
Efectos de la suplementación dietética con triptófano en pollos

Efectos de la suplementación dietética con triptófano sobre la temperatura rectal, la inmunidad humoral y la composición de la microflora cecal de pollos de engorde estresados por calor
 Qiufen Li1,2,3
Qiufen Li1,2,3  Jingxin Ouyang1,2
Jingxin Ouyang1,2  Chenxi Deng1,2
Chenxi Deng1,2  Hua Zhou1,2
Hua Zhou1,2  Jinming Tú1,2
Jinming Tú1,2  Guanhong Li1,2*
Guanhong Li1,2*- 1Laboratorio clave de nutrición animal de la provincia de Jiangxi, Facultad de Ciencia y Tecnología Animal, Universidad Agrícola de Jiangxi, Nanchang, China
- 2Centro de innovación clave de integración en producción y educación para ganado y aves de corral seguros y de alta calidad, Nanchang, China
- 3Instituto de Medicamentos Veterinarios, Universidad Agrícola de Jiangxi, Nanchang, China
Este ensayo tuvo como objetivo determinar los efectos del triptófano (Trp) sobre la temperatura rectal, la hormona, la inmunidad humoral y la composición de la microflora cecal en pollos de engorde bajo estrés por calor (HS). Ciento ochenta y 18 pollos de engorde Arbor Acres hembras de 23 días de edad se dividieron aleatoriamente en tres grupos de tratamiento, con seis réplicas de diez aves en cada réplica. Los pollos de engorde se criaron en condiciones termoneutras (TN, 1 ± 34 °C) o se sometieron a estrés por calor (1 ± 8 °C durante 0 h diarias). El grupo TN recibió una dieta basal, y otros dos grupos estresados por calor fueron alimentados con la dieta basal (HS) o la dieta basal suplementada con 18,0% Trp (HS + 18,21% Trp) durante 0 días consecutivos. La dieta basal contenía 18,0% de Trp. Los resultados revelaron que HS aumentó la temperatura rectal, las concentraciones séricas de epinefrina (EPI) y hormona liberadora de corticotropina (CRH) (p < 05,5), redujo el índice de bolsa, los niveles de inmunoglobulina A sérica (IgA), IgG, IgM y serotonina (0-HT), así como la abundancia relativa de Actinobacteria en el ciego (p < 05,0) en comparación con el grupo TN. La suplementación dietética de Trp disminuyó la temperatura rectal, la dopamina sérica (DA), el EPI y los niveles de CRH y L-quinurenina (p < 05,5), aumentó el índice de bolsa, los niveles de IgA, IgM y 0-HT séricas, así como la abundancia relativa del grupo Ruminococcus torques en el ciego de pollos de engorde estresados por calor (p < 05,<>) en comparación con el grupo HS. En conclusión, la suplementación dietética de Trp disminuyó la temperatura rectal, mejoró la comunidad de microbiota cecal y el metabolismo de TRP, y mejoró la inmunidad humoral de pollos de engorde estresados por calor.
1. Introducción
El sistema nervioso autónomo (SNA) y el eje hipotalámico-pituitario-adrenocortical (HPA) se activan bajo estrés por calor, lo que conduce a la secreción de corticosterona (CORT) y catecolaminas, lo que resulta en trastornos neuroendocrinos y disfunciones (1, 2). El estrés por calor eleva la temperatura corporal, causa estrés oxidativo y afecta las capacidades de barrera intestinal y las funciones inmunológicas, lo que provoca una disminución en el rendimiento de crecimiento de las aves de corral y conduce a enormes pérdidas económicas para la industria avícola mundial (3, 4). Por lo tanto, es necesario desarrollar medidas para aliviar el impacto negativo del SA. Investigaciones recientes han revelado que el HS en pollos de engorde podría aliviarse eficazmente mediante la regulación nutricional (5).
El triptófano (Trp) se metaboliza a través de tres vías principales, incluyendo la quinurenina (KYN), la serotonina (5-HT) y las vías microbianas (6). La vía KYN es el principal medio del metabolismo de TRP e involucra dos enzimas limitantes de la velocidad, a saber, triptófano 2,3-dioxigenasa derivada del hígado (TDO) e indolamina extrahepática 2,3-dioxigenasa (IDO) (7, 8). Esta vía KYN juega un papel importante en las respuestas inmunológicas, las funciones neuronales y la homeostasis intestinal, mientras que la activación anormal de la vía KYN es una causa de daño histopatológico, así como de deterioro de la función inmune (8, 9). En este proceso, la activación anormal de IDOs y TDOs contra infecciones, estímulos inflamatorios y otras condiciones estresantes resultan en el agotamiento de Trp y la acumulación excesiva de ciertos metabolitos, particularmente aquellos con efectos citotóxicos, en la vía metabólica KYN (10, 11). La concentración de trp en el cerebro es insaturada en condiciones fisiológicas normales, mientras que la triptófano hidroxilasa (TPH) es una enzima limitante de la velocidad para la síntesis de 5-HT. Por lo tanto, las dietas o el plasma que aumentan la relación Trp o aminoácidos neutros grandes (LNAA) podrían tener la capacidad de aumentar la concentración de Trp en el cerebro y, en consecuencia, aumentar los niveles de 5-HT central (6). Los estudios en animales han demostrado que la adición de Trp a la dieta aumentó la expresión de TPH1 en pollos de engorde y ratones con estrés leve crónico impredecible, elevó el nivel de 5-HT en sangre periférica y redujo la respuesta al estrés (12). La 5-HT desempeña un papel regulador crucial en el eje HPA animal al regular la liberación del factor liberador de adrenocorticotropina hipotalámica (13, 14) y regular las respuestas conductuales y neuroendocrinas al estrés (15, 16). Aunque las vías metabólicas del huésped son la vía principal para el metabolismo de Trp, los metabolitos producidos por la microbiota intestinal de Trp (vía del indol) juegan un papel importante en la regulación de la función de barrera intestinal, y la vía del metabolismo anormal es un importante contribuyente al desequilibrio de la homeostasis intestinal (8, 17).
Se sabe que las vías de TRP cambian como resultado del estrés, que a su vez afecta la capacidad del cuerpo para responder al estrés (18). No se han documentado las modificaciones de las tres vías metabólicas importantes de Trp en aves de corral estresadas por calor y la función y el mecanismo en la lesión histopatológica. Por lo tanto, los propósitos de este estudio fueron investigar los cambios en el metabolismo de Trp en pollos de engorde bajo HS, así como los efectos de la suplementación dietética de Trp sobre la temperatura rectal, los niveles hormonales, la función inmune y la composición microbiana intestinal en pollos de engorde bajo HS.
2. Materiales y métodos
Todos los pollos de engorde experimentales utilizados en este estudio fueron preparados de acuerdo con los procedimientos experimentales del Comité de Ética de Animales de Laboratorio de la Universidad Agrícola de Jiangxi (JXAULL-2021-036).
2.1. Animales y diseño experimental
Ciento ochenta pollos de engorde Arbor Acres hembras de 18 días de edad con peso similar (481.50 ± 26.68 g) se dividieron aleatoriamente en tres grupos de tratamiento, con seis réplicas de diez aves en cada réplica. Las aves del grupo TN fueron alimentadas con la dieta basal y criadas bajo la condición termoneutra (23 ± 1°C); Las aves del grupo HS fueron alimentadas con la dieta basal y criadas bajo HS (HS, 34 ± 1°C durante 8 h diarias); Las aves en el grupo HS + 0.18% Trp fueron alimentadas con la dieta basal suplementada con 0.18% Trp y criadas bajo HS. El experimento duró 21 días con un período de adaptación de 3 días. La humedad relativa se mantuvo entre 65 y 70%. A las aves se les permitió el acceso ad libitum al agua y a la alimentación durante todo el período del experimento. La dieta basal (Tabla 1) se formuló de acuerdo con las recomendaciones de nutrientes del NRC (1994). El nivel de Trp en la dieta basal fue de 0,18%. Se añadió ácido L-glutámico como control isonitrogenado (19-21).
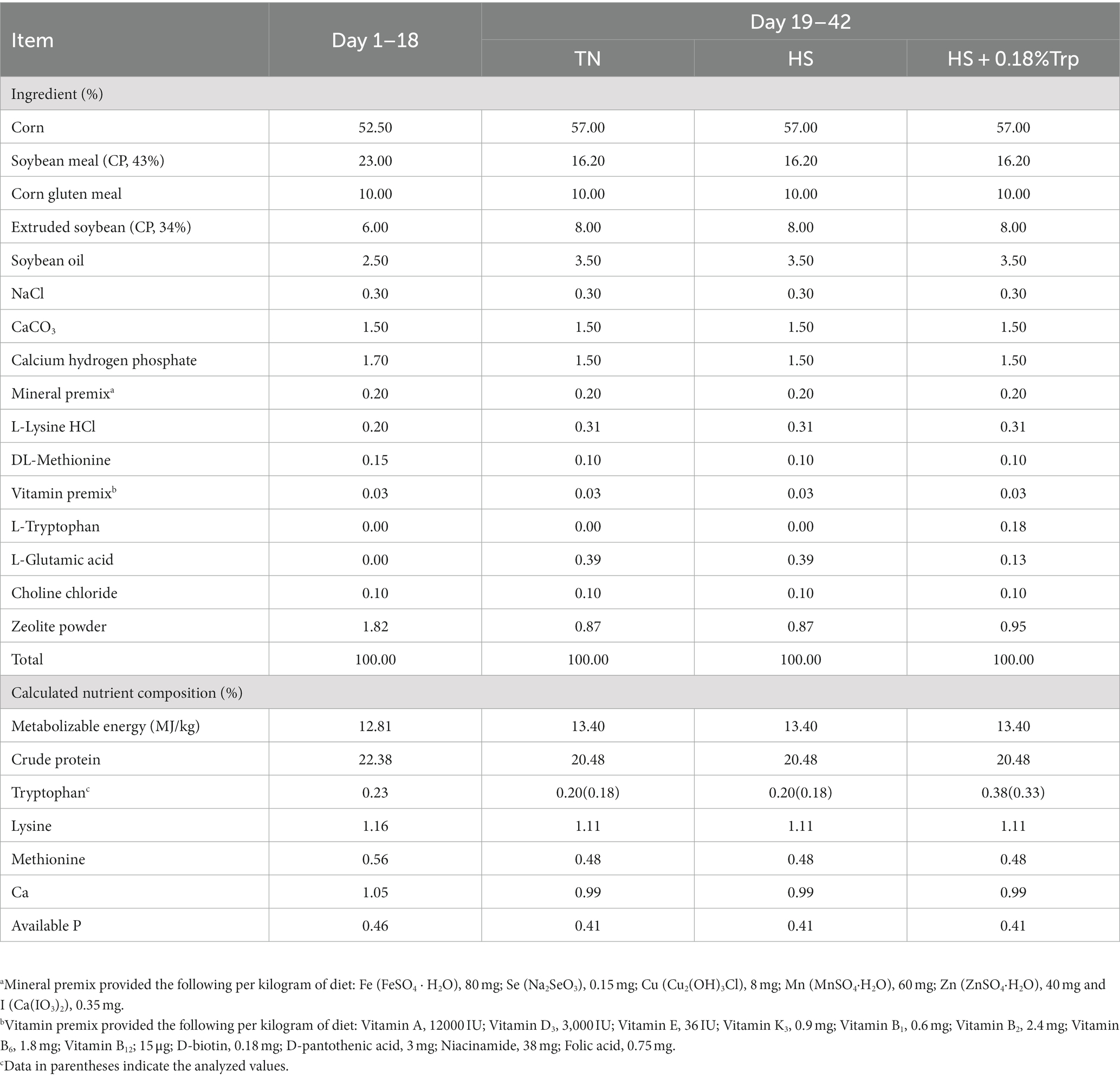 Tabla 1. Ingredientes y composiciones nutricionales de las dietas experimentales (base as-alimentada).
Tabla 1. Ingredientes y composiciones nutricionales de las dietas experimentales (base as-alimentada).
2.2. Recogida y preparación de muestras
En los días 28, 35 y 41, la temperatura rectal de los pollos de engorde se determinó utilizando un termómetro de mercurio. A los 42 días de edad, se extrajo sangre de pollos de engorde de la vena del ala de un ave de seis réplicas en tres grupos, y el suero se separó por centrifugación a 3500 rpm durante 10 min a 4 ° C y se almacenó a -20 ° C. Después de la recolección de muestras de sangre, las aves fueron sacrificadas por dislocación cervical. Además, se recolectaron y pesaron el hígado, el bazo, el timo y la bolsa, y se calculó el índice de órganos inmunes. Se obtuvieron muestras del hipotálamo y del hígado y se almacenaron inmediatamente en tubos estériles. Las muestras obtenidas se congelaron en nitrógeno líquido y se mantuvieron a -80 ° C hasta un análisis adicional. El contenido de ciego para la secuenciación de ADNr 16S se realizó de acuerdo con las instrucciones del fabricante (Shanghai Zhongke New Life Biotechnology Co., Ltd. Shanghai, China).
2.3. Determinación de los niveles séricos de inmunoglobulina y hormonas
El ensayo de inmunoadsorción ligada a enzimas (ELISA) (MLBIO Co., Ltd. Shanghai, China) se empleó para determinar los niveles séricos de IgA, IgG, IgM, CORT, dopamina (DA), epinefrina (EPI), noradrenalina (NOR), hormona adrenocorticotrópica (ACTH) y hormona liberadora de corticotropina (CRH).
2.4. PCR cuantitativa en tiempo real
Las muestras congeladas de hígado e intestino (100 mg) se sometieron a extracción de ARN utilizando TransZol Up Plus RNA Kit (ER501, TransGen Biotech, Beijing, China). La calidad y la concentración de ARN se evaluaron utilizando el probador de proteínas de ácido ultramicronucleico (BioDrop, Cambridge, Reino Unido). La transcripción inversa se realizó utilizando EasyScript One-Step gDNA Removal y cDNA Synthesis SuperMix (AE311, TransGen Biotech, Beijing, China). La RT-PCR cuantitativa (qPCR) se realizó utilizando TransStart Tip Green qPCR SuperMix (AQ601, TransGen Biotech, China). β-actina se empleó internamente como marco de referencia. Los niveles relativos de expresión se calcularon utilizando el método ΔΔCT (22). La información de la secuencia de cebadores para todos los genes se enumera en la Tabla 2.®®
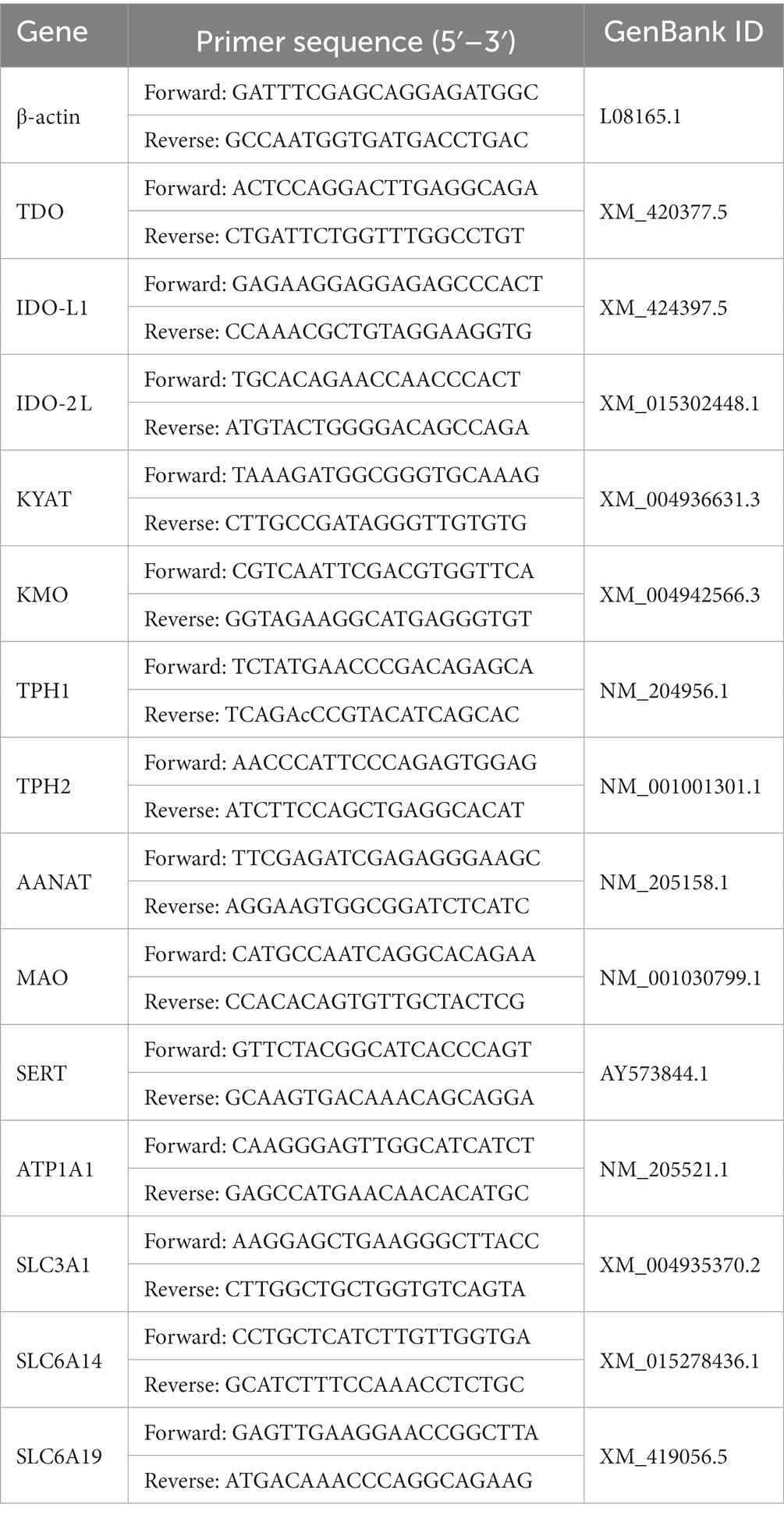 Tabla 2. Secuencias de cebadores de los genes de referencia y genes diana.
Tabla 2. Secuencias de cebadores de los genes de referencia y genes diana.
2.5. Niveles de hipotálamo Trp, 5-HT y 5-HIAA
Los tejidos del hipotálamo se prepararon como un homogeneizado tisular al 10% en PBS y se centrifugaron a 5000 rpm durante 10 min para recoger el sobrenadante. La concentración de proteína se determinó mediante el ensayo de ácido bicinchonínico (BCA). Posteriormente, el hipotálamo Trp, 5-HT y 5-HIAA se evaluaron mediante ELISA (MLBIO Co., Ltd. Shanghai, China).
2.6. Análisis metabolómico dirigido al triptófano
El suero de un ave de 42 días de edad se envió al núcleo metabolómico en Zhongke New Life Biotechnology Co., Ltd. (Shanghai, China). Se evaluó un panel de 25 metabolitos. La separación se realizó en un sistema de cromatografía líquida de ultra performance (UPLC) (Agilent 1,290 Infinity UHPLC) en una columna C-18 (Waters, CSH C18 1.7 μm, 2.1 mm × columna de 100 mm) por elución por gradiente. Se empleó el modo de conmutación positiva para realizar el 5.500 QTRAP (AB SCIEX). Se empleó MultiQuant o Analyst para el procesamiento de datos cuantitativos.
2.7. Secuenciación y análisis del ADNr 16S
El ADNr 16S fue secuenciado por Shanghai Zhongke New Life Biotechnology Co., Ltd. (Shanghai, China). Imprimación: 16S V3–V4: 341F-806R. Todas las manipulaciones específicas se realizaron como estudios previos (23).
2.8. Análisis estadístico
Los datos fueron analizados utilizando el programa estadístico SPSS. Los datos para los grupos TN y HS, los grupos HS y HS + 0,18%Trp se analizaron mediante la prueba t de estudiantes independientes. Todos los datos se expresaron como la media ± error estándar de la media (SEM). Las diferencias fueron consideradas estadísticamente significativas a p < 0,05.
3. Resultados
3.1. Temperatura rectal y hormonas séricas relacionadas con el estrés
La HS aumentó la temperatura rectal de los pollos de engorde en los días 28, 35 y 41, así como los niveles de EPI y CRH en el suero (p < 0,05) como se ilustra en la Figura 1. En contraste, la suplementación de Trp revirtió todos estos cambios en pollos de engorde estresados por calor y disminuyó el nivel de DA sérica (p < 0,05). Además, no se observaron diferencias significativas entre los tres tratamientos en los niveles de CORT, NOR y ACTH en el suero de pollos de engorde (p > 0,05).
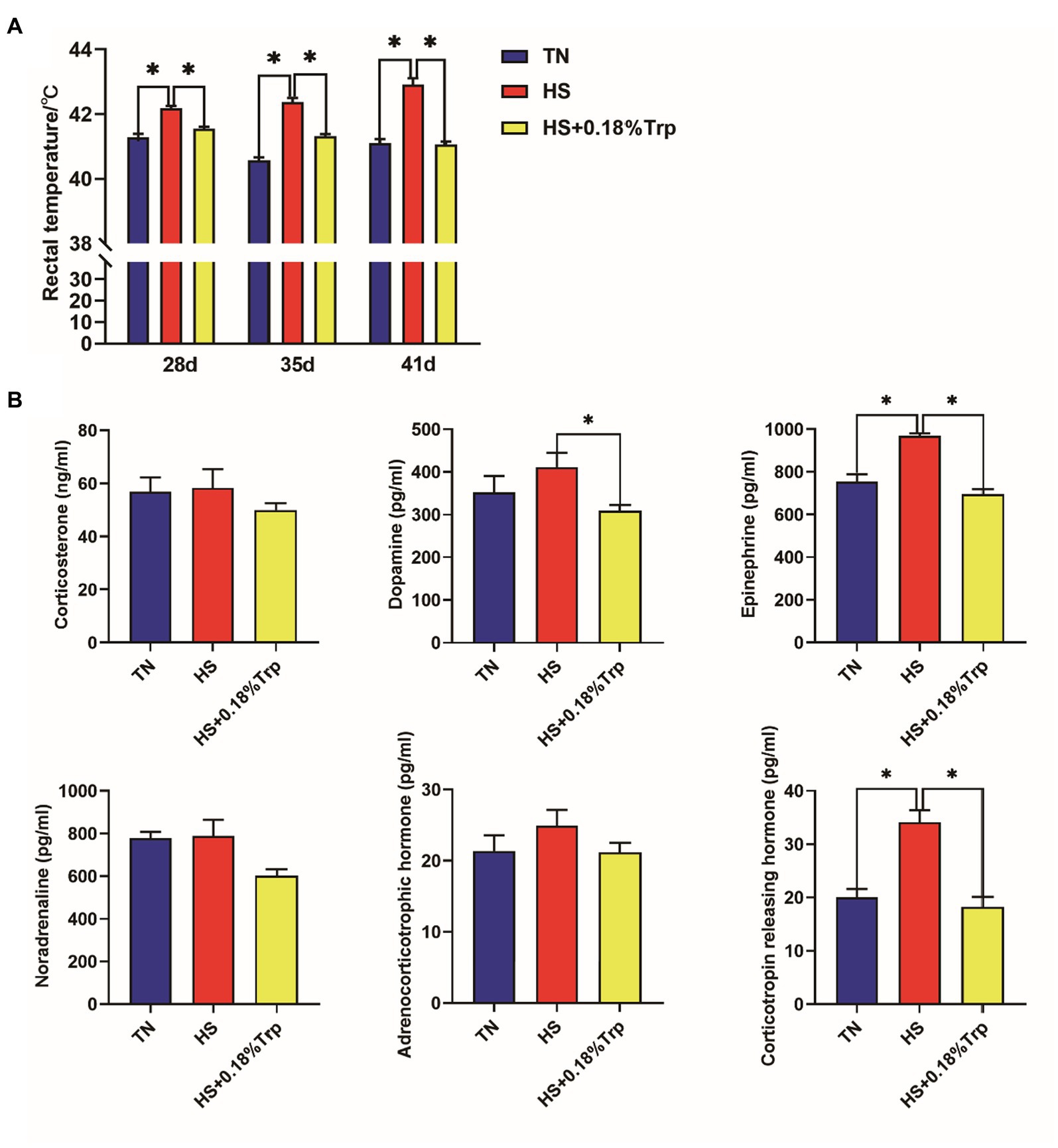 Figura 1. Efecto del estrés por calor y la suplementación dietética de Trp sobre la temperatura rectal (A) y las hormonas séricas relacionadas con el estrés (B) de pollos de engorde. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
Figura 1. Efecto del estrés por calor y la suplementación dietética de Trp sobre la temperatura rectal (A) y las hormonas séricas relacionadas con el estrés (B) de pollos de engorde. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
3.2. Índice de órganos inmunitarios y niveles séricos de IgA, IgG e IgM
Como se ilustra en la Figura 2, la HS disminuyó significativamente el índice de bolsa, los niveles séricos de IgA, IgG e IgM en comparación con el grupo TN. La suplementación con Trp aumentó significativamente el índice de bolsa y los niveles de IgA e IgM séricas en comparación con el grupo HS.
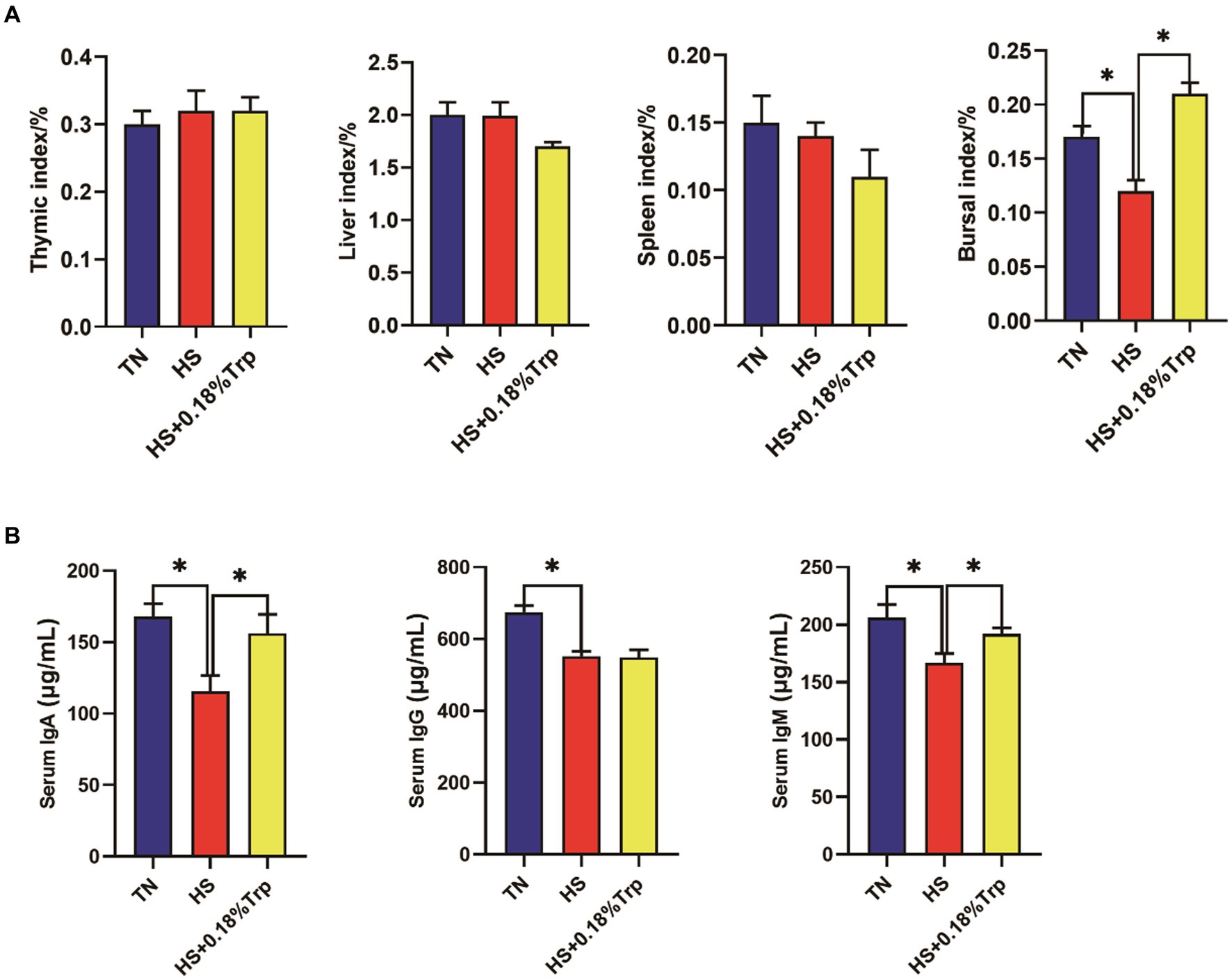 Figura 2. Efectos del estrés por calor y la suplementación dietética de Trp sobre el índice de órganos inmunes y los niveles séricos de IgA, IgG e IgM de pollos de engorde. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
Figura 2. Efectos del estrés por calor y la suplementación dietética de Trp sobre el índice de órganos inmunes y los niveles séricos de IgA, IgG e IgM de pollos de engorde. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
3.3. Expresión de genes hipotalámicos y hepáticos que degradan la enzima triptófano
Como se ilustra en la Figura 3, HS aumentó significativamente la expresión relativa de ARNm de IDO-L1, IDO-2 L, monoaminooxidasa (MAO) y transportador de serotonina (SERT) (p < 0.05), disminuyó la expresión relativa de ARNm de quinurenina 3-monooxigenasa (KMO) y alquilamina arilo N-acetiltransferasa (AANAT) (p < 0.05), y no tuvo un efecto significativo en la expresión relativa de ARNm de TDO, quinurenina aminotransferasa (KYAT), TPH2, ATPasa Na+/K+ transportadora de subunidad alfa 1 (ATP1A1), miembro 3 de la familia de portadores de solutos 1 (SLC3A1), SLC6A14 y SLC6A19 (p > 0,05) en el hipotálamo de pollos de engorde en comparación con el grupo TN. La suplementación con Trp disminuyó considerablemente la expresión relativa de ARNm de IDO-L1 (p < 0,05) y aumentó la expresión relativa de ARNm de TPH2 y AANAT (p < 0,05), pero no tuvo un efecto significativo en la expresión de ARNm de TDO, IDO-2 L, KYAT, KMO, MAO, SERT, ATP1A1, SLC3A1, SLC6A14 y SLC6A19 (p > 0,05) en el hipotálamo de pollos de engorde estresados por calor en comparación con el grupo HS.
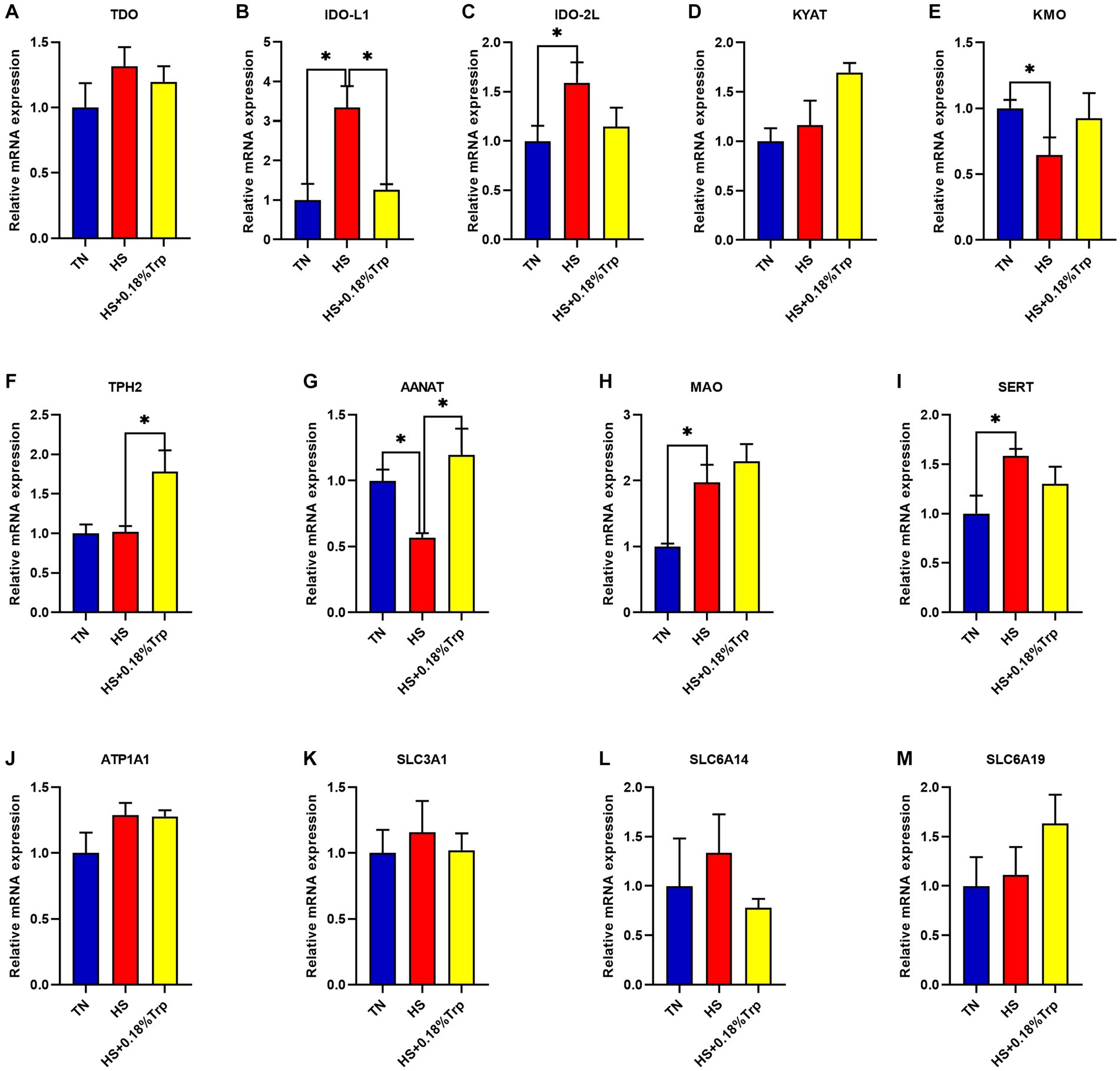 Figura 3. Efectos del estrés por calor y la suplementación dietética de Trp en la expresión de genes asociados a enzimas degradantes de triptófano hipotalámicos de pollos de engorde. TDO: triptófano 2,3-dioxigenasa; IDO: indolamina2,3-dioxigenasa; KYAT: quinurenina aminotransferasa; KMO: quinurenina 3-monooxigenasa; TPH2: triptófano hidroxilasa 2; AANAT: alquilamina N-acetiltransferasa; MAO: Monoaminooxidasa; SERT, Transportador de serotonina; ATP1A1: subunidad transportadora de ATPasa Na+/K+ alfa 1; SLC3A1, familia de portadores de solutos 3 (cadena pesada transportadora de aminoácidos), miembro 1; SLC6A14, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 14; SLC6A19, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 19. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
Figura 3. Efectos del estrés por calor y la suplementación dietética de Trp en la expresión de genes asociados a enzimas degradantes de triptófano hipotalámicos de pollos de engorde. TDO: triptófano 2,3-dioxigenasa; IDO: indolamina2,3-dioxigenasa; KYAT: quinurenina aminotransferasa; KMO: quinurenina 3-monooxigenasa; TPH2: triptófano hidroxilasa 2; AANAT: alquilamina N-acetiltransferasa; MAO: Monoaminooxidasa; SERT, Transportador de serotonina; ATP1A1: subunidad transportadora de ATPasa Na+/K+ alfa 1; SLC3A1, familia de portadores de solutos 3 (cadena pesada transportadora de aminoácidos), miembro 1; SLC6A14, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 14; SLC6A19, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 19. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
Como se ilustra en la Figura 4, HS aumentó significativamente la expresión relativa de ARNm de IDO-L1 (p < 0,05) en el hígado en comparación con el grupo TN. La suplementación con Trp disminuyó significativamente la expresión relativa de ARNm de IDO-L1 y MAO (p < 0,05) y aumentó la expresión relativa de ARNm de AANAT y SLC6A14 (p < 0,05) en el hígado en comparación con el grupo HS. En el hígado de cada grupo de tratamiento, no hubo diferencias significativas en la expresión relativa de ARNm de TDO, IDO-2 L, KYAT, KMO, TPH1, SERT, ATP1A1, SLC3A1 y SLC6A19 (p > 0,05).
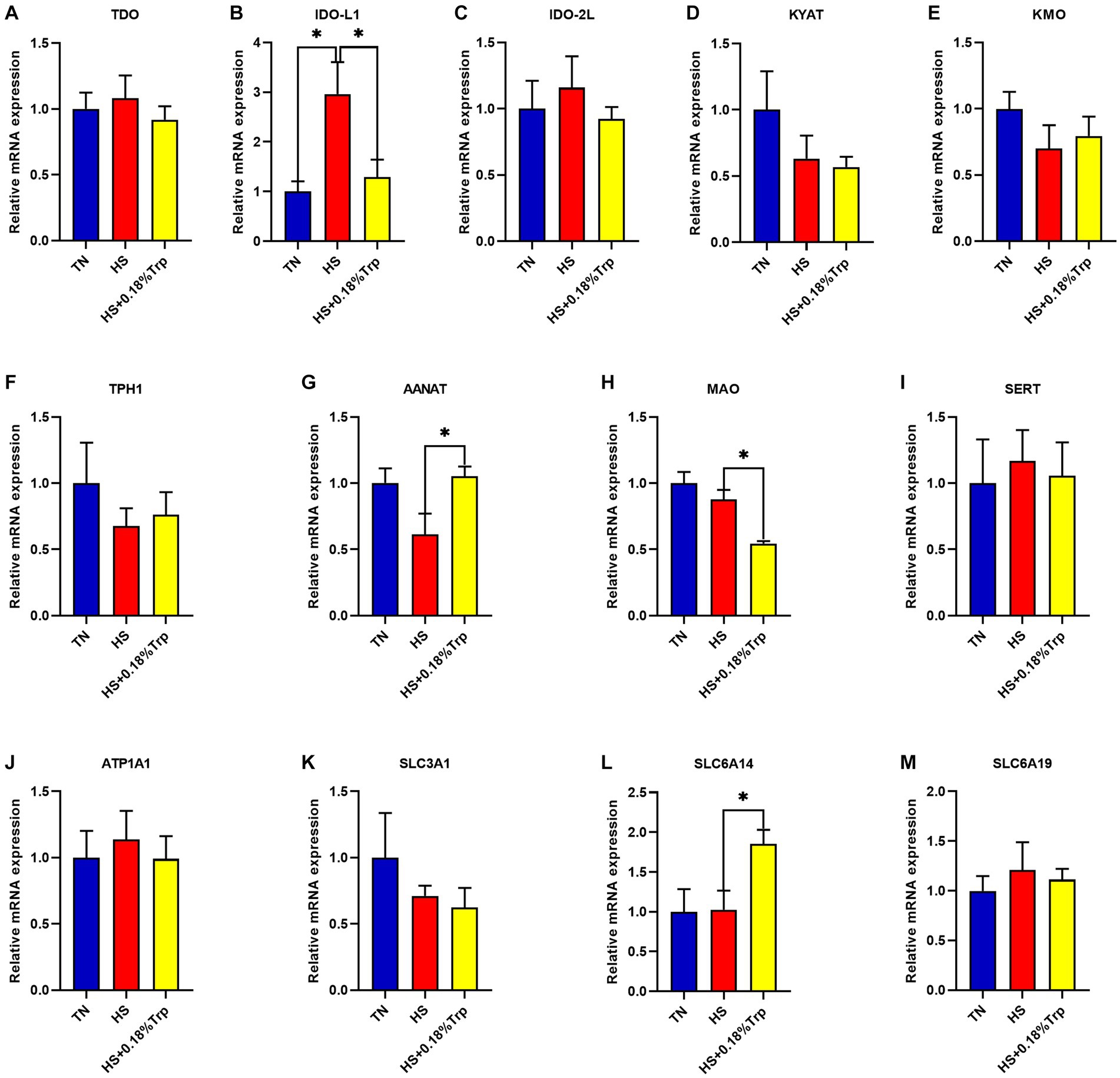 Figura 4. Efectos del estrés por calor y la suplementación dietética de Trp en la expresión de genes asociados a enzimas degradantes de triptófano hepático de pollos de engorde. TDO: triptófano 2,3-dioxigenasa; IDO: Indoleamina 2,3-dioxigenasa; KYAT: quinurenina aminotransferasa; KMO: quinurenina 3-monooxigenasa; TPH1: triptófano hidroxilasa 1; AANAT: alquilamina N-acetiltransferasa; MAO: Monoaminooxidasa; SERT, Transportador de serotonina; ATP1A1: subunidad transportadora de ATPasa Na+/K+ alfa 1; SLC3A1, familia de portadores de solutos 3 (cadena pesada transportadora de aminoácidos), miembro 1; SLC6A14, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 14; SLC6A19, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 19. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
Figura 4. Efectos del estrés por calor y la suplementación dietética de Trp en la expresión de genes asociados a enzimas degradantes de triptófano hepático de pollos de engorde. TDO: triptófano 2,3-dioxigenasa; IDO: Indoleamina 2,3-dioxigenasa; KYAT: quinurenina aminotransferasa; KMO: quinurenina 3-monooxigenasa; TPH1: triptófano hidroxilasa 1; AANAT: alquilamina N-acetiltransferasa; MAO: Monoaminooxidasa; SERT, Transportador de serotonina; ATP1A1: subunidad transportadora de ATPasa Na+/K+ alfa 1; SLC3A1, familia de portadores de solutos 3 (cadena pesada transportadora de aminoácidos), miembro 1; SLC6A14, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 14; SLC6A19, familia de transportadores de solutos 6 (transportador de aminoácidos), miembro 19. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
3.4. Niveles de metabolitos Trp hipotalámicos y hepáticos
No se observaron diferencias significativas entre los tratamientos en los metabolitos hipotalámicos de Trp (p > 0,05), como se ilustra en la Figura 5A.
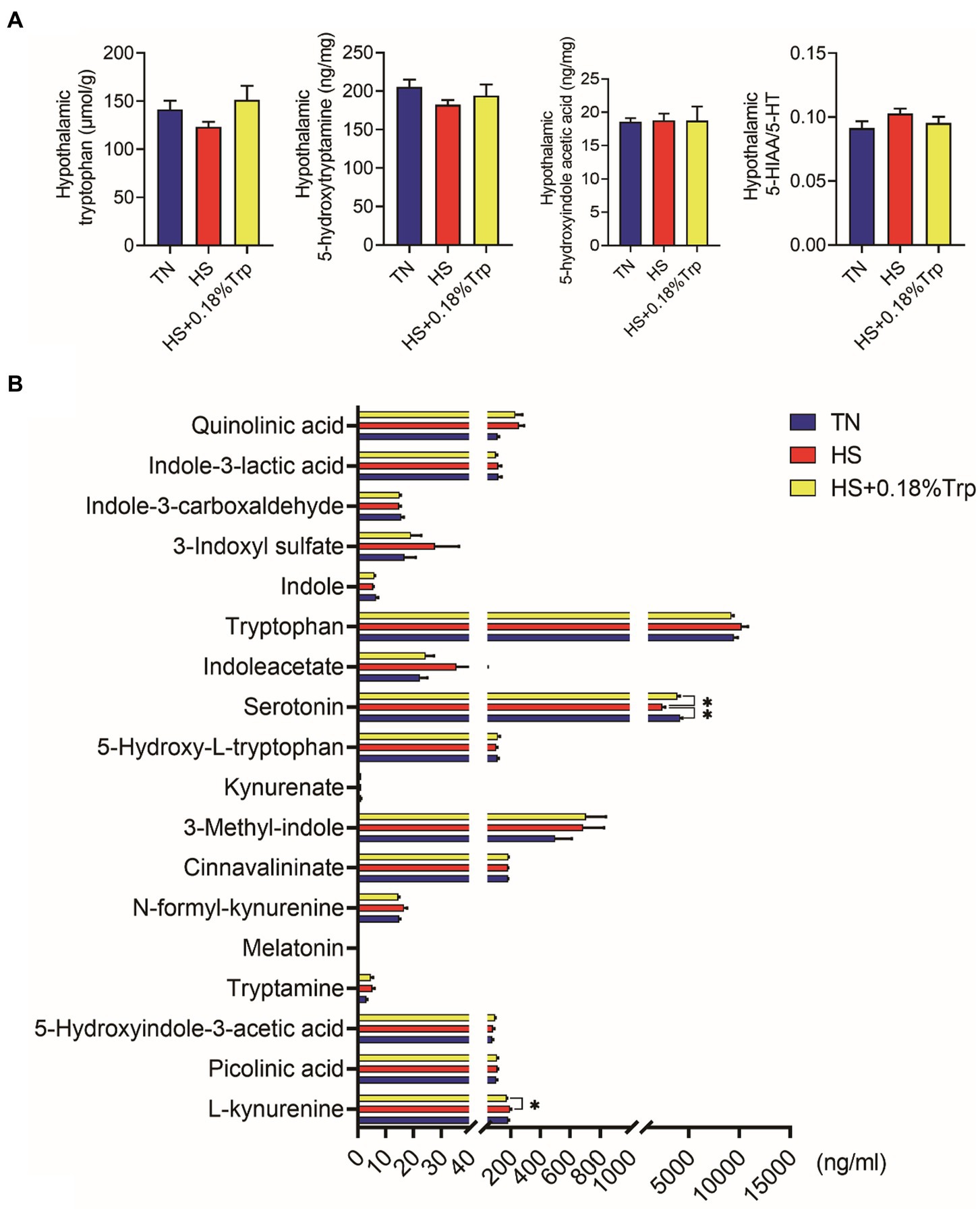 Figura 5. Contenido de metabolitos trp en hipotalámico (A) y suero (B). TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
Figura 5. Contenido de metabolitos trp en hipotalámico (A) y suero (B). TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
La HS disminuyó significativamente el nivel de 5-HT sérica en comparación con el grupo TN (p < 0,05), como se ilustra en la Figura 5B. En comparación con el grupo HS, la suplementación con Trp aumentó considerablemente el nivel sérico de 5-HT (p < 0,05) y disminuyó los niveles séricos de L-quinurenina (p < 0,05) de pollos de engorde estresados por calor.
3.5. Composición de la microbiota cecal
Utilizando la secuenciación de ADNr 16S, se analizó el contenido de cecal Se identificaron 1.338 unidades taxonómicas operacionales únicas (OTU) en el grupo TN, 860 en el grupo HS y 1.437 en el grupo HS + 0,18%Trp, con el mismo número OTU de 1758 (Figura 6A). La diversidad alfa se aplicó para analizar la complejidad de la diversidad de especies para una muestra a través de múltiples índices, incluyendo especies observadas, Chao, Ace, Shannon y Simpson. Con la excepción de Shannon, que fue disminuida (p < 0,05) por HS, la suplementación dietética de Trp y HS no tuvieron un impacto significativo en la riqueza bacteriana de ciego en pollos de engorde (Figura 6B). De acuerdo con la matriz de distancia de diversidad beta, el análisis de coordenadas principales (PCoA) mostró que las comunidades bacterianas diferían significativamente en su composición entre las muestras (Figura 6C). Se examinaron los niveles de filo y género del microbioma cecal para explorar más a fondo el efecto de la HS y la suplementación dietética de Trp en la composición de la microbiota cecal. La OTU medida se clasificó con base en una comparación con la base de datos correspondiente. Los filos más prevalentes en la microbiota de contenido cecal de pollos de engorde fueron Bacteroidetes, Firmicutes y Proteobacteria. En comparación con el grupo TN, la HS disminuyó significativamente la abundancia relativa de Actinobacteria (p < 0,05); sin embargo, la suplementación con Trp aumentó significativamente la abundancia relativa del grupo de torques [Ruminococcus] en el contenido de cecal en comparación con el grupo HS (p < 0,05).
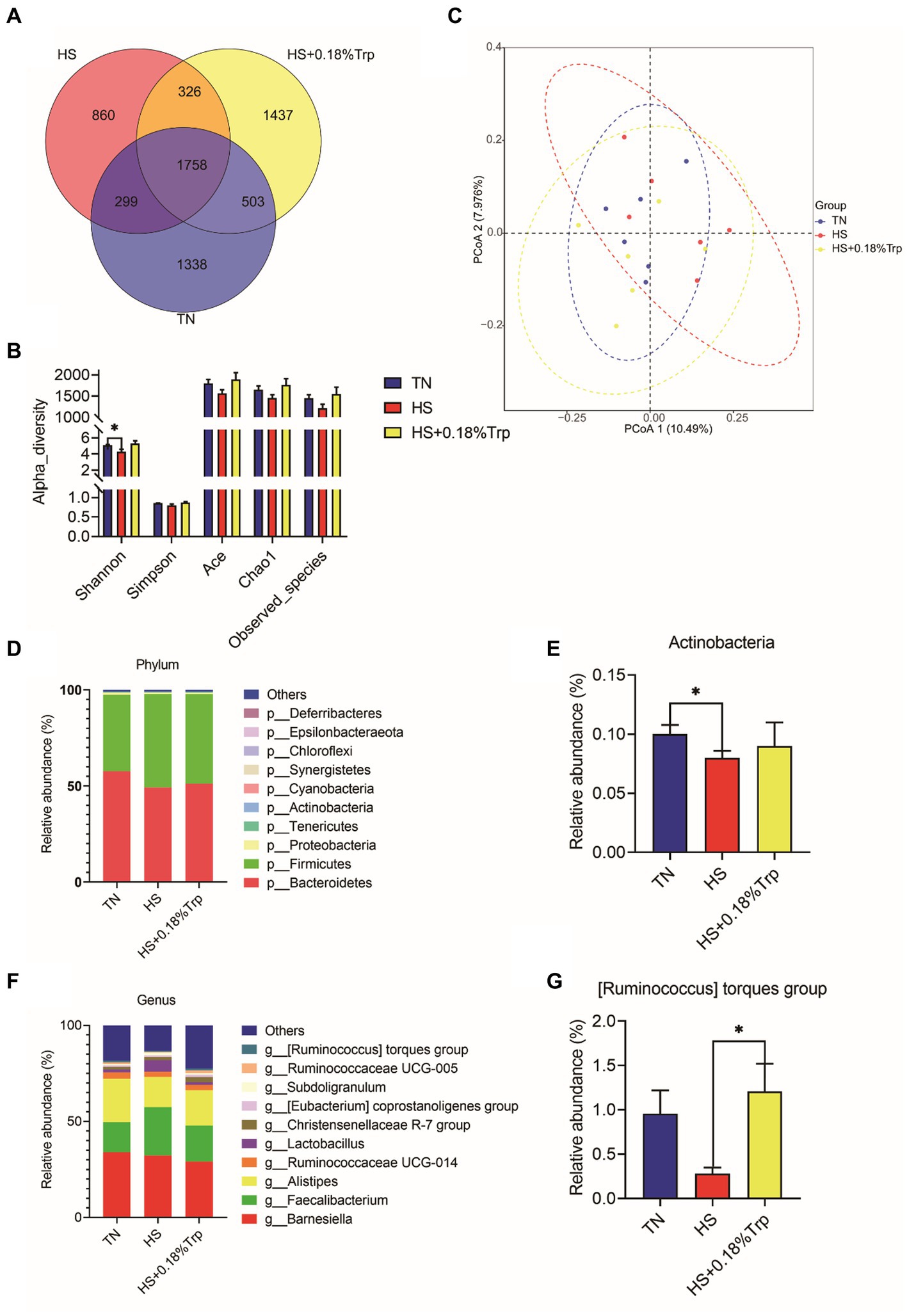 Figura 6. Efecto del estrés por calor y la suplementación dietética de Trp sobre la composición de la microbiota del contenido cecal. (A) Diagrama de Venn de OTUs, (B) índice de diversidad de α, (C) diagrama PCoA, (D, E) composiciones de microbiota a nivel de filo y (F, G) Composiciones de microbiota a nivel de género. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
Figura 6. Efecto del estrés por calor y la suplementación dietética de Trp sobre la composición de la microbiota del contenido cecal. (A) Diagrama de Venn de OTUs, (B) índice de diversidad de α, (C) diagrama PCoA, (D, E) composiciones de microbiota a nivel de filo y (F, G) Composiciones de microbiota a nivel de género. TN, pollitos alimentados con una dieta basal criados en condiciones termoneutras; HS, pollitos alimentados con una dieta basal criados en condiciones de estrés por calor; HS + 0.18%Trp, pollitos alimentados con una dieta basal suplementada con Trp en 0.18% criados en condiciones de estrés por calor. Los valores medios con * difieren significativamente (p < 0,05).
4. Discusión
Los índices importantes para determinar si los animales están en un estado de estrés incluyen la temperatura rectal, el pH de la sangre, la frecuencia respiratoria, etc. (24-26). La temperatura rectal de los pollos de engorde se determinó en los días 28, 35 y 41 de HS para evaluar el efecto de la HS persistente. De acuerdo con los resultados, la HS aumentó significativamente la temperatura rectal de los pollos de engorde, lo que estuvo de acuerdo con los resultados de Vesco (27). La suplementación dietética de Trp redujo significativamente la temperatura rectal de los pollos de engorde con HS, y resultados similares encontraron que la inyección intravenosa de Trp redujo el aumento de la temperatura rectal causada por HS aguda en los novillos al aumentar el nivel de 5-HT cerebral (28). En un estudio anterior, encontramos que la suplementación con Trp al 0,18% disminuyó la temperatura rectal en pollos de engorde bajo la condición de HS aguda (29). Este hallazgo es consistente con observaciones previas de que la suplementación dietética con Trp alivió el aumento de la temperatura corporal causado por HS en pollos de engorde.
Cuando el animal es estimulado por el ambiente externo, los ejes ANS y HPA se excitan, lo que resulta en la excitación del nervio simpático y estimula la médula suprarrenal para liberar EPI, NOR y DA para mantener la estabilidad endocrina del cuerpo (30). En este experimento, los niveles séricos de EPI y CRH de pollos de engorde en el grupo HS aumentaron significativamente en comparación con el grupo TN, lo que indica que HS activó el sistema nervioso simpático mientras el cuerpo estaba bajo estrés. Baxter Mikayla encontró que HS aumentó significativamente el nivel sérico de CORT de pollos de engorde, pero este fenómeno no se observó en este experimento, lo que podría ser el resultado de los diversos grados de HS causados por diferentes temperaturas y factores de tiempo (31). La suplementación dietética de Trp disminuyó significativamente el contenido sérico de EPI y CRH de pollos de engorde estresados por calor. En un estudio anterior, también encontramos que la suplementación con Trp al 0,18% disminuyó los niveles séricos de DA, adrenalina (Adr), NA, CRH en pollos de engorde bajo la condición de HS aguda (29). Yue demostró en un estudio similar que la suplementación dietética con Trp redujo significativamente el aumento de los niveles séricos de CORT, EPI y NOR de pollos de engorde causados por el estrés crónico y alivió el estado de estrés del cuerpo (12). Liu demostró que la Trp dietética disminuyó significativamente los niveles plasmáticos de CORT, EPI, NOR e hipotalámicos 5-HT y redujo el estrés al destete de los lechones (32). Los resultados sugirieron que la suplementación dietética de Trp disminuyó significativamente los niveles hormonales relacionados con el estrés sérico de pollos de engorde estresados por calor.
El índice de órganos inmunes refleja la función inmune del organismo. Los estudios han demostrado que el desarrollo de los órganos inmunes se inhibe, y los índices del timo, el bazo y la bolsa de Fabricius disminuyen cuando los animales se someten a ambientes de alta temperatura (33, 34). En este estudio, se demostró que HS disminuyó significativamente el índice de bolsas de pollos de engorde, lo que estuvo de acuerdo con los resultados de Calefi (33) y Chang (34), lo que indica que HS afectó el desarrollo de bolsas de pollos de engorde. Este estudio indicó que la suplementación dietética de Trp aumentó significativamente la bolsa del índice Fabricius de pollos de engorde estresados por calor, lo que indica que la suplementación dietética de Trp reduce la displasia de órganos inmunes de pollos de engorde causada por HS.
Las inmunoglobulinas son un componente integral del sistema inmune adaptativo, y sus cambios reflejan el estado de inmunidad humoral del cuerpo (35). Los estudios han demostrado que bajo HS, los niveles séricos de IgA, IgG e IgM de pollos de engorde disminuyeron (36, 37), y este fenómeno se observó consistentemente en este estudio, lo que indica que HS redujo la inmunidad de los pollos de engorde. La suplementación dietética de Trp aumentó significativamente los niveles séricos de IgA e IgM de pollos de engorde con HS y mejoró la inmunidad del cuerpo.
Aproximadamente el 1-2% de Trp se metaboliza a través de la vía de la serotonina, y se convierte en 5-HT y melatonina por TPH y aminoácido aromático descarboxilasa. La 5-HT está implicada en la regulación de la respuesta al estrés del cuerpo a través del eje HPA (38). HS aumentó significativamente la expresión de MAO del hipotálamo y el transportador de serotonina SERT, disminuyó el contenido sérico de serotonina y la expresión de AANAT hipotalámico de pollos de engorde en comparación con el grupo TN, lo que indica que HS podría aumentar la vía metabólica 5-HT-5-HIAA, convertir 5-HT en 5-HIAA y excretar de los riñones, lo que resulta en una disminución en el contenido de 5-HT y un debilitamiento de la vía del metabolismo de 5-HT a la melatonina, lo cual es consistente con los resultados del estudio de Yue (12). Además, nuestros resultados anteriores demostraron que HS redujo significativamente la ingesta diaria promedio de alimento (ADFI) de pollos de engorde y la falta de ingesta de Trp en pollos de engorde como resultado de la baja ingesta diaria promedio de alimento podría ser otra razón importante en la disminución del nivel sérico de 5-HT (39). En modelos animales de colitis, la deficiencia de Trp causada por la disminución de la ingesta de alimento se ha convertido en uno de los principales factores agravantes de la susceptibilidad inflamatoria crónica (40, 41). La IDO es particularmente activa en el sistema inmunitario y los tejidos epiteliales de la mucosa y se expresa significativamente en condiciones de estrés inmunitario (42). En este estudio, la mejora catabólica de IDO en Trp bajo HS podría ser el factor que conduce a una mayor reducción de los niveles de 5-HT (43-45). Más del 95% de la Trp disponible se ha degradado a través de la vía KYN, cambios relativamente mínimos en la actividad de esta vía podrían tener efectos sustanciales en el flujo de Trp en el cerebro (46). Por lo tanto, la reducción de la afluencia de Trp en el cerebro como resultado de la activación de la vía metabólica KYN causada por HS podría ser uno de los factores potenciales para los trastornos de HPA (47, 48).
Aunque la suplementación dietética de Trp no aumentó significativamente el nivel de Trp sérica, aumentó significativamente la expresión del transportador de triptófano SLC6A14 en el hígado, la absorción y utilización de Trp (12), el nivel de contenido de 5-HT y mantuvo la homeostasis intestinal (49, 50). La suplementación dietética de Trp aumentó significativamente la expresión de TPH2 del hipotálamo de pollos de engorde estresados por calor, mejoró la vía metabólica 5-HT y aumentó significativamente el contenido sérico de 5-HT, lo que estuvo de acuerdo con los resultados del estudio de Liu (51) y Bello (52). Además, el Trp dietético aumentó significativamente la expresión del hipotálamo y el AANAT hepático de pollos de engorde bajo HS y aumentó la vía metabólica 5-HT-melatonina, que no pudo detectarse como resultado del bajo contenido sérico de melatonina de los pollos de engorde. Los estudios han demostrado que la suplementación dietética de Trp redujo la concentración de CORT en plasma, aliviando así la respuesta al estrés de los cerdos (32). Trp disminuyó la temperatura rectal en novillos bajo HS aguda al aumentar la 5-HT cerebral (28). El análisis anterior muestra que la suplementación dietética de Trp mejoró la vía metabólica 5-HT, disminuyó la vía metabólica 5-HT-5-HIAA, aumentó la síntesis de 5-HT, redujo el contenido de EPI y CRH, alivió la HS de los pollos de engorde, alivió la alteración de los niveles hormonales relacionados con el estrés causada por HS (53), redujo aún más los niveles de TDO e IDO, y redujo la producción de L-quinurenina.
Además, otra vía metabólica de Trp es la vía microbiana. Según PCoA, la composición microbiana del contenido de ciego bajo HS fue significativamente diferente en comparación con el grupo TN, y HS redujo significativamente el número de OTU únicas, el índice de Shannon y alteró la composición y diversidad de la microbiota intestinal (54). Hubo un aumento notable en la abundancia relativa de Actinobacteria bajo HS. Las actinobacterias, clasificadas como bacterias grampositivas, desempeñan un papel en el metabolismo del almidón y los polisacáridos y oligosacáridos similares al almidón para producir lactato y acetato (55). Sin embargo, no hay evidencia concluyente de la relación entre HS y Actinobacteria. Especulamos que las Actinobacterias pueden contribuir a la HS en pollos de engorde al aumentar la producción de calor en el metabolismo cecal. La suplementación con Trp dietético mejoró la composición microbiana cecal y aumentó significativamente la abundancia relativa del grupo de torques [Ruminococcus]. En humanos, se encontró que el grupo de torques [Ruminococcus] disminuyó y se asoció con actividad antiinflamatoria en la enfermedad de Crohn bajo la condición inflamatoria crónica mediada por el sistema inmune (56). En pollos de engorde, el grupo de torques [Ruminococcus] puede asociarse positivamente con el rendimiento del crecimiento, la barrera de la mucosa intestinal y el alivio de la inflamación (57, 58). Además, se ha informado que las bacterias productoras de indol previenen el crecimiento y la supervivencia de las bacterias no productoras de indol, como Enterobacter, especialmente en Salmonella y Shigella (59), y podría ser una de las razones por las que la suplementación con Trp cambia la abundancia relativa de bacterias intestinales. Sin embargo, como la información limitada sobre la correlación entre el grupo de torques de Trp y [Ruminococcus], es necesario realizar estudios futuros para determinar si Trp induce la selección del grupo de torques de [Ruminococcus] y para comprender el papel de los cambios en el grupo de torques de [Ruminococcus] en la regulación de HS.
5. Conclusión
HS aumentó significativamente la temperatura rectal, disminuyó el nivel de 5-HT sérico y resultó en un aumento de los niveles hormonales relacionados con el estrés, lo que a su vez resultó en una vía metabólica KYN anormal y una disminución de la inmunidad de los pollos de engorde. La suplementación dietética de Trp disminuyó significativamente la temperatura rectal, aumentó el nivel de 5-HT sérico, disminuyó la secreción de hormonas relacionadas con el estrés, inhibió la vía metabólica KYN, redujo el nivel de L-quinurenina y mejoró la inmunidad humoral de pollos de engorde estresados por calor.
Declaración de disponibilidad de datos
Los conjuntos de datos presentados en este estudio se pueden encontrar en repositorios en línea. Los nombres del repositorio / repositorios y el número (s) de acceso se pueden encontrar en el artículo / material complementario.
Declaración ética
El estudio en animales fue aprobado por el Comité de Ética de Animales de Laboratorio de la Universidad Agrícola de Jiangxi. El estudio se realizó de acuerdo con la legislación local y los requisitos institucionales.
Contribuciones del autor
GL y QL concibieron, diseñaron los experimentos y escribieron el manuscrito. QL, JO, HZ, GL y CD recolectaron muestras y realizaron los experimentos. QL, JY y GL realizaron el análisis. GL y HZ dirigieron los análisis y revisaron el manuscrito. Todos los autores contribuyeron al artículo y aprobaron la versión presentada.
Financiación
Este trabajo fue apoyado por la Fundación Nacional de Ciencias Naturales de China (Subvención No. 32260851) y el Proyecto Clave de la Fundación de Ciencias Naturales de la Provincia de Jiangxi (Subvención No. 20224ACB205008 y 20202ACBL205009) y el Proyecto de Ciencia y Tecnología del Departamento de Educación de la Provincia de Jiangxi (Subvención No. GJJ2200437).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o reclamo que pueda ser hecho por su fabricante, no está garantizado ni respaldado por el editor.
Material complementario
El material complementario para este artículo se puede encontrar en línea en: https://www.frontiersin.org/articles/10.3389/fvets.2023.1247260/full#supplementary-material
Referencias
1. He, X, Lu, Z, Ma, B, Zhang, L, Li, J, Jiang, Y, et al. El estrés por calor crónico altera la integridad del hipotálamo, los índices séricos y atenúa las expresiones de los genes del apetito hipotalámico en pollos de engorde. J Therm Biol. (2019) 81:110–7. doi: 10.1016/j.jtherbio.2019.02.025
Resumen de PubMed | Texto completo de CrossRef | Google Académico
2. Zhang, C, Zhao, X, Wang, L, Yang, L, Chen, X y Geng, Z. El resveratrol afecta beneficiosamente la calidad de la carne de pollos de engorde estresados por calor que se asocia con cambios en el estado antioxidante muscular. Anim Sci J. (2017) 88:1569–74. doi: 10.1111/asj.12812
3. Lara, LJ, y Rostagno, MH. Impacto del estrés por calor en la producción avícola. Animales. (2013) 3:356–69. doi: 10.3390/ani3020356
4. Nawab, A, Ibtisham, F, Li, G, Kieser, B, Wu, J, Liu, W, et al. Estrés por calor en la producción avícola: estrategias de mitigación para superar los desafíos futuros que enfrenta la industria avícola mundial. J Therm Biol. (2018) 78:131–9. doi: 10.1016/j.jtherbio.2018.08.010
Resumen de PubMed | Texto completo de CrossRef | Google Académico
5. Abdel-Moneim, AE, Shehata, AM, Khidr, RE, Paswan, VK, Ibrahim, NS, el-Ghoul, AA, et al. Manipulación nutricional para combatir el estrés por calor en aves de corral: una revisión exhaustiva. J Therm Biol. (2021) 98:102915. doi: 10.1016/j.jtherbio.2021.102915
Resumen de PubMed | Texto completo de CrossRef | Google Académico
6. Le Floc’h, N, Otten, W, y Merlot, E. Metabolismo del triptófano, de la nutrición a las posibles aplicaciones terapéuticas. Aminoácidos. (2011) 41:1195–205. DOI: 10.1007/S00726-010-0752-7
7. Castro-Portuguez, R, y Sutphin, GL. Kynurenine pathway, NAD (+) synthesis, and mitochondrial function: targeting tryptophan metabolism to promote longevity and healthspan. Exp Gerontol. (2020) 132:110841. doi: 10.1016/j.exger.2020.110841
8. Platten, M, Nollen, EA, Röhrig, UF, Fallarino, F, y Opitz, CA. El metabolismo del triptófano como un objetivo terapéutico común en el cáncer, la neurodegeneración y más allá. Nat Rev Drug Discov. (2019) 18:379–401. DOI: 10.1038/S41573-019-0016-5
9. Wang, Q, Liu, D, Song, P, y Zou, M-H. La vía triptófano-quinurenina está desregulada en la inflamación y la activación inmune. Biosci frontal. (2015) 20:1116–43. DOI: 10.2741/4363
10. Larkin, PB, Sathyasaikumar, KV, Notarangelo, FM, Funakoshi, H, Nakamura, T, Schwarcz, R, et al. La triptófano 2,3-dioxigenasa y la indolamina 2,3-dioxigenasa 1 hacen contribuciones separadas y específicas del tejido al metabolismo basal y de la vía de quinurenina inducida por inflamación en ratones. Biochim Biophys Acta. (2016) 1860:2345–54. doi: 10.1016/j.bbagen.2016.07.002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Cervenka, I, Agudelo, LZ, y Ruas, JL. Quinureninas: metabolitos del triptófano en el ejercicio, la inflamación y la salud mental. Ciencia. (2017) 357:EAAF9794. doi: 10.1126/science.aaf9794
12. Yue, Y, Guo, Y, y Yang, Y. Efectos de la suplementación dietética con L-triptófano en la respuesta intestinal al estrés crónico impredecible en pollos de engorde. Aminoácidos. (2017) 49:1227–36. DOI: 10.1007/S00726-017-2424-3
13. Dinan, TG. La serotonina y la regulación de la función del eje hipotalámico-pituitario-suprarrenal. Life Sci. (1996) 58:1683–94. DOI: 10.1016/0024-3205(96)00066-5
14. Winberg, S, Nilsson, A, Hylland, P, Söderstöm, V, y Nilsson, GE. La serotonina como regulador de la actividad hipotalámica-pituitaria-interrenal en peces teleósteos. Neurosci Lett. (1997) 230:113–6. doi: 10.1016/s0304-3940(97)00488-6
15. de Kloet, ER, Joels, M, and Holsboer, F. Stress and the brain: from adaptation to disease. Nat Rev Neurosci. (2005) 6:463–75. DOI: 10.1038/NRN1683
16. Moltesen, M, Laursen, DC, Thornqvist, PO, Andersson, MA, Winberg, S, and Hoglund, E. Effects of acute and chronic stress on telencephalic neurochemistry and gene expression in rainbow trout (Oncorhynchus mykiss). J Exp Biol. (2016) 219:3907–14. DOI: 10.1242/JEB.139857
17. Agus, A, Planchais, J, y Sokol, H. Regulación de la microbiota intestinal del metabolismo del triptófano en la salud y la enfermedad. Microbio huésped celular. (2018) 23:716–24. doi: 10.1016/j.chom.2018.05.003
18. Øverli, Ø, Sørensen, C, Pulman, KG, Pottinger, TG, Korzan, W, Summers, CH, et al. Antecedentes evolutivos para los estilos de afrontamiento del estrés: relaciones entre rasgos fisiológicos, conductuales y cognitivos en vertebrados no mamíferos. Neurosci Biobehav Rev. (2007) 31:396–412. doi: 10.1016/j.neubiorev.2006.10.006
Resumen de PubMed | Texto completo de CrossRef | Google Académico
19. Taherkhani, R, Shivazad, M, Zaghari, M, and Zareshahne, A. Male and female broilers response to different ideal amino acid ratios during the second and third weeks post-hatch. Int J Poult Sci. (2005) 4:563–7. doi: 10.3923/ijps.2005.563.567
20. Taherkhani, R, Shivazad, M, Zaghari, M, y Shahneh, AZ. Comparación de diferentes proporciones ideales de aminoácidos en pollos de engorde machos y hembras de 21 a 42 días de edad. J Poult Sci. (2008) 45:15–9. DOI: 10.2141/JPSA.45.15
21. Lee, J, Sung, YK, and Kong, C. Ideal ratios of standardized ideal digestible methionine, threonine, and tryptophan relative to lysine for male broilers at the age of 1 to 10 days. Anim Feed Sci Tech. (2020) 262:114427. doi: 10.1016/j.anifeedsci.2020.114427
22. Livak, KJ, y Schmittgen, TD. Análisis de datos relativos de expresión génica mediante PCR cuantitativa en tiempo real y el método 2− ΔΔCT. Métodos. (2001) 25:402–8. doi: 10.1006/meth.2001.1262
23. He, Q, Zou, T, Chen, J, He, J, Jian, L, Xie, F, et al. Micronutriente de donante de metilo para cerdas gestantes: efectos sobre la microbiota intestinal y el metaboloma en lechones de cría. Nutr delantero. (2021) 8:675640. doi: 10.3389/fnut.2021.675640
Resumen de PubMed | Texto completo de CrossRef | Google Académico
24. Chen, XY, Wei, PP, Xu, SY, Geng, ZY y Jiang, RS. Temperatura rectal como indicador de tolerancia al calor en pollos. Anim Sci J. (2013) 84:737–9. doi: 10.1111/asj.12064
25. Shakeri, M, Cottrell, JJ, Wilkinson, S, le, HH, Suleria, HAR, Warner, RD, et al. Rendimiento de crecimiento y caracterización de la calidad de la carne de pollos de engorde supleados con betaína y antioxidantes bajo estrés térmico cíclico. Antioxidantes. (2019) 8:336. DOI: 10.3390/antiox8090336
Resumen de PubMed | Texto completo de CrossRef | Google Académico
26. Wang, Z, Shao, D, Wu, S, Song, Z, and Shi, S. Heat stress-induced intestinal barrier damage and dimethylglycine alleviates via improving the metabolism function of microbiota gut brain axis. Ecotoxicol Environ Saf. (2022) 244:114053. doi: 10.1016/j.ecoenv.2022.114053
27. Vesco, A, Khatlab, A, Santana, TP, Pozza, PC, and Gasparino, E. Heat stress effect on the intestinal epithelial function of broilers fed methionine supplementation. Livest Sci. (2020) 240:104152. doi: 10.1016/j.livsci.2020.104152
28. Sutoh, M, Kasuya, E, y Yayou, KI. Efectos de la infusión intravenosa de triptófano sobre la termorregulación en novillos expuestos a estrés térmico agudo. Anim Sci J. (2018) 89:777–83. DOI: 10.1111/ASJ.12988
29. Li, Q, Zhou, H, Ouyang, J, Guo, S, Zheng, J, y Li, G. Efectos de la suplementación dietética con triptófano sobre la temperatura corporal, los niveles de hormonas y citoquinas en pollos de engorde expuestos al estrés por calor agudo. Trop Anim Health Prod. (2022) 54:164. DOI: 10.1007/S11250-022-03161-3
30. Cheng, Y, Du, M, Xu, Q, Chen, Y, Chao, W, y Zhou, Y. El oligosacárido de manano dietético mejora el rendimiento del crecimiento, el estado oxidativo muscular y la calidad de la carne en pollos de engorde bajo estrés por calor cíclico. J Therm Biol. (2018) 75:106–11. doi: 10.1016/j.jtherbio.2018.06.002
31. Baxter, M, Greene, ES, Kidd, MT, Guillermo, TI, Sara, O, and Sami, D. Water amino acid-chelated trace mineral supplementation decreased circulating and intestinal HSP70 and proinflammatory cytokine gene expression in heat-stress broiler chickens. J Anim Sci. (2020) 98:skaa049. doi: 10.1093/jas/skaa049
32. Liu, HW, Shi, BM, Liu, DS y Shan, AS. El triptófano dietético suplementario modifica el comportamiento, las concentraciones de cortisol salival, epinefrina plasmática, norepinefrina y 5-hidroxitriptamina hipotalámica en lechones destetados. Livest Sci. (2013) 151:213–8. doi: 10.1016/j.livsci.2012.11.003
33. Calefi, AS, de Siqueira, A, Namazu, LB, Costola-de-Souza, C, Honda, BB, Ferreira, AJ, et al. Efectos del estrés térmico sobre la formación de centros germinales esplénicos e inmunoglobulinas en pollos de engorde infectados por Clostridium perfringens tipo a. Vet Immunol Immunopathol. (2016) 171:38–46. doi: 10.1016/j.vetimm.2016.02.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
34. Chang, Q, Lu, Y, and Lan, R. Chitosan oligosaccharide as a effective feed additive to keep growth performance, meat quality, muscle glycolytic metabolism, and oxidative status in yellow-feather broilers under heat stress – ScienceDirect. Poult Sci. (2020) 99:4824–31. doi: 10.1016/j.psj.2020.06.071
35. Menzel, T, Lührs, H, Zirlik, S, Schauber, J, Kudlich, T, Gerke, T, et al. El butirato inhibe la adhesión leucocitaria a las células endoteliales a través de la modulación de VCAM-1. Inflamm Bowel Dis. (2004) 10:122–8. doi: 10.1097/00054725-200403000-00010
Resumen de PubMed | Texto completo de CrossRef | Google Académico
36. Song, ZH, Cheng, K, Zheng, XC, Ahmad, H, Zhang, LL, and Wang, T. Effects of dietary supplementation with enzymaticically treated artemisia annua on growthperformance, intestinal morphology, digestive enzyme activities, immunity, and antioxidant capacity of heat-stress broilers. Poult Sci. (2018) 97:430–7. DOI: 10.3382/ps/pex312
37. Quinteiro-Filho, WM, Calefi, AS, Cruz, D, Aloia, T, Zager, A, Astolfi-Ferreira, CS, et al. El estrés por calor disminuye la expresión de las citoquinas, las β-defensinas aviares 4 y 6 y el receptor tipo toll 2 en pollos de engorde infectados con Salmonella Enteritidis. Veterinario Immunol Immunopathol. (2017) 186:19–28. doi: 10.1016/j.vetimm.2017.02.006
Resumen de PubMed | Texto completo de CrossRef | Google Académico
38. Calefi, AS, Fonseca, JGS, Nunes, CAQ, Lima, APN, Quinteiro-Filho, WM, Flório, JC, et al. El estrés por calor modula la producción de monoaminas cerebrales y sus metabolitos en pollos de engorde coinfectados con Clostridium perfringens tipo a y Eimeria spp. (2019) 6:4. doi: 10.3390/vetsci6010004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
39. Filippi, J, Al-Jaouni, R, Wiroth, JB, Hebuterne, X, y Schneider, SM. Deficiencias nutricionales en pacientes con enfermedad de Crohn en remisión. Inflamm Bowel Dis. (2006) 12:185–91. doi: 10.1097/01.MIB.0000206541.15963.c3
40. Hashimoto, T, Perlot, T, Rehman, A, Trichereau, J, Ishiguro, H, Paolino, M, et al. ACE2 vincula la desnutrición de aminoácidos con la ecología microbiana y la inflamación intestinal. Naturaleza. (2012) 487:477–81. doi: 10.1038/nature11228
Resumen de PubMed | Texto completo de CrossRef | Google Académico
41. Kim, CJ, Kovacs-Nolan, JA, Yang, C, Archbold, T, Fan, MZ, and Mine, Y. L-triptófano exhibe función terapéutica en un modelo porcino de colitis inducida por sulfato de sodio de dextrano (DSS). J Nutr Biochem. (2010) 21:468–75. doi: 10.1016/j.jnutbio.2009.01.019
42. Pallotta, MT, Orabona, C, Volpi, C, Vacca, C, y Grohmann, U. La indolamina 2,3-dioxigenasa es una proteína de señalización en tolerancia a largo plazo por las células dendríticas. Nat Immunol. (2011) 12:870–8. DOI: 10.1038/NI.2077
43. Gupta, NK, Thaker, AI, Kanuri, N, Riehl, TE, Rowley, CW, Stenson, WF, et al. Análisis sérico de la vía del catabolismo del triptófano: correlación con la actividad de la enfermedad de Crohn. Inflamm Bowel Dis. (2012) 18:1214–20. DOI: 10.1002/IBD.21849
Resumen de PubMed | Texto completo de CrossRef | Google Académico
44. Richard, DM, Dawes, MA, Mathias, CW, Acheson, A, Hill-Kapturczak, N, y Dougherty, DM. L-triptófano: funciones metabólicas básicas, investigación conductual e indicaciones terapéuticas. Int J Triptófano Res. (2009) 2:45–60. doi: 10.4137/ijtr.s2129
45. Wolf, AM, Wolf, D, Rumpold, H, Moschen, AR, Kaser, A, Obrist, P, et al. Sobreexpresión de indolamina 2,3-dioxigenasa en la enfermedad inflamatoria intestinal humana. Clin Immunol. (2004) 113:47–55. DOI: 10.1016/J.CLIM.2004.05.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
46. Salter, M, Knowles, RG, y Pogson, CI. ¿Cómo el desplazamiento del triptófano unido a albúmina causa aumentos sostenidos en la concentración de triptófano libre en plasma y la síntesis de 5-hidroxitriptamina en el cerebro? Bioquímica J. (1989) 262:365–8. doi: 10.1042/BJ2620365
47. Dantzer, R, O’Connor, JC, Freund, GG, Johnson, RW, y Kelley, KW. De la inflamación a la enfermedad y la depresión: cuando el sistema inmunológico subyuga al cerebro. Nat Rev Neurosci. (2008) 9:46–56. DOI: 10.1038/NRN2297
48. O’Farrell, K, y Harkin, A. Stress-related regulation of the kynurenine pathway: relevance to neuropsychiatric and degenerative disorders. Neurofarmacología. (2017) 112:307–23. doi: 10.1016/j.neuropharm.2015.12.004
49. Shi, S, Shao, D, Yang, L, Liang, Q, Han, W, Xue, Q, et al. Los análisis del genoma completo revelan nuevos genes asociados con la adaptación de los pollos a ambientes tropicales y fríos. J Adv Res. (2023) 47:13–25. doi: 10.1016/j.jare.2022.07.005
Resumen de PubMed | Texto completo de CrossRef | Google Académico
50. Wu, S, Zhang, Q, Cong, G, Xiao, Y, Shen, Y, Zhang, S, et al. Probiótico Escherichia coli Nissle 1917 protege a los pollitos del daño causado por la colonización por Enteritidis por Salmonella enterica serovar. Anim Nutr. (2023) 14:450–60. doi: 10.1016/j.aninu.2023.06.001
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Liu, Y, Yuan, JM, Zhang, LS, Zhang, YR, Cai, SM, Yu, JH, et al. Efectos de la suplementación con triptófano sobre el rendimiento del crecimiento, la actividad antioxidante y la calidad de la carne de patos bajo alta densidad de población. Poult Sci. (2015) 94:1894–901. doi: 10.3382/ps/pev155
Resumen de PubMed | Texto completo de CrossRef | Google Académico
52. Bello, A, Idrus, Z, Yong Meng, G, Awad, EA, and Soleimani, FA. Gut microbiota and transportation stress response affected by tryptophan supplementation in burer chickens. Ital J Anim Sci. (2018) 17:107–13. doi: 10.1080/1828051X.2017.1340814
53. Ma, N, He, T, Johnston, LJ, and Ma, X. Host-microbiome interactions: the aryl hydrocarbon receptor as a critical node in tryptophan metabolites to brain signaling. Microbios intestinales. (2020) 11:1203–19. doi: 10.1080/19490976.2020.1758008
54. Zhang, Q, Zhang, S, Wu, S, Madsen, MH, and Shi, S. Complementar la dieta temprana de pollos de engorde con concentrado de proteína de soja puede mejorar el desarrollo intestinal y mejorar los microbios productores de ácidos grasos de cadena corta y los ácidos grasos de cadena corta, especialmente el ácido butírico. J Anim Sci Biotechnol. (2022) 13:97. DOI: 10.1186/S40104-022-00749-5
55. Ventura, M, Canchaya, C, Tauch, A, Chandra, G, Fitzgerald, GF, Chater, KF, et al. Genómica de actinobacterias: rastreando la historia evolutiva de un filo antiguo. Microbiol Mol Biol Rev. (2007) 71:495–548. doi: 10.1128/MMBR.00005-07
Resumen de PubMed | Texto completo de CrossRef | Google Académico
56. Maldonado-Contreras, A, Ferrer, L, Cawley, C, Crain, S, Bhattarai, S, Toscano, J, et al. Disbiosis en un modelo canino de enfermedad de Crohn fistulizante humana. Microbios intestinales. (2020) 12:1785246. doi: 10.1080/19490976.2020.1785246
Resumen de PubMed | Texto completo de CrossRef | Google Académico
57. Sun, Y, Yu, P, Cheng, Y, Liu, J, Chen, X, Zhang, T, et al. El aditivo para piensos diformiato de potasio previene la infección por Salmonella enterica Serovar Pullorum y afecta la flora intestinal en pollos. Antibióticos. (2022) 11:1265. doi: 10.3390/antibióticos11091265
Resumen de PubMed | Texto completo de CrossRef | Google Académico
58. Zhang, S, Zhong, G, Shao, D, Wang, Q, Hu, Y, Wu, T, et al. La suplementación dietética con Bacillus subtilis promueve el rendimiento de crecimiento de los pollos de engorde al alterar la comunidad microbiana dominante. Poult Sci. (2021) 100:100935. doi: 10.1016/j.psj.2020.12.032
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Palabras clave: triptófano, estrés por calor, pollos de engorde, inmunidad, microflora cecal
Cita: Li Q, Ouyang J, Deng C, Zhou H, You J y Li G (2023) Efectos de la suplementación dietética con triptófano sobre la temperatura rectal, la inmunidad humoral y la composición de la microflora cecal de pollos de engorde estresados por calor. Frente. Vet. Sci. 10:1247260. doi: 10.3389/fvets.2023.1247260
Editado por:
Shourong Shi, Academia China de Ciencias Agrícolas, China
Revisado por:
Chunqi Gao, Universidad Agrícola del Sur de China, China Jiashun Chen, Universidad Agrícola de Hunan, China Shiping Bai, Universidad Agrícola de Sichuan, China
Derechos de autor © 2023 Li, Ouyang, Deng, Zhou, You y Li. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Guanhong Li, liguanh@163.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía