
Reducción de la producción de metano a partir de heces almacenadas de vacas lecheras

Reducción de la producción de metano a partir de heces almacenadas de vacas lecheras por Asparagopsis taxiformis
 Mohammad Ramin1*,
Mohammad Ramin1*,  Juana C. C. Chagas1,
Juana C. C. Chagas1,  Yash Pal1,
Yash Pal1,  Rebecca Danielsson2,
Rebecca Danielsson2,  Petra Fant1 y
Petra Fant1 y  Sophie J. Krizsan2
Sophie J. Krizsan2- 1Departamento de Nutrición y Gestión Animal, Universidad Sueca de Ciencias Agrícolas, Umeå, Suecia
- 2Departamento de Nutrición y Gestión Animal, Universidad Sueca de Ciencias Agrícolas, Uppsala, Suecia
El objetivo fue evaluar si el metano (CH4) la producción a partir de heces almacenadas de vacas previamente suplementadas con Asparagopsis taxiformis (AT) en su dieta fue menor en comparación con las heces de vacas no suplementadas con AT. También se investigó la posibilidad de reducir aún más el CH4 producción mediante la adición de AT a las heces almacenadas de las vacas. Se proporcionaron muestras fecales de un ensayo de alimentación (durante dos períodos diferentes) de cuatro vacas divididas en dos grupos diferentes. Un grupo fue suplementado con AT a un nivel de 0,5% de la ingesta total de materia orgánica, y el otro grupo no fue suplementado con AT. Se estableció un diseño factorial de 2 × 2 en el laboratorio para la incubación de heces. Las muestras fecales de los dos grupos de vacas se dividieron en dos submuestras que no recibieron adición de AT o la adición de AT a un nivel de 0,5% de OM incubado. Esto resultó en cuatro tratamientos con dos réplicas por período. El mismo diseño se repitió durante el segundo período. En total, se incubaron 400 g de muestras fecales frescas en frascos de suero de 1 L durante 9 semanas a 39°C en un baño maría. CH4 y la producción total de gas se midió en los días 1, 4 y 7 y posteriormente cada 2ª semana hasta el final del período de incubación. CH entérico4 la producción mostró una reducción significativa (61%) cuando se complementó AT en la dieta de vacas lecheras. Encontramos que CH4 la producción de las heces de vacas lecheras suplementadas con AT en su dieta fue solo numéricamente menor (P = 0.61). La adición de AT a las heces de las vacas lecheras redujo significativamente el CH4 producción de las heces en un 44% en comparación con las heces sin AT. No se observaron diferencias en los perfiles de la comunidad bacteriana y arqueal de las muestras fecales entre las vacas alimentadas con TA y las no alimentadas con AT. Este estudio concluye que la adición de AT a las heces almacenadas puede reducir eficazmente el CH4 producción a partir de las heces de las vacas lecheras.
Introducción
Ha habido muchos esfuerzos para disminuir la huella de carbono de las actividades humanas para frenar el cambio climático en todo el mundo. Dentro del sector ganadero, el metano entérico (CH4) las emisiones del sector de los rumiantes desempeñan un papel clave. Rodeado de rumiantes, el sector lácteo es uno de los contribuyentes más importantes a CH4 Emisiones. Se estima que el ganado rumiante emite alrededor de un tercio de CH antropogénico4 (Jackson et al., 2020). La mayoría de los CH4 producido por una vaca lechera es de fermentación entérica, pero la segunda fuente más grande de CH4 es la descomposición de las heces de las vacas (Agencia de Protección Ambiental de los Estados Unidos, 2006). Se estima que el estiércol almacenado en el clima europeo frío-templado es responsable de ~ 12% del CH total4 emisiones del sistema lácteo (Hindrichsen et al., 2015). Sin embargo, el estiércol también se utiliza para la producción de biogás, que es beneficioso para el medio ambiente a través de la producción de energía renovable (Huhtanen et al., 2021). La contribución del estiércol a las emisiones de gases de efecto invernadero (GEI) depende de muchos factores, como la composición química del estiércol, las condiciones en las que se almacena el estiércol y la temperatura (Hassanat y Benchaar, 2019). Por ejemplo, las heces incubadas de vacas alimentadas frente a no alimentadas con aceite de colza produjeron 3.45 y 3.85 L, respectivamente, de CH4 por kg de materia seca de material incubado (Ramin et al., 2021). Las estrategias dietéticas, como la alimentación con alto contenido de concentrados, disminuirán la producción entérica de CH4 al tiempo que aumenta la producción fecal de materia orgánica potencialmente digerible (pdOM; Huhtanen et al., 2021). Esto aumentará la producción de CH4 del estiércol a medida que aumenta la cantidad de sustrato fermentable que termina en las heces (Møller et al., 2014). En Suecia y algunos otros países nórdicos, la metodología de Nivel 2 del IPCC se utiliza para predecir CH4 producción a partir de estiércol (Grupo Intergubernamental de Expertos sobre el Cambio Climático, 2006). Las ecuaciones de predicción desarrolladas por el IPCC no tienen en cuenta los efectos del uso de compuestos inhibidores o la inducción de cambios en la composición de las heces debido a cambios en la dieta. Esto conducirá a una sobreestimación/subestimación de CH4 producción a partir de estiércol.
Se han reportado varias estrategias para disminuir el CH entérico4 producción de vacas lecheras. Recientemente, una macroalga roja nativa de áreas tropicales, Asparagopsis taxiformis (AT), fue identificada como un CH entérico natural.4 mitigador. El compuesto halogenado más explotado que se encuentra en AT es el bromoformo (CHBr3). El bromoformo mitiga el CH4 producción en el rumen al perjudicar el paso final en la metanogénesis (Duin et al., 2016). Stefenoni et al. (2021) mostraron una disminución de ~65% en CH4 producción de vacas lecheras alimentadas con TA mediante la inclusión de TA al 0,5% de la ingesta de materia seca (DMI). Sin embargo, existen preocupaciones sobre el uso de AT en la dieta de las vacas lecheras, ya que se han identificado rastros de CHBr3 en la leche de vacas lecheras (Stefenoni et al., 2021), y un estudio reciente mostró daños inflamatorios en la pared ruminal de vacas lecheras suplementadas con AT (Muizelaar et al., 2021). Una solución más práctica para usar AT para reducir CH4 Las emisiones podrían ser la adición de AT al estiércol de las vacas lecheras. Solo unos pocos estudios en la literatura presentan estrategias para disminuir los GEI del estiércol, y la mayoría de ellos están relacionados con las emisiones de óxido nitroso (N2O).
El primer objetivo de este estudio fue estimar CH4 producción a partir de heces almacenadas de vacas previamente suplementadas o no suplementadas con AT en su dieta. El segundo objetivo fue evaluar si CH4 La producción podría mitigarse agregando AT a las heces de las vacas lecheras.
Planteamos la hipótesis de que las heces de vacas suplementadas con AT en su dieta producirían menos CH4 que las heces de vacas no suplementadas con AT y que la adición de AT a las heces de las vacas incubadas podría afectar la metanogénesis durante la descomposición de las heces, lo que resulta en una disminución de CH4 producción.
Materiales y métodos
Recolectamos muestras fecales en relación con un ensayo de alimentación realizado en la granja experimental Röbäcksdalen de la Universidad Sueca de Ciencias Agrícolas en Umeå (63 ° 45 ‘N, 20 ° 17’ E) que tuvo como objetivo evaluar el efecto de la suplementación de AT a una dieta basada en ensilaje de pasto alimentada a vacas lecheras. Se pueden encontrar más detalles sobre el ensayo de alimentación en el estudio de Krizsan et al. (2023). En resumen, seis vacas lecheras Nordic Red con 122 ± 13,7 días (media ± DE) en leche, paridad 2,7 ± 0,52, y produciendo 36 ± 2,5 kg de leche/día al inicio del experimento fueron asignadas a un diseño de cambio cuadrado latino que comprende dos tratamientos dietéticos (dieta suplementada y no suplementada con AT). Las vacas se mantenían en un establo aislado con acceso gratuito a agua dulce. Las vacas fueron alimentadas con una dieta basal que consistía en ensilaje de pasto y un concentrado comercial en una proporción de 60:40 sobre una base de materia seca. La dieta basal no se complementó o se complementó con 0,5% de TA sobre una base de ingesta total de materia orgánica (OM). Las dietas fueron alimentadas a las vacas como una ración mixta total (TMR) ad libitum. Las dietas se mezclaron utilizando el mezclador TMR (Nolan A / S) y se entregaron a los comederos tres veces al día mediante un vagón de alimentación automática. La ingesta de alimento se monitorizó con alimentadores de control de ingesta de forraje (Insentec B.V., Marknesse, Países Bajos).
CH entérico4 la producción se midió utilizando el sistema GreenFeed (C-Lock Inc., Rapid City, SD, EUA), según lo descrito por Huhtanen et al. (2015). El GreenFeed se configuró para permitir que las seis vacas en el experimento (previamente entrenadas) visitaran el sistema a intervalos de 5 horas. Para atraer a los animales a visitar el GreenFeed regularmente, ocho gotas de ~ 50 g (cada gota) de concentrado granulado, el mismo concentrado comercial (Komplett Amin 180; Lantmännen Lantbruk AB, Malmö, Suecia) como en el TMR, se dieron a las vacas. El intervalo establecido entre cada gota fue de 40 s.
Se recogieron muestras fecales durante dos períodos de dos vacas no suplementadas con AT (control) y de dos vacas suplementadas con AT a un nivel del 0,5% de la ingesta de OM. Recolectamos muestras fecales de solo dos vacas por tratamiento, ya que teníamos espacio limitado en el laboratorio para mediciones de CH fecal.4 producción. Se estableció un diseño factorial de 2 × 2 para la incubación de heces en el laboratorio. Cada muestra fecal (cuatro en total) se dividió en dos submuestras. Cada submuestra recibió uno de los dos tratamientos; sin adición de AT a la muestra fecal (control) o adición de AT a la muestra fecal a un nivel de 0,5% de OM incubado. Esto dio lugar a cuatro tratamientos con dos réplicas por tratamiento por período. El mismo diseño se repitió durante el segundo período. El diseño experimental aplicado a la incubación de heces para medir CH fecal4 producción se muestra en la Figura 1.
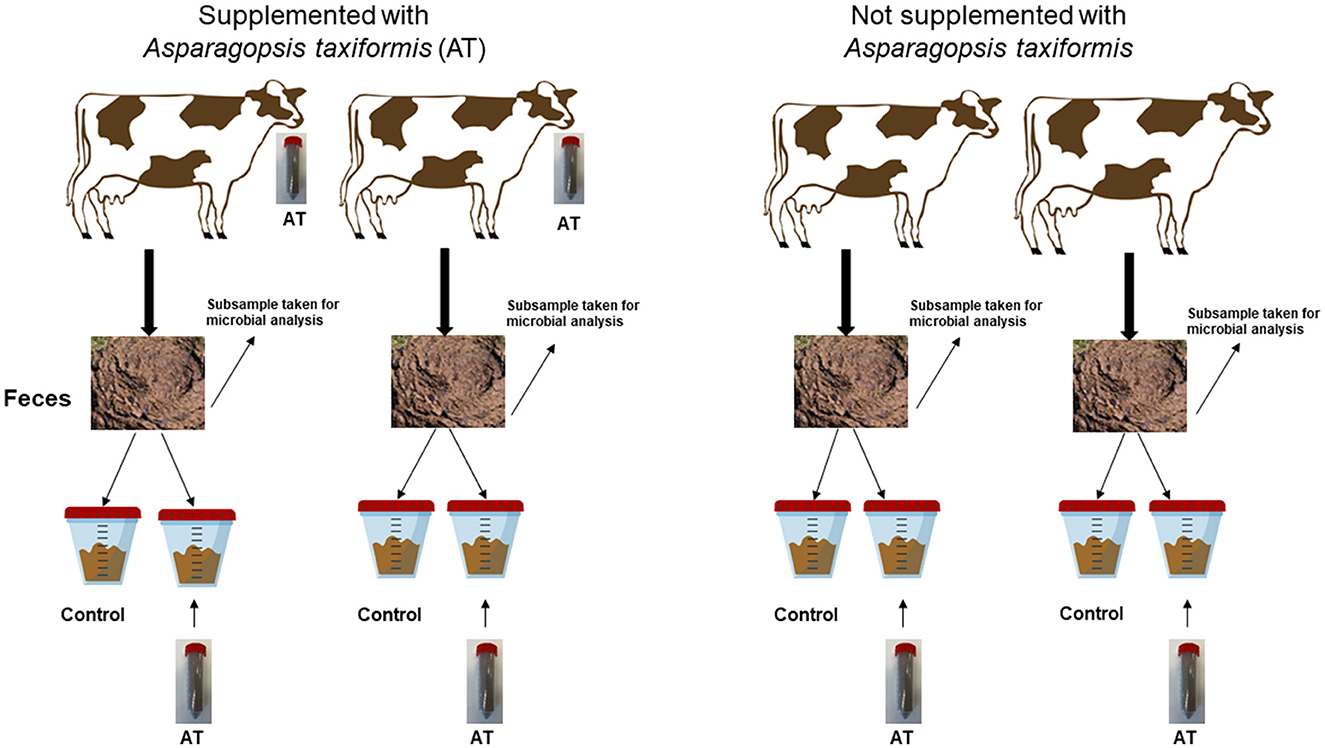 Figura 1. Una visión esquemática del diseño experimental aplicado a la incubación de heces para medir CH fecal4 producción.
Figura 1. Una visión esquemática del diseño experimental aplicado a la incubación de heces para medir CH fecal4 producción.
Se utilizó el método descrito por Ramin et al. (2021) para medir CH4 producción a partir de heces. En resumen, 400 g de muestra fecal fresca recolectada de las vacas se incubaron en ocho botellas de suero de 1 L durante 9 semanas a 39 ° C en un baño de agua. Se añadió Asparagopsis taxiformis a cuatro de los frascos (Figura 1). Las botellas se agitaban suavemente tres veces por semana. Los frascos de suero de 1 L se conectaron a bolsas de muestreo de lámina inerte herméticas al gas (Supel)TM-Supelco, Estados Unidos). El metano y la producción total de gas se midieron en los días 1, 4 y 7 y luego cada 2ª semana hasta el final de la incubación.
CH acumulativo4 la producción se calculó sobre la base de la suma de CH4 producción en cada punto de tiempo de la siguiente manera:
Metano (ml) = HSCH4 × 600 + Bolsa CH4×Gas total producido (ml),Metano(Ml) = HSCH4 × 600 + Bolsa CH4 ×Gas total producido(Ml),
donde HSCH4 es el espacio de cabeza CH4 concentración; bolsa CH4 es el CH4 concentración en la bolsa de muestreo; y 600 es el volumen de espacio de cabeza en las botellas de suero. La concentración de CH4 en la muestra de gas se determinó inyectando 0,2 ml de gas extraído del espacio de cabeza/bolsa de muestreo en un cromatógrafo de gases utilizando una jeringa estanca al gas, como se describe en Ramin y Huhtanen (2012).
Las muestras fecales de las cuatro vacas seleccionadas para las incubaciones fecales se secaron a 105°C durante 16 h. Posteriormente, las muestras se molieron utilizando un molino de corte (SM 300, Retsch Ltd., Haan, Alemania) para pasar a través de un tamiz de 1 mm para el análisis de fibra detergente neutra (NDF) y OM. Las submuestras también se molieron para pasar un tamiz de 2 mm para el análisis de NDF no digerible (iNDF). La ceniza se determinó en muestras fecales secas por combustión a 500 °C durante 4 h (AOAC International, 2000). La materia orgánica se calculó como 1.000 cenizas. La concentración de NDF de muestras fecales se determinó utilizando α-amilasa estable al calor y sulfito de sodio (Mertens, 2002) en un analizador de fibra Ankom200 (Ankom Technology Corp., Macedon, NY, EUA). El NDF no digerible se determinó como muestras duplicadas de 2 g que se pesaron en bolsas de poliéster de 11 μm de tamaño de poro (Krizsan et al., 2015). Las bolsas se incubaron durante 288 h en el rumen de tres vacas canuladas que fueron alimentadas con una dieta basada en ensilaje de pasto, y la iNDF se expresó sin cenizas (Huhtanen et al., 1994). El NDF potencialmente digerible (pdNDF, g/kg MS) se calculó como la diferencia entre NDF y iNDF. El detergente neutro soluble (NDS) se calculó como la diferencia entre OM y NDF. El pdOM se calculó como la suma de NDS y pdNDF.
Se recolectaron submuestras de 15 g de las heces de vacas suplementadas con y sin AT en la dieta, se congelaron a -80 ° C y se enviaron a Novogene, Reino Unido, para el análisis de la comunidad microbiana. El ADN comunitario se aisló utilizando el kit de ADN en heces del amplificador TIAN, según las instrucciones del fabricante con ligeras modificaciones. El ADN extraído de las heces se sometió a la amplificación del gen 16S rRNA (región V4), dirigida tanto a bacterias como a arqueas utilizando el par de cebadores 515F (GTGBCAGCMGCCGCGGTAA) / 805R (GACTACHVGGGTATCTAATCC). El producto amplificado se utilizó además para la secuenciación de meta-códigos de barras utilizando la estrategia PE250 en la plataforma de secuenciación Illumina (Hi Seq 2500), y las lecturas de secuenciación sin procesar se procesaron utilizando USEARCH versión 11 (Edgar, 2010), según Pal et al. (2022). Los archivos OTU se utilizaron además para la clasificación utilizando el clasificador RDP versión 2.11, versión de la base de datos 11.5 (Cole et al., 2014), y la imagen final se renderizó en MEGAN-LR (Huson et al., 2018). Las OTU que representan taxones arqueales se extrajeron utilizando MEGAN-LR para la representación gráfica (Huson et al., 2018).
Ingesta de materia seca, hidrógeno entérico y CH entérico4 los datos de producción se sometieron al análisis de varianza utilizando el procedimiento PROC GLM en SAS (SAS Inc. 2002–2003, Release 9.4 SAS Inst. Inc., Cary, NC, USA) aplicando el siguiente modelo:
YIJK=μ+DYo+Pj+Ck+ εIJK,YIJK=μ+DYo+Pj+Ck+ εIJK,
donde μ = la media global, DYo = el efecto del tratamiento dietético (suplementado con TA o no), Pj = el efecto del período, Ck = el efecto de la vaca, y εIJK = error residual aleatorio.
Se adoptó el siguiente modelo (PROC GLM) utilizando el paquete estadístico de SAS (9.4) para el análisis de CH4 y la producción total de gas a partir de heces almacenadas:
YIJK=μ+FYo+Unj+Pk+(FA)Ij+ εIJK,YIJK=μ+FYo+Unj+Pk+(FA)Ij+ εIJK,
donde μ = la media global, FYo = el efecto del nivel Yo de factor F (vaca) suplementado con AT a dieta o no, Aj = el efecto del nivel j del factor A añadiendo AT a las heces o no, Pk = el efecto del nivel k del período, (FA)Ij = el efecto de la interacción entre F y A, y εIJK = error residual aleatorio.
Resultados
La composición química de las heces de vacas con o sin suplementación de AT en su dieta se presenta en la Tabla 1. El contenido de pdOM en las heces de vacas suplementadas con AT fue bastante similar a las heces de vacas no suplementadas con AT (Tabla 1). La suplementación de AT a la dieta de vacas lecheras disminuyó el CH entérico diario4 producción, g/d (P = 0,01) en un 61% y DMI (P = 0,03) en 3,7 kg/d (Tabla 2). El rendimiento de metano también disminuyó (P = 0,02) de las vacas suplementadas con AT (9 g/kg DMI más bajo) que de las vacas no suplementadas con AT (Tabla 2). Como consecuencia de la reducción de CH4 producción de vacas suplementadas con AT en su dieta, la producción de hidrógeno fue mayor (P = 0,02) para las vacas suplementadas con AT (5,02 vs. 1,07 g/d, respectivamente; Tabla 2). La producción de metano fue numéricamente menor (P = 0,61) en las heces de vacas suplementadas con AT en su dieta en comparación con las heces de vacas no suplementadas con AT en su dieta, 3,73 vs. 4,13 L/kg MS, respectivamente (Tabla 3). Adición de AT a las heces de vacas reducidas (P < 0,01) CH4 producción de las heces en un 44% en comparación con las heces sin la adición de AT, 2,84 vs. 5,04 L/kg MS (Tabla 3). La producción total de gas también fue menor (P = 0,03) en las heces con la adición de AT que en las heces sin la adición de AT (Tabla 3).
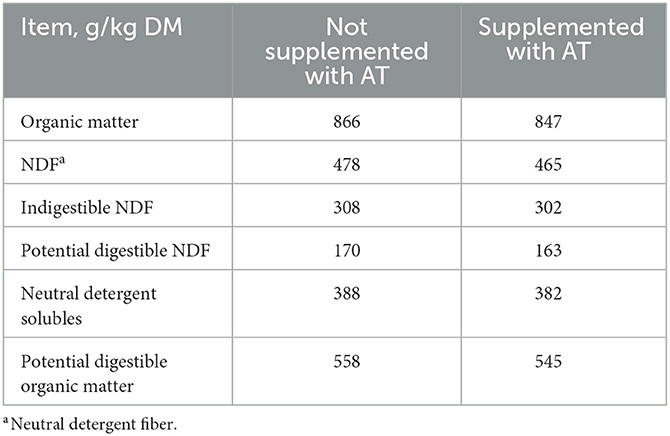 Tabla 1. Composición química de las heces de vacas (n = 4) no suplementadas o complementadas con Asparagopsis taxiformis (AT) en su dieta.
Tabla 1. Composición química de las heces de vacas (n = 4) no suplementadas o complementadas con Asparagopsis taxiformis (AT) en su dieta.
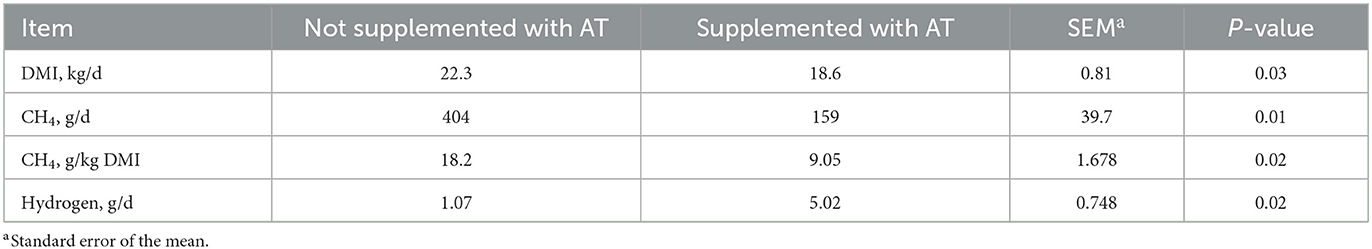 Tabla 2. Efecto de la suplementación de Asparagopsis taxiformis (AT) a la dieta de vacas lecheras (n = 4) sobre la ingesta de materia seca (DMI) y metano entérico (CH4) producción.
Tabla 2. Efecto de la suplementación de Asparagopsis taxiformis (AT) a la dieta de vacas lecheras (n = 4) sobre la ingesta de materia seca (DMI) y metano entérico (CH4) producción.
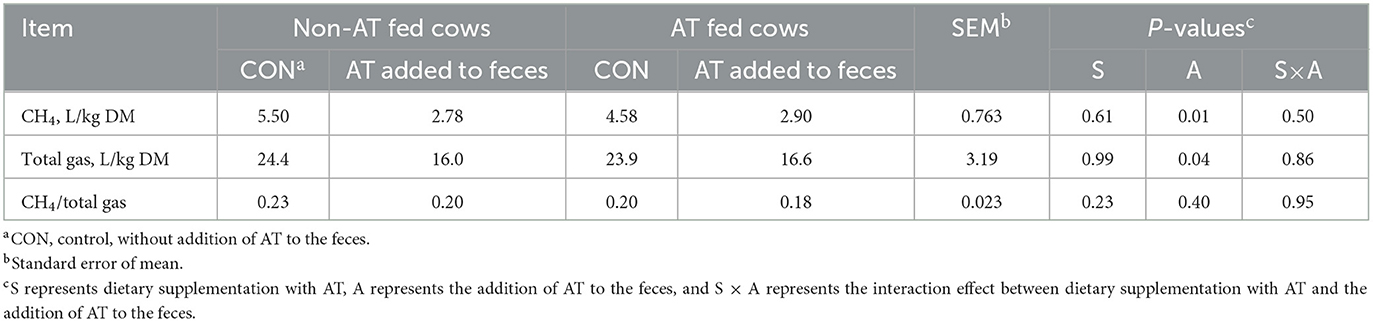 Tabla 3. Metano (CH4) producción a partir de heces almacenadas (n = 8) de vacas suplementadas o no con Asparagopsis taxiformis (AT) en su dieta y con o sin adición de AT a las heces.
Tabla 3. Metano (CH4) producción a partir de heces almacenadas (n = 8) de vacas suplementadas o no con Asparagopsis taxiformis (AT) en su dieta y con o sin adición de AT a las heces.
En ocho muestras, se observaron un total de 823.839 lecturas limpias con una longitud de lectura promedio de 102.980. Se observaron un total de 19.638 OTU, entre las cuales 61 OTU se clasificaron como arqueas y 19.181 OTU como bacterias. No se observó ninguna variación significativa en los perfiles de comunidades arqueales y bacterianas de muestras de heces de vacas suplementadas con dietas suplementadas con AT y no AT. Firmicutes y Bacteroidetes fueron los filos bacterianos más dominantes, seguidos por Proteobacteria, Actinobacteria y Acidobacteria, constituyendo más de ~95% de la abundancia relativa total de OTU (Figura 2A). Además, Gemmatimonadetes, Synergistetes y Planctomycetes solo se observaron en muestras que no eran AT. Euryarchaeota fue el único filo arqueal observado en muestras AT y no AT. A nivel de género, Methanobrevibacter, Methanosphaera y Methanocorpusculum (dentro del filo Euryarchaeota) estuvieron presentes en muestras AT y no AT (Figura 2B). La riqueza promedio de especies (Chao-1) fue mayor en el grupo alimentado con AT en comparación con el grupo no alimentado con AT, mientras que no se observaron diferencias contrastantes en la diversidad alfa (Simpson y Shannon) y la diversidad beta (datos no mostrados).
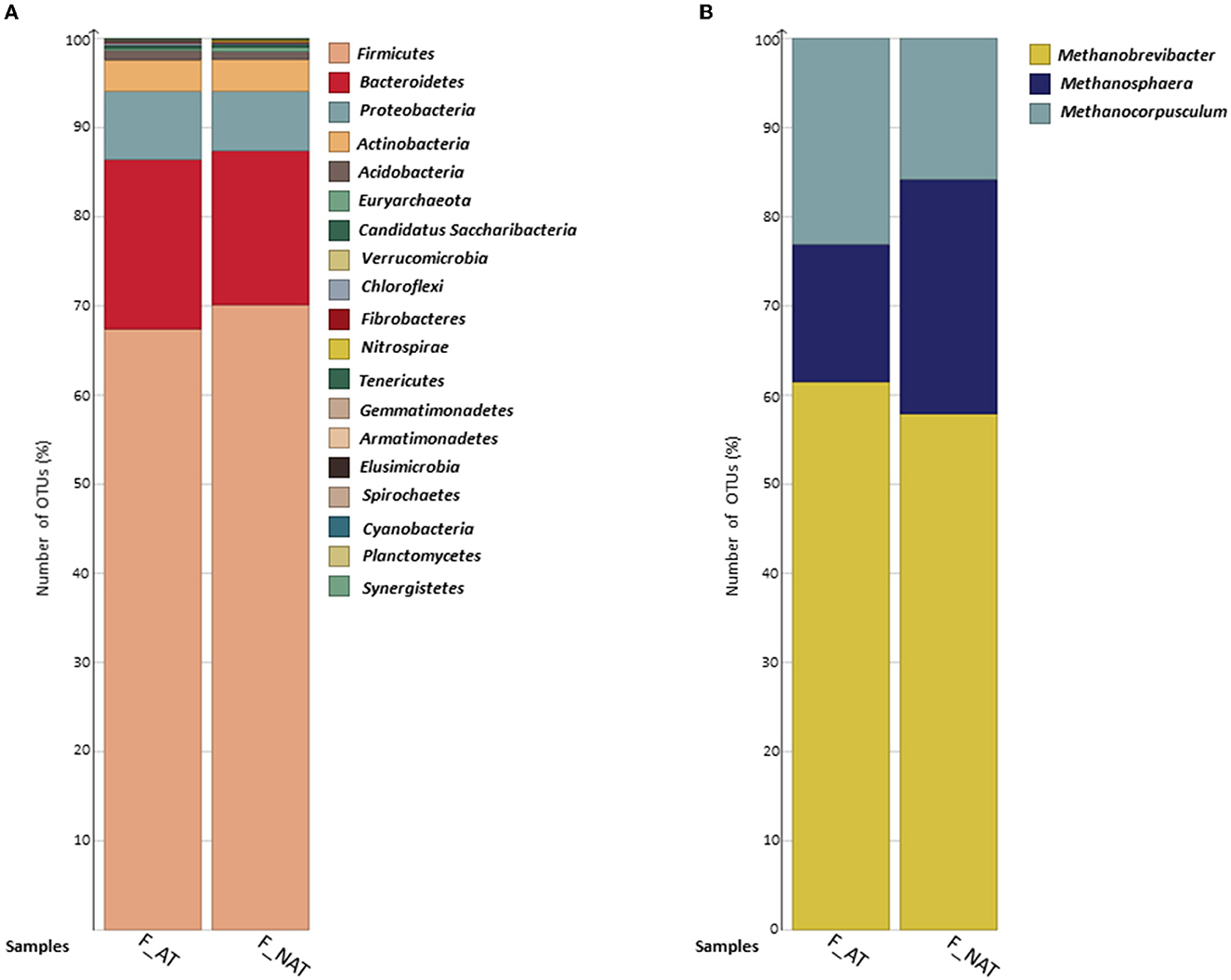 Figura 2. El gráfico de barras representa la distribución porcentual de OTU (A) bacterianas y (B) arqueales a nivel de filo y género, respectivamente. Muestras AT (n = 4) y NAT (n = 4). Las OTU arqueales se extrajeron del total de OTU observadas para una mejor visualización. AT, heces de vacas suplementadas con Asparagopsis taxiformis en su dieta; NAT, heces de vacas no suplementadas con Asparagopsis taxiformis en su dieta.
Figura 2. El gráfico de barras representa la distribución porcentual de OTU (A) bacterianas y (B) arqueales a nivel de filo y género, respectivamente. Muestras AT (n = 4) y NAT (n = 4). Las OTU arqueales se extrajeron del total de OTU observadas para una mejor visualización. AT, heces de vacas suplementadas con Asparagopsis taxiformis en su dieta; NAT, heces de vacas no suplementadas con Asparagopsis taxiformis en su dieta.
Discusión
Demostramos que la suplementación de AT a la dieta de las vacas lecheras nórdicas reduce significativamente el DMI y el CH entérico4 producción. También encontramos que agregar AT a las heces de las vacas lecheras disminuye aún más CH4 producción. La producción de metano de las heces de las vacas que previamente han sido suplementadas con AT en su dieta no disminuye significativamente en comparación con las heces de vacas no suplementadas con AT. Sin embargo, la adición de AT a las heces mostró que alrededor del 44% de CH4 La producción podría reducirse a partir de las heces de las vacas lecheras.
Hasta la fecha, AT es el inhibidor de origen natural más prometedor de CH4 producción de vacas lecheras (Wasson et al., 2022). El CH más reportado4 El inhibidor del compuesto activo en AT es CHBr3, y actúa inhibiendo la enzima metil-coenzima M reductasa (MCR), que cataliza el paso final en la metanogénesis en arqueas ruminales (Duin et al., 2016). Dado que el compuesto principal reduce CH4 en AT es CHBr3, como lo muestran Stefenoni et al. (2021) en vacas lecheras, la adición de otros compuestos halogenados es un método factible para reducir CH4 producción a partir de estiércol también. Inhibición de CH4 La producción de digestores anaeróbicos es una preocupación relevante hoy en día para el sector ganadero que pretende utilizar el estiércol en sus campos (Wasson et al., 2022). Además de CH4, N2O es otro GEI que tiene un efecto significativo sobre el calentamiento global con un potencial de calentamiento global 265 veces mayor que el dióxido de carbono (Grupo Intergubernamental de Expertos sobre el Cambio Climático, 2014). En comparación con CH4 producción, N2O podría mitigarse más fácilmente del estiércol, ya que la optimización dietética del consumo de proteínas conducirá a una mejor utilización del nitrógeno y dará como resultado una menor pérdida de nitrógeno para el estiércol (Novak y Fiorelli, 2010). Sin embargo, la CH entérica atenuante4 La producción de vacas lecheras parece ser más desafiante, ya que también podría afectar la productividad de los animales. Además, podría ser más fácil reducir las emisiones de CH4 de estiércol, aunque una menor cantidad de CH4 se produce a partir del estiércol en comparación con la fermentación entérica. También debemos tener en cuenta que complementar las vacas lecheras con TA y los beneficios ambientales relacionados dependerá de la aceptación por parte del productor del costo actual de TA (Wasson et al., 2022).
También podrían probarse y utilizarse otros compuestos halogenados, por ejemplo, 3-nitrooxipropanol (3NOP). Utilizamos AT como el CH4 inhibidor en este estudio ya que 3NOP y AT ambos tienen el mismo modo de acción y bloquean la enzima en el proceso de fermentación en los metanógenos, produciendo CH4 en el rumen. El uso de 3NOP podría ser una opción más barata tanto para alimentar al animal como para agregar estiércol para mitigar el CH4 producción. El efecto de 3NOP en CH4 sin embargo, la inhibición en las vacas lecheras es menor que el efecto de la AT. En un estudio realizado por Hristov et al. (2015), la CH entérica4 la producción se redujo en ~ 30% de las vacas lecheras suplementadas con 3NOP en comparación con el grupo de control. Suplementar las vacas lecheras con TA también tiene efectos potencialmente negativos en su rendimiento de producción. Encontramos que el DMI se redujo significativamente, similar a los hallazgos de Stefenoni et al. (2021). Además, encontraron que la producción de leche disminuyó, lo que también está respaldado por los hallazgos de Krizsan et al. (2023), quienes mostraron una disminución en el rendimiento de leche corregido energéticamente de vacas suplementadas con AT. El uso de AT en la dieta también podría limitar la adopción del mercado si se observan efectos secundarios negativos de la suplementación con AT (Wasson et al., 2022). En un estudio reciente, Muizelaar et al. (2021) informaron que la alimentación de AT a las vacas lecheras mostró signos de inflamación en el rumen.
Una suposición es que las estrategias dietéticas que reducen el CH entérico4 la producción se compensa con un aumento de CH4 producción a partir del estiércol, que se denomina compensación entre CH entérico y fecal4 producción. Esto podría deberse a la presencia de carbohidratos solubles (sólidos volátiles) en las heces, que luego se fermentan y se convierten en CH4. Benchaar et al. (2015) encontraron que la adición de aceite de linaza a las vacas lecheras alimentadas con una dieta basada en ensilaje de maíz causó una disminución en el CH entérico4 producción (−26%). Sin embargo, Hassanat y Benchaar (2019) encontraron que CH4 La producción (L / kg OM) de las heces fue mayor en las vacas alimentadas con una dieta basada en ensilaje de maíz suplementada con aceite de linaza debido a la derivación de sustratos fermentables en las heces. Sin embargo, en este estudio, pdOM (también podría denominarse sólidos volátiles) en las heces fue bastante similar entre el grupo de vacas suplementadas con AT en su dieta y el grupo de control de vacas no suplementadas con AT. Esto también está en línea con las diferencias no significativas en CH4 producción de heces entre los mismos dos grupos de vacas.
Los estudios que analizan la comunidad arqueal y bacteriana en las heces de vacas suplementadas con AT en su dieta son raros. Uno de los objetivos del presente estudio fue evaluar si la población de las comunidades bacterianas y arqueales diferirá entre las heces de vacas suplementadas con AT y las heces de vacas no suplementadas con AT. A nivel de phyla, Firmicutes fue el grupo más abundante, seguido de Bacteroidetes, que está de acuerdo con estudios previos donde se han examinado las comunidades microbianas en las heces de vacas lecheras (Callaway et al., 2010; Hagey y otros, 2019; Williamson et al., 2022). En el estudio de Williamson et al. (2022), hubo claras diferencias entre la comunidad microbiana en el rumen y la comunidad microbiana en las heces, y la principal diferencia se relacionó con un aumento de Firmicutes en relación con Bacteroidetes. Sin embargo, para la comunidad bacteriana, no se observaron diferencias en las heces de vacas suplementadas con AT o no suplementadas con AT. Henderson et al. (2015) realizaron un estudio con 742 muestras ruminales de composición de la comunidad microbiana del intestino anterior de 35 países, y encontraron que bacterias y arqueas similares dominaban en casi todas las muestras.
El efecto de AT está relacionado principalmente con la inhibición de la metanogénesis en la comunidad arqueal, y los efectos adicionales sobre la población bacteriana no están bien estudiados. En este estudio, las arqueas solo fueron representadas por Euryarchaeota a nivel de phyla, lo que está en línea con el estudio de Jin et al. (2017). La mayoría de los metanógenos en el estiércol líquido se originan en el intestino del animal, y Methanobrevibacter es, en general, el género más abundante en las vacas lecheras, tanto en el rumen como en las heces (Henderson et al., 2015; Jin et al., 2017). En este estudio, se encontró que Methanobrevibacter es el género más abundante en las heces de las vacas, independientemente de si sus dietas se complementaron con AT o no. La similitud observada entre los tratamientos en la comunidad microbiana de estiércol se alinea con la falta de un efecto sobre CH4 producción (L/kg MS) a partir de heces.
Se anticipa que habrá una distinción notable en los tratamientos como resultado de la inhibición de la metanogénesis en metanógenos dentro del rumen, lo que posteriormente conducirá a cambios observables en las heces. Sin embargo, se ha demostrado que el CH4-compuesto inhibidor presente en AT, CHBr3, se metaboliza rápidamente en el rumen (Romero et al., 2022) y que no se excreta en el estiércol de los animales (Muizelaar et al., 2021). Debido a la rápida degradación de CHBr3 en el rumen (Muizelaar et al., 2021), es posible que no haya un efecto sobre los metanógenos activos en las heces.
Nuestros resultados mostraron que el efecto inhibitorio de complementar las dietas de las vacas lecheras con TA fue menor en CH4 producción de heces comparada con el efecto sobre CH4 Producción a partir de fermentación entérica. Asparagopsis taxiformis produce una amplia gama de diferentes compuestos halogenados (Woolard et al., 1979), y la potencia del efecto inhibitorio de estos diferentes compuestos varía (Lanigan, 1972). Nuestros resultados revelaron que CH4 La producción de las heces de las vacas suplementadas con AT en su dieta fue numéricamente menor en comparación con las heces de vacas no suplementadas con AT. Sin embargo, debemos señalar que este estudio fue un estudio piloto; aportó nuevos conocimientos sobre la reducción de CH4 producción a partir de estiércol. Se necesita más investigación con un mayor número de vacas en experimentos futuros para confirmar los resultados de este estudio. La producción de metano podría reducirse aún más mediante la adición de TA al estiércol. No obstante, también se requieren más estudios para dilucidar las interacciones entre los compuestos halogenados de los TA y el microbioma de las heces.
Conclusión
Este estudio concluyó que el CH entérico diario4 producción y CH4 el rendimiento se redujo cuando AT se complementó con la dieta de las vacas lecheras Nordic Red. Sin embargo, debe tenerse en cuenta que el DMI también se vio afectado y reducido por la suplementación dietética de TA. La producción de metano a partir de las heces podría reducirse hasta en un 44% con la adición de AT a las heces almacenadas. No se observaron diferencias en la distribución de las comunidades bacterianas y arqueales entre las heces de las vacas que fueron suplementadas con AT en su dieta y las que no lo fueron. Se necesitan más estudios para investigar las interacciones entre los compuestos halogenados de los TA y el microbioma fecal, con el fin de proporcionar una comprensión más profunda de sus efectos.
Declaración de disponibilidad de datos
Los conjuntos de datos presentados en este estudio se pueden encontrar en repositorios en línea. Los nombres del repositorio / repositorios y el número (s) de acceso se pueden encontrar en: https://www.ncbi.nlm.nih.gov/genbank/, PRJNA906285.
Declaración ética
El manejo de animales en este experimento fue aprobado por el Comité de Ética Sueco sobre Investigación Animal (Dnr A 32-16), representado por el Tribunal de Apelación de Norrland del Norte en Umeå, y el experimento se llevó a cabo de acuerdo con las leyes y regulaciones que rigen los experimentos realizados con animales vivos en Suecia.
Contribuciones del autor
MR: conceptualización, análisis formal, adquisición de fondos, metodología, administración del proyecto, recursos, supervisión y redacción del manuscrito. JC: metodología y ayudó en la redacción del manuscrito. YP: metodología, análisis microbiano y redacción del manuscrito. RD: análisis microbiano y edición del manuscrito. PF: análisis estadístico y redacción y edición del manuscrito. SK: conceptualización y redacción del manuscrito. Todos los autores contribuyeron al artículo y aprobaron la versión presentada.
Financiación
Este estudio fue financiado en parte por el Proyecto RumenPredict (FACCE ERA-GAS) como parte del Programa de Investigación e Innovación Horizonte 2020 de la UE y Carl Tryggers Stiftelsen, Suecia (número de proyecto: 24837801).
Reconocimientos
Agradecemos al personal que trabaja en la granja que ayudó con el ensayo in vivo y al personal del laboratorio por su apoyo en la realización de este estudio.
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o reclamo que pueda ser hecho por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
Benchaar, C., Hassanat, F., Martineau, R. y Gervais, R. (2015). Suplementación con aceite de linaza a vacas lecheras alimentadas con ensilaje de trébol rojo o dietas a base de ensilaje de maíz: efectos sobre la producción de metano, fermentación ruminal, digestibilidad de nutrientes, equilibrio de N y producción de leche. J. Dairy Sci. 98, 7993–8008. DOI: 10.3168/JDS.2015-9398
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Callaway, T. R., Dowd, S. E., Edrington, T. S., Anderson, R. C., Krueger, N., Bauer, N., et al. (2010). Evaluación de la diversidad bacteriana en el rumen y las heces de ganado alimentado con diferentes niveles de granos secos de destilería más solubles utilizando pirosecuenciación de amplicón FLX codificado por etiqueta bacteriana. J. Anim. 12, 3977–3983. DOI: 10.2527/JAS.2010-2900
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Cole, J. R., Wang, Q., Fish, J. A., Chai, B., McGarrell, D. M., Sun, Y., et al. (2014). Ribosomal Database Project: datos y herramientas para el análisis de rRNA de alto rendimiento. Ácidos nucleicos res. 42, D633–D642. doi: 10.1093/nar/gkt1244
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Duin, E. C., Wagner, T., Shima, S., Prakash, D., Cronin, B., Yáñez-Ruiz, D. R., et al. (2016). Modo de acción descubierto para la reducción específica de las emisiones de metano de los rumiantes por la pequeña molécula 3-nitrooxipropanol. Proc. Natl. Acad. Sci. U. S. A. 113, 6172–6177. doi: 10.1073/PNAS.1600298113
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Edgar, R. C. (2010). Busque y agrupe órdenes de magnitud más rápido que BLAST. Bioinformática 26, 2460–2461. DOI: 10.1093/bioinformatics/btq461
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Hagey, J. V., Bhatnagar, S., Heguy, J. M., Karle, B. M., Price, P. L., Meyer, D., et al. (2019). Comunidades microbianas fecales en una gran cohorte representativa de vacas lecheras de California. Frente. Microbiol. 10, 1093. doi: 10.3389/fmicb.2019.01093
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Hassanat, F. y Benchaar, C. (2019). Emisiones de metano del estiércol de vacas lecheras alimentadas con trébol rojo o dietas basadas en ensilaje de maíz complementadas con aceite de linaza. J. Lácteos. Ciencia. 102, 11766–11776. DOI: 10.3168/JDS.2018-16014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Henderson, G., Cox, F., Ganesh, S., Jonker, A., Young, W., Census, G. R., et al. (2015). La composición de la comunidad microbiana del rumen varía con la dieta y el huésped, pero se encuentra un microbioma central en un amplio rango geográfico. 5, 14567. DOI: 10.1038/SEP14567
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Hindrichsen, I. K., Wettstein, H. R., Machmüller, A., Jörg, B. y Kreuzer, M. (2015). Efecto de la composición de carbohidratos de los concentrados de alimentos sobre la emisión de metano de las vacas lecheras y su purín. Entorno Mont. Asses. 107, 329–350. DOI: 10.1007/S10661-005-3008-3
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Hristov, A. N., Oh, J., Giallongo, F., Frederick, T., Harper, M., Weeks, H., et al. (2015). Un inhibidor disminuyó persistentemente la emisión entérica de metano de las vacas lecheras sin ningún efecto negativo en la producción de leche. Proc. Natl. Acad. Sci. U. S. A 112, 10663–10668. doi: 10.1073/pnas.1504124112
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Huhtanen, P., Cabezas-Garcia, E. H., Utsumi, S. y Zimmerman, S. (2015). Comparación de métodos para determinar las emisiones de metano de las vacas lecheras en condiciones de granja. J. Dairy Sci. 98, 3394–3409. DOI: 10.3168/JDS.2014-9118
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Huhtanen, P., Kaustell, K. y Jaakkola, S. (1994). El uso de marcadores internos para predecir la digestibilidad total y el flujo duodenal de nu-triens en bovinos con seis dietas diferentes. Anim. Feed Sci. Technol. 48, 211–227. doi: 10.1016/0377-8401(94)90173-2
Huhtanen, P., Krizsan, S. J. y Ramin, M. (2021). Modelización de la producción fecal y la composición de nutrientes, y la emisión máxima de metano del estiércol de las vacas lecheras. Anim. Feed Sci. Tech. 282, 115120. doi: 10.1016/j.anifeedsci.2021.115120
Huson, D. H., Albrecht, B., Bagci, C., Bessarab, I., Górska, A., Jolic, D., et al. (2018). MEGAN-LR: los nuevos algoritmos permiten un binning preciso y una fácil exploración interactiva de lecturas largas y contigs metagenómicos. Biol. Directo. 13, 6. DOI: 10.1186/S13062-018-0208-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Grupo Intergubernamental de Expertos sobre el Cambio Climático (2006). «Capítulo 10 emisiones de la gestión del ganado y el estiércol», en las Directrices del IPCC para los inventarios nacionales de gases de efecto invernadero, vol. 4. Agriculture, Forestry and Other Land Use, preparado por el Programa Nacional de Inventarios de Gases de Efecto Invernadero, eds S. Eggleston, L. Buendia, K. Miwa, T. Ngara y K. Tanabe (IGES). Disponible en línea en: http://www.ipcc-~nggip.iges.or.jp/public/2006gl/vol4.html (consultado el 25 de febrero de 2023).
Grupo Intergubernamental de Expertos sobre el Cambio Climático (2014). «AR5. Cambio climático 2014: mitigación», en Contribución del Grupo de Trabajo III al Quinto Informe de Evaluación del Panel Intergubernamental sobre el Cambio Climático, eds O. Edenhofer, et al. (Nueva York, NY: Cambridge University Press). Disponible en línea en: https://www.ipcc.ch/report/ar5/wg3/ (consultado el 7 de noviembre de 2022).
Jackson, R. B., Saunois, M., Bousquet, P., Canadell, J. G., Poulter, B., Stavert, A. R., et al. (2020). El aumento de las emisiones antropogénicas de metano proviene igualmente de fuentes agrícolas y de combustibles fósiles. Lett 15, 1002. DOI: 10.1088/1748-9326/AB9ED2
Jin, D., Kang, K., Wang, H., Wang, Z., Xue, B., Wang, L., et al. (2017). Efectos de la suplementación dietética de levadura seca activa sobre la diversidad de arqueas metanogénicas fecales en vacas lecheras. Anaerobio 44, 78–86. doi: 10.1016/j.anaerobe.2017.02.007
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Krizsan, S. J., Ramin, M., Chagas, J. C. C., Halmemies-Beauchet-Filleau, A., Singh, A., Schnürer, A., et al. (2023). Los efectos sobre el microbioma ruminal y la calidad de la leche de las vacas lecheras complementaron la macroalga Asparagopsis taxiformis en una dieta basada en ensilaje de pasto. Frente. Anim. Sci. Anim. Nutr. 2023, 1112969. doi: 10.3389/fanim.2023.1112969
Krizsan, S. J., Rinne, M., Nyholm, L. y Huhtanen, P. (2015). Nuevas recomendaciones para la determinación ruminal in situ de fibra detergente neutra indigerible. Anim. Feed Sci. Technol. 205, 31–41. doi: 10.1016/j.anifeedsci.2015.04.008
Lanigan, G. (1972). Metabolismo de los alcaloides pirrolizidina en el rumen ovino. IV. Efectos del hidrato de cloral y los metanos halogenados sobre la metanogénesis ruminal y el metabolismo alcaloide en ovejas fistuladas. Cosecha. Pasto Sci. 23, 1085–1091. doi: 10.1071/AR9721085
Mertens, D. R. (2002). Determinación gravimétrica de fibra detergente neutra tratada con amilasa en piensos con reflujo en vasos de precipitados o crisoles: estudio colaborativo. J. AOAC Int. 85, 1217–1240.
Møller, H. B., Moset, V., Brask, M., Weisbjerg, M. R. y Lund, P. (2014). Composición de heces y rendimiento de metano derivado del estiércol de vacas lecheras: influencia de la dieta con enfoque en el suplemento de grasa y el tipo de forraje. Ambiente. 94, 36–43. doi: 10.1016/j.atmosenv.2014.05.009
Muizelaar, W., Groot, M., van Duinkerken, G., Peters, R. y Dijkstra, J. (2021). Estudio de seguridad y transferencia: transferencia del bromoformo presente en Asparagopsis taxiformis a la leche y orina de vacas lecheras lactantes. Alimentos 10, 584. doi: 10.3390/foods10030584
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Novak, S. M. y Fiorelli, J. L. (2010). Gases de efecto invernadero y emisiones de amoníaco de sistemas orgánicos mixtos de cultivos y productos lácteos: una revisión crítica de las opciones de mitigación. Agron. Sostener. Desarrollador. 30, 215–236. DOI: 10.1051/AGRO/2009031
Pal, Y., Mayilraj, S. y Krishnamurthi, S. (2022). Explorar la distribución distinta de las comunidades arqueales en sitios contaminados con explosivos. Biomoléculas 12, 489. DOI: 10.3390/biom12040489
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Ramin, M., Chagas, J. C., Smidt, H., Gomez Exposito, R. y Krizsan, S. (2021). Emisiones de metano entéricas y fecales de vacas lecheras alimentadas con dietas de ensilaje de pasto o maíz complementadas con aceite de colza. Animales 11, 1322. DOI: 10.3390/ANI11051322
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Ramin, M., y Huhtanen, P. (2012). Desarrollo de un método in vitro para la determinación de la cinética de producción de metano utilizando un sistema de gas in vitro totalmente automatizado: un enfoque de modelado. Anim. Feed Sci. Technol. 174, 190–200. doi: 10.1016/j.anifeedsci.2012.03.008
Romero, P., Belanche, A., Hueso, R., Ramos-Morales, E., Salwen, J. K., Kebreab, E., et al. (2022). «In vivo rumen microbial degradation of bromoform and the impact on rumen fermentation», en 8th Annual Global Greenhouse Gas & Animal Agriculture Conference, (Orlando, FL), 291.
Stefenoni H. A. Räisänen S. E. Cueva S. F. Wasson D. E. Lage C. F. A. Melgar A. . . (2021). Efectos de la macroalga Asparagopsis taxiformis y las hojas de orégano sobre la emisión de metano, la fermentación ruminal y el rendimiento de la lactancia de las vacas lecheras. J. Dairy Sci. 104, 4157–4173. DOI: 10.3168/JDS.2020-19686
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Agencia de Protección Ambiental de los Estados Unidos (2006). Antropogénico global no-CO2 Emisiones de gases de efecto invernadero: 1990-2020, EPA 430-R-06-003. Washington DC: Agencia de Protección Ambiental de los Estados Unidos.
Wasson, D. E., Yarish, C. y Hristov, A. N. (2022). Mitigación entérica del metano a través de la suplementación con Asparagopsis taxiformis y posibles alternativas de algas. Frente. Anim. Sci. 3, 999338. doi: 10.3389/fanim.2022.999338
Williamson, J. R., Callaway, T. R., Lourenco, J. M. y Ryman, V. E. (2022). Caracterización de la microbiota ruminal, fecal y láctea en vacas lecheras lactantes. Frente. Microbiol. 13, 984119. doi: 10.3389/fmicb.2022.984119
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Cita: Ramin M, Chagas JCC, Pal Y, Danielsson R, Fant P y Krizsan SJ (2023) Reducción de la producción de metano a partir de heces almacenadas de vacas lecheras por Asparagopsis taxiformis. Frente. Sostener. Sistema de Alimentos. 7:1187838. DOI: 10.3389/FSUFS.2023.1187838
Recibido: 16 de marzo de 2023; Aprobado: 09 Junio 2023;
Publicado: 13 julio 2023.
Editado por:
Arif Reza, Universidad de Stony Brook, Estados Unidos
Revisado por:
Wenbing Tan, Academia China de Investigación de Ciencias Ambientales, China
Ashish Kumar Das, Universidad Jashore de Ciencia y Tecnología, Bangladesh
Derechos de autor © 2023 Ramin, Chagas, Pal, Danielsson, Fant y Krizsan. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Mohammad Ramin, mohammad.ramin@slu.se
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía