
Función olfativa canina asociada con la salud, el manejo y los posibles vínculos con la microbiota

Cuando la nariz no lo sabe: función olfativa canina asociada con la salud, el manejo y los posibles vínculos con la microbiota
 Eileen K. Jenkins1,
Eileen K. Jenkins1,  Mallory T. DeChant2 y
Mallory T. DeChant2 y  Erin B. Perry2*
Erin B. Perry2*- 1Programa de Educación Veterinaria de Primer Año de Posgrado, Actividad de Salud Pública – Fort Bragg, Ejército de los Estados Unidos, Fort Bragg, NC, Estados Unidos
- 2Departamento de Ciencia Animal, Alimentos y Nutrición, Facultad de Ciencias Agrícolas, Universidad del Sur de Illinois, Carbondale, IL, Estados Unidos
El impacto de la salud, el manejo y la microbiota en la función olfativa en los caninos no se ha examinado en revisión. La característica más importante del canino de detección es su sentido del olfato. Los receptores olfativos se encuentran principalmente en los etmocornetes de la cavidad nasal. El órgano vomeronasal es un sitio adicional de detección de olores que detecta señales químicas que estimulan cambios conductuales y / o fisiológicos. Los avances recientes en la genética del olfato sugieren que los cambios genéticos, junto con la anatomía única y el flujo de aire de la nariz canina, son responsables de la macrosmia de la especie. La inflamación, las alteraciones en el flujo sanguíneo y la hidratación, y las enfermedades sistémicas alteran el olfato y pueden afectar la eficiencia de trabajo de los caninos de detección. La literatura científica contiene abundante información sobre el impacto potencial de los productos farmacéuticos en el olfato en humanos, pero solo se han estudiado esteroides, antibióticos y agentes anestésicos en el canino. Los factores estresantes físicos, como el ejercicio, la falta de acondicionamiento y la alta temperatura ambiente, afectan el olfato directa o indirectamente en el canino. Se ha demostrado que el contenido de grasa en la dieta, la cantidad de alimentos por comida y el horario de las comidas afectan el olfato en ratones y perros. La microbiota gastrointestinal (GI) probablemente afecta el olfato a través de la comunicación bidireccional entre el tracto gastrointestinal y el cerebro, y la microbiota se ve afectada por el ejercicio, la dieta y el estrés. El objetivo de esta revisión de la literatura es discutir los efectos específicos de la salud, el manejo y los cambios de microbiota en el rendimiento olfativo en caninos de trabajo.
Introducción
La extraordinaria capacidad olfativa del canino ha sido utilizada durante mucho tiempo por los humanos para la identificación y discriminación de olores (1). Se ha informado que la capacidad del canino para la detección de olores es de 10,000 a 100,000 veces mayor que la del humano promedio, y el límite inferior canino de detectabilidad para compuestos orgánicos volátiles es una parte por billón (ppt) (2). Este sentido elevado da a los caninos la capacidad de detectar una gran cantidad de compuestos químicos que contienen moléculas que muestran diferencias sutiles en las estructuras estereoisoméricas (3). Esta sensibilidad, la capacidad única de detectar un olor objetivo entre una miríada de olores en un entorno operativo (4) y la capacidad del perro para aprender mediante el condicionamiento operante (5) ha hecho del canino de trabajo un componente intrínseco de las funciones policiales, militares, de búsqueda y rescate, médicas y de asistencia / servicio en todo el mundo. Sin embargo, a pesar de la naturaleza crítica del servicio que brindan nuestros socios caninos, la evidencia relacionada con la salud y el rendimiento olfativo está subrepresentada en la literatura científica. El objetivo de esta revisión es discutir los efectos de las decisiones de manejo relacionadas con la dieta y el acondicionamiento físico, la atención médica y los cambios en la microbiota sobre el rendimiento olfativo en caninos de trabajo.
Salud y enfermedad
Anatomía del olfato
Para manejar adecuadamente la salud del perro de detección, uno debe comprender la anatomía y fisiología asociada con el olfato. Los principales componentes del sistema olfativo son la cavidad nasal, el epitelio olfativo y los receptores, el órgano vomeronanasal (VNO) y el bulbo olfatorio. La cavidad nasal se compone de dos cámaras separadas por el tabique nasal, que están altamente vascularizadas, suministradas principalmente por la arteria esfenopalatina. Cada cámara de la cavidad nasal contiene tres cornetes (nasocornato, maxilocornete y etmocornete) (6) que contribuyen al aumento de la superficie mucosa. Sin embargo, el área total de la superficie mucosa puede estar fuertemente influenciada por el tamaño y la forma del hocico en el canino (7). Los cornetes nasales se proyectan desde las paredes laterales de la cámara y contienen una red de venas tortuosas. Medial y dorsal a los cornetes es la hendidura olfativa, donde se desvía el 5-15% del aire inhalado, y múltiples nervios craneales terminan. A medida que se produce la inhalación, el aire llega primero al maxilocorneto donde hay un pequeño número de neuronas sensoriales olfativas. El aire continúa fluyendo hacia los etmocornetes y los senos paranasales y luego se dirige hacia la faringe (6). La congestión de cornetes altera el flujo de aire en la hendidura olfativa, afectando el olfato. La congestión de cornetes se reduce con el ejercicio, la hipercapnia y el aumento del tono simpático, mientras que aumenta con el aire frío, los irritantes químicos, la hipocapnia y el aumento del tono parasimpático. Algunos olores / productos químicos en el aire pueden estimular las terminaciones nerviosas libres del trigémino en la mucosa nasal, que causan sensaciones como calor, frescura, nitidez, pero no olor (8). La detección del olor ocurre solo a través del epitelio olfativo y los nervios olfativos.
El epitelio olfativo está compuesto por neurepitelio que recubre la placa cribiforme, el tabique dorsal, los cornetes dorsales y medios, y el epitelio columnar pseudoestratificado, con millones de células receptoras olfativas (OR) (ORC). El epitelio olfativo también contiene células sustentaculares de soporte que regulan la composición de la mucosa nasal, sirven como aislantes entre los ORC y protegen el epitelio del daño de los agentes inhalados (9). La capa mucosa de la mucosa nasal se deriva de las glándulas de Bowman incrustadas en el epitelio olfativo; Esta capa mucosa mantiene los niveles normales de humedad nasal y atrapa los odorantes (10). La percepción olfativa normal depende de esta área receptora húmeda (9).
Las células receptoras olfativas se proyectan directamente al bulbo olfatorio, con axones que terminan en los glomérulos del bulbo olfatorio (11). Los ORCs tienen cilios que tienen receptores de olor superficial; Los ORC humanos tienen aproximadamente 25 cilios por ORC, pero los perros tienen cientos de cilios por ORC, lo que permite la detección de concentraciones significativamente más pequeñas de odorantes en los caninos. Hay más de 220 millones de quirófanos en la cavidad nasal canina, que permiten que una gran cantidad de olores se unan (12). Solo hay un tipo de OR por ORC, y la intensidad del olor es proporcional al número de ORC activados; ORC también tienen receptores para hormonas y neurotransmisores. Las neuronas olfativas solo viven de 30 a 60 días, pero a diferencia de otras células sensoriales de mamíferos, las ORC se regeneran constantemente (13). El número y el tipo de ORC presentes en un perro individual están dictados por la raza, la genética y el entrenamiento (7, 14-17); Este concepto será explorado más adelante en el manuscrito.
Incrustados en la membrana de los cilios ORC hay porciones extracelulares que se unen al odorante, y porciones intracelulares acopladas a la proteína G. Cuando un odorante se une a la porción extracelular del receptor, la subunidad A de la proteína G se rompe, activando la adenilciclasa, que posteriormente convierte el ATP en cAMP. cAMP amplifica la señal entrante del odorante activando múltiples canales dependientes de sodio (11). La apertura de dos pasos de los canales de sodio cerrados causa despolarización, y el potencial de acción resultante se transmite a través del bulbo olfatorio. Cada odorante se reconoce por una combinación única de quirófanos activados (18). La capacidad del perro de detección para reconocer adecuadamente los olores se basa en esta función.
El VNO se encuentra a lo largo de la cara ventrorostral del tabique nasal, es bilateralmente simétrico y actúa como un sitio adicional de detección de olores (19). Las neuronas sensoriales VNO detectan señales químicas que estimulan cambios conductuales y/o fisiológicos (20), proporcionan una vía neuronal alternativa al hipotálamo y son muy lentas para adaptarse a los olores. El VNO contiene epitelio receptor y epitelio no receptor, que difieren estructuralmente en los tipos de fibras nerviosas y tipos de células incrustadas (21). El VNO funciona en la detección de olores no volátiles, especialmente feromonas, y se cree que desempeña un papel en el comportamiento social y la reproducción (21).
El bulbo olfatorio es una estructura pareada, que funciona principalmente como una estación de relevo y para filtrar la entrada sensorial (6). Hay aproximadamente 1.000 axones ORC por neurona de segundo orden, lo que resulta en una amplificación significativa de la señal de olor. Las células mitrales del bulbo olfatorio proyectan una dendrita primaria a un glomérulo y un axón a la corteza olfativa. El bulbo olfatorio se encuentra debajo de los lóbulos frontales, por encima de la placa cribiforme en los seres humanos, pero se encuentra más rostralmente en otros mamíferos, lo que puede desempeñar un papel en la mejora del olfato en los mamíferos inferiores (19). La corteza olfativa se encuentra dentro de los lóbulos temporales mediales y se comunica directamente con la corteza cerebral. La corteza olfativa funciona para recibir información sensorial del bulbo olfatorio, permite la conciencia consciente del olor, la identificación del olor, la memoria del olor y la localización del olor en los mamíferos inferiores. El bulbo olfatorio tiene un papel sensorial (procesamiento inicial de la información olfativa) y un papel modulador en el cerebro anterior, el hipotálamo y el sistema límbico (22). La vía olfativa de los caninos se demuestra en las Figuras 1 y 2.
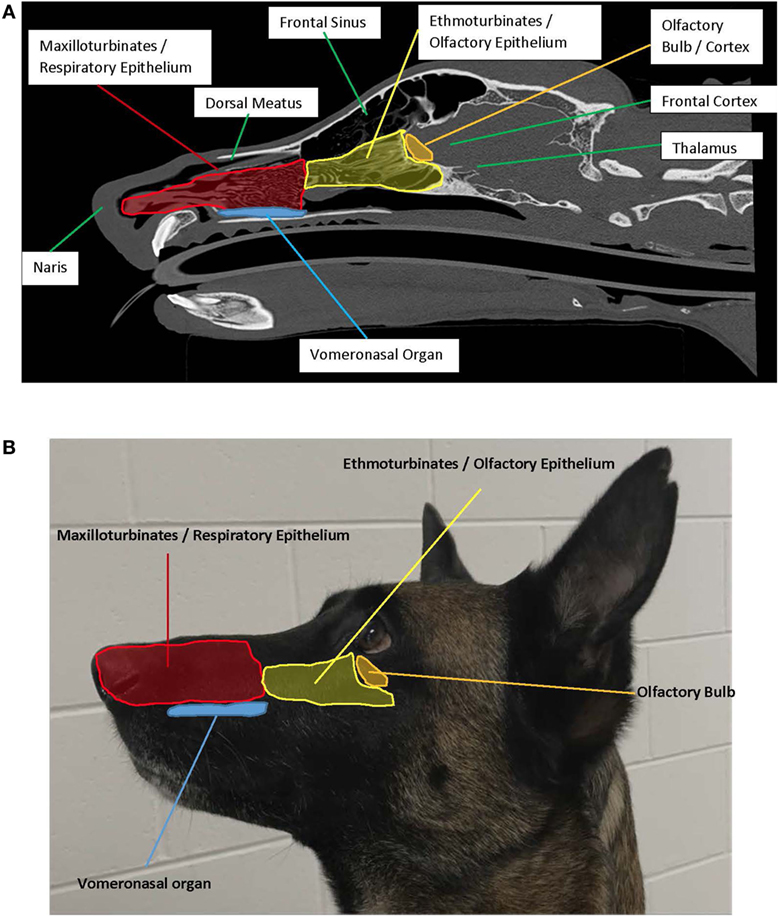 FIGURA 1. (A) Plano sagital izquierdo destacando la anatomía asociada al olfato. Crédito de la foto: Adrien-Maxence Hespel, Universidad de Tennessee. (B) Vista exterior izquierda que muestra la colocación de estructuras interiores asociadas con el olfato.
FIGURA 1. (A) Plano sagital izquierdo destacando la anatomía asociada al olfato. Crédito de la foto: Adrien-Maxence Hespel, Universidad de Tennessee. (B) Vista exterior izquierda que muestra la colocación de estructuras interiores asociadas con el olfato.
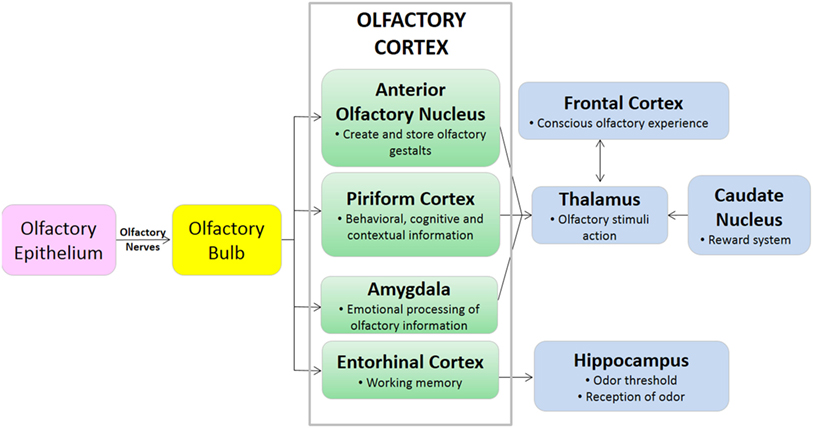 FIGURA 2. Diagrama de la vía que muestra el proceso de señalización olfativa.
FIGURA 2. Diagrama de la vía que muestra el proceso de señalización olfativa.
Las áreas cerebrales olfativas del cerebro se dividen en dos categorías funcionales: la neocortical (por ejemplo, complejo orbitofrontal) que regula la percepción consciente del olor, y la límbica (23). El sistema límbico es una colección de estructuras cerebrales que regulan colectivamente el olfato, la memoria, el comportamiento y la motivación. Los componentes del sistema límbico incluyen el bulbo olfatorio, el hipocampo, la amígdala y el complejo entorrinal, entre otros. El tamaño y la función del sistema límbico varían según las especies de mamíferos, pero en todas las especies el sistema límbico tiene componentes olfativos y no olfativos (24). La isocorteza del cerebro regula funciones de orden superior como la percepción sensorial y la cognición. Mientras que los primates, incluidos los humanos, tienen una relación inversa entre el isocórtex y el volumen del sistema límbico, los carnívoros terrestres, incluidos los caninos, tienen altos volúmenes relativos tanto del isocórtex como del sistema límbico (24). Estas diferencias anatómicas en los volúmenes de los componentes cerebrales pueden ser parcialmente responsables de las diferencias en la capacidad olfativa entre humanos y caninos.
Fisiología del olfato
En comparación con los humanos, los perros tienen un área de superficie significativamente mayor de epitelio olfativo, con aproximadamente un 30% más de quirófanos que pueden reconocer una variedad mucho mayor de olores. Los perros también tienen la capacidad de una excelente localización del olor, incluso en presencia de un olor de fondo significativo, probablemente debido al mayor tamaño de la cavidad nasal en comparación con otras especies (25) y los patrones únicos de flujo de aire creados por el olfateo (26). La capacidad de encontrar la fuente del olor, incluso en presencia de olores competitivos, hace que el perro de detección sea un socio crítico en muchas operaciones militares, policiales y de búsqueda y rescate.
Durante la inspiración, el 12-13% del flujo de aire viaja a la porción olfativa de la nariz, y el flujo de aire restante se dirige hacia la nasofaringe donde sale de la cavidad nasal (26). Los perros han mejorado el muestreo del flujo de aire y la recolección de olores a través de la inhalación activa, que es la producción de respiraciones cortas y agudas a 4-7 Hz, independientemente del tamaño del cuerpo canino (26). El perro promedio inhala 30 ml de aire por fosa nasal por inhalación (19), y el aire se inhala desde el frente y se exhala hacia un lado como se ve en la Figura 3; Esto permite un muestreo más eficiente de los odorantes. Cuando un canino está olfateando, el aire dentro de aproximadamente 1 cm de la fosa nasal se atrae hacia la narina (26), y el flujo de aire de alta velocidad se transporta a la cavidad nasal dorsal donde gira 180 ° y fluye hacia atrás sobre los etmocornetes. Cada fosa nasal toma muestras de aire por separado, produciendo muestras de olor bilateral que ayudan en la localización de la fuente de olor (26). A diferencia de los humanos y otras especies microsmóticas, el aire no entra ni sale del receso olfativo del perro durante la espiración, lo que resulta en una exposición prolongada de aire inspirado a los quimiorreceptores del epitelio olfativo y una estimulación olfativa continua durante todo el ciclo respiratorio (26). Para el canino de trabajo, la exploración activa durante la búsqueda de «nariz hacia abajo, cola hacia arriba» (ver Figura 4) y la localización eficiente de la fuente de olor son fundamentales para completar la misión.
 FIGURA 3. Vías para la inhalación y exhalación y flujo de aire asociado con el olfato (flechas azules = inhalado; flechas rojas = exhalado). Los caninos usan preferentemente la fosa nasal derecha para oler olores de excitación conespecíficos y olores novedosos, y la fosa nasal izquierda para oler olores familiares, estímulos no aversivos y olores de excitación heteroespecíficos.
FIGURA 3. Vías para la inhalación y exhalación y flujo de aire asociado con el olfato (flechas azules = inhalado; flechas rojas = exhalado). Los caninos usan preferentemente la fosa nasal derecha para oler olores de excitación conespecíficos y olores novedosos, y la fosa nasal izquierda para oler olores familiares, estímulos no aversivos y olores de excitación heteroespecíficos.
 FIGURA 4. Caninos de desastres que realizan trabajos de búsqueda que muestran la postura típica de «nariz abajo, cola hacia arriba» asociada con el olfato activo. Crédito de la foto a Tracy Darling.
FIGURA 4. Caninos de desastres que realizan trabajos de búsqueda que muestran la postura típica de «nariz abajo, cola hacia arriba» asociada con el olfato activo. Crédito de la foto a Tracy Darling.
Las condiciones ambientales, como la humedad relativa y la presión barométrica, pueden tener impactos directos en el olfato, además de los impactos que esos factores tienen en la generación y el movimiento del olor en sí. Philpott et al. (27) informaron que los umbrales olfativos en humanos eran independientes de la temperatura ambiente, la humedad máxima y el flujo nasal inspiratorio máximo. Un estudio posterior y más amplio reportado por Kuehn et al. (28) determinó posteriormente que el nivel umbral olfativo se vio afectado en condiciones hipobáricas, y los umbrales olfativos fueron más bajos (el sentido del olfato mejoró) en un ambiente húmedo. Los perros de búsqueda y rescate funcionan mejor cuando la humedad relativa es alta (29), posiblemente debido a la mejora de la humedad nasal y la captura de olores. La humedad, pero no la lluvia, aumentó la eficiencia de los perros en las tareas de seguimiento y búsqueda al aumentar la intensidad del olor (30) y mejoró la detección olfativa de feromonas, lo que llevó a una mayor actividad de apareamiento durante la temporada del monzón (31).
La inhalación es ventajosa en comparación con la inhalación normal porque produce un flujo laminar unidireccional hacia el meato dorsal y el epitelio sensorial de los etmocornetes (26, 32), aumenta la sensibilidad a los olores (32), impulsa la actividad en la corteza olfativa y afecta la intensidad e identificación del olor (33). Los patrones de flujo de aire nasal descritos por Craven et al. (26) mejoran la agudeza olfativa en el perro, pero no explican completamente la macrosmia, la capacidad mejorada de oler, en el canino. Lawson et al. (34) describieron el transporte de tipos específicos de odorantes y el impacto posterior en el olfato. Los patrones de deposición de olores corresponden a la organización anatómica de las neuronas OR: los odorantes altamente solubles se depositan en la parte frontal de la hendidura olfativa (meato dorsal y tabique nasal), mientras que los odorantes moderadamente solubles o insolubles se depositan en toda la hendidura olfativa (34). Esta combinación de organización anatómica de las neuronas OR y los patrones de flujo de aire inducidos durante el olfateo son probablemente responsables de la macrosmia ampliamente demostrada en los caninos de trabajo. Los caninos se mueven más lentamente y el período de olfateo dura tres veces más durante la fase decisiva del seguimiento olfativo (el «hallazgo»), en comparación con la fase inicial de búsqueda y las fases de seguimiento (35). Concha et al. (36) demostraron que los patrones de olfateo en caninos de trabajo pueden usarse para diferenciar las respuestas negativas verdaderas de las falsas negativas. Los perros entrenados para detectar olores pasaron significativamente menos tiempo olfateando muestras negativas verdaderas (sin olor; sin respuesta de alerta), con solo un episodio de olfateo para muestras negativas verdaderas (36). Para los perros de detección, estas características de olfateo pueden resultar en un trabajo de detección más eficiente durante un ciclo de trabajo largo.
Los perros han demostrado repetidamente la «especialización hemisférica», es decir, el procesamiento cerebral específico del hemisferio de estímulos emocionales, acústicos y olfativos (37, 38). A diferencia de otros sentidos, las vías olfativas ascienden desde el punto de detección (cavidad nasal) hasta el punto de percepción en el cerebro (corteza olfativa) ipsilateralmente: la entrada sensorial de la fosa nasal derecha se entrega al hemisferio derecho del cerebro y la entrada sensorial de la fosa nasal izquierda se entrega al hemisferio izquierdo (37). Los caninos usan preferentemente la fosa nasal derecha para oler olores de excitación conespecíficos y olores novedosos, entregando información sensorial al hemisferio cerebral derecho, que procesa estímulos amenazantes y alarmantes. Los caninos usan preferentemente la fosa nasal izquierda para olfatear olores familiares y estímulos no aversivos como alimentos, así como olores de excitación heteroespecíficos (como muestras de sudor inducidas por el miedo humano) (37). D’Aniello et al. demostraron que la especialización hemisférica y la quimioseñalización mejoran la comunicación de los estados emocionales (incluido el estrés) entre perros y humanos (39). Para los perros de detección, esto probablemente significa que los olores objetivo se procesan a través de la fosa nasal izquierda.
Las conexiones anatómicas entre las vías olfativas de la amígdala y la corteza piriforme y el sistema límbico subyacen a la interconexión entre el olfato y la memoria (9). El olfato y otras formas de aprendizaje/memoria están reguladas por las mismas reglas neurobiológicas (40). En los caninos de trabajo, la memoria del olfato es de importancia crítica: ¿cuándo comienza la memoria del olor? ¿Cuántos olores pueden recordar los caninos? ¿Cuánto tiempo recuerdan los caninos los olores entrenados? ¿Cuánto tiempo puede un perro mantener el rendimiento olfativo sin entrenamiento? Los caninos aprenden el olor a partir del período prenatal, debido a la influencia de la dieta materna en la composición del líquido amniótico (41), pero la memoria de olor aprendida parece disiparse a las 10 semanas de edad (42). Se ha demostrado que el olfato y la cognición se deterioran con la edad en el canino, pero no existe una edad específica en la que el perro deje de aprender (40). Williams y Johnston (43) determinaron que los caninos podían aprender fácilmente y posteriormente identificar 10 olores en una tarea de búsqueda. Dado que los caninos domésticos han demostrado la capacidad de aprender y recordar más de 200 palabras (44) y los nombres de más de 2,000 juguetes (45), es probable que los caninos de trabajo puedan recordar mucho más de 10 olores. La durabilidad de la memoria en olores entrenados no ha sido ampliamente estudiada, pero Johnston (46) demostró que en los caninos de detección de explosivos no hubo un deterioro sistemático en el rendimiento de detección durante un máximo de 4 meses. Los métodos de entrenamiento pueden afectar la durabilidad de la memoria de olor, o al menos la señalización del canino al manejador cuando se detecta un olor específico. Si las alertas no se refuerzan, o si el canino realiza varias búsquedas sin detectar un olor entrenado, la alerta o el comportamiento de búsqueda pueden extinguirse (47). Sin embargo, no está claro si la memoria se mantiene cuando se extingue el comportamiento de alerta o búsqueda o cuál es la duración máxima del tiempo que un canino puede mantener la memoria del olor sin entrenamiento (47).
La genética se reconoce cada vez más como un componente crítico del olfato en los caninos, con una revisión exhaustiva publicada en otros lugares (7). Hay cuatro tipos de receptores involucrados con el olfato y la quimiosensación en el perro: OR, receptores vomeronasales, receptores asociados a aminas traza y proteínas similares al receptor del péptido formilo. La mayoría de las investigaciones sobre la genética del olfato canino se han centrado en los genes OR. El repertorio canino OR está compuesto por 1.094 genes, aproximadamente tres veces más que un humano. Se cree que este gran repertorio genético está relacionado con la macrosmia evidente en los caninos, produciendo una amplia gama de OR que permiten la detección de amplias gamas de odorantes (48). En el canino, aproximadamente el 20% de los genes OR son pseudogenes funcionalmente inactivos, pero el porcentaje de pseudogenes varía según la raza y es significativamente menor que las especies microsmóticas como los humanos, en las que hasta el 50% de los genes olfativos son pseudogenes (49). El polimorfismo de los genes OR también puede afectar la capacidad olfativa y la sensibilidad en razas e individuos. Tacher et al. (15) informaron que tanto el porcentaje de pseudogenes como la frecuencia de polimorfismo genético específico variaban según la raza, y especularon que los cambios genéticos pueden contribuir a las diferencias en las capacidades olfativas entre razas e individuos. Esto puede ofrecer una idea de las «líneas de trabajo» dentro de algunas razas que tienden a producir frecuencias más altas de caninos de detección exitosos que otras.
La literatura actual contiene información contradictoria sobre la capacidad olfativa específica de la raza. Jezierski et al. (50) demostraron que los pastores alemanes eran significativamente mejores en la detección de narcóticos que los labradores y terriers. En contraste, Hall et al. (5) informaron que los Pugs superaron consistentemente a los pastores y galgos alemanes en tareas de adquisición olfativa y discriminación. Polgar et al. (51) informaron que los perros de «grupo olfativo» (por ejemplo, basset hound, puntero alemán, etc.) se desempeñaron mejor en una tarea de detección natural que los perros «sin olor» (por ejemplo, galgo inglés, sabueso afgano, etc.) y perros de «nariz corta» (por ejemplo, Cavalier King Charles spaniel, Boston terrier, etc.). Se necesita investigación adicional para determinar si las capacidades olfativas específicas de la raza están correlacionadas con el polimorfismo genético o si el rendimiento olfativo es más una función de atributos conductuales como la motivación inherente (es decir, el impulso) y la capacidad de entrenamiento.
Hiposmia: enfermedad y medicación
La hiposmia, definida como disminución de la sensación del olfato, se caracteriza como tipo I, II o III. La hiposmia tipo I es la incapacidad de reconocer los olores correctamente. La hiposmia tipo II es una disminución cuantitativa en la capacidad de reconocer odorantes, reconocida en caninos de trabajo como cambio en el umbral o falta persistente de alertar a odorantes previamente entrenados. La hiposmia tipo III es una disminución en la estimación de la magnitud de los olores; Este tipo de hiposmia solo es reconocible en humanos. Las causas de la hiposmia pueden clasificarse ampliamente como trastornos conductivos, pérdidas sensoriales o causas neuronales (52). La hiposmia conductiva resulta de la incapacidad de los odorantes para llegar a la mucosa olfativa, por ejemplo, inflamación nasal, exceso de producción de moco y obstrucción física por masas que ocupan espacio (pólipos, neoplasia, etc.) (53). La hiposmia sensorial es causada por daño a la mucosa olfativa, por ejemplo, virus, exposición a sustancias químicas tóxicas o agentes industriales, y neoplasia (54-56). La hiposmia neural es causada por lesiones del sistema nervioso central o periférico, específicamente la corteza olfativa, el bulbo olfatorio y los nervios craneales I (olfativo) y V (trigémino), por ejemplo, lesión en la cabeza (57).
Enfermedad
El impacto de la enfermedad en el olfato ha sido ampliamente documentado en la medicina humana. De hecho, «la degradación en el sentido del olfato es una condición centinela, particularmente para enfermedades neurodegenerativas como el Alzheimer» (58). Las afecciones asociadas con hiposmia o anosmia en humanos incluyen trastornos congénitos (por ejemplo, síndrome de Kallaman), trastornos endocrinos o metabólicos, infecciones, inflamación, trastornos neurológicos que incluyen traumatismo craneal, procesos locales, exposición ocupacional al polvo y productos químicos y materiales tóxicos, edad avanzada y uremia (8, 57, 59–61). La hiposmia y la anosmia son frecuentemente autoinformadas en la medicina humana, pero el autoinforme puede representar de forma exagerada la prevalencia real de hiposmia en humanos. La prevalencia de hiposmia en caninos es desconocida, pero la hiposmia en caninos de detección podría ser catastrófica.
Cuando un canino está expuesto a temperaturas corporales prolongadas y aumentadas debido a extremos ambientales o de esfuerzo sin la capacidad de aclimatarse adecuadamente, puede ocurrir una lesión térmica (62). La termorregulación se ve comprometida durante el estrés por calor; El canino aumenta la frecuencia respiratoria (es decir, jadeo) y la frecuencia cardíaca para compensar y enfriar el cuerpo. El jadeo disminuye la eficiencia olfativa en los caninos y puede conducir a hiposmia relevante. Los factores exógenos que pueden contribuir al estrés térmico y aumentar la probabilidad de comprometer el olfato incluyen la falta de aclimatación a un ambiente nuevo, la humedad elevada, la falta de acceso al agua y la mala ventilación (63). La lesión por calor probablemente conduce a un compromiso olfativo, pero se desconoce la magnitud y la duración del olfato alterado; Se indican estudios adicionales.
Al igual que en los humanos, la enfermedad puede afectar el olfato en los caninos. Aunque hay investigaciones caninas limitadas disponibles, la infección viral con moquillo canino (64) y las infecciones por el virus de la parainfluenza canina (65) han causado alteraciones en el olfato. El virus de la parainfluenza canina aumentó la inflamación nasal y las secreciones mucosas, causando una hiposmia conductora al reducir el contacto entre los odorantes y los receptores olfativos o del trigémino en la cavidad nasal. Además, la inflamación nasal, como la causada por el moquillo canino o los virus de la parainfluenza, conduce a la congestión vascular en la mucosa respiratoria, alterando los patrones de flujo de aire en la cavidad nasal (65). También se ha informado que la enfermedad endocrina (p. ej., hiperadrenocorticismo, diabetes mellitus e hipotiroidismo) y la enfermedad neurológica (p. ej., meningoencefalitis granulomatosa y tumores nasales) causan hiposmia en caninos (55); El mecanismo exacto de la hiposmia en estos estados de enfermedad no se conoce, pero es probable que sea neural. Recientemente, la vomeronasalitis se asoció con agresión intraespecífica en gatos (66). Asproni teorizó que la inflamación presente en el VNO y la cavidad nasal deterioró la función del epitelio sensorial y la comunicación intraespecífica, pero no examinó la función olfativa en los gatos estudiados. Dada nuestra comprensión de la VNO y la fisiología nasal, es probable que la vomeronasalitis cause hiposmia sensorial y comunicación intraespecífica interrumpida en gatos y posiblemente en perros. El trauma es una causa bien documentada de hiposmia neural en las personas, pero el impacto del traumatismo craneal en el olfato en los perros aún no se ha estudiado. Si un canino de detección experimenta traumatismo craneal asociado con falta de conciencia, se indica la evaluación del olfato (67). La función olfativa disminuye con la edad en los seres humanos a través de una variedad de mecanismos que incluyen alteración de la congestión nasal, daño acumulativo al epitelio olfativo, disminución de las enzimas mucosas, pérdida de selectividad de ORCs y enfermedad neurodegenerativa (68). Se encontraron cambios similares relacionados con la edad en el sistema olfativo de perros mayores de 14 años y fueron prominentes en perros mayores de 17 años (69). Los perros más viejos tuvieron una disminución en el número de ORCs y el número de cilios en ORCs. Curiosamente, los perros mayores demostraron cambios cerebrales seniles como amiloidosis cerebrovascular en el bulbo olfatorio, pero no en la mucosa olfativa. La hiposmia inducida por la enfermedad, pero no inducida por la edad, en humanos es generalmente reversible, posiblemente porque las neuronas olfativas se regeneran fácilmente, pero no se puede predecir la duración de la hiposmia y la normalización de la función (54, 68); Esto también es probablemente cierto en los caninos.
Productos farmacéuticos
La hiposmia tipo II es común en humanos durante o después de la terapia farmacéutica (70); La hiposmia suele ser bilateral y temporal. La lista de productos farmacéuticos conocidos por inducir hiposmia en humanos es larga, incluyendo: anestésicos, antiarrítmicos, antihistamínicos, antimicrobianos, medicamentos antiproliferativos e inmunosupresores, medicamentos endocrinos, medicamentos gastrointestinales (GI), medicamentos neurológicos y AINE (8, 57, 59). Los productos farmacéuticos con frecuencia causan hiposmia a través del deterioro de la unión del olor al quirófano o lesión al quirófano (hiposmia sensorial), o a través del deterioro neurológico (hiposmia neural).
La información más relevante sobre los productos farmacéuticos que afectan el olfato canino se extrapola de la medicina humana. El metabolismo del zinc está directamente relacionado con la función olfativa tanto en humanos como en animales de laboratorio. Las nanopartículas de zinc, cuando se agregan a los explosivos, mejoran la respuesta odorante en caninos entrenados de una manera dependiente de la dosis (70). La quelación del zinc, sin embargo, causa hiposmia sensorial a nivel de quirófano. Algunos fármacos cardiovasculares como los inhibidores de la enzima convertidora de angiotensina (ECA-I) (por ejemplo, captopril) quelan zinc y causan hiposmia en humanos (8); Este efecto no ha sido estudiado en caninos. Se ha documentado que los anestésicos causan hiposmia en humanos; el impacto en el olfato en caninos se está investigando actualmente en la Universidad de Auburn.
Los antimicrobianos como el metronidazol y la doxiciclina se recetan comúnmente a los caninos que trabajan para tratar la diarrea y las enfermedades transmitidas por vectores, respectivamente. Se ha informado que el metronidazol causa hiposmia en humanos (8) y disminuye el rendimiento olfato en caninos de detección (71). Se ha informado que la doxiciclina causa hiposmia en humanos (60), pero no causa hiposmia en caninos de detección (71). Jenkins et al. observaron que el 50% de los perros entrenados para la detección de explosivos demostraron una elevación en el umbral olfativo cuando se les administró metronidazol en dosis altas durante 10 días, pero la administración de doxiciclina en dosis estándar durante 10 días no afectó el olfato. La hiposmia inducida por metronidazol no se pudo predecir en función del sexo masculino o femenino, el estado de castración o la edad, pero la hiposmia fue temporal, ya que el umbral olfativo volvió a la normalidad dentro de los 10 días posteriores a la interrupción del metronidazol. Se deben considerar intervenciones médicas alternativas cuando sea apropiado antes del uso de metronidazol para perros de detección; Si se debe usar metronidazol, debe usarse a la dosis eficaz más baja durante el menor tiempo posible.
Los esteroides pueden causar hiposmia en humanos (8) y en caninos (72). Ezeh administró altas dosis de dexametasona o hidrocortisona combinada con desoxicorticosterona a perros de laboratorio y observó hiposmia sin signos clínicos aparentes después de 7 y 18 días de tratamiento, respectivamente. La hiposmia inducida por esteroides observada en perros se atribuyó a la elevación en el umbral de detección olfativa. Sin embargo, los estudios en humanos con inflamación nasal demostraron que la administración de esteroides orales y/o intranasales a veces mejoró el olfato, probablemente debido a la resolución de la inflamación nasal (73-75). Por lo tanto, los veterinarios y los cuidadores caninos deben sopesar cuidadosamente la necesidad clínica de esteroides contra los efectos potenciales sobre el olfato. Los mecanismos de la hiposmia inducida por productos farmacéuticos incluyen el deterioro de la unión del olor a través de la alteración de la cantidad o calidad del moco (por ejemplo, antihistamínicos), la inhibición del recambio / regeneración normal de las neuronas olfativas (por ejemplo, esteroides y quimioterápicos), vasoconstricción nasal (por ejemplo, descongestionantes), efectos enzimáticos asociados de los medicamentos (ECA-I), niveles alterados de GMP cíclico (bloqueadores de la fosfodiesterasa) y quelación del zinc (medicamentos cardíacos) (76).
Dada la escasez de investigación sobre la hiposmia inducida por productos farmacéuticos en caninos, los manejadores, entrenadores y veterinarios que cuidan a los perros de detección deben tener precaución con los productos farmacéuticos que se sabe que causan hiposmia en humanos. También es importante considerar qué medicamentos pueden ser biotransformados por la microbiota gastrointestinal cuando se habla de atención médica para caninos que trabajan. Se debe considerar la información sobre la reducción, las reacciones hidrolíticas y otras reacciones químicas para los medicamentos comúnmente prescritos y sus impactos asociados en la microbiota y el olfato. El umbral olfatorio y la capacidad de discriminación deben probarse en cualquier perro de detección que haya sido tratado con productos farmacéuticos inductores de hiposmia antes de regresar al trabajo.
Administración
Hay una miríada de factores que pueden mejorar o comprometer el rendimiento de los caninos de trabajo. La frecuencia, la intensidad y la duración de los ciclos de trabajo deben considerarse antes de tomar decisiones de gestión, particularmente en lo que respecta a la agudeza olfativa. Los perros de detección (explosivos, narcóticos, búsqueda y rescate) son diferentes de los perros deportivos (agilidad, caza, trineo) y se miden con criterios de rendimiento muy diferentes (ver Tabla 1).
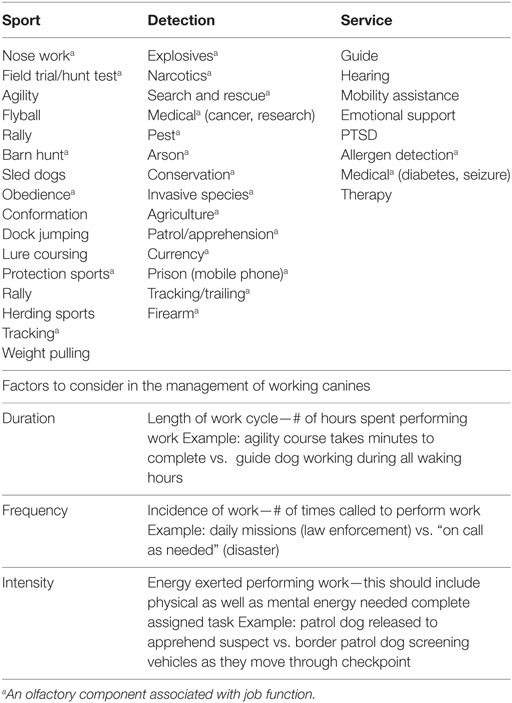 TABLA 1. Categorías de caninos de trabajo y disciplinas típicas asociadas a cada uno.
TABLA 1. Categorías de caninos de trabajo y disciplinas típicas asociadas a cada uno.
Las condiciones que pueden alterar el potencial de trabajo de un perro incluyen la cría y selección, la aptitud y el acondicionamiento regulares, y el desarrollo de un régimen dietético que cumpla con los requisitos de nutrientes y utilice ingredientes de calidad. Maximizar la función olfativa debe ser primordial en las decisiones relacionadas con los perros de detección. En la Tabla 2 se presenta un resumen de las publicaciones seleccionadas asociadas con el rendimiento canino de trabajo.
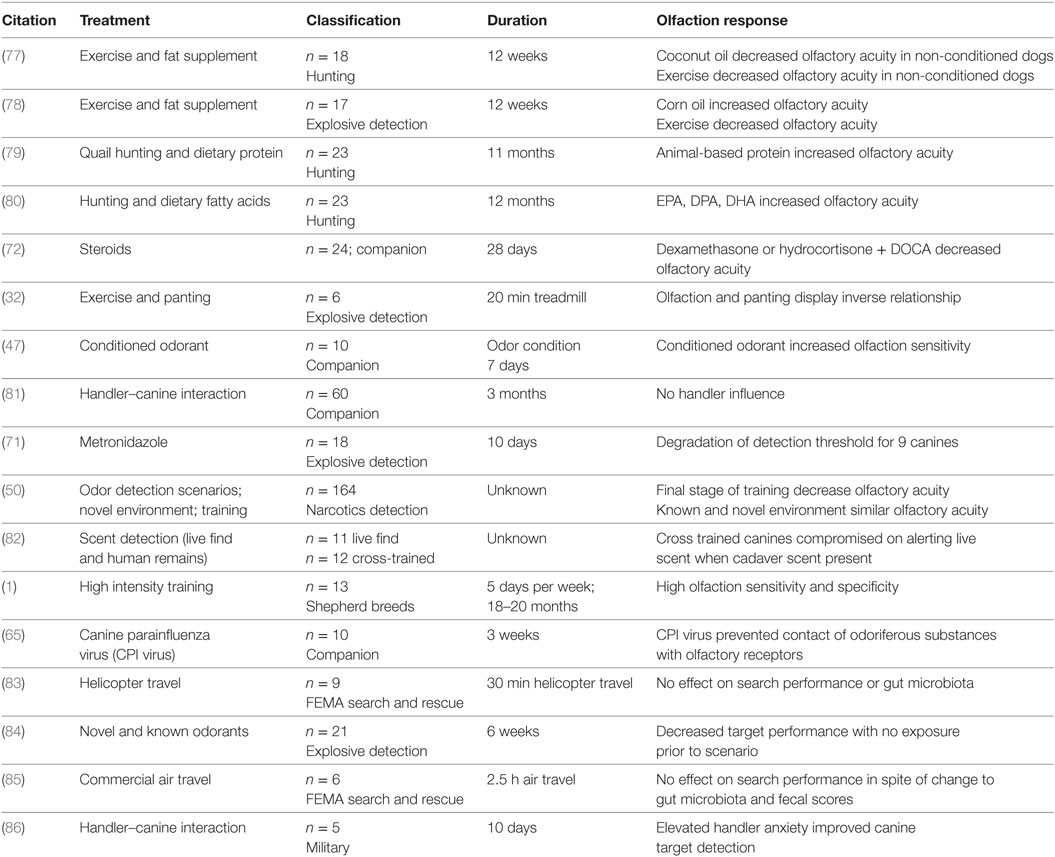 TABLA 2. Resumen de los estudios seleccionados que informan los efectos sobre el olfato/rendimiento asociados con el tratamiento o la atención médica.
TABLA 2. Resumen de los estudios seleccionados que informan los efectos sobre el olfato/rendimiento asociados con el tratamiento o la atención médica.
Acondicionamiento y entrenamiento
Como era de esperar, el entrenamiento y el acondicionamiento fisiológico afectan significativamente el rendimiento olfativo. Se ha informado de una disminución de las tasas de búsqueda utilizando perros de detección certificados en ruedas aromáticas después del ejercicio (78); Esto probablemente se explica por el aumento del jadeo que generalmente ocurre después del ejercicio. Los caninos que fueron condicionados físicamente mantuvieron una mayor agudeza olfativa en comparación con los caninos que no fueron condicionados físicamente cuando ambos grupos fueron desafiados con ejercicio. Los caninos no condicionados mostraron una disminución del 63,6% en la sensibilidad olfativa después del ejercicio (78). Los caninos físicamente acondicionados tienen una frecuencia cardíaca de ejercicio más baja en comparación con sus contrapartes no condicionadas y esta condición cardiovascular mejorada puede contribuir a un mejor rendimiento termorregulador y, posteriormente, disminuir la necesidad de jadear (32). Otros trabajos de apoyo han demostrado que un programa de entrenamiento riguroso conduce a altas frecuencias de alertas de objetivos correctos (32). Inmediatamente después del ejercicio físico extremo, hay una reducción en la tasa de olfateo y un aumento de la tasa de jadeo que resultan en un rendimiento olfativo reducido (32). Esto puede explicarse por el hecho de que los caninos no condicionados jadean más fuerte durante el ejercicio intenso en lugar de respirar por la nariz, lo que disminuye la cantidad de odorantes que pasan sobre el epitelio olfativo en la cavidad nasal (77). Parece claro que el acondicionamiento físico (específicamente en lo que respecta a minimizar el jadeo) puede apoyar un mejor olfato en el perro de detección.
Las técnicas de entrenamiento de detección de olores también pueden afectar directamente el olfato. Wang et al. (16) y Youngentob y Kent (17) demostraron que los perros desarrollan más quirófanos para los odorantes con los que son entrenados regularmente. Gerritsen y Hank (14) también informaron que la renovación celular de ORC no es estática: el nuevo tipo de ORC de reemplazo se desencadena por olores familiares. Los odorantes simples y los odorantes complejos inducen diferentes respuestas neuronales en perros detectores de olores. Wilson y Stevenson (87) teorizaron que la plasticidad sináptica cortical se ve reforzada por la experiencia con odorantes (simples o complejos) en una variedad de condiciones. Gerritsen y Hank (14) sugirieron además que los perros aprenderán olores complejos más rápidamente si primero son entrenados en componentes individuales del olor, pero los resultados varían entre los estudios. Fischer-Tenhagen et al. (88) encontraron que los perros de detección entrenados con mezclas de olor que contenían el olor objetivo tenían indicaciones más correctas cuando el olor objetivo se probaba en un nuevo contexto, que los perros entrenados en un olor de referencia puro. Estos datos proporcionan evidencia científica para el concepto de entrenamiento tradicional de «probar» a los perros de detección con el uso de elementos de distracción. La resonancia magnética funcional del sistema olfativo en perros de olor entrenados indicó que la concentración de olor afecta la activación cerebral: la baja concentración de olor resultó en la activación cerebral unilateral, mientras que la alta concentración de olor resultó en la activación cerebral bilateral (58). Además del tipo de olor y la frecuencia, las técnicas de entrenamiento afectan la sensibilidad y la discriminación del olfato. El acondicionamiento pavloviano mejoró significativamente la adquisición de olores (89) y mejoró la resistencia a los disruptores (90). Los sistemas de recompensa continua funcionaron mejor para adquirir un comportamiento, como aprender a discriminar un olor específico, y las recompensas intermitentes funcionaron mejor para mantener un comportamiento aprendido (40). Se necesita más investigación para determinar el impacto del entrenamiento del olor simple versus complejo, el impacto de la concentración de olores en el aprendizaje y la interacción de la genética y el entrenamiento en el rendimiento en perros de detección.
Hidratación
El manejo del perro de detección en el campo a menudo puede implicar la mitigación de la deshidratación y la fatiga. La deshidratación de la membrana mucosa nasal produce una disminución de la actividad enzimática y una disminución de la fluidez de la membrana, alterando la transducción de neuroseñales y la función del receptor odorante. Una combinación de disminución del flujo de aire y deshidratación de la capa mucosa puede disminuir significativamente las capacidades de detección de olores en el canino de trabajo (77). Se informó que la deshidratación en los caninos de búsqueda y rescate ocurrió en perros que trabajaban después de los ataques terroristas del 9/11 (91, 92), el terremoto de Haití (93) y el deslizamiento de tierra de Washington (94). Un estudio reciente examinó tres estrategias de intervención para la hidratación de los caninos (95). Se utilizaron caninos de inspección de vehículos de la patrulla fronteriza (búsquedas de alta frecuencia y baja intensidad) para investigar los beneficios del agua, la solución electrolítica oral o los líquidos subcutáneos para la rehidratación de los caninos que trabajan en condiciones de calor. Los autores no informaron beneficios claros para ninguna de las estrategias examinadas, pero notaron que el consumo voluntario de la solución electrolítica oral con sabor fue mayor en comparación con el agua sola. El aumento del consumo voluntario de líquidos contribuyó a mejorar la hidratación. No se observaron beneficios asociados con el uso de líquidos subcutáneos. Por el contrario, la hidratación con líquidos subcutáneos se asoció con un aumento de la creatinina que se observó que indicaba deshidratación o daño muscular potencial. Los autores no proporcionaron información sobre los regímenes dietéticos y los comportamientos registrados no se vieron afectados por la estrategia de hidratación. No se pudo cuantificar el olfato como medida del rendimiento; Las pruebas de olfato estandarizadas no fueron posibles debido a la naturaleza operativa del estudio de campo. Este estudio demuestra que la deshidratación en el campo es una preocupación que merece más investigación, especialmente cuando se considera en relación con los posibles desafíos olfativos.
La recuperación térmica se mejoró cuando se usó una dieta baja en proteínas aderezada con aceite de maíz en labradores ejercitados en cintas de correr (96). Los autores informaron temperaturas corporales centrales más bajas 10 y 20 minutos después del ejercicio y temperaturas rectales más bajas en perros alimentados con una dieta de mantenimiento cubierta con aceite de maíz en comparación con perros que consumen la ración de rendimiento sin aceite de maíz. La agudeza olfativa no se midió en este estudio. Por el contrario, las tasas de caza en English Pointers mejoraron en perros alimentados con una dieta más alta en proteínas y grasas (31:21%), en comparación con una dieta que contiene menos proteínas y grasas (26:17%) (79). No se investigó la recuperación térmica. Los factores asociados con la fatiga no se informaron en ninguno de los estudios. La extrapolación entre los estudios es un desafío debido a la diferencia en los métodos, ingredientes y parámetros medidos, pero la recuperación térmica y el impacto olfativo deben sopesarse mucho en las decisiones con respecto a las dietas para perros de detección.
Contenido de nutrientes
Los requerimientos nutricionales para los atletas caninos han sido examinados previamente en revisión (97). Mullis et al. (98) examinaron los requisitos de energía de mantenimiento específicos de los perros de detección e informaron que eran aproximadamente el doble del requerimiento de energía en reposo conocido (RER = 70 kcal × BW0,75 kg). Los autores no notaron diferencias en los requisitos de energía según la raza, la edad o el género, pero informaron un efecto significativo asociado con el número de búsquedas realizadas. Esto es particularmente interesante porque el trabajo realizado por estos perros simplemente requería que estuvieran activos y alertas; No se informó como físicamente extenuante. Los hallazgos en estos perros sugieren que puede haber un requerimiento de energía inexplicable asociado con el enfoque / atención mental requerido por los caninos de trabajo. La duración, la frecuencia y la intensidad del trabajo probablemente afecten los requisitos de energía para el canino de trabajo. Se desconoce el impacto de la esterilización quirúrgica en el olfato, pero la esterilización de las perras galgo de carreras no produjo cambios en el rendimiento general, la motivación o la velocidad de carrera (99).
El ejercicio y la dieta parecen estar inextricablemente vinculados al rendimiento canino, pero hay pocos estudios que examinen la relación entre estos elementos del manejo del perro de detección. Los Bracos Ingleses retenidos del ejercicio y alimentados con una dieta suplementada con aceite de coco parecían experimentar un olfato comprometido, pero los perros ejercitados mantuvieron la agudeza olfativa (77). Los autores informaron una mayor sensibilidad olfativa para todos los perros ejercitados, independientemente de la fuente de grasa dietética (sebo de res; sebo de res + aceite de maíz; sebo de res + aceite de coco). Angle et al. (78) demostraron beneficios para el olfato al usar dietas complementadas con aceite de maíz y ejercicio.
La mejora del olfato observada con el aumento del contenido de ácidos grasos poliinsaturados (PUFA) en la dieta también se ha informado en estudios con roedores (100). Los estudios con roedores también se han utilizado para medir la sensibilidad olfativa asociada con el estado nutricional y han reportado una mejora del olfato asociada con el ayuno (101, 102) y el olfato comprometido como resultado de la saciedad (101, 103). Se cree que estos hallazgos están relacionados con la hormona inductora del apetito grelina, que contribuye al comportamiento exploratorio y olfateador y mejora la sensibilidad olfativa (104). Este trabajo de importancia crítica demuestra un vínculo potencial entre el ayuno y la mejora del rendimiento en los perros de detección. Los informes anecdóticos de entrenadores experimentados a menudo han incluido recomendaciones para dejar que los perros trabajen con hambre; Estos datos pueden proporcionar evidencia de esta técnica de entrenamiento canino de larga data. Se ha documentado que la comida es una recompensa más efectiva que los elogios o las caricias, pero no se ha comparado con la efectividad de los juguetes (105). Hall et al. (90) informaron respuestas inconsistentes para los perros a los que se les ofreció alimentación antes de la sesión cuando se realizaron pruebas de discriminación de olores. Para las disciplinas de detección que requieren que los perros trabajen independientemente del manejador (desastre, explosivos), el uso del ayuno para mejorar el comportamiento exploratorio y olfateador puede ser una herramienta de entrenamiento útil para examinar. Se necesitan más estudios para determinar la titulación adecuada de la dieta para maximizar el olfato, la duración del tiempo de ayuno necesario y los posibles impactos en el rendimiento olfativo.
Dieta y comportamiento
La relación entre la dieta y el comportamiento ha sido bien estudiada en otras especies, pero pocos estudios han examinado la relación entre la dieta y el comportamiento en caninos (106). El ácido docosahexaenoico (DHA) es necesario para el desarrollo neurológico óptimo en cachorros, y concentraciones más bajas de DHA se han asociado con la agresión en perros pastores alemanes (40). Los AGPI son esenciales para la función de la membrana y el control del estrés oxidativo, especialmente en el hipocampo del cerebro, el área responsable del aprendizaje asociativo (40). Hennessy et al. (107) informaron una reducción en la hormona adrenocorticotrópica tras la exposición a nuevos estímulos para aquellos alimentados con una dieta premium (44% de proteína de origen animal) en comparación con aquellos alimentados con una dieta de mantenimiento (17% de proteína de origen animal). Otros estudios han demostrado una reducción en la agresión territorial en perros propiedad del cliente alimentados con una dieta baja en proteínas (106-109). DeNapoli et al. (110) informaron que las dietas bajas en proteínas con triptófano suplementario redujeron la agresión en perros. Sechi et al. (111) utilizaron una estrategia de intervención dietética de suplementación nutracéutica (incluyendo triptófano) en perros con trastornos de conducta. Informaron un aumento posterior en la serotonina, la dopamina y las β-endorfinas que indican una reducción de la agresión, y una reducción del cortisol plasmático y la noradrenalina que indica marcadores reducidos de estrés. Estos estudios ofrecen una visión de la aplicación potencial de la manipulación dietética para el manejo del estrés y la agresión. La necesidad de que los caninos de trabajo operen sin agresión en ambientes estresantes justifica una mayor investigación en esta área. Sin embargo, la reducción de la proteína dietética podría ser una tarea peligrosa y este tema requeriría una extensa investigación antes del uso de esta estrategia de mitigación para perros en el campo.
Microbiota
Entendiendo los microbios
La comunidad microbiana GI es un ecosistema complejo que contiene bacterias, hongos, arqueas y protozoos. Las mejoras en las técnicas moleculares, como la secuenciación de próxima generación, han aumentado nuestro estudio y, posteriormente, nuestra comprensión tanto de la composición como de la función de la microflora gastrointestinal. Sin embargo, sigue habiendo muchas preguntas sin respuesta sobre los impactos asociados con los cambios en la microbiota en la salud general y el rendimiento del canino de trabajo.
A medida que se publican más estudios que destacan los cambios en la microbiota gastrointestinal, es cada vez más importante comprender cómo se miden esos cambios y cómo se presentan esos datos (112-115). Microflora, microbiota y microbioma son palabras que parecen impregnar la discusión en muchas comunidades científicas. Microflora es un término que se refiere a la comunidad colectiva (hongos, arqueas, protozoos, bacterias) en cuestión. Las bacterias se conocen como la «microbiota». Los estudios que hacen referencia al término microbioma generalmente describen el genoma de la microbiota y generalmente incluyen información sobre subproductos de la fermentación (AGV, pH, etc.), así como información genética sobre los componentes de la comunidad (116).
Los estudios de microbiota generalmente se presentan visualmente para responder preguntas relacionadas con la taxonomía, como (1) ¿Cuántas y qué comunidades microbianas están presentes? (2) ¿Cuál es la diversidad de la población? La diversidad taxonómica generalmente se representa utilizando la diversidad alfa y beta. La diversidad alfa (diversidad dentro de una muestra dada) se representa típicamente como una curva de rarefacción y describe la uniformidad y riqueza de una muestra dada (117). Es más probable que falten especies microbianas raras en muestras pequeñas, por lo tanto, la riqueza es un factor importante a considerar para conjuntos de datos pequeños. Alternativamente, la diversidad beta (diversidad entre muestras) se utiliza para medir la similitud taxonómica basada en la distancia filogenética (118). La diversidad beta también proporciona una evaluación visual de la abundancia (ponderada) o la presencia (no ponderada) de taxones dados y se representa utilizando una gráfica PCoA. Otras técnicas para la representación visual de datos incluyen mapas de calor o análisis jerárquico de conglomerados.
Aunque una discusión exhaustiva sobre los procedimientos asociados con la secuenciación microbiana está más allá del alcance de este trabajo, es importante comprender que la selección del cebador y la región objetivo del gen de ARN 16s son críticos (119, 120) y pueden causar una variación significativa en los resultados y la posterior interpretación de los datos generados. Estas técnicas son independientes del cultivo y han permitido a los investigadores mejorar en gran medida nuestra comprensión de la microbiología gastrointestinal. Los datos se ven muy afectados por varios factores, incluidas las técnicas de secuenciación, los cebadores, la selección de la región hipervariable correcta y otros. Los enfoques inconsistentes utilizados en muchos estudios publicados han hecho que sea extremadamente difícil hacer comparaciones entre conjuntos de datos y continúan desafiando la interpretación.
Las técnicas tradicionales dependientes del cultivo (es decir, la secuenciación de Sanger) han permitido a los investigadores investigar la presencia de patógenos específicos y son útiles para identificar especies comúnmente asociadas con enfermedades gastrointestinales como Salmonella, Campylobacter jejuni o Clostridium perfringens. Sin embargo, estas técnicas son limitadas en su aplicabilidad en comparación con los métodos moleculares actuales (es decir, la secuenciación de próxima generación) que facilitan la identificación taxonómica y las aplicaciones metagenómicas (121). La caracterización integral y la identificación de la comunidad requeridas para el perfil microbiano del tracto gastrointestinal requieren las técnicas más sensibles asociadas con la secuenciación de próxima generación y se ha convertido en el estándar aceptado para los estudios microbianos.
Equilibrio microbiano
El ecosistema microbiano gastrointestinal alberga comunidades significativamente diferentes dentro de cada compartimento (122, 123). Los filos predominantes en los caninos de trabajo son similares a otras especies monogástricas y están típicamente dominados por Firmicutes y Bacteroidetes. La caracterización de la comunidad microbiana GI colectiva y la función asociada está más allá del alcance de este trabajo y se ha informado previamente en otros lugares (114, 122, 123). Los grupos bacterianos residentes dentro del tracto gastrointestinal juegan un papel intrínseco en la regulación de la homeostasis; Su papel en la regulación de la inmunidad innata del huésped ha sido bien descrito (124-126). La microbiota comprende parte de la barrera lumínica intestinal, contribuyendo a la protección del ecosistema gastrointestinal a través de la competencia por nutrientes y sitios de adhesión y secretando compuestos que se cree que inhiben la colonización de microbios no residentes (127). Esto puede explicar por qué los cachorros generalmente tienen más riesgo de enfermedad gastrointestinal asociada con patógenos como C. jejuni, ya que su perfil bacteriano aún puede no estar lo suficientemente maduro como para proporcionar suficiente protección o disuasión (128).
La variación de la estructura de la comunidad microbiana entre individuos está constantemente presente (129). Se ha demostrado que la edad, la raza y el género afectan el perfil microbiano en múltiples especies (130-132). También se ha demostrado que la cohabitación de humanos y perros afecta a la comunidad microbiana (133). Los autores concluyeron que los factores que afectan la homeostasis microbiana no son los mismos para las comunidades oral y gastrointestinal en comparación con la comunidad de la piel. Estos datos sugieren que los cambios en el IG están relacionados con otros factores hasta ahora desconocidos. Estas variaciones reportadas deben considerarse al evaluar los datos microbianos publicados. Los estudios que incluyen perros en varios grupos de edad, razas y con ambos sexos deben analizarse en consecuencia para tener en cuenta la variación asociada con esos factores.
Desequilibrio microbiano
Si bien es relativamente fácil predecir los factores que afectarán a la microbiota (edad, sexo, raza, uso de antibióticos, viajes), es un poco más difícil predecir los impactos asociados al perro. La evidencia actual sugiere que las alteraciones en la comunidad microbiana GI pueden alterar fundamentalmente la estructura y función de la luz GI; Esto se ha denominado «síndrome del intestino permeable» con revisión previa en otros lugares (134). Esta condición describe los cambios físicos en la luz intestinal asociados con cambios en el perfil microbiano y es particularmente preocupante debido al potencial de alteración inmunológica y translocación bacteriana que resulta en endotoxemia. Se cree que los subproductos de la fermentación microbiana saludable, específicamente los ácidos grasos de cadena corta (AGCC), proporcionan energía para el huésped y contribuyen a la mediación entre el ecosistema microbiano y la activación del sistema inmunológico (135).
Los altos niveles de diversidad bacteriana generalmente se asocian con una buena salud; La disminución de la diversidad se ha reportado consistentemente con resultados negativos para la salud en humanos, como obesidad, diabetes y enfermedades gastrointestinales (136). Se han reportado reducciones en los filos Firmicutes y Bacteroidetes, que son típicamente dominantes, junto con aumentos concomitantes en Proteobacteria en perros diagnosticados con enfermedad inflamatoria gastrointestinal crónica (137). Minamoto et al. (138) demostraron impactos microbianos ligeramente diferentes, pero eso puede deberse a la variación inherente a las diferentes técnicas, razas y edades de los perros muestreados. El desarrollo de un índice de disbiosis (DI) ha ofrecido una herramienta de diagnóstico para categorizar los datos microbianos en una proporción simple que refleja la microbiota normal (DI < 0) o la microbiota indicativa de enteropatías crónicas (DI > 0) (139). Desafortunadamente, el uso de este índice requiere pruebas de laboratorio y está limitado por su conjunto de datos inicial muy pequeño. Sin embargo, el concepto proporciona un paso importante en la dirección de evaluar las muestras fecales de diagnóstico con recomendaciones para el tratamiento y las intervenciones dietéticas.
La comunicación bidireccional que se produce entre el cerebro y el intestino (eje microbiota-intestino-cerebro) proporciona una idea de la disbiosis que se ha informado como resultado del estrés ambiental (140). El estrés asociado con los viajes, el cambio en el entorno y el esfuerzo físico son comunes en el canino que trabaja. Los cambios en la microbiota fecal de los caninos en funcionamiento después del transporte en cabina a través de una aerolínea comercial tuvieron un impacto tanto en la abundancia como en el tipo de bacterias y se acompañaron de una puntuación fecal más baja (85). Por el contrario, cuando los investigadores examinaron los efectos del estrés de viaje en helicóptero en caninos de trabajo, la naturaleza relativamente corta del factor estresante (carga caliente y 30 minutos de vuelo) no produjo ningún efecto sobre la microbiota (83). En particular, ambos estudios no informaron ningún efecto sobre el rendimiento según lo determinado por el tiempo total de búsqueda o los comportamientos de estrés previamente identificados. La duración y el tipo de viaje requerido para inducir disbiosis microbiana no se ha examinado en caninos de trabajo.
Modificación dietética de la microbiota
Si bien los estudios en perros son limitados, algunos datos han mostrado resultados prometedores para la manipulación microbiana mediante el uso de diferentes suplementos de fibra en la comunidad microbiana y la producción resultante de AGCC (112, 113, 132, 141-144).
Los investigadores que examinaron el uso de fructooligosacáridos informaron una mejor producción de butirato, un ácido graso volátil beneficioso para la reparación de colonocitos y células epiteliales, así como reducciones en C. perfringens, un microbio potencialmente patógeno. Un segundo estudio arrojó resultados similares junto con un mayor número de bifidobacterias, un microbio potencialmente beneficioso (144). Otro trabajo en perros de trineo alimentados con un simbiótico (pre y probióticos combinados) informó una disminución de la incidencia de diarrea (141). Si los investigadores pueden desarrollar estrategias de mitigación dietética que reduzcan o prevengan consistentemente el malestar gastrointestinal, esto puede beneficiar a los perros que trabajan en escenarios de campo con acceso limitado a la intervención veterinaria. El uso de suplementos dietéticos que pueden mitigar o prevenir la aparición de malestar gastrointestinal justifica estudios adicionales.
La dieta ha sido identificada durante mucho tiempo como el factor dominante que afecta la estructura de la comunidad microbiana (112, 113, 144-148). Lo que no sabemos es qué impacto tiene el tamaño y la frecuencia de las comidas en la microbiota gastrointestinal. Los manejadores con frecuencia deben ajustar los horarios y tamaños de las comidas para los perros de detección a lo largo del curso de una misión. Los datos en caballos han demostrado un efecto sobre la microbiota gastrointestinal asociado con la frecuencia y el tamaño de las comidas (148); No se sabe si se observaría un impacto similar en el canino monogástrico. La información que aclare los posibles impactos en la microbiota sería útil para manejar las preocupaciones asociadas con la diarrea en el campo.
Microflora y olfato
Se ha informado que el nicho microbiano densamente poblado en el tracto gastrointestinal desempeña un papel clave en la regulación del comportamiento y la función cerebral. El eje microbiota-intestino-cerebro influye en la neurotransmisión y el comportamiento. Por lo tanto, podría ser la clave en las intervenciones nutricionales para mantener la salud del cerebro y el olfato (149), con una modulación microbiana temprana que resulta en impactos a largo plazo en la fisiología y el comportamiento relacionados con el estrés (150). Dada la naturaleza relativamente inexplorada de la comunicación que se produce entre la microbiota intestinal y el sistema de respuesta al estrés del cerebro, parece razonable preguntarse si las alteraciones de la microbiota intestinal podrían desempeñar un papel en la reducción del estrés, como lo demuestra la exhibición de comportamientos de estrés en el perro.
El epitelio olfativo generalmente se ha pasado por alto con respecto al papel potencial de los microorganismos en el desarrollo y la eficiencia de la transducción de olores. Los OR están formados por muchas proteínas receptoras acopladas a proteínas G que identifican moléculas odoríferas volátiles (151). Originalmente se pensaba que los quirófanos solo se encontraban en el epitelio olfativo. En el tracto gastrointestinal, se han identificado OR en células enterocromafines; estos receptores pueden afectar la secreción de serotonina en respuesta a moléculas fragantes con efectos posteriores sobre la motilidad GI (152). La serotonina también juega un papel crítico para el procesamiento de información olfativa, ya que el bulbo olfatorio está compuesto de fibras serotoninérgicas y recientemente se demostró que regula eficazmente el flujo de procesamiento olfativo en ratones (153). Dado el vínculo entre la microbiota gastrointestinal y la regulación de la serotonina, parece probable que exista una relación entre la microbiota GI y la detección de olores, aunque aún se desconoce (154).
La estructura de la comunidad de la microbiota nasal se ha relacionado con la función olfativa (155). Los sujetos humanos demostraron diferencias en la microbiota de las personas evaluadas para la función olfativa con deficiencias relacionadas con la presencia de microbios productores de ácido butírico (155). Estos hallazgos sugieren que la composición microbiana del pasaje nasal puede potencialmente dar forma o alterar el rendimiento olfativo. Las implicaciones del rendimiento olfativo alterado asociado con las fluctuaciones bacterianas en la nariz son significativas. Se informó que la microbiota nasal de los perros con rinitis crónica y neoplasia nasal difiere en la estructura de la comunidad en comparación con los perros sanos (156). Isaiah et al. (157) identificaron un efecto asociado con el tipo de trabajo en la microbiota nasal canina. A pesar de que todos los perros fueron alojados en una sola instalación y alimentados con una sola dieta, los investigadores informaron diferencias en la diversidad alfa para los caninos que estaba relacionada con el tipo de trabajo (estela de vapor, patrulla y narcóticos, explosivos). No se reportaron diferencias en la diversidad beta, lo que sugiere que la riqueza de especies, pero no la estructura de la comunidad bacteriana, se vio afectada por el trabajo realizado por los perros en cada grupo (157).
Se ha detectado un OR específico (OR51E1) en cerdos a lo largo de todo el tracto gastrointestinal desde el cardias gástrico hasta el recto (152). OR51E1 se colocaliza con un marcador celular enteroendocrino a lo largo del tracto gastrointestinal y se expresó en la mayor densidad en el duodeno. Las células enteroendocrinas duodenales son la principal fuente de péptido inhibidor gástrico y colecistoquinina. Las células duodenales enteroendocrinas están equipadas con múltiples receptores conectados a sabores dulces y amargos. La expresión génica OR51E1 en bulbos olfatorios ha demostrado mecanismos de retroalimentación, activación diferencial de factores de transcripción y regulación epigenética. Las hormonas circulantes que controlan la ingesta de alimentos y el equilibrio energético modulan el epitelio olfativo, y la ablación de las neuronas sensoriales olfativas en ratones los protegió de la obesidad inducida por la dieta (158). Hay varios factores como la edad y la dieta que afectan el microambiente luminal intestinal y la microbiota intestinal modula la expresión génica OR51E1 en los tejidos del tracto gastrointestinal (152).
Direcciones futuras y preguntas sin respuesta
Carecemos de datos basados en evidencia realizados en caninos de trabajo que nos permitan investigar a fondo los vínculos entre los cambios de microbiota y cualquier posible rendimiento (es decir, relacionado con el olfato) o sequalae de salud. Sabemos que la dieta puede cambiar la microbiota e impactar el olfato en otras especies. Lo que aún no sabemos es qué mecanismo (si es que existe) existe que vincula el olfato con la microbiota. Cuando se considera la comunidad microbiana única que alberga el perro individual, ¿explica eso por qué el olfato solo se vio comprometido en el 50% de los perros que recibieron metronidazol (71)? ¿Es posible que la reducción de Firmicutes experimentada por los perros que recibieron metronidazol proporcione la clave del desafío olfativo que experimentaron (114)? Si el olfato se mejora como resultado del ayuno (102) y la saciedad reduce el rendimiento olfativo (103), ¿deberíamos repensar el momento de nuestros programas de alimentación? ¿Qué impactos tendrá ese ayuno en la microbiota del canino que trabaja? El impacto crítico del trabajo realizado por estos caninos requiere una comprensión mucho más profunda de todas las cosas que podrían obstaculizar su desempeño laboral. Una investigación más exhaustiva de los factores asociados con los cambios microbianos y los impactos asociados en el desempeño laboral (es decir, el olfato) es vital.
Contribuciones del autor
MD desarrolló el concepto y escribió el primer borrador del manuscrito. EJ y EP escribieron secciones principales del documento en su forma actual. Todos los autores leyeron, editaron y aprobaron el manuscrito final.
Renuncia
Las opiniones expresadas en este documento son las del autor (es) y no reflejan la política oficial del Departamento Médico del Ejército de los Estados Unidos, el Departamento del Ejército, el Departamento de Defensa o el Gobierno de los Estados Unidos.
Declaración de conflicto de intereses
Esta revisión se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Reconocimientos
Los autores agradecen a Tracy Darling, RVT, VTS (SAIM), Adrien-Maxence Hespel, DVM, MS, DACVR, Nicholas Jenkins, MAS, MEd, y Kathleen Kelsey, MS, MBA, por su ayuda en el desarrollo de figuras y tablas.
Referencias
1. Marchal S, Bregeras O, Puaux D, Gervais R, Ferry B. El entrenamiento riguroso de perros conduce a una alta precisión en la coincidencia del olor humano con el rendimiento de la muestra. PLoS One (2016) 11(2):e0146963. doi:10.1371/journal.pone.0146963
2. Walker D, Walker J, Cavnar P, Taylor J, Pickel D, Hall S, et al. Cuantificación naturalista de la sensibilidad olfativa canina. Appl Anim Behav Sci (2006) 97:241–54. doi:10.1016/j.applanim.2005.07.009
3. Riezzo I, Neri M, Rendine M, Bellifemina A, Cantatore S, Fiore C, et al. Perros cadáveres: ¿mito no científico o dispositivos biológicos fiables? Forensic Sci Int (2014) 244:213–21. doi:10.1016/j.forsciint.2014.08.026
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Angle C, Waggoner LP, Ferrando A, Haney P, Passier T. Detección canina del volátilema: una revisión de las implicaciones para la detección de patógenos y enfermedades. Front Vet Sci (2016) 3:47. doi:10.3389/fvets.2016.00047
Resumen de PubMed | Texto completo de CrossRef | Google Académico
5. Hall NJ, Glenn K, Smith DW, Wynne CD. Actuación de pugs, pastores alemanes y galgos (Canis lupus familiaris) en una tarea de discriminación de olores. J Comp Psychol (2015) 129:237–46. doi:10.1037/a0039271
6. Evans HE, de Lahunta A. Miller’s Anatomy of the Dog. 4ª ed. St. Louis, MO: Elsevier Saunders (2013).
7. Quignon P, Galibert F. Genética del olfato canino. En: Jezierski T, Ensminger J, Papet LE, editores. Ciencia y Derecho del Olfato Canino. Boca Raton, FL: CRC Press (2016). págs. 39–48.
8. Bromley SM. Trastornos del olfato y el gusto: un enfoque de atención primaria. Am Fam Physician (2000) 61:427–36.
10. Menco B, Morrison E. Morfología del epitelio olfativo de los mamíferos: forma, estructura fina, función y patología. En: Doty RL, editor. Manual de Olfato y Gustation. Nueva York, NY: Wiley Blackwell (2003). págs. 17–49.
11. Sala JE. Libro de texto de fisiología médica de Guyton y Hall. 12ª ed. Filadelfia, PA: Elsevier (2011).
12. Uemura EE. Fundamentos de Neuroanatomía y Neurofisiología Canina. 1ª ed. Danvers, MA: Blackwell (2015).
13. Parnafes-Gazit I. Variables que influyen en el comportamiento y la eficiencia de detección en perros detectores de explosivos. Tel Aviv-Yafo, Israel: Universidad de Tel Aviv (2005).
14. Gerritsen R, Hank R. K9 Scent Training: A Manual for Training Your Identification, Tracking, and Detection Dog. Vancouver, BC: Brush Education, Inc (2015).
15. Tacher S, Quignon P, Rimbault M, Dreano S, Andre C, Galibert F. Polimorfismo de secuencia del receptor olfativo dentro y entre razas de perros. J Heredity (2005) 96:812–6. doi:10.1093/jhered/esi113
16. Wang HW, Wysocki CJ, Gold GH. Inducción de la sensibilidad olfativa del receptor en ratones. Science (1993) 260:998–1000. doi:10.1126/science.8493539
Resumen de PubMed | Texto completo de CrossRef | Google Académico
17. Youngentob SL, Kent PF. Mejora de los patrones de actividad mucosa inducidos por olores en ratas entrenadas en una tarea de identificación de olores. Brain Res (1995) 670:82–8. doi:10.1016/0006-8993(94)01275-M
Resumen de PubMed | Texto completo de CrossRef | Google Académico
18. Buck LB, Bargmann CI. Olor y gusto: los sentidos químicos. En: Kandel ER, editor. Principios de la Ciencia Neural. 5ª ed. Nueva York, NY: McGraw Hill (2013). págs. 712–35.
19. Harper RJ, Furton KG. Detección biológica de explosivos. En: Yinon J, editor. Técnicas de detección antiterrorista de explosivos. Ámsterdam: Elsevier (2007). págs. 395–431.
20. Dennis JC, Allgier JG, Desouza LS, Eward WC, Morrison EE. Inmunohistoquímica del órgano vomeronasal. J Anat (2003) 202(6):515–24. doi:10.1046/j.1469-7580.2003.00190.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
22. Caín DP. El papel del bulbo olfatorio en los mecanismos límbicos. Psychol Bull (1974) 81:654–71. doi:10.1037/h0036954
23. Soudry Y, Lemogne C, Malinvaud D, Consoli SM, Bonfils P. Sistema olfativo y emoción: sustratos comunes. Eur Ann Otorhinolaryngol Head Neck Dis (2011) 128:18–23. doi:10.1016/j.anorl.2010.09.007
Resumen de PubMed | Texto completo de CrossRef | Google Académico
24. Reep RL, Finlay BL, Darlington RB. El sistema límbico en la evolución del cerebro de los mamíferos. Brain Behav Evol (2007) 70:57–70. doi:10.1159/000101491
Resumen de PubMed | Texto completo de CrossRef | Google Académico
25. Barrios AW, Sánchez-Quinteiro P, Salazar I. Perro y ratón: hacia una visión equilibrada del sistema olfativo de los mamíferos. Neuroanat frontal (2014) 8:106. doi:10.3389/fnana.2014.00106
Resumen de PubMed | Texto completo de CrossRef | Google Académico
26. Craven BA, Paterson EG, Settles GS. La dinámica de fluidos del olfato canino: patrones únicos de flujo de aire nasal como explicación de la macrosmia. J R Soc Interface (2010) 7(47):933–43. doi:10.1098/rsif.2009.0490
27. Philpott C, Goodenough P, Passant C, Robertson A, Murty G. El efecto de la temperatura, la humedad y el flujo nasal inspiratorio máximo sobre los umbrales olfativos. Clin Otolaryngol Allied Sci (2004) 29:24–31. doi:10.1111/j.1365-2273.2004.00760.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
28. Kuehn M, Welsch H, Zahnert T, Hummel T. Los cambios de presión y humedad afectan la función olfativa. Eur Arch Otorhinolaryngol (2008) 265:299–302. doi:10.1007/s00405-007-0446-2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
30. Gutzwiller KJ. Minimizar los sesgos inducidos por perros en la investigación de aves de caza. Wildlife Soc B (1990) 18:351–6.
31. Sen Majumder S, Bhadra A. Cuando el amor está en el aire: entender por qué los perros tienden a aparearse cuando llueve. PLoS One (2015) 10:e0143501. doi:10.1371/journal.pone.0143501
Resumen de PubMed | Texto completo de CrossRef | Google Académico
32. Gazit I, Terkel J. Detección de explosivos por perros rastreadores después de una actividad física extenuante. Appl Anim Behav Sci (2003) 81:149–61. doi:10.1016/S0168-1591(02)00274-5
33. Mainland J, Sobel N. El olfateo es parte de la percepción olfativa. Chem Senses (2006) 31:181–96. doi:10.1093/chemse/bjj012
Resumen de PubMed | Texto completo de CrossRef | Google Académico
34. Lawson MJ, Craven BA, Paterson EG, Settles GS. Un estudio computacional del transporte y deposición de odorantes en la cavidad nasal canina: implicaciones para el olfato. Chem Senses (2012) 37:553–66. doi:10.1093/chemse/bjs039
Resumen de PubMed | Texto completo de CrossRef | Google Académico
35. Thesen A, Steen JB, Doving KB. Comportamiento de los perros durante el seguimiento olfativo. J Exp Biol (1993) 180:247–51.
36. Concha A, Mills DS, Feugier A, Zulch H, Guest C, Harris R, et al. Usar el comportamiento de olfateo para diferenciar las respuestas negativas verdaderas de las falsas negativas en perros entrenados con detección de olores. Chem Senses (2014) 39:749–54. doi:10.1093/chemse/bju045
Resumen de PubMed | Texto completo de CrossRef | Google Académico
37. Siniscalchi M. El olfato y el cerebro canino. En: Jezierski T, Ensminger J, Papet LE, editores. Ciencia y Derecho del Olfato Canino. Boca Raton, FL: Taylor & Francis Group (2016). págs. 31–7.
38. Siniscalchi M, Sasso R, Pepe AM, Dimatteo S, Vallortigara G, Quaranta A. Sniffing with the right nasal tril: lateralization of response to odor stimuli by dogs. Anim Behav (2011) 82:399–404. doi:10.1016/j.anbehav.2011.05.020
39. D’Aniello B, Semin GR, Alterisio A, Aria M, Scandurra A. Interspecies transmission of emotional information via chemosignals: from humans to dogs (Canis lupus familiaris). Anim Cogn (2018) 21:67–78. doi:10.1007/s10071-017-1139-x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
40. En general KL. Ese perro es más inteligente de lo que crees: avances en la comprensión del aprendizaje canino, la memoria y la cognición. Top Companion Anim Med (2011) 26:2–9. doi:10.1053/j.tcam.2011.01.001
Resumen de PubMed | Texto completo de CrossRef | Google Académico
41. Wells DL, Hepper PG. Aprendizaje olfativo prenatal en el perro doméstico. Anim Behav (2006) 72:681–6. doi:10.1016/j.anbehav.2005.12.008
42. Hepper PG, Wells DL. Aprendizaje olfativo perinatal en el perro doméstico. Chem Senses (2006) 31:207–12. doi:10.1093/chemse/bjj020
Resumen de PubMed | Texto completo de CrossRef | Google Académico
43. Williams M, Johnston JM. Entrenar y mantener el rendimiento de los perros (Canis familiaris) en un número creciente de discriminaciones de olores en un entorno controlado. Appl Anim Behav Sci (2002) 78:55–65. doi:10.1016/S0168-1591(02)00081-3
44. Kaminski J, Call J, Fischer J. Word learning in a domestic dog: evidence for «Fast Mapping». Science (2004) 304:1682–3. doi:10.1126/science.1097859
45. Pilley JW, Reid AK. Border collie comprende los nombres de objetos como referentes verbales. Behav Processes (2011) 86:184–95. doi:10.1016/j.beproc.2010.11.007
Resumen de PubMed | Texto completo de CrossRef | Google Académico
46. Johnston JM. Capacidades de detección canina: implicaciones operativas de los hallazgos recientes de investigación y desarrollo. Auburn, AL: Universidad de Auburn (1999).
47. Hall NJ, Wynne CD. Aprendizaje olfativo canino y comportamiento. En: Jezierski T, Ensminger J, Papet LE, editores. Ciencia y Derecho del Olfato Canino. Boca Raton, FL: CRC Press (2016). págs. 123–37.
48. Quignon P, Rimbault M, Robin S, Galibert F. Genetics of canine olfaction and receptor diversity. Mamm Genome (2012) 23:132–43. doi:10.1007/s00335-011-9371-1
Resumen de PubMed | Texto completo de CrossRef | Google Académico
49. Aloni R, Olender T, Lancet D. Ancient genomic architecture for mammalian olfactory receptor clusters. Genoma Biol (2006) 7:R88. doi:10.1186/gb-2006-7-10-r88
Resumen de PubMed | Texto completo de CrossRef | Google Académico
50. Jezierski T, Adamkiewicz E, Walczak M, Sobczynska M, Gorecka-Bruzda A, Ensminger J, et al. La eficacia de la detección de drogas por perros policía totalmente entrenados varía según la raza, el nivel de entrenamiento, el tipo de droga y el entorno de búsqueda. Forensic Sci Int (2014) 237:112–8. doi:10.1016/j.forsciint.2014.01.013
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Polgar Z, Kinnunen M, Ujvary D, Miklosi A, Gacsi M. Una prueba de la capacidad olfativa canina: comparando varias razas de perros y lobos en una tarea de detección natural. PLoS One (2016) 11:e0154087. doi:10.1371/journal.pone.0154087
Resumen de PubMed | Texto completo de CrossRef | Google Académico
52. Henkin RI. Trastornos del gusto y el olfato inducidos por drogas. Incidencia, mecanismos y manejo relacionados principalmente con el tratamiento de la disfunción sensorial de los receptores. Drug Saf (1994) 11:318–77. doi:10.2165/00002018-199411050-00004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
53. Seiden AM, Duncan HJ. El diagnóstico de una pérdida olfativa conductiva. Laringoscopio (2001) 111:9–14. doi:10.1097/00005537-200101000-00002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
54. Malaty J, Malaty IA. Trastornos del olfato y del gusto en atención primaria. Am Fam Physician (2013) 88:852–9.
56. Seiden AM. Pérdida olfativa postviral. Otolaryngol Clin North Am (2004) 37:1159–66. doi:10.1016/j.otc.2004.06.007
Resumen de PubMed | Texto completo de CrossRef | Google Académico
57. McNeill EJM, Carrie S. Olfactory dysfunction – assessment and management. J ENT Masterclass (2009) 2:68–73.
58. Jia H, Pustovyy OM, Waggoner P, Beyers RJ, Schumacher J, Wildey C, et al. Resonancia magnética funcional del sistema olfativo en perros conscientes. PLoS One (2014) 9:e86362. doi:10.1371/journal.pone.0086362
Resumen de PubMed | Texto completo de CrossRef | Google Académico
59. Scott AE. Características clínicas de los trastornos del gusto y el olfato. Ear Nose Throat J (1989) 68:297–298,301,304–210assim.
60. Barco JA, Chavez EM. Sentidos especiales: trastornos del gusto y el olfato de Hamilton. Ontario: BC Decker (2001).
61. Yousem DM, Geckle RJ, Bilker W, McKeown DA, Doty RL. Evaluación por RM de pacientes con hiposmia congénita o anosmia. AJR Am J Roentgenol (1996) 166:439–43. doi:10.2214/ajr.166.2.8553963
Resumen de PubMed | Texto completo de CrossRef | Google Académico
63. Andress M, Goodnight ME. Golpe de calor en un perro de trabajo militar. US Army Med Dep J (2013) 1: 34–7.
64. Myers LJ, Hanrahan LA, Swango LJ, Nusbaum KE. Anosmia asociada con moquillo canino. Am J Vet Res (1988) 49:1295–7.
65. Myers LJ, Nusbaum KE, Swango LJ, Hanrahan LN, Sartin E. Dysfunction of sense of smell caused by canine parainfluenza virus infection in dogs. Am J Vet Res (1988) 49:188–90.
66. Asproni P, Cozzi A, Verin R, Lafont-Lecuelle C, Bienboire-Frosini C, Poli A, et al. Patología y comportamiento en medicina felina: investigando el vínculo entre vomeronasalitis y agresión. J Feline Med Surg (2016) 18: 997–1002. doi:10.1177/1098612X15606493
Resumen de PubMed | Texto completo de CrossRef | Google Académico
67. Otto CM. Efectos de la enfermedad en el olfato canino. En: Jezierski T, Ensminger J, Papet LE, editores. Ciencia y Derecho del Olfato Canino. Boca Raton, FL: Taylor & Francis Group (2016). págs. 31–7.
68. Doty RL, Kamath c. Las influencias de la edad en el olfato: una revisión. Front Psychol (2014) 5:20. doi:10.3389/fpsyg.2014.00020
Resumen de PubMed | Texto completo de CrossRef | Google Académico
69. Hirai T, Kojima S, Shimada A. Cambios relacionados con la edad en el sistema olfativo de los perros. Neuropathol Appl Neurobiol (1996) 22(6):532–9. doi:10.1111/j.1365-2990.1996.tb01132.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
70. Moore CH, Pustovyy O, Dennis JC, Moore T, Morrison EE, Vodyanoy VJ. Las respuestas olfativas a los olores asociados a explosivos se ven reforzadas por nanopartículas de zinc. Talanta (2012) 88:730–3. doi:10.1016/j.talanta.2011.11.024
Resumen de PubMed | Texto completo de CrossRef | Google Académico
71. Jenkins EK, Lee-Fowler TM, Angle TC, Behrend EN, Moore GE. Efectos de la administración oral de metronidazol y doxiciclina sobre las capacidades olfativas de los perros detectores de explosivos. Am J Vet Res (2016) 77:906–12. doi:10.2460/ajvr.77.8.906
Resumen de PubMed | Texto completo de CrossRef | Google Académico
72. Ezeh PI, Myers LJ, Hanrahan LA, Kemppainen RJ, Cummins KA. Efectos de los esteroides sobre la función olfativa del perro. Physiol Behav (1992) 51:1183–7. doi:10.1016/0031-9384(92)90306-M
Resumen de PubMed | Texto completo de CrossRef | Google Académico
73. Blomqvist EH, Lundblad L, Bergstedt H, Stjarne P. Estudio controlado con placebo, aleatorizado, doble ciego que evalúa la eficacia del aerosol nasal de propionato de fluticasona para el tratamiento de pacientes con hiposmia / anosmia. Acta Otolaryngol (2003) 123:862–8. doi:10.1080/00016480310002140
Resumen de PubMed | Texto completo de CrossRef | Google Académico
74. Golding-Wood DG, Holmstrom M, Darby Y, Scadding GK, Lund VJ. El tratamiento de la hiposmia con esteroides intranasales. J Laryngol Otol (1996) 110:132–5. doi:10.1017/S0022215100132967
Resumen de PubMed | Texto completo de CrossRef | Google Académico
75. Heilmann S, Huettenbrink KB, Hummel T. Local and systemic administration of corticosteroids in the treatment of olfactory loss. Am J Rhinol (2004) 18:29–33.
76. Lotsch J, Geisslinger G, Hummel T. Sniffing out pharmacology: interactions of drugs with human olfaction. Trends Pharmacol Sci (2012) 33:193–9. doi:10.1016/j.tips.2012.01.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
77. Altom EK, Davenport GM, Myers LJ, Cummins KA. Efecto de la fuente de grasa dietética y el ejercicio en la capacidad de detección de olores de los atletas caninos. Res Vet Sci (2003) 75:149–55. doi:10.1016/S0034-5288(03)00071-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
78. Angle CT, Wakshlag JJ, Gillette RL, Steury T, Haney P, Barrett J, et al. Los efectos del ejercicio y la dieta sobre la capacidad olfativa en perros de detección. J Nutr Sci (2014) 3:e44. doi:10.1017/jns.2014.35
Resumen de PubMed | Texto completo de CrossRef | Google Académico
79. Davenport GM, Kelley RL, Altom EK, Lepine AJ. Efecto de la dieta en el rendimiento de caza de los punteros ingleses. Vet Ther (2001) 2:10–23.
80. Davenport G, Kelley R, Altom E, Lepine A. Métodos dietéticos para la mejora del rendimiento canino. Publicación de solicitud de patente de los Estados Unidos No.: 0194478 A1 (2003).
81. Hegedus D, Balint A, Miklosi A, Pngracz P. Los propietarios no influyen en las elecciones de los perros en una tarea de apuntar visualmente de dos opciones. Comportamiento (2013) 150(3):427–43. doi:10.1163/1568539X-00003060
82. Lit L, Crawford CA. Efectos del entrenamiento de paradigmas en el rendimiento del perro de búsqueda. Appl Anim Behav Sci (2006) 98(3):277–92. doi:10.1016/j.applanim.2005.08.022
83. Perry E, Gulson N, Liu Cross TW, Swanson KS. Efectos fisiológicos del estrés relacionados con los viajes en helicóptero en caninos de búsqueda y rescate de la Agencia Federal para el Manejo de Emergencias. J Nutr Sci (2017) 6:e28. doi:10.1017/jns.2017.25
Resumen de PubMed | Texto completo de CrossRef | Google Académico
84. Porritt F, Shapiro M, Waggoner P, Mitchell E, Thomson T, Nicklin S, et al. Disminución del rendimiento de los perros de búsqueda en tareas repetitivas y estrategias de mitigación. Appl Anim Behav Sci (2015) 166:112–22. doi:10.1016/j.applanim.2015.02.013
85. Venable EB, Bland SD, Holscher HD, Swanson KS. Efectos del estrés de los viajes aéreos en el microbioma canino: un estudio piloto. Int J Vet Health Sci Res (2016) 4: 132–9. doi:10.19070/2332-2748-1600028
86. Zubedat S, Aga-Mizrachi S, Cymerblit-Sabba A, Shwartz J, Fiko Leon J, Rozen S, et al. Interfaz humano-animal: los efectos del estrés del manipulador en el rendimiento de los caninos en una tarea de detección de explosivos. Appl Anim Behav (2014) 158:69–75. doi:10.1016/j.applanim.2014.05.004
87. Wilson DA, Stevenson RJ. El papel fundamental de la memoria en la percepción olfativa. Trends Neurosci (2003) 26:243–7. doi:10.1016/S0166-2236(03)00076-6
Resumen de PubMed | Texto completo de CrossRef | Google Académico
88. Fischer-Tenhagen C, Johnen D, Heuwieser W, Becker R, Schallschmidt K, Nehls I. Percepción del olor por perros: evaluación de dos enfoques de entrenamiento para el aprendizaje del olor de perros rastreadores. Chem Senses (2017) 42:435–41. doi:10.1093/chemse/bjx020
Resumen de PubMed | Texto completo de CrossRef | Google Académico
89. Hall NJ, Smith DW, Wynne CD. Efecto de la preexposición al olor en la adquisición de una discriminación de olor en perros. Learn Behav (2014) 42:144–52. doi:10.3758/s13420-013-0133-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
90. Hall NJ, Smith DW, Wynne CD. El condicionamiento pavloviano mejora la resistencia a la interrupción de los perros que realizan una discriminación de olores. J Exp Anal Behav (2015) 103:484–97. doi:10.1002/jeab.151
91. Otto CM, Downend AB, Moore GE, Daggy JK, Ranivand DL, Reetz JA, et al. Vigilancia médica de perros de búsqueda desplegados en el World Trade Center y el Pentágono: 2001-2006. J Environ Health (2010) 73:12–21.
92. Slensky KA, Drobatz KJ, Downend AB, Otto CM. Deployment morbidity among search-and-rescue dogs used after the September 11, 2001, terrorist attacks. J Am Vet Med Assoc (2004) 225:868–73. doi:10.2460/javma.2004.225.868
Resumen de PubMed | Texto completo de CrossRef | Google Académico
93. Gordon LE. Lesiones y enfermedades entre los perros urbanos de búsqueda y rescate desplegados en Haití después del terremoto del 12 de enero de 2010. J Am Vet Med Assoc (2012) 240:396–403. doi:10.2460/javma.240.4.396
Resumen de PubMed | Texto completo de CrossRef | Google Académico
94. Gordon LE. Lesiones y enfermedades entre los perros de búsqueda y recuperación y búsqueda y rescate certificados por la Agencia Federal para el Manejo de Emergencias desplegados en Oso, Washington, después del deslizamiento de tierra de la Ruta Estatal 22 del 2014 de marzo de 530. J Am Vet Med Assoc (2015) 247: 901–8. doi:10.2460/javma.247.8.901
Resumen de PubMed | Texto completo de CrossRef | Google Académico
95. Otto CM, Hare E, Nord JL, Palermo SM, Kelsey KM, Darling TA, et al. Evaluación de tres estrategias de hidratación en perros de detección que trabajan en un ambiente caluroso. Front Vet Sci (2017) 4:174. doi:10.3389/fvets.2017.00174
Resumen de PubMed | Texto completo de CrossRef | Google Académico
96. Ober J, Gillette RL, Angle TC, Haney P, Fletcher DJ, Wakshlag JJ. Los efectos de concentraciones variables de proteínas y grasas en la dieta sobre los gases en sangre, la química sérica hematológica y la temperatura corporal antes y después del ejercicio en perros perdigueros de Labrador. Front Vet Sci (2016) 3:59. doi:10.3389/fvets.2016.00059
Resumen de PubMed | Texto completo de CrossRef | Google Académico
97. Wakshlag J, Shmalberg J. Nutrition for working and service dogs. Vet Clin North Am Small Anim Pract (2014) 44:719–740,vi. doi:10.1016/j.cvsm.2014.03.008
Resumen de PubMed | Texto completo de CrossRef | Google Académico
98. Mullis RA, Witzel AL, Price J. Maintenance energy requirements of odor detection, explosive detection and human detection working dogs. PeerJ (2015) 3:e767. doi:10.7717/peerj.767
Resumen de PubMed | Texto completo de CrossRef | Google Académico
99. Payne RM. El efecto de la esterilización en el rendimiento de las carreras de galgos hembras. Veterinario J (2013) 198:372–5. doi:10.1016/j.tvjl.2013.08.026
Resumen de PubMed | Texto completo de CrossRef | Google Académico
100. Seebungkert B, Lynch JW. Efectos de los ácidos grasos poliinsaturados en los canales K+ y Na+ dependientes de voltaje en neuronas receptoras olfativas de rata. Eur J Neurosci (2002) 16:2085–94. doi:10.1046/j.1460-9568.2002.02288.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
101. Aime P, Duchamp-Viret P, Chaput MA, Savigner A, Mahfouz M, Julliard AK. El ayuno aumenta y la saciedad disminuye la detección olfativa de un olor neutro en ratas. Behav Brain Res (2007) 179:258–64. doi:10.1016/j.bbr.2007.02.012
Resumen de PubMed | Texto completo de CrossRef | Google Académico
102. Soria-Gomez E, Bellocchio L, Reguero L, Lepousez G, Martin C, Bendahmane M, et al. El sistema endocannabinoide controla la ingesta de alimentos a través de procesos olfativos. Nat Neurosci (2014) 17:407–15. doi:10.1038/nn.3647
Resumen de PubMed | Texto completo de CrossRef | Google Académico
103. Critchley HD, Rolls ET. El hambre y la saciedad modifican las respuestas de las neuronas olfativas y visuales en la corteza orbitofrontal de los primates. J Neurophysiol (1996) 75:1673–86. doi:10.1152/jn.1996.75.4.1673
Resumen de PubMed | Texto completo de CrossRef | Google Académico
104. Tong J, Mannea E, Aime P, Pfluger PT, Yi CX, Castaneda TR, et al. La grelina mejora la sensibilidad olfativa y la olfateación exploratoria en roedores y humanos. J Neurosci (2011) 31:5841–6. doi:10.1523/JNEUROSCI.5680-10.2011
Resumen de PubMed | Texto completo de CrossRef | Google Académico
105. Feuerbacher EN, Wynne CD. La mayoría de los perros domésticos (Canis lupus familiaris) prefieren la comida a las caricias: población, contexto y efectos de horario en la elección concurrente. J Exp Anal Behav (2014) 101:385–405. doi:10.1002/jeab.81
Resumen de PubMed | Texto completo de CrossRef | Google Académico
106. Dodman NH, Reisner I, Shuster L, Rand W, Luescher UA, Robinson I, et al. Efecto del contenido de proteínas dietéticas sobre el comportamiento en perros. J Am Vet Med Assoc (1996) 208:376–9.
107. Hennessy MB, Voith VL, Hawke JL, Young TL, Centrone J, McDowell AL, et al. Efectos de un programa de interacción humana y alteraciones en la composición de la dieta sobre la actividad del eje hipotalámico-pituitario-suprarrenal en perros alojados en un refugio público de animales. J Am Vet Med Assoc (2002) 221:65–71. doi:10.2460/javma.2002.221.65
Resumen de PubMed | Texto completo de CrossRef | Google Académico
108. Bauer JE. La naturaleza esencial de los ácidos grasos omega-3 en la dieta en perros. J Am Vet Med Assoc (2016) 249: 1267–72. doi:10.2460/javma.249.11.1267
109. Zicker SC, Jewell DE, Yamka RM, Milgram NW. Evaluación del aprendizaje cognitivo, la memoria, las funciones psicomotoras, inmunológicas y retinianas en cachorros sanos alimentados con alimentos fortificados con aceite de pescado rico en ácido docosahexaenoico de 8 a 52 semanas de edad. J Am Vet Med Assoc (2012) 241:583–94. doi:10.2460/javma.241.5.583
Resumen de PubMed | Texto completo de CrossRef | Google Académico
110. DeNapoli JS, Dodman NH, Shuster L, Rand WM, Gross KL. Efecto del contenido de proteínas dietéticas y la suplementación con triptófano sobre la agresión de dominancia, la agresión territorial y la hiperactividad en perros. J Am Vet Med Assoc (2000) 217:504–8. doi:10.2460/javma.2000.217.504
Resumen de PubMed | Texto completo de CrossRef | Google Académico
111. Sechi S, Di Cerbo A, Canello S, Guidetti G, Chiavolelli F, Fiore F, et al. Efectos en perros con trastornos del comportamiento de una dieta nutracéutica comercial sobre el estrés y los parámetros neuroendocrinos. Veterinario Rec (2017) 180:18. doi:10.1136/vr.103865
Resumen de PubMed | Texto completo de CrossRef | Google Académico
112. Middelbos IS, Vester Boler BM, Qu A, White BA, Swanson KS, Fahey GC Jr. Caracterización filogenética de comunidades microbianas fecales de perros alimentados con dietas con o sin fibra dietética suplementaria utilizando 454 pirosecuenciación. PLoS One (2010) 5:e9768. doi:10.1371/journal.pone.0009768
Resumen de PubMed | Texto completo de CrossRef | Google Académico
113. Panasevich MR, Kerr KR, Dilger RN, Fahey GC Jr, Guerin-Deremaux L, Lynch GL, et al. Modulación del microbioma fecal de perros adultos sanos mediante la inclusión de fibra de patata en la dieta. Br J Nutr (2015) 113:125–33. doi:10.1017/S0007114514003274
Resumen de PubMed | Texto completo de CrossRef | Google Académico
114. Suchodolski J, Olson E, Honneffer J, Guard B, Blake A, Mustafa A, et al. Efectos de una dieta de proteínas hidrolizadas y metronidazol sobre el microbioma fecal y el metaboloma en perros sanos. J Vet Intern Med (2016) 30:1455. doi:10.1111/jvim.13952
115. Wakshlag JJ, Simpson KW, Struble AM, Dowd SE. Las características fecales negativas se asocian con alteraciones del pH y de la flora fecal durante el cambio en la dieta del perro. Int J Appl Res Vet Med (2011) 9:278–83.
116. Stulberg E, Fravel D, Proctor LM, Murray DM, LoTempio J, Chrisey L, et al. Una evaluación de la investigación del microbioma de EE. UU. Nat Microbiol (2016) 1:15015. doi:10.1038/nmicrobiol.2015.15
Resumen de PubMed | Texto completo de CrossRef | Google Académico
117. Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, et al. Diversidad de la flora microbiana intestinal humana. Science (2005) 308:1635–8. doi:10.1126/science.1110591
Resumen de PubMed | Texto completo de CrossRef | Google Académico
118. Lozupone C, Knight R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol (2005) 71:8228–35. doi:10.1128/AEM.71.12.8228-8235.2005
Resumen de PubMed | Texto completo de CrossRef | Google Académico
119. Foster JA, Moore JH, Gilbert JA, Bunge J. Microbiome studies: analytical tools and techniques. En: Altman RB, Dunker AK, Hunter L, Murray TA, Klein TE, editores. Bioinformática 2012. Kohala Coast, Hawái: World Scientific (2012). págs. 200–2.
120. Rintala A, Pietila S, Munukka E, Eerola E, Pursiheimo JP, Laiho A, et al. Los resultados del análisis de la microbiota intestinal dependen en gran medida de la región diana del gen 16S rRNA, mientras que el impacto de la extracción de ADN es menor. J Biomol Tech (2017) 28:19–30. doi:10.7171/jbt.17-2801-003
Resumen de PubMed | Texto completo de CrossRef | Google Académico
121. van Dijk EL, Auger H, Jaszczyszyn Y, Thermes C. Ten years of next-generation sequencing technology. Tendencias Genet (2014) 30:418–26. doi:10.1016/j.tig.2014.07.001
Resumen de PubMed | Texto completo de CrossRef | Google Académico
122. Hooda S, Minamoto Y, Suchodolski JS, Swanson KS. Estado actual del conocimiento: el microbioma gastrointestinal canino. Anim Health Res Rev (2012) 13:78–88. doi:10.1017/S1466252312000059
Resumen de PubMed | Texto completo de CrossRef | Google Académico
123. Suchodolski JS, Ruaux CG, Steiner JM, Fetz K, Williams DA. Evaluación de la variación cualitativa en la microflora bacteriana entre compartimentos del tracto intestinal de perros mediante el uso de una técnica de huella molecular. Am J Vet Res (2005) 66:1556–62. doi:10.2460/ajvr.2005.66.1556
Resumen de PubMed | Texto completo de CrossRef | Google Académico
124. Kinross JM, Darzi AW, Nicholson JK. Interacciones microbioma-huésped intestinal en la salud y la enfermedad. Genoma Med (2011) 3:14. doi:10.1186/gm228
Resumen de PubMed | Texto completo de CrossRef | Google Académico
125. Palm NW, de Zoete MR, Flavell RA. Interacciones inmuno-microbiota en la salud y la enfermedad. Clin Immunol (2015) 159:122–7. doi:10.1016/j.clim.2015.05.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
126. Sekirov I, Russell SL, Antunes LC, Finlay BB. La microbiota intestinal en la salud y la enfermedad. Physiol Rev (2010) 90:859–904. doi:10.1152/physrev.00045.2009
Resumen de PubMed | Texto completo de CrossRef | Google Académico
127. Kanauchi O, Matsumoto Y, Matsumura M, Fukuoka M, Bamba T. Los efectos beneficiosos de la microflora, especialmente los anaerobios obligados, y sus productos sobre el ambiente colónico en la enfermedad inflamatoria intestinal. Curr Pharm Des (2005) 11:1047–53. doi:10.2174/1381612053381675
Resumen de PubMed | Texto completo de CrossRef | Google Académico
128. Torre E, Tello M. Factores que influyen en la excreción fecal de Campylobacter jejuni en perros sin diarrea. Am J Vet Res (1993) 54:260–2.
129. Hildebrand F, Nguyen TL, Brinkman B, Yunta RG, Cauwe B, Vandenabeele P, et al. Los enterotipos asociados a la inflamación, el genotipo del huésped, la jaula y los efectos interindividuales impulsan la variación de la microbiota intestinal en ratones de laboratorio comunes. Genoma Biol (2013) 14:R4. doi:10.1186/gb-2013-14-1-r4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
130. Dougal K, de la Fuente G, Harris PA, Girdwood SE, Pinloche E, Geor RJ, et al. Caracterización de la comunidad bacteriana fecal en caballos adultos y ancianos alimentados con una dieta alta en fibra, alta en aceite o alta en almidón mediante pirosecuenciación 454. PLoS One (2014) 9:e87424. doi:10.1371/journal.pone.0087424
Resumen de PubMed | Texto completo de CrossRef | Google Académico
131. Mariat D, Firmesse O, Levenez F, Guimaraes V, Sokol H, Dore J, et al. La relación Firmicutes/Bacteroidetes de la microbiota humana cambia con la edad. BMC Microbiol (2009) 9:123. doi:10.1186/1471-2180-9-123
Resumen de PubMed | Texto completo de CrossRef | Google Académico
132. Simpson JM, Martineau B, Jones WE, Ballam JM, Mackie RI. Caracterización de poblaciones bacterianas fecales en caninos: efectos de la edad, raza y fibra dietética. Microb Ecol (2002) 44:186–97. doi:10.1007/s00248-002-0001-z
Resumen de PubMed | Texto completo de CrossRef | Google Académico
133. Song SJ, Lauber C, Costello EK, Lozupone CA, Humphrey G, Berg-Lyons D, et al. Los miembros de la familia que cohabitan comparten microbiota entre sí y con sus perros. Elife (2013) 2:e00458. doi:10.7554/eLife.00458
Resumen de PubMed | Texto completo de CrossRef | Google Académico
134. Peterson LW, Artis D. Células epiteliales intestinales: reguladores de la función de barrera y la homeostasis inmune. Nat Rev Immunol (2014) 14:141–53. doi:10.1038/nri3608
Resumen de PubMed | Texto completo de CrossRef | Google Académico
135. Koh A, De Vadder F, Kovatcheva-Datchary P, Backhed F. De la fibra dietética a la fisiología del huésped: ácidos grasos de cadena corta como metabolitos bacterianos clave. Celda (2016) 165:1332–45. doi:10.1016/j.cell.2016.05.041
Resumen de PubMed | Texto completo de CrossRef | Google Académico
136. Mosca A, Leclerc M, Hugot JP. Diversidad de la microbiota intestinal y enfermedades humanas: ¿deberíamos reintroducir depredadores clave en nuestro ecosistema? Microbiol frontal (2016) 7:455. doi:10.3389/fmicb.2016.00455
Resumen de PubMed | Texto completo de CrossRef | Google Académico
137. Honneffer JB, Minamoto Y, Suchodolski JS. Alteraciones de la microbiota en la inflamación gastrointestinal aguda y crónica de gatos y perros. World J Gastroenterol (2014) 20:16489–97. doi:10.3748/wjg.v20.i44.16489
Resumen de PubMed | Texto completo de CrossRef | Google Académico
138. Minamoto Y, Otoni CC, Steelman SM, Buyukleblebici O, Steiner JM, Jergens AE, et al. Alteración de la microbiota fecal y de los perfiles de metabolitos séricos en perros con enfermedad inflamatoria intestinal idiopática. Gut Microbes (2015) 6:33–47. doi:10.1080/19490976.2014.997612
Resumen de PubMed | Texto completo de CrossRef | Google Académico
139. AlShawaqfeh MK, Wajid B, Minamoto Y, Markel M, Lidbury JA, Steiner JM, et al. Un índice de disbiosis para evaluar los cambios microbianos en muestras fecales de perros con enteropatía inflamatoria crónica. FEMS Microbiol Ecol (2017) 93:fix136. doi:10.1093/femsec/fix136
Resumen de PubMed | Texto completo de CrossRef | Google Académico
140. Lyte M, Vulchanova L, Brown DR. Estrés en la superficie intestinal: catecolaminas e interacciones mucosa-bacteria. Cell Tissue Res (2011) 343:23–32. doi:10.1007/s00441-010-1050-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
141. Gagne JW, Wakshlag JJ, Simpson KW, Dowd SE, Latchman S, Brown DA, et al. Efectos de un simbiótico sobre la calidad fecal, las concentraciones de ácidos grasos de cadena corta y el microbioma de perros de trineo sanos. BMC Vet Res (2013) 9:246. doi:10.1186/1746-6148-9-246
Resumen de PubMed | Texto completo de CrossRef | Google Académico
142. Kuzmuk KN, Swanson KS, Tappenden KA, Schook LB, Fahey GC Jr. La dieta y la edad afectan la morfología intestinal y las concentraciones del producto final fermentativo del intestino grueso en perros adultos mayores y jóvenes. J Nutr (2005) 135:1940–5. doi:10.1093/jn/135.8.1940
Resumen de PubMed | Texto completo de CrossRef | Google Académico
143. Middelbos IS, Fastinger ND, Fahey GC Jr. Evaluación de oligosacáridos fermentables en dietas alimentadas a perros en comparación con los estándares de fibra. J Anim Sci (2007) 85:3033–44. doi:10.2527/jas.2007-0080
Resumen de PubMed | Texto completo de CrossRef | Google Académico
144. Swanson KS, Grieshop CM, Flickinger EA, Bauer LL, Chow J, Wolf BW, et al. Los fructooligosacáridos y Lactobacillus acidophilus modifican las poblaciones microbianas intestinales, las digestibilidades de nutrientes totales del tracto y las concentraciones de catabolito de proteína fecal en perros adultos sanos. J Nutr (2002) 132:3721–31. doi:10.1093/jn/132.12.3721
145. Rastall RA. Bacterias en el intestino: amigos y enemigos y cómo alterar el equilibrio. J Nutr (2004) 134:2022S–6S. doi:10.1093/jn/134.8.2022S
Resumen de PubMed | Texto completo de CrossRef | Google Académico
146. Superchi P, Saleri R, Ossiprandi MC, Riccardi E, Passaglia E, Cavalli V, et al. La suplementación dietética de zeolita natural (chabazita/phillipsita) influye en la microbiota fecal y el estado oxidante de los perros de trabajo. Ital J Anim Sci (2016) 16:115–21. doi:10.1080/1828051X.2016.1261008
147. Swanson KS, Grieshop CM, Flickinger EA, Bauer LL, Healy HP, Dawson KA, et al. Los fructooligosacáridos y mananooligosacáridos suplementarios influyen en la función inmune, las digestibilidades de nutrientes ileal y del tracto total, las poblaciones microbianas y las concentraciones de catabolitos proteicos en el intestino grueso de los perros. J Nutr (2002) 132:980–9. doi:10.1093/jn/132.5.980
148. Venable EB, Fenton KA, Braner VM, Reddington CE, Halpin MJ, Heitz SA, et al. Efectos del manejo de la alimentación sobre la microbiota cecal equina. J Equine Vet Sci (2017) 49: 113–21. doi:10.1016/j.jevs.2016.09.010
149. Sandhu KV, Sherwin E, Schellekens H, Stanton C, Dinan TG, Cryan JF. Alimentación del eje microbiota-intestino-cerebro: dieta, microbioma y neuropsiquiatría. Transl Res (2017) 179:223–44. doi:10.1016/j.trsl.2016.10.002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
150. Foster JA, Rinaman L, Cryan JF. Estrés y eje intestino-cerebro: regulación por el microbioma. Neurobiol Stress (2017) 7: 124-36. doi:10.1016/j.ynstr.2017.03.001
151. Flegel C, Manteniotis S, Osthold S, Hatt H, Gisselmann G. Perfil de expresión de receptores olfativos ectópicos determinados por secuenciación profunda. PLoS One (2013) 8:e55368. doi:10.1371/journal.pone.0055368
Resumen de PubMed | Texto completo de CrossRef | Google Académico
152. Priori D, Colombo M, Clavenzani P, Jansman AJ, Lalles JP, Trevisi P, et al. El receptor olfativo OR51E1 está presente a lo largo del tracto gastrointestinal de los cerdos, se colocaliza con las células enteroendocrinas y está modulado por la microbiota intestinal. PLoS One (2015) 10:e0129501. doi:10.1371/journal.pone.0129501
Resumen de PubMed | Texto completo de CrossRef | Google Académico
153. Dugue GP, Mainen ZF. Cómo la serotonina gates flujo de información olfativa. Nat Neurosci (2009) 12:673–5. doi:10.1038/nn0609-673
154. Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, et al. Las bacterias autóctonas de la microbiota intestinal regulan la biosíntesis de serotonina del huésped. Cell (2015) 161:264–76. doi:10.1016/j.cell.2015.02.047
Resumen de PubMed | Texto completo de CrossRef | Google Académico
155. Koskinen K, Reichert JL, Hoier S, Schachenreiter J, Duller S, Moissl-Eichinger C, et al. El microbioma nasal refleja y potencialmente da forma a la función olfativa. Sci Rep (2018) 8:1296. doi:10.1038/s41598-018-19438-3
Resumen de PubMed | Texto completo de CrossRef | Google Académico
156. Tress B, Dorn ES, Suchodolski JS, Nisar T, Ravindran P, Weber K, et al. Microbioma bacteriano de la nariz de perros sanos y perros con enfermedad nasal. PLoS One (2017) 12:e0176736. doi:10.1371/journal.pone.0176736
Resumen de PubMed | Texto completo de CrossRef | Google Académico
157. Isaiah A, Hoffmann AR, Kelley R, Mundell P, Steiner JM, Suchodolski JS. Caracterización de la microbiota nasal y oral de perros de detección. PLoS One (2017) 12:e0184899. doi:10.1371/journal.pone.0184899
Resumen de PubMed | Texto completo de CrossRef | Google Académico
158. Riera CE, Tsaousidou E, Halloran J, Follett P, Hahn O, Pereira MMA, et al. El sentido del olfato afecta la salud metabólica y la obesidad. Cell Metab (2017) 26:198–211.e5. doi:10.1016/j.cmet.2017.06.015
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Cita: Jenkins EK, DeChant MT and Perry EB (2018) When the Nose Doesn’t Know: Canine Olfactory Function Associated With Health, Management, and Potential Links to Microbiota. Frente. Vet. Sci. 5:56. doi: 10.3389/fvets.2018.00056
Recibido: 28 de noviembre de 2017; Aprobado: 05 Marzo 2018;
Publicado: 29 Marzo 2018
Editado por:
Cynthia M. Otto, Universidad de Pensilvania, Estados Unidos
Revisado por:
Mitsuaki Ohta, Universidad de Agricultura de Tokio, Japón
Federica Pirrone, Università degli Studi di Milano, Italia
Derechos de autor: © 2018 Jenkins, DeChant y Perry. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Erin B. Perry, erin.perry@siu.edu
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía