
Niveles de calcio y colecalciferol en gallinas ponedoras

Niveles de calcio y colecalciferol en gallinas ponedoras de fase tardía: efectos sobre los rasgos productivos, la calidad del huevo, la bioquímica sanguínea y las respuestas inmunes
 Youssef A. Attia1*,
Youssef A. Attia1*,  Mohammed A. Al-Harthi1 y
Mohammed A. Al-Harthi1 y  Hayam M. Abo El-Maaty2*
Hayam M. Abo El-Maaty2*- 1Departamento de Agricultura, Facultad de Ciencias Ambientales, Universidad Rey Abdulaziz, Jeddah, Arabia Saudita
- 2Departamento de Producción Avícola, Facultad de Agricultura, Universidad de Mansoura, Mansoura, Egipto
Los rasgos productivos y la inmunidad en las gallinas ponedoras disminuyen drásticamente durante la fase tardía de la puesta debido al envejecimiento, lo que afecta negativamente el metabolismo y el estado hormonal de los animales. La influencia de los niveles de Ca (3.5, 4.0 y 4.5%) y / o colecalciferol [Vitamina D3 (VD3) suplementación (dieta de 800, 1,000 y 1,200 UI / kg o como total de 3,800, 4,000 y 4,200 IC VD3) sobre el rendimiento, la calidad del huevo, la bioquímica sanguínea y la inmunidad de las ponedoras de huevo marrón. Trescientas sesenta ponedoras de huevos H&N Brown (60 semanas de edad) fueron asignadas al azar en nueve tratamientos nutricionales de cinco réplicas (jaulas) de ocho gallinas cada una. La dieta de control en este experimento contenía un nivel de Ca del 3,5% con 800 UI de VD3. La adición de VD3 a 1.000 y 1.200 UI a 3,5 y 4% de Ca las dietas significativamente (P ≤ 0,05) aumentaron significativamente la tasa de puesta, la masa de huevos y la tasa de conversión alimenticia (FCR) en comparación con la dieta de control en 3,5% y 800 U de VD3. Además de esto, la adición de VD3 a 800 y 1,200 UI a 3.5% de nivel de Ca, las dietas mejoraron la puntuación de la unidad de Haugh. Se observaron resultados similares en las mediciones de calidad de la cáscara de huevo y la ceniza de tibia. Aumentar la concentración de Ca de 3,5 a 4 y 4,5% y aumentar la VD3 niveles de 800 a 1,000 o 1,200 UI aumentaron significativamente y de manera similar la proteína sérica total y la globulina. Además, VD3 a 1.000 UI aumentó la albúmina sérica, en comparación con 800 UI. El aumento del nivel de Ca aumentó la IgA, y los niveles de 4 y 4,5% de Ca aumentaron de manera similar la IgG y la α-2 globulina en comparación con la dieta de 3,5% de Ca. VD3 adición a 1.200 UI a la dieta de 4% Ca aumentó significativamente la γ-globulina en comparación con 1.000 UI, pero disminuyó la β-globulina. El aumento del nivel de Ca al 4% redujo significativamente los triglicéridos séricos, y la lipoproteína de muy baja densidad y la relación triglicéridos/lipoproteínas de alta densidad disminuyeron con dietas de nivel de 4 y 4,5% de Ca. El aumento del nivel de Ca causó un aumento gradual de la catalasa, que aumentó notablemente con VD3 suplementación a 1.200 UI. El estrógeno plasmático aumentó considerablemente con VD3 suplementación al 3,5% Ca, pero los niveles de hormona paratiroidea no se vieron afectados. En conclusión, el aumento de los niveles de Ca en la dieta de las gallinas ponedoras al 4% durante la fase de producción tardía podría ser una herramienta útil para mejorar el rendimiento de la puesta, la calidad de la cáscara del huevo, la puntuación de la unidad de Haugh y el estado fisiológico e inmunológico. Además, VD3 con una dieta de 1.000 UI/kg a 3,5% de Ca mejoró el rendimiento de las gallinas alimentadas con 3,5% de Ca, lo que demuestra que el impacto potencial de VD3 depende de las concentraciones de Ca.
Introducción
A nivel mundial, tanto los huevos como la carne son fuentes aceptables y económicas de proteínas para la nutrición humana (1). Los híbridos modernos de gallinas ponedoras producen más de 320 huevos por año. Los huevos contienen 10% de cáscara, y la cáscara de huevo contiene 40% de Ca; por lo tanto, las gallinas requieren una cantidad considerable de Ca para la formación de cáscaras de huevo. La cáscara de huevo protege los huevos y es responsable de mantener su calidad interna desde la oviposición hasta su uso por parte de los consumidores (2). Obviamente, la calidad del huevo y particularmente la calidad de la cáscara disminuyen con la edad de las gallinas ponedoras durante la fase tardía (60 semanas o más) de la producción (3, 4). La disminución en la calidad de la cáscara de huevo durante la última etapa de producción podría determinarse por un aumento en el tamaño del huevo, reducciones en el metabolismo de nutrientes, principalmente de Ca, y en las hormonas reproductivas, especialmente estrógenos (5). El calcio es un componente crítico en la nutrición de las gallinas ponedoras, y su disponibilidad depende de la absorción intestinal de la dieta. Aún así, el esqueleto es una fuente esencial de Ca durante la noche cuando la absorción intestinal ha cesado (5, 6). El aumento de la rotura y la pérdida de huevos son los mayores problemas en la industria del huevo de mesa, causando pérdidas económicas sustanciales, posiblemente debido a la baja calidad de la cáscara del huevo (6, 7). No solo se produce una baja calidad de la cáscara de huevo, sino también la pérdida de resistencia ósea durante el período de puesta tardía (5, 8). La evidencia reciente ha demostrado que las proteínas están integradas en la regulación de las proteínas que impulsan la calcificación de la calidad y la función de la cáscara en las gallinas ponedoras (9, 10).
El consumo adecuado de Ca por parte de las gallinas ponedoras es fundamental para garantizar una calidad confiable de la cáscara de huevo. Sin embargo, el impacto del nivel de Ca en la dieta en la producción y calidad de los huevos es contradictorio en la literatura, y algunos datos se informaron hace varias décadas. Además, ha habido una mejora significativa en la tasa de puesta debido al progreso genético y al uso de herramientas biotecnológicas en la cría de animales, junto con la conciencia de los consumidores sobre la calidad del huevo, que requiere mayor atención al producir huevos funcionales (5, 11). En la literatura, los requisitos de Ca para las gallinas ponedoras dependen de la edad, la fase de producción, la temperatura ambiental, la cepa y las concentraciones de Ca, P y vitamina D (VD) en la dieta (5). Por ejemplo, los rasgos cualitativos del peso del huevo y la tibia de las gallinas ponedoras alimentadas con 3,5% de Ca (3,77 g / d) fueron más altos que los alimentados con 3,0% (3,29 g / d), pero el aumento del nivel de Ca al 4% (4,31 g / d) no afectó la calidad de la cáscara del huevo o el peso de la tibia (12). Además, hace más de dos décadas se recomendaba un requerimiento diario de Ca de 3,75 g (13, 14). Por otro lado, el porcentaje óptimo de Ca para la formación de cáscara de huevo fue citado como 4,73% (15). Las directrices recientes de las empresas reproductoras recomiendan una ingesta diaria de Ca de 4,10 g durante la fase temprana de producción de huevos (19 semanas), y esto aumenta gradualmente con la edad hasta alcanzar el 4,4 g % para las ponedoras Lohaman Brown-Classic en la fase tardía de producción a las 65 semanas o más (16). Por otro lado, el NRC (17) recomienda una ingesta diaria de 3,75 g para las capas de cáscara de huevo blanca y marrón.
Colecalciferol (VD)3) y sus dos metabolitos son esenciales para la calcificación de la cáscara del huevo y el hueso y tienen influencias hormonales e inmunes (3, 4, 18). VD3 Se puede formar en la piel de la dermis y la epidermis a partir de 7-dihidrocolesterol bajo luz ultravioleta o se puede ofrecer como un componente dietético (3, 19). Hoy en día, las gallinas ponedoras se crían bajo el sistema extensivo y se alojan en jaulas cerradas para diferentes perspectivas, como la creciente intensidad de la producción, las ganancias agrícolas, la producción de huevos limpios y la mejora de las condiciones de cría. Las gallinas alojadas en estas condiciones no están expuestas a la luz natural adecuada para transformar el 7-dihidrocolesterol a niveles apropiados para la síntesis suficiente de VD.3. Por lo tanto, VD3 generalmente se agrega a los feeds de capas; Estos son críticos para la homeostasis del calcio y para mantener el rendimiento de puesta, la calcificación ósea y la formación de cáscaras de huevo (5, 14, 20). La biosíntesis de la forma activa 1, 25-dihidroxicolecalciferol (1,25-(OH)2D3) de VD3 fue revisado por Geng et al. (20). Esto ocurrió en el hígado y el riñón en dos pasos, mediados por enzimas 25-hidroxilasa1 y α-hidroxilasa (5).
El nivel común de VD3 en las dietas de ponedoras es de aproximadamente 2,200 UI / kg (21, 22), mientras que las guías comerciales de criadores de huevos recomiendan 2,500 UI / kg en dietas (14), y 3,000-5,000 UI / kg es recomendado por los productores comerciales (18). La proteína de unión a Ca implicada en el transporte activo de Ca a través de la pared intestinal requiere VD3 (18 y 22). En la literatura, VD3 tiene muchos beneficios para la salud, inmunológicos y fisiológicos (5), y es una vitamina vital que desempeña un papel considerable en el desarrollo de la salud muscular, esquelética y en el mantenimiento de la homeostasis del calcio y el fósforo (18, 23, 24). La formación de cáscara de huevo y la salud del hueso en gallinas ponedoras es esencial e implica la integración entre el metabolismo de Ca, P y VD3 (5, 18, 25). Rodriguez-Lecompte et al. (26) y Manolagas et al. (27) han reportado que VD3 o la forma activa de VD3, 25-(OH)D, ambos tienen fuertes propiedades inmunomoduladoras con una ayuda final gradual de las células T (Th2).
Las gallinas ponedoras híbridas comerciales producen huevos a un ritmo mayor debido al aumento del potencial genético y la mejora en las estrategias agrícolas y nutricionales. Por consiguiente, en conjunto, los factores que afectan a las necesidades de Ca y VD3 de las gallinas ponedoras necesitan más investigación, particularmente durante la fase tardía de la producción, debido a una disminución en el rendimiento de la puesta, la calidad de los huevos y las cáscaras, y la adaptación fisiológica e inmunológica. Por lo tanto, planteamos la hipótesis de que el Ca y el VD3 las necesidades de las gallinas ponedoras aumentaron durante la última etapa de producción y aumentaron el Ca y/o VD3 puede mejorar la producción y la calidad de los huevos y los beneficios para los criadores de gallinas ponedoras. Por lo tanto, este trabajo examina la integración entre diferentes concentraciones de Ca (3.5, 4.0 y 4.5%) y a (VD).3) Dieta de 800, 1,000, 1,200 UI / kg sobre los rasgos productivos, la bioquímica de la sangre y la respuesta inmune de las gallinas ponedoras durante la fase tardía de la puesta de huevos marrones.
Materiales y métodos
El comité científico del Departamento de Producción Avícola, Facultad de Agricultura, Universidad de Mansoura, aprobó el presente experimento. El número de protocolo era DF-715-155-1441 H. El comité recomendó que el cuidado y manejo del animal mantuviera los derechos y el bienestar y minimizara el estrés (Directiva 2010/63/UE).
Gallinas ponedoras y cría
La investigación se realizó en la Unidad de Investigación Avícola, Centro Qalabsho de Investigaciones y Experimentos Agrícolas, Facultad de Agricultura, Universidad de Mansour, Egipto, de julio a septiembre de 2018. Este es el período más caluroso del año en Egipto; la humedad relativa y la temperatura ambiente durante el período experimental oscilaron entre 54 y 70% y 22.8 y 35.6 ° C, respectivamente. Las aves estuvieron expuestas a condiciones climáticas cálidas, y las gallinas mostraron síntomas de altas temperaturas ambiente, como acostarse en jaulas, aletear las alas y jadear. Sin embargo, no discutimos el estrés por calor en la sección Resultados y Discusión, debido a la falta de un grupo de control para el estrés por calor que se mantuvo bajo temperatura óptima (25 ° C).
360 ponedoras H&N Brown Nick (60 semanas) fueron asignadas al azar en nueve tratamientos, que consistían en cinco réplicas de ocho gallinas. Las gallinas se criaban en jaulas de ponedoras abiertas (ocho gallinas/jaula con un comedero y dos pezones); El tamaño de cada jaula era de 60 × 120 × 50 cm. Las gallinas fueron sometidas a un programa de luz de 16:8 horas de ciclo de luz/oscuridad y recibieron alimento de puré y agua del grifo ad libitum.
Dietas experimentales
Nueve dietas experimentales, que consisten en tres niveles de Ca (3.5, 4.0 y 4.5%) con tres niveles de vitamina D suplementada3 a la dosis de una dieta de 800, 1,000, 1,200 UI / kg o como total de 3,800, 4,000 y 4,200 IC VD3 fueron alimentados durante el período experimental de 60 a 72 semanas de edad (Tabla 1). La dieta de control en este experimento fue que contenía un nivel de Ca del 3,5% con 800 UI de VD.3.
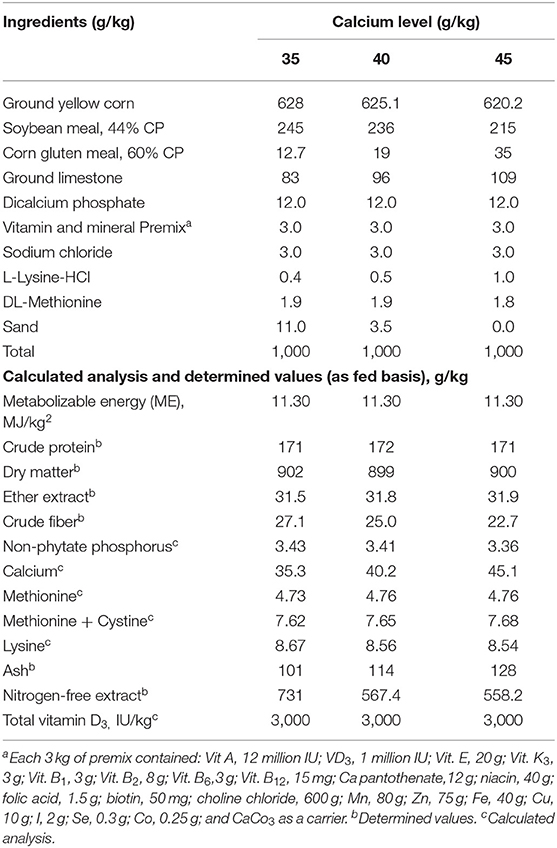 TABLA 1. Formulación y análisis próximos de las dietas experimentales (g/kg) sobre base de materia seca alimentadas a cepas de ponedoras de huevo marrón de 60 a 72 semanas de edad.
TABLA 1. Formulación y análisis próximos de las dietas experimentales (g/kg) sobre base de materia seca alimentadas a cepas de ponedoras de huevo marrón de 60 a 72 semanas de edad.
El VD total3 Las cantidades en las dietas probadas fueron 3,800, 4,000 y 4,200 UI / kg. El VD en los piensos se encuentra en forma de VD2, que tiene una pequeña actividad para las aves de corral (17); por lo tanto, no se tuvo en cuenta en el cálculo del VD total3 en la dieta. El colecalciferol VD3 era de grado alimenticio en polvo, un producto de Polifar Group Limited, Nanjing, Jiangsu, China. Las dietas fueron formuladas de acuerdo con NRC (17). Las dietas cumplieron o superaron las necesidades de nutrientes especificadas en la nueva guía de manejo de H&N (28). Las dietas experimentales se analizaron utilizando los métodos oficiales de análisis (29) materia seca, proteína cruda, extracto de éter, fibra cruda y ceniza.
Rendimiento de colocación
Los registros diarios de la tasa de puesta (LR), la ingesta de alimento (DFI), el peso del huevo (EW), la masa del huevo (EM) y la tasa de mortalidad se registraron y utilizaron para calcular los índices de producción en un período de 28 días de la siguiente manera:
El cambio de peso corporal (BWC) también se estimó a partir de las diferencias entre los pesos corporales inicial y final durante el período de prueba.
Mediciones de la calidad del huevo
Los huevos (30 huevos por tratamiento) se seleccionaron al azar a las 72 semanas de edad para representar por igual todas las réplicas y se utilizaron en la determinación de las características de calidad del huevo exterior e interior, según lo citado por Burke y Attia (30) y Attia et al. (31, 32). Estas características involucraron EW y sus componentes comparativos (albúmina, yema y cáscara), unidades Haugh, puntaje de color de la yema (YCS), índice de yema (YI), grosor de la cáscara (ST) e índice de forma de huevo (ESI). La calidad de la cáscara del huevo se midió en tres puntos diferentes de la cáscara en el punto medio y en los dos extremos de los huevos, y los valores se utilizaron para el cálculo del valor medio del grosor de la cáscara del huevo. El peso de la cáscara por unidad de superficie (SWUSA) se determinó dividiendo el peso de la cáscara por el área de superficie del huevo (ESA; cm2). Estas mediciones se realizaron según lo informado por Burke y Attia (30) y Attia et al. (31, 32).
Las imágenes de microscopía electrónica de barrido (SEM) de las cáscaras de huevo se tomaron de acuerdo con Stefanello et al. (33) utilizando dos muestras de la cáscara de huevo de cada huevo recolectadas a las 72 semanas de edad. El número de cáscaras de huevo fue una por réplica de cada uno de los nueve tratamientos. La cáscara de huevo libre de membrana de cáscara se obtuvo después de romper los huevos. La membrana de la cubierta se eliminó por inmersión de las muestras en una solución de hidróxido de sodio al 0,15%, cloruro de sodio al 4,12% e hipoclorito de sodio al 6%. Se utilizó agua del grifo para lavar las conchas, que luego se secaron al aire a temperatura ambiente (27 ° C). Las imágenes se realizaron utilizando 0,5 cm2 de la cáscara de huevo sin membrana de cada réplica de cada tratamiento utilizando un instrumento Shimadzu SS-550 Super Scan (Shimadzu Corporation, EVISA, Kyoto, Japón).
Parámetros sanguíneos
Se recolectaron cinco muestras de sangre de mililitros de la vena del ala de seis gallinas por grupo de 72 semanas en dos tubos de sangre, con y sin heparina. Las gallinas utilizadas para medir los componentes de la sangre se seleccionaron con huevos de cáscara dura en el útero. Las muestras de sangre se centrifugaron a 1.716 g durante 15 min para separar el plasma y el suero, que se mantuvieron a 20 °C hasta el análisis. Se determinaron proteína sérica total, albúmina, globulina, triglicéridos (Trig.), colesterol total (CT) y lipoproteína-colesterol de alta densidad (HDL-C). Se midieron el calcio plasmático (Ca) y el P (Pi) inorgánico, la capacidad antioxidante total (TAC), el malondialdehído (MDA), la superóxido dismutasa (SOD) y la catalasa (CAT). Aspartato aminotransferasa (AST); Alanina aminotransferasa (ALT); fosfato alcalino (AlkP); α, β y γ-globulina; también se determinaron inmunoglobulina IgG, IgM, IgA, estrógeno y hormona paratiroidea (PTH). Las mediciones se realizaron utilizando kits de diagnóstico comerciales (34) citados por Attia et al. (35, 36). El título de anticuerpos para la influenza aviar y el virus de la enfermedad de Newcastle se determinó utilizando kits comerciales de ensayo inmunoabsorbente ligado a enzimas (ELISA) según lo citado por Attia et al. (35, 36).
Órganos internos reproductivos
Cinco gallinas fueron seleccionadas al azar de cada tratamiento y sacrificadas de acuerdo con los métodos islámicos (35). Después del sangrado completo, se abrieron las gallinas y se diseccionaron el ovario y el tracto reproductivo. Luego se registró el peso de las partes del ovario y el oviducto (infundíbulo, magnum, istmo y útero) y se estimaron sus proporciones en relación con el peso corporal vivo de las gallinas.
Análisis estadístico
El procesamiento analítico de los resultados se realizó mediante un análisis bidireccional de varianza (Ca, VD3, y su interacción) del procedimiento GLM del Sistema de Análisis Estadístico (37) utilizando la réplica como unidad experimental. Los datos se transformaron en arcseno para normalizar la distribución. Las diferencias entre las medias se probaron mediante la prueba de Student-Newman-Keuls en P < 0,05 (37). Los valores de p entre 0,10 y <0,05 fueron considerados como una tendencia.
Resultados y discusión
Rendimiento de las gallinas ponedoras
La Tabla 2 muestra la influencia de Ca y/o VD3 nivel en los rasgos de producción de las gallinas ponedoras durante la fase tardía. El BW inicial de las gallinas ponedoras no fue diferente entre los grupos experimentales, lo que indica una distribución aleatoria de gallinas; por lo tanto, no se presentaron los datos. Los resultados sugieren que Ca y/o VD3 Los niveles tuvieron un efecto significativo en la mayoría de los rasgos de rendimiento de puesta, excepto en la ingesta de alimento. EW se incrementó gradualmente con el aumento de los niveles de Ca. El efecto de interacción indica que el aumento del nivel de Ca de 3.5 a 4 o 4.5% mejoró de manera similar LR, EM, FCR, EPI y BWG. El aumento del nivel de Ca del 3,5 al 4% no influyó en la EW, pero un aumento adicional al 4,5% aumentó la EW; por lo tanto, la diferencia entre los grupos de 3,5 y 4,5% de Ca fue significativa. Estos resultados revelan que el 4% de Ca fue adecuado para el rendimiento de capas de 60-72 semanas de edad y confirmaron los requisitos de Ca sugeridos (4,1% durante 45-70 semanas de edad) en la guía de gestión de H&N (28) y los de Rao y Roland (38) y Zhang y Coon (39). Estos pueden diferir debido al papel del Ca en la regulación de las hormonas reproductivas y el crecimiento de los ovarios (5). Nascimento et al. (40) encontraron que no había interacción entre las diferentes fuentes de vitamina D en una dieta de 2.000 UI/kg de VD3, 25-(OH)D3, 1,25-(OH)2D3, y cuatro niveles de calcio de 2,85 a 5,25%, con intervalos de 0,80% en todos los rasgos de producción de huevo. Sin embargo, se observó un componente cuadrático significativo del análisis de contraste para el nivel de calcio en LR y FCR, mostrando mejores características productivas a 4,12 y 4,09% Ca, respectivamente. En contraste, el nivel de Ca no afectó el EW. No obstante, la fuente de VD3 influenciado (P < 0.05) LR, FCR y EW, mostrando mejores resultados de gallinas ponedoras con 25-(OH)D3 y colecalciferol (40).
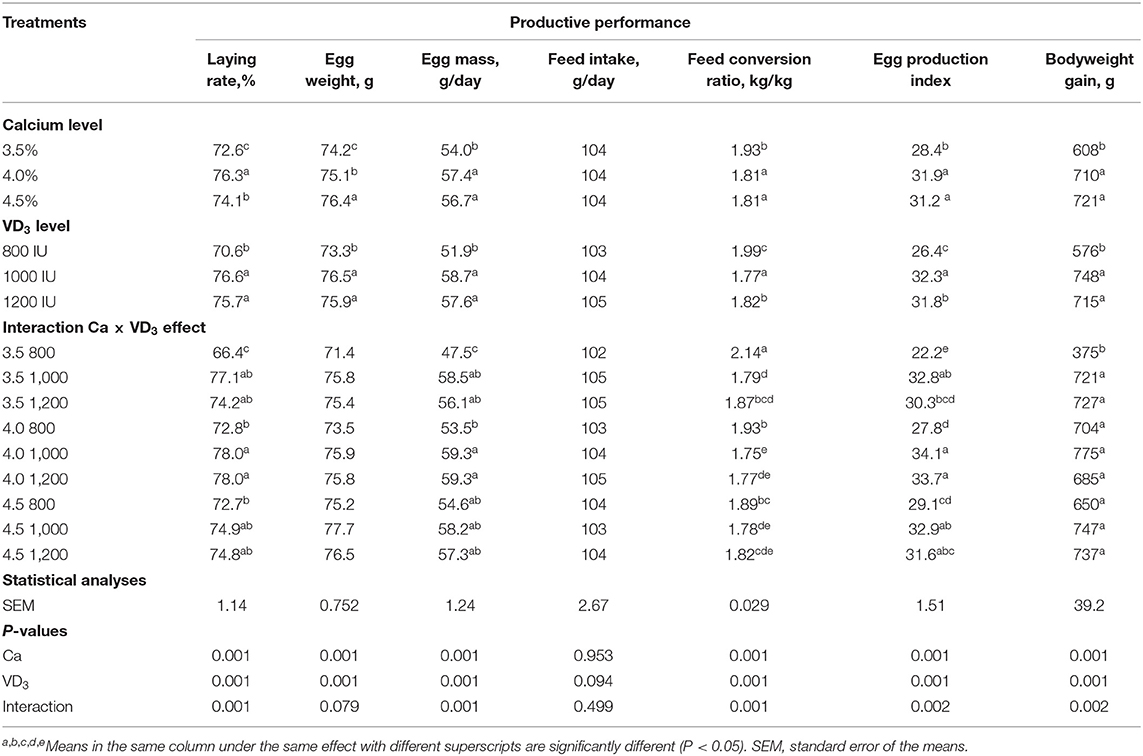 TABLA 2. Rendimiento productivo de las ponedoras de huevos marrones alimentadas con tres niveles de Ca, complementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
TABLA 2. Rendimiento productivo de las ponedoras de huevos marrones alimentadas con tres niveles de Ca, complementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
Se observó que de VD3 en una dieta de 1,000 y 1,200 UI / kg a 3.5 y 4% de Ca las dietas aumentaron significativamente LR, EM y mejoraron la FCR. Además, VD3 la suplementación con una dieta de 1.000 y 1.200 UI/kg al 3,5% de Ca aumentó notablemente el BWG de las gallinas ponedoras (Tabla 2). Aumento de VD3 a 1.000 UI aumentó significativamente la EW, y el efecto fue saturado a este nivel. La adición de VD3 a 1.000 y 1.200 UI a diferentes niveles de Ca aumentó el EPI de las gallinas en diferentes niveles de Ca, excepto para las gallinas alimentadas con 4,5% de Ca suplementadas con una dieta de 1.200 UI / kg. Estos resultados revelan que la suplementación con 1.000 UI de VD3/kg dieta o un total de 4.000 UI/kg es adecuado para los rasgos de producción de huevos de gallinas alimentadas con un nivel de 3,5% de Ca.
Las presentes observaciones indican que VD3 contenido en la dieta que contiene 3.5% con un total de VD3 a 3.000 UI/kg (Tabla 1) no fue adecuado para las gallinas ponedoras H&N Brown en la fase tardía. Cabe mencionar que la guía de gestión nutricional de H&N Brown recomienda 2.500 UI para la colocación durante el período de puesta sin recomendaciones y/o consideraciones específicas para cada etapa de la puesta. Además, 3,000–5,000 UI de VD3 para gallinas ponedoras durante diferentes etapas de producción se recomendó (18). Es evidente que a medida que el metabolismo de los nutrientes disminuye con la edad (5, 41), el VD3 Los requisitos deben volver a investigarse en relación con la etapa de colocación teniendo en cuenta el impacto del envejecimiento en la VD.3 requisitos y debido a VD3 y sus metabolitos desempeñan un papel vital en la absorción, deposición y excreción de Ca (5, 21, 42). Asimismo, otros autores (5, 24, 27) mostraron hallazgos similares. El VD3 es esencial para la formación de proteínas unidas a Ca, que están involucradas en el transporte activo de Ca a través del intestino (28), y en la lipoproteína, el precursor de la formación de la yema (5, 20, 21).
Rasgos de calidad del huevo
Los resultados indican que Ca y la interacción entre Ca y VD3 tiene efectos significativos en los rasgos de calidad del huevo (Tabla 3).
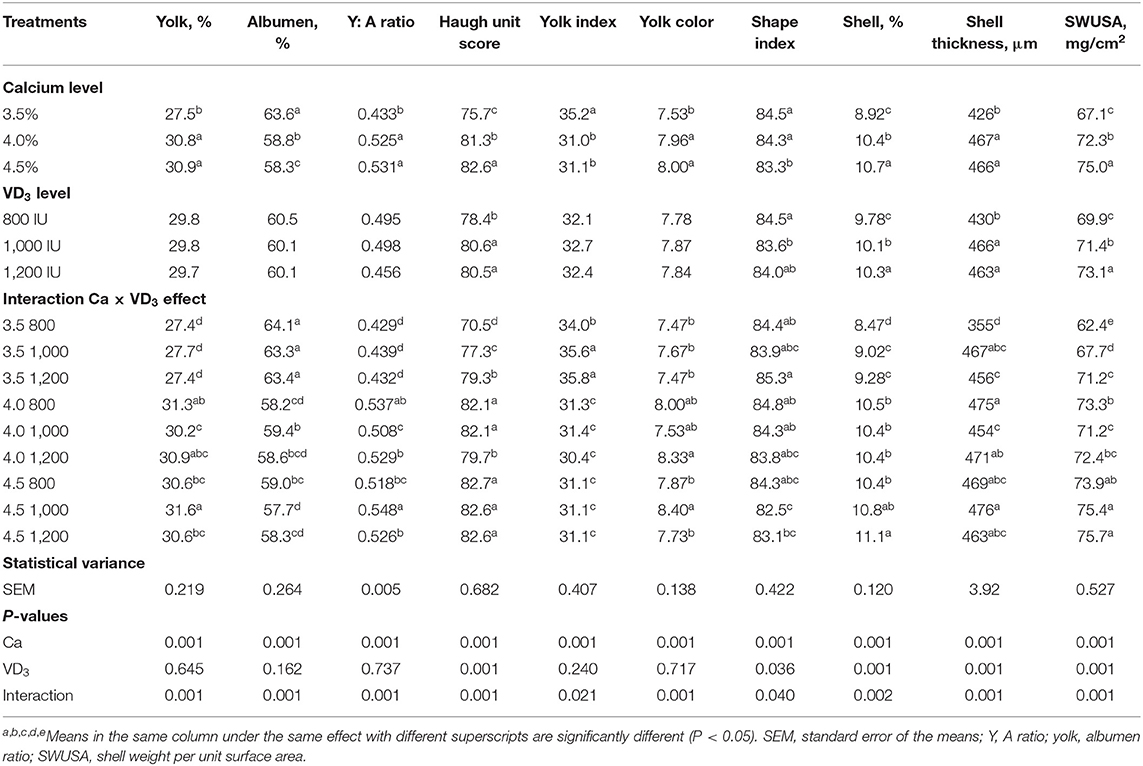 TABLA 3. Rasgos exteriores e interiores de la calidad del huevo marrón Ponedoras de huevo alimentadas con tres niveles de Ca suplementado con tres niveles de VD3 de 60 a 72 semanas de edad.
TABLA 3. Rasgos exteriores e interiores de la calidad del huevo marrón Ponedoras de huevo alimentadas con tres niveles de Ca suplementado con tres niveles de VD3 de 60 a 72 semanas de edad.
Efectos sustanciales de VD3 solo se registraron la puntuación de la unidad Haugh, el índice de forma y los rasgos de calidad de la concha. Del mismo modo, las fuentes de VD como VD3, y 25-(OH) D3 a 2.000 UI aumentó la puntuación de la unidad de Haugh, siguiendo a Nascimento et al. (40).
La interacción entre el nivel de Ca y VD3 revela que el aumento del nivel de Ca de 3.5 a 4 o 4.5% aumentó notablemente y de manera similar el porcentaje de yema, la relación yema: albúmina, la puntuación de la unidad Haugh, el porcentaje de concha, ST y SWUSA. Los resultados indicaron que el Ca al 4% fue adecuado para mejorar las características de la calidad del huevo durante la última etapa de la producción de huevos y el aumento del nivel de Ca al 4,5% no tuvo efectos positivos. La mejora en los rasgos de calidad del huevo y la cáscara de las gallinas alimentadas con 4% de Ca sugiere una mayor disponibilidad de Ca para la formación de cáscara de huevo, de acuerdo con otros investigadores en este campo (16, 40). Esto puede deberse al papel del Ca en la regulación de las hormonas reproductivas y el crecimiento de los ovarios y su papel vital en la formación de la cáscara del huevo y como agente crítico para preservar la calidad del huevo (5, 16, 21). Se citó que cada aumento en la cantidad de Ca disponible puede mejorar la calidad de la cáscara y la resistencia a la rotura de huesos, con un efecto lineal (40, 43).
La suplementación con VD3 a 1,000 UI a 4% de Ca la dieta redujo sustancialmente el porcentaje de yema de gallinas en 4% de Ca, en comparación con el mismo grupo de Ca con 800 UI de VD3, y la relación yema:albúmina, en comparación con 800 y 1.200 UI. Sin embargo, 1.000 UI de VD3 aumento del porcentaje de yema, proporción yema: albúmina y color de la yema de aquellos alimentados con 4.5% de Ca en comparación con los otros grupos con el mismo nivel de Ca. El índice vitelino aumentó significativamente debido a la suplementación de 3,5% de Ca con 1.000 y 1.200 UI de VD3 en comparación con 800 UI añadidas al mismo nivel de Ca.
El porcentaje de albúmina mostró un marcado aumento debido a VD3 suplementación a 1,000 UI a 4.0% Ca, en comparación con las 800 UI de VD3 suplementado al mismo nivel de Ca. Por otro lado, 1.000 UI de VD3 disminuyó significativamente el porcentaje de albúmina de gallinas alimentadas con 4,5% de Ca, en comparación con 800 UI de VD3 añadido al mismo nivel de Ca. Estos cambios son contrarios a los cambios mostrados en el porcentaje de yema.
La unidad Haugh, el espejo de la calidad de la albúmina, mostró un aumento considerable gradualmente, con un aumento de VD3 adición a 3.5% de dieta Ca, pero 1,200 UI de VD3 al 4% de dieta Ca disminuyendo significativamente la unidad de Haugh, en comparación con los otros niveles de VD3 suplementado al mismo nivel de Ca. El efecto de VD3 depende de las concentraciones del nivel de Ca. El índice de forma del huevo más alto fue de gallinas alimentadas con un nivel de 3.5% de Ca suplementado con 1,200 UI, mientras que el más pequeño fue de gallinas alimentadas con 4.5% de Ca suplementadas con 1,000 UI de VD3. La fortificación con VD3 a 1.000 y 1.200 UI a 3,5% de Ca la alimentación aumentó sustancialmente el porcentaje de la cáscara y el grosor de la cáscara de manera similar en comparación con los de 800 UI de VD3. El aumento en el SWUSA fue lineal, tal vez debido a la corrección del área de superficie del huevo (31, 32).
El efecto de VD3 en la calidad de la cáscara de huevo de las gallinas alimentadas con niveles adecuados de Ca (4 y 4,5%) fue menos pronunciada y dependió del tipo de mediciones. Estos resultados demostraron que el impacto de VD3 La calidad de la cáscara de huevo depende de que el nivel de Ca en la dieta sea más pronunciado con una ingesta inadecuada de Ca, según otras investigaciones (5, 21, 27). Los metabolitos de VD son esenciales para la proteína de unión a Ca que participa en el transporte activo de Ca a través de la pared intestinal, que es esencial para la formación de la cáscara del huevo (5, 21, 23). La formación de proteínas de unión a Ca en diferentes tejidos (intestino, riñón y útero) requirió metabolitos de VD en ambas etapas transcripcionales y posttranscripcionales. Las proteínas de unión al calcio mejoran la absorción de calcio en el intestino, la recuperación de la orina y la deposición de la cáscara (17, 20).
Las imágenes del microscopio electrónico (Figuras 1-9) mostraron un aumento en la ultraestructura de la cáscara del huevo debido al aumento del nivel de Ca y VD3 suplementación, como se manifiesta por la diferente distribución de botones mamilares, que se hizo más grande a medida que los niveles dietéticos de Ca y VD3 aumentó (Figuras 2–9). La cáscara del huevo, evaluada por microscopía electrónica, consta de varias regiones morfológicamente diferentes y también contiene miles de poros de intercambio de gases. La capa más externa es la cutícula que actúa como una barrera física contra el agua y la contaminación bacteriana. Los sitios de nucleación específicos en la superficie externa de la membrana de la capa externa atraen sales de calcio para la formación de las capas calcificadas (cono o capa mamilar), que pueden estar influenciadas por la actividad enzimática, y los minerales traza actúan como cofactores de dicho proceso. Cada columna de empalizada crece a partir de un botón mamilar, y a medida que avanza el mecanismo de calcificación, proporcionan una mayor resistencia a la cáscara (44).
 FIGURA 1. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con la dieta de control de 3.5% Ca con 800 UI VD3/kg dieta. La figura muestra una baja densidad de botones mamilares, lo que resulta en una reducción de la fuerza, de acuerdo con el grosor de cáscara de huevo significativamente más bajo y SWUSA (P < 0.05) registrado en este grupo experimental. Es importante subrayar la gran área intersticial relativa entre las formaciones mamilares, lo que hace que el huevo sea más susceptible a romperse a lo largo de estas grietas.
FIGURA 1. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con la dieta de control de 3.5% Ca con 800 UI VD3/kg dieta. La figura muestra una baja densidad de botones mamilares, lo que resulta en una reducción de la fuerza, de acuerdo con el grosor de cáscara de huevo significativamente más bajo y SWUSA (P < 0.05) registrado en este grupo experimental. Es importante subrayar la gran área intersticial relativa entre las formaciones mamilares, lo que hace que el huevo sea más susceptible a romperse a lo largo de estas grietas.
 FIGURA 2. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.0% de Ca con 800 UI de VD3/kg de dieta. En comparación con la Figura 1, se puede detectar un ligero aumento en el número de botones mamilares asociados con un aumento del tamaño más grande.
FIGURA 2. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.0% de Ca con 800 UI de VD3/kg de dieta. En comparación con la Figura 1, se puede detectar un ligero aumento en el número de botones mamilares asociados con un aumento del tamaño más grande.
 FIGURA 3. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.5% de Ca con 800 UI de VD3/kg de dieta. Es posible observar un aumento en el número de botones mamilares, en comparación con la Figura 1, al aumentar solo los niveles de Ca.
FIGURA 3. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.5% de Ca con 800 UI de VD3/kg de dieta. Es posible observar un aumento en el número de botones mamilares, en comparación con la Figura 1, al aumentar solo los niveles de Ca.
 FIGURA 4. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 3.5% de Ca con 1,000 UI de VD3/kg dieta. Esta figura muestra una distribución similar de botones mamilares, pero el área intersticial parece menos evidente que en la Figura 1.
FIGURA 4. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 3.5% de Ca con 1,000 UI de VD3/kg dieta. Esta figura muestra una distribución similar de botones mamilares, pero el área intersticial parece menos evidente que en la Figura 1.
 FIGURA 5. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.0% de Ca con 1,000 UI de VD3/kg dieta. En esta figura, es posible observar una mayor área intersticial relativa entre las formaciones mamilares que influye negativamente en la fuerza de la cáscara del huevo. El área intersticial más alta es un indicador del grado de elasticidad de la cáscara del huevo.
FIGURA 5. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.0% de Ca con 1,000 UI de VD3/kg dieta. En esta figura, es posible observar una mayor área intersticial relativa entre las formaciones mamilares que influye negativamente en la fuerza de la cáscara del huevo. El área intersticial más alta es un indicador del grado de elasticidad de la cáscara del huevo.
 FIGURA 6. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.5% de Ca con 1,000 UI de VD3/kg dieta. En comparación con la Figura 5, se observó una reducción del área intersticial entre los botones mamilares, pero no se detectaron diferencias en cuanto a la densidad de los botones mamilares.
FIGURA 6. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.5% de Ca con 1,000 UI de VD3/kg dieta. En comparación con la Figura 5, se observó una reducción del área intersticial entre los botones mamilares, pero no se detectaron diferencias en cuanto a la densidad de los botones mamilares.

 FIGURA 8. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.0% de Ca con 1.200 UI-VD3/kg dieta. En esta figura, es posible observar la mayor densidad y porcentaje de botones mamilares en los huevos de gallinas ponedoras alimentadas con la dieta con 4.0 y 4.5% Ca y 1,200 UI VD3, de acuerdo con el espesor de proyectil más alto (P < 0,05) y SWUSA.
FIGURA 8. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.0% de Ca con 1.200 UI-VD3/kg dieta. En esta figura, es posible observar la mayor densidad y porcentaje de botones mamilares en los huevos de gallinas ponedoras alimentadas con la dieta con 4.0 y 4.5% Ca y 1,200 UI VD3, de acuerdo con el espesor de proyectil más alto (P < 0,05) y SWUSA.
 FIGURA 9. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.5% de Ca con 1,200 UI de VD3/kg dieta. En esta figura, es posible observar la mayor densidad y porcentaje de botones mamilares en los huevos de gallinas ponedoras alimentadas con la dieta con 4.0 y 4.5% Ca y 1,200 UI VD3, de acuerdo con el espesor de proyectil más alto (P < 0,05) y SWUSA.
FIGURA 9. Foto SEM de la cáscara de huevo de gallinas ponedoras alimentadas con una dieta que contiene 4.5% de Ca con 1,200 UI de VD3/kg dieta. En esta figura, es posible observar la mayor densidad y porcentaje de botones mamilares en los huevos de gallinas ponedoras alimentadas con la dieta con 4.0 y 4.5% Ca y 1,200 UI VD3, de acuerdo con el espesor de proyectil más alto (P < 0,05) y SWUSA.
El aumento de la resistencia de las cáscaras, así como la reducción de la pérdida de huevos, es un objetivo importante que tiene importancia económica en términos comerciales. Se ha considerado que la calidad de la cáscara del huevo se ve afectada por los componentes orgánicos, particularmente las proteínas (9, 10) y los componentes inorgánicos (45). Por lo tanto, la capa de empalizada comprende aproximadamente dos tercios del espesor total de la cáscara (46). En este sentido, la capa de empalizada mostró una reducción lineal en el número de botones mamilares en la cáscara de huevo de las gallinas alimentadas, aumentando los niveles de Mn, Zn y Cu (33). Como informaron Stefanello et al. (33), la suplementación con oligoelementos orgánicos puede ejercer una influencia en la formación de la capa de empalizada, lo que resulta en botones mamilares más grandes. Nuestros resultados mostraron que el grupo alimentado con una dieta que contenía 3,5% de Ca suplementada con 800 UI de VD3 tenía un desorden en la distribución de los botones mamilares en la superficie interna de la carcasa (Figura 1). A partir de estos resultados, podemos especular que la capa de empalizada y el número de botones mamilares presentes en la cáscara también pueden influir en la calidad de la cáscara.
Contenido de minerales plasmáticos y tibiales
La Tabla 4 muestra la influencia del Ca y VD dietéticos3 sobre minerales plasmáticos y características de la tibia. Ca y/o VD dietético3 no afectaron los niveles plasmáticos de Ca, Pi y Ca:Pi.
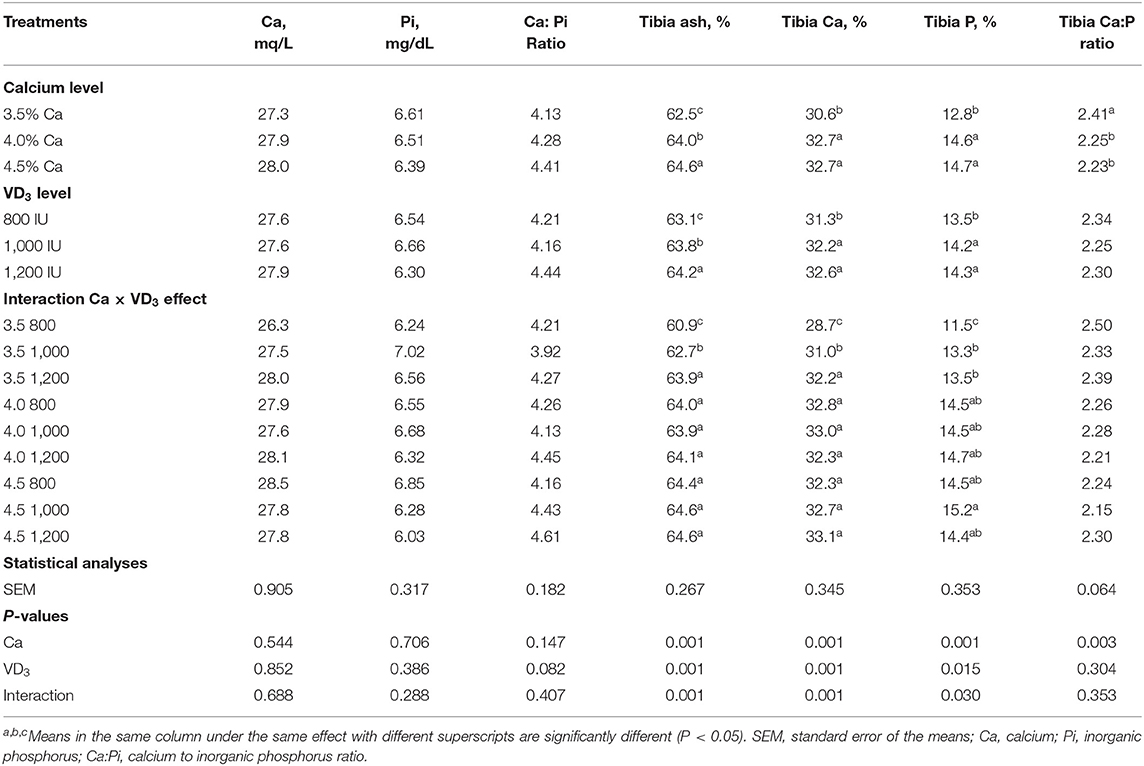 TABLA 4. Minerales del plasma sanguíneo y características de la tibia de ponedoras de huevo marrón de 72 semanas alimentadas con tres niveles de Ca suplementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
TABLA 4. Minerales del plasma sanguíneo y características de la tibia de ponedoras de huevo marrón de 72 semanas alimentadas con tres niveles de Ca suplementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
Esto indica que Ca y VD3 en las dietas control (3,5% Ca y 800 UI de VD suplementada3 o como VD total3 con una dieta de 3.800 UI/kg) fueron adecuados para mantener la concentración plasmática de Ca y Pi en niveles normales. Además, Nascimento et al. (20, 40) demostraron que no hubo efectos de las fuentes de VD a 2.000 UI de VD3, 25- (OH) D3, y 1, 25 (OH)2 D3 en plasma Ca. Además, Susanna et al. (47) observaron que la VD dietética3 a 3.000 UI de una sola fuente o a 1.500 UI de cada VD3 y 25 (OH) D3 no afectó el nivel plasmático de Ca, pero disminuyó el Pi plasmático de gallinas ponedoras de 34 semanas de edad. Esto puede deberse al control hormonal de Ca y Pi a través de la paratiroides y la calcitonina (21, 41, 48).
Se encontró que la ceniza, Ca y Pi se vieron afectadas significativamente por el Ca y el VD dietéticos.3 Pero el impacto interfirió con la interacción entre las dos variables. Además, la tibia Ca: Pi se vio afectada solo por los niveles nutricionales de Ca, lo que demuestra que la relación Ca: Pi disminuyó significativamente de manera similar debido a que el Ca aumentó por encima del 3.5%. El aumento del nivel de Ca de 3.5 a 4 y 4.5% dentro de las dietas no complementadas aumentó significativamente y de manera similar la ceniza de tibia, Ca y Pi, mostrando saturación de respuesta al 4% Ca. Esto sugiere que el 4% de Ca fue adecuado para la calcificación ósea (5, 37-41). La suplementación del nivel de 3,5% de Ca con VD3 a 1.000 y 1.200 UI aumentó significativamente la ceniza de tibia y Ca de manera gradual y aumentó de manera similar Pi. Esto indica que la suplementación con VD3 es beneficioso para gallinas ponedoras a partir de las 60 semanas de edad para la calcificación ósea (16, 20). Estos pueden deberse al papel de VD3 y sus metabolitos en la matriz ósea (20, 41). Se observó que el aumento de la EV3 La suplementación a 1.000 y 1.200 UI dentro de los niveles de 4 y 4,5% de Ca no afectó la ceniza de tibia y los contenidos de Ca y Pi. Estos resultados indican que la respuesta a VD3 depende de los niveles de Ca en la dieta (5, 14).
Metabolitos de lípidos séricos
El efecto de diferentes Ca y/o VD dietéticos3 Los niveles en la mayoría de los metabolitos lipídicos séricos sanguíneos no fueron significativos, excepto para los triglicéridos, vLDL y la relación triglicéridos:HDL y se acercaron significativos para HDL (Tabla 5).
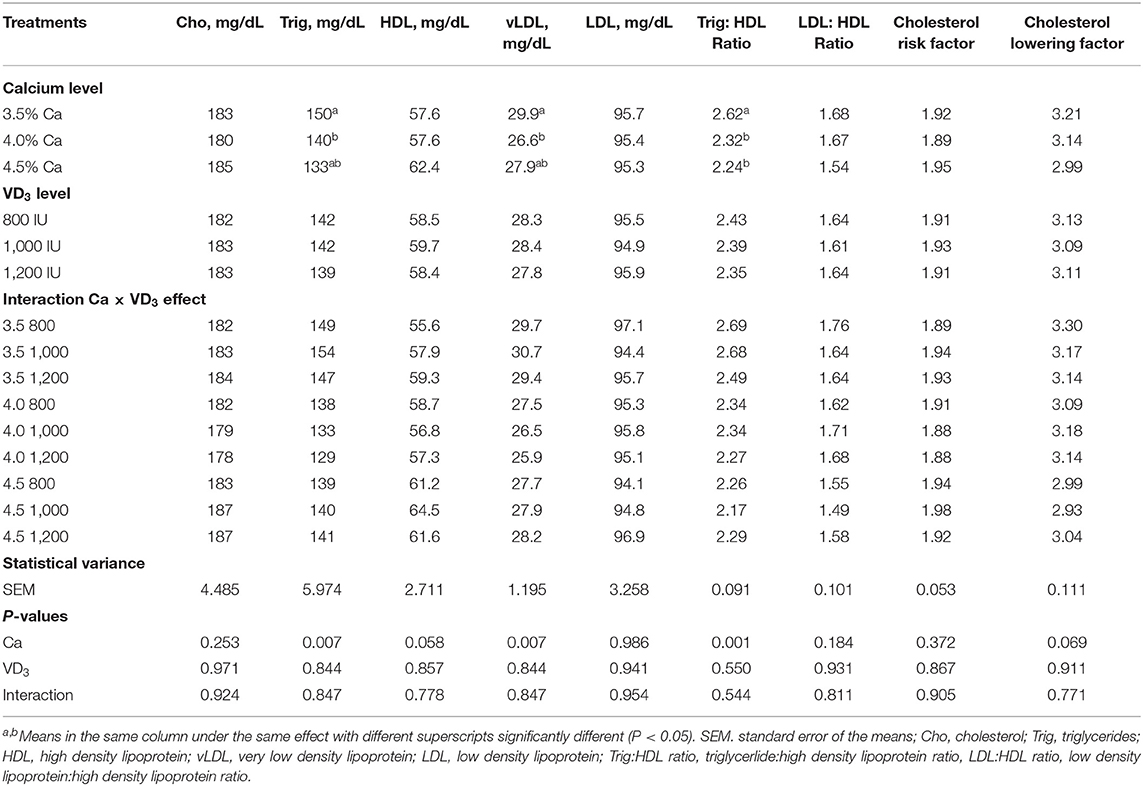 TABLA 5. Metabolitos lipídicos séricos sanguíneos de ponedoras de huevo marrón alimentadas con tres niveles de Ca suplementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
TABLA 5. Metabolitos lipídicos séricos sanguíneos de ponedoras de huevo marrón alimentadas con tres niveles de Ca suplementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
Los resultados demostraron que los niveles de Ca al 4% disminuyeron considerablemente los triglicéridos y vLDL en comparación con el 3,5% de Ca. Además, las concentraciones de 4 y 4,5% de Ca disminuyeron sustancialmente los triglicéridos séricos: HDL, en comparación con el 3,5% de Ca. Además, esto se asocia con un aumento en HDL con el aumento del nivel de Ca. Estas disminuciones podrían atribuirse al aumento en la producción diaria de masa de huevos de gallinas alimentadas con 4 y 4,5% de Ca, debido al uso de lipoproteínas para la formación de yema. Las lipoproteínas, particularmente la yema de lipoproteína de muy baja densidad (VLDLy) se formatearon bajo la influencia del estrógeno. El precursor de la yema se sintetiza en el hígado y se transporta al ovario para la formación de la yema, que aumentaría con el aumento de LR (1, 26, 49).
Fracciones de proteínas séricas y enzimas del índice hepático
La Tabla 6 muestra el impacto de diferentes Ca y VD3 niveles en fracciones de proteínas séricas e índices hepáticos para enzimas de fuga. No hubo efectos de diferentes niveles de Ca sobre la albúmina sérica, AST, ALT y fosfatasa alcalina. El aumento en el nivel de Ca indujo un aumento similar en la proteína sérica total, globulina (proteína inmune específica) y α-2-globulina en comparación con 3.5% Ca, pero disminuyó de manera similar la albúmina sérica (agente inmune no específico) / relación globulina y la relación AST: ALT. Estos resultados indican que el Ca es un elemento esencial para la inmunidad de las gallinas ponedoras, como se manifiesta por el aumento de la proteína sérica total y la globulina (50-52). La deficiencia de Ca en gallinas ponedoras puede causar enfermedades óseas como fatiga de las ponedoras de jaulas y osteoporosis (1, 5, 18, 26).
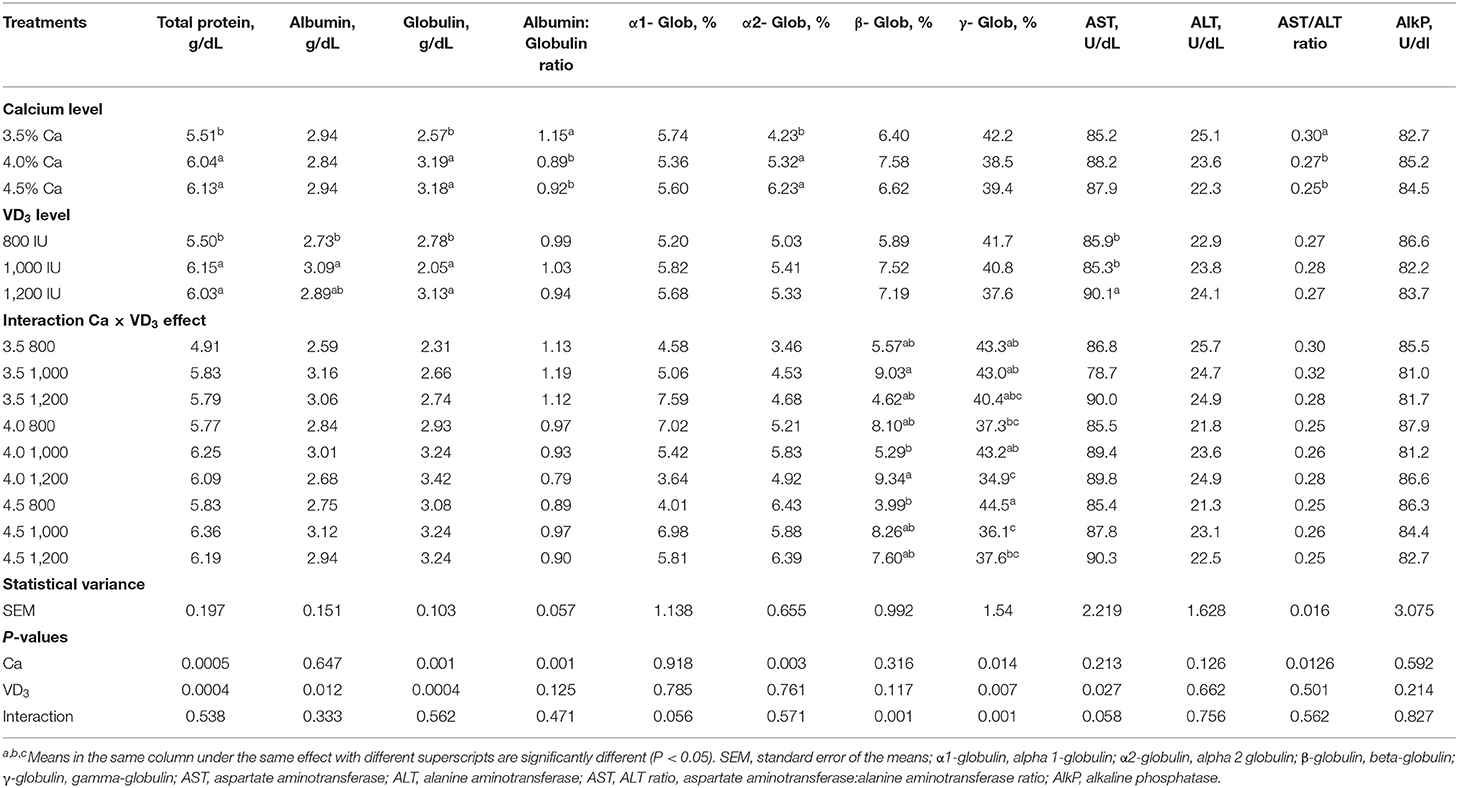 TABLA 6. Fracciones de proteína sérica sanguínea e índices de enzimas de fuga hepática de ponedoras de huevo marrón alimentadas con tres niveles de Ca, complementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
TABLA 6. Fracciones de proteína sérica sanguínea e índices de enzimas de fuga hepática de ponedoras de huevo marrón alimentadas con tres niveles de Ca, complementadas con tres niveles de VD3 de 60 a 72 semanas de edad.
VD dietético3 no influyeron en la relación albúmina/globulina, α-1- y α-2-globulina, β-globulina, γ-globulina, los niveles de ALT, la relación AST:ALT y el nivel de fosfatasa alcalina. Además, las fracciones de proteínas séricas y las enzimas de fuga del índice hepático no se ven afectadas por la interacción entre las dos variables.
Se encontró que el aumento de VD3 a una dieta de 1.000 y 1.200 UI/kg aumentó sustancialmente el total de proteínas y globulina, en comparación con una dieta de 800 y 1.000 UI/kg, aumentó la albúmina sérica en comparación con una dieta de 800 UI/kg. La actividad inmunomoduladora, antiinflamatoria y anticoccidios de VD3 o sus metabolitos han sido reportados en pollos (20, 23–26, 51, 52) y células de pollo (25, 27, 28). El anticuerpo es de naturaleza proteómica, y el aumento en los niveles totales de proteína y globulina apoyó la hipótesis de que el aumento de Ca y VD3 Mejora la inmunidad de las gallinas ponedoras, tal vez debido a la protección contra las enfermedades óseas [raquitismo, osteoporosis y fatiga de las ponedoras de jaulas (20, 41, 53)]. VD tiene una tarea esencial en el mantenimiento de la inmunidad y la comunicación entre los sistemas de inmunidad adaptativa e innata al afectar los receptores vitamínicos de VD y activar las enzimas (20, 54, 55).
El efecto de interacción indica que la β-globulina más alta se observó en grupos alimentados con 3,5 y 4% de Ca, suplementados con 1.000 y 1.200 UI de VD3, respectivamente, mientras que los valores más bajos fueron de gallinas alimentadas con 4 y 4,5% de Ca, suplementadas con 1.000 y 800 UI de VD3respectivamente.
La γ-globulina, el anticuerpo material, fue significativamente mayor en las gallinas alimentadas con 4,5% de Ca que en las gallinas alimentadas con 4% de Ca cuando se compararon los grupos no suplementados. Además, suplementación de 4% Ca con 1,200 UI VD3 disminuyó significativamente la γ-globulina, en comparación con 1.000 UI de VD3 complementando la misma concentración de Ca. Además, la suplementación de 4,5% de Ca con 1.000 y 1.200 UI de VD3 disminuyó significativamente la γ-globulina sérica en comparación con la suplementación con 800 UI del mismo nivel de Ca.
Título de inmunoglobulina y anticuerpos
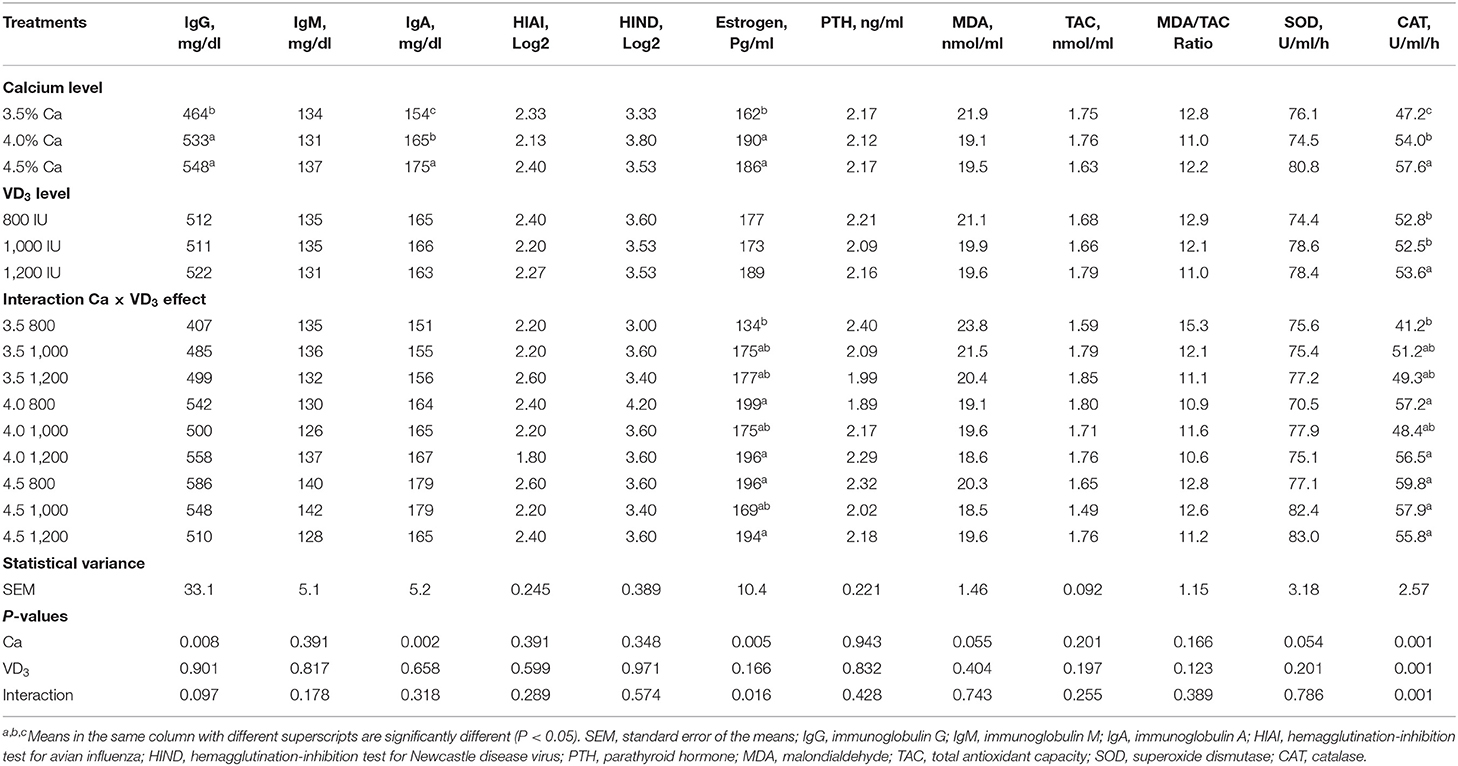
TABLA 7. Parámetros de inmunidad del suero sanguíneo y algunas hormonas de las ponedoras de huevo marrón alimentadas con tres niveles de Ca suplementado con tres niveles de VD3 de 60 a 72 semanas de edad.
La falta de efectos significativos de VD3 en la mayoría de las respuestas inmunitarias (el tipo de globulina, inmunoglobulinas y título de anticuerpos), en lugar de proteína sérica total, albúmina y globulina, puede indicar que la dieta de control suplementada con 800 UI, o un VD total3 de 3.800 UI, contiene VD adecuado3 para mantener la función inmune básica. Esto puede compensar el efecto del colecalciferol suplementado sobre la inmunidad celular y humoral. Esto se estableció aún más por la falta de enfermedades óseas como la osteoporosis y la fatiga de las ponedoras en jaulas en gallinas ponedoras mantenidas en jaulas en condiciones climáticas cálidas. Tales trastornos indican VD dietética inadecuada3 (1, 20, 26).
En la literatura, VD3 se informa que tiene muchas propiedades, como actividades antioxidantes, inmunomoduladoras, antiinflamatorias, antivirales, antibacterianas, antialérgicas y de prevención del cáncer (10–12). En aves de corral, el papel de VD3 en Ca y Pi el metabolismo es fundamental para el desarrollo de la formación de hueso y cáscara de huevo. En este sentido, Rodríguez-Lecompte et al. (26) demostraron que tanto VD3 y 25-(OH) D tienen fuertes efectos inmunomoduladores, como el aumento de la respuesta positiva de las células T auxiliares (Th2) y la autofagia. Las gallinas ponedoras mantenidas a altas temperaturas ambientales, como las observadas en este documento, son altamente susceptibles a enfermedades infecciosas, debido a la baja inmunidad (49). Aslam et al. (56) encontraron que la deficiencia de VD disminuye la respuesta inmune celular en pollos de engorde. La literatura informa que VD3 o 25-hidroxicolecalciferol tiene una actividad antiinflamatoria en las células inmunes de las aves después de la administración de lipooligosacáridos [LPS (20, 57-60)]. A pesar de esto, se ha prestado poca atención a la influencia de la VD dietética.3 suplementación o su deficiencia en la respuesta inmune y la bioquímica sanguínea de gallinas ponedoras desafiadas con LPS, o los mecanismos de acción, aunque los efectos anticancerígenos y antiinflamatorios de VD3 están bien documentados (17, 61–64).
Hormonas plasmáticas
El efecto del nivel de Ca y la interacción entre el nivel de Ca y VD3 se observó en los niveles plasmáticos de estrógeno (Tabla 7). Está claro que el aumento del nivel de Ca de 3,5 a 4 o 4,5% aumentó el estrógeno plasmático. La PTH plasmática no fue influenciada por Ca y/o VD dietética3 Concentraciones. Esto indica que la dieta control (3,5% Ca y 3.800 UI VD3) contiene Ca y VD adecuados3 para mantener el nivel normal de PTH, que, junto con 1,25-(OH)2D3, y controla la absorción de calcio en el canal digestivo y la reabsorción de calcio de los huesos y la excreción para mantener el equilibrio de Ca (1, 21, 26).
Se encontró que el aumento de los niveles de Ca a 4 y 4,5% dentro de los grupos no suplementados significativamente, y de manera similar, aumentó el E plasmático2 en comparación con el nivel del 3,5%. No hubo cambios significativos en el estrógeno plasmático dentro de cada nivel de Ca o en diferentes niveles de Ca como resultado de la suplementación con VD3 cuando se compararon los niveles correspondientes. La E más baja2 fue en gallinas alimentadas con 3,5% de Ca suplementadas con 800 UI de VD3, mientras que el más alto fue de gallinas alimentadas con 4 y 4,5% de Ca, suplementadas con 800 y 1.200 UI de VD3. Estos hallazgos demuestran que un nivel de Ca del 4% fue adecuado cuando se complementó con 800 UI de VD3 para mejorar E2. El metabolismo del Ca en gallinas ponedoras está controlado por E2, PTH y calcitonina (5, 21), y el aumento de E2 se asoció con el aumento del rendimiento y la calidad de la cáscara de huevo (Tabla 7).
Estado antioxidante
No se encontró ningún efecto en la mayoría de los índices antioxidantes (TAC, MDA y SOD) con niveles de Ca, excepto para CAT. Ca y VD dietéticos3 concentración afectó a la CAT plasmática, pero el efecto fue confundido por la interacción significativa entre Ca dietético y VD3 (Tabla 7). Se encontró que el aumento de la VD3 nivel a 1.000 y 1.200 UI en el grupo de 3,5% Ca aumentó CAT en cierta medida. No hay diferencias significativas en los niveles de TAC entre los diferentes VD3 niveles o dentro o entre diferentes niveles de Ca. Estos resultados muestran que el impacto de VD3 en CAT depende de la concentración de Ca en la dieta. Por lo tanto, las enzimas antioxidantes como CAT necesitan una cantidad adecuada de Ca y VD3 para ser mantenidas para gallinas ponedoras de 60–72 semanas de edad.
Ovario y órganos reproductivos
Los resultados del peso relativo de los órganos ováricos y reproductivos afectados por diferentes Ca y VD3 las concentraciones se presentan en la Tabla 8.
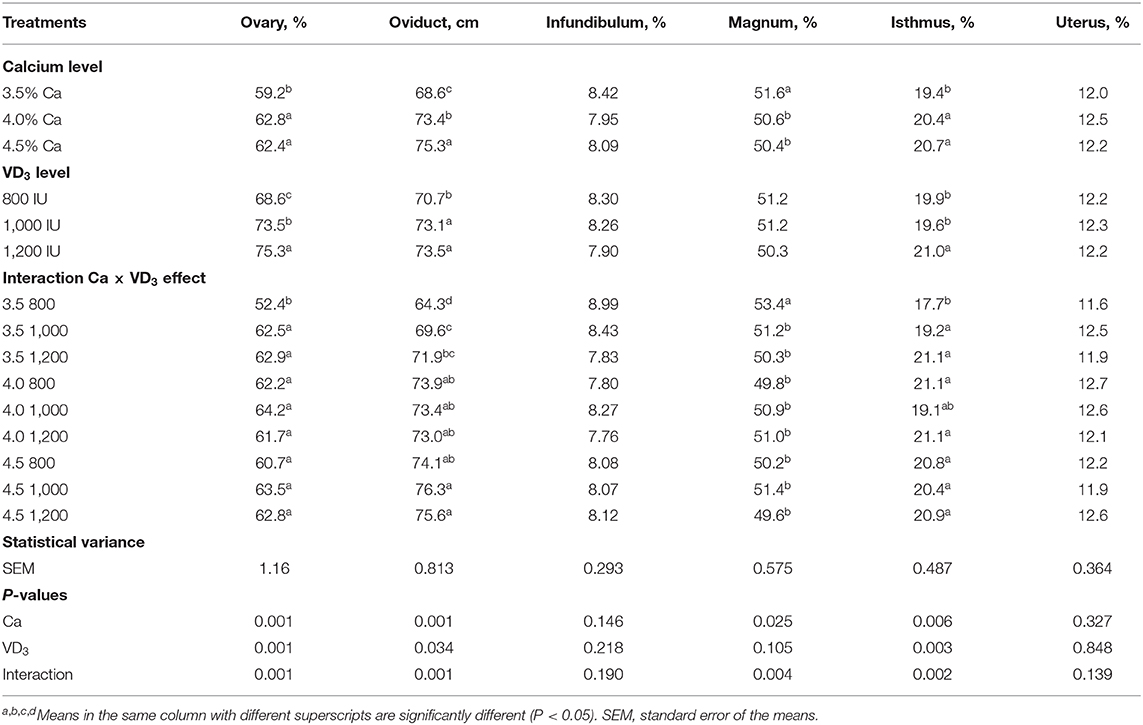 TABLA 8. La longitud de las partes del sistema reproductor femenino de las ponedoras de huevos marrones alimentadas con tres niveles de Ca suplementados con tres niveles de VD3 de 60 a 72 semanas de edad.
TABLA 8. La longitud de las partes del sistema reproductor femenino de las ponedoras de huevos marrones alimentadas con tres niveles de Ca suplementados con tres niveles de VD3 de 60 a 72 semanas de edad.
El nivel de Ca en la dieta tuvo un impacto (P < 0,05) en el porcentaje del ovario, magnum, el istmo y la longitud absoluta del oviducto (cm), y VD3 Las concentraciones mostraron una marcada influencia en el peso relativo del ovario y el istmo y la longitud del oviducto. Estos impactos fueron confundidos por la interacción entre Ca y VD3.
Los resultados de la interacción indican que el aumento de los niveles de Ca con los grupos suplementados con 800 UI de VD3 aumentó significativamente el peso relativo de ovario e istmo y la longitud absoluta del oviducto, en comparación con la dieta control que contenía 3,5% de Ca y 3.800 UI de EV total3, pero disminuyó el porcentaje magnum. Se encontró que elevar el VD3 concentraciones a 1.000 (4.000 UI como VD total3) y 1200 UI (4.200 UI como VD total3) dentro del nivel de Ca del 3,5% elevó de manera similar los pesos relativos del ovario y el istmo y la longitud absoluta del oviducto, pero disminuyó el peso relativo del magnum. Estos cambios en el ovario y el oviducto, excepto el magnum, reflejaron los cambios positivos en E.2 y mejoró el rendimiento de puesta de las gallinas alimentadas con 3,5% de Ca cuando se suplementaron con 1.000 o 1.200 UI de VD3.
En conclusión, aumentar los niveles dietéticos de Ca en gallinas ponedoras hasta un 4% durante la fase de producción tardía podría ser una herramienta útil para mejorar el rendimiento de la puesta, la calidad de la cáscara del huevo, la unidad Haugh y el estado fisiológico e inmunológico. Además, suplementando dietas 3.5% Ca con 1,000 UI de VD3 o un total de 4.000 UI/kg de dieta VD3 mejora del rendimiento de las gallinas alimentadas con un nivel de Ca del 3,5% durante la última etapa de producción (60-72 semanas de edad), lo que demuestra que el impacto de VD3 depende de las concentraciones dietéticas de Ca.
Declaración de disponibilidad de datos
Todos los conjuntos de datos generados para este estudio se incluyen en el artículo / material complementario.
Declaración ética
Los procedimientos experimentales fueron aprobados por la Universidad Rey Abdulaziz, Jeddah, Arabia Saudita bajo el número de protocolo (DF-715-155-1441H) que recomienda los derechos de los animales, el bienestar y el estrés mínimo y no causó ningún daño o sufrimiento a los animales de acuerdo con el Real Decreto número M59 en 14/9/1431H.
Contribuciones del autor
Todos los autores enumerados han hecho una contribución sustancial, directa e intelectual al trabajo, y lo aprobaron para su publicación.
Financiación
Este proyecto fue financiado por el Decanato de Investigación Científica (DSR), Universidad Rey Abdulaziz, Jeddah, bajo la Subvención No. (DF-715-155-1441 H).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Reconocimientos
Los autores agradecen al DSR, la Universidad Rey Abdulaziz, por su apoyo técnico y financiero.
Referencias
1. Oluyemi JA, Roberts FA. Producción avícola en climas cálidos y húmedos. 2ª ed. Ibadan: Spectrum Books Limited (2000).
2. Pizzolante CC, Saldanha ESPB, Lagana C, Kakimoto SK, Toghashi CK. Efecto de los niveles de calcio y el tamaño de la retícula de piedra caliza en la calidad del huevo de las ponedoras semipesadas en su segundo ciclo de producción. Braz J Poult Sci. (2009) 11:79–86. doi: 10.1590/S1516-635X2009000200002
3. Saunders-Blades JL, Korver DR. Efecto de la edad de gallina y la fuente materna de vitamina D sobre el rendimiento, la incubabilidad, la densidad mineral ósea y la progenie in vitro de la función inmune innata temprana. Poult Sci. (2015) 94:1233–46. doi: 10.3382/ps/pev002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Keshavarz KA. Comparación entre colecalciferol y 25-OH-colecalciferol en el rendimiento y la calidad de la cáscara de huevo de gallinas alimentadas con diferentes niveles de calcio y fósforo. Poult Sci. (2003) 82:1415–22. doi: 10.1093/ps/82.9.1415
5. Roland, DA. Calidad de la cáscara de huevo III: requisitos de calcio y fósforo de los Leghorns comerciales. Mundos Poult Sci J. (1986) 42:154–65. doi: 10.1079/WPS19860012
6. Al-Batshan HA, Scheideler SE, Black BL, Garlich JD, Anderson KE. La captación duodenal de calcio, la ceniza del fémur y la calidad de la cáscara de huevo disminuyen con la edad y aumentan después de la muda. Poult Sci. (1994) 73:1590–6. DOI: 10.3382/PS.0731590
Resumen de PubMed | Texto completo de CrossRef | Google Académico
7. Saunders-Blades JL, MacIsaac JL, Korver DR, Anderson DM. El efecto de la fuente de calcio y el tamaño de partícula en el rendimiento de producción y la calidad ósea de las gallinas ponedoras. Poult Sci. (2009) 88:338–53. DOI: 10.3382/PS.2008-00278
Resumen de PubMed | Texto completo de CrossRef | Google Académico
8. Gregory NG, Wilkins LJ. Huesos rotos en aves domésticas: manejo y procesamiento de daños en gallinas en batería al final de la puesta. Br Poult Sci. (1989) 30:555–62. DOI: 10.1080/00071668908417179
Resumen de PubMed | Texto completo de CrossRef | Google Académico
9. Marie P, Labas V, Brionne A, Harichaux G, Hennequet-Antier C, Nys Y, et al. La proteómica cuantitativa y el análisis bioinformático proporcionan una nueva visión de la función de las proteínas durante la biomineralización de la cáscara de huevo aviar. J Proteómica. (2015) 113:178–93. doi: 10.1016/j.jprot.2014.09.024
Resumen de PubMed | Texto completo de CrossRef | Google Académico
10. Marie P, Labas V, Brionne A, Harichaux G, Hennequet-Antier C, Rodriguez-Navarro AB, et al. La proteómica cuantitativa proporciona nuevos conocimientos sobre las funciones de la proteína de la matriz de la cáscara de huevo de gallina durante los eventos primarios de mineralización y la fase de calcificación activa. J Proteómica. (2015) 126:140–54. doi: 10.1016/j.jprot.2015.05.034
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Attia YA, Al-Harthi MA, Shiboob MM. Evaluación de la calidad y el contenido de nutrientes de los huevos de mesa de diferentes fuentes en el mercado minorista. Ital J Anim Sci. (2014) 13:3269. doi: 10.4081/ijas.2014.3294
12. Keshavarz K, Scott ML, Blanchard J. El efecto de la solubilidad y el tamaño de partícula de las fuentes de calcio en la calidad de la cáscara y la mineralización ósea. J Appl Poult Res. (1993) 2:259–67. DOI: 10.1093/JAPR/2.3.259
13. Clunies M, Park D, Leeson S. Metabolismo del calcio y fósforo y formación de cáscara de huevo de gallinas alimentadas con diferentes cantidades de calcio. Poult Sci. (1992) 71:482–9. DOI: 10.3382/PS.0710482
Resumen de PubMed | Texto completo de CrossRef | Google Académico
14. Wallner-Pendleton E, Scheideler SE. The Influence of Varying Levels of Calcium and Vitamin D3 in the Mature Laying Hen’s Diet: Effect on Egg Production, Shell Quality, Bone Ash and Urolithiasis. The Nebraska Poultry Report. University of Nebr Coop Ext EC, Lincoln (1996).
15. Roush WB, Mylet M, Rosenberger JL, Derr J. Investigation of calcium and available phosphorus requirement of laying hens by response surface methodology. Poult Sci. (1986) 65:964–70. doi: 10.3382/ps.0650964
16. Tierzucht L, Layers LBC. Lohaman-Brown-Classic Layers Management–Guide Cage Housing. (2016). Available online at: https://www.ltz.de/ (accessed March 20, 2018).
17. Dale N. National Research Council Nutrient Requirements of Poultry 9th Revised Edition. Washington, DC: National Academy Press (1994).
18. Directrices 2016 para materiales nutricionales de salud de animales domésticos. DSM Vitamin Supplementation Guidelines 2016 for Animal Nutrition. (2016). págs. 1–26. Disponible en línea en: https://www.dsm.com/content/dam/dsm/anh/en_US/documents/Vitamin_Supp_Guidelinespdf.?download=a1debc5c-f2e3-43c5-b782-64b7257f55b21584479483674 (consultado el 25 de marzo de 2019).
19. Stevens VI, Blair R, Salmon RE. Efectos de VD3, calcio y fósforo en el crecimiento y desarrollo óseo de pavos de mercado. Poult Sci. (1984) 63:1571–85. DOI: 10.3382/PS.0631571
Resumen de PubMed | Texto completo de CrossRef | Google Académico
20. Geng Y, Ma Q, Wang ZD, Guo Y. La suplementación dietética con vitamina D3 protege a las gallinas ponedoras contra el estrés inmunológico inducido por lipopolisacáridos. Nutr Metab. (2018) 15:58. DOI: 10.1186/S12986-018-0293-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
21. Scott ML, Nesheim MC, Young RJ. Nutrición del pollo. Ithaca, NY: Scott ML and Associates (1982).
22. Soares JH Jr, Kerr JM, Gray RW. 25-Hydroxycholicalciferol en nutrición avícola. Poult Sci. (1995) 74:1919–34. DOI: 10.3382/PS.0741919
Resumen de PubMed | Texto completo de CrossRef | Google Académico
23. Overbergh L, Decallonne B, Valckx D, Verstuyf A, Depovere J, Laureys J, et al. Identificación y regulación inmune de la 25-hidroxivitamina D-1-alfa-hidroxilasa en macrófagos murinos. Clin Exp Immunol. (2000) 120:139–46. doi: 10.1046/j.1365-2249.2000.01204.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
24. Gorman S, Buckley AG, Ling KM, Berry LJ, Fear VS, Stick SM, et al. La suplementación con vitamina D de ratones inicialmente deficientes en vitamina D disminuye la inflamación pulmonar con efectos limitados sobre la integridad epitelial pulmonar. Physiol Rep. (2017) 15:e13371. doi: 10.14814/phy2.13371
Resumen de PubMed | Texto completo de CrossRef | Google Académico
26. Rodriguez-Lecompte JC, Yitbarek A, Cuperus T, Echeverry H, van Dijk A. El efecto inmunomodulador de la vitamina D en pollos depende de la dosis y está influenciado por los niveles de calcio y fósforo. Poult Sci. (2016) 95:2547–56. doi: 10.3382/ps/pew186
Resumen de PubMed | Texto completo de CrossRef | Google Académico
27. Manolagas SC, Provvedini DM, Tsoukas CD. Interacciones de 1, 25-dihidroxi VD3 y el sistema inmune. Mol Celular Endocrinol. (1985) 43:113–22. doi: 10.1016/0303-7207(85)90074-7
28. H&N International Brown Nick, Brown Egg Layers. Nueva guía de gestión – H&N International GmbH. (2016). p. 76. Disponible en línea en: https://www.hn-int.com/eng-wAssets/docs/managementguides/Layers-englisch/HN_MG_Brown-Nick_EN.pdf (consultado el 25 de mayo de 2019).
29. AOAC. Métodos oficiales de análisis. Washington, DC: Asociación de Químicos Analíticos Oficiales (2004).
30. Burke, WH, Attia YA. Muda de cuernos de pierna blancos de peine único con el uso de la formulación de depósito de lupron de acetato de leuprolida. Poult Sci. (1995) 73:1226–32. DOI: 10.3382/PS.0731226
Resumen de PubMed | Texto completo de CrossRef | Google Académico
31. Attia YA, Burke WH, Yamani KA. Respuesta de las gallinas reproductoras de pollos de engorde a la muda forzada por manipulaciones hormonales y dietéticas. Poult Sci. (1994) 73:245–58. DOI: 10.3382/PS.0730245
Resumen de PubMed | Texto completo de CrossRef | Google Académico
32. Attia YA, Burke WH, Yamani KA, Jensen LS. Asignaciones de energía y rendimiento de los reproductores de pollos de engorde 2- hembras. Poult Sci. (1995) 74:261–70. DOI: 10.3382/PS.0740261
33. Stefanello C, Santos TC, Murakami AE, Martins EN, Carneiro TC. Rendimiento productivo, calidad de la cáscara de huevo y ultraestructura de la cáscara de huevo de gallinas ponedoras alimentadas con dietas complementadas con minerales traza orgánicos. Poult Sci. (2014) 93:104–13. DOI: 10.3382/PS.2013-03190
Resumen de PubMed | Texto completo de CrossRef | Google Académico
34. Spectrum, Corp for Biotech SAE, Egypt Spectrum Diagnostic Kits. (2016) Disponible en línea en: http://www.spectrum-diagnostics.com/new/
35. Attia YA, Al-Khalaifah HS, Abd El-Hamid HE, Al-Harthi MA, El-shafey AA. Efecto de diferentes niveles de multienzimas sobre la respuesta inmune, hematología sanguínea y bioquímica, estado antioxidante e histología de órganos de pollos de engorde alimentados con dietas estándar y de baja densidad. Front Vet Sci. (2020) 6:510. DOI: 10.3389/fvets.2019.00510
36. Attia YA, Al-Khalifa H, Ibrahim MS, Abd Al-Hamid AE, Al-Harthi MA, El-Naggar Asmaa. Componentes hematológicos y bioquímicos de la sangre, enzimas antioxidantes, inmunidad y órganos linfoides de pollos de engorde supleados con propóleos, polen de abeja y oligosacáridos de manano de forma continua o intermitente. Poult Sci. (2017) 96:4182–92. DOI: 10.3382/ps/pex173
37. SAS. Versión de estadísticas de la Guía del usuario de SAS.® 10ª ed. Cary, NC: SAS Institute Inc (2004).
38. Rao KS, Roland DA. Influencia del nivel de calcio dietético y el tamaño de partícula de la fuente de calcio en la solubilización de calcio in vivo por leghorns comerciales. Poult Sci. (1989) 68:1499–505. doi: 10.3382/ps.0681499
Resumen de PubMed | Texto completo de CrossRef | Google Académico
39. Zhang B, Coon CN. The relationship of calcium intake, source, size, solubility in vitro and in vivo, and gizzard limestone retention in laying hens. Poult Sci. (1997) 76:1702–6. doi: 10.1093/ps/76.12.1702
40. Nascimento GR, Murakami AE, Guerra AFQM, Ospinas-Rojas IC, Ferreira MFZ, Fanhani JC. Efecto de diferentes fuentes de vitamina D y niveles de calcio en la dieta de las ponedoras en el segundo ciclo de puesta. Braz J Poult Sci. (2014) 16:37–42. doi: 10.1590/1516-635×160237-42
41. Elaroussi MA, Forte LR, Eber SL, Biellier HV. Homeostasis del calcio en la gallina ponedora 1 edad y efectos del calcio en la dieta. Poult Sci. (1994) 73:1581–9. DOI: 10.3382/PS.0731581
Resumen de PubMed | Texto completo de CrossRef | Google Académico
42. Duran MR, Chen C, Kim WK. Efectos de la vitamina d y el calcio para la prevención de la osteoporosis en diversas etapas de la vida de las gallinas ponedoras-revisión. Inter J Poult Sci. (2018) 17:405–9. doi: 10.3923/ijps.2018.405.409
43. Sauveur B. Lésions osseuses et articulaires des pattes des volailles: rôles de l’alimentation. Prod Anim. (1988) 11:35–45.
44. Nys Y, Gautron J. Estructura y formación de la cáscara del huevo. En: Huopalahti R, López-Fandiño R, Anton M, Schade R, editores. Compuestos bioactivos del huevo. Berlín: Springer (2007). págs. 99–102.
45. El-Safty SA. Análisis de regresión gradual para mediciones ultraestructurales de la calidad de la cáscara de huevo en dos razas locales de pollo. Egipto Poult Sci. (2004) 24:189–203.
46. Fathi MM, Zein El-Dein A, El-Safty SA, Radwan LM. Uso de microscopía electrónica de barrido para detectar las variaciones ultraestructurales en la calidad de la cáscara de huevo de las razas de pollos Fayoumi y Dandarawi. Int J Poult Sci. (2007) 6:236–41. DOI: 10.3923/IJPS.2007.236.241
47. Susanna K, Fröhlich E, Gebhardt-Henrich GS, Schäublin H, Pfulg A, Zweifel R et al. Efectos de la suplementación dietética con vitamina D3 sintética y 25-hidroxicolecalciferol sobre los niveles de calcio y fosfato en sangre y el rendimiento en gallinas ponedoras. ArchGeflügelk. (2011) 75:179–84.
48. Baimbridg KG, Taylor TG. El papel de la calcitonina en el control de la hipercalcemia en las aves domésticas (Gallus domesticus). Comp. Bioch. Phys. Parte A Phys. (1981) 68:647–51. DOI: 10.1016/0300-9629(81)90372-8
49. Rosemary L, Walzem RJ, Hansen DL, Williams RL. Inducción de estrógenos de Hamilton del ensamblaje VLDLy en gallinas ponedoras. J Tuerca. (1999) 129:467S−72S. doi: 10.1093/jn/129.2.467S
Resumen de PubMed | Texto completo de CrossRef | Google Académico
50. Garlich J, Brake J, Parkhurst CR, Thaxton JP, Morgan GW. Perfil fisiológico de las ponedoras enjauladas durante un año de producción, muda y postmuda: producción de huevos, calidad de la cáscara del huevo, hígado, fémur y parámetros sanguíneos. Poult Sci. (1984) 63:339–33. DOI: 10.3382/PS.0630339
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Barreda DR, Konowalchuk JD, Rieger AM, Wong ME, Havixbeck JJ. Triennial growth symposium–novel roles for vitamin D in animal immunity and health. J Anim Sci. (2014) 92:930–8. doi: 10.2527/jas.2013-7341
Resumen de PubMed | Texto completo de CrossRef | Google Académico
52. Attia YA, Abd El-Hamid AE, Abedalla AA, Berika MA, El-Gandy MF, Sahin K, et al. Efecto de la betaína, la vitamina C y la vitamina E sobre la calidad del huevo, la incubabilidad y los marcadores de las funciones hepáticas y renales en gallinas reproductoras de doble propósito expuestas al estrés crónico por calor. Europ Poult Sci. (2018) 82. doi: 10.1399/eps2017171
53. Suaini NH, Zhang Y, Vuillermin PJ, Allen KJ, Harrison LC. Modulación inmune por la vitamina D y su relevancia para la alergia alimentaria. Nutrición. (2015) 7:6088–108 doi: 10.3390/nu7085271
Resumen de PubMed | Texto completo de CrossRef | Google Académico
54. Adams JS, Hewison M. Acciones inesperadas de la vitamina D: nuevas perspectivas sobre la regulación de la inmunidad innata y adaptativa. Nat Clin Pract Endocrinol Metab. (2008) 4:80–90. DOI: 10.1038/NCPENDMET0716
Resumen de PubMed | Texto completo de CrossRef | Google Académico
55. Li YC, Chen YZ, Liu WC, Thadhani R. MicroRNA-mediated mechanism of vitamin D regulation of innate immune response. J Esteroides Biochem Mol Biol. (2014) 144PA:81–6. doi: 10.1016/j.jsbmb.2013.09.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
56. Aslam SM, Garlich JD, Qureshi MA. La deficiencia de vitamina D altera las respuestas inmunes de los pollos de engorde. Poult Sci. (1998) 77:842–9. doi: 10.1093/ps/77.6.842
Resumen de PubMed | Texto completo de CrossRef | Google Académico
57. Lu L, Li SM, Zhang L, Liu XQ, Li DY, Zhao XL, et al. Expresión de beta defensinas en intestinos de pollos inyectados con vitamina D3 y lipopolisacárido. Genet Mol Res. (2015) 14:3330–7. doi: 10.4238/2015.abril.13.12
Resumen de PubMed | Texto completo de CrossRef | Google Académico
58. Morris A, Shanmugasundaram R, Lilburn MS, Selvaraj RK. 25- La suplementación con hidroxicolecalciferol mejora el rendimiento del crecimiento y disminuye la inflamación durante una inyección experimental de lipopolisacáridos. Poult Sci. (2014) 93:1951–6. DOI: 10.3382/PS.2014-03939
Resumen de PubMed | Texto completo de CrossRef | Google Académico
59. Morris A, Shanmugasundaram R, McDonald J, Selvaraj RK. Efecto del tratamiento in vitro e in vivo con 25-hidroxivitamina D en macrófagos, células T y pollos ponedoras durante un desafío de coccidios. J Anim Sci. (2015) 93:2894–903. DOI: 10.2527/jas.2014-8866
Resumen de PubMed | Texto completo de CrossRef | Google Académico
60. Shojadoost B, Behboudi S, Villanueva AI, Brisbin JT, Ashkar AA, Sharif S. La vitamina D3 modula la función de los macrófagos de pollo. Res Vet Sci. (2015) 100:45–51. doi: 10.1016/j.rvsc.2015.03.009
Resumen de PubMed | Texto completo de CrossRef | Google Académico
61. Krishnan AV, Feldman D. Mecanismos de las acciones anticancerígenas y antiinflamatorias de la vitamina D. Annu Rev Pharmacol Toxicol. (2011) 51: 311–36. doi: 10.1146/annurev-pharmtox-010510-100611
Resumen de PubMed | Texto completo de CrossRef | Google Académico
62. Xu S, Chen YH, Tan ZX, Xie DD, Zhang C, Xia MZ, et al.3 El pretratamiento alivia el estrés oxidativo renal en la lesión renal aguda inducida por lipopolisacáridos. J Esteroides Biochem Mol Biol. (2015) 152:133–41. doi: 10.1016/j.jsbmb.2015.05.009
Resumen de PubMed | Texto completo de CrossRef | Google Académico
63. Di Rosa M, Malaguarnera M, Nicoletti F, Malaguarnera L. Vitamina D3: un inmunomodulador útil. Inmunología. (2011) 134:123–39. doi: 10.1111/j.1365-2567.2011.03482.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
64. Zhang L, Lu L, Li S, Zhang G, Ouyang L, Robinson K, et al. 1,25-Dihidroxivitamina-D3 induce la expresión del gen beta-defensina aviar en pollos. PLoS ONE. (2016) 11:E0154546. doi: 10.1371/journal.pone.0154546
Resumen de PubMed | Texto completo de CrossRef | Google Académico
Palabras clave: gallinas ponedoras, calcio, VD3, calidad del huevo, respuestas inmunológicas, bioquímica sanguínea, microscopio electrónico
Cita: Attia YA, Al-Harthi MA y Abo El-Maaty HM (2020) Niveles de calcio y colecalciferol en gallinas ponedoras de fase tardía: efectos sobre rasgos productivos, calidad del huevo, bioquímica sanguínea y respuestas inmunes. Frente. Vet. Sci. 7:389. DOI: 10.3389/fvets.2020.00389
Recibido: 30 de marzo de 2020; Aprobado: 01 Junio 2020;
Publicado: 30 julio 2020.
Editado por:
Andrea Serra, Universidad de Pisa, Italia
Revisado por:
Sebastian Knaga, Universidad de Ciencias de la Vida de Lublin, Polonia
Mohamed E. Abd El-Hack, Universidad Zagazig, Egipto
Derechos de autor © 2020 Attia, Al-Harthi y Abo El-Maaty. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Youssef A. Attia, yaattia@kau.edu.sa; Hayam M. Abo El-Maaty, hayam151@yahoo.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía