
Suplementación dietética con β-mananasa y probióticos como estrategia para mejorar el rendimiento de las gallinas ponedoras

Suplementación dietética con β-mananasa y probióticos como estrategia para mejorar el rendimiento de las gallinas ponedoras y la calidad de los huevos
 Camila Lopes Carvalho1,2
Camila Lopes Carvalho1,2  Inés Andretta1*
Inés Andretta1*  Gabriela Miotto Galli1
Gabriela Miotto Galli1  Thais Bastos Stefanello1
Thais Bastos Stefanello1  Nathalia de Oliveira Telesca Camargo1 Ricardo Evandro Mendes3
Nathalia de Oliveira Telesca Camargo1 Ricardo Evandro Mendes3  Giovanna Pelisser3
Giovanna Pelisser3  Balasubramanian Balamuralikrishnan4 Raquel Melchor1
Balasubramanian Balamuralikrishnan4 Raquel Melchor1  Marcos Kipper5
Marcos Kipper5- 1Departamento de Ciencia Animal, Facultad de Agronomía, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brasil
- 2Departamento de Patología, Farmacología y Medicina Zoológica, Facultad de Medicina Veterinaria, Universidad de Gante, Merelbeke, Bélgica
- 3Laboratorio de Patología Veterinaria, Instituto Federal Catarinense, Concórdia, Brasil
- 4Departamento de Ciencia y Biotecnología de los Alimentos, Universidad de Sejong, Seúl (República de Corea)
- 5Elanco Animal Health, São Paulo, Brasil
El objetivo de este estudio fue evaluar el impacto de la β-mananasa y los probióticos en el rendimiento, la bioquímica sérica, los rasgos morfométricos intestinales y la calidad de los huevos frescos de las gallinas ponedoras. Un total de 120 jaulas, que albergaban gallinas ponedoras livianas (36 semanas de edad), fueron asignadas aleatoriamente a cuatro tratamientos diferentes. Estos tratamientos incluyeron un grupo control alimentado con dietas no suplementadas; dietas suplementadas con 300 g/tonelada de beta-mananasa; dietas suplementadas con 50 g/tonelada de probiótico; o dietas que contengan 300 g/tonelada de β-mananasa y 50 g/tonelada de probióticos. El ensayo tuvo una duración de 26 semanas y se dividió en tres fases productivas, cada una con una duración de 28 días. La inclusión de β-mananasa resultó en una mejora significativa en la tasa de puesta en un 11% (p < 0,05) en comparación con el tratamiento control. Del mismo modo, la adición de probióticos también mejoró la tasa de puesta en un 7% (p < 0,05), así como la suplementación con aditivos combinados (11,5%). Los aditivos combinados mostraron un aumento en las masas de huevos, y la asociación de aditivos mejoró en un 13,9% (p < 0,001) en contraste con el tratamiento control. En general, la β-mananasa y los aditivos combinados utilizados durante el período de suplementación dieron lugar a mejoras en el peso de los huevos frescos. Estos beneficios se observaron después de un período de 14 semanas sin suplementación (p < 0,05). Además, se observaron diferencias significativas en la bioquímica sérica y en las masas de huevos de las aves que fueron alimentadas con dietas que contenían ambos aditivos (β-mananasa + probióticos) en comparación con el grupo control. Parámetros como el ácido úrico, el colesterol total y los triglicéridos mostraron variaciones notables. La altura de las vellosidades: profundidad de la cripta mostró diferencias con los aditivos combinados (β-mananasa + probióticos). La β-mananasa mejoró la gravedad específica, la altura, la longitud y el pH de la yema, y los rasgos de color de la yema en comparación con el tratamiento de control. El uso de probióticos ayudó a mejorar la altura de la yema, el pH y la puntuación de color. Además, los aditivos combinados (β-mananasa + probióticos) mejoran la altura, la longitud, el peso, el pH y el pH de la yema. Por lo tanto, la incorporación de β-mananasa y probióticos en las dietas de las gallinas ponedoras demuestra ser una estrategia altamente efectiva para mejorar la tasa de puesta y el estado de salud general, al mismo tiempo que eleva ciertos atributos de calidad de los huevos frescos.
1 Introducción
En 2019, la Organización Mundial de la Salud (OMS) lanzó una campaña en la que pedía a los gobiernos que adoptaran medidas para contener la resistencia a los antimicrobianos. El uso inadecuado de antibióticos tanto en medicina humana como en producción animal se ha convertido en un problema de salud pública, que se ha ido agravando (1). El uso de antibióticos es menos frecuente en aves ponedoras debido a la posibilidad de residuos en los huevos. Aun así, el uso de aditivos alimentarios es una posible alternativa para mejorar la productividad, el estado sanitario e incluso la calidad de los huevos.
Los probióticos fueron definidos por Fuller (2) como un suplemento compuesto por microorganismos vivos que benefician al huésped y mejoran su equilibrio microbiano intestinal. Los mecanismos de acción de los probióticos se producen a través de diferentes procesos, que pueden o no estar asociados. Los efectos físicos surgen de la exclusión competitiva o de la competencia por los sitios de unión en la mucosa intestinal. Las bacterias beneficiosas presentes en los probióticos forman una barrera física protectora, evitando que los patógenos oportunistas ocupen los mismos sitios de unión. Los efectos biológicos se producen debido a la presencia de bacterias anaeróbicas en los probióticos. Estas bacterias promueven un ambiente de baja tensión de oxígeno dentro del intestino, lo que inhibe el crecimiento de patógenos, creando un ambiente desfavorable para su supervivencia y reproducción. Los efectos químicos se observan a través de la producción de bacteriocinas por parte de las bacterias probióticas. Las bacteriocinas son sustancias antimicrobianas que son eficaces contra diversos patógenos, inhibiendo aún más su crecimiento y reduciendo su impacto en el organismo. Además, los probióticos pueden tener un efecto nutricional, proporcionando nutrientes esenciales y promoviendo un entorno intestinal saludable, lo que favorece el crecimiento y el mantenimiento de bacterias beneficiosas (3-6).
La suplementación enzimática es una estrategia valiosa para mejorar la salud intestinal al mitigar los efectos de los componentes antinutricionales (7). La β-mananasa, en particular, puede ayudar a los animales no rumiantes a digerir polisacáridos sin almidón, que de otro modo pueden dificultar la digestibilidad de los nutrientes (8-11). Estos polisacáridos, principalmente β-mananos, son abundantes en las paredes celulares de las plantas y se encuentran comúnmente en ingredientes de alimentos para animales como la soja (12). β-mananos también están presentes en la superficie de los microorganismos, activando el sistema inmunitario innato del animal y provocando la activación de monocitos, macrófagos, células dendríticas y un aumento de la producción de citoquinas. En consecuencia, esto incurre en un gasto energético innecesario y una mayor respuesta inflamatoria (13). Al hidrolizar β-mananos, esta enzima mejora la digestibilidad del manano, aumenta la población de bacterias beneficiosas, fortalece la inmunidad, mejora la digestión y absorción de nutrientes y restringe la proliferación de patógenos potenciales en el intestino (7).
Aunque las ventajas mencionadas anteriormente son dignas de mención, es importante señalar que la mayoría de los datos existentes se derivaron de diferentes categorías de aves de corral, específicamente pollos de engorde. Además, ambos aditivos funcionan de manera complementaria, lo que sugiere el potencial de efectos sinérgicos cuando se combinan en la suplementación alimentaria. Actualmente no hay literatura disponible que describa los efectos combinados de estos aditivos. Por lo tanto, el objetivo de este estudio fue evaluar si la suplementación de gallinas ponedoras comerciales con β-mananasa y probióticos solos o en combinación podría mejorar el rendimiento, el estado de salud y la calidad del huevo.
2 Materiales y métodos
2.1 Animales, alojamiento y diseño experimental
El protocolo experimental descrito fue aprobado por el Comité Institucional de Ética en el Uso de Animales (CEUA/UFRGS) bajo el número de protocolo 39783. Las unidades experimentales consistieron en gallinas seleccionadas al azar de una granja comercial ubicada en Salvador do Sul, Rio Grande do Sul, Brasil. La granja albergaba aproximadamente 28.000 gallinas ponedoras ligeras del linaje Hyline W 36, de 36 semanas de edad. Para el ensayo, se utilizaron 120 jaulas, cada una con cuatro aves, como réplicas. Estas réplicas se asignaron aleatoriamente a los cuatro tratamientos. Los tratamientos consistieron en: (1) Tratamiento control (CON): Este grupo recibió una dieta basal sin ningún tipo de suplementación con aditivos adicionales. (2) Tratamiento con β-mananasa (BMA): Las aves de este grupo recibieron la dieta control suplementada con 300 g/tonelada de β-mananasa. (3) Tratamiento con probióticos (PRO): Las aves de este grupo recibieron la dieta de control suplementada con 50 g/tonelada de un aditivo probiótico multicepa. (4) Tratamiento con β-mananasa + probiótico (BMA + PRO): Las aves de este grupo recibieron la dieta control suplementada con 300 g/tonelada de β-mananasa y 50 g/tonelada de un probiótico multicepa.
La β-mananasa empleada en este ensayo fue Hemicell™ HT, procedente de Elanco Animal Health en São Paulo, Brasil. Es una enzima exógena derivada de la fermentación de la bacteria Paenibacillus lentus. El aditivo probiótico utilizado en este ensayo fue el concentrado de Protexin™, también proporcionado por Elanco Animal Health en São Paulo, Brasil. El probiótico comprende una combinación de cepas bacterianas beneficiosas, que incluyen: Lactobacillus acidophilus (2,06 × 108 UFC/g), L. bulgaricus (2,06 × 108 UFC/g), L. plantarum (1,26 × 108 UFC/g), L. rhamnosus (2,06 × 108 UFC/g), Bifidobacterium bifidum (2,0 × 108 UFC/g), Enterococcus faecium (6,46 × 108 UFC/g) y Streptococcus thermophilus (4,10 × 108 UFC/g).
El experimento duró 26 semanas. Durante los primeros 84 días del ensayo, las aves recibieron suplementos. Para facilitar la evaluación, este período se dividió en tres fases distintas: fase 1 (36-40 semanas), fase 2 (41-44 semanas) y fase 3 (45-48 semanas). Al finalizar el período de suplementación, todas las aves pasaron a la dieta de control durante un período de 14 semanas. Posteriormente, se realizó una nueva evaluación en la semana 62 para valorar los resultados.
La dieta basal (Tabla 1) se formuló como un alimento a base de harina de maíz-soya para cumplir con los requerimientos nutricionales genéticos (14). Para tener en cuenta la ausencia de β-mananasa y/o aditivos probióticos, se incorporó material inerte en forma de caolín en el alimento basal. Durante todo el período experimental, las aves tuvieron acceso ad libitum a alimento y agua. Se utilizaron bebederos de tetina y comederos de canaleta, lo que garantizó el acceso sin restricciones para las aves.
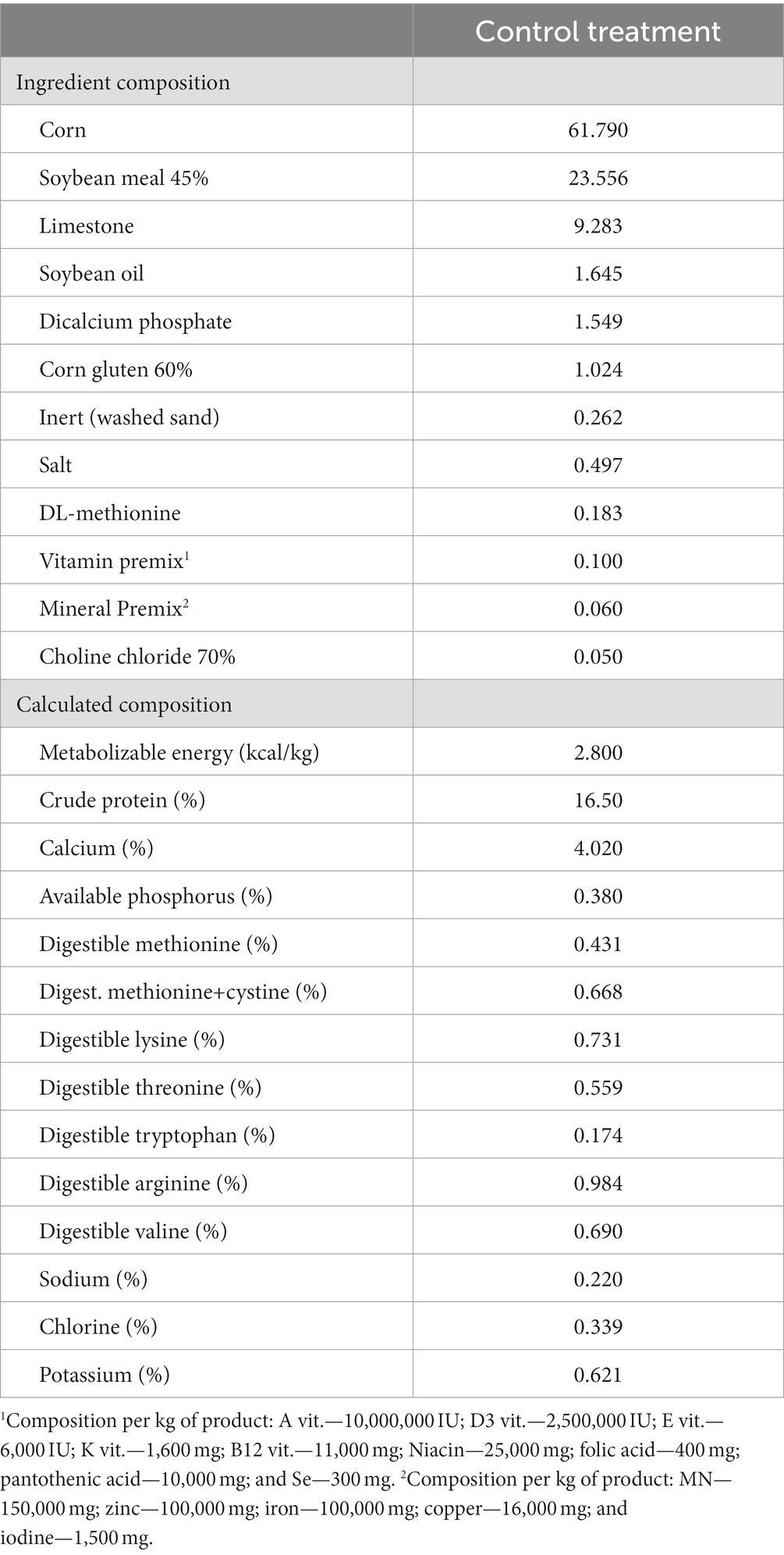 Tabla 1. Composición de la dieta de control.
Tabla 1. Composición de la dieta de control.
Las aves se alojaron en galpones convencionales que estaban orientados en dirección este-oeste. Estos cobertizos se construyeron con pisos de concreto y paredes de mampostería, y malla de alambre se extendía hasta el techo. Para garantizar un confort térmico óptimo, los cobertizos se equiparon con cortinas laterales ajustables, que se gestionaron en función de las condiciones climáticas imperantes. El régimen de iluminación siguió un horario de 16 h de luz a partir de las 04:00 pm hasta las 08:00 am seguido de 8 h de oscuridad cada día, proporcionando un patrón de iluminación consistente para las aves. A lo largo de toda la duración del experimento, las aves fueron alojadas en jaulas de alambre galvanizado. Estas jaulas medían 100 cm de largo, 40 cm de ancho y 45 cm de alto, ofreciendo una superficie de suelo de 500 cm2 por gallina.
2.2 Análisis de rendimiento
Se evaluó la producción de huevos en las semanas 4, 8 y 12 en 120 jaulas con cuatro aves cada una, correspondientes a 30 repeticiones por tratamiento. Todos los huevos producidos se pesaron individualmente. La tasa de puesta y la masa de huevos se calcularon considerando todos los huevos (incluidos los huevos no comercializables) para cada réplica (jaula). El coeficiente de variabilidad se calculó para cada jaula considerando el peso individual de todos los huevos producidos cada semana. El mismo procedimiento se adoptó para las masas de huevos [Masa de huevos = peso medio (g) × porcentaje de producción de huevos %]. Debido a las limitaciones de manejo relacionadas con el sistema comercial, la medición de la ingesta de alimento no fue posible en este estudio. Por esa razón, tampoco se evaluó la conversión alimenticia.
2.3 Grado de suciedad de las cáscaras de huevo
Todos los huevos producidos en cada repetición en las semanas 4, 8 y 12 fueron inspeccionados individualmente para detectar la presencia de heces en las cáscaras, que fueron clasificadas por el mismo observador a través del análisis visual como huevos limpios (ausentes; puntuación 0), presencia menor (puntuación 1) y presencia mayor (puntuación 3 y 4). Durante el análisis de los datos, las puntuaciones 3 y 4 fueron consideradas conjuntamente debido a la baja casuística de la puntuación 3.
2.4 Bioquímica sérica
Al final de la semana 12, se recogieron muestras de sangre de ocho aves seleccionadas al azar en cada tratamiento. Las muestras se obtuvieron de la vena cubital de las aves, las cuales fueron escogidas de diferentes jaulas. Para la obtención del suero, la sangre se recolectó en tubos sin anticoagulante. Las muestras de sangre recogidas se sometieron a centrifugación a 3.500 rpm durante 10 min. Este proceso permitió la separación del suero de otros componentes de la sangre. El suero resultante se recolectó cuidadosamente y se congeló a -20 °C para preservar su integridad para su posterior análisis bioquímico.
Para el análisis bioquímico, las muestras de suero congeladas fueron procesadas y examinadas utilizando el Analizador Bioquímico Bio-Plus 2000®, fabricado por Bioplus en São Paulo, Brasil. Se utilizaron kits comerciales de Wiener Lab Group, São Paulo, Brasil, para medir varios parámetros, incluyendo proteína total, albúmina, ácido úrico, colesterol total, triglicéridos, glucosa, fosfatasa alcalina, alanina aminotransferasa y aspartato aminotransferasa.
2.5 Prueba de parasitología
Al final del período experimental, se recolectaron muestras de excrementos de tres aves en cada una de las 10 jaulas. La recolección se realizó con cuidado, asegurando que las muestras se procesaran dentro de las 2 horas posteriores a la recolección para mantener su integridad. Se empleó la técnica de flotación centrífuga (15) para procesar las muestras de excretas. Para iniciar el proceso, se diluyó una submuestra de 1 g de excreta en 15 mL de solución de sacarosa. Esta muestra diluida se sometió a centrifugación durante 5 min. Después de la centrifugación, se utilizó un microscopio óptico para examinar la muestra en un portaobjetos de vidrio. El microscopio se ajustó a aumentos de 10, 40 y 100x para garantizar un recuento preciso de ooquistes.
2.6 Análisis morfométricos intestinales
De acuerdo con las normas de bienestar animal y eutanasia descritas en las directrices de práctica de la eutanasia del Consejo Nacional para el Control de la Experimentación Animal (16), seis aves por tratamiento fueron sacrificadas de forma humanitaria utilizando la luxación cervical. Después de la eutanasia, se recolectaron muestras de 2 cm del duodeno, yeyuno y ciego de cada ave. Estas muestras de tejido se almacenaron cuidadosamente en frascos que contenían una solución de formaldehído al 10% para preservar su integridad estructural.
Los portaobjetos histológicos se prepararon a partir de las muestras de tejido recolectadas y se tiñeron con hematoxilina y eosina (H&E) archivadas. Para capturar las imágenes histológicas, se utilizó una cámara ocular digital de microcámara, junto con un microscopio trinocular biológico modelo TNB-41 T-PL a un aumento de 40x. Para determinar la profundidad de la cripta y la longitud de las vellosidades, se midió una línea desde la base de la cripta hasta la parte superior para determinar la profundidad de la cripta. Para la longitud de las vellosidades, se trazó una línea recta desde la punta de las vellosidades hasta la parte superior de las criptas. Se empleó el software ImageJ incluido con Java 64.1.8_0 de 172 bits para mediciones precisas.
2.7 Calidad de los huevos frescos
El último día de las semanas 4, 8 y 12, se recolectaron aleatoriamente un total de 15 óvulos frescos de cada tratamiento en cada fase para evaluar su calidad. Los huevos rotos se excluyeron de esta evaluación. Para determinar la gravedad específica, se midió el peso de los huevos tanto en el aire como en el agua, siguiendo el principio de Arquímedes.
La altura de la albúmina se determinó midiendo tres puntos diferentes en la albúmina, cada uno a 10 mm de distancia de la yema, utilizando un calibrador digital (TMX PD-150, China). El promedio de estas medidas se utilizó para calcular la Unidad de Haugh (HU), empleando la ecuación desarrollada por Haugh (17).
La calidad de la yema se evaluó mediante el cálculo del índice vitelino, que es la relación entre la altura de la yema y el diámetro de la yema, utilizando la fórmula: YI = altura de la yema (mm)/diámetro de la yema (mm). La altura de la yema (mm) se midió con un altímetro, mientras que el diámetro de la yema (mm) se midió con un calibrador digital.
Las medidas de ancho y alto de la yema (mm) se obtuvieron utilizando un calibrador digital (TMX PD-150, China). El índice de yema se calculó mediante la fórmula: índice de yema = peso de la yema/ancho de la yema.
Para evaluar el color de la yema, se empleó el abanico colorimétrico Roche (DSM, São Paulo, Brasil). Este ventilador utiliza un sistema de puntuación que va desde 1 (que representa un color amarillo claro) hasta 15 (que indica un tono naranja rojizo). Además, se empleó un dispositivo espectrofotómetro (Delta Vista modelo 450G, Delta Color, São Leopoldo, Brasil) para esta evaluación, proporcionando coordenadas colorimétricas de luminosidad (L*), intensidad de rojo (a*) e intensidad de amarillo (b*). El croma, que representa el color real de la yema para el análisis, se estimó utilizando la siguiente ecuación: C = √(a2 + b2).
Una vez separadas la yema y la albúmina, se midieron los pesos de ambos componentes. Para asegurar la uniformidad, las porciones densas y fluidas de la albúmina, así como la yema, se homogeneizaron durante 20 s. Para evaluar los niveles de pH, se empleó un medidor de pH digital (Kasvi modelo k39-2014B, Paraná, Brasil).
El contenido sólido total de la albúmina y la yema se evaluó individualmente. Para ello, se pesaron 5 g de albúmina y yema en crisoles de porcelana presecados. A continuación, los crisoles que contenían las muestras se colocaron en un horno a 60 °C durante 12 h. Tras el proceso de secado, los crisoles se volvieron a pesar para determinar el peso de la albúmina y la yema secas. Para obtener el contenido final de sólidos, los crisoles se sometieron a una temperatura más alta de 105 °C durante otras 12 h. Después de este paso de secado, los crisoles se pesaron nuevamente para calcular con precisión el contenido sólido total de la albúmina y la yema.
Para determinar el peso de la concha, se siguió un procedimiento separado. Las conchas se separaron cuidadosamente, se lavaron a fondo y se secaron. Posteriormente, las conchas secas se pesaron con precisión para obtener su peso.
2.8 Análisis estadístico
El análisis de los datos se realizó utilizando el programa estadístico SAS (v 9.3, SAS Institute Inc., Cary, NC). Las unidades experimentales variaron entre las respuestas, pero brevemente, fue la jaula para el rendimiento, el ave para las respuestas bioquímicas e intestinales, y cada huevo para la evaluación de la calidad. Los datos se sometieron a pruebas de normalidad y homocedasticidad y luego se sometieron a análisis de varianza utilizando PROC MIXED, excepto el coeficiente de varianza del peso del huevo, que se analizó utilizando PROC GLIMMIX. Todos los modelos estadísticos incluyeron el efecto fijo de los tratamientos y el error. Los datos de rendimiento se analizaron considerando medidas repetidas a lo largo del tiempo. La calidad de los óvulos se analizó considerando también el efecto aleatorio de la fase en el modelo, aunque aquí solo se presentan medias agrupadas debido a la falta de interacción entre el tratamiento y la fase. Las diferencias de medias eventuales se compararon mediante la prueba de Tukey con una probabilidad del 5 y el 10%.
3 Resultados
Las mediciones de temperatura y humedad se obtuvieron utilizando un datalogger. Los valores promedio registrados para las temperaturas mínimas y máximas fueron de 18 y 36°C, respectivamente. Los valores medios de humedad relativa del aire oscilaron entre el 35,8 y el 94,7%. Este estudio se llevó a cabo en Salvador do Sul (región sur, Brasil), que experimenta en gran medida un clima subtropical. La ejecución del proyecto comenzó en diciembre (verano) y duró hasta marzo (verano-otoño). A lo largo de todo el ensayo, los animales mostraron un rendimiento consistente con las expectativas de su genotipo específico. Además, los animales se mantuvieron en buen estado de salud durante todo el período experimental, ya que no se observaron problemas de salud o enfermedades graves.
3.1 Rendimiento y grado de suciedad de las cáscaras de huevo
En la fase 1, los grupos BMA y BMA + PRO presentaron un 12% más de tasa postural en comparación con el CON (p < 0,001, Tabla 2). En la fase 2, todos los tratamientos suplementados tuvieron mayores tasas de puesta (21%) en comparación con CON (p < 0,001). En la fase 3, el grupo alimentado con BMA+PRO fue un 5% superior a CON (p < 0,001), mientras que los grupos BMA y PRO fueron intermedios en relación a CON y BMA+PRO. En el período total, todos los tratamientos tuvieron una tasa de puesta más alta (p < 0,001) en comparación con CON, en la que BMA + PRO tuvo la tasa de puesta más alta seguida de BMA y PRO.
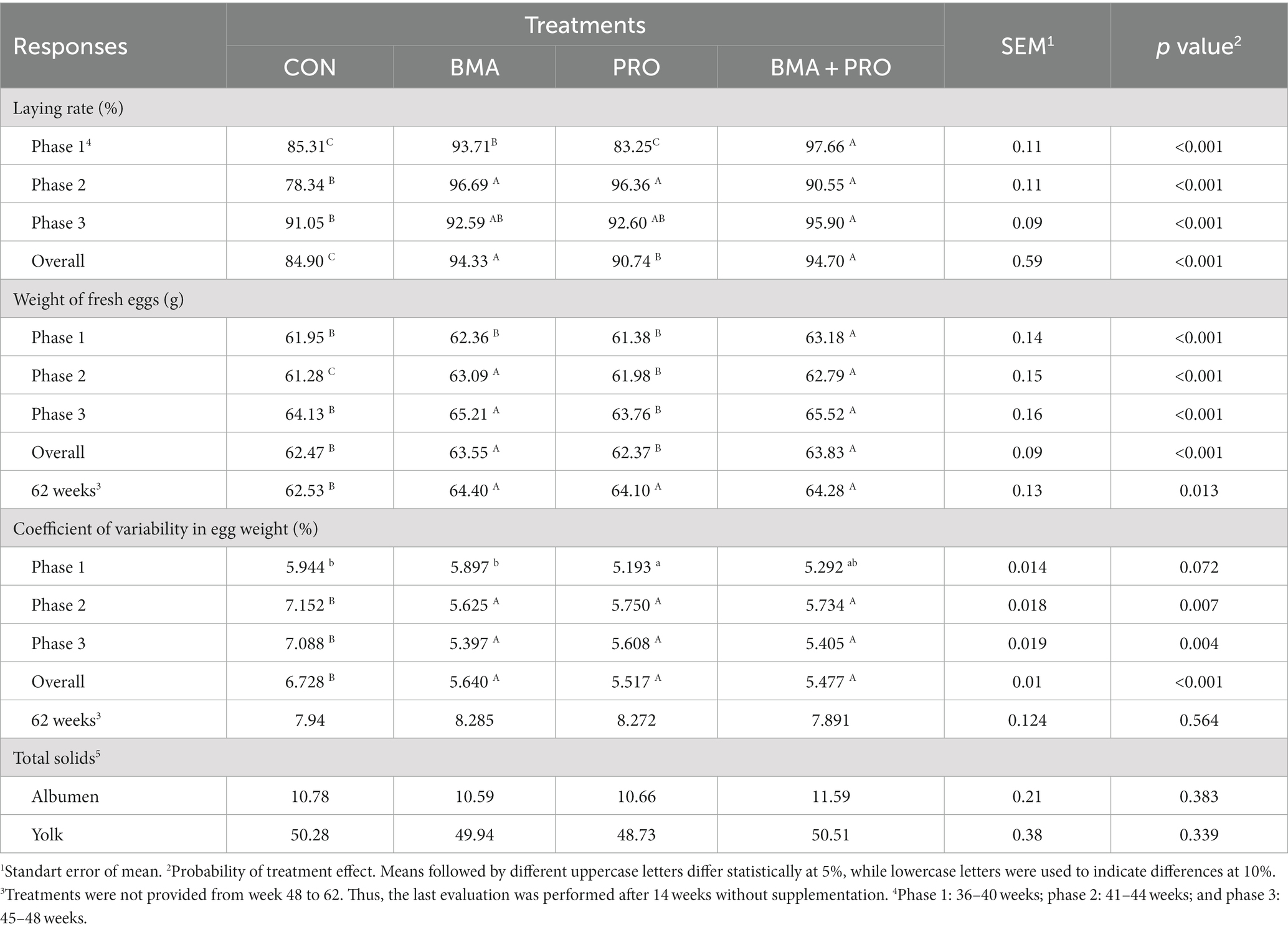 Tabla 2. Rendimiento de gallinas ponedoras alimentadas con dietas suplementadas con β-mananasa (BMA) y/o probióticos (PRO).
Tabla 2. Rendimiento de gallinas ponedoras alimentadas con dietas suplementadas con β-mananasa (BMA) y/o probióticos (PRO).
En cuanto al peso del huevo, el grupo BMA + PRO difirió del CON, con mayor peso del huevo (p < 0,001) en la fase 1. En la fase 2, todos los tratamientos difirieron (p < 0,001) de CON, siendo BMA y BMA + PRO similares entre sí. En la fase 3, los tratamientos PRO y BMA + PRO difirieron de CON (p < 0,001); sin embargo, el grupo PRO fue similar al control que también ocurre en el período general. En la semana 62, todos los tratamientos diferían (p = 0,013) de la CON.
Todos los tratamientos difirieron (p < 0,001) del CON en cuanto al coeficiente global de variabilidad del peso del huevo, observándose los valores más bajos en el grupo BMA + PRO. Se observó un efecto tendencial (p = 0,072) en la fase 1, con el menor coeficiente de variabilidad atribuido al tratamiento con PRO. En las fases 2 (p = 0,007) y 3 (p = 0,004) todos los tratamientos difirieron de CON, observándose valores más bajos en el grupo BMA. Sin embargo, en la semana 62 (después de la retirada del tratamiento), no se observaron diferencias significativas entre los grupos (p = 0,564).
En cuanto a las masas de huevos (Tabla 3), en las fases 1 y 3 (p < 0,001), los tratamientos BMA y BMA + PRO fueron diferentes al grupo CON, mostrando mayores masas de huevos. En la fase 2 y en general (p < 0,001), todos los tratamientos aumentaron las masas de huevos en comparación con el CON.
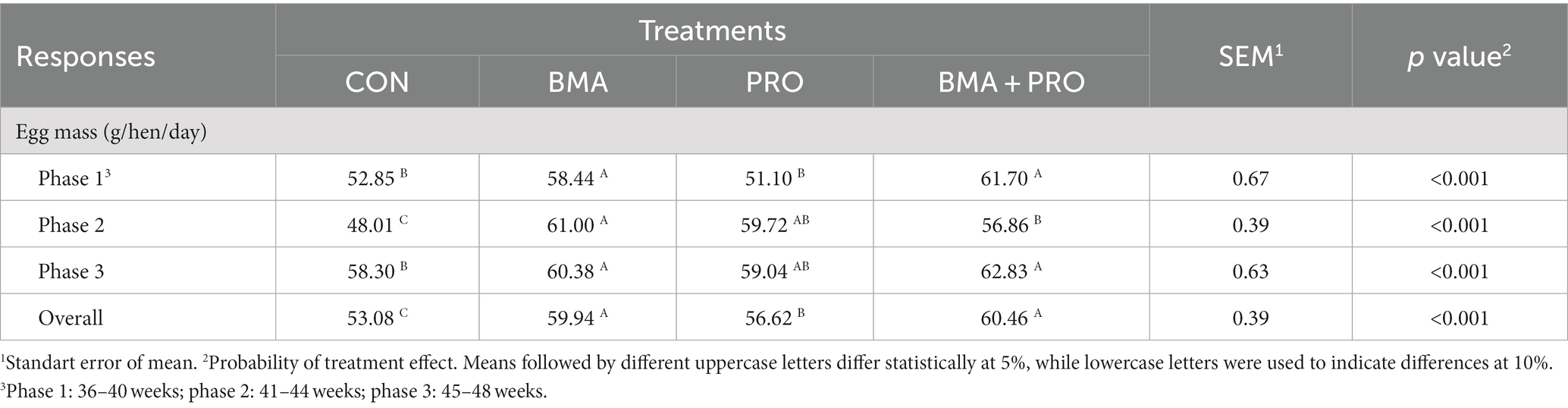 Tabla 3. Masas de huevos de gallinas ponedoras alimentadas con dietas suplementadas con β-mananasa (BMA) y/o probióticos (PRO).
Tabla 3. Masas de huevos de gallinas ponedoras alimentadas con dietas suplementadas con β-mananasa (BMA) y/o probióticos (PRO).
La ocurrencia de huevos limpios en las aves tratadas difirió de la CON en todas las fases del experimento (p < 0.05, Figura 1). Los grupos BMA, PRO y BMA+PRO fueron superiores a CON en todas las fases. En la semana 62 de producción, después de 14 semanas sin suplementación, la ocurrencia de huevos limpios aún difiere (p < 0,001) en los grupos BMA y BMA + PRO en comparación con CON.
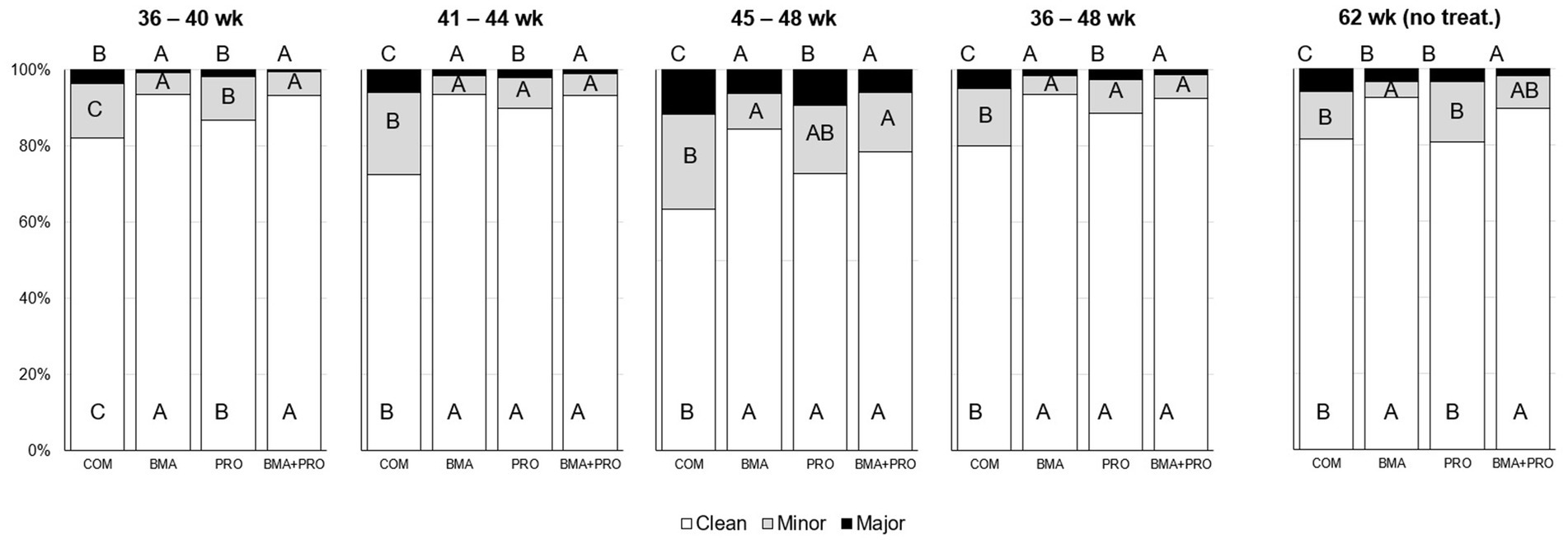 Figura 1. Presencia de huevos limpios o presencia menor/mayor de heces (%) en huevos de gallinas ponedoras alimentadas con β-mananasa y/o probióticos. Se realizaron comparaciones entre tratamientos en cada período. La probabilidad de efecto del tratamiento fue de p < 0,001 para todas las respuestas, excepto para el período de 45 a 48 semanas en el que todas las respuestas mostraron p < 0,05. Las diferentes letras mayúsculas difieren estadísticamente en un 5%. No se proporcionaron tratamientos desde la semana 48 hasta la 62. Por lo tanto, la última evaluación se realizó después de 14 semanas sin suplementación.
Figura 1. Presencia de huevos limpios o presencia menor/mayor de heces (%) en huevos de gallinas ponedoras alimentadas con β-mananasa y/o probióticos. Se realizaron comparaciones entre tratamientos en cada período. La probabilidad de efecto del tratamiento fue de p < 0,001 para todas las respuestas, excepto para el período de 45 a 48 semanas en el que todas las respuestas mostraron p < 0,05. Las diferentes letras mayúsculas difieren estadísticamente en un 5%. No se proporcionaron tratamientos desde la semana 48 hasta la 62. Por lo tanto, la última evaluación se realizó después de 14 semanas sin suplementación.
3.2 Bioquímica sérica
El ácido úrico difirió del CON (p < 0,001) en todos los tratamientos, observándose los valores más bajos en BMA + PRO, BMA y PRO, respectivamente (Tabla 4). El colesterol total y los triglicéridos difirieron del CON (p < 0,001), mostrándose valores menores en los tratamientos suplementados. Los niveles de glucosa sérica fueron mayores en las aves CON (p < 0,001) en relación con los tratamientos BMA y BMA + PRO. El tratamiento PRO no difirió del CON. La fosfatasa alcalina sérica fue mayor (p = 0,007) en los tratamientos PRO y BMA + PRO que en los CON. La BMA fue similar a la CON y otros tratamientos en este estudio. La alanina aminotransferasa fue mayor en CON (p = 0,005) que en los tratamientos PRO y BMA + PRO.
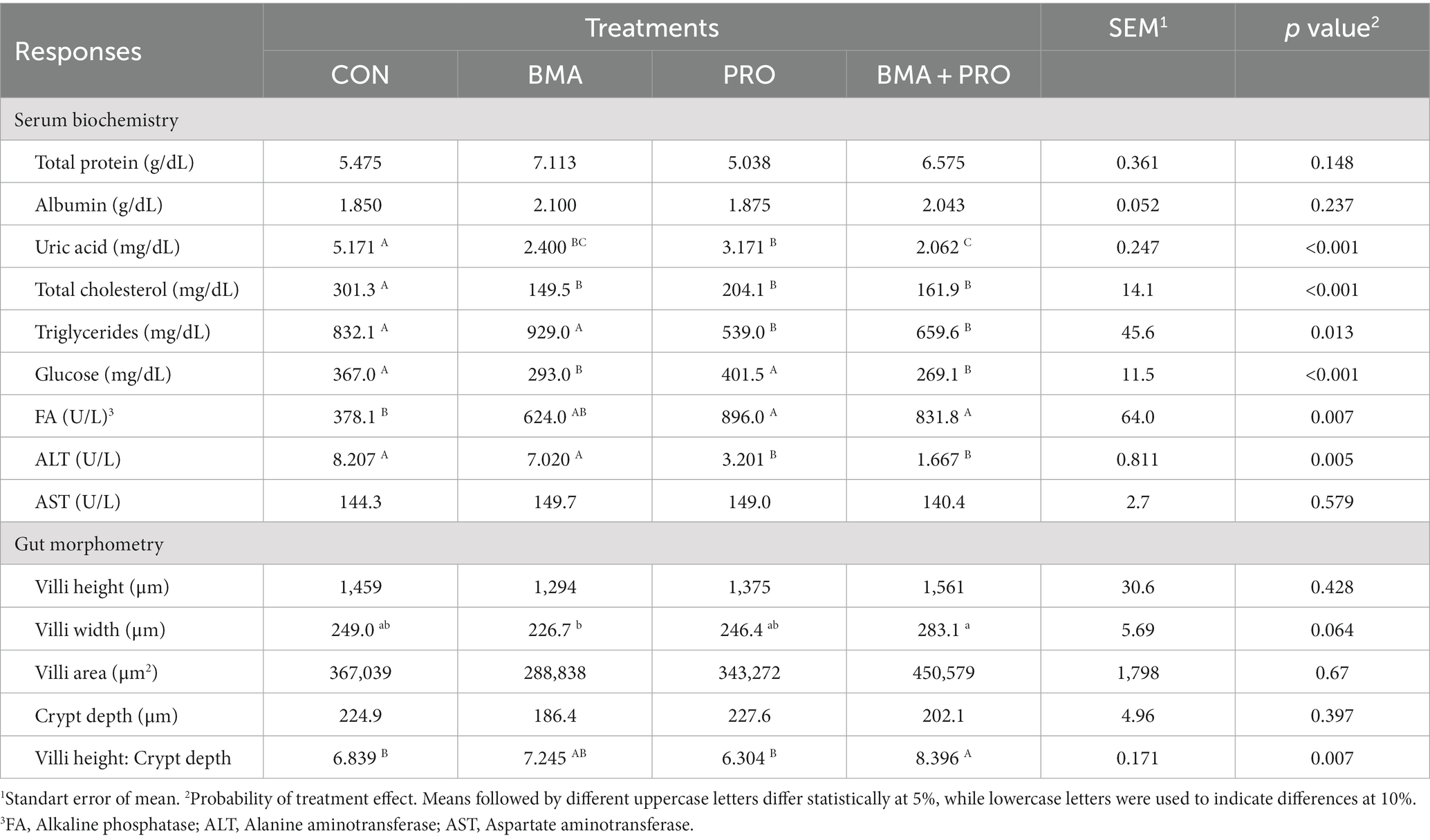 Tabla 4. Bioquímica sérica y morfometría intestinal de gallinas ponedoras alimentadas con β-mananasa y/o probióticos.
Tabla 4. Bioquímica sérica y morfometría intestinal de gallinas ponedoras alimentadas con β-mananasa y/o probióticos.
No se observaron diferencias significativas en la aspartato aminotransferasa (p = 0,579) y en la proteína total (p = 0,148). Tampoco hubo diferencias significativas en la albúmina sérica (p = 0,237).
3.3 Morfometría intestinal y análisis parasitológico
No se observaron diferencias en la altura de las vellosidades, el área de las vellosidades y la profundidad de las criptas entre los tratamientos. El ancho de las vellosidades tendió (p = 0,064) a ser menor en el tratamiento con BMA en comparación con el control, mientras que el tratamiento con BMA + CON tendió a ser superior al CON y el tratamiento PRO fue similar al CON. La relación entre la altura de las vellosidades y la profundidad de la cripta fue significativa (p = 0,007), observándose la mayor relación en el tratamiento BMA + PRO en comparación con el CON. El tratamiento PRO fue similar al CON y BMA similar al CON y BMA + PRO.
No se encontraron parásitos ni ooquistes en las muestras de excreción fresca, incluido el tratamiento de control. Esta condición no permitió evaluar el eventual efecto de los tratamientos sobre los desafíos parasitarios.
3.4 Calidad de los huevos frescos
El grupo BMA mostró mayor gravedad específica (p < 0,001) en comparación con el CON (Tabla 5), con valores más altos. Además, se observaron pesos de cáscara más altos (p = 0,009) en los grupos BMA y BMA + PRO en comparación con CON.
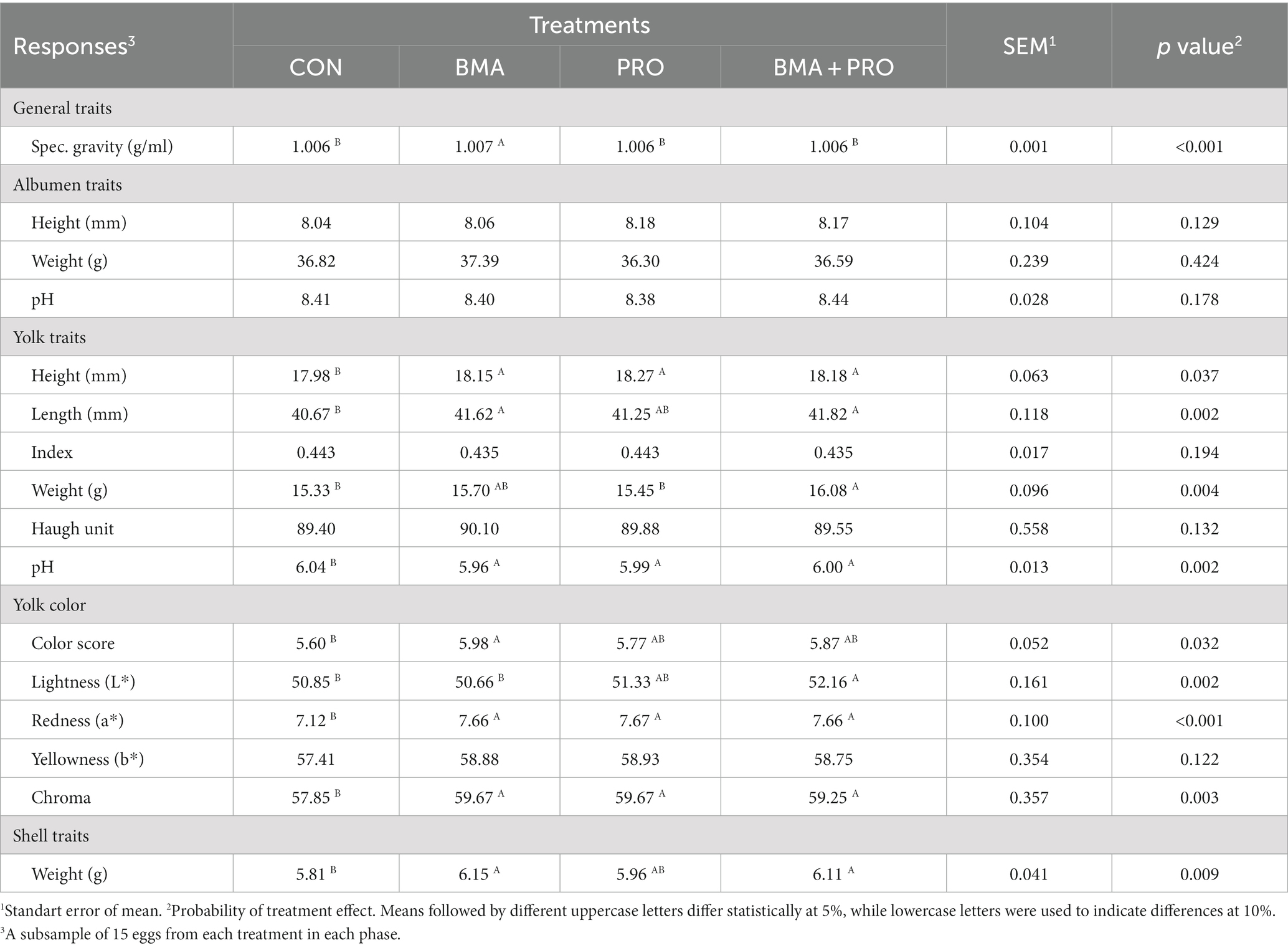 Tabla 5. Calidad de los huevos frescos de gallinas ponedoras alimentadas con dietas suplementadas con β-mananasa (BMA) y/o probióticos (PRO).
Tabla 5. Calidad de los huevos frescos de gallinas ponedoras alimentadas con dietas suplementadas con β-mananasa (BMA) y/o probióticos (PRO).
La altura de la yema mostró diferencias significativas en los grupos BMA, PRO y BMA + PRO en comparación con CON (p = 0,037), todos con valores más altos. El ancho de la yema también fue mayor (p = 0,002) en los grupos BMA y BMA + PRO en comparación con CON. El peso de la yema fue mayor en el grupo BMA + PRO (p = 0,004) en comparación con el grupo CON. El pH de la yema, por su parte, difirió del CON en todos los grupos (p = 0,002), mostrando valores de pH más bajos.
El grupo BMA mostró una mayor puntuación de color en la evaluación colorimétrica de Roche en comparación con el CON (p = 0,032). En cuanto a los valores de luminosidad (color L*), el grupo BMA+PRO fue superior al grupo CON (p = 0,002). Tales hallazgos indican menor luminosidad, es decir, eran opacos ya que transmiten menos luz. Se observaron valores de mayor intensidad de rojo (color A*) y croma en todos los grupos suplementados que en CON (p < 0,001; Figura 2).
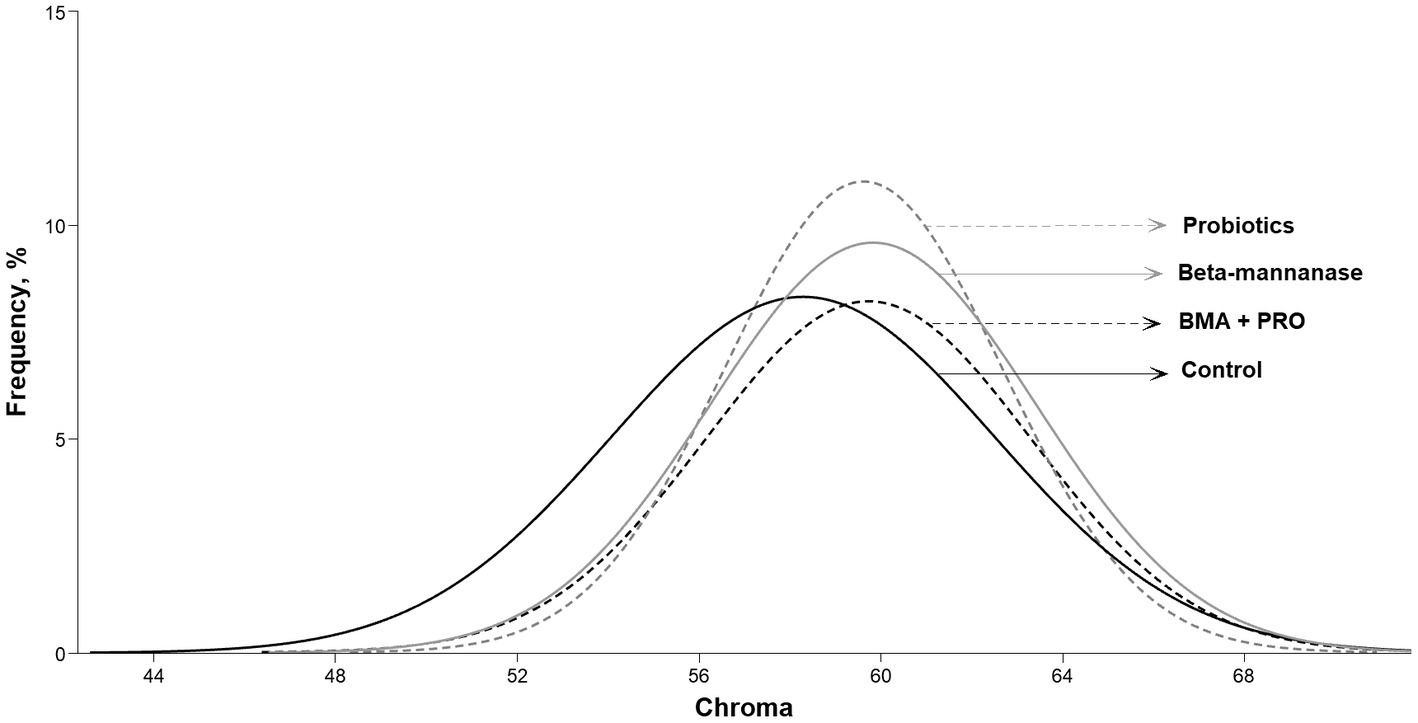 Figura 2. Frecuencia de diferentes índices de croma en yemas de huevo de gallinas ponedoras alimentadas con β-mananasa y/o probióticos.
Figura 2. Frecuencia de diferentes índices de croma en yemas de huevo de gallinas ponedoras alimentadas con β-mananasa y/o probióticos.
4 Discusión
4.1 Rendimiento y grado de suciedad de las cáscaras de huevo
En el presente estudio, se observó una mejora en las tasas generales de puesta en todos los tratamientos en comparación con el grupo de control. Este hallazgo se alinea con investigaciones previas realizadas por Zhan et al. (18), quienes informaron un aumento significativo en las tasas de puesta en aves cuando se alimentan con probióticos, particularmente cuando se administran a una dosis de 5 × 104 ufc/g de Clostridium butiricum. De manera similar, Ribeiro et al. (19) documentaron un aumento sustancial en las tasas de puesta con una dosis de 8 × 105 ufc/g de Bacillus subtilis, mientras que Saleh et al. (20) observaron una mejoría al utilizar 0.05% de Aspergillus awamori. Los probióticos han sido reconocidos por su impacto positivo en la producción de huevos, lo que se atribuye a varios mecanismos beneficiosos. Estos incluyen una mayor absorción de nutrientes (19), una mejor función inmunológica, una reducción del estrés en las aves y la promoción de la salud intestinal (18), que son efectos que pueden ayudar a explicar los resultados encontrados en este estudio. En este contexto, es importante mencionar que el presente estudio se desarrolló en condiciones de granja comercial. Dicha condición generalmente indica que los animales están expuestos a ciertos desafíos (bioseguridad, medio ambiente, número de animales, entre otros) que serían limitados en condiciones experimentales a pequeña escala. Ambos tipos de estudios son relevantes y pueden proporcionar datos útiles. Sin embargo, esta característica debe tenerse en cuenta a la hora de interpretar los resultados.
En referencia a la β-mananasa, nuestros hallazgos actuales muestran una mayor tasa de puesta en comparación con el tratamiento control. Estos resultados concuerdan con los resultados de Zheng et al. (21), quienes reportaron un aumento significativo en la producción de huevos entre las gallinas ponedoras que fueron suplementadas con β-mananasa en dietas bajas en energía. En particular, los valores fueron similares a los de las dietas altas en energía sin enzimas. Datos similares también fueron encontrados por Wu et al. (22). Estos hallazgos pueden estar asociados con el hecho de que la β-mananasa, al evitar la respuesta inmune desencadenada por la β-mananasa, redirige la energía y los nutrientes para el rendimiento del ave (23).
El estudio actual observó un aumento significativo en el peso del huevo, alineándose con investigaciones previas de Khan et al. (24), Alaquil et al. (25) y Mikulski et al. (26) en dietas suplementadas con probióticos. Por otra parte, en lo que respecta a la β-mananasa, nuestros hallazgos concuerdan con los resultados de Ryu et al. (27), al utilizar 0,8 g de β-mananasa/kg, se observó un aumento en el peso del huevo en comparación con el control. En cuanto al coeficiente de variabilidad del peso del huevo, se observó una producción estable y predecible, lo que facilita los procesos y aumenta la rentabilidad al disminuir el número de huevos desclasificados.
Con respecto a las masas de huevos, nuestro estudio reveló un aumento en las masas de huevos en todos los tratamientos en comparación con el control. Estos resultados concuerdan con Ryu et al. (27) que observaron mayores masas de huevos utilizando β-mananasa. Este efecto puede atribuirse a la mejora de la digestión y la utilización de los componentes de la dieta, lo que conduce a una mayor absorción y utilización de nutrientes para la producción de huevos. En cuanto a los probióticos, Saleh et al. (20), Alaquil et al. (25) y Ribeiro et al. (19) también observaron mayores masas de huevos en comparación con el grupo control. Se sabe que los probióticos fomentan una microbiota intestinal equilibrada, lo que puede influir positivamente en el metabolismo y la absorción de nutrientes, contribuyendo posteriormente a mejorar la producción de masa de huevos.
La mayor ocurrencia de huevos limpios en las aves tratadas observada en este estudio difirió del control en todas las fases del experimento. El uso de β-mananasa puede explicarse por la probable disminución de la viscosidad de las heces. Los polisacáridos solubles sin almidón aumentan la viscosidad de la digesta al aumentar la retención de agua, lo que perjudica la difusión y el transporte de nutrientes. Daskiran et al. (28) demostraron que las dietas que utilizaban β-mananasa reducían significativamente el agua de producción fecal total en pollos de engorde. Asimismo, Mehri et al. (29) demostraron que la viscosidad de la digesta del yeyuno de pollos de engorde disminuyó en dietas con la enzima. Los huevos más limpios observados en el tratamiento con probióticos pueden atribuirse probablemente a la mayor estabilidad de la microbiota intestinal y a un menor recuento de bacterias oportunistas. Higgins et al. (30) y Deng et al. (31) observaron una disminución de las colonias de Salmonella sp. en aves suplementadas con probióticos. Aalaei et al. (32) encontraron que la adición de probióticos multicepa redujo la presencia de E. coli en pollos de engorde, y con ello, fue posible reducir la diarrea en las aves. Por lo tanto, en el presente estudio, la reducción de la diarrea podría ser la causa de la disminución de los huevos sucios del tratamiento PRO. Hasta donde sabemos, este es el primer estudio que evalúa la presencia de huevos sucios en gallinas ponedoras alimentadas con β-mananasa sola o combinada con probióticos.
4.2 Bioquímica sérica
El ácido úrico es el principal producto del metabolismo del nitrógeno en las aves, que se sintetiza en el hígado y los riñones. Los trastornos de la función renal pueden aumentar la concentración de ácido úrico en el suero y el plasma de las aves (33), así como las temperaturas elevadas (34). Los niveles bajos de ácido úrico también indican un menor recambio proteico (35), es decir, menores pérdidas endógenas de nitrógeno y amoníaco. En el presente estudio, que se llevó a cabo en verano, las aves se enfrentaron a altas temperaturas, y aun así los valores de ácido úrico encontrados en la sangre fueron menores en todos los tratamientos en comparación con el control, lo que puede indicar una mejora en la salud de las aves y una mejor eficiencia en la utilización de proteínas debido a los aditivos.
Además, en relación con el metabolismo de las proteínas, el ácido úrico puede dilucidar los fenómenos observados de aumento del peso de la albúmina y la masa de los huevos en este estudio, potencialmente atribuibles a una mayor deposición de proteínas dentro de estos huevos.
Los niveles de colesterol total y triglicéridos fueron significativamente más bajos en los grupos que recibieron probióticos y los aditivos combinados (BMA + PRO) en comparación con el grupo control. Estos hallazgos son consistentes con investigaciones previas (20, 36, 37) en aves que fueron suplementadas con probióticos. Además, las aves que fueron alimentadas con β-mananasa exhibieron niveles más bajos de colesterol total, lo que se alinea con los resultados reportados por Karimi y Shokrollari (38), quienes observaron una disminución en los niveles de colesterol LDL. Los niveles séricos de colesterol y triglicéridos reflejan el metabolismo de los lípidos. Saleh et al. (39) informaron que uno de los posibles mecanismos de reducción del colesterol por probióticos ocurre a través de la producción de HMG-CoA reductasa (3-hidroxi-3-metil-glutaril-CoA reductasa), que reduce la deposición de grasa abdominal al influir en la actividad de la enzima lipasa sensible a las hormonas y la enzima malato deshidrogenasa en los tejidos adiposos (40). Además, uno de los supuestos mecanismos de los probióticos ocurre a través de la reducción de la síntesis de ácidos biliares hepáticos (41). Las bacterias del ácido láctico, como las que se encuentran en el producto probado, tienen la capacidad de reducir el colesterol en el torrente sanguíneo (42). La disminución del colesterol por la β-mananasa puede explicarse por el efecto hipolipidémico de la enzima, que reduce la absorción de lípidos (38, 43).
En cuanto a la glucosa sérica, nuestros hallazgos concuerdan con Tang et al. (44). La disminución de la glucosa por β-mananasa puede explicarse por el hecho de que esta enzima estimula la secreción de insulina (45), lo que puede estimular el comportamiento de ingesta de alimento y, en consecuencia, estar relacionado con el aumento de la producción de huevos.
El aumento de la fosfatasa alcalina sérica concuerda con los hallazgos de Yalcin et al. (37). Y la alanina aminotransferasa superior también está de acuerdo con Saleh et al. (39) y Tang et al. (44). La concentración sérica de enzimas hepáticas como la fosfatasa alcalina y la alanina aminotransferasa puede proporcionar información sobre el daño a tejidos y órganos (46). La fosfatasa alcalina también se asocia con el metabolismo del calcio y el fósforo y con la participación en actividades osteoblásticas y condrogénicas. Por lo tanto, el aumento de esta enzima se asocia con el crecimiento óseo, la consolidación de fracturas y la fase de preovulación y calcificación medular en pollos (33). Además, los cambios en los niveles de fosfatasa alcalina pueden indicar que el hueso medular promueve el calcio durante la formación de cáscaras de huevo y almacena calcio cuando no hay óvulo en el útero (46). En relación con la alanina aminotransferasa en aves, se cree que puede estar elevada debido al daño a múltiples tejidos, lo que dificulta su interpretación (47). En el presente estudio, los resultados observaron que las aves alimentadas con los grupos PRO y BMA + PRO presentaron valores más altos de fosfatasa alcalina sérica, lo que indica una mejor salud para estas aves. Los valores más bajos de alanina aminotransferasa observados en este estudio pueden indicar un metabolismo más eficiente de estas aves debido a un menor daño hepático, lo que puede explicar los resultados positivos en el rendimiento.
4.3 Morfometría intestinal y análisis parasitológico
Las mediciones de altura y profundidad de las criptas se utilizan a menudo para evaluar la integridad intestinal. La altura de las vellosidades indica una mayor área para la absorción de nutrientes y una cripta más profunda indica que hay una mayor renovación del tejido (48, 49). En el presente estudio, el grupo tratado con BMA+PRO mostró una mayor relación altura vellosas: profundidad de criptas, demostrando una mejoría en la salud intestinal.
Estudios previos han mostrado diferencias significativas en la relación entre la altura de las vellosidades y la profundidad de las criptas (36) en el intestino de gallinas ponedoras alimentadas con probióticos. A pesar de que no existen estudios en relación a la β-mananasa en gallinas ponedoras, en base a estos resultados, se cree que este aditivo puede beneficiar la salud intestinal de las aves. La mayor relación vellosidades:criptas en estos grupos, ya que se asocia a una mayor superficie para la absorción de nutrientes, puede explicar el mejor comportamiento de estas aves, especialmente en relación con el peso y la masa de huevos.
4.4 Calidad de los huevos frescos
Es crucial reconocer que la composición de la cáscara del huevo consiste principalmente en carbonato de calcio, junto con carbonato de magnesio y fosfato de calcio, entre otros componentes. El equilibrio entre los iones calcio y fósforo juega un papel vital en la formación de la cáscara del huevo (50). El peso específico es un indicador de la proporción de la cáscara en relación con otros componentes del huevo. Está estrechamente asociado con el grosor de la cáscara y, en consecuencia, con la deposición de carbonato de calcio. La evaluación de la gravedad específica proporciona información sobre la calidad de la carcasa. Además, el peso de la cáscara se puede utilizar para respaldar los hallazgos obtenidos de las mediciones de gravedad específica y evaluar el metabolismo del calcio.
En nuestro estudio, observamos una mayor gravedad específica y un aumento del peso de la cáscara de huevo en el grupo BMA en comparación con el CON. Estos hallazgos sugieren una mayor calidad de la cáscara y una menor probabilidad de rotura durante la manipulación (51, 52). La mejora de la gravedad específica y el aumento del peso de la cáscara del huevo indican una mayor integridad y resistencia de la cáscara, que son atributos deseables en términos de calidad y comerciabilidad del huevo.
El peso de la yema y la albúmina muestran una correlación positiva con el peso del huevo. Los huevos con pesos más altos tienden a tener mayores masas de yema y albúmina en comparación con aquellos con pesos más bajos. El peso del huevo está influenciado por varios factores, como la heredabilidad, la edad y el peso de las aves (53). Además, el peso del huevo influye fuertemente en los requerimientos de proteínas en la dieta (54). En este estudio, observamos niveles más bajos de ácido úrico en aves alimentadas con el grupo BMA en comparación con el grupo CON, lo que sugiere una reducción del recambio proteico. Se sabe que los β-mananos, que se encuentran en la dieta, reducen la viscosidad e inhiben la acción enzimática (55). La adición de β-mananasa en el grupo BMA facilitó la actividad enzimática al descomponer los β-mananos, lo que podría conducir a un aumento de la absorción de proteínas. Este mecanismo puede explicar el mayor peso de la yema observado en el grupo BMA. Otra hipótesis está asociada con el efecto reductor de la viscosidad de la β-mananasa (56). Esta actividad enzimática altera la estructura de las micelas (57, 58), que son compuestos lipídicos de gran importancia ya que se depositan en la yema. La disminución de la viscosidad causada por la β-mananasa podría mejorar la formación o función de las micelas, contribuyendo a un aumento del peso de la yema.
Los resultados relacionados con el color de la yema demostraron una menor luminosidad en el grupo BMA+PRO en comparación con el CON, lo que indica un aspecto más opaco y una menor transmisión de luz. Además, los grupos PRO y BMA exhibieron colores amarillentos y rojizos deseables, que se consideran atractivos para los consumidores (59). La pigmentación de la yema se produce a través de la absorción de pigmentos carotenoides presentes en la dieta del ave (60). El maíz, por ejemplo, contiene carotenoides como la xantofila, la luteína y la zeaxantina (61). Estos compuestos lipofílicos e insaturados se acumulan en la yema, que tiene la mayor concentración de grasa en el huevo (62). Una hipótesis para los cambios de color observados es que la β-mananasa puede mejorar la absorción de nutrientes y/o aumentar la producción de micelas, que son responsables de transportar carotenoides y acumularlos en la yema. Sin embargo, se necesitan más estudios para validar esta hipótesis.
Cabe mencionar que parámetros como el pH, la altura y el ancho de la yema son indicadores de la frescura del huevo (63, 64). En este ensayo, los tratamientos dieron como resultado una disminución de los valores de pH y un aumento de la altura y el ancho de la yema, lo que indica una mejor calidad y frescura del huevo.
5 Conclusión
El presente estudio proporciona evidencia de que la suplementación con β-mananasa, probióticos y su combinación en el alimento puede mejorar significativamente el rendimiento (tasa de puesta y peso de los huevos frescos) y la salud intestinal (relación entre la altura de las vellosidades y la profundidad de la cripta). Además, la suplementación de estos aditivos en la alimentación de las aves de corral conduce a la aparición de huevos más homogéneos en términos de peso, más limpios y con una mejor calidad.
Contribuciones de los autores
CC, IA y MK: conceptualización, metodología, validación y análisis formal. CC, GG, NC, TB, RiM y GP: investigación. IA y MK: recursos, visualización, administración de proyectos y captación de fondos. CC e IA: curación de datos. CC: redacción: preparación del borrador original. CC, IA, GG, BB y MK: redacción, revisión y edición. IA, RaM y MK: supervisión. Todos los autores contribuyeron al artículo y aprobaron la versión enviada.
Reconocimientos
Agradecemos al Consejo Nacional de Desarrollo Científico y Tecnológico (CNPq) y a la Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) por financiar parcialmente este estudio. También agradecemos a Granja Petry por la donación de los huevos y a Elanco por la donación de los productos.
Conflicto de intereses
MK fue empleado de Elanco Animal Health, São Paulo, Brasil.
El resto de los autores declaran que la investigación se llevó a cabo en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un potencial conflicto de intereses.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio se incluyen en el artículo/material complementario, las consultas posteriores pueden dirigirse al autor correspondiente.
Declaración ética
El estudio en animales fue aprobado por el protocolo experimental descrito y aprobado por el Comité Institucional de Ética en el Uso de Animales (CEUA/UFRGS) bajo el protocolo número 39783. El estudio se llevó a cabo de acuerdo con la legislación local y los requisitos institucionales.
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o afirmación que pueda ser hecha por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Organización Mundial de la Salud (2020). No hay tiempo que esperar: Asegurar el futuro de las infecciones por resistencia a los medicamentos. https://www.who.int/publications/i/item/no-time-to-wait-securing-the-future-from-drug-resistant-infections (consultado en septiembre de 2022).
2. Fuller, R. Probióticos en el hombre y los animales. J Appl Bacteriol. (1989) 66:365–78. doi: 10.1111/j.1365-2672.1989.tb05105.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
3. Callaway, TR, Edrington, TS, Anderson, RC, Harvey, RB, Genovese, KJ, Kennedy, CN, et al. Probióticos, prebióticos y exclusión competitiva para la profilaxis frente a enfermedades bacterianas. Anim Health Res Rev. (2008) 9:217–25. doi: 10.1017/s1466252308001540
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Duggan, C, Gannon, J y Walker, WA. Nutrientes protectores y alimentos funcionales para el tracto gastrointestinal. ACJN. (2002) 75:789–808. doi: 10.1093/ajcn/75.5.789
5. Stahl, CH, Callaway, TR, Lincoln, LM, Lonergan, SM y Genovese, KJ. Actividades inhibitorias de las colicinas contra las cepas de Escherichia coli responsables de la diarrea post-destete y la enfermedad edema en cerdos. Agentes antimicrobianos quimiotera. (2004) 48:3119–21. doi: 10.1128/FAAC.48.8.3119-3121.2004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
6. Wu, X, Vallance, BA, Boyer, L, Bergstrom, KSB, Walker, J, Madsen, K, et al. Saccharomyces boulardii mejora la colitis inducida por Citrobacter rodentium a través de acciones sobre los factores de virulencia bacteriana. Am J Physiol Gastrointest Liver Physiol. (2008) 294:G295-306. doi: 10.1152/ajpgi.00173.2007
Resumen de PubMed | Texto completo de CrossRef | Google Académico
7. Saeed, M, Ayasan, T, Alagawany, M, El-Hack, M, Abdel-Latif, M, y Patra, AK. El papel de la ß-mananasa (Hemicela) en la mejora de la productividad, la salud y el medio ambiente de las aves de corral. Braz J Poult Sci. (2019) 21:1–8. doi: 10.1590/1806-9061-2019-1001
8. Balamuralikrishnan, B, Lee, JH, y Kim, IH. Efectos de la suplementación dietética con β-mananasa de harina de soja en el rendimiento de los cerdos destetados. Anim Nutr Feed Technol. (2018) 18:13–23. doi: 10.2527/jam2016-0921
9. Balamuralikrishnan, B, Ingale, SL, Park, JH, Rathi, PC, Shanmugam, S, y Kim, IH. La inclusión de β-mananasa en la dieta mejora el rendimiento y la digestibilidad ileal y reduce la viscosidad de la digesta ileal de los pollos de engorde alimentados con una dieta basada en harina de maíz y soja. Poult Sci. (2018) 97:3097–101. doi: 10.3382/ps/pey157
Resumen de PubMed | Texto completo de CrossRef | Google Académico
10. Mohammadigheisar, M, Shouldice, VL, Balasubramanian, B y Kim, I. Efecto de la suplementación dietética de β-mananasa sobre el rendimiento del crecimiento, las características de la canal, la microflora de excrementos, los componentes sanguíneos y la digestibilidad ileal de nutrientes en pollos de engorde. Anim Biosci. (2021) 34:1342–9. doi: 10.5713/ab.20.0355
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Balamuralikrishnan, B, Lee, SI, y Kim, IH. Inclusión dietética de diferentes probióticos complejos multicepa; Efectos sobre el rendimiento en pollos de engorde. Hno. Poult Sci. (2017) 58:83–6. doi: 10.1080/00071668.2016.1257112
Resumen de PubMed | Texto completo de CrossRef | Google Académico
12. Jackson, ME. Mananasa, alfa-galactosidada y pectinasa En: MR Bedford y GG Partridge, editores. Enzimas en la nutrición de animales de granja. Wallingford, Inglaterra: CABI Publishing (2001). 54–84.
13. Hsiao, HY, Anderson, DM y Dale, NM. Niveles de β-manano en harina de soja. Poult Sci. (2006) 85:1430–2. doi: 10.1093/ps/85.8.1430
Resumen de PubMed | Texto completo de CrossRef | Google Académico
14. Hy-line (2020). Guía de gestión para ponedoras comerciales W-36. https://www.hyline.com/filesimages/Hy-Line-Products/Hy-Line-Product-PDFs/W-36/36%20COM%20ENG.pdf (consultado en septiembre de 2022).
16. Brasil (2013). Resolução Normativa n° 13, de 20 de dezembro de 2013. Diretrizes para a prática de eutanásia do CONCEA: MCTI. Ministério da Ciência, Tecnologia e Inovação. 2013, Brasilia, Distrito Federal, Brasil.
17. Haugh, RR. La unidad de Haugh para medir la calidad de los huevos. Aves de corral de huevo de EE. UU. Magaz. (1937) 43:552–5.
18. Zhan, HQ, Dong, XY, Li, LL, Zheng, YX, Gong, YJ y Zou, XT. Efectos de la suplementación dietética con Clostridium butyricum sobre el rendimiento de las ponedoras, la calidad del huevo, los parámetros séricos y la microflora cecal de gallinas ponedoras en la fase tardía de producción. Poult Sci. (2019) 98:896–903. doi: 10.3382/ps/pey436
Resumen de PubMed | Texto completo de CrossRef | Google Académico
19. Ribeiro, RV Jr, Albino, LFT, Rostagno, HS, Barreto, SLT, Hannas, MI, Harrington, D, et al. Efectos de la suplementación dietética de los niveles de Bacillus subtilis sobre el rendimiento, la calidad del huevo y la humedad de los excrementos de las ponedoras. Anim Feed Sci Technol. (2014) 195:142–6. doi: 10.1016/j.anifeedsci.2014.06.001
20. Saleh, AA, Gálik, B, Arpálová, H, Capcarová, M, Kalafová, A, L·imko, M, et al. Efecto sinérgico de la alimentación con aspergillus awamori y bacterias lácticas sobre el rendimiento, las características del huevo, el colesterol de la yema de huevo y el perfil de ácidos grasos en gallinas ponedoras. Ital J Anim Sci. (2016) 16:132–9. doi: 10.1080/1828051X.2016.1269300
21. Zheng, L, Cho, SH, Kang, CW, Lee, KW, Kim, KE y An, BK. Efectos de la β-mananasa en el rendimiento de la producción de huevos, la calidad del huevo, la microbiota intestinal, la viscosidad y la concentración de amoníaco en gallinas ponedoras. Braz J Poult Sci. (2020) 22:1–8. doi: 10.1590/1806-9061-2019-1180
22. Wu, G, Bryant, MM, Voitle, RA, y Roland, DA. Efectos de la β-mananasa en dietas de maíz-soja en leghorns comerciales en gallinas de segundo ciclo. Poult Sci. (2005) 84:894–7. doi: 10.1093/ps/84.6.894
Resumen de PubMed | Texto completo de CrossRef | Google Académico
23. Klasing, KC. Nutrición y sistema inmunológico. Hno. Poult Sci. (2007) 48:525–37. doi: 10.1080/00071660701671336
24. Khan, SH, Atif, M, Mukhtar, N, Rehman, A y Fareed, G. Efectos de la suplementación con probióticos multienzimáticos y multiespecies sobre el rendimiento de la producción, la calidad de los huevos, el nivel de colesterol y el sistema inmunológico en gallinas ponedoras. J Appl Anim Res. (2011) 39:386–98. doi: 10.1080/09712119.2011.621538
25. Alaqil, AA, Abbas, AO, El-Beltagi, HS, El-Atty, HKA, Mehaisen, GMK y Moustafa, ES. La suplementación dietética del probiótico Lactobacillus acidophilus modula los niveles de colesterol, la respuesta inmunitaria y el rendimiento productivo de las gallinas ponedoras. Animales. (2020) 10:1–12. doi: 10.3390/ani10091588
26. Mikulski, D, Jankowski, J, Mikulska, M y Demey, V. Efectos de la suplementación con probióticos dietéticos (Pediococcus acidilactici) sobre el rendimiento productivo, la calidad del huevo y la composición corporal en gallinas ponedoras alimentadas con dietas que varían en densidad energética. Poult Sci. (2020) 99:2275–85. doi: 10.1016/j.psj.2019.11.046
Resumen de PubMed | Texto completo de CrossRef | Google Académico
27. Ryu, MH, Hosseindoust, A, Kim, JS, Choi, YH, Lee, SH, Kim, MJ, et al. La β-Mannanase derivada de Bacillus Subtilis WL-7 mejora el rendimiento de las gallinas ponedoras comerciales alimentadas con dietas bajas o altas a base de Mannan. J Poult Sci. (2017) 54:212–7. doi: 10.2141/jpsa.0160021
Resumen de PubMed | Texto completo de CrossRef | Google Académico
28. Daskiran, M, Teeter, RG, Fodge, D, y Hsiao, HY. Una evaluación de los efectos de la endo-beta-D-mananasa (Hemicell) sobre el rendimiento de los pollos de engorde y el uso de energía en dietas que varían en el contenido de B-manano. Poult Sci. (2004) 83:662–8. doi: 10.1093/ps/83.4.662
Resumen de PubMed | Texto completo de CrossRef | Google Académico
29. Mehri, M, Adibmoradi, M, Samie, A y Shivazad, M. Efectos de la b-mananasa en el rendimiento de los pollos de engorde, la morfología intestinal y el sistema inmunológico. Afr J Biotechnol. (2010) 9:6221–8.
30. Higgins, SE, Wolfeden, AD, Téllez, G, Hargis, BM, y Porter, TE. Perfil transcripcional de la expresión génica cecal en pollitos neonatos desafiados por probióticos y Salmonella. Poult Sci. (2011) 90:901–13. doi: 10.3382/ps.2010-00907
Resumen de PubMed | Texto completo de CrossRef | Google Académico
31. Deng, Z, Han, D, Wang, Y, Wang, Q, Yan, X, Wang, S, et al. Lactobacillus casei protege la mucosa intestinal del daño en los pollitos causado por Salmonella pullorum mediante la regulación de la inmunidad y la vía de señalización Wnt y el mantenimiento de la abundancia de microbiota intestinal. Poult Sci. (2021) 100:101283–14. doi: 10.1016/j.psj.2021.101283
Resumen de PubMed | Texto completo de CrossRef | Google Académico
32. Aalaei, M, Khatibjoo, A, Zaghari, M, Taherpour, K, Gharaei, MA y Soltani, M. Comparación de los efectos de los probióticos de una o varias cepas en el rendimiento de las reproductoras de pollos de engorde, la producción de huevos, la calidad de los huevos y la incubabilidad. Hno. Poult Sci. (2018) 59:531–8. doi: 10.1080/00071668.2018.1496400
Resumen de PubMed | Texto completo de CrossRef | Google Académico
33. Campbell, TW. Bioquímica Clínica das Aves En: MA Thrall, G Weiser, RW Allison y TW Campell, editores. Hematología y Bioquímica Clínica Veterinária. São Paulo, Brasil: Roca (2014). 1233–66.
34. Qaid, MM, y Al-Garadi, MA. Metabolismo de proteínas y aminoácidos en aves de corral durante y después del estrés por calor: una revisión. Animales. (2021) 11:1167. doi: 10.3390/ani11041167
Resumen de PubMed | Texto completo de CrossRef | Google Académico
35. Ran, J, Ma, J, Liu, Y, Tan, R, Liu, H y Lao, G. La dieta baja en proteínas inhibe la síntesis de ácido úrico y atenúa el daño renal en ratas diabéticas inducidas por estreptozotocina. J Diabetes Res. (2014) 2014:1–10. doi: 10.1155/2014/287536
Resumen de PubMed | Texto completo de CrossRef | Google Académico
36. Song, D, Wang, YW, Lu, ZX, Wang, WW, Miao, HJ, Zhou, H, et al. Efectos de la suplementación dietética de Enterococcus fecalis microencapsulado y el extracto de semilla de Camellia oleifera sobre el rendimiento de la ponedora, la calidad del huevo, los parámetros bioquímicos séricos y la diversidad de la microflora cecal en gallinas ponedoras. Poult Sci. (2019) 98:2880–7. doi: 10.3382/ps/pez033
Resumen de PubMed | Texto completo de CrossRef | Google Académico
37. Yalçin, S, Yalçin, S, Uzunoglu, K, Duyum, HM y Eltan, Ö. Efectos del autolisado de levadura en la dieta (Saccharomyces cerevisiae) y la semilla de comino negro (Nigella sativa L.) sobre el rendimiento, las características del huevo, algunas características sanguíneas y la producción de anticuerpos de gallinas ponedoras. Livest Sci. (2010) 90:1695–701. doi: 10.1002/jsfa.4004
38. Karimi, K, y Shokrollah, B. Respuestas lipidémicas de pollos de engorde machos a dietas basadas en harina de trigo y soja suplementadas con enzimas con varios niveles de energía metabolizable. Pak J Biol Sci. (2013) 16:1295–302. doi: 10.3923/pjbs.2013.1295.1302
Resumen de PubMed | Texto completo de CrossRef | Google Académico
39. Saleh, AA, Hayashi, K y Ohtsuka, A. Efecto sinérgico de la alimentación con aspergillus Awamori y Saccharomyces Cerevisiae sobre el rendimiento del crecimiento en pollos de engorde; Promoción del metabolismo proteico y modificación del perfil de ácidos grasos en el músculo. J Poult Sci. (2013) 50:242–50. doi: 10.2141/jpsa.0120153
40. Mersmann, HJ. Lipasas liposensibles a lipoproteínas y hormonas en tejido adiposo porcino. J Anim Sci. (1998) 76:1396. doi: 10.2527/1998.7651396x
41. De Smet, P, De Boever, S y Verstraete, W. Reducción del colesterol en cerdos a través de una mayor actividad bacteriana de la sal biliar hidrolasa. BJN. (1998) 79:185–94. doi: 10.1079/bjn19980030
Resumen de PubMed | Texto completo de CrossRef | Google Académico
42. Jin, LZ, Ho, YW, Abdullah, N y Jalaludin, S. Rendimiento del crecimiento, poblaciones microbianas intestinales y colesterol sérico de pollos de engorde alimentados con dietas que contienen cultivos de lactobacilos. Poult Sci. (1998) 77:1259–65. doi: 10.1093/ps/77.9.1259
Resumen de PubMed | Texto completo de CrossRef | Google Académico
43. Korolenko, TA, Bgatova, NP, Ovsyukova, MV, Shintyapina, A y Vetvicka, V. Efectos hipolipidémicos de β-glucanos, mananos y fucoidanos: mecanismo de acción y sus perspectivas de aplicación clínica. Moléculas. (2020) 25:1819. doi: 10.3390/moléculas25081819
Resumen de PubMed | Texto completo de CrossRef | Google Académico
44. Tang, SGH, Sieo, CC, Ramasamy, K, Saad, WZ, Wong, HK y Ho, YW. Rendimiento, respuestas bioquímicas y hematológicas, y pesos relativos de los órganos de gallinas ponedoras alimentadas con dietas suplementadas con prebióticos, probióticos y simbióticos. BMC Vet Res. (2017) 13:1–12. doi: 10.1186/s12917-017-1160-y
45. Jackson, ME, Fodge, DW, y Hsiao, HY. Efectos de la beta-mananasa en dietas de harina de maíz y soja sobre el rendimiento de las gallinas ponedoras. Poult Sci. (1999) 78:1737–41. doi: 10.1093/ps/78.12.1737
Resumen de PubMed | Texto completo de CrossRef | Google Académico
46. Aguafuertes, RJ. Logística del calcio en la gallina ponedora. J Nutr. (1987) 117:619–28. doi: 10.1093/jn/117.3.619
Resumen de PubMed | Texto completo de CrossRef | Google Académico
47. Harr, KE. Química clínica de las especies de aves acompañantes: una revisión. Veterinario Clin Pathol. (2002) 31:140–51. doi: 10.1111/j.1939-165x.2002.tb00295.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
48. Lei, K, Li, YL, Yu, DY, Rajput, IR y Li, WF. Influencia de la inclusión dietética de Bacillus licheniformis en el rendimiento de las gallinas ponedoras, la calidad del huevo, las actividades enzimáticas antioxidantes y la función de barrera intestinal de las gallinas ponedoras. Poult Sci. (2013) 92:2389–95. doi: 10.3382/ps.2012-02686
Resumen de PubMed | Texto completo de CrossRef | Google Académico
49. Chen, JF, Xu, MM, Kang, KL, Tang, SG, He, CQ, Qu, XY, et al. Efectos y efectos combinatorios de Bacillus subtilis y montmorillonita sobre el estado de salud intestinal en gallinas ponedoras. Poult Sci. (2020) 99:1311–9. doi: 10.1016/j.psj.2019.11.016
Resumen de PubMed | Texto completo de CrossRef | Google Académico
50. Oliveira, BL, y Oliveira, DD. Qualidade e Tecnologia de Ovos. Lavras, Brasil: Universidade Federal de Lavras (2013). 223 págs.
51. Carnicero, GD y Miles, RD (2021). Gravedad específica del huevo: diseño de un programa de monitoreo. Universidad de Florida. Disponible en: https://edis.ifas.ufl.edu/publication/VM044 (consultado en agosto de 2021).
52. Gordon, RW, y Roland, DA. Influencia de la fitasa suplementaria en la utilización de calcio y fósforo en gallinas ponedoras. Poult Sci. (1998) 77:290–4. doi: 10.1093/ps/77.2.290
Resumen de PubMed | Texto completo de CrossRef | Google Académico
53. Ledvinka, Z, Zita, L, y Klesalová, L. Calidad del huevo y algunos factores que influyen en ella: una revisión. Sci Agric Bohem. (2012) 43:46–52.
54. Shim, MY, Song, E, Billard, L, Aggrey, SE, Pesti, GM y Sodsee, P. Efectos de los niveles equilibrados de proteínas dietéticas en la producción de huevos y los parámetros de calidad de los huevos de ponedoras comerciales individuales. Poult Sci. (2013) 92:2687–96. doi: 10.3382/ps.2012-02569
55. Moreira, LRSF, y Filho, EX. Una visión general de la estructura del manano y de los sistemas enzimáticos que degradan el manano. Aplicación Microbiol Biotechnol. (2008) 79:165–78. doi: 10.1007/s00253-008-1423-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
56. Lattimer, JM, y Haub, MD. Efectos de la fibra dietética y sus componentes en la salud metabólica. Nutrientes. (2010) 2:1266–89. doi: 10.3390/nu2121266
Resumen de PubMed | Texto completo de CrossRef | Google Académico
57. Anachkov, SE, Georgieva, GS, Abezgauz, L, Danino, D, y Kralchevsky, PA. Pico de viscosidad debido a la transición de forma de micelas similares a gusanos a micelas: efecto del ácido dodecanoico. Langmuir. (2018) 34:4897–907. doi: 10.1021/acs.langmuir.8b00421
Resumen de PubMed | Texto completo de CrossRef | Google Académico
58. Kamranfar, P, y Jamialahmadi, M. Efecto de la transición de forma de micela de surfactante en la viscosidad de la microemulsión y su aplicación en procesos mejorados de recuperación de petróleo. J Mol Liq. (2014) 198:286–91. doi: 10.1016/j.molliq.2014.07.009
59. Bessei, W. Comportamiento de las gallinas ponedoras en sistemas de grupos pequeños desde el punto de vista del bienestar animal. Arch für Geflugelkd. (2010) 74:6–12.
60. Garcia, EA, Mendes, AA, Pizzolante, CC, Gonçalves, HC, Oliveira, RP, y Silva, MA. Efeitos dos níveis de cantaxantina na dieta sobre o desempenho e qualidade dos ovos de poedeiras comerciais. Rev Bras Cien Avi. (2002) 4:1. doi: 10.1590/S1516-635X2002000100007
61. Perry, A, Rasmussen, H, y Johnson, EJ. Contenido de xantofila (luteína, zeaxantina) en frutas, verduras y productos de maíz y huevo. J Comida Compos Anal. (2009) 22:9–15. doi: 10.1016/j.jfca.2008.07.006
62. Cardoso, S.L. Fotofísica de carotenoides e o papel antioxidante de β-caroteno. Química Nova. (1997) 20:535–40. doi: 10.1590/S0100-40421997000500014
63. Feddern, V, Prá, MC, Mores, R, Nicoloso, RS, Coldebella, A, y Abreu, PG. Evaluación de la calidad del huevo en diferentes condiciones de almacenamiento, temporadas y cepas de gallinas ponedoras. Ciênc Agrotec. (2017) 41:322–33. doi: 10.1590/1413-70542017413002317
64. Huang, Q, Qiu, N, Ma, MH, Jin, YG, Yang, H, Geng, F, et al. Estimación de la frescura del huevo utilizando como indicador la S-ovoalbúmina. Poult Sci. (2012) 91:739–43. doi: 10.3382/ps.2011-01639
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Palabras clave: aditivos, indicadores bioquímicos, calidad del huevo, alimentación, salud intestinal, nutrición
Cita: Carvalho CL, Andretta I, Galli GM, Bastos Stefanello T, Camargo NdOT, Mendes RE, Pelisser G, Balamuralikrishnan B, Melchior R y Kipper M (2023) Suplementación dietética con β-mananasa y probióticos como estrategia para mejorar el rendimiento de las gallinas ponedoras y la calidad de los huevos. Frente. Vet. Sci. 10:1229485. doi: 10.3389/fvets.2023.1229485
Editado por:
Ozgur Kaynar, Universidad de Kastamonu, Türkiye
Revisado por:
Alicia Zem Fraga, Universidade Estadual de São Paulo, Brasil Cheila Roberta Lehnen, Universidade Estadual de Ponta Grossa, Brasil
Derechos de autor © 2023 Carvalho, Andretta, Galli, Bastos Stefanello, Camargo, Mendes, Pelisser, Balamuralikrishnan, Melchior y Kipper. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia Creative Commons Attribution License (CC BY).
*Correspondencia: Inés Andretta, ines.andretta@ufrgs.br
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, ni las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o afirmación que pueda hacer su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía