de sangre. Durante cada punto de muestreo, se tomó una muestra total de 5 ml de sangre de la vena yugular de cada cordero utilizando tubos de vacío que no contenían anticoagulante. A continuación, las muestras de sangre se centrifugaron a 3.500 × g y 4 °C durante 15 min para recoger plasma. El plasma se almacenó a -20 °C para su posterior análisis.
2.5 Análisis de orina, heces y muestras de sangre
Las muestras de alimento y heces se analizaron para MS (método 934.01), materia orgánica (MO, método 942.05), PC (método 990.93), extracto de éter (EE, método 920.39), calcio (Ca) y fósforo (P) referencia a los procedimientos AOAC (28). Además, se determinaron los contenidos de fibra detergente neutro (FDN) y fibra detergente ácida (FDA) utilizando un analizador de fibra ANKOM (A2000i, Ankom Technology Corp., Macedonia, Nueva York, EE. UU.). Para calcular la digestibilidad aparente se utilizaron los contenidos de composición química en el alimento y las heces, así como el IMD y el peso fecal (29).
Las muestras de plasma recogidas el día 40 antes de la mañana se utilizaron para medir el contenido de parámetros bioquímicos, incluyendo glucosa (GLU), bilirrubina total (T-Bil), bilirrubina directa (D-Bil), alanina transaminasa (ALT), aspartato transaminasa (AST), fosfatasa alcalina (ALP), glutamil transferasa (GT), proteína total (TP), triglicéridos (TG), colesterol total (CT) y albúmina (ALB), con un analizador bioquímico automático (ZY KHB-1280, Huaren Biotechnology Co., Ltd., Nanjing, China). Las muestras de plasma recogidas en puntos dinámicos se utilizaron para la determinación de BA, HA y nitrógeno ureico. Las concentraciones de BA y HA en plasma y orina se analizaron mediante cromatografía líquida de alta resolución (HPLC) siguiendo los procedimientos de Kubota et al. (30). Brevemente, se transfirieron 100 μL de plasma (200 μL de orina después de una dilución de 10 veces) a otro tubo de centrífuga, y se añadieron 200 μL (400 μL de orina) de proteína precipitada de acetonitrilo que contenía ácido o-clorobenzoico como objetivo interno. Las muestras se agitaron y mezclaron durante 20 s, y se centrifugaron a 9.500 × g durante 1 min. Posteriormente, se recogieron 10 μL de mezcla y se midieron mediante cromatógrafo líquido. Condiciones de determinación: Se utilizó la columna de intercambio catiónico débilmente ácida IC YS-50 de 4,6 × 125 mm; caudal 1,0 mL/min; temperatura de la columna 30°C; longitud de onda de detección 235 nm; una elución lineal; El tamaño de la muestra fue de 10 μL. Además, se determinó nitrógeno ureico utilizando un kit comercial (NO. RATA-A 7170 Huaying, Pekín, China) referencia a las instrucciones.
2.6 Análisis estadístico
Todos los datos se analizaron con el procedimiento ANOVA de una vía del software estadístico SPSS (versión 22.0 para Windows; SPSS, Chicago, EE.UU.), con cada cordero como unidad experimental. Se completaron contrastes polinómicos ortogonales para detectar los efectos lineales y cuadráticos de los niveles de ácido benzoico. Se realizó la prueba de Duncan para determinar las diferencias entre cuatro tratamientos. Los datos se presentaron como media y error estándar de media. El nivel de significancia se indicó en p ≤ 0,05 y se declaró una tendencia en 0,05 < p ≤ 0,10. Además, los cambios dinámicos de BA, HA y nitrógeno ureico se utilizaron para dibujar gráficos de líneas discontinuas utilizando el software GraphPad Prism (versión 8.0 para Windows; GraphPad Prism, San Diego, Estados Unidos).
3 Resultados
3.1 Comportamiento del crecimiento de los corderos destetados
Como se muestra en la Tabla 2, el peso corporal inicial y el peso corporal en el día 30 no mostraron diferencia significativa (p > 0,05) entre todos los grupos. Sin embargo, el peso corporal final mostró tendencia a la variación cuadrática (p = 0,006), y el grupo 1 BA tuvo el valor más alto. El IMD y la GMD fueron similares (p > 0,05) en todos los grupos de 1 a 30 días, y la F:G fue significativamente menor en el grupo de 1BA en comparación con el grupo de 0 BA (p < 0,05). De 31 a 60 días, el IMD y el GMD de 1 grupo BA fueron mayores (p < 0,05) que los de los grupos 0 BA y 1,5 BA, mientras que F:G mostró una tendencia opuesta. No se observaron diferencias significativas (p > 0,05) de IMD en todos los grupos durante todo el período experimental. En comparación con los grupos de 0 BA y 1,5 BA, la GMD en el grupo de 1 BA se elevó significativamente (p < 0,05) de d 1 a 60. La F:G del grupo 1 BA tuvo valor mínimo y menor (p < 0,05) que los grupos 0 y 1,5 BA.
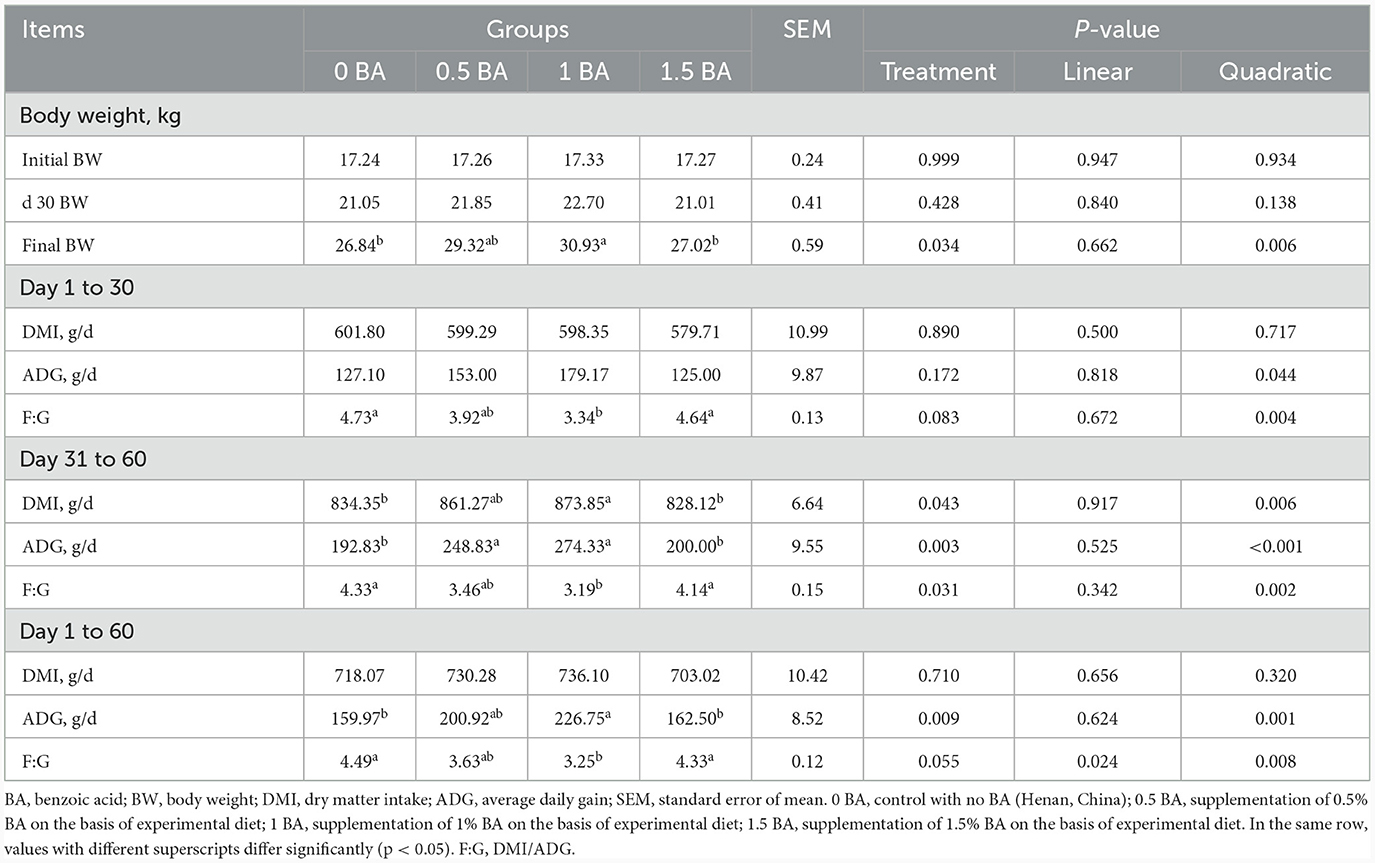 Tabla 2. Efectos de la suplementación con ácido benzoico sobre la ingesta de materia seca y la ganancia media diaria de corderos destetados.
Tabla 2. Efectos de la suplementación con ácido benzoico sobre la ingesta de materia seca y la ganancia media diaria de corderos destetados.
3.2 Digestibilidad de los nutrientes de los corderos destetados
Obviamente, la digestibilidad aparente de PC, EE, Ca y P fue similar (p > 0,05) entre cuatro grupos. La digestibilidad de la MS, MO y FDA del grupo 1 BA fue mayor (p < 0,05) que la del grupo 0 BA (Tabla 3). En comparación con otros grupos, la digestibilidad de la FDN en el grupo 1 BA aumentó significativamente (p < 0,05).
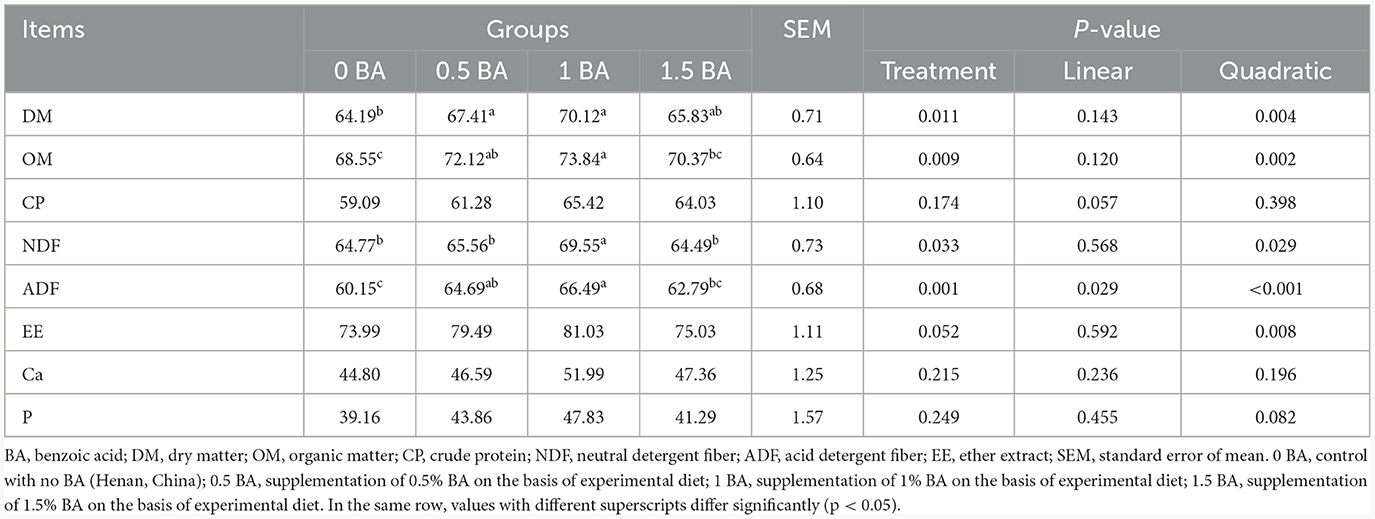 Tabla 3. Efectos de la suplementación con ácido benzoico sobre la digestibilidad de los nutrientes de los corderos destetados.
Tabla 3. Efectos de la suplementación con ácido benzoico sobre la digestibilidad de los nutrientes de los corderos destetados.
3.3 pH urinario y excreción de nitrógeno de los corderos destetados
No se encontraron diferencias evidentes (p > 0,05) del pH urinario, el excremento, la excreción total de nitrógeno y la excreción de nitrógeno de otros ingredientes entre los cuatro grupos (Tabla 4). El HA y el nitrógeno hipurato en los grupos de 1 y 1,5 BA fueron mayores (p < 0,05) que en los grupos de 0 y 0,5 BA. Asimismo, en comparación con el grupo 0 BA, la excreción de AH y la excreción de nitrógeno hipurato en los tratamientos con BA aumentaron significativamente (p < 0,05). Se observó una tendencia opuesta del nitrógeno ureico entre el grupo 0 BA y los tratamientos con BA. Sin embargo, la excreción de nitrógeno ureico fue similar (p > 0,05) entre todos los grupos. Además, el nitrógeno total y el nitrógeno de otros ingredientes en orina del grupo 1,5 BA fueron mayores (p < 0,05) que los grupos 0 y 0,5 BA.
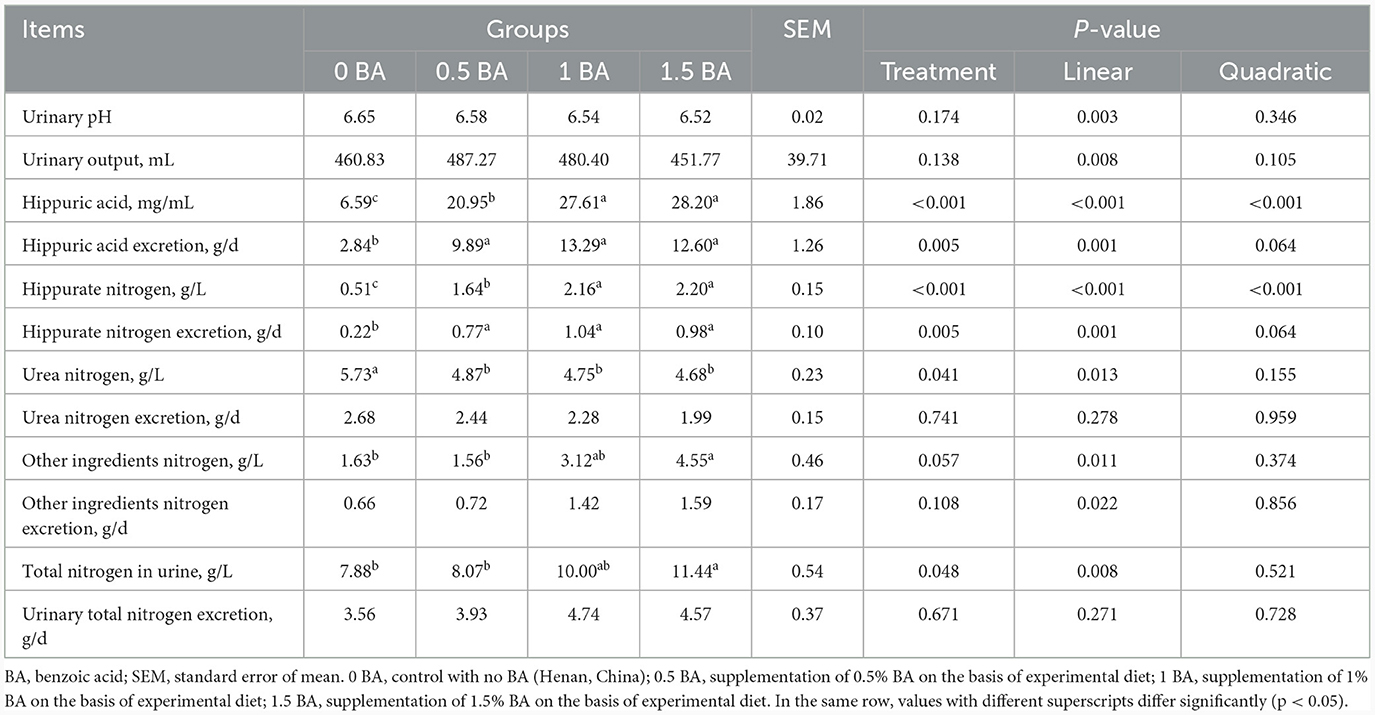 Tabla 4. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre el pH urinario y la excreción de nitrógeno en corderos destetados.
Tabla 4. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre el pH urinario y la excreción de nitrógeno en corderos destetados.
3.4 Análisis de correlación de nitrógeno hipurato y nitrógeno ureico en corderos destetados
Como se puede observar en la Figura 1, el contenido de nitrógeno HA en la orina de los corderos se correlacionó significativamente negativamente con el contenido de nitrógeno ureico (p = 0,0052), y la cantidad de nitrógeno HA excretado se correlacionó significativamente positivamente con la cantidad de nitrógeno ureico excretado (p = 0,0242).
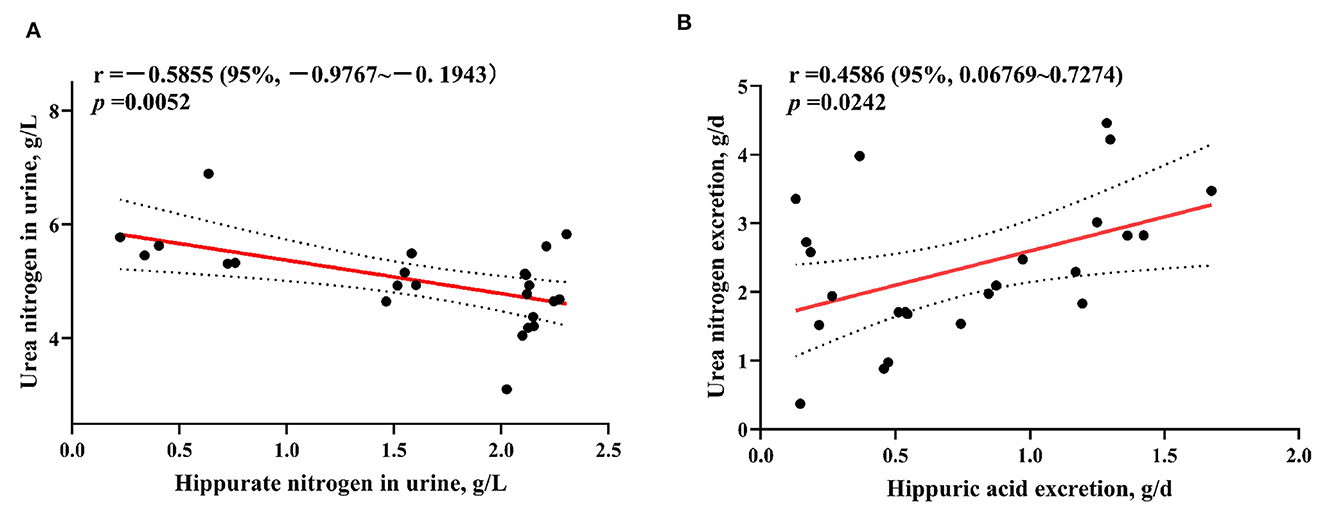 Figura 1. Análisis del contenido de nitrógeno hipurato y nitrógeno ureico en la orina de corderos destetados (A) y la correlación entre el nitrógeno hipurato y la excreción de nitrógeno ureico (B).
Figura 1. Análisis del contenido de nitrógeno hipurato y nitrógeno ureico en la orina de corderos destetados (A) y la correlación entre el nitrógeno hipurato y la excreción de nitrógeno ureico (B).
3.5 Metabolismo del nitrógeno de los corderos destetados
No hubo diferencia significativa (p > 0,05) de nitrógeno fecal, nitrógeno urinario y retención de nitrógeno entre todos los grupos (Tabla 5). Sin embargo, la ingesta de nitrógeno en los grupos 0,5 y 1 BA aumentó significativamente (p < 0,05) en comparación con los grupos 0 y 1,5 BA.
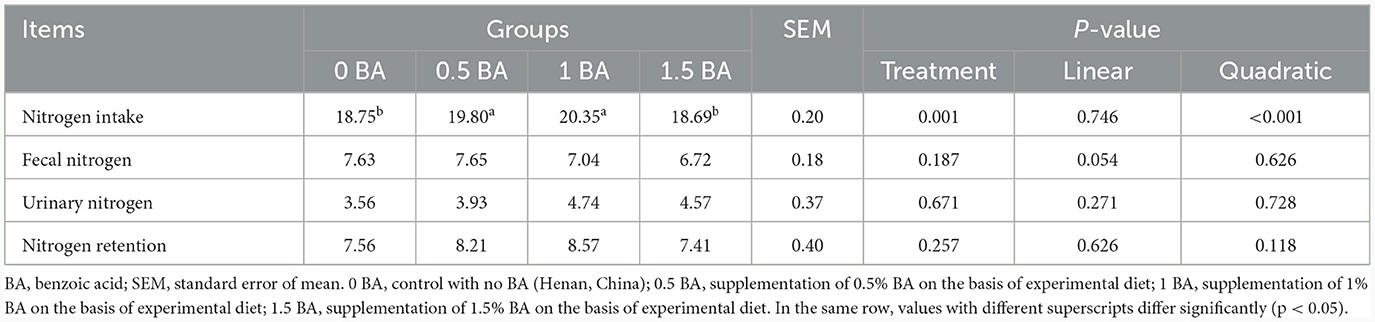 Tabla 5. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre el metabolismo del nitrógeno en corderos destetados (g/d).
Tabla 5. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre el metabolismo del nitrógeno en corderos destetados (g/d).
3.6 Parámetros bioquímicos sanguíneos de los corderos destetados
Las concentraciones de GLU, T-Bil, D-Bil, ALT, AST, ALP, GT, TP, TG y CT en plasma fueron similares (p > 0,05) entre cuatro grupos (Tabla 6). En comparación con los grupos de 0 BA y 1,5 BA, el contenido plasmático de ALB de 1 grupo de BA aumentó significativamente (p < 0,05).
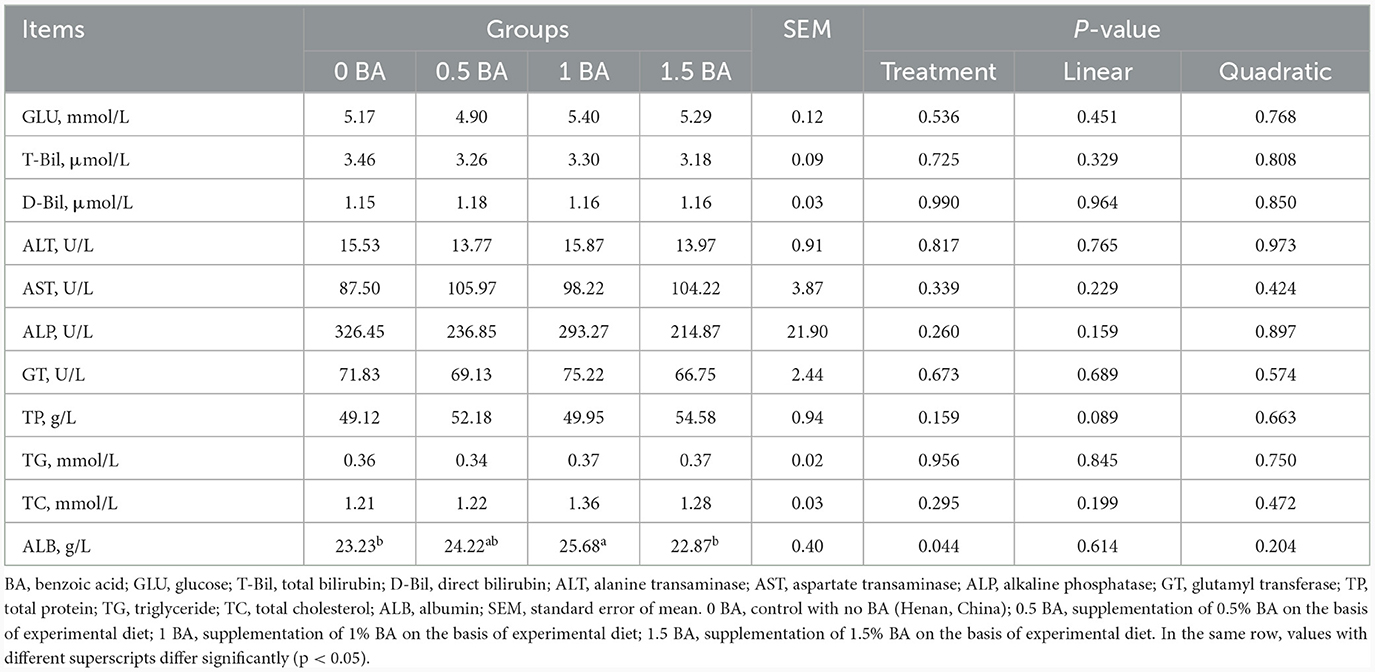 Tabla 6. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre los parámetros bioquímicos sanguíneos en corderos destetados.
Tabla 6. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre los parámetros bioquímicos sanguíneos en corderos destetados.
3.7 Cambios dinámicos del ácido benzoico, el ácido hipúrico y el nitrógeno ureico en el plasma de corderos destetados
Como se muestra en la Figura 2A, 1 h después de la alimentación matutina, la concentración plasmática de BA del grupo 1 BA alcanzó el valor máximo y fue mayor (p < 0,05) que otros grupos, y luego comenzó a disminuir. A 1 post-alimentación, la concentración de BA en el grupo de 1,5 BA aumentó significativamente (p < 0,05) en comparación con los grupos de 0 y 0,5 BA. Durante todo el proceso de muestreo, el contenido de BA en los grupos de 0 y 0,5 BA estuvo siempre en un nivel más bajo. Del mismo modo, la concentración de AH en plasma de los grupos 1 y 1,5 BA fue mayor (p < 0,05) que la del grupo 0 BA de 1 a 4 h después de la alimentación matutina (Figura 2B). Después de que los corderos suplementados con BA, la concentración plasmática de AH aumentó gradualmente y alcanzó el valor más alto 1 h después de la alimentación y luego comenzó a mostrar un cambio de fluctuación. La concentración de AH de los grupos 0, 0,5 y 1,5 BA a las 4 h fue cercana a las 0 h correspondientes. Por el contrario, a las 3 h después de la alimentación, la concentración de nitrógeno ureico en el plasma del grupo 0 BA fue mayor (p < 0,05) que la del grupo 1,5 BA (Figura 2C). No se observaron diferencias evidentes (p > 0,05) en la concentración de nitrógeno ureico en otros momentos entre todos los grupos.
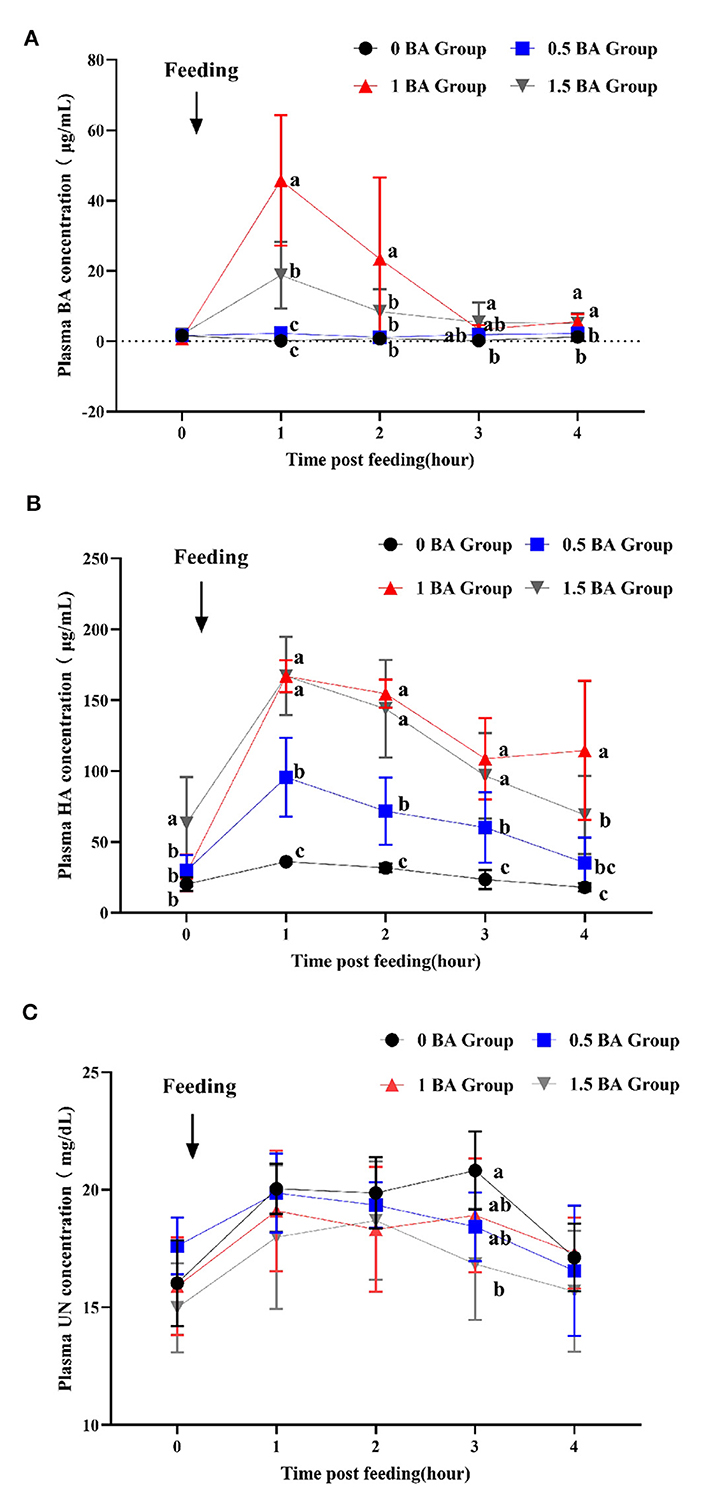 Figura 2. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre los cambios dinámicos del ácido benzoico (A), el ácido hipúrico (B) y el nitrógeno ureico (C) en plasma en corderos destetados. BA: ácido benzoico; HA: ácido hipúrico; ONU, nitrógeno ureico. 0 BA, control sin BA (Henan, China); 0,5 BA, suplementación de 0,5% BA sobre la base de la dieta experimental; 1 BA, suplementación con 1% de BA sobre la base de una dieta experimental; 1,5 BA, suplementación de 1,5% BA sobre la base de una dieta experimental. Las diferentes letras (a, b y c) representan diferencias estadísticamente significativas (P < 0,05).
Figura 2. Efectos de la suplementación con diferentes niveles de ácido benzoico sobre los cambios dinámicos del ácido benzoico (A), el ácido hipúrico (B) y el nitrógeno ureico (C) en plasma en corderos destetados. BA: ácido benzoico; HA: ácido hipúrico; ONU, nitrógeno ureico. 0 BA, control sin BA (Henan, China); 0,5 BA, suplementación de 0,5% BA sobre la base de la dieta experimental; 1 BA, suplementación con 1% de BA sobre la base de una dieta experimental; 1,5 BA, suplementación de 1,5% BA sobre la base de una dieta experimental. Las diferentes letras (a, b y c) representan diferencias estadísticamente significativas (P < 0,05).
4 Discusión
La nutrición del cordero es un tema cada vez más importante en la producción ovina actual. El manejo de la alimentación de los corderos tiene una influencia a largo plazo en el rendimiento futuro de la producción de la calidad de la carne de las ovejas de engorde (31). Debido a la inmadurez del tracto gastrointestinal, los corderos se ven fácilmente afectados por microorganismos dañinos. Después del destete, los corderos experimentan cambios en el tipo de alimento y el patrón de cría, lo que resulta en una disminución de la digestibilidad de los nutrientes y la tasa de crecimiento (32). Aliviar el estrés del destete de los corderos es de gran importancia para mejorar el crecimiento saludable de los corderos. Como ácido carboxílico aromático, el BA tiene múltiples beneficios para la salud, incluidas actividades antibacterianas y antiinflamatorias (18). Anteriormente, una investigación ha encontrado que la suplementación dietética de BA podría aumentar la GMD de los cerdos destetados, pero no tenía una diferencia obvia de eficiencia alimenticia (33). En el estudio actual, la suplementación con 1% de BA aumentó significativamente la GMD y la eficiencia alimenticia en comparación con los grupos de 0 BA y 1,5 BA. La razón diferente puede ser que los animales utilizados en el experimento eran diferentes. BA puede producir productos de esterificación con bacterias, que luego afectan el metabolismo de las bacterias patógenas. Además, el BA puede interferir con la actividad del ADN de las bacterias patógenas, disminuyendo así su crecimiento (34). Estos efectos son propicios para reducir la tasa de incidencia de diarrea y aliviar el impacto negativo en la tasa de crecimiento de los corderos causado por el estrés del destete. Nuestros resultados mostraron que la adición apropiada (1%) de BA mejoró el rendimiento del crecimiento de los corderos destetados. La mejora del rendimiento del crecimiento de los animales suele asociarse a una mayor digestibilidad aparente y a un mayor metabolismo del nitrógeno. Por lo tanto, realizamos el siguiente experimento para estudiar la influencia del BA en la digestibilidad aparente y el metabolismo del nitrógeno de los corderos destetados.
El tracto gastrointestinal de los corderos destetados tiene una secreción insuficiente de enzimas digestivas, por lo que la digestibilidad de los nutrientes de los corderos es baja (35). En nuestro estudio, la suplementación dietética de BA con 0,5 y 1% aumentó significativamente la ingesta de nutrientes de los corderos. En general, los piensos típicos alimentados con rumiantes jóvenes tienen valores relativamente altos de capacidad de fijación de ácidos, lo que afecta a la ingesta de nutrientes de los animales (36). Un estudio anterior encontró que la suplementación dietética con BA podría disminuir la capacidad de unión de ácidos de los alimentos (37), lo que podría explicar los efectos positivos de BA en la ingesta de nutrientes. La digestibilidad de la MS y la MO son parámetros clave para reflejar la capacidad de utilización de los piensos por parte de los animales (38). En el presente estudio, la digestibilidad de MS y MO en el grupo 1 BA mostró los valores más altos y fueron significativamente elevadas en comparación con el grupo 0 BA, lo que indica que los corderos a los que se les administró 1% BA pudieron obtener más nutrientes y acelerar el crecimiento, lo que se correspondió con los resultados de ADG. El ácido orgánico es una alternativa eficaz para mejorar la digestibilidad de los nutrientes en la producción de la industria animal. El ácido orgánico tiene efectos multifuncionales, incluyendo la reducción del pH gastrointestinal, el aumento del tiempo de retención gastrointestinal, la estimulación de las secreciones pancreáticas y la promoción de la morfología gastrointestinal, mejorando así la digestibilidad de los nutrientes (39, 40). Nuestros resultados podrían estar relacionados con la capacidad de BA para regular el pH gastrointestinal y los niveles de enzimas digestivas y mejorar la morfología intestinal. En el futuro, los efectos de la BA en el desarrollo gastrointestinal de los corderos necesitan una investigación en profundidad. Además, también encontramos que la digestibilidad de la FDN y la FDA de los grupos 1 BA fueron mayores que las de los grupos 0 y 1,5 BA. En los rumiantes, la comunidad microbiana en el rumen es responsable de la utilización de la fibra bruta. Desafortunadamente, no se ha reportado el efecto del BA sobre la comunidad microbiana ruminal. Un estudio reciente ha verificado que la suplementación dietética de BA podría estabilizar la fluctuación de la microbiota causada por el estrés del destete y acelerar la rápida colonización de las bacterias dominantes en el intestino de los lechones (41). A pesar de que la comunidad microbiana entre los animales monogástricos y los rumiantes existió de manera diferente, estos resultados indicaron que el BA podría afectar a la comunidad microbiana. Especulamos que los efectos positivos de la BA sobre la digestibilidad de la FDN y la FDA podrían estar asociados con la regulación de la microbiota en el tracto gastrointestinal. Por último, la actividad de las enzimas digestivas juega un papel importante en la digestión de los nutrientes (42). Por lo tanto, se necesitan más experimentos para explorar los efectos del BA en la comunidad microbiana y la actividad de las enzimas digestivas de los corderos destetados.
Anteriormente, los estudios han informado que con la elevación de la suplementación con BA en la dieta, el pH urinario se redujo linealmente (43) y, además, la concentración de AH en orina aumentó linealmente (44, 45), lo que también mostró hallazgos similares en nuestra investigación. La razón de la reducción del pH urinario puede deberse a la elevación del contenido urinario de AH. Después de la absorción, el BA se conjuga con glicina a través de la glicina-N-acilasa, y luego se transforma en HA en el hígado (46). Además, la concentración de nitrógeno ureico en los tratamientos de suplementación con BA al 1% disminuyó significativamente (3 h), lo que sugiere que la conversión de nitrógeno de los corderos 0 BA fue baja. Como se mencionó anteriormente, la síntesis de AH y el ciclo de urea ocurren en las mitocondrias hepáticas de los rumiantes, y juntos consumen NH +4 4+ producido a partir del metabolismo de los aminoácidos de la dieta (47). Por lo tanto, cuando el BA entra en el cuerpo del animal, ¿el aumento de la síntesis de AH tiene el efecto de reducir la producción de urea? En el presente estudio, los resultados mostraron que con el aumento de la ingesta de BA, el contenido urinario de AH y la excreción de AH aumentaron linealmente, mientras que el contenido urinario de nitrógeno ureico disminuyó linealmente. Al mismo tiempo, se encontró que existía una correlación significativa entre la excreción urinaria de nitrógeno ureico y la excreción urinaria de nitrógeno AH. Este resultado confirmó nuestra hipótesis previa de que la elevación de la excreción de AH mediante la adición de BA a la dieta redujo la excreción de nitrógeno ureico. Sin embargo, hubo resultados insatisfactorios en cuanto a la producción total de nitrógeno urinario. Además del nitrógeno ácido hipúrico y el nitrógeno ureico, también hay nitrógeno producido por derivados de purinas en el nitrógeno total urinario. En estudios previos de toros cruzados, se encontró que el nitrógeno producido por derivados de purinas (como el nitrógeno alantoína) aumentaba linealmente a medida que aumentaba la ingesta de alimento (48). En nuestro experimento, la excreción de nitrógeno de otros componentes de los corderos destetados aumentó significativamente, y la excreción total de nitrógeno urinaria insatisfactoria podría deberse a la mayor ingesta de alimento de los corderos con 0,5 y 1% de BA. Además, de acuerdo con nuestros datos no publicados, después de alimentar con BA a corderos lactantes, los metabolitos del contenido de yeyuno se enriquecieron principalmente en las vías metabólicas de pirimidina y purina, lo que indica que la ingesta de BA también afectó el metabolismo de los derivados de purina en corderos. Un estudio previo en conejos encontró que la ingesta de benzoato de sodio condujo a un aumento en la excreción urinaria de AH y una disminución en la excreción urinaria de nitrógeno ureico (49), lo que fue consistente con los resultados de nuestro experimento. Nousiainen et al. (50) informaron que el aumento del contenido de nitrógeno ureico en orina y sangre indicaba una reducción de la eficiencia del nitrógeno de las vacas. Los grupos de 0,5 y 1 BA mostraron una mayor ingesta de nitrógeno en comparación con los grupos de 0 y 1,5 BA. De acuerdo con nuestros resultados, la suplementación adecuada de BA en la dieta podría mejorar la utilización del nitrógeno de los corderos destetados, lo que fue útil para promover el crecimiento.
Recientemente, un estudio informó que la mejora de la GMD se atribuyó a la elevada retención de nitrógeno en cerdos alimentados con dietas con BA (51). En el presente estudio, no encontramos diferencias obvias en la retención de nitrógeno entre todos los grupos. Sin embargo, el grupo 1 BA tuvo el valor más alto de retención de nitrógeno. Un estudio anterior encontró que la suplementación dietética de BA podría mejorar la morfología intestinal y regular al alza la expresión del gen del péptido similar al glucagón 2 en la mucosa intestinal (37), lo que podría mejorar la retención de nitrógeno en animales alimentados con raciones de BA. También encontramos que la adición de 1.5% de BA a la ración basal disminuyó la ingesta de nitrógeno y redujo la utilización de nitrógeno en los corderos, lo que afectó negativamente el rendimiento del crecimiento. La posible razón podría estar relacionada con la elevación de la síntesis de glicina necesaria para transformar BA en HA en el hígado, lo que podría afectar el metabolismo de los aminoácidos y reducir la concentración de aminoácidos para la síntesis de proteínas (46). La utilización excesiva de proteínas y aminoácidos para la transformación del AH reduce la retención de nitrógeno. Así, según nuestros resultados, el nivel adecuado de suplementación de BA en la dieta de corderos fue del 1%.
Como índices importantes relacionados con el estado de salud de los animales, los cambios de la bioquímica sérica se pueden utilizar para estimar el metabolismo fisiológico del cuerpo y las funciones de los órganos (52). Los contenidos de T-Bil, D-Bil, ALT, AST, ALP y GT en plasma se pueden utilizar para evaluar la función hepática, y los cambios en las concentraciones de GLU, TG y TC se asocian con el metabolismo de las grasas. Además, las concentraciones plasmáticas de TP y ALB son parámetros clave del metabolismo de las proteínas (52). En el presente experimento, los contenidos plasmáticos de GLU, T-Bil, D-Bil, ALT, AST, ALP, GT, TP, TG y TC fueron similares entre cuatro tratamientos, lo que sugiere que la suplementación con BA no tuvo efectos adversos sobre el metabolismo lipídico y la función hepática de los corderos destetados. Un estudio anterior encontró que la suplementación con BA aumentó significativamente el contenido de ALB en la sangre y redujo el contenido de nitrógeno ureico (53). De acuerdo con estudios previos, nuestro estudio encontró que la suplementación con BA al 1% aumentó el contenido plasmático de ALB y redujo la concentración de nitrógeno ureico, lo que indica que BA podría mejorar el anabolismo de la proteína hasta cierto punto, lo que favoreció el crecimiento de los corderos.
Estudios previos han verificado que el AH se forma por una reacción de biosíntesis de BA y puede ser excretado de la orina del cuerpo dentro de las 4 h (54, 55). Estudiamos por primera vez los cambios dinámicos de BA, HA y nitrógeno ureico en plasma de corderos destetados alimentados con BA. Los resultados mostraron que después de la alimentación con BA 1 h, las concentraciones plasmáticas de BA y HA alcanzaron el valor más alto y luego disminuyeron gradualmente, y los grupos de 1 y 1,5 BA fueron más altos que otros grupos. El BA se absorbe a través del intestino del animal y entra en el torrente sanguíneo, donde luego se transporta al hígado para su metabolismo (13). Nuestros hallazgos indicaron que después de que el BA fue transportado al hígado, el proceso de producción de BA y glicina bajo la catálisis de la enzima fue muy rápido. En los animales, el BA y la glicina son catalizados por enzimas para formar HA, que luego se excreta en la orina. Además, el BA también puede ser metabolizado por los microbios intestinales (56, 57). La comunidad microbiana de los corderos aún se encontraba en etapa de desarrollo, y los niveles más altos de BA y HA pueden tener efectos positivos en la comunidad microbiana. Sin embargo, aún es necesario explorar el posible mecanismo de acción. Como los principales productos del metabolismo de las proteínas del cuerpo de los animales, el nitrógeno ureico son índices importantes para reflejar la utilización de proteínas. La disminución de estos contenidos indica que se mejora la utilización de proteínas, lo que contribuye a la deposición de nitrógeno y, además, el alto contenido de nitrógeno ureico también puede tener efectos adversos en la salud animal (58). En este estudio, la suplementación dietética de BA disminuyó efectivamente el contenido de nitrógeno ureico plasmático después de la alimentación matutina de 2 a 3 h, lo que estuvo en línea con el contenido de nitrógeno ureico urinario mencionado anteriormente. Este resultado favoreció la mejora del metabolismo del nitrógeno de los corderos. La investigación futura debería prestar más atención a los posibles mecanismos de BA en el metabolismo del nitrógeno de los corderos.
En conjunto, la suplementación con BA tuvo la capacidad de mejorar el metabolismo del nitrógeno de los corderos disminuidos, lo que favoreció la promoción del crecimiento de los corderos. Sin embargo, es necesario reconocer algunas limitaciones en el presente estudio. En primer lugar, la vía de absorción de BA en el tracto gastrointestinal requiere una mayor elucidación. En el experimento posterior, exploraremos el posible mecanismo de acción utilizando estudios in vitro e in vivo. Además, se necesitan estudios futuros para comprender las funciones de la comunidad microbiana en el tracto gastrointestinal mediante la metagenómica o la culturomica, así como la función del huésped mediante transcriptómica, para proporcionar más información sobre el papel de los microorganismos en el órgano digestivo de los corderos destetados y su respuesta a la suplementación con BA en la dieta.
5 Conclusiones
La suplementación con BA (1%) mejoró la GMD, la eficiencia alimenticia y la digestibilidad de MS, MO, FDN y FDA de los corderos destetados. Además, la suplementación dietética con BA al 1% aumentó significativamente el contenido y la excreción urinaria de AH, así como la ingesta de nitrógeno. Después de la suplementación con BA, se redujeron los contenidos de nitrógeno ureico plasmático y urinario. Por lo tanto, según nuestros hallazgos, la suplementación adecuada de BA (1%) en la dieta mejora el rendimiento del crecimiento y el metabolismo del nitrógeno de los corderos destetados.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio se incluyen en el artículo/material complementario, las consultas posteriores pueden dirigirse al autor correspondiente.
Declaración ética
Todos los procedimientos relacionados con el cuidado y manejo de los animales utilizados en este experimento fueron autorizados (número de protocolo: 2020022) por el Comité Institucional de Cuidado y Uso de Animales de la Universidad Agrícola de Xinjiang (Urumqi, Xinjiang, China). El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones de los autores
WZ: Conceptualización, Investigación, Visualización, Escritura – borrador original, Escritura – revisión y edición. SS: Investigación, Visualización, Escritura – revisión y edición. YaqZ: Investigación, Escritura – revisión y edición. YanZ: Escritura – borrador original. JW: Investigación, Escritura, revisión y edición. ZL: Investigación, Escritura – Revisión y Edición. KY: Conceptualización, Curación de datos, Adquisición de fondos, Metodología, Administración de proyectos, Supervisión, Redacción, revisión y edición.
Financiación
El/los autor/es declara(n) haber recibido apoyo financiero para la investigación, autoría y/o publicación de este artículo. Esta investigación fue financiada por el Programa Clave de Investigación y Desarrollo del 14º Plan Quinquenal Nacional (2021YFD1600702-2).
Reconocimientos
Agradecemos el apoyo de Xinjiang Shangpin Meiyang Technology Co., Ltd., China.
Conflicto de intereses
JW y ZL fueron empleados de Xinjiang Shangpin Meiyang Technology Co., Ltd.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Worden D, Hailu G, Jones K, Lee YN. Los efectos de la agrupación en las valoraciones de los productores ganaderos de los rasgos respetuosos con el medio ambiente disponibles a través de la selección genómica. Can J Agric Econ-Rev Can Agroecon. (2022) 70:263–86. doi: 10.1111/cjag.12322
2. Cai XH, Qin YF, Yan BJ, Shi WJ. Identificación de explotaciones ganaderas con riesgo potencial de contaminación ambiental mediante el uso de un modelo de retorno de estiércol de ganado a las tierras cultivadas. Environ Sci Pollut Res. (2023) 30:103062–72. doi: 10.1007/s11356-023-29681-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
3. Martín-Hernández E, Montero-Rueda C, Ruiz-Mercado GJ, Vaneeckhaute C, Martín M. Evaluación tecnoeconómica multiescala de sistemas de recuperación de nitrógeno para explotaciones ganaderas. Sustain Prod Consump. (2023) 41:49–63. doi: 10.1016/j.spc.2023.07.028
Resumen de PubMed | Texto completo de Crossref | Google Académico
4. van der Weerden TJ, Noble AN, Beltran I, Hutchings NJ, Thorman RE, de Klein CAM, et al. Influencia de los factores clave en los factores de emisión de amoníaco y óxido nitroso para las excretas depositadas por el ganado y el estiércol aplicado a la tierra. Sci Total Environ. (2023) 889:164066. doi: 10.1016/j.scitotenv.2023.164066
Resumen de PubMed | Texto completo de Crossref | Google Académico
5. Soares JR, Souza BR, Mazzetto AM, Galdos MV, Chadwick DR, Campbell EE, et al. Mitigación de las emisiones de óxido nitroso en sistemas de pastoreo a través de inhibidores de la nitrificación: un meta-análisis. Nutr Cycl Agroecosyst. (2023) 19:10256–8. doi: 10.1007/s10705-022-10256-8
6. Cheng LX, Zhang XM, Reis S, Ren CC, Xu JM, Gu BJ, et al. El cambio del 12% de la producción ganadera de monogástricos a rumiantes puede reducir las emisiones e impulsar la producción de cultivos para 525 millones de personas. Nat Comida. (2023) 4:190–210. doi: 10.1038/s43016-022-00661-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
7. Jin D, Zhao SG, Zheng N, Beckers Y, Wang JQ. Metabolismo de la urea y regulación por la ureasa bacteriana ruminal en rumiantes: una revisión. Ann Anim Sci. (2018) 18:303–18. doi: 10.1515/aoas-2017-0028
8. Ewaoluwagbemiga EO, Bee G, Kasper C. Análisis genético de la eficiencia proteica y su asociación con el rendimiento y los rasgos de calidad de la carne bajo una dieta restringida en proteínas. Genet Sel Evol. (2023) 55:16. doi: 10.1186/s12711-023-00812-3
Resumen de PubMed | Texto completo de Crossref | Google Académico
9. Seleem MS, Wu ZH, Xing CQ, Zhang Y, Hanigan MD, Bu DP. Impactos de la suplementación con metionina y lisina encapsulada en el rumen y la baja proteína dietética en la eficiencia del nitrógeno y el rendimiento de la lactancia de las vacas lecheras. J. Dairy Sci. (2023). doi: 10.3168/jds.2023-23404
Resumen de PubMed | Texto completo de Crossref | Google Académico
10. Hristov AN, Bannink A, Crompton LA, Huhtanen P, Kreuzer M, McGee M, et al. Revisión invitada: nitrógeno en la nutrición de rumiantes: una revisión de las técnicas de medición. J. Dairy Sci. (2019) 102:5811–52. doi: 10.3168/jds.2018-15829
Resumen de PubMed | Texto completo de Crossref | Google Académico
11. McCoard S.A., Pacheco D. La importancia del N-carbamoilglutamato en la producción de rumiantes. J Anim Sci Biotechnol. (2023) 14:17. DOI: 10.1186/s40104-023-00854-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
12. Ticinesi A, Guerra A, Nouvenne A, Meschi T, Maggi S. Desenredando la complejidad de la nutrición, la fragilidad y las vías microbianas intestinales durante el envejecimiento: un enfoque en el ácido hipúrico. Nutrientes. (2023) 15:05118. doi: 10.3390/nu15051138
Resumen de PubMed | Texto completo de Crossref | Google Académico
13. Ulaszewska M, García-Aloy M, Vázquez-Manjarrez N, Soria-Florido MT, Llorach R, Mattivi F, et al. Biomarcadores de ingesta de alimentos para bayas y uvas. Genes Nutr. (2020) 15:35. doi: 10.1186/s12263-020-00675-z
Resumen de PubMed | Texto completo de Crossref | Google Académico
14. De Simone G, Balducci C, Forloni G, Pastorelli R, Brunelli L. Ácido hipúrico: ¿podría convertirse en un barómetro de la fragilidad y los síndromes geriátricos? Ageing Res Rev. (2021) 72:101466. doi: 10.1016/j.arr.2021.101466
Resumen de PubMed | Texto completo de Crossref | Google Académico
15. Roble BW. Algunos cambios químicos en los constituyentes nitrogenados de la orina cuando se orinan en los pastos. J Agr Sci. (1952) 42:162–71. doi: 10.1017/S0021859600058767
16. Bristow AW, Whitehead DC, Cockburn JE. Constituyentes nitrogenados en la orina de bovinos, ovinos y caprinos. J Sci Food Agr. (1992) 59:387–94. doi: 10.1002/jsfa.2740590316
17. Rychen G, Aquilina G, Azimonti G, Bampidis V, de Lourdes Bastos M, Bories G, et al. Seguridad y eficacia de VevoVitall (ácido benzoico) como aditivo alimentario para especies porcinas menores. EFSA J. (2017) 15:e05026. doi: 10.2903/j.efsa.2017.5026®
Resumen de PubMed | Texto completo de Crossref | Google Académico
18. Süloglu AK, Koçkaya EA, Selmanoglu G. Toxicidad del benzoato de bencilo como aditivo alimentario y agente farmacéutico. Toxicol Ind Health. (2022) 38:221–33. doi: 10.1177/07482337221086133
Resumen de PubMed | Texto completo de Crossref | Google Académico
19. Gao Z, Yu B, Zheng P, He J, Mao X, Yu J, et al. Efectos del ácido benzoico sobre la microflora intestinal y los metabolitos de lechones. Chin J Anim Nutr. (2014) 26:1044–54.
20. Chen J, Chen D, Yu B, He J, Mao X, Yu J, et al. Efectos del ácido benzioc sobre el rendimiento del crecimiento, los índices de órganos y el pH del contenido gastrointestinal de lechones destetados. Chin J Anim Nutr. (2015) 27:238–46.
21. Mao XB, Yang Q, Chen DW Yu B, He J. El ácido benzoico utilizado como aditivos para alimentos y piensos puede regular las funciones intestinales. Biomed Res Int. (2019) 2019:6. doi: 10.1155/2019/5721585
Resumen de PubMed | Texto completo de Crossref | Google Académico
22. Diao H, Zheng P, Yu B, He J, Mao XB Yu J, Chen DW. Efectos de la suplementación dietética con ácido benzoico sobre la estructura morfológica intestinal y la microflora en lechones destetados. Livest Sci. (2014) 167:249–56. doi: 10.1016/j.livsci.2014.05.029
23. Hailemariam S, Zhao SG, He Y, Wang JQ. Transporte e hidrólisis de urea en el rumen: una revisión. Anim Nutr. (2021) 7:989–96. doi: 10.1016/j.aninu.2021.07.002
24. Consejo NR. Requerimientos de nutrientes de los pequeños rumiantes. Washington, DC: The National Academies Press (2007).
25. Williams MS, Mandell IB, Bohrer BM, Wood KM. Los efectos de la alimentación con ácido benzoico y/o levadura viva activa (Saccharomyces cerevisiae) sobre el rendimiento del ganado vacuno, el comportamiento alimentario y las características de la canal. Transl Anim Sci. (2021) 5:txab143. doi: 10.1093/tas/txab143
Resumen de PubMed | Texto completo de Crossref | Google Académico
26. O’Meara FM, Gardiner GE, O’Doherty JV, Lawlor PG. Efecto de la inclusión dietética de ácido benzoico (VevoVitall) en la calidad microbiana de los piensos líquidos y en el crecimiento y la calidad de la canal de los cerdos de engorde. Livest Sci. (2020) 237:104043. doi: 10.1016/j.livsci.2020.104043®
27. López MC, Fernández C. Partición energética y oxidación del sustrato por cabras murciano-granadinas durante la mitad de la lactancia alimentadas con cáscaras de soja y mezcla de piensos con gluten de maíz como sustituto del grano de maíz. J Lácteos Sci. (2013) 96:4542–52. doi: 10.3168/jds.2012-6473
Resumen de PubMed | Texto completo de Crossref | Google Académico
28. AOAC. Asociación de Químicos Analíticos Oficiales Métodos Oficiales de Análisis. Arlington, VA (2010).
29. Aguiar FD, Bezerra LR, Cordao MA, Cavalcante ITR, de Oliveira JPF, do Nascimento RR, et al. Efectos del aumento de los niveles de taninos totales sobre la ingesta, digestibilidad y equilibrio de nitrógeno, agua y energía en corderos de pelo. Animales. (2023) 13:2497. doi: 10.3390/ani13152497
Resumen de PubMed | Texto completo de Crossref | Google Académico
30. Kubota K, Horai Y, Kushida K, Ishizaki T. Determinación del ácido benzoico y el ácido hipúrico en plasma y orina humanos mediante cromatografía líquida de alta resolución. J Chromatogra B. (1988) 425:67–75. doi: 10.1016/0378-4347(88)80007-0
Resumen de PubMed | Texto completo de Crossref | Google Académico
31. Guo YX, Yang RC, Duan CH, Wang Y, Hao QH, Ji SK, et al. Efecto de los residuos opuestos a la dioscorea sobre el rendimiento del crecimiento, los parámetros sanguíneos, la fermentación ruminal y la comunidad bacteriana ruminal en corderos destetados. J Integr Agric. (2023) 22:1833–46. doi: 10.1016/j.jia.2022.10.002
32. Mao HL Ji WW, Yun Y, Zhang YF Li ZF, Wang C. Influencia de la suplementación con probióticos en el rendimiento del crecimiento, las variables plasmáticas y la comunidad bacteriana ruminal del cordero con retraso en el crecimiento. Microbiol frontal. (2023) 14:1216534. doi: 10.3389/fmicb.2023.1216534
Resumen de PubMed | Texto completo de Crossref | Google Académico
33. Warner AJ, DeRouchey JM, Tokach MD, Woodworth JC, Goodband RD, Gebhardt JT. Efecto del carbonato de calcio añadido sin y con ácido benzoico sobre el rendimiento del crecimiento de los cerdos destetados, la materia seca fecal y las concentraciones de Ca y P en sangre. Transl Anim Sci. (2023) 7:txad055. doi: 10.1093/tas/txad055
Resumen de PubMed | Texto completo de Crossref | Google Académico
34. Rohatgi A, Gupta P. Derivados del ácido benzoico como potentes agentes antibiofilm contra el biofilm de Klebsiella pneumoniae. J Biosci Bioeng. (2023) 136:190–7. doi: 10.1016/j.jbiosc.2023.06.011
Resumen de PubMed | Texto completo de Crossref | Google Académico
35. Abdelsattar MM, Vargas-Bello-Pérez E, Zhuang YM, Fu YZ, Zhang NF. Impacto de la suplementación dietética con ácido β-hidroxibutírico sobre el rendimiento, la digestibilidad de los nutrientes, el desarrollo de los órganos y los indicadores de estrés sérico en cabritos destetados precozmente. Anim Nutr. (2022) 9:16–22. doi: 10.1016/j.aninu.2021.11.003
Resumen de PubMed | Texto completo de Crossref | Google Académico
36. Wang Y, Lu N, Tian X, Yang J, Chen Y, Bai C. Efectos de los niveles de capacidad de unión de ácidos en la dieta sobre el rendimiento y la digestibilidad aparente de los nutrientes de los corderos. Chin J Anim Nutr. (2009) 21:488–92.
37. Diao H, Gao Z, Yu B, Zheng P, He J, Yu J, et al. Efectos del ácido benzoico (Vevo Vitall) sobre el rendimiento y la fisiología digestiva yeyunal en cerdos jóvenes. J Anim Sci Biotechnol. (2016) 7:32. doi: 10.1186/s40104-016-0091-y
Resumen de PubMed | Texto completo de Crossref | Google Académico
38. Ma J, Fan X, Zhang W, Zhou G, Yin F, Zhao Z, et al. El extracto de semilla de uva como aditivo alimentario mejora el rendimiento del crecimiento, la fermentación ruminal y la inmunidad de los terneros destetados. Animales. (2023) 13:1876. doi: 10.3390/ani13111876
Resumen de PubMed | Texto completo de Crossref | Google Académico
39. Han MM, Chen BB, Dong YY, Miao ZQ, Su Y, Liu C, et al. Evaluación de los ácidos orgánicos líquidos sobre el rendimiento, el pH del quimo, la utilización de nutrientes y la microbiota intestinal en pollos de engorde con alta densidad de población. Animales. (2023) 13:17. doi: 10.3390/ani13020257
Resumen de PubMed | Texto completo de Crossref | Google Académico
40. Li ZQ, Liu SH, Zhao YR, Wang JY, Ma XK. Los ácidos orgánicos compuestos podrían mejorar el rendimiento del crecimiento, la inmunidad y las propiedades antioxidantes, así como la salud intestinal al alterar el perfil de la microbiota de los lechones destetados. J Anim Sci. (2023) 101:12. doi: 10.1093/jas/skad196
Resumen de PubMed | Texto completo de Crossref | Google Académico
41. Wei K, Yang X, Zhao H, Chen H, Bei W. Efectos de la aplicación combinada de ácido benzoico y 1-monolaurina sobre el rendimiento del crecimiento, la digestibilidad de los nutrientes, el microbioma intestinal y los niveles de factores inflamatorios en lechones destetados. Manejo de la salud de Porc. (2023) 9:46. doi: 10.1186/s40813-023-00339-5
Resumen de PubMed | Texto completo de Crossref | Google Académico
42. Ma J, Zhu X, Wang Z, Yu X, Hu R, Wang X, et al. La suplementación con glutamina afectó a la comunidad bacteriana intestinal y a la fermentación, lo que llevó a una mejor digestibilidad de los nutrientes en los yaks con retraso en el crecimiento. FEMS Microbiol. Ecol. (2021) 97:fiab084. DOI: 10.1093/femsec/fiab084
Resumen de PubMed | Texto completo de Crossref | Google Académico
43. Halas D, Hansen CF, Hampson DJ, Mullan BP, Kim JC, Wilson RH, et al. La suplementación dietética con ácido benzoico mejora la digestibilidad ileal aparente del nitrógeno total y aumenta la altura de las vellosidades y la diversidad microbiana cecal en los cerdos destetados. Anim Feed Sci Technol. (2011) 163:261–7. doi: 10.1016/j.anifeedsci.2010.11.012
44. Gräber T, Kluge H, Hirche F, Broz J, Stangl GI. Efectos del ácido benzoico y el benzoato de sodio en la dieta sobre el rendimiento, el equilibrio de nitrógeno y minerales y la excreción de ácido hipúrico de lechones. Arch Anim Nutr. (2012) 66:227–36. doi: 10.1080/1745039X.2012.676812
Resumen de PubMed | Texto completo de Crossref | Google Académico
45. Kluge H, Broz J, Eder K. Efectos del ácido benzoico en la dieta sobre el pH urinario y la digestibilidad de los nutrientes en cerdas lactantes. Livest Sci. (2010) 134:119–21. doi: 10.1016/j.livsci.2010.06.116
46. Bühler K, Wenk C, Broz J, Gebert S. Influencia del ácido benzoico y el nivel de proteínas dietéticas en el rendimiento, el metabolismo del nitrógeno y el pH urinario en cerdos de engorde y engorde. Arch Anim Nutr. (2006) 60:382–9. doi: 10.1080/17450390600884369
Resumen de PubMed | Texto completo de Crossref | Google Académico
47. Schuba J, Südekum KH, Pfeffer E, Jayanegara A. Excreción de nitrógeno fecal, ureico urinario y no ureico urinario por cuatro especies de rumiantes influenciadas por la ingesta de nitrógeno en la dieta: un metaanálisis. Livest Sci. (2017) 198:82–8. doi: 10.1016/j.livsci.2017.01.017
48. George SK, Dipu MT, Mehra UR, Verma AK, Singh P. Influencia del nivel de ingesta de alimento en la concentración de derivados de purinas en muestras de manchas urinarias y suministro de nitrógeno microbiano en toros cruzados. Australas Asiáticas J Anim Sci. (2006) 19:1291–7. doi: 10.5713/ajas.2006.1291
49. Lewis HB. Estudios sobre la síntesis de ácido hipúrico en el organismo animal. Síntesis de ácido hipúrico en conejos con una dieta libre de glicocoles. J Biol Chem. (1914) 17:503–8. doi: 10.1016/S0021-9258(18)88391-0
50. Nousiainen J, Shingfield KJ, Huhtanen P. Evaluación del nitrógeno ureico de la leche como diagnóstico de la alimentación con proteínas. J Lácteos Sci. (2004) 87:386–98. doi: 10.3168/jds. S0022-0302(04)73178-1
Resumen de PubMed | Texto completo de Crossref | Google Académico
51. Choi H, Chen Y, Longo F, Kim SW. Efectos comparativos del ácido benzoico y el benzoato de sodio en dietas para cerdos de destete sobre el rendimiento del crecimiento y la acidificación de la digesta y la orina. J Anim Sci. (2023) 101:skad116. doi: 10.1093/jas/skad116
Resumen de PubMed | Texto completo de Crossref | Google Académico
52. Huang YH, Yan Q, Jiang MC, Guo S, Li HW, Lin M, et al. El aditivo Astragalus membranaceus mejora los parámetros bioquímicos séricos y el rendimiento reproductivo en vacas lecheras posparto. Front Vet Sci. (2022) 9:952137. doi: 10.3389/fvets.2022.952137
Resumen de PubMed | Texto completo de Crossref | Google Académico
53. Shu Y, Yu B, He J, Yu J, Zheng P, Yuan ZC, et al. El exceso de suplementos de ácido benzoico en la dieta conduce a un retraso del crecimiento, anomalías hematológicas y lesiones orgánicas en los lechones. Livest Sci. (2016) 190:94–103. doi: 10.1016/j.livsci.2016.06.010
54. Shin MY, Shin C, Choi JW, Lee J, Lee S, Kim S. Perfil farmacocinético del propilparabeno en humanos después de la administración oral. Environ Int. (2019) 130:9. doi: 10.1016/j.envint.2019.104917
Resumen de PubMed | Texto completo de Crossref | Google Académico
55. Baba S, Akira K, Suzuki H, Imachi M. Uso de la espectroscopia de resonancia magnética nuclear y el marcaje selectivo C para la investigación farmacocinética en el hombre: detección de la conversión de ácido benzoico en ácido hipúrico. Biol Pharm Bull. (1995) 18:643–7. doi: 10.1248/bpb.18.643
Resumen de PubMed | Texto completo de Crossref | Google Académico
56. Wang Y, Chiba LI, Huang C, Torres IM, Wang L, Welles EG. Efecto de la complejidad de la dieta, los complejos multienzimáticos, los aceites esenciales y el ácido benzoico en los cerdos destetados. Livest Sci. (2018) 209:32–8. doi: 10.1016/j.livsci.2017.12.007
57. Han X, Li MM, Sun LJ, Liu XJ, Yin Y, Hao JY, et al. El ácido p-hidroxibenzoico mejora la colitis al mejorar la barrera mucosa de una manera dependiente de la microbiota intestinal. Nutrientes. (2022) 14:5383. doi: 10.3390/nu14245383
Resumen de PubMed | Texto completo de Crossref | Google Académico
58. Recavarren MI, Milán GD. La velocidad y el patrón de la infusión de urea en el rumen de las hembras altera el equilibrio de nitrógeno y el amoníaco plasmático. J Anim Physiol Anim Nutr. (2014) 98:1047–53. doi: 10.1111/jpn.12168
Resumen de PubMed | Texto completo de Crossref | Google Académico
Palabras clave: ácido benzoico, corderos destetados, ácido hipúrico, rendimiento de crecimiento, metabolismo del nitrógeno
Cita: Zhang W, Sun S, Zhang Y, Zhang Y, Wang J, Liu Z y Yang K (2024) La suplementación con ácido benzoico mejora el rendimiento del crecimiento, la digestibilidad de los nutrientes y el metabolismo del nitrógeno de los corderos destetados. Frente. Vet. Sci. 11:1351394. doi: 10.3389/fvets.2024.1351394
Recibido: 06 de diciembre de 2023; Aceptado: 29 de enero de 2024;
Publicado: 09 Febrero 2024.
Editado por:
Rui Hu, Universidad Agrícola de Sichuan, China
Revisado por:
Adham Al-Sagheer, Universidad de Zagazig, Egipto
Bai Yan, Universidad de Qinghai, China
Derechos de autor © 2024 Zhang, Sun, Zhang, Zhang, Wang, Liu y Yang. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Kailun Yang, ykl@xjau.edu.cn
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía