
Lactobacillus plantarum GX17 beneficia el rendimiento del crecimiento y mejora las funciones de la barrera intestinal / flora intestinal

Lactobacillus plantarum GX17 beneficia el rendimiento del crecimiento y mejora las funciones de la barrera intestinal / flora intestinal entre pollos de engorde de plumas amarillas
 Yangyan Yin1,2,3,
Yangyan Yin1,2,3,  Yuying Liao1,2, Jun Li1,2, Zhe Pei4, Leping Wang1,2,3, Yan Shi1,2,3, Hongyan Peng1,2,3, Yizhou Tan1,2,3, Changting Li1,2, Huili Bai1,2, Chunxia Ma1,2, Yu Gong5, Tianchao Wei3* y Hao Peng1,2*
Yuying Liao1,2, Jun Li1,2, Zhe Pei4, Leping Wang1,2,3, Yan Shi1,2,3, Hongyan Peng1,2,3, Yizhou Tan1,2,3, Changting Li1,2, Huili Bai1,2, Chunxia Ma1,2, Yu Gong5, Tianchao Wei3* y Hao Peng1,2*- 1Guangxi Key Laboratory of Veterinary Biotechnology, Guangxi Veterinary Research Institute, Nanning, China
- 2Laboratorio clave de China (Guangxi)-Asociación de Naciones del Sudeste Asiático (ASEAN) Prevención y Control Transfronterizo de Enfermedades Animales, Ministerio de Agricultura y Asuntos Rurales de China, Nanning, China
- 3Instituto de Ciencia y Tecnología Animal, Universidad de Guangxi, Nanning, China
- 4Virginia Tech, Departamento de Ingeniería, Blacksburg, Nueva York, NY, Estados Unidos
- 5Estación Provincial de Gestión de Recursos Genéticos de Ganadería y Avicultura de Guizhou, Guiyang, China
Lactobacillus plantarum se ha encontrado recientemente para ser una fuente natural de bacterias aditivas para piensos con grandes ventajas en la seguridad alimentaria y el bienestar animal. El descubrimiento de nuevas cepas con potenciación de aplicaciones comerciales podría beneficiar a la industria avícola local y, en particular, apoyar a los agricultores chinos. En este estudio, probamos una nueva cepa recientemente aislada de Lactobacillus plantarum GX17 como aditivo para piensos en el rendimiento de crecimiento y las funciones de barrera intestinal de pollitos chinos de plumas amarillas de 1 día de edad. Tan bueno como otros probióticos comerciales, la alimentación con Lactobacillus plantarum GX17 mostró mejoras significativas en las respuestas inmunes humorales y mejoró el efecto inmune después de la vacunación para la vacuna contra la enfermedad de Newcastle o la vacuna contra la influenza aviar. Este estudio también encontró que la alimentación con Lactobacillus plantarum GX17 mejoró la relación alimentación-peso y causó un aumento significativo de la longitud de las vellosidades a la relación de profundidad de la cripta. Además, Lactobacillus plantarum GX17 reguló significativamente la expresión de ARNm de CLDN, MUC2 y TLR2, todos los cuales son genes de barrera asociados al yeyuno, lo que indica una mejora de las funciones de barrera intestinal al mejorar la unión estrecha entre las células del epitelio. Estos resultados son comparables a los efectos de la alimentación con probióticos complejos comerciales que mejoran los niveles de expresión de CLDN, ocludina, MUC2, TLR2 y TLR4. En términos de mantener la salud intestinal, los probióticos complejos comerciales aumentaron la abundancia relativa de Parabacteroides y Romboutsia, mientras que Lactobacillus plantarum GX17 aumentó la abundancia relativa de Pseudoflavonifractor. Nuestros datos sugieren que Lactobacillus plantarum GX17 podría mejorar la absorción intestinal de nutrientes y, por lo tanto, mejorar el rendimiento de crecimiento de los pollitos chinos de plumas amarillas. En conclusión, en comparación con los probióticos complejos comerciales, Lactobacillus plantarum GX17 tiene efectos más positivos sobre el rendimiento de crecimiento y la función de barrera intestinal de los pollos de plumas amarillas, y se puede utilizar como aditivo para piensos.
Introducción
Hoy en día, la velocidad con la que los patógenos pueden desarrollar tolerancia a los antibióticos puede ser incluso más rápida que el descubrimiento de nuevos compuestos antimicrobianos. Se ha convertido en un gran desafío para la salud pública a nivel mundial. Los residuos de antibióticos en los productos animales y el abuso de antibióticos causaron la aparición de cepas resistentes y son las dos razones principales de la evolución de los patógenos. China y la Unión Europea prohibieron el uso de antibióticos promotores del crecimiento como aditivos para piensos en 2020 y 2006, respectivamente (1), lo que ayudó a controlar la propagación de cepas resistentes a los antibióticos (2). Sin embargo, evitar el uso de antibióticos en los piensos ha introducido nuevos desafíos, incluido el mantenimiento del rendimiento del crecimiento y la supresión de infecciones bacterianas en pollos de engorde (3). Por lo tanto, se ha vuelto crucial desarrollar alternativas seguras y confiables a los antibióticos en la industria avícola. Los probióticos han sido ampliamente estudiados como una alternativa potencial a los antibióticos promotores del crecimiento, y para la búsqueda de cepas bacterianas calificadas ha atraído mucha atención recientemente en el campo.
Con efectos positivos en la prevención y el control de enfermedades, así como en la promoción del crecimiento, los probióticos se han utilizado ampliamente en las industrias ganadera y avícola (4-8). Lactobacillus plantarum es una de las bacterias del ácido láctico más utilizadas. Tiene efectos probióticos positivos y puede mejorar el rendimiento de crecimiento de los pollos de engorde (9-11) y tuvo buenos efectos en la prevención y el control de enfermedades (12). Por ejemplo, agregar cantidades optimizadas de Lactobacillus plantarum para alimentar aumentó la ingesta diaria promedio, el peso corporal, la materia seca, el nitrógeno y la energía total de los pollos de engorde en términos de digestibilidad aparente del intestino completo, redujo significativamente la cantidad de E. coli y aumentó el número de bacterias del ácido láctico (13). Además, Lactobacillus plantarum exhibió actividad antibacteriana contra Salmonella in vitro: inhibió significativamente el crecimiento, la adhesión y la invasión de Salmonella, al tiempo que redujo en gran medida su capacidad de formación de biopelículas (14). Además, la alimentación de Lactobacillus plantarum podría reducir el número de Salmonella typhimurium y ralentizar las respuestas inflamatorias a la infección en pollos de engorde neonatales mediante la modulación de la expresión de miRNA (15). Sin embargo, la seguridad de los probióticos está en debate y necesita ser explorada más a fondo. Se ha informado que tomar suplementos probióticos entre las personas mayores de 69 años de edad conduce no solo a un aumento de las bacterias intestinales potencialmente beneficiosas, sino también a un aumento en la activación de respuestas inmunes inespecíficas (16). Si bien los huéspedes inmunocomprometidos pueden beneficiarse más de la suplementación con probióticos, estos grupos también pueden tener un mayor riesgo de efectos adversos, como el desarrollo de sepsis, debido a su capacidad reducida para eliminar los microorganismos (17). Aunque comúnmente se informa que los probióticos protegen la barrera intestinal, puede haber casos en los que los probióticos no solo no restauran la barrera intestinal, sino que también promueven la translocación o las infecciones autoinducidas (18). Mientras tanto, no se ha estudiado el uso de Lactobacillus plantarum como aditivo en el agua potable para pollos de pollos de engorde chinos nativos de plumas amarillas. Los pollos de engorde de plumas amarillas tienen un peso corporal más pequeño, un ciclo de crecimiento más largo y una sabrosa calidad de la carne, lo que los hace adecuados para su uso en la cocina. Por lo tanto, en comparación con los pollos de engorde de plumas blancas (vendidos principalmente a KFC, McDonald’s y otras cadenas de restaurantes de comida rápida), los pollos de engorde de plumas amarillas son más populares entre los consumidores chinos con un mercado de 4.43 millones de pollos de engorde durante 2020 (19). Como los pollos de engorde de plumas amarillas generalmente se crían en granjas de corral, y el tiempo de reproducción es de dos a tres veces más largo que el de los pollos de engorde de plumas blancas, son más susceptibles a la infección por bacterias dañinas (20), y la demanda de productos antibacterianos alternativos es más urgente en comparación con los pollos de engorde de plumas blancas de cultivo a gran escala.
Una nueva cepa de Lactobacillus plantarum GX17 ha sido aislada e identificada en nuestro laboratorio y fue examinada in vitro por sus buenos efectos antibacterianos, pero su papel como aditivo alimenticio en el rendimiento del crecimiento de pollos de engorde y la función de barrera no está claro. Este estudio tuvo como objetivo comparar el rendimiento de crecimiento y el estado de salud de los pollos de engorde al alimentarlos con Lactobacillus plantarum GX17 o la dieta de probióticos complejos comerciales. El objetivo también fue explorar la cooperación entre un solo probiótico y un probiótico complejo comercial para investigar el potencial de Lactobacillus plantarum GX17 en aplicaciones de producción ganadera y avícola. Este estudio fue diseñado y realizado durante un ensayo de alimentación de 42 días. Se investigaron los efectos de Lactobacillus plantarum GX17 sobre los parámetros de crecimiento, índices inmunológicos, morfología intestinal yeyunal, función de barrera yeyunal y flora intestinal del ciego de pollos de engorde. Este estudio proporciona datos teóricos para los efectos de aplicación de Lactobacillus plantarum GX17 en la calidad y productividad de pollos de engorde de plumas amarillas.
Materiales y método
Fuentes de materiales
Todos los procedimientos con animales se realizaron de acuerdo con los protocolos aprobados por el Formulario Institucional de Inspección Ética Experimental de Animales del Instituto de Investigación Veterinaria de Guangxi, Nanning, China (8/2014/JU). El probiótico, Lactobacillus plantarum GX17, con una concentración final de alimentación de 1.5 × 10 ~ 9 UFC / animal, fue mantenido por el Laboratorio Clave de Biotecnología Veterinaria del Instituto de Investigación Veterinaria de Guangxi, Guangxi, China. El reactivo probiótico compuesto comercial fue comprado a una compañía biológica local (Shandong Baolaililai Biological Engineering Co., Ltd.).
Diseño experimental y cría de animales
Un total de 225 polluelos de plumas amarillas de 1 día de edad se dividieron aleatoriamente en tres grupos (15×5 animales por grupo) con un peso corporal similar: el grupo de alimentación de Lactobacillus plantarum GX17 (grupo LP), el grupo de alimentación de probióticos complejos comerciales (grupo S) y el grupo control (Grupo C). El grupo C fue alimentado con la dieta basal sin probióticos; el grupo LP fue alimentado con la dieta basal con Lactobacillus plantarum GX17 (1.5×10~9 UFC/cada uno) en el agua potable; el grupo S fue alimentado con la dieta basal y el agua potable con probióticos compuestos comerciales (1.5×10 ~ 9 UFC / cada uno). Las dietas de prueba se formularon de acuerdo con el Estándar de Alimentación de Pollo (Protocolo Estándar Nacional Chino: NY/T33-2004) que combinaba el ciclo de crecimiento y las características de la raza de los pollos de engorde de plumas amarillas. La Tabla 1 resume la composición y los niveles nutricionales de estas dietas (todas las dietas fueron fortificadas sin antibióticos). Los pollos de engorde de plumas amarillas se mantuvieron a una temperatura inicial de 31 ° C y se redujeron gradualmente en 2 ° C / semana hasta 25 ° C al final de la tercera semana. El peso corporal (PC) y la ingesta de alimento (FI) de todos los animales de prueba se registraron el último día del experimento.
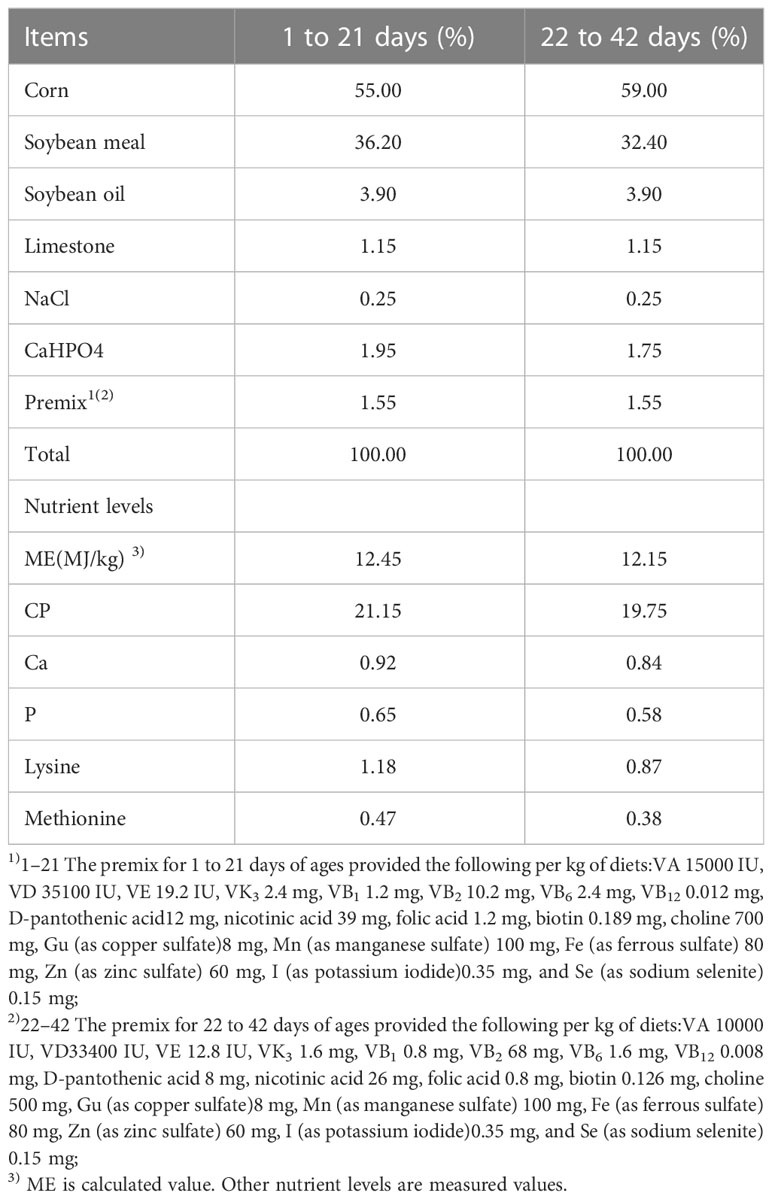 Cuadro 1 Composición de dietas basales (base aire-seca).
Cuadro 1 Composición de dietas basales (base aire-seca).
La evaluación de los resultados de crecimiento
El estado, el apetito y la defecación de todos los pollitos de prueba se observaron y registraron diariamente. Cada grupo de polluelos se pesó el último día del experimento y se ayunó durante 12 h sin agua antes de pesar. Se registró la cantidad de alimento suministrado, la cantidad de alimento restante y la cantidad de alimento perdido en la última etapa. El peso inicial, el peso final, la ingesta de alimento y la cantidad sobrante de cada etapa se registraron con precisión durante el experimento, y se calcularon la ganancia diaria promedio (ADG), la ingesta diaria promedio de alimento (ADFI) y la tasa de conversión alimenticia (FCR). En este estudio se utilizó la siguiente fórmula: Aumento de peso diario promedio = (peso promedio al final del ensayo – peso promedio al principio)/número de días en el ensayo; Ingesta diaria media de alimento = consumo total de alimento durante el período de prueba/(número de días de prueba × número de pollos); y Índice de conversión alimenticia = ingesta diaria promedio de alimento / ganancia de peso diaria promedio.
La determinación de los índices de órganos inmunes
En el día 21 y el día 42 del experimento, tres pollos de engorde fueron seleccionados al azar por réplica para ser pesados y sacrificados. El bazo, el timo y la bolsa se extirparon cuidadosamente, aislando el tejido adiposo, cada uno de los órganos inmunes se pesó y calculó para el índice de órganos inmunes. Los índices de órganos inmunes fueron generalmente el peso del órgano (mg) por 10 g de peso corporal utilizando las siguientes fórmulas: Índice del timo = [peso del timo (g) * 103]/[peso del pollo vivo (g)/10] = [peso del timo (g)/peso del pollo vivo (g)] * 104. Índice del bazo = [peso del bazo (g) * 103]/[peso del pollo vivo (g)/10] = [peso del bazo (g)/peso del pollo vivo (g)] * 104. Índice de bolsa = [peso de la bolsa(g)* 103]/[peso del pollo vivo(g)/10] = [peso de la bolsa(g)/peso del pollo vivo(g)]*104.
Ensayo de inmunoanticuerpos
Los procedimientos de inmunización comprendieron la inmunización por inyección subcutánea de la vacuna MD a 1 día de edad, la inmunización ojo-intranasal de 7 días con nueva vacuna combinada de rama (cepa LaSota + cepa H120) y la nueva vacuna triple tributaria de 14 días de edad 0,5 ml inyección subcutánea (cepa LaSota + cepa M41 + cepa H2). En el 7ésimo y 28ésimo Un día después de la inmunización, tres animales fueron seleccionados al azar por réplica. Se recogió sangre de las venas del viento y se aisló suero y se analizó la potencia de los anticuerpos contra la enfermedad de Newcastle y la gripe aviar H9. El ensayo de inhibición de la hemaglutinación se utilizó para valorar las respuestas de anticuerpos.
Medición de indicadores inmunológicos séricos
Tres pollos fueron seleccionados aleatoriamente por réplica el día 21 y el día 42 y ayunaron durante 8 horas, después de lo cual se recolectaron 5 ml de sangre de la vena del ala de cada animal utilizando un tubo de recolección de sangre no anticoagulada. Las muestras de sangre entera se asentaron durante 30 minutos a temperatura ambiente y luego se centrifugaron a 3000 rpm / minuto durante 20 minutos. La capa superior de suero se tomó y se almacenó a -80 ° C. Se utilizó el método ELISA para la determinación de moléculas de IgG, IgA, IgM, CD4 y CD8, y el proceso de operación y cálculo se llevó a cabo en estricta conformidad con las instrucciones del kit. Los kits fueron comprados a Jiangsu Jingmei Technology Co.
Observaciones morfológicas para el intestino duodenal y yeyunal
El día 21 y el día 42, respectivamente, tres pollos por réplica fueron seleccionados al azar y sacrificados después del ayuno nocturno. Se recogieron muestras de tejido intestinal duodenal y yeyunal, se enjuagaron suavemente con tampón PBS para eliminar el contenido intestinal y luego se colocaron en fijador de paraformaldehído al 4% para la fijación. En el Instituto de Medicina Tradicional China de Guangxi, se prepararon secciones de tejido y se midieron bajo el microscopio. Se tomaron y calcularon datos de la altura de las vellosidades duodenales y yeyunales y de la profundidad de la cripta, y se recogió la relación altura de las vellosidades/profundidad de la cripta (VCR). Para cada muestra, se tomaron dos secciones consecutivas y se seleccionaron aleatoriamente tres campos de visión diferentes. Las alturas de las vellosidades (μm) se midieron desde la punta hasta la unión de la cripta de las vellosidades, y la profundidad de la cripta se definió como la profundidad de la invaginación de las vellosidades adyacentes. Las razones se calcularon utilizando el valor medio de cada grupo para una comparación adicional (Figura 1).
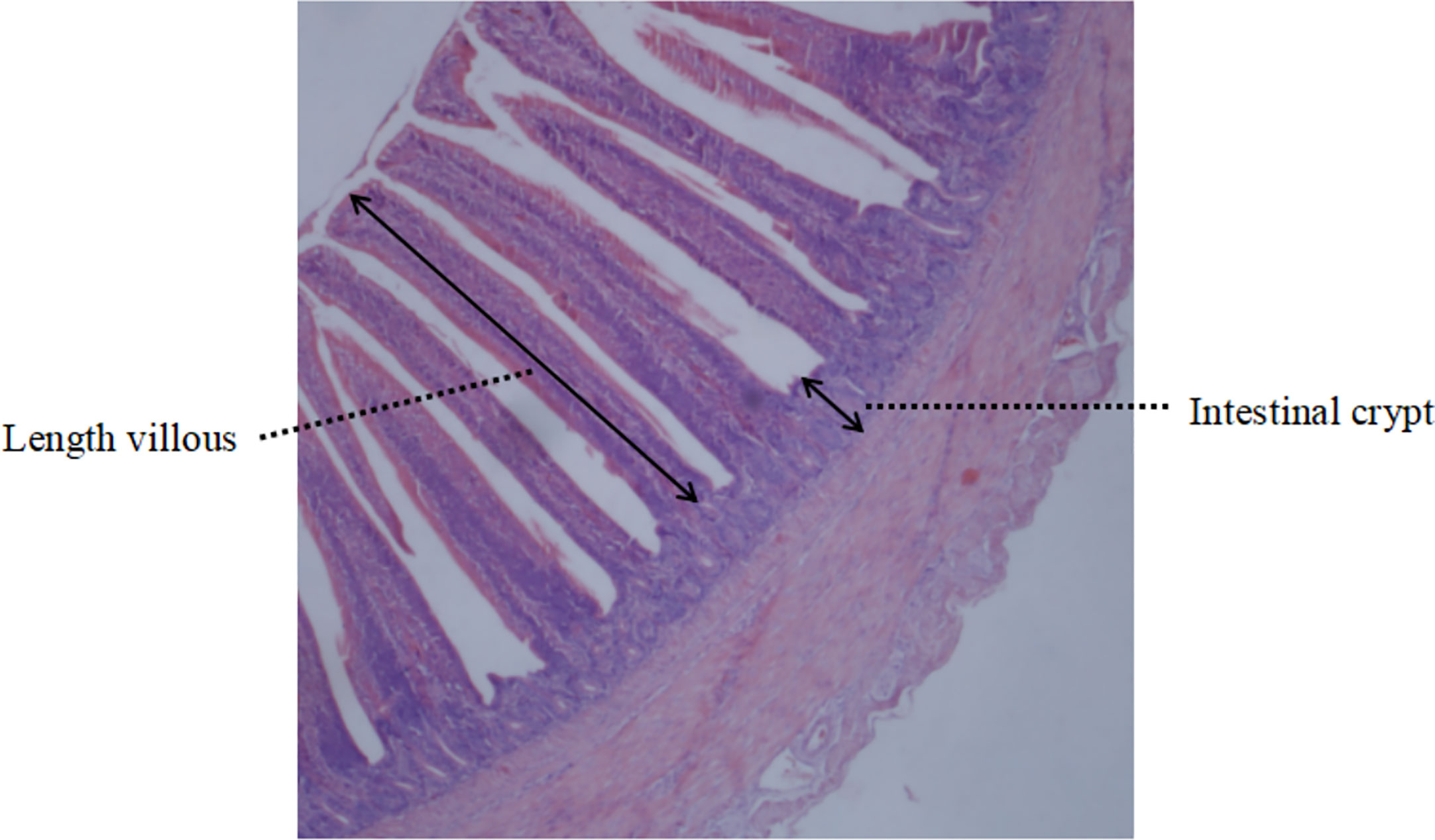 Figura 1 Diagrama esquemático de las vellosidades intestinales.
Figura 1 Diagrama esquemático de las vellosidades intestinales.
La expresión del ARNm de la proteína yeyunal por el método RT-qPCR
En el día 42, tres pollos por réplica fueron seleccionados al azar y sacrificados después del ayuno nocturno. Se recogieron muestras de tejido yeyunal y se colocaron en nitrógeno líquido. Para la extracción de ARN (realizada a 4 °C), se agregaron 100 mg de intestino vacío a 900 μL de solución salina y luego se molieron con un molinillo de tejido, y las muestras se sometieron a extracción de ARN de acuerdo con el procedimiento de extracción del kit de tejido ARNpure. La transcripción inversa se realizó utilizando el kit de síntesis de ADNc HiFiScript. La temperatura de reacción fue de 42 ° C durante 15 minutos y la reacción se terminó aumentando la temperatura a 85 ° C durante 5 segundos. Se utilizó el kit UltraSYBR Miture para cuantificar el nivel de expresión de ARNm relevante de la muestra. Cada reacción contenía 1 μL de plantilla de ADNc, 0,5 μL de cada cebador, mezcla maestra (12,5 μL) y 10,5 μL ddH2O para un volumen total de 25 μL. La PCR cuantitativa fluorescente (qPCR) se realizó mediante el siguiente procedimiento: 95°C durante 5 min, seguido de 40 ciclos (95°C durante 15 s y 60°C durante 1 min). Después de la amplificación, la curva de fusión se realizó incubando la reacción a 60 ° C durante 30 s y luego aumentando la temperatura de 60 ° C a 95 ° C a una velocidad de 0.2 ° C / s. Todas las secuencias de cebadores y condiciones de prueba se muestran en la Tabla 2.
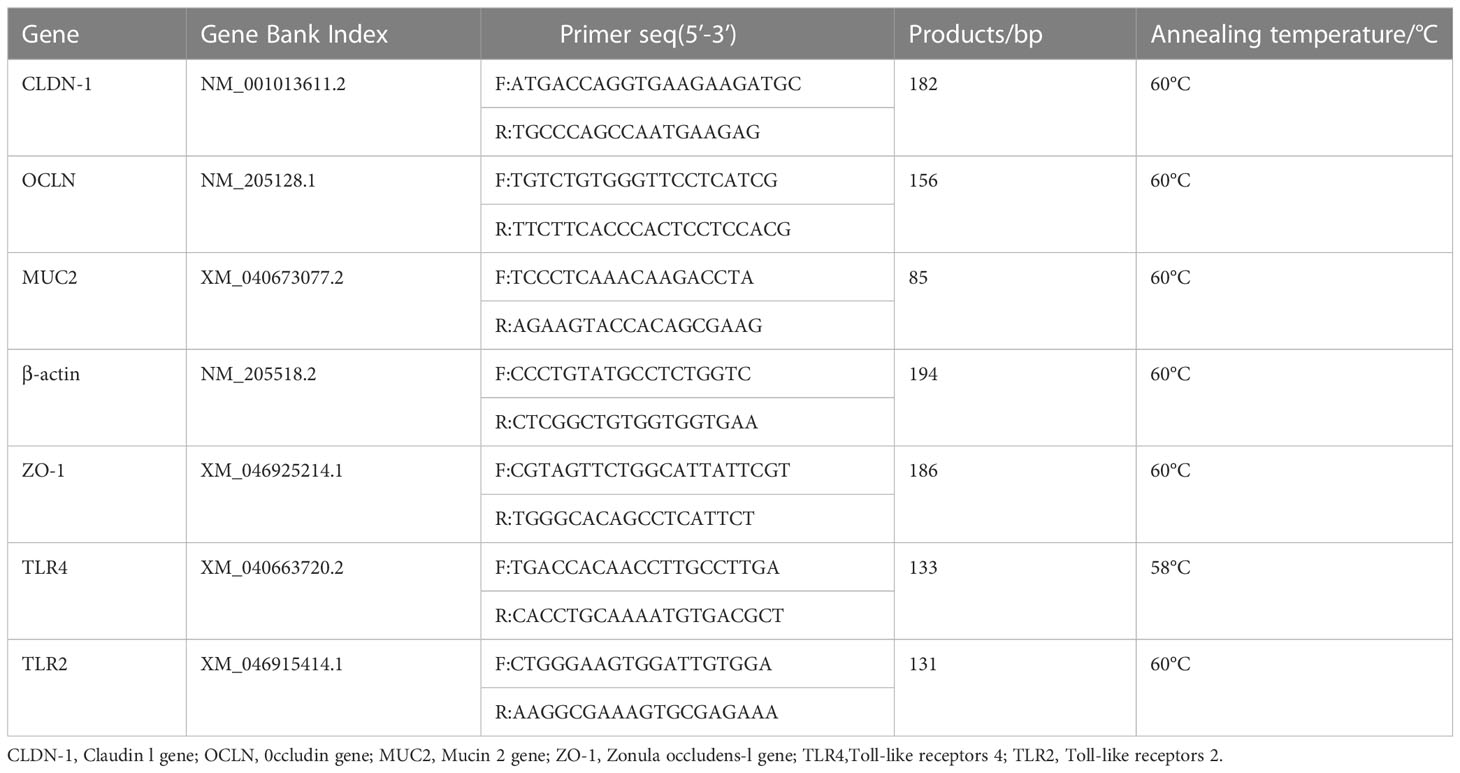 Cuadro 2 PCR cuantitativa fluorescente.
Cuadro 2 PCR cuantitativa fluorescente.
El análisis del ADNr 16S para la población de flora intestinal
En el día 42, tres pollos de cada grupo de prueba fueron seleccionados al azar y sacrificados después del ayuno nocturno, momento en el que se recolectaron los contenidos cecales. El ADN genómico total se extrajo utilizando el kit de extracción de ADN del genoma bacteriano (Beijing Kangwei Century Biotechnology Co., Ltd.) siguiendo las instrucciones del fabricante. La concentración y la integridad del ADN se midieron con Implen NanoPhotometer y electroforesis en gel de agarosa. El ADN extraído se almacenó a -20 ° C hasta su posterior procesamiento. El ADN extraído se utilizó como plantilla para la amplificación por PCR de genes bacterianos 16S rRNA con los cebadores con código de barras y Takara Ex Taq (Takara). Para el análisis de la diversidad bacteriana, las regiones variables V3-V4 (o V4-V5) de los genes 16S rRNA se amplificaron con cebadores universales 343F (5′-TACGGRAGGCAGCAG-3′) y 798R (5′-AGGGTATCTAATCCT-3′) y 907R (5′-CCGTCAATTCMTTTRAGTTT-3′) para las regiones V3-V4.
La calidad del Amplicón se visualizó mediante electroforesis en gel de agarosa. Los productos de PCR se purificaron con perlas AMPure XP (Agencourt) y se amplificaron para otra ronda de PCR. Después de ser purificado con las perlas AMPure XP nuevamente, el amplicón final se cuantificó utilizando Qubit dsDNA Assay Kit (Thermo Fisher Scientific, EE. Las concentraciones se ajustaron para la secuenciación. La secuenciación se realizó en un Illumina NovaSeq 6000 con lecturas de extremo pareado de 250 pb. (Illumina Inc., San Diego, CA; OE Biotech Company; Shanghai, China).
La secuenciación de la biblioteca y el procesamiento de datos fueron realizados por OE biotech Co., Ltd. (Shanghai, China). Los datos de secuenciación sin procesar estaban en formato FASTQ. Las lecturas de extremo emparejado se procesaron previamente utilizando el software Cutadapt para detectar y cortar el adaptador. Después del recorte, las lecturas de extremo emparejado se filtraron para secuencias de baja calidad, se eliminaron el ruido, se fusionaron y se detectaron, y las lecturas de quimera se cortaron utilizando DADA2 con los parámetros predeterminados de QIIME2 (2020.11). Finalmente, la salida del software lee el representante y la tabla de abundancia ASV. La lectura representativa de cada ASV se seleccionó utilizando el paquete QIIME2. Todas las lecturas representativas fueron anotadas y criticadas contra la base de datos Silva (Versión 138) usando q2-feature-classifier con los parámetros predeterminados.
Se utilizó el software QIIME2 para el análisis de diversidad alfa y beta. La diversidad microbiana en las muestras se estimó utilizando la diversidad alfa que incluye el índice Chao1 y el índice de Shannon. Luego se utilizó el paquete R para analizar las diferencias significativas entre los diferentes grupos utilizando la prueba estadística ANOVA.
Análisis estadístico
Los datos experimentales fueron analizados estadísticamente utilizando SPSS 25.0. Los datos se presentaron como medias y se agruparon SEM. Los efectos del tratamiento dietético sobre las variables medidas se analizaron mediante ANOVA unidireccional seguido de la prueba de comparación múltiple de Duncan para comparar entre grupos. Las diferencias se consideraron significativas a P < 0,05.
Resultados
Lactobacillus plantarum GX17 mostró efectos positivos en el rendimiento de crecimiento de pollos de engorde de plumas amarillas
Como se muestra en la Figura 2, aunque no hubo diferencias significativas entre el aumento de peso diario promedio de los pollos de engorde de plumas amarillas en los grupos LP y S en comparación con el grupo Control, durante todo el período de prueba, el aumento de peso diario promedio de los pollos de engorde de plumas amarillas en los grupos LP y S que fueron alimentados con probióticos aumentó en 0.927 ± 1.33 g y 0.926 ± 1.52 g respectivamente en comparación con los controles. La ingesta diaria promedio de alimento de pollos de engorde de plumas amarillas en los grupos LP y S con probióticos agregados disminuyó significativamente en aproximadamente 1.497 ± 0.17 g y 1.714 ± 0.13 g (P<0.001). La ingesta media diaria de alimento de los grupos LP y S con probióticos fue significativamente menor, alrededor de 1.497 ± 0.17 g y 1.714 ± 0.13 g (P<0.001). Por lo tanto, después de 42 días de alimentación con probióticos, la tasa de conversión alimenticia fue significativamente menor en los grupos LP y S (P<0.001).
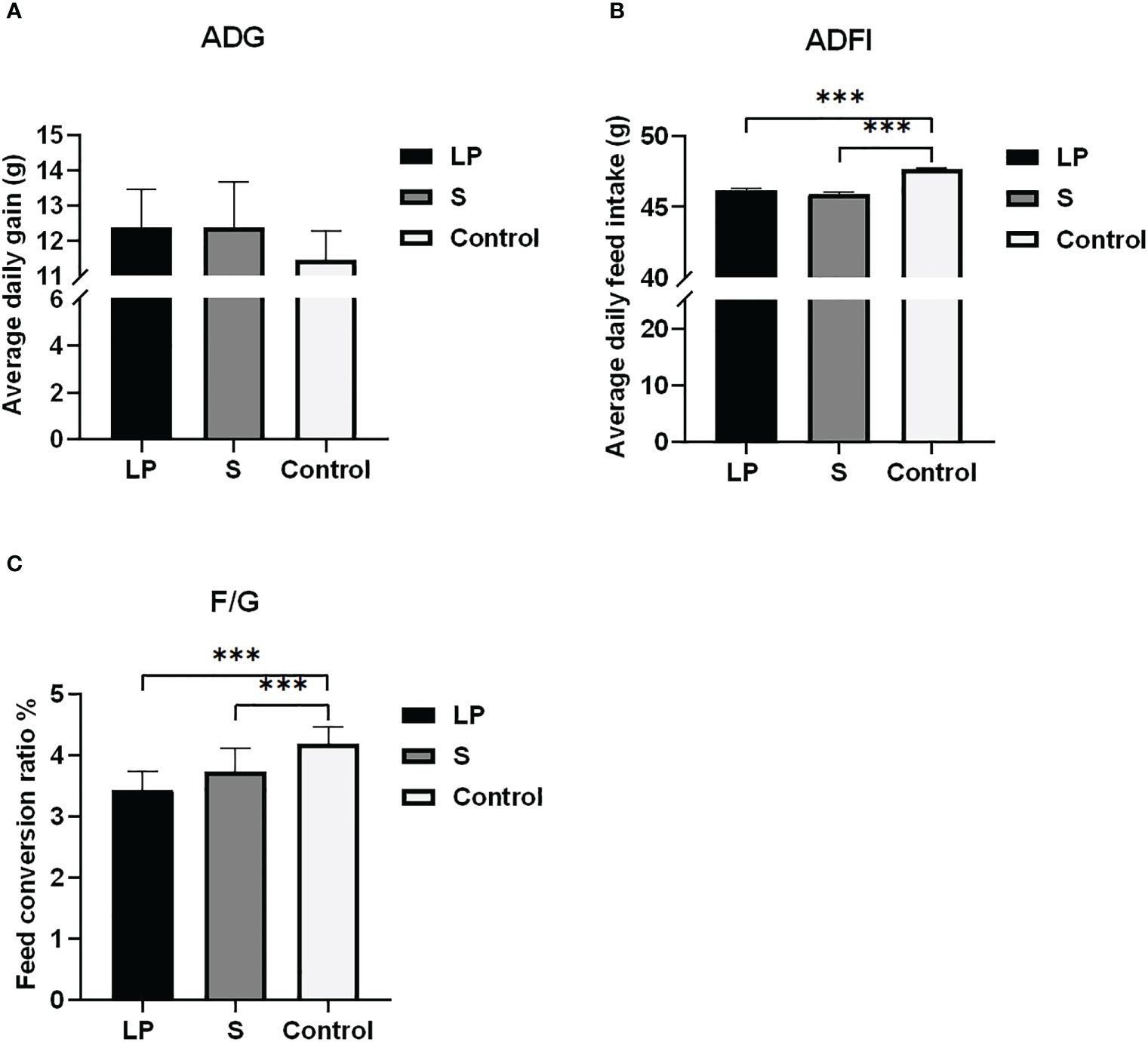 Figura 2 Efectos de Lactobacillus plantarum GX17 en el rendimiento de crecimiento de pollos de engorde. (A) La ganancia diaria promedio de los tres grupos de pollos de engorde de plumas amarillas en el día 42; (B) Ingesta diaria promedio de alimento de tres grupos de pollos de engorde de plumas amarillas en el día 42; (C) La relación entre el alimento y la ganancia de pollos de engorde de plumas amarillas en los tres grupos (***p<0.001).
Figura 2 Efectos de Lactobacillus plantarum GX17 en el rendimiento de crecimiento de pollos de engorde. (A) La ganancia diaria promedio de los tres grupos de pollos de engorde de plumas amarillas en el día 42; (B) Ingesta diaria promedio de alimento de tres grupos de pollos de engorde de plumas amarillas en el día 42; (C) La relación entre el alimento y la ganancia de pollos de engorde de plumas amarillas en los tres grupos (***p<0.001).
Lactobacillus plantarum GX17 mejoró las respuestas inmunes entre los pollos de engorde de plumas amarillas
Como se muestra en la Figura 3, Lactobacillus plantarum GX17 no tuvo un efecto significativo sobre el índice de órganos inmunes de pollos de engorde de plumas amarillas (P > 0.05). Sin embargo, mejoró la potencia de anticuerpos de pollos de engorde de plumas amarillas inmunizados con la vacuna contra la enfermedad de Newcastle y la vacuna contra la gripe aviar (Figura 4) en comparación con el grupo de control. La potencia de anticuerpos de la enfermedad de Newcastle fue significativamente mayor en el grupo LP en el 7º día después de la inmunización secundaria con un aumento de aproximadamente 1,08 potencia (P = 0,013). La potencia de anticuerpos de la gripe aviar fue significativamente mayor en el grupo S, con un aumento de aproximadamente 1,65 potencias (p = 0,009). No hubo diferencias significativas en la potencia de anticuerpos del virus de la enfermedad de Newcastle y el virus de la gripe aviar entre los grupos LP y S en el día 28 después de la inmunización secundaria (P>0,05). La alimentación de Lactobacillus Plantarum GX17 también mostró efectos positivos sobre el contenido de inmunoglobulina en el suero de pollos de engorde de plumas amarillas (Figura 5). En el día 21, en ambos grupos que fueron alimentados con Lactobacillus plantarum GX17 o probióticos complejos comerciales, los contenidos de IgM, IgG, CD4 y CD8 entre los grupos no cambiaron (P>0,05); aunque el grupo S tenía un mayor contenido de IgA (comparar con el grupo LP: P=0,005; comparar con los controles: P=0,043). Sin embargo, al día 42 de alimentación, CD4 fue significativamente mayor en los grupos LP y S en comparación con el grupo Control, con un aumento del 102% y 121%, respectivamente (P<0,001), mientras que los contenidos de IgM, IgA, IgG y CD8 no cambiaron significativamente (P>0,05).
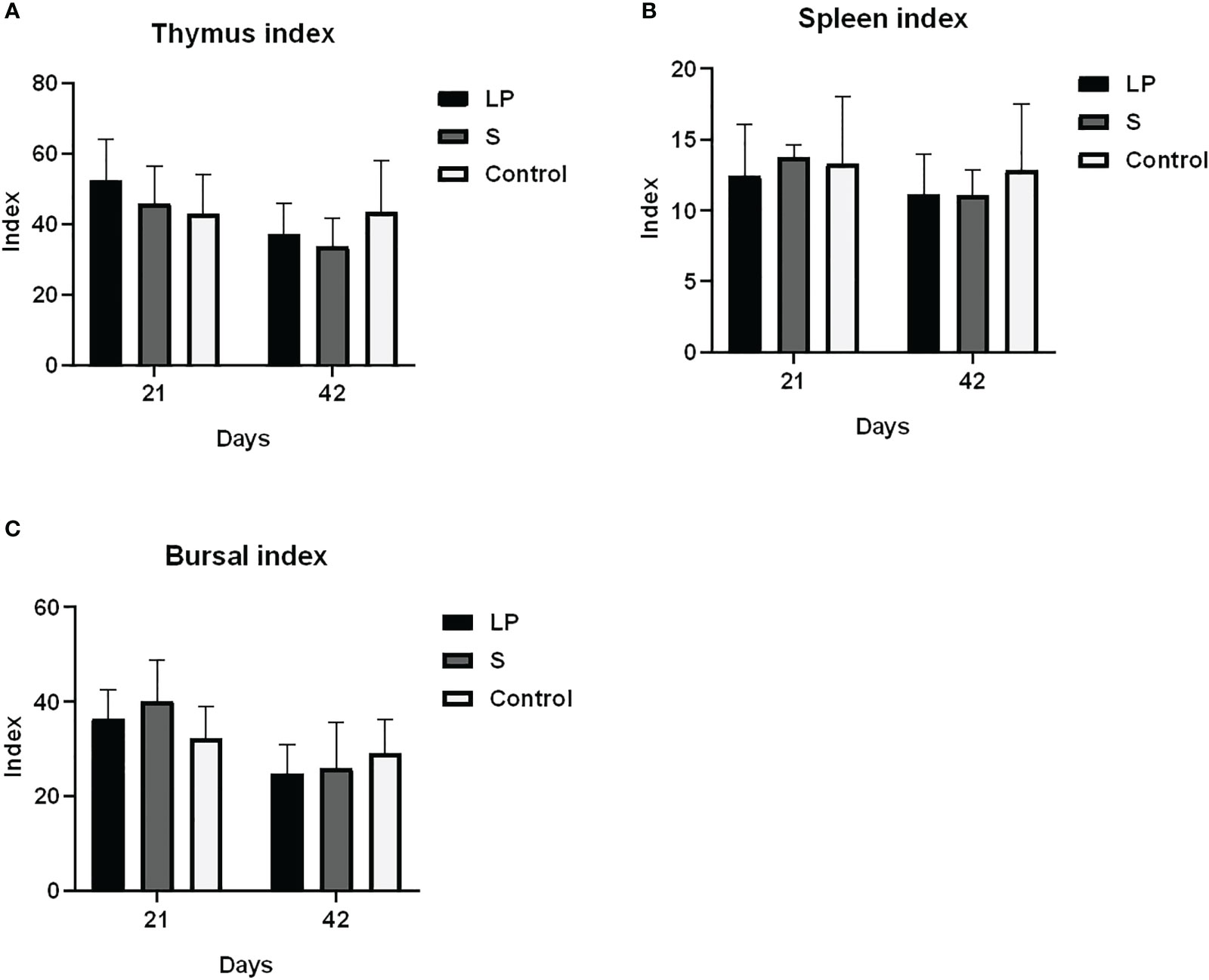 Figura 3 Efectos de Lactobacillus plantarum GX17 sobre el índice de órganos inmunes de pollos de engorde de plumas amarillas. (A) índice del timo (B) índice del bazo (C) índice de las bolsas.
Figura 3 Efectos de Lactobacillus plantarum GX17 sobre el índice de órganos inmunes de pollos de engorde de plumas amarillas. (A) índice del timo (B) índice del bazo (C) índice de las bolsas.
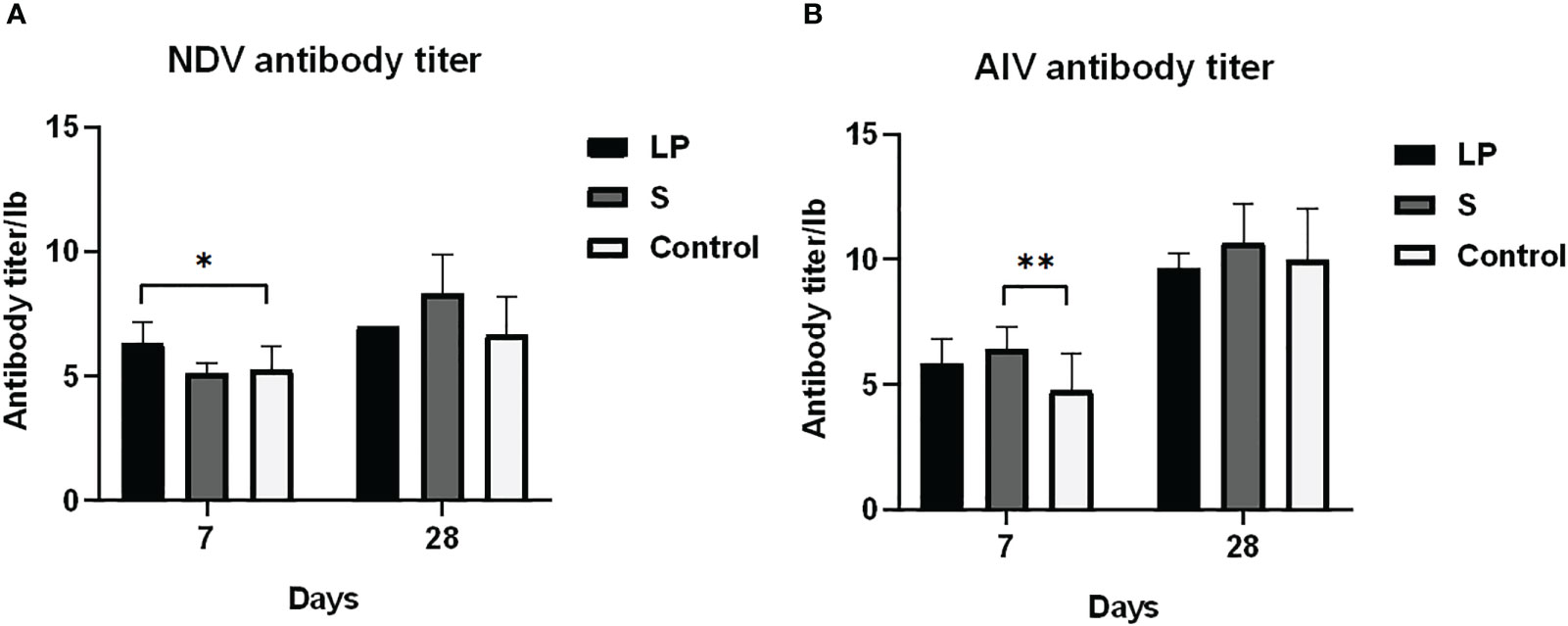 Figura 4 Título de anticuerpos de Lactobacillus plantarum GX17 después de la inmunización contra pollos de engorde de plumas amarillas. (7 y 28 días después de la inmunización) (A) Título de anticuerpos contra la enfermedad de Newcastle (B) título de anticuerpos contra la influenza aviar (*p<0.05; **p<0.01).
Figura 4 Título de anticuerpos de Lactobacillus plantarum GX17 después de la inmunización contra pollos de engorde de plumas amarillas. (7 y 28 días después de la inmunización) (A) Título de anticuerpos contra la enfermedad de Newcastle (B) título de anticuerpos contra la influenza aviar (*p<0.05; **p<0.01).
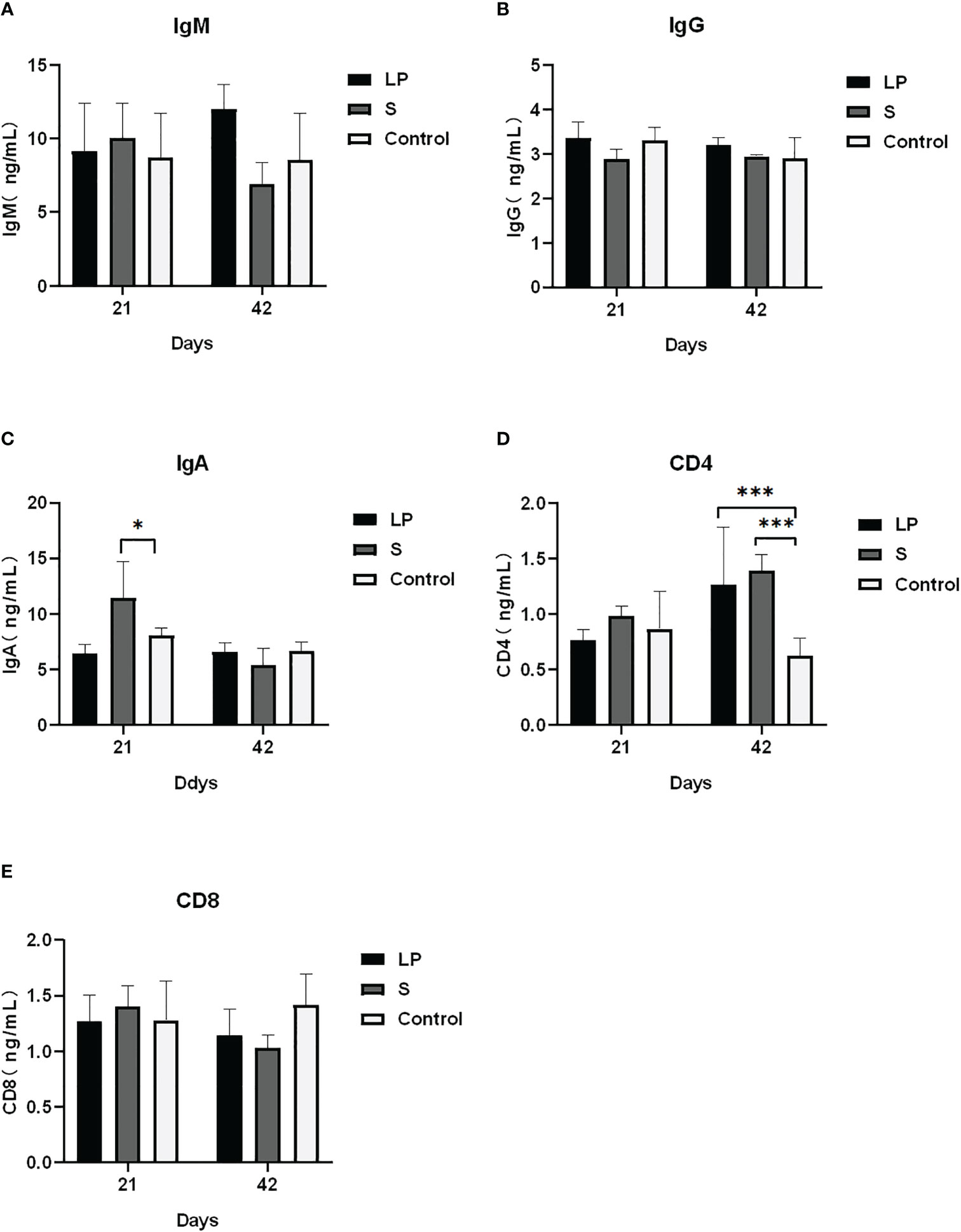 Figura 5 Efecto de Lactobacillus plantarum GX17 sobre el contenido sérico de inmunoglobulina de pollos de engorde de plumas amarillas. (*p<0,05; ***p<0,001).
Figura 5 Efecto de Lactobacillus plantarum GX17 sobre el contenido sérico de inmunoglobulina de pollos de engorde de plumas amarillas. (*p<0,05; ***p<0,001).
Lactobacillus plantarum GX17 mejoró la salud intestinal de pollos de engorde
Lactobacillus plantarum GX17 promovió el crecimiento de vellosidades intestinales, facilitando así la absorción de nutrientes. En el grupo S, después de alimentar probióticos durante 21 días, la relación de VCR duodenal aumentó en un 34,7% (P = 0,017) y la profundidad de la cripta duodenal aumentó en un 31,9% (P <0,001) en comparación con los controles. En el grupo LP, aunque el valor de VCR duodenal tuvo una tendencia de aumento del 22,4% (P = 0,051, >0,05), no fue estadísticamente significativo en comparación con el grupo control; La mejora en la profundidad de la cripta duodenal tampoco fue significativa. En yeyuno, el grupo S mostró una mejora significativa del valor de VCR (+43,9%, p = 0,006), aunque la longitud de las vellosidades y la profundidad de la cripta mantuvieron un nivel similar al de los controles. Curiosamente, cuando la duración del tratamiento probiótico se extendió a 42 días, no se observaron diferencias significativas en la longitud de las vellosidades, la profundidad de la cripta o los valores de VCR entre los tres grupos, excepto que el valor de VCR del grupo LP aumentó en un 34,1% (p = 0,049) (Figura 6). Esto sugiere que los probióticos pueden tener un efecto facilitador más fuerte para el crecimiento de pollos de engorde de plumas amarillas más jóvenes, mientras que los cambios morfológicos disminuyeron cuando el yeyuno y el duodenal maduraron. Sin embargo, la alimentación de probióticos durante más tiempo, particularmente GX17, podría tener ventajas para mejorar la expresión de los genes de barrera yeyunal. Después de alimentar a Lactobacillus plantarum GX17 durante 42 días, los niveles de CLDN, MUC2 y TLR2 fueron significativamente más altos en aproximadamente 2.99, 103 y 22.37 veces, respectivamente, en el grupo LP en comparación con los controles (P < 0.001). Para la ocludina, ZO-1 y TLR4, los tres mostraron una tendencia al aumento en comparación con los controles, aunque el resultado no fue estadísticamente significativo. Cuando se alimentó con probióticos complejos comerciales en el grupo S, el nivel de CLDN, ocludina, MUC2, TLR2 y TLR4 aumentó significativamente en 3.04, 4.96, 74.98, 60.30 y 6.57 pliegues en comparación con los controles (P < 0.001), mientras que solo el contenido de ZO-1 no cambió significativamente.
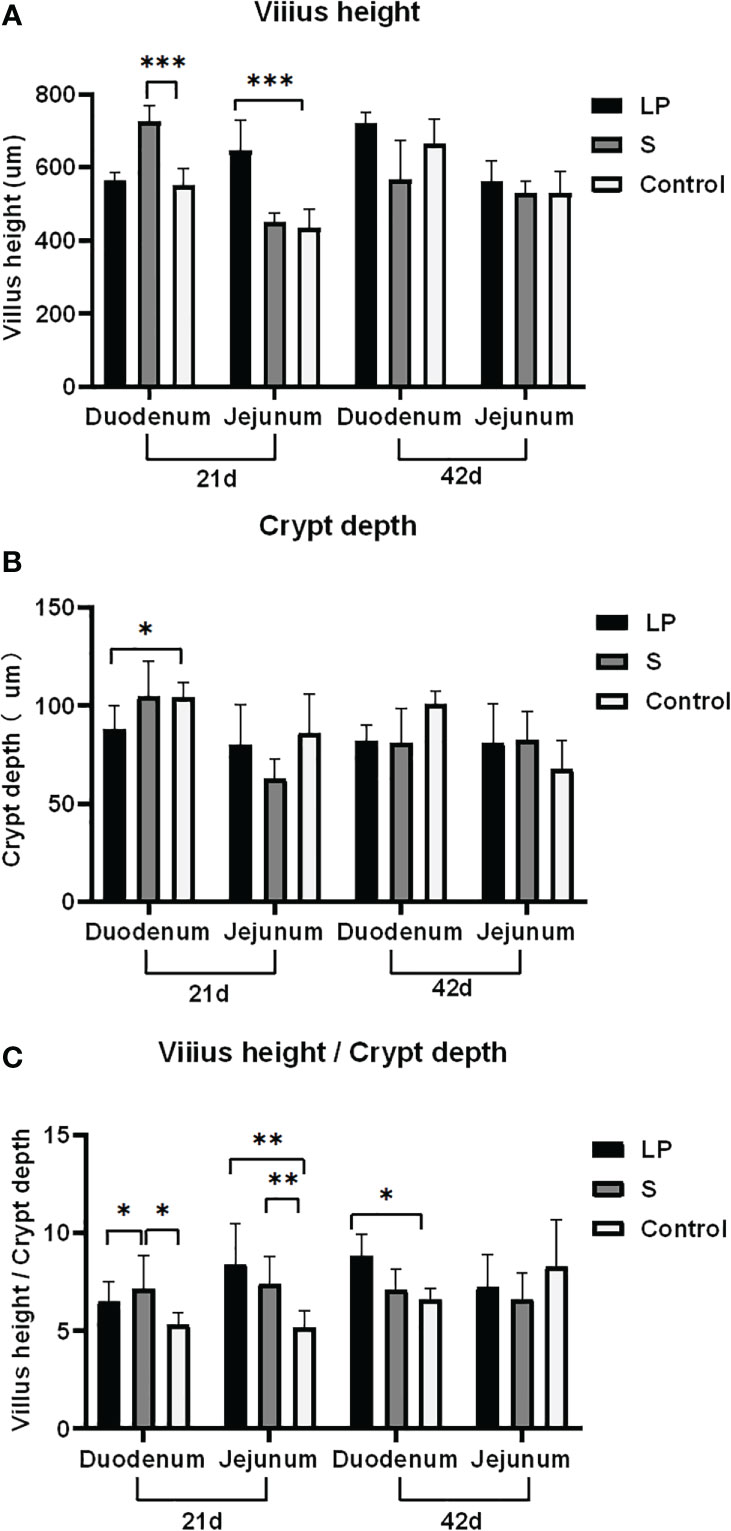
Figura 6 Efecto de Lactobacillus plantarum GX17 sobre las vellosidades intestinales del duodeno y yeyuno en pollos de engorde de plumas amarillas. (*p<0,05; **p<0,01; ***p<0,001).
Efectos de la alimentación con Lactobacillus plantarum GX17 sobre la abundancia de flora intestinal entre pollos de engorde de plumas amarillas
Para dilucidar los mecanismos subyacentes de la potenciación resistente a la enfermedad entre pollos de engorde de plumas amarillas que fueron alimentados con probióticos, exploramos los cambios de la microbiota del ciego bajo diversas condiciones de prueba. Analizamos la diversidad de su microbiota cecum a nivel de filo y género para caracterizar la dinámica de la distribución taxonómica microbiana. Los datos mostraron que a nivel de filo, todos los grupos estaban dominados por Bacteroidetes, Firmicutes y Proteobacteria (Figura 7A). Además, comparamos la composición bacteriana del ciego a nivel de género: se encontró que otro conjunto de géneros, que incluyen Alistipes, Bacteroides, Clostridia y Ruminococcus, eran más dominantes (Figura 7B). El análisis ANOVA mostró que a nivel de género, la abundancia relativa de Parabacteroides y Romboutsia en el grupo S fue 3,66 y 6,17 veces mayor que en el grupo LP, mientras que estos resultados fueron 2,64 y 1,70 veces mayores que los controles, respectivamente. Todos estos niveles crecientes fueron estadísticamente significativos, aunque no hubo diferencias significativas entre los grupos LP y Control. La abundancia relativa de Pseudoflavonifractor en el grupo LP fue 2,10 y 3,66 veces mayor que en los grupos S y Control, respectivamente, (Figuras 8, 9).
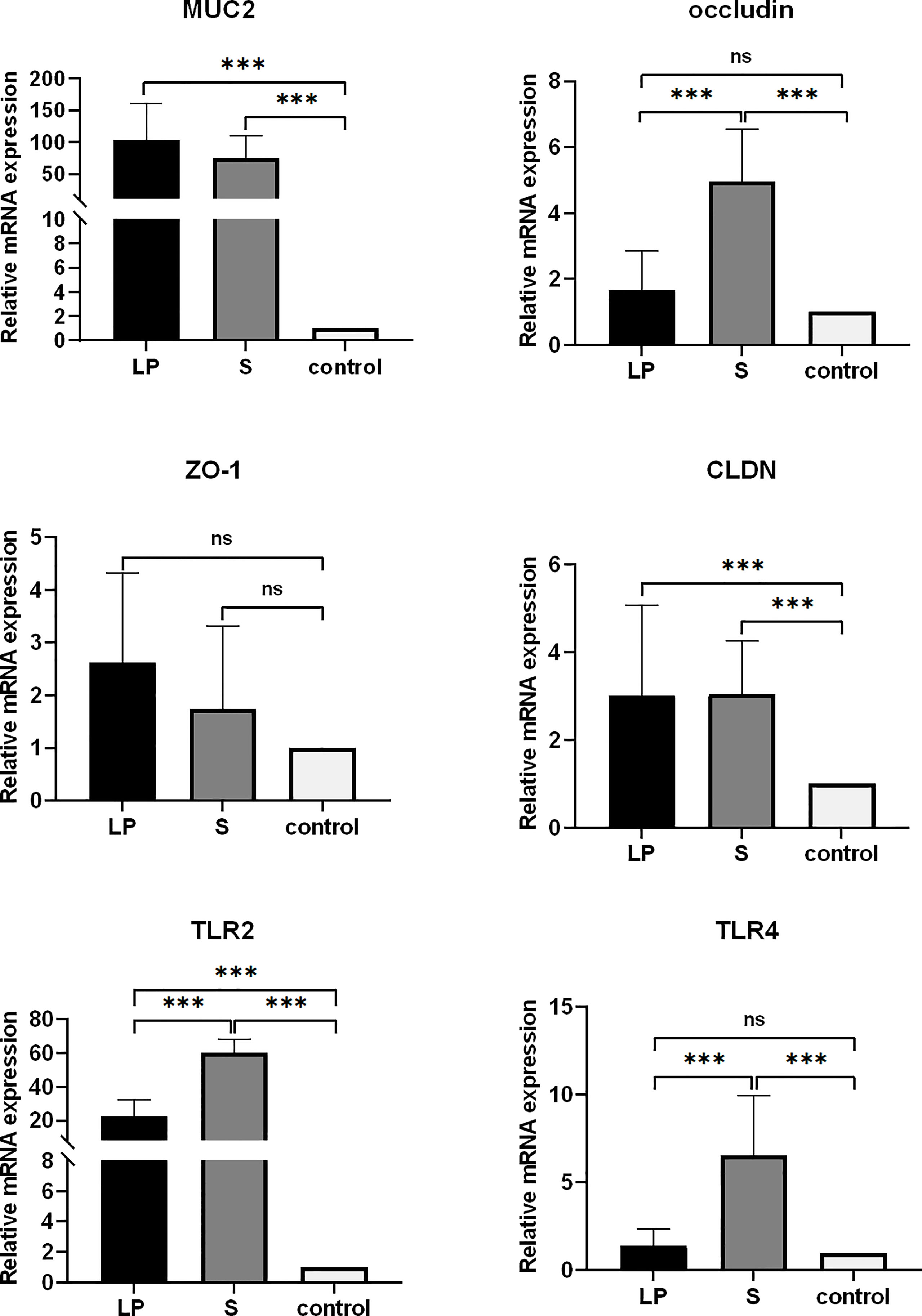 Figure 7 mRNA expression of jejunal barrier related genes (42 days) (all Control values are normalized) CLDN-1: Claudin l gene; OCLN: 0ccludin gene; MUC2: Mucin 2 gene; ZO-1: Zonula occludens-l gene;TLR4:Toll-like receptors 4; TLR2: Toll-like receptors 2 (***p<0.001) ns, no significance.
Figure 7 mRNA expression of jejunal barrier related genes (42 days) (all Control values are normalized) CLDN-1: Claudin l gene; OCLN: 0ccludin gene; MUC2: Mucin 2 gene; ZO-1: Zonula occludens-l gene;TLR4:Toll-like receptors 4; TLR2: Toll-like receptors 2 (***p<0.001) ns, no significance.
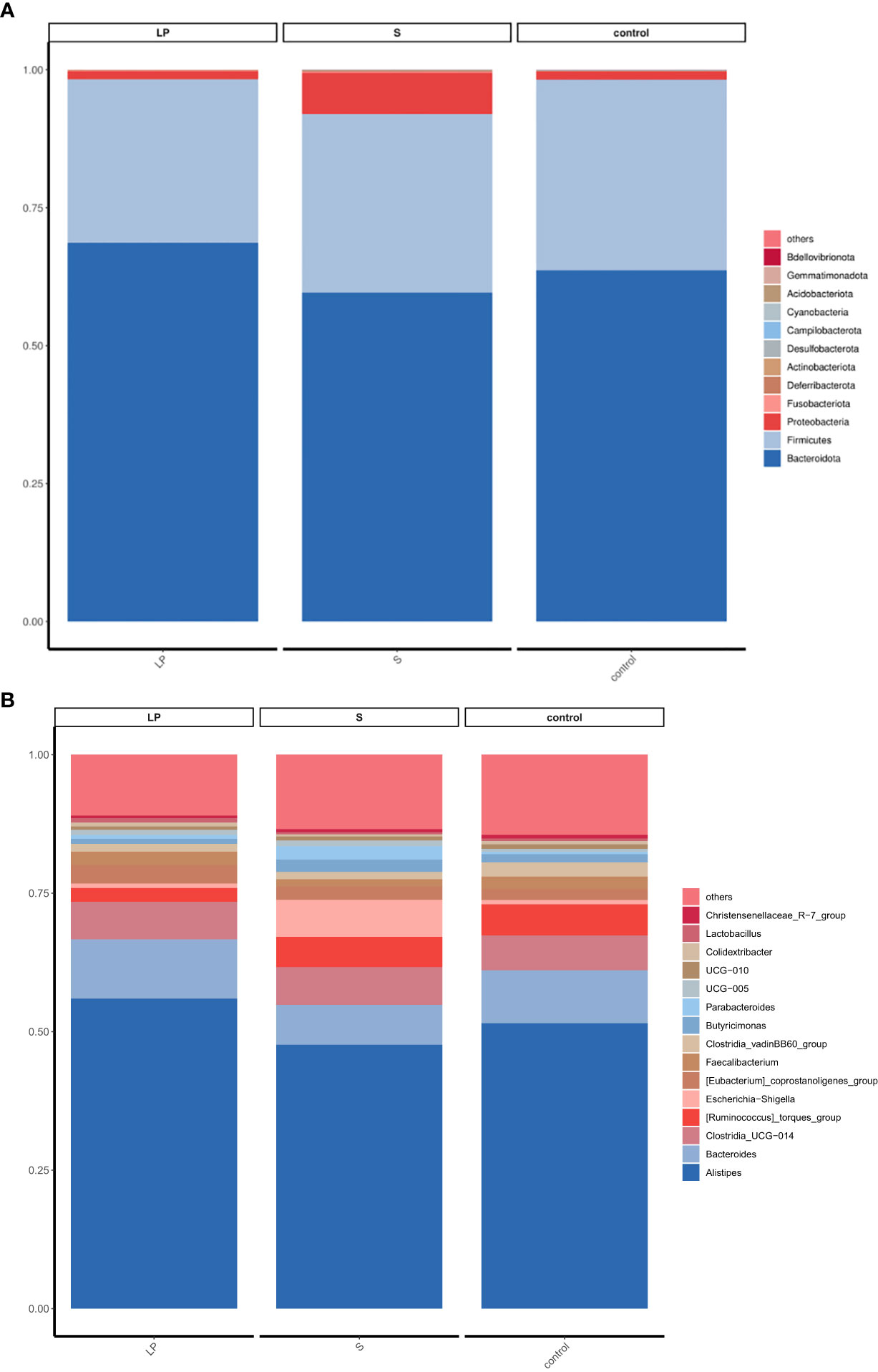 Figura 8 Diferencia de abundancia relativa a nivel de puerta y género (42d): (A) nivel de filo; (B) nivel de género.
Figura 8 Diferencia de abundancia relativa a nivel de puerta y género (42d): (A) nivel de filo; (B) nivel de género.
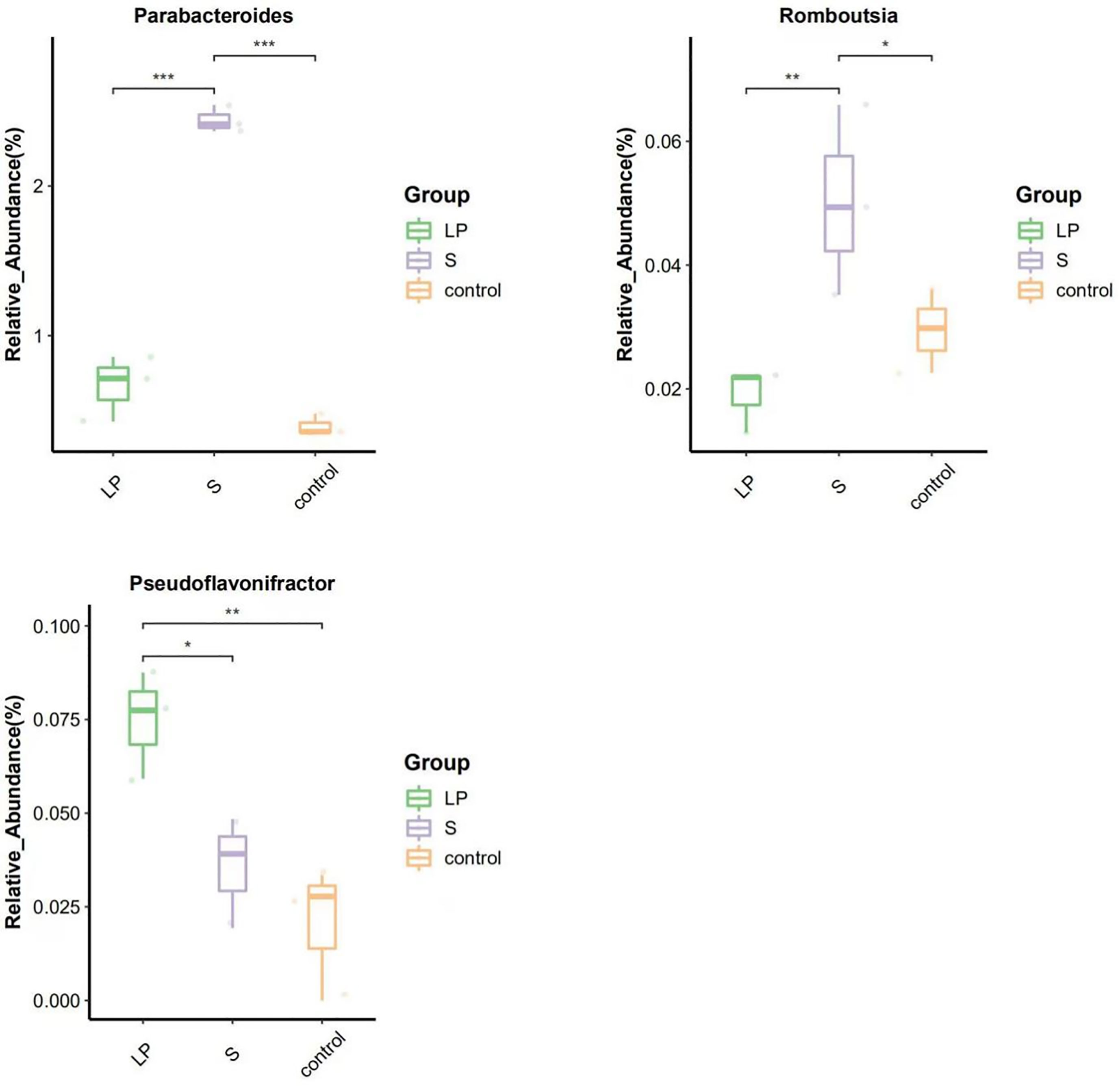 Figura 9 Diferencia en la abundancia de especies Top10 boxplot (42d) a nivel de género. (∗p<0,05; ∗∗p<0,01; ∗∗∗p<0,001).
Figura 9 Diferencia en la abundancia de especies Top10 boxplot (42d) a nivel de género. (∗p<0,05; ∗∗p<0,01; ∗∗∗p<0,001).
Discusión
El aumento de peso diario promedio, la ingesta de alimento y la relación alimento-peso son indicadores importantes del rendimiento del crecimiento del ganado. Se ha demostrado que la adición de Lactobacillus plantarum B1 a la dieta no tiene un efecto significativo en el rendimiento de los pollos de engorde en el período inicial, pero durante el período de desarrollo, el aumento de peso diario promedio de los pollos de engorde tratados con Lactobacillus plantarum aumenta significativamente y se mejora la tasa de conversión alimenticia (21). Yeo y Kim et al. (22) encontraron que el aumento de peso diario promedio de los pollos de engorde alimentados con probióticos aumentó significativamente al comienzo del período de crecimiento y no cambió significativamente en el período posterior. No hubo cambios significativos en las etapas posteriores. En el presente estudio, se encontró que la alimentación de Lactobacillus plantarum GX17 y preparaciones probióticas complejas comerciales no tuvo un efecto significativo en el rendimiento de crecimiento de pollos de engorde de 1 a 42 días de edad, lo que es consistente con los hallazgos de Al-Khalaifa (23) et al. Sin embargo, la relación alimento-peso de pollos de engorde suplementados con Lactobacillus plantarum GX17 y probióticos complejos comerciales fue significativamente menor a los 1-42 días de edad, por lo que se presume que el efecto beneficioso de este probiótico en pollos de engorde es el resultado de la alimentación a largo plazo, y se requiere alimentación a largo plazo para el uso posterior de la producción para lograr un mejor rendimiento de crecimiento. La alimentación de Lactobacillus plantarum GX17 también se observó para reducir la ingesta de alimento en este estudio, lo que es consistente con algunos estudios donde la suplementación con probióticos redujo la ingesta de alimento (24), y Chen (25) et al. encontraron de manera similar que la adición de probióticos a las dietas de pollos de engorde redujo la ingesta de alimento. Sin embargo, algunos científicos también han observado una mejor ingesta de alimento después de la suplementación de probióticos en la dieta, y los probióticos redujeron el tiempo de vaciado gástrico, lo que condujo a una mayor ingesta de alimento (26, 27), y en lugar de reducir la proporción de alimento a carne, se agregaron más costos de alimentación. En un análisis exhaustivo, la alimentación de Lactobacillus plantarum GX17 tiene una tendencia a mejorar el rendimiento de crecimiento de los pollos de engorde, pero su período de acción puede tener diferentes efectos dependiendo de la cepa, el contenido y la ruta de alimentación.
La morfología intestinal, incluida la altura de las vellosidades duodeno, yeyuno e íleon, la profundidad de la cripta y la relación entre la altura de las vellosidades y la profundidad de la cripta, puede reflejar la salud intestinal de los pollos de engorde. El aumento de la altura de las vellosidades del intestino delgado ayuda a aumentar el área de contacto entre el intestino delgado y los nutrientes y mejora la capacidad de digestión y absorción del organismo animal, mientras que la oscilación fuerte y regular de las vellosidades intestinales también puede prevenir la colonización de bacterias dañinas en el intestino y estabilizar el equilibrio de la microbiota (28). La función principal de la cripta es secretar jugos digestivos, y una disminución en su profundidad indica un aumento en la madurez celular y su capacidad para secretar jugos digestivos (29). El aumento en la altura de las vellosidades y la altura de las vellosidades en la relación entre la profundidad de la cripta se relacionó directamente con el aumento del recambio epitelial (30). Arifin et al. encontraron que la alimentación de preparaciones probióticas a pollos de engorde resultó en un aumento significativo en la longitud y anchura de las vellosidades intestinales pequeñas, junto con un aumento en el peso corporal y una disminución en la relación alimento / peso (31). Song et al. (32) observaron que la adición de una mezcla probiótica de Bacillus licheniformis, Bacillus subtilis y Lactobacillus plantarum a la dieta mejoró el acortamiento de las vellosidades intestinales inducido por el estrés por calor y la profundización de la profundidad de la cripta en los pollitos. En este experimento, se encontró que la alimentación de Lactobacillus plantarum GX17 y la mezcla probiótica comercial aumentaron significativamente la longitud de las vellosidades del intestino delgado y aumentaron la proporción de la altura de las vellosidades a la profundidad de la cripta, lo que es consistente con los resultados de muchos otros estudios. Esto indica que la adición de Lactobacillus plantarum GX17 a la dieta puede mejorar el rendimiento de crecimiento de los pollos de engorde al mejorar el resultado intestinal, mejorando así la absorción intestinal de nutrientes y reduciendo la proporción de alimento a carne.
La barrera mucosa intestinal, formada por el epitelio intestinal, es parte de la composición inmune natural del intestino, y sus células epiteliales intestinales son la primera línea de defensa contra patógenos extraños (33). La función de barrera de la superficie mucosa se encuentra principalmente en la membrana plasmática de las células epiteliales, que es resistente a muchas sustancias hidrófilas debido a su falta de proteínas de transporte específicas. La función de barrera se pierde significativamente en el contacto directo con antígenos de la mucosa o agentes citotóxicos, pero la vía de célula a célula está protegida y funciona debido a la presencia de monómeros epiteliales. Y una de las funciones está regulada por el complejo de unión apical, que es una colección de uniones estrechas y uniones adherentes adyacentes (AJ) (34). Las uniones estrechas intercelulares (TJ) exhiben una estructura molecular compleja que consiste en diferentes tipos de proteínas transmembrana unidas a uniones citoplasmáticas que contribuyen a la unión a las células vecinas, involucrando interacciones entre proteínas de unión citoplasmática (por ejemplo, cintas de ocludina) y proteínas de unión de membrana integral (por ejemplo, ocludinas y claudinas). Estas proteínas responden a diferentes señales fisiopatológicas a través de constantes modificaciones reguladoras (35).
Se ha encontrado que una variedad de probióticos mejora la función de barrera intestinal al aumentar las proteínas correspondientes a la unión estrecha en presencia o ausencia de infección intestinal, así como al aumentar la producción de mucina y proteínas de choque térmico y modular las vías de señalización (33, 36). Ciertos lactobacilos atenúan la interrupción de la barrera al regular al alza las proteínas TJ. Lactobacillus acidophilus y Lactobacillus plantarum aumentan la expresión de proteínas de cierre en modelos in vivo e in vitro, respectivamente (37, 38). Además de aumentar la expresión de la proteína ocludina, Lactobacillus plantarum también indujo el reposicionamiento apical de ZO-1 y ocludina mediante la estimulación de TLR2 (39, 40). Izuddin encontró que la adición de Lactobacillus plantarum RG14 a la dieta de corderos destetados regulaba al alza la expresión de TJP-1, CLDN-1 y CLDN-4 ARNm en el yeyuno de cordero y mejoraba la integridad de la barrera intestinal (41). Se ha demostrado que Lactobacillus plantarum G83 puede aumentar la expresión de ARNm de Claudina-1, Zo-1 y Ocludina-1 ileal al inducir alteraciones de la microbiota, mejorando así la barrera intestinal y defendiéndose contra E. coli K88 productora de enterotoxinas (42). Wang et al. (43) encontraron que Lactobacillus plantarum ZLP001 podía atenuar las proteínas TJ inducidas por endotoxinas (claudina-1, ocludina y ZO-1) y regulaba negativamente la expresión y secreción de las citoquinas proinflamatorias IL-6 e IL-8 y el factor de necrosis tumoral α. Zijian et al. encontraron que los niveles de expresión reducidos de ZO-1, claudina-1 y ocludina causados por una dieta alta en grasas y alta en fructosa se redujeron después de que el tratamiento con Lactobacillus plantarum NA136 volvió a los niveles normales (44). En el presente estudio, Lactobacillus plantarum GX17 mostró el mismo efecto; en comparación con los pollos de engorde sin adición de probióticos, Lactobacillus plantarum GX17 reguló al alza la expresión de ARNm de CLDN, ocludina y ZO-1 sin cambios significativos en la expresión de ARNm, mientras que la expresión de ARNm de CLDN, ocludina y MUC2 fue significativamente mayor en pollos de engorde suplementados con probióticos complejos comerciales (Lactobacillus). La diferencia en su expresión génica de barrera puede estar relacionada con las especies bacterianas y el efecto de los probióticos individuales versus complejos no es exactamente el mismo. MUC2 es la principal mucina producida por los cuprocitos y es un componente importante de la capa de moco que cubre el epitelio intestinal. Se ha demostrado que la suplementación al final de la dieta con probióticos aumentó la expresión del gen MUC2 en pollos de engorde (45, 46). También se ha encontrado que la regulación a la baja de la expresión de MUC2 inducida por endotoxinas fue antagonizada por la adición de Bacillus subtilis (47). En el presente estudio, se encontró que la adición de Lactobacillus plantarum GX17 a la dieta aumentó significativamente la expresión del gen MUC2 en el yeyuno de pollos de engorde.
Los TLR son un receptor de proteína transmembrana identificado por Lemaitre en su estudio sobre el desarrollo embrionario de Drosophila, que fue demostrado por estudios posteriores para tener una función de señalización celular y un papel clave en el proceso de infección y respuesta inmune. Xueer Gui et al. (48) encontraron que la expresión relativa de los ARNm TLR2, TLR4, TRAF-6, Myd88 y AP-1 en el intestino de los pollos de engorde aumentó significativamente mediante la adición de probióticos complejos a la dieta. En este experimento, se encontró que la adición de probióticos complejos a la dieta aumentó significativamente la expresión relativa de ARNm TLR2 y TLR4, y la adición de Lactobacillus plantarum GX17 aumentó significativamente la expresión de ARNm TLR2. Los efectos beneficiosos de los probióticos sobre la función de barrera intestinal están mediados, al menos en parte, por la estimulación tisular de TLRs, particularmente TLR2, y por cambios secuenciales en la expresión y localización de proteínas de unión estrecha. Hay nuevos estudios que muestran que los TLR interactúan con la microbiota intestinal y las células epiteliales en uniones estrechas. Dada su interacción directa con las uniones estrechas intestinales, los TLR desempeñan un papel diferente y crucial en la regulación de las uniones estrechas intestinales como un posible objetivo terapéutico probiótico (49). Un estudio encontró que Caco-2 mostró una fuerte redistribución apical de ZO-1 y un aumento de la actividad intracelular de la proteína quinasa C (PKC) después del tratamiento con ligandos TLR-2 (50). Esto puede sugerir que la estimulación TLR2 promueve modificaciones estructurales en uniones estrechas al promover la actividad PKC. Por lo tanto, los niveles insuficientes de ligandos TLR2 protectores del huésped pueden afectar la función de barrera. Por ejemplo, la administración de fármacos antimicrobianos puede afectar la función de barrera al reducir la población luminal de comensales intestinales grampositivos como Lactobacillus spp. y Bifidobacterium spp., ambas fuentes importantes de ligandos estimulantes de TLR2 (51). Por lo tanto, la homeostasis intestinal y la función de barrera requieren un delicado equilibrio entre la simbiosis y la activación de TLR inducida por patógenos. Esta posibilidad también existe en el presente estudio, donde Lactobacillus plantarum GX17 y las bacterias probióticas complejas mejoraron la función de barrera intestinal al aumentar la expresión de TLR2 seguida de la inducción de la expresión de proteínas de unión estrecha para mejorar la integridad de la barrera intestinal, pero el mecanismo exacto de acción aún no se ha descubierto en estudios adicionales.
Las bacterias intestinales juegan un papel importante en el mantenimiento de la salud del organismo, que incluye la síntesis de vitamina B, la mejora de las funciones digestivas y neurológicas, y la promoción de la angiogénesis (52). Debido a que los pollitos recién nacidos carecen de contacto con aves adultas, las colonias microbianas ambientales pueden ser críticas para el desarrollo inmunológico de los pollitos de la planta de incubación, que potencialmente se extiende a lo largo de toda su vida útil y puede influir en el desarrollo de la microbiota gastrointestinal (53). En este experimento, los probióticos se alimentaron a pollos de engorde de plumas amarillas justo después de la eclosión para estudiar el efecto de los probióticos en la flora gastrointestinal de los pollitos. La comunidad microbiana en el tracto gastrointestinal de los pollos de engorde tiene más de 900 especies de bacterias (54). Desempeñan un papel crucial en la digestión, la degeneración de toxinas, la eliminación de patógenos, la estimulación del sistema inmunológico y la actividad endocrina (55). Firmicutes, Bacteroides y Proteobacteria son los tres filos más comunes en el ciego del pollo, lo que se confirmó en el presente estudio ya que la estructura de la flora intestinal fue similar entre todos los grupos experimentales y la flora dominante principal también fueron los tres filos más comunes mencionados anteriormente. El Parabacteroides es también el patógeno de la enfermedad inflamatoria intestinal en los seres humanos, y también se correlaciona con el metabolismo de los carbohidratos, la obesidad y la secreción de ácidos grasos de cadena corta (56). Se ha encontrado que los ácidos grasos de cadena corta son importantes para mantener las funciones intestinales saludables, y el ácido butírico es la principal fuente de energía para las células epiteliales del colon (57). Además, el AGCC podría suprimir la expresión de factores de virulencia de patógenos bacterianos (58). También se ha encontrado que Parabacteroides puede producir acetato, que atenúa la pancreatitis aguda en la familia acetil heparinasa al reducir la infiltración de neutrófilos (59). La Romboutsia se encuentra generalmente en el intestino de los animales y se ha asociado con su estado de salud. La disminución dramática en su abundancia en la mucosa intestinal se considera como un indicador microbiano que representa ciertas enfermedades. En animales sanos, la abundancia relativa de Romboutsia es significativamente mayor que en el tejido del cáncer colorrectal. Por lo tanto, la ausencia de bacterias de este género es potencialmente un marcador de patología mucosa (60, 61). Además, Pseudoflavonifractor es una bacteria colonizadora de ciego común con proteínas derivadas de alcohol deshidrogenasas de clase IV que influyen en la vía final de producción de butirato (62). En este estudio, la abundancia relativa de Parabacteroides y Romboutsia fue significativamente mayor en el intestino de pollos de engorde de plumas amarillas alimentados con probióticos complejos comerciales, y la abundancia relativa de Pseudoflavonifractor fue significativamente mayor en el intestino de pollos de engorde de plumas amarillas alimentados con Lactobacillus plantarum GX17. Estos datos sugieren que los probióticos complejos comerciales y Lactobacillus plantarum GX17 son capaces de mantener la salud del ciego de los pollos de engorde de plumas amarillas al promover la proliferación de bacterias específicas.
Se ha informado que el peso del timo, el bazo y la bolsa reflejan el estado inmunológico de los pollitos, y el aumento de la masa absoluta y relativa de estos órganos indica la mejora de la inmunidad celular y humoral (63). Aunque la aplicación de potenciadores inmunes puede mejorar las funciones inmunes de las aves de corral al tiempo que mejora su resistencia a las enfermedades infecciosas, la mayoría de los potenciadores inmunes se sintetizan químicamente con efectos secundarios significativos, problemas de residuos de medicamentos y otros riesgos de seguridad alimentaria. Estudios previos han demostrado que alimentar a los pollos de engorde con probióticos podría aumentar significativamente su índice de órganos inmunes, lo que indica una mejora de su función inmune (64). Aunque algunos otros estudios sostuvieron el resultado opuesto de que el crecimiento del bazo de pollos de engorde se inhibió después de 21 días de alimentación con compuestos probióticos comerciales (65). La alimentación de Lactobacillus plantarum GX17 no estimuló ni inhibió el crecimiento de órganos inmunes, lo que indica que los pollos de engorde tienen una fuerte capacidad de adaptación a la colonización de la cepa GX17. Dado que la cepa GX17 no produce ningún efecto inmune adverso en pollos de engorde, podría ser un aditivo alimentario potencial apto para la aplicación diaria en pollitos jóvenes. Según nuestros datos, la suplementación con probióticos podría mejorar la función de barrera intestinal y mejorar la inmunidad intestinal, lo que conduce al aumento del nivel de anticuerpos al mejorar el tracto intestinal, y el aumento de microorganismos beneficiosos también puede ayudar a mejorar el efecto después de la vacunación.
El nivel de anticuerpos inmunes es un indicador común de la función inmune, entre los cuales el nivel de anticuerpos contra la enfermedad de Newcastle y la influenza aviar pueden reflejar mejor la respuesta inmune humoral específica de los pollos. Producidas por linfocitos B derivados de la médula ósea, lgG, IgM y lgA son inmunoglobulinas con efectos antibacterianos, antivirales y antitoxínicos (66), que son las principales moléculas efectoras de la inmunidad humoral y son esenciales en la inmunidad humoral. La capacidad inmune del cuerpo puede reflejarse por el nivel de inmunoglobulinas en el suero, que también es un indicador importante para evaluar los cambios en la función inmune de los animales. Se ha demostrado que los probióticos no solo promueven el desarrollo de los órganos inmunes del organismo, sino que también tienen un efecto modulador sobre las vacunas y mejoran la función inmune humoral específica del organismo (67). Talebi et al. (68) informaron que la administración de probióticos junto con la vacunación produjo una mejor respuesta de anticuerpos. En el presente experimento, la alimentación con Lactobacillus plantarum GX17 aumentó significativamente la potencia de anticuerpos de la enfermedad de Newcastle en el día 7 después de la inmunización y los resultados del estudio son consistentes con los datos de Talazadeh et al. (69) que muestran un aumento significativo en los títulos de anticuerpos contra la vacuna contra la enfermedad de Newcastle después de alimentar a los probióticos, pero ningún cambio significativo en los títulos de anticuerpos contra la vacuna contra el virus de la influenza aviar. Esto puede deberse al hecho de que la alimentación con probióticos podría mejorar el nivel de anticuerpos del organismo contra algunos antígenos específicos. Mientras tanto, Isolauri et al. (70) encontraron que la alimentación de Lactobacillus casei durante la vacunación contra el rotavirus aumenta la cantidad de células secretoras de anticuerpos IgM e IgA del organismo, lo que aumenta la producción de anticuerpos. En el presente estudio, los pollos de engorde de plumas amarillas alimentados con Lactobacillus plantarum GX17 tenían niveles significativamente más altos de moléculas CD4 en el suero a los días 28 y 42 después de la segunda vacunación en comparación con el grupo control (solo vacunados). Se ha informado que los probióticos desempeñan un papel clave en la estimulación del desarrollo del sistema inmune durante su entrada inicial y colonización del intestino, con una amplia gama de funciones de mejora inmunológica, como la activación de las células inmunes, el aumento en el número y la promoción en la maduración de las células T / B, la mejora del reconocimiento inmunológico, la inducción de la expresión de factores inmunes relevantes, y activando todo el sistema inmunológico del huésped (71, 72). Se ha demostrado que Lactobacillus mejora la respuesta de inmunidad humoral de los pollos a la vacuna viva contra la enfermedad de Newcastle, particularmente durante el período de choque térmico (73). En un ensayo con conejos destetados, se observó que la alimentación de Lactobacillus licheniformis mejoró significativamente las respuestas de inmunidad humoral y la vacunación después de la alimentación con probióticos y fue superior a la vacunación sola (74). En el presente experimento se demostró que Lactobacillus plantarum GX17 utilizado sinérgicamente con la vacuna mostró una mejor mejora de la inmunidad celular y humoral que la vacuna sola. Se demostró fuertemente que la alimentación con Lactobacillus plantarum GX17 podría mejorar la función inmune humoral de pollos de engorde y mejorar el efecto inmune de la enfermedad de Newcastle y las vacunas contra la influenza aviar al mismo tiempo.
Declaración de disponibilidad de datos
Las contribuciones originales presentadas en el estudio están disponibles públicamente. Estos datos se pueden encontrar en el repositorio del NCBI, número de acceso: PRJNA950182.
Declaración ética
El estudio en animales fue revisado y aprobado por el Formulario Institucional de Inspección Ética Experimental con Animales del Instituto de Investigación Veterinaria de Guangxi, Nanning, China.
Contribuciones del autor
YYY: Conceptualización, metodología, investigación, redacción: preparación del borrador original y adquisición de fondos. YYL: metodología, software, análisis formal, curación de datos y redacción: preparación del borrador original. JL: metodología, software, análisis formal, curación de datos y redacción: preparación del borrador original. ZP: análisis formal, redacción: revisión y edición, e investigación. LPW: metodología, redacción: preparación del borrador original y recursos. YS: metodología, investigación, redacción, revisión y edición. HYP: metodología e investigación. YZT: metodología e investigación. CTL: software. HLB: software. CXM: redacción, revisión y edición. YG: escritura, revisión y edición. TCW: Conceptualización, análisis formal, recursos, supervisión y administración de proyectos. HP: Conceptualización, análisis formal, recursos, supervisión, administración de proyectos y adquisición de fondos. Todos los autores contribuyeron al artículo y aprobaron la versión presentada. Todos los autores han leído y aceptado la versión publicada del manuscrito.
Financiación
La investigación fue financiada por el Programa de Investigación y Desarrollo de Guangxi (AB21238003; AB21220005-4); Proyecto Especial para la Construcción del Sistema Técnico de la Industria Nacional de Aves Acuáticas (CARS-42-55); Programa clave de investigación y desarrollo del Distrito de Nanning (20212023); Programa clave de investigación y desarrollo del distrito de Liangqing( 202118); Proyecto de Construcción del Equipo de Innovación de Guangxi del Sistema Nacional de Tecnología de la Industria Agrícola Moderna (nycytxgxcxtd); Programa clave de investigación y desarrollo de Fangchengang (AB21014016); Programa clave de investigación y desarrollo de Jiangnan (20220620-2); y el Programa de Investigación y Desarrollo (20210102) del Distrito de Wuming. Deseamos reconocer el apoyo técnico del proyecto de mejoramiento de animales y plantas agrícolas de Guizhou (NY [2018]016).
Conflicto de intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Nota del editor
Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo, o reclamo que pueda ser hecho por su fabricante, no está garantizado ni respaldado por el editor.
Referencias
1. Guo M, Li M, Zhang C, Zhang X, Wu Y. La administración dietética del bacillus subtilis mejora las respuestas inmunes y la resistencia a enfermedades en pollos. Microbiol frontal (2020) 11. DOI: 10.3389/fmicb.2020.01768
2. Pyw A, Cxm A, Prz C, Pyc B, Ys D, Pfhe F, et al. Cambios en la resistencia a la colistina y la abundancia de mcr-1 en escherichia coli de origen animal y humano tras la prohibición de aditivos positivos para colistina en China: un estudio comparativo epidemiológico. Lancet Infect Dis (2020) 20 doi: 10.1016/S1473-3099(20)30149-3
3. Khalique A, Zeng D, Shoaib M, Wang H, Ni X. Probióticos que mitigan la enteritis necrótica subclínica (SNE) como alternativas potenciales a los antibióticos en aves de corral. Amb Express (2020) 10(1):50. DOI: 10.1186/S13568-020-00989-6
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Ray SM, Ghule S, Muthukumar S, Banik A, Maji C. Efectos de la suplementación dietética de un probiótico de una sola cepa y una cepa múltiple sobre el rendimiento del crecimiento y la histomorfología intestinal de pollos de engorde comerciales. Int J Poultry Sci (2020) 19(8):363–71. doi: 10.3923/ijps.2020.363.371
5. Aziz HN, Ahmed HZO, Hassan HA, Mustafa AN. (2019). Effects of the probiotic miaclost (Bacillus subtilis and entrococus feacium) on growth performance, hematological values and small intestinal morphology of broiler chicks, en: IOP Conference Series: Earth and Environmental Science, (Published under licence by IOP Publishing Ltd), vol. 388.
6. Eslam A, Mootaz A, Khaled G. Efecto del probiótico en el rendimiento del crecimiento, rasgos de la canal y parámetros de salud clínica de pollos de engorde criados bajo estrés por calor en el alto Egipto. SVU-International J Veterinary Sci (2019) 2(2):27–44. doi: 10.21608/svu.2019.11221.1012
7. Redweik GAJ, Stromberg ZR, Van Goor A, Mellata M. Bacterias gramnegativas – escherichia coli; estudios de la Universidad Estatal de Iowa en el área de Escherichia coli descritos (Protección contra escherichia coli patógena aviar y salmonella Kentucky exhibida en pollos que recibieron probióticos y vacuna viva contra la salmonela). En: Food weekly news. Estados Unidos: Oxford University Press.
8. Asahara T. Efecto preventivo de las bifidobacterias probióticas contra las infecciones por escherichia coli y salmonella productoras de toxina shiga. Biociencia Microflora (2010) 2010:29. doi: 10.12938/bifidus.29.11
9. Jinfeng H, Yongwei W, Dan S, Zixian L, Zhenglin D, Haijiang M, et al. Efectos de Clostridium butyricum y Lactobacillus plantarum sobre el rendimiento del crecimiento, la función inmune y el nivel de ácidos grasos volátiles de la digesta cecal en pollos de engorde. Food Agric Immunol (2018) 29(1):797–807. doi: 10.1080/09540105.2018.1457013
10. Peng Q, Zeng XF, Zhu JL, Wang S, Liu XT, Hou CL, et al. Bacterias grampositivas; nuevos hallazgos sobre lactobacillus plantarum de la Universidad de Saskatchewan resumidos (Efectos de lactobacillus plantarum B1 dietético sobre el rendimiento de crecimiento, microbiota intestinal y perfiles de ácidos grasos de cadena corta en pollos de engorde). Agric Week (2016) 95(4):893–900. doi:10.3382/ps/pev435
11. Incharoen T, Charoensook R, Onoda S, Tatrakoon W, Numthuam S, Pechkong T. Los efectos de la suplementación con lactobacillus plantarum l-137 matada por calor sobre el rendimiento del crecimiento, la morfología intestinal y la expresión génica relacionada con el sistema inmunitario en pollos de engorde. Anim Feed Sci Technol (2019) 2019:257. doi: 10.1016/j.anifeedsci.2019.114272
12. Baikui W, Yuanhao Z, Yulong M, Li G, Xiang L, Shujie X, et al. La suplementación dietética con lactobacillus plantarum mejora el compromiso del rendimiento del crecimiento mediante la modulación de los ácidos grasos de cadena corta y la disbiosis intestinal en pollos de engorde bajo clostridium perfringens Challenge&13. Frente Nutr (2021) 8. doi: 10.3389/fnut.2021.706148
13. Yang Y, Park JH, Kim IH. Efecto del probiótico que contiene lactobacillus plantarum sobre el rendimiento de crecimiento, la digestibilidad de nutrientes y la microbiota fecal en cerdos destetados. Can Sci Publishing65 Auriga Drive Suite 203 Ottawa ON K2E 7W6 (2020) 11(8):2232. doi: 10.3390/ani11082232
14. Mohanty D, Panda S, Kumar S, Ray P. Evaluación in vitro de la adherencia y la propiedad antiinfecciosa del probiótico lactobacillus plantarum DM 69 contra la salmonela enterica. Microb Pathog (2018) 126: 212–7. doi: 10.1016/j.micpath.2018.11.014
Resumen de PubMed | Texto completo de CrossRef | Google Académico
15. Chen Q, Chao T, Ma S, Zhou L, Zhao L, Xin Z. Involvement of MicroRNAs in probiotics-induced reduction of the cecal inflammation by salmonella typhimurium. Frente Immunol (2017) 2017:8. doi: 10.3389/fimmu.2017.00704
16. Gill HS, Rutherfurd KJ, Cross ML. La suplementación con probióticos dietéticos mejora la actividad de las células asesinas naturales en los ancianos: una investigación de los cambios inmunológicos relacionados con la edad. J Clin Immunol (2001) 21:264–71. doi: 10.1023/A:1010979225018
Resumen de PubMed | Texto completo de CrossRef | Google Académico
17. Mater DDG, Langella P, Corthier GE, Rard, Flores MI, Eacute A. Una cepa de lactobacilos probióticos puede adquirir resistencia a la vancomicina durante el tránsito digestivo en ratones. J Mol Microbiol Biotechnol (2008) 14:123–27. DOI: 10.1159/000106091
Resumen de PubMed | Texto completo de CrossRef | Google Académico
18. Boyle RJ, Robins-Browne RM, Tang ML. Uso de probióticos en la práctica clínica: ¿cuáles son los riesgos? Am J Clin Nutr (2006) 83:1256–64. doi: 10.1093/ajcn/83.6.1256
Resumen de PubMed | Texto completo de CrossRef | Google Académico
19. Wang Y, Wang Y, Wang B, Mei X, Jiang S, Li W. El ácido protocatecúico mejoró el rendimiento del crecimiento, la calidad de la carne y la salud intestinal de los pollos de engorde chinos de plumas amarillas. Poult Sci (2019) 98:3138–49. DOI: 10.3382/ps/pez124
Resumen de PubMed | Texto completo de CrossRef | Google Académico
20. Zhu C, Huang K, Bai Y, Feng X, Gong L, Wei C, et al. La suplementación dietética con berberina mejora el rendimiento del crecimiento y modula la composición y función de la microbiota cecal en pollos de engorde de plumas amarillas. Poult Sci (2021) 100:1034–48. doi: 10.1016/j.psj.2020.10.071
Resumen de PubMed | Texto completo de CrossRef | Google Académico
21. Peng Q, Zeng XF, Zhu JL, Wang S, Qiao SY. Efectos de lactobacillus plantarum B1 dietético sobre el rendimiento de crecimiento, la microbiota intestinal y los perfiles de ácidos grasos de cadena corta en pollos de engorde. Poultry Sci (2016) 95:893. DOI: 10.3382/ps/pev435
22. Yeo J, Kim K. Efecto de las dietas de alimentación que contienen un antibiótico, un probiótico o extracto de yuca sobre el crecimiento y la actividad de la ureasa intestinal en pollos de engorde. Poultry Sci (1997) 76:381. doi: 10.1093/ps/76.2.381
23. Al-Khalaifa H, Al-Nasser A, Al-Surayee T, Al-Kandari S, Mohammed A. Efecto de los probióticos y prebióticos dietéticos en el rendimiento de los pollos de engorde. Ciencias avícolas (2020). doi: 10.1016/j.psj.2020.12.054
24. Falaki M, Shargh MS, Dastar B, Zrehdaran S. Efectos de diferentes niveles de probióticos y prebióticos sobre el rendimiento y las características de la canal de pollos de engorde. J Anim Veterinary Adv (2011) 10:2390–95. doi: 10.3923/javaa.2011.378.384
25. Olnood CG, Beski SSM, Choct M, Iji PA. Nuevos probióticos: sus efectos sobre el rendimiento del crecimiento, el desarrollo intestinal, la comunidad microbiana y la actividad de los pollos de engorde. Anim Nutr (2015) 1:184–91. doi: 10.1016/j.aninu.2015.07.003
Resumen de PubMed | Texto completo de CrossRef | Google Académico
26. Altaf-ur-Rahman, Khan S, Khan D, Hussain M, Ahmed S, Sohail SM, et al. Uso de probióticos en la alimentación de pollos de engorde en la fase inicial. Sarhad J Agric (2009) 25(3):469–73.
27. Abdel-Raheem SM, Abd-Allah SMS. El efecto de la suplementación dietética única o combinada de manano oligosacárido y probióticos sobre el rendimiento y las características de sacrificio de pollos de engorde. Int J Poultry Sci (2011) 10:854–62. doi: 10.3923/ijps.2011.854.862
28. Caspary WF. Fisiología y fisiopatología de la absorción intestinal. Am J Clin Nutr (1992) 55(1):299S–308S. doi: 10.1093/ajcn/55.1.299s
Resumen de PubMed | Texto completo de CrossRef | Google Académico
29. Zhang LR. Efectos de tres probióticos sobre el crecimiento, desarrollo del intestino delgado y microorganismo ciego de pollitos [maestro]. Jiangsu, China: Universidad de Yangzhou (2018).
30. Fan YK, Croom J, Christensen VL, Black BL, Eisen EJ. Captación de glucosa yeyunal y consumo de oxígeno en pollos de pavo seleccionados para un rápido crecimiento. Poult (1997) 76:1738–45. doi: 10.1093/ps/76.12.1738
31. Arifin M, Pramono VJ. El efecto del suplemento sinbiótico como reemplazo del antibiótico promotor del crecimiento sobre el crecimiento y el tamaño de las vellosidades intestinales en pollos de engorde. Jurnal Sain Veteriner (2015) 32(2). doi: 10.22146/JSV.6555
32. Song J, Xiao K, Ke LY, Jiao FL, Hu HC, Diao YQ, et al. Efecto de una mezcla probiótica sobre la microflora intestinal, la morfología y la integridad de la barrera de los pollos de engorde sometidos a estrés por calor. Poultry Sci (2014) 93(3):581–8. DOI: 10.3382/PS.2013-03455
33. Thomas CM, Versalovic J. Comunicación probiótico-huésped: modulación de las vías de señalización en el intestino. Gut Microbes (2010) 1:148. doi: 10.4161/gmic.1.3.11712
Resumen de PubMed | Texto completo de CrossRef | Google Académico
34. Shruti P, Sapna S, Prabhanshu T. Papel de la integridad de la barrera y las disfunciones en el mantenimiento del intestino sano y sus resultados de salud &13. Frente Physiol (2021) 12. DOI: 10.3389/FPHYS.2021.715611
35. Philip M. Cummins. Ocludina: Una proteína, muchas formas. Mol Cell Biol (2012) 32(2):242–50. doi: 1128/MCB.06029-11
36. Otte JM, Podolsky DK. Modulación funcional de enterocitos por microorganismos grampositivos y gramnegativos. Am J Physiol Gastrointest Liver Physiol (2004) 286:G613. doi: 10.1152/ajpgi.00341.2003
Resumen de PubMed | Texto completo de CrossRef | Google Académico
37. Qin HL, Shen TY, Gao ZG, Fan XB, Hang XM, Jiang YQ, et al. Efecto de lactobacillus sobre la microflora intestinal y la función de barrera de las ratas con infección abdominal. World J Gastroenterol (2005) 11(17):2591–6. doi: 10.3748/wjg.v11.i17.2591
Resumen de PubMed | Texto completo de CrossRef | Google Académico
38. Anderson RC, Cookson AL, Mcnabb WC, Park Z, Mccann MJ, Kelly WJ, et al. Lactobacillus plantarum MB452 mejora la función de la barrera intestinal al aumentar los niveles de expresión de genes involucrados en la formación de la unión estrecha. BMC Microbiol (2010) 2010:10. doi: 10.1186/1471-2180-10-316
39. Blackwood BP, Yuan CY, Wood DR, Nicolas JD, Grothaus JS, Hunter CJ. Las especies de lactobacilos probióticos fortalecen la función de barrera intestinal y la integridad de la unión estrecha en la enterocolitis necrotizante experimental. J Probiotics Health (2017) 5 (1): 159. doi: 10.4172/2329-8901.1000159
Resumen de PubMed | Texto completo de CrossRef | Google Académico
40. Karczewski J, Troost FJ, Konings I, Dekker J, Wells JM. Regulación de las proteínas epiteliales de unión estrecha humana por lactobacillus plantarum in vivo y efectos protectores sobre la barrera epitelial. Am J Physiol Gastrointestinal Liver Physiol (2010) 298: 851-59. doi: 10.1152/ajpgi.00327.2009
41. Izuddin WI, Loh TC, Foo HL, Samsudin AA, Humam AM. Postbiotic l. plantarum RG14 mejora el crecimiento del epitelio ruminal, el estado inmunológico y regula al alza la función de barrera intestinal en corderos post-destete. Sci Rep (2019) 9(1):9938. DOI: 10.1038/S41598-019-46076-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
42. Yi Z, Xueqin N, Ling D, Lili N, Qian L, Yan Z, et al. Lactobacillus plantarum BSGP201683 mejora la barrera intestinal del ratón gigante asociado a la microbiota panda infectado por escherichia coli K88 enterotoxigénica. Probióticos Proteínas antimicrobianas (2020) 13(3):664–76. DOI: 10.1007/S12602-020-09722-Y
Resumen de PubMed | Texto completo de CrossRef | Google Académico
43. Jing W, Haifeng J, Sixin W, Hui L, Wei Z, Dongyan Z, et al. Probiótico lactobacillus plantarum promueve la función de barrera intestinal mediante el fortalecimiento del epitelio y la modulación de la microbiota intestinal. Microbiol frontal (2018) 2018:9. DOI: 10.3389/fmicb.2018.01953
44. Zijian Z, Long C, Yujuan Z, Chao W, Cuicui D, Ge Y, et al. Lactobacillus plantarum NA136 mejora la enfermedad del hígado graso no alcohólico mediante la modulación de la microbiota intestinal, la mejora de la integridad de la barrera intestinal y la atenuación de la inflamación. Appl Microbiol Biotechnol (2020) 104(12):5273–82. DOI: 10.1007/S00253-020-10633-9
Resumen de PubMed | Texto completo de CrossRef | Google Académico
45. Aliakbarpour HR, Chamani M, Rahimi G, Sadeghi AA, Qujeq D. El bacillus subtilis y los probióticos de las bacterias del ácido láctico influyen en la expresión génica de la mucina intestinal, la histomorfología y el rendimiento del crecimiento en pollos de engorde. Asian-Australas J Anim Sci (2012) 25:1285–93. doi: 10.5713/ajas.2012.12110
Resumen de PubMed | Texto completo de CrossRef | Google Académico
46. Smirnov A, Pérez R, Amit-Romach E, Sklan D, Uni Z. La dinámica de la mucina y las poblaciones microbianas en el intestino delgado del pollo se modifican por la suplementación con probióticos dietéticos y antibióticos promotores del crecimiento. J Nutr (2005) 135:187–92. doi: 10.1093/jn/135.2.187
Resumen de PubMed | Texto completo de CrossRef | Google Académico
47. Gadde UD, Oh S, Lee Y, Davis E, Zimmerman N, Rehberger T, et al. Los microbios alimentados directamente a base de bacilo subtilis dietético alivian el estrés inmunológico intestinal inducido por LPS y mejoran la expresión génica de barrera intestinal en pollos de engorde comerciales. Res Vet Sci (2017) 114: 236-43. doi: 10.1016/j.rvsc.2017.05.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
48. Gui XE, Wang Z, Li ST, He MC, Zhu J, Feng SB, et al. Efecto de los probióticos compuestos derivados de pollo sobre la inmunoglobulina y la vía del receptor tipo toll de pollos de engorde de plumas blancas. Acta Agriculturae Zhejiangensis (2020) 32:1609–14. doi: 10.3969/j.issn.1004-1524.2020.09.09
49. Ziegler AL. Probióticos, prebióticos y uniones estrechas epiteliales: un enfoque prometedor para modular la función de barrera intestinal. Int J Mol Sci (6729) 22(13):6729. DOI: 10.3390/IJMS22136729
50. Cario E, Gerken G, Podolsky DK. Toll-like receptor 2 mejora la integridad de la barrera epitelial intestinal asociada a ZO-1 a través de la proteína quinasa c. Gastroenterology (2004) 127:224–38. doi: 10.1053/j.gastro.2004.04.015
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Nord CE, Sillerstrom E, Wahlund E. Efecto de la tigeciclina en la microflora orofaríngea e intestinal normal. Antimicrobial Agents Chemotherapy (2006) 50:3375. doi: 10.1128/AAC.00373-06
Resumen de PubMed | Texto completo de CrossRef | Google Académico
52. Zhang YJ, Li S, Gan RY, Zhou T, Xu DP, Li HB. Impactos de las bacterias intestinales en la salud humana y las enfermedades. Int J Mol Sci (2015) 16(4):7493–519. DOI: 10.3390/IJMS16047493
Resumen de PubMed | Texto completo de CrossRef | Google Académico
53. Oakley BB, Lillehoj HS, Kogut MH, Kim WK, Maurer JJ, Pedroso A, et al. El microbioma gastrointestinal del pollo. FEMS Microbiol Lett (2014) 360:100–12. doi: 10.1111/1574-6968.12608
Resumen de PubMed | Texto completo de CrossRef | Google Académico
54. Stanley D, Hughes RJ, Moore RJ. Microbiota del tracto gastrointestinal del pollo: influencia en la salud, la productividad y la enfermedad. Appl Microbiol Biotechnol (2014) 98(10):4301–10. DOI: 10.1007/S00253-014-5646-2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
55. Zoetendal EG, von Wright A, Vilpponen-Salmela T, Ben-Amor K, Akkermans AD, de Vos WM. Las bacterias asociadas a la mucosa en el tracto gastrointestinal humano se distribuyen uniformemente a lo largo del colon y difieren de la comunidad recuperada de las heces. Appl Environ Microbiol (2002) 68:3401–07. doi: 10.1128/AEM.68.7.3401-3407.2002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
56. Cui Y, Zhang L, Wang X, Yi Y, Shan Y, Liu B, et al. Funciones de los parabacteroides intestinales en la salud humana y las enfermedades. FEMS Microbiol Lett (2022) 369(1):fnac072. doi: 10.1093/femsle/fnac072
Resumen de PubMed | Texto completo de CrossRef | Google Académico
57. Fleming SE, Fitch MD, DeVries S, Liu ML, Kight C. Utilización de nutrientes por células aisladas de yeyuno de rata, ciego y colon. J Nutr (1991) 121(6):869–78. doi: 10.1093/jn/121.6.869
Resumen de PubMed | Texto completo de CrossRef | Google Académico
58. Boyen F, Haesebrouck F, Vanparys A, Volf J, Mahu M, Immerseel FV, et al. Los ácidos grasos recubiertos alteran las propiedades de virulencia de la salmonella typhimurium y disminuyen la colonización intestinal de los cerdos. Vet Microbiol (2008) 132:319–27. doi: 10.1016/j.vetmic.2008.05.008
Resumen de PubMed | Texto completo de CrossRef | Google Académico
59. Lei Y, Tang L, Liu S, Hu S, Wu L, Liu Y, et al. Parabacteroides produce acetato para aliviar la pancreatitis aguda exacerbada por heparanasa mediante la reducción de la infiltración de neutrófilos. Microbioma (2021) 9:115. DOI: 10.1186/S40168-021-01065-2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
60. Mangifesta M, Mancabelli L, Milani C, Gaiani F, De Angelis N, De Angelis GL, et al. La microbiota mucosa de los pólipos intestinales revela supuestos biomarcadores de cáncer colorrectal. entific Rep (2018) 8(1):13974. DOI: 10.1038/S41598-018-32413-2
61. Ricaboni D, Mailhe M, Khelaifia S, Raoult D, Million M. Romboutsia timonensis, una nueva especie aislada del intestino humano. Nuevas infecciones por microbios (2016) 12:6–07. doi: 10.1016/j.nmni.2016.04.001
Resumen de PubMed | Texto completo de CrossRef | Google Académico
62. Polansky O, Sekelova Z, Faldynova M, Sebkova A, Sisak F, Rychlik I. Importantes vías metabólicas y procesos biológicos expresados por la microbiota cecal del pollo. Appl Environ Microbiol (2015) 82:1569. doi: 10.1128/AEM.03473-15
Resumen de PubMed | Texto completo de CrossRef | Google Académico
63. Rivas LA, Fabricant J. Indicaciones de inmunodepresión en pollos infectados con varias cepas del virus de la enfermedad de Marek. Avian Dis (1988) 32(1):1–8. DOI: 10.2307/1590941
Resumen de PubMed | Texto completo de CrossRef | Google Académico
64. Wu F, Xi YM, Tang B, Wang Q, Yang Z, Yang BK, et al. Efectos de la dieta de probióticos mixtos y ácidos orgánicos con diferentes niveles de proteína cruda sobre la función inmune y la actividad antioxidante en pollos de engorde de pies verdes. Anim Husbandry Veterinary Med (2016) 6(01):10–5. doi: CNKI:SUN:XMYS.0.2016-01-003
65. Ye SL, Li ZY, Li HM, Zhang XY, Fan SR, Li XP, et al. Efectos de los probióticos compuestos en los órganos digestivos e inmunes e indicadores séricos de pollos de engorde de plumas blancas. Anim Husbandry Veterinary Med (2021) 53(02):43–9.
66. Hao YR, Jie HL, Chen Q, Niu LQ, Hua CF, Geng YL, et al. Los probióticos mejoran el rendimiento de crecimiento de los pollos de engorde y alivian su inflamación hepática causada por LPS. Anim Husbandry Veterinary Med (2019) 2019: 6. doi: CNKI: SUN: XMYS.0.2019-04-011
67. Ning YC, Qiao HX, Zheng M, Ma H. Efecto del uso combinado de probióticos y gynostemma pentaphyllum en el rendimiento de pollos de engorde. Chin J Anim Sci (2012) 4(21):61–4. doi: 10.3969/j.issn.0258-7033.2012.21.016
68. Talazadeh F, Mayahi M, Zeinali S. El efecto del probiótico aviar Aquablend incluyendo lactobacillus, streptococcus y bifidobacterium en la respuesta sistémica de anticuerpos contra Newcastle y la vacuna contra la enfermedad de la gripe en pollos de engorde. Int J Enteric Pathog (2016) 4:10–35689. doi: 10.17795/ijep.35689
69. Talebi A, Amirzadeh B, Mokhtari B, Gahri H. Effects of a multi-strain probiotic (PrimaLac) on performance and antibody responses to Newcastle disease virus and infectious bursal disease virus vaccination in buriler chickens. Avian Pathol (2008) 37:509–12. DOI: 10.1080/03079450802356995
Resumen de PubMed | Texto completo de CrossRef | Google Académico
70. Isolauri E, Joensuu J, Suomalainen H, Luomala M, Vesikari T. Improved immunogenicity of oral d x RRV reassortant rotavirus vaccine by lactobacillus casei GG. Vacuna (1995) 13:310. doi: 10.1016/0264-410X(95)93319-5
Resumen de PubMed | Texto completo de CrossRef | Google Académico
71. Haghighi HR, Gong J, Gyles CL, Hayes MA, Sharif S. Los probióticos estimulan la producción de anticuerpos naturales en pollos. Clin Vaccine Immunol (2006) 13:975–80. doi: 10.1128/CVI.00161-06
Resumen de PubMed | Texto completo de CrossRef | Google Académico
72. Ohland CL, Macnaughton WK. Bacterias probióticas y función de barrera epitelial intestinal. AJP Gastrointestinal Liver Physiol (2010) 298:G807–19. doi: 10.1152/ajpgi.00243.2009
73. Sohail MU, Ijaz A, Yousaf MS, Ashraf K, Zaneb H, Aleem M, et al. Alivio del estrés por calor cíclico en pollos de engorde mediante la suplementación dietética de manano-oligosacárido y probióticos a base de lactobacilos: dinámica de cortisol, hormonas tiroideas, colesterol, proteína C reactiva e inmunidad humoral. Poultry Sci (2010) 89:1934–38. DOI: 10.3382/PS.2010-00751
Palabras clave: Lactobacillus plantarum, pollo de plumas amarillas, rendimiento de crecimiento, inmunidad, población de flora intestinal
Cita: Yin Y, Liao Y, Li J, Pei Z, Wang L, Shi Y, Peng H, Tan Y, Li C, Bai H, Ma C, Gong Y, Wei T y Peng H (2023) Lactobacillus plantarum GX17 beneficia el rendimiento del crecimiento y mejora las funciones de la barrera intestinal / flora intestinal entre pollos de engorde de plumas amarillas. Frente. Immunol. 14:1195382. doi: 10.3389/fimmu.2023.1195382
Recibido: 28 de marzo de 2023; Aprobado: 06 Junio 2023;
Publicado: 03 julio 2023.
Editado por:
Yigang Xu, Universidad A & F de Zhejiang, China
Revisado por:
Dandan Liu, Universidad de Yangzhou, China LI Jianmei, Universidad de Emory, Estados Unidos
Shaohui Wang, Academia China de Ciencias Agrícolas, China
Derechos de autor © 2023 Yin, Liao, Li, Pei, Wang, Shi, Peng, Tan, Li, Bai, Ma, Gong, Wei y Peng. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Hao Peng, hpeng2006@163.com; Tianchao Wei, tcwei88@126.com
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía