
Utilidad de las enzimas alimenticias y derivados de levadura en pollos de engorde

Utilidad de las enzimas alimenticias y derivados de levadura para mejorar los efectos nocivos de la coccidiosis en la salud intestinal y la función en pollos de engorde
 Elías G. Kiarie1*†,
Elías G. Kiarie1*†,  Haley Leung1,
Haley Leung1,  Reza Akbari Moghaddam Kakhki1†,
Reza Akbari Moghaddam Kakhki1†,  Rob Patterson2 y
Rob Patterson2 y  John R. Barta3
John R. Barta3- 1Departamento de Biociencias Animales, Universidad de Guelph, Guelph, ON, Canadá
- 2Departamento de Servicios Técnicos e Innovación, Canadian Bio-Systems Inc., Calgary, AL, Canadá
- 3Departamento de Patobiología, Universidad de Guelph, Guelph, ON, Canadá
Las lesiones necróticas inducidas por coccidiosis afectan la capacidad digestiva y la función de barrera en concurrencia con un mayor riesgo de infecciones bacterianas secundarias. La industria ha tenido éxito en el control de la coccidiosis con anticoccidiales y vacunación. Sin embargo, las preocupaciones sobre las especies de Eimeria resistentes a los anticoccidiales, las brechas en la vacunación y la restricción de antibióticos están estimulando la investigación y la aplicación de estrategias alternativas y / o complementarias para el control de la coccidiosis. El objetivo de este trabajo es evaluar la literatura sobre la utilidad de las enzimas alimenticias y los derivados de levaduras en la modulación de la coccidiosis. Las enzimas alimenticias pueden complementar las enzimas endógenas (proteasa, amilasa y lipasa) que pueden volverse insuficientes en las aves afectadas por la coccidiosis. La coccidiosis en el intestino delgado superior crea condiciones que mejoran la eficacia de la fitasa y hay informes que indican que la fitasa suplementaria puede mitigar el impacto negativo de la coccidiosis en la calidad ósea. El aumento de los ácidos grasos intestinales de cadena corta debido a las enzimas que degradan la fibra suplementaria se ha relacionado con una menor capacidad de supervivencia de Eimeria. Hay evidencia de que la levadura entera (viva o muerta) y los derivados pueden modular la coccidiosis. Se ha demostrado que las propiedades de inmunomudulación de los derivados de levadura mejoran la inmunidad celular y humoral en modelos de desafío de Eimeria, que es fundamental para la efectividad de la vacunación coccidial. Además, los nucleótidos de levadura han demostrado ser beneficiosos para estimular la curación de la superficie de la mucosa intestinal. Otro trabajo novedoso ha demostrado que ciertas células de levadura pueden producir derivados con compuestos anticoccidiales eficaces para atenuar el desprendimiento de ooquistes. La superficie de las células de levadura también ha demostrado ser un vehículo eficaz de administración de la vacuna oral Eimeria. En general, si bien se justifica una mayor investigación de refinamiento para abordar las inconsistencias en las respuestas y la aplicación comercial, existe evidencia de que las enzimas alimenticias y los derivados de levadura podrían complementar las estrategias para mantener la función intestinal para reforzar el rendimiento del crecimiento en pollos de engorde comprometidos con coccidiosis. Sin embargo, los pollos de engorde reciben dietas que contienen varios aditivos para piensos con distintos modos de acción y, sin embargo, hay escasez de datos empíricos sobre las respuestas esperadas. Las evaluaciones futuras deben considerar combinaciones de aditivos para documentar las respuestas de los animales y las posibles sinergias.
Introducción
Se estima que la población humana mundial alcanzará los 9.6 millones para 2050, durante este período, se espera que la producción de pollos de engorde crezca un 121% para satisfacer la demanda de proteína animal (1). Sin embargo, el sector de producción de proteína animal está bajo presión para producir productos alimenticios de manera ética, ambientalmente sostenible y saludable. Por ejemplo, la agricultura animal utiliza cantidades significativas de antibióticos para la terapia, la prevención de infecciones bacterianas y la promoción del crecimiento. Existe una creciente preocupación en todo el mundo sobre el uso indiscriminado de antibióticos y la vinculación con la aparición de patógenos resistentes a los antibióticos. Estas preocupaciones han requerido el cese legislativo y de los consumidores y/o restricciones en el uso de antibióticos para estimular el crecimiento (AGP). Además, existe una creciente demanda por parte de los consumidores de productos avícolas especiales criados con regímenes orgánicos, de dieta vegetal y de pastoreo. Estos cambios en las formas de producción avícola están trayendo nuevos desafíos y exacerbando los antiguos relacionados con la salud de las aves, el bienestar animal y las regulaciones. En el contexto de la salud y la nutrición de las aves de corral, las principales preocupaciones son el aumento de la incidencia de enfermedades entéricas como la coccidiosis, la enteritis necrótica, la alteración de la digestión de nutrientes y la absorción, lo que en última instancia conduce a una eficiencia deficiente en la conversión alimenticia, así como a un aumento de la mortalidad y la condena en la planta de procesamiento. Todos estos aspectos convergen para reivindicar la importancia del control efectivo y la prevención de patógenos entéricos para garantizar la seguridad alimentaria y la seguridad de una población humana en crecimiento.
Causada por parásitos protozoarios del filo apicomplexan, género Eimeria, la coccidiosis está inexplicablemente vinculada al avance y la modernización de la producción avícola y el impacto global anual asciende a más de $ 3 mil millones en pérdidas de morbilidad y mortalidad (2, 3). Los protozoos invaden las células intestinales como parte del ciclo de vida, lo que lleva a una digestión y absorción deficientes, función de barrera e infecciones bacterianas secundarias (4-6). Los parásitos exhiben notables sitios específicos de especies de desarrollo y focos de patología dentro del tracto intestinal (5, 7-9). Eimeria acervulina, E. máximos, y E. Tenella es la especie que se encuentra con mayor frecuencia en los sistemas de producción de pollos de engorde comerciales (8). Las altas densidades animales observadas en estos sistemas de producción son favorables para la transmisión de Eimeria (8). La infección por Eimeria también exacerba la proliferación intestinal de patógenos como Clostridium, perfringens y Salmonella enterica serovars Enteritidis o Typhimurium (9-12). De ello se deduce que la coccidiosis no solo tiene implicaciones en la salud de las aves, sino que también puede comprometer la seguridad alimentaria (13).
En reconocimiento de los efectos negativos de la coccidiosis en la producción avícola, la industria ha desarrollado y adoptado durante mucho tiempo anticoccidiales o vacunación viva o combinaciones de estas estrategias para el control (6, 14). Sin embargo, la preocupación por las especies de Eimeria resistentes a los anticoccidiales y la preocupación pública por el uso de medicamentos en la producción animal está limitando las opciones de quimioterapia (15). La vacunación depende del ciclo óptimo de Eimeria a través de cada parvada, es intensiva en manejo y la protección cruzada a cepas de tipo salvaje no es 100% efectiva (11, 16, 17). Además, la vacunación implica el suministro de especies vivas de Eimeria dentro del primer día de vida de los pollitos, lo que puede aumentar el riesgo de trastornos entéricos (18). Existen numerosos aditivos alimentarios alternativos a las estrategias tradicionales de control de la coccidiosis que se afirma que atenúan o remedian el daño intestinal estructural y funcional ocasionado por la coccidiosis (19). La intención de esta revisión es evaluar el conjunto de datos publicados sobre la función de las enzimas alimenticias y los derivados de levaduras en la modulación de la coccidiosis.
Enzimas alimenticias
La propuesta de aplicación de enzimas exógenas en la nutrición avícola fue sugerida inicialmente hace casi 100 años (20), sin embargo, el costo prohibitivo no permitió su aplicación en nutrición animal hasta muchas décadas después (21). Las xilanasas y las β-glucanasas fueron pioneras en las enzimas alimenticias comerciales para tratar los alimentos viscosos problemáticos, como la cebada y el trigo (22-25). Los primeros experimentos mostraron que la suplementación de estas enzimas en dietas ricas en alimentos viscosos mejoró la digestibilidad, el rendimiento del crecimiento y redujo los costos de alimentación (21-23). Estos estudios ayudaron a los científicos a comprender los modos de acción y estimularon nuevos esfuerzos de investigación y desarrollo para innovar nuevas actividades dirigidas a sustratos específicos y estabilizadas para soportar los rigores del procesamiento de alimentos y las condiciones gastrointestinales (26). De hecho, la utilidad de las enzimas alimenticias en la nutrición de los no rumiantes es ampliamente aceptada (21-23, 25, 27). Las enzimas alimenticias se aplican en gran medida en los programas de alimentación monogástrica bajo la premisa de que los animales no pueden digerir el 100% de los componentes de la dieta. Por ejemplo, los pollos de engorde excretan entre el 25 y el 30% de la materia seca ingerida en el estiércol (28). Esto se debe a factores antinutricionales (ANF) como el ácido fítico o fracciones no digeribles por las condiciones y la matriz de enzimas digestivas en el TGI (29, 30). La mayoría de las enzimas comerciales para piensos se desarrollan y aplican para atacar tales ANF (25). Además, la aplicación en aves jóvenes está impulsada por el hecho de que el tracto gastrointestinal no está bien desarrollado debido a (1) un sistema inmune inmaduro, (2) capacidad secretora endécima endógena limitada y (3) microbiota intestinal inestable (31-33). Por lo tanto, la adopción original de la tecnología de enzimas alimenticias en la nutrición avícola fue degradar el ANF en los piensos y complementar las enzimas endógenas en el intestino de los animales comprometidos, particularmente los pollitos recién nacidos.
Evolución del papel de las enzimas alimenticias en la nutrición avícola
La presión sobre los costos de alimentación es y seguirá siendo un factor decisivo para la producción avícola rentable y sostenible, y las enzimas de la alimentación tienen un papel establecido en la reducción de los costos de alimentación al aumentar la flexibilidad de las opciones de ingredientes de la alimentación. Además, la necesidad de reducir la excreción de nutrientes en la cadena de valor de la proteína animal eleva la utilidad de la enzima alimentaria en las operaciones avícolas. Sin embargo, cuestiones emergentes como la restricción en el uso de AGP han estimulado nuevas direcciones y perspectivas para la aplicación de enzimas para piensos. La evidencia emergente gira en torno a la evaluación de las enzimas alimenticias como parte de un programa integrado de gestión de la salud intestinal (24). La peculiaridad es que la microbiota intestinal se nutre de nutrientes luminales (dietéticos y/o endógenos) (34). Debido a las diferencias en la preferencia del sustrato y los requisitos de crecimiento, la composición y estructura de la digesta influye en gran medida en el microbioma GIT (24). De ello se deduce que el perfil del microbioma y la función metabólica reflejan en parte la composición del alimento (34). Por lo tanto, es plausible que la manipulación de la digestibilidad de la dieta influya en el microbioma GIT (24). Además, las enzimas degradadoras de fibra podrían liberar productos de hidrólisis «prebióticos» que pueden modular la microbiota intestinal (24, 35-37).
Levadura entera y derivados
Las levaduras son unicelulares, de 5-10 μm de tamaño, microorganismos eucariotas pertenecientes al reino de los hongos (38). Las levaduras son importantes en muchos ecosistemas complejos y están involucradas en interacciones simbióticas, mutualistas, parasitarias y competitivas con otros microorganismos. Desde la primera observación de A. van Leeuwenhoek en 1680 y el descubrimiento de su función en la fermentación por Louis Pasteur en la década de 1850, los humanos han explotado la levadura para la producción de alimentos y bebidas, entre muchas otras aplicaciones durante eones (39). Curiosamente, aunque hay más de 1.000 especies conocidas de levadura, muy pocas son explotadas comercialmente (40). La mayoría de las especies de levadura no son dañinas ni beneficiosas, y se sabe que pocas son patógenas para los seres humanos y/o los animales (40). El género Saccharomyces tiene ~ 20 especies que son de importancia industrial significativa, por ejemplo, etanol, pan, proteína unicelular y producción de vitaminas (40). Se ha estimado que la producción mundial anual de Saccharomyces cerevisiae supera la producción de todos los demás microorganismos industriales (41). Candida utilis (anteriormente clasificada como Torulopsis utilis) y comercialmente conocida como «Levadura Torula» es única ya que utiliza azúcares pentosa, por lo que es muy útil en el procesamiento de pulpa de madera a papel. Otra levadura importante es Kluyveromyces marxianus o la «levadura de suero de leche» para el procesamiento de productos lácteos. Aunque la explotación comercial de la levadura se basa principalmente en los procesos de fermentación tradicionales, el avance en biología molecular ha abierto enormes oportunidades para desarrollar cepas de levadura para diversas aplicaciones. Por ejemplo, Komagataella (Pichia) pastoris, S. cerevisiae, Ogataea (Hansenula) polymorpha, para la producción heteróloga de proteínas (40-42).
Hay muchos ingredientes para piensos asociados a la levadura y aditivos para piensos que se producen, comercializan y aplican en la agricultura animal en todo el mundo (43). Los principales ingredientes de los piensos, como los granos de destilería con solubles (DDGS), la levadura de cerveza, la levadura de suero y los coproductos de panadería, se derivan de los procesos de fermentación de la levadura (44, 45). Las levaduras se utilizan como fuentes ricas de proteínas, minerales, vitaminas (particularmente vitaminas B) y otros nutrientes para humanos y animales. Se ha sugerido que la producción de proteína unicelular a partir de levadura tiene enormes ventajas en relación con las plantas, los animales y otras fuentes microbianas de proteína debido a su rápida tasa de crecimiento en una amplia variedad de sustratos, incluidos los desechos industriales y agrícolas (46). Además, el tamaño celular relativamente grande y la capacidad de floculación de las levaduras las hace más fáciles de cosechar que las bacterias en medios de fermentación (46). Otros productos especiales de levadura incluyen selenio de levadura y levadura Phaffia rhodozyma que mejora el color de la carne en salmón y trucha (43). Las levaduras y productos nutricionales se utilizan en la industria de piensos como fuentes de aminoácidos y micronutrientes (43). Sin embargo, la utilidad de los productos de levadura en la agricultura animal ha evolucionado para explotar sus atributos funcionales. De particular interés son los componentes funcionales del contenido celular, como péptidos, enzimas, nucleótidos y constituyentes de la pared celular como β-glucanos, glicoproteínas, mananos y quitina (47, 48). Las secciones siguientes describirán brevemente los atributos funcionales de la levadura y los derivados que han demostrado influir en la salud y el estado inmunológico de las aves de corral.
Vive la levadura entera
Muchas especies de levadura son reconocidas como seguras por muchas autoridades reguladoras, como el estado de Presunción Calificada de Seguridad Alimentaria asignado por la Autoridad Europea de Seguridad Alimentaria, la Asociación Americana de Funcionarios de Control de Alimentos y la Agencia Canadiense de Inspección de Alimentos. Sin embargo, en general, la mayoría de los aditivos probióticos comerciales para piensos para aves de corral son de preparaciones bacterianas, por ejemplo, (49-53). Los pocos probióticos no bacterianos (levadura o hongos) incluyen Aspergillus oryzae (54, 55), Candida pintolopesii (54), Candida saitoana, Saccharomyces bourlardii (56) y S. cerevisiae (57). Podría decirse que los probióticos a base de levadura son indispensables en la nutrición de rumiantes por su efectividad en la modulación del microbioma ruminal (58, 59). La levadura seca activa es una de las levaduras viables más comunes utilizadas como probióticos en la producción ganadera. Los probióticos de levadura que se utilizan en la agricultura animal como aditivos para piensos suelen contener materiales portadores como piedra caliza, cáscaras de arroz y / o solubles de destilería. Estos productos suelen contener 5 × 109 unidades formadoras de colonias por gramo que representan el 20-25% de las UFC de levadura seca pura y activa (43). Los probióticos comerciales a base de levadura se fabrican principalmente en forma seca y se han planteado preocupaciones sobre su estabilidad en los procesos de fabricación de piensos (55, 60-62). Por ejemplo, el alimento suplementado con células de levadura activas se sometió a peletización (82 ° C) o extrusión (72 ° C para 31 s) (61). La peletización no afectó el recuento total de levadura, pero el número viable de levadura se redujo 10 veces. Sin embargo, la extrusión redujo los recuentos de levadura totales y viables. La mayoría de las dietas de las aves de corral se someten a un riguroso procesamiento de piensos, incluida la reducción del tamaño de partícula y el procesamiento hidrotérmico para mejorar la eficiencia y la higiene del alimento (26). Lo que sugiere que se esperaría que la supervivencia de las células de levadura desprotegidas sea baja en los alimentos para aves de corral sometidos a procesamiento hidrotérmico.
Derivados de levadura
Los derivados de levadura se denominan colectivamente cultivos de levadura y se componen en gran medida de una combinación de biomasa de levadura y productos de fermentación producidos en procesos de fermentación condicionados. También pueden estar presentes trazas de células de levadura residuales viables. Su producción implica la inoculación de medios de cultivo específicos con células de levadura vivas y la posterior fermentación en condiciones específicas, sobre las cuales se seca posteriormente todo el medio fermentado. La masa cosechada a menudo se formula en aditivo para piensos o se somete a un procesamiento posterior para producir productos especiales. La producción de productos especiales se considera un diferenciador clave de muchos aditivos funcionales para piensos a base de levaduras disponibles para la industria avícola. Como organismos heterótrofos, la energía y el metabolismo del carbono están íntimamente interconectados, lo que da a las células de levadura la capacidad de producir una amplia variedad de derivados dependiendo de la composición de los medios de fermentación y las condiciones de fermentación (63). De ello se deduce que la producción de cultivo de levadura puede manipularse para producir aditivos alimentarios únicos que contengan derivados únicos o combinados beneficiosos para la nutrición y la salud animal.
Enzimas
Las enzimas fueron descubiertas por primera vez por el químico francés Anselme Payen en 1833 (64). Décadas más tarde, Louis Pasteur concluyó que la fermentación estaba correlacionada con la vida y la organización de las células de levadura, pero no con la muerte celular (65). Se puede argumentar entonces que las levaduras fueron los organismos pioneros para la producción de enzimas, sin embargo, el mercado de piensos está dominado por bacterias y enzimas derivadas de hongos filamentosos (21, 24, 25, 37). Esto se debe principalmente a que los sistemas de expresión microbiana no por levadura son ventajosos en términos de ciertos desarrollos de productos y procesos (66). Sin embargo, con el avance en biotecnología algunas levaduras por ejemplo, K. (Pichia) pastoris, S. cerevisiae, O. (Hansenula) Polymorpha y algunas otras especies de levadura se han desarrollado para la producción industrial de enzimas y proteínas (66-68). Sin embargo, estas proteínas de levadura refinadas se aplican en la producción de productos químicos especializados, como los productos intermedios farmacéuticos (69).
Nucleótidos
La concentración total de ácidos nucleicos en la levadura entera varía de 3 a 12% de peso de células secas (70, 71). Las levaduras también son ricas en nucleasas endógenas y proteasas que pueden degradar ácidos nucleicos, ADN y ARN en nucleótidos a través de la autólisis (71, 72). Al controlar el pH, la temperatura y la duración, así como el uso de aditivos como la sal y las enzimas exógenas, la autólisis de las células de levadura puede optimizarse y estandarizarse para una calidad constante del producto (73). Estas modificaciones se utilizan cada vez más para producir productos de nucleótidos de levadura para diversas aplicaciones industriales. Por ejemplo, en condiciones normales de autólisis, el ARN se degrada principalmente a tres nucleótidos primarios, sin embargo, bajo hidrólisis enzimática controlada, se producen 5 nucleótidos principales de guanina, adenina, citosina y uracilo (73, 74).
Componentes de la pared celular (JOC)
La JOC representa alrededor del 15 al 20% del peso seco de la levadura y es rica en β-glucanos y mananos, así como trazas de quitina. Estructuralmente, la JOC está formada por una capa interna de β-glucanos y mananos insolubles, una capa intermedia de β-glucanos solubles y la capa externa de glicoproteína (75). Sin embargo, debe tenerse en cuenta que estas capas no son discretas, sino que forman estructuras complejas que son recalcitrantes a la descomposición (75). β-1,3 glucanos con enlaces de rama β-1,6 son el principal componente polisacárido en la JOC y se ha demostrado que muestran efectos inmunomoduladores (76). En este contexto, existe un creciente interés en la extracción refinada de β-glucanos a través de métodos mecánicos (por ejemplo, molienda de perlas, sonicación, homogeneización a alta presión) y no mecánicos (por ejemplo, termólisis, choque osmótico, químicos y enzimáticos) (75, 77, 78). Los mananos consisten en α-1,6 enlaces con cadenas laterales de manosa en α-1,2 enlaces (79).
Utilidad de las enzimas alimenticias y derivados de levadura en la mitigación de los efectos negativos de la coccidiosis
Modelos experimentales de desafío
Las pérdidas de producción, el aumento de la mortalidad, la reducción del bienestar animal y el aumento del riesgo de contaminación de los productos avícolas debido a enfermedades entéricas son motivo de gran preocupación para la industria avícola (9, 10, 12, 13). Hay muchas investigaciones que utilizaron modelos de desafío de patógenos entéricos para examinar la efectividad de un aditivo para piensos o una estrategia dietética (80, 81). Este modelo in vivo permite la evaluación de un aditivo alimentario dado en el contexto de un agente patógeno infeccioso que forma parte del ecosistema gastrointestinal. Sin embargo, en términos de identificar los factores predisponentes más influyentes, un modelo de infección confiable y reproducible es crítico. La infección por coccidios con ooquistes vivos esporulados a través de sonda oral (cultivo) o contaminación por hojarasca se usa comúnmente en modelos experimentales de coccidiosis y es razonablemente confiable en general (82). Sin embargo, existen variaciones en los enfoques experimentales relacionados con la dosificación, la especificación de la especie, el momento y la composición (de tipo silvestre o atenuada) para la sobredosis de vacunación y la coinfección con Clostridium perfringen, entre otros (82, 83). Para la investigación nutricional, es imperativo tener un modelo de coccidiosis que no solo sea reproducible, sino que se dirija a secciones del intestino que tengan ramificaciones significativas en la digestión y absorción de nutrientes.
Nuestro laboratorio ha desarrollado un modelo para examinar los efectos de la coccidiosis en la digestión y las capacidades de absorción y los efectos posteriores en la ecología de GIT. La razón es utilizar este modelo de infección para probar estrategias dietéticas durante la fase aguda y las fases de recuperación (84-88). El enfoque general es desafiar submuestras de aves en un corral con dosis altas (100.000 E. acervullina y 60.000 ooquistes esporulados de E. maxima) para generar lesiones macroscópicas y el resto de aves con una dosis baja (25.000 E. acervulina y 5.000 ooquistes esporulados de E. maxima) para examinar las consecuencias de la digestión y absorción de nutrientes alterada. Brevemente, los parásitos Eimeria se propagan y purifican según Shirley (89) y la dosis se basa en ensayos de titulación en el laboratorio de parasitología del Dr. John Barta (Universidad de Guelph). Como se muestra en la Figura 1, hemos sido capaces de reproducir puntuaciones de lesión consistentes en alineación con la biología de los organismos de desafío, lo que indica una alta reproducibilidad del modelo de desafío. La destrucción de Eimeria del revestimiento intestinal da como resultado un rendimiento de crecimiento deficiente, excrementos sueltos y muerte en casos extremos (5, 14). El fracaso de los animales parasitados para crecer se debe en parte a la pérdida de apetito y la malabsorción de nutrientes (84, 86, 92-94). Los daños estructurales y funcionales al intestino delgado están indicados por la histomorfología, las enzimas digestivas, los transportadores de nutrientes y la retención de nutrientes en nuestro modelo (84, 86, 87). Además, la infección por E. acervulina y E. maxima regula la expresión de enzimas digestivas y transportadores de nutrientes (84, 87, 93-95). Las secciones siguientes evaluarán la literatura donde se utilizó el desafío de coccidiosis para evaluar el impacto de las enzimas alimenticias y los derivados de levadura en la modulación de la expresión de coccidiosis.
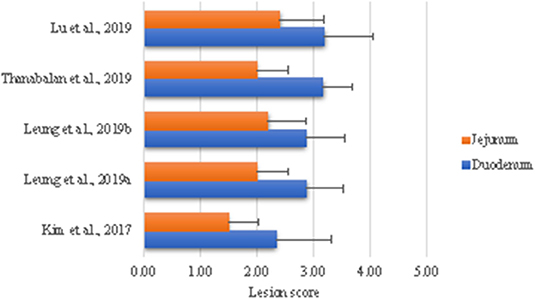 FIGURA 1. Puntuaciones de lesiones intestinales en pollos de engorde desafiados con Eimeria. Todas las aves fueron desafiadas con 100.000 ooquistes esporulados de E. acervullina y 60.000 E. maxima el día 10 de edad, con excepción de Kim et al. (84) en cuyo caso las aves fueron desafiadas el día 5. Se realizó una necropsia a las aves 5 días después del desafío para evaluar las puntuaciones de las lesiones descritas por Price et al. (90) utilizando una escala de 0 (ninguna) a 4 [alta; Johnson y Reid (91)].
FIGURA 1. Puntuaciones de lesiones intestinales en pollos de engorde desafiados con Eimeria. Todas las aves fueron desafiadas con 100.000 ooquistes esporulados de E. acervullina y 60.000 E. maxima el día 10 de edad, con excepción de Kim et al. (84) en cuyo caso las aves fueron desafiadas el día 5. Se realizó una necropsia a las aves 5 días después del desafío para evaluar las puntuaciones de las lesiones descritas por Price et al. (90) utilizando una escala de 0 (ninguna) a 4 [alta; Johnson y Reid (91)].
Utilidad de las enzimas alimenticias en la modulación de la coccidiosis en pollos de engorde
La coccidiosis reduce eficazmente la digestión y absorción de nutrientes relacionados con la anorexia en concurrencia con el daño intestinal morfológico y funcional (Figura 2) (84, 86, 87). El aumento de la mucogénesis y el recambio de enterocitos (84), así como los cambios metabólicos posteriores a la absorción y la activación del sistema inmunológico, probablemente influyen en las necesidades de nutrientes de los pollos de engorde (9). Por ejemplo, la infección por Eimeria aumentó la proliferación de células de la mucosa yeyunal en un 40% en concomitante con el aumento de la profundidad de la cripta, lo que indica que las aves priorizaron el desarrollo intestinal después del insulto intestinal (84). También se observó un aumento en la proliferación celular en la base de la cripta de pollos desafiados por Eimeria (96). En general, el requerimiento de energía de mantenimiento aumenta en proporción al tamaño corporal metabólico a medida que el ave madura. Sin embargo, se ha demostrado que la coccidiosis aumentó notablemente el requerimiento de energía de mantenimiento. Por ejemplo, Leung et al. (86), observaron una disminución del 16% en la energía asignada al aumento de peso corporal (medido en eficiencia calórica) en un ave de 35 días desafiada con E. acervulina y E. Maxima al día 10 de edad. La energía necesaria para el desarrollo de la inmunidad fue del 5% en aves sanas en comparación con el 28% para las aves con problemas de coccidiosis y este costo se elevó desproporcionadamente a medida que las aves envejecían (97).
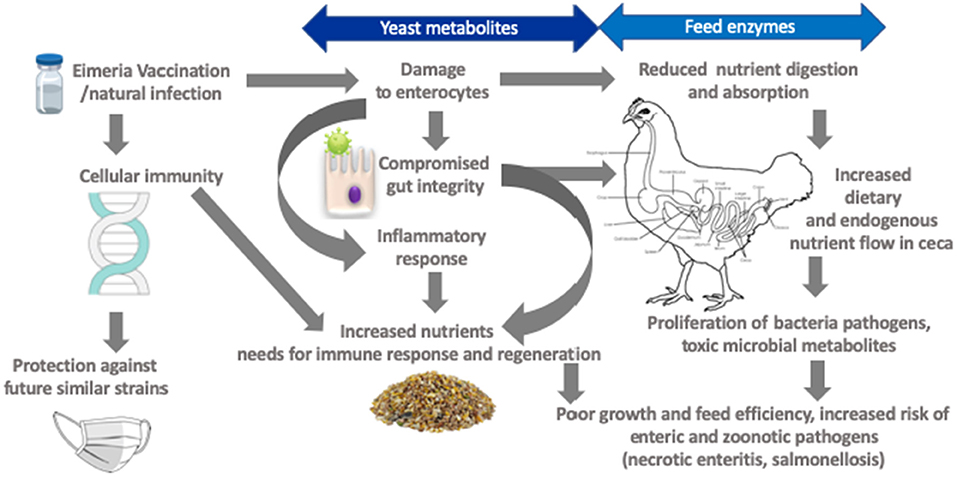 FIGURA 2. Marco para objetivos susceptibles de derivados de levaduras y enzimas alimenticias para mitigar los efectos negativos de la coccidiosis.
FIGURA 2. Marco para objetivos susceptibles de derivados de levaduras y enzimas alimenticias para mitigar los efectos negativos de la coccidiosis.
Con disminución de la capacidad digestiva y elevada ineficiencia en la utilización de energía en aves afectadas por coccidiosis; Es plausible que las enzimas exógenas suplementarias del alimento puedan ser beneficiosas. Sin embargo, pocos estudios investigaron el impacto de la suplementación de enzimas alimenticias para complementar las enzimas endógenas (p.ej., amilasa, proteasa, lipasa) en aves con riesgo de coccidiosis. La suplementación dietética con una proteasa redujo el impacto negativo de una infección por coccidiosis (E. acervulina, E. maxima y E. tenella) en el aumento de peso corporal en pollos de engorde, pero no tuvo efectos sobre las lesiones y la eliminación de ooquistes (98). En contraste, Parker et al. (99) mostraron que una mezcla de enzimas (amilasa, proteasa y xilanasa) alimentada a pollos de engorde vacunados contra la coccidiosis no tuvo ningún efecto sobre la digestibilidad de los nutrientes ileales o el rendimiento del crecimiento, pero redujo las puntuaciones de las lesiones. El daño intestinal asociado a E. acervulina y E. maxima se han relacionado con efectos adversos sobre la salud ósea principalmente porque infectan el duodeno y el yeyuno superior, los principales sitios de absorción de minerales (85, 100, 101). En este contexto, el papel de la fitasa suplementaria en la utilización de minerales también se ha investigado en el modelo de infección por coccidios (102, 103). La peculiaridad es que el pH del duodeno de las aves sanas es de 6,0 o mayor, pero se reduce a <5 en las aves infectadas con coccidiosis (104). Se cree que un pH duodenal más bajo mejora la eficacia de la fitasa vinculada al rango óptimo de pH (2.5 y 5.5) para la degradación efectiva del fitato (105). De hecho, la fitasa aumentó el rendimiento de crecimiento y la concentración de ceniza tibial en presencia o ausencia de E. acervulina (102). La coccidiosis redujo el rendimiento del crecimiento y la absorción de calcio y fósforo, lo que resultó en una reducción de la resistencia ósea, sin embargo, la suplementación con fitasa no mitigó los efectos negativos de la coccidiosis en la utilización de fósforo (106). La suplementación de fitasa, proteasa y xilanasa solos o en combinación no mitigó la reducción en el rendimiento de crecimiento en pollos de engorde expuestos a una vacuna mixta contra coccidios (107).
Utility of Yeast Probiotics in Modulating Coccidiosis in Broilers
There are limited studies on yeast probiotic (e.g., S. boulardii) supplementation in broilers (111, 112). The common approach is a blend of yeast probiotics with bacterial cultures on the premise that beneficial effects of probiotics are genus, species and strain specific and use of multi-strain and multi-species might be more effective than mono-strain probiotics (54, 55). Indeed, some investigations have shown that co-supplementing yeast and bacterial probiotics enhanced their survival and growth (113, 114). Moreover, aggregation of lactobacillus with yeasts enhanced tolerance in gastric or intestinal juices (115). For example, a supplement containing Lactobacillus acidophilus, Bacillus subtilis, S. cerevisiae, and A. oryzae improved live body weight gain linked to enhanced nutrients utilization and intestinal microbial modulation (55). A combination of yeast and (L. acidophilus and Streptococcus faecium) enhanced growth performance in broilers through increased digestion and absorption of nutrients (116). Lactobacillus fermentum and S. cerevisiae was shown to modulate intestinal immune system without negative effects on growth performance in broilers (57). In contrast, a blend (Lactobacillus plantarum, Lactobacillus delbrueckii ssp. bulgaricus, L. acidophilus, Lactobacillus rhamnosus, Bifidobacterium bifidum, Streptococcus salivarius ssp. thermophilus, Enterococcus faecium, A. oryzae, and Candida pintolepesii) did not ameliorate negative effects of delayed feed access in newly hatched chicks on growth performance and gastrointestinal physiology (54). A challenge of evaluating studies of probiotic blends is that experimental design does not always incorporate single strains to characterize responses of each strain vs. combination.
Saccharomyces cerevisiae var. boulardii is one of the most researched non-bacterial probiotics with proven benefits in various human gastrointestinal disease models (117). This yeast was originally isolated from litchi fruit in Indochina by Henri Boulard in 1920 and has been used for treatment of intestinal diseases in children and adults since the 1950’s (117, 118). Several mechanisms have been suggested as to the broad health-promoting effects of consuming yeast probiotics in humans and span from local general trophic effects to action on both innate and/or adaptive immunity (117, 119, 120). Clinical trials included mitigation of antibiotic-associated diarrhea; Clostridium difficile diarrhea, irritable bowel syndrome and inflammatory bowel diseases (117, 121). Folignè et al. (117) tested six yeast strains for anti-inflammatory potential and demonstrated that yeast-mediated protection seems to take place predominantly at the level of the intestinal mucosa. The authors indicated that prophylactic reinforcement and therapeutic restoration of barrier function by changing the luminal environment stimulated the mucosal barrier. This work extended previous observations that showed yeast probiotics enhanced epithelial integrity and reduced bacterial translocation in various sepsis models (122, 123). The mode of action of yeasts in controlling enteric diseases have not been elucidated but have been associated with release of antimicrobial peptides, acidification of surrounding environment, alteration of inflammatory and immune responses or destruction of toxic factors (63).
Aunque el uso de levadura probiótica para controlar las enfermedades entéricas en humanos se ha estudiado ampliamente, se ha prestado poca atención a las enfermedades entéricas de los animales de granja. En un estudio, los pollos de engorde fueron alimentados con 1 o 100 g de S. boulardii/kg de alimento y desafiados con S. typhimurium (124). Los autores observaron un 70% de colonización en la cecá del ave control vs. 20 o 5% de colonización en aves alimentadas con 1 o 100 g, respectivamente. Un estudio innovador investigó la actividad anticoccidial de un compuesto (s) aislado del cultivo de levadura Meyerozyma guilliermondii (125). Se demostró que los compuestos tienen actividad anticoccidial contra los ooquistes de E. tenella bajo simulación in vitro relacionada con la viabilidad reducida de los ooquistes. En otras investigaciones, S. cerevisiae se investigó como vehículo de administración de vacunas orales de Eimeria proporcional a la administración exitosa anterior de vacunas virales orales (126, 127). En este contexto, una proteína micronema (EtMic2) de E. tenella que está íntimamente involucrada en la invasión de la célula huésped (128) se expresó en la superficie de células vivas de S. cerevisiae (129). Las células de levadura enteras sin o con EtMic2 en sus superficies se utilizaron como una vacuna oral viva contra el desafío de E. tenella en pollitas. Se observó una diseminación de ooquistes significativamente menor y puntuaciones de lesión en las aves que recibieron levadura EtMic2 en relación con el control o las aves que recibieron levadura sin proteína EtMic2 (129). El grupo EtMic2 también mostró mayores aumentos de peso. En general, la utilidad de la vacunación contra la coccidiosis en la industria se basa principalmente en el desarrollo de inmunidad celular para la protección contra la exposición posterior a Eimeria (130). Sin embargo, se ha demostrado que los antígenos de superficie inmunodominantes identificados en E. acervulina y E. maxima provocan respuestas de anticuerpos medibles, particularmente la producción de IgA, además de estimular la inmunidad mediada por células (131). Por lo tanto, existe la posibilidad de que los anticuerpos (generados por la inmunización viva o contra antígenos purificados de Eimeria específicos de la etapa) inhiban el desarrollo del parásito (128, 129, 132). Las aplicaciones de campo de los compuestos anticoccidiales secretados por las levaduras y/o la utilidad de la levadura para administrar vacunas deben investigarse para su aplicación comercial como complemento para fármacos anticoccidiales y/o como adyuvante para las vacunas contra los cocos.
Utilidad de los extractos de levadura ricos en nucleótidos en la modulación de la coccidiosis en pollos de engorde
Como unidades básicas del ADN y el ARN, los nucleótidos están presentes en todas las células vivas (133, 134). Con un suministro adecuado de energía y aminoácidos, los nucleótidos pueden sintetizarse de novo. Sin embargo, pueden volverse condicionalmente esenciales durante la enfermedad, los períodos de ingesta limitada de alimento o el rápido crecimiento (133, 134). Estas condiciones son proporcionales al daño gastrointestinal ocasionado por la infección por Eimeria (9). Se han reportado evaluaciones de extractos de levadura ricos en nucleótidos sobre crecimiento, eficiencia alimenticia y desarrollo intestinal y salud en pollos de engorde (86, 87, 135, 136). Sin embargo, hay estudios limitados que evalúan los efectos de la suplementación con nucleótidos en la dieta sobre el desarrollo de órganos inmunes en pollos de engorde desafiados con Eimeria. Se demostró que los bebés humanos alimentados con nucleótidos tenían una mayor producción de IgA (137) relacionada con el requerimiento de linfocitos para nucleótidos exógenos (138). Además, se ha demostrado que la suplementación con nucleótidos dietéticos en bebés afecta otras funciones inmunes, como la activación de células NK y macrófagos, la producción de citoquinas esplénicas y el número de células secretoras de anticuerpos (138). Leung et al. (87) evaluaron el efecto de la suplementación de suplementos de nucleótidos de levadura rica en nucleótidos sobre el rendimiento del crecimiento, la histomorfología intestinal, la expresión de genes intestinales seleccionados y la actividad microbiana durante la fase aguda del desafío de Eimeria (7 días después del desafío). Los nucleótidos suplementarios mejoraron la histomorfología yeyunal y la expresión del transportador de nutrientes transportador de aminoácidos catiónicos 1 (CAT1). Curiosamente, el impacto de los nucleótidos y Eimeria fue independiente de la comunidad de microbiota pero interactivo de la actividad microbiana. De manera instructiva, hubo una tendencia a disminuir la diversidad alfa con la suplementación con nucleótidos proporcional a la observada con la provisión de antimicrobianos (139). Estos cambios en la diversidad alfa y en las poblaciones de microbiota pueden tener impactos a largo plazo o pueden amplificarse aún más con el tiempo. Los efectos también podrían caer en cascada en la producción de ácidos grasos volátiles y cambiar el pH cecal y, posteriormente, factores como la histomorfología y el desarrollo del sistema inmunológico. En estudios posteriores, Leung et al. (86) alimentaron pollos de engorde con extracto de levadura rico en nucleótidos y los desafiaron con Eimeria en d 10 post-nacimiento. La concentración de IgA plasmática y mucosa y los pesos de los órganos inmunes (bolsa, bazo y timo) se determinaron en d 5 y 25 después del desafío. No hubo efectos en d 5 mediciones post-desafío, sin embargo, las aves alimentadas con nucleótidos de levadura mostraron bursa más pesada en d 25 post-desafío. Parece que la suplementación con nucleótidos puede atenuar algunos de los efectos negativos de Eimeria, sin embargo, se justifican más investigaciones para determinar el período óptimo de suplementación y la concentración y el efecto de los nucleótidos individuales.
Es importante considerar la relación entre Eimeria y Clostridium perfringens en la evaluación del papel de las enzimas alimenticias en el control de la coccidiosis. La infección por Eimeria aumenta las pérdidas endógenas de proteínas plasmáticas y mucina que nutre C. perfringens (9). El gran flujo de materiales nitrogenados en la ceca promueve la producción de metabolitos tóxicos, por ejemplo, tioles, aminas, amoníaco e indoles (108), pero lo más importante es que el alto pH de la digesta de ceca promueve la proliferación de patógenos como C. perfringens (9, 12). Las enzimas alimenticias pueden modular la ecología de la TGI reduciendo los nutrientes no digeridos y la producción de oligosacáridos con posibles efectos prebióticos (Figura 2) (24). Por ejemplo, la suplementación con mezcla de enzimas (amilasa, proteasa y xilanasa) apoyó la ecología intestinal que redujo las puntuaciones de lesiones intestinales, particularmente en la ceca relacionada con perfiles microbianos alterados en pollos de engorde vacunados con coccidios (99). Los autores interpretaron que, aunque la mezcla de enzimas no influyó en la digestibilidad ileal de los nutrientes, alteró las características de la digesta de tal manera que se alteraron las comunidades de microbiota de la ceca. La suplementación con fitasa no tuvo efectos sobre la excreción de ooquistes en pollos de engorde vacunados con cocos y no tratados sometidos a provocación de Eimeria, pero redujo las puntuaciones de lesión (106). El efecto de las enzimas suplementarias sobre las puntuaciones de lesión en pollos de engorde afectados por coccidiosis se ha atribuido a la producción de ácidos grasos volátiles (34). Un elegante estudio de Ruff et al. demostró que la coccidiosis redujo el pH luminal del intestino delgado pero aumentó el pH de la ceca (109). Por lo tanto, es relevante que se haya demostrado que las enzimas suplementarias del alimento aumentan la concentración de ácidos grasos volátiles como los ácidos acético y butírico con la reducción concomitante del pH de la ceca digesta (29). Se demostró que el ácido acético es proporcional al fármaco anticoccidial (Amprolium) para suprimir los efectos negativos asociados a la coccidiosis sobre el rendimiento del crecimiento (110). En conjunto, estos estudios sugirieron que las enzimas suplementarias afectan de alguna manera la supervivencia o la extensión del daño intestinal.
Utilidad de los probióticos de levadura en la modulación de la coccidiosis en pollos de engorde
Los componentes de la pared celular de levadura (β-glucanos y mananos) se han relacionado con la modulación del sistema inmune (140, 141), la unión a toxinas y células patógenas (142) y las interacciones con los constituyentes intestinales (143, 144). Hooge (145), informó un metanálisis de las dietas suplementadas con componentes de la pared celular de S. cerevisiae var. boulardii, 29 experimentos) o dietas de control positivo suplementadas con antibióticos (promotor del crecimiento antimicrobiano, 21 experimentos). Los metanálisis revelaron una magnitud pequeña (<2% de mejora frente a control negativo) pero un impacto significativo de la JOC en el aumento de peso corporal y la tasa de conversión alimenticia, pero ninguna diferencia entre el control positivo (145). Sin embargo, las aves alimentadas con JOC mostraron una mortalidad 21,4 y 18,1% menor en comparación con el control negativo y positivo, respectivamente. Los productos de pared celular de levadura pueden servir como patrones moleculares asociados a microbios y modular la expresión de receptores de reconocimiento de patrones (PRRS) (136, 146). De hecho, la JOC suplementaria mostró un rendimiento de crecimiento proporcional y respuestas de habitabilidad a la bacitracina de zinc y la salinomicina en un modelo de coinfección por Eimeria y C. perfringens (147). Los macrófagos de pollo están involucrados en las respuestas inmunes adaptativas a través de la interacción con Eimeria en la mucosa intestinal (148). Se ha demostrado que los antígenos de superficie inmunodominantes identificados en E. acervulina y E. maxima provocan respuestas de anticuerpos mensurables, siendo IgA el isotipo más importante (131). Por lo tanto, aunque E. acervulina y E. maxima desafío redujeron la IgA de la mucosa yeyunal en un 33% a los 5 días después del desafío, la concentración aumentó en un 16% a los 25 días después del desafío (86). Otro estudio indicó que el desafío de E. acervulina y E. maxima aumentó la IgA yeyunal en un 64% a los 5 días posteriores al desafío (149). Aunque los mecanismos específicos de acción intestinal de la IgA sobre las infecciones coccidiales todavía son objeto de investigación, se ha especulado que la IgA reduce el desarrollo de esporozoítos o merozoítos y previene la invasión de la célula huésped (150, 151). La relevancia es que se sabe que los componentes de la JOC son inmunomoduladores y se ha demostrado que la suplementación dietética aumenta las secreciones locales de IgA de la mucosa, así como las respuestas inmunes celulares y humorales (Figura 2) (152).
Es plausible que los componentes de la JOC puedan modular las respuestas inmunes mediadas por células y humores contra las infecciones coccidiales (130). La JOC dietética (1 o 10 g/kg) redujo la gravedad de la infección y la eliminación de ooquistes de una sola E. tenella o mezcla de E. acervulina, E. maxima y E. tenella desafío en pollos de engorde (153, 154). La suplementación dietética con 5 g/kg de derivados autolizados de S. cerevisiae estimuló la secreción de IgA de la mucosa intestinal, las respuestas inmunes humorales y mediadas por células, y redujo el desprendimiento de ooquistes en pollos de engorde sometidos a vacunación contra coccidios (152). Sin embargo, la JOC (0,5 g/kg) alimentada sola o en combinación con tanino (0,5 g/kg) no redujo la gravedad de la infección con una mezcla de E. acervulina, E. maxima y E. tenella en pollos de engorde. Los pollos de engorde alimentados con un suplemento (1-2 g / kg) que contenía levadura muerta entera mostraron una disminución en el desprendimiento de ooquistes, así como un aumento en la producción de óxido nítrico de macrófagos y la producción de citoquinas inflamatorias (155). Se probó una mezcla refinada de JOC y β-glucanos y un extracto de levadura cruda en pollos de engorde sometidos a 10 veces la vacuna Coccivac B (156). Aunque ambos productos no tuvieron impacto en el rendimiento del crecimiento en relación con el control, las aves alimentadas con mezcla refinada tuvieron una menor expresión de IL-6 en la mucosa ileal y las alimentadas con levadura cruda mostraron una mejor inmunoglobulina G sérica (156). Estos resultados contradictorios están posiblemente relacionados con las diferencias en la inclusión de JOC en el pienso y las dosis de inoculación de Eimeria spp.
Las investigaciones en la programación del desarrollo han contribuido a nuestra comprensión de la interfaz de la descendencia materna y continúan planteando preguntas importantes con respecto al manejo nutricional de los animales reproductores (157, 158). En las especies aviares, el desarrollo de la inmunocompetencia se inicia durante la embriogénesis (159). El papel protector de los anticuerpos maternos es crítico debido a la naturaleza precoz de las especies aviares (33). Los anticuerpos del reproductor se depositan en el huevo y continúan funcionando en los primeros años de vida de los pollitos (33). Se ha investigado el papel de la inmunidad materna en las infecciones coccidiales en pollitos (160, 161). Por ejemplo, la infección de gallinas reproductoras con Eimeria maxima indujo la producción de anticuerpos específicos del parásito que se transfirieron a los pollitos (160). Estos anticuerpos fueron altamente protectores, mediando hasta un 97% de reducción en la diseminación de ooquistes en crías desafiadas. Recientemente demostramos que la alimentación de los reproductores de pollos de engorde con levadura producto rico en componentes de la pared celular hidrolizada enzimática aumentó la deposición de IgA en las yemas de huevo para incubar (149). Los datos indicaron además que la alimentación de la pared celular de levadura hidrolizada a los reproductores de pollos de engorde y a los pollitos mejoró la histomorfología yeyunal independiente del desafío de Eimeria (149).
Perfeccionamiento adicional y direcciones futuras
La producción mundial de aves de corral se ha triplicado a una producción anual de 90 millones de toneladas de carne de pollo y 1,1 billones de huevos (http://www.fao.org/faostat/) en las últimas dos décadas. Se espera una mayor expansión en respuesta a la creciente población humana. El sector también está bajo presión para producir productos alimenticios de manera ética, ambientalmente sostenible y saludable. Por ejemplo, el uso indiscriminado de antibióticos es un problema global de actualidad con un énfasis creciente en estrategias alternativas para la prevención y el control efectivos de patógenos entéricos. Aquí hay una descripción general de la coccidiosis, sus implicaciones en la salud y función intestinal y los objetivos susceptibles de modulación por enzimas alimenticias y derivados de levaduras (Figura 2). En gran medida, la industria avícola ha aceptado ampliamente el uso de enzimas alimenticias para mejorar la digestión del alimento. Es plausible que las enzimas exógenas suplementarias del alimento puedan contrarrestar la disminución de la capacidad digestiva en las aves afectadas por coccidiosis. Las enzimas alimenticias podrían modular la ecología de la ceca al reducir el flujo de nutrientes no digeridos y promover la fermentación ácida. Crear efectivamente condiciones que reduzcan la capacidad de supervivencia de la proliferación de Eimeria y C. perfringens. La levadura y los derivados de levadura se han asociado con la alteración de las respuestas inflamatorias e inmunes. Existe una gran oportunidad para desarrollar una nueva generación de derivados de levadura con características optimizadas en términos de interacción con Eimeria y modulación del sistema inmune del huésped. El papel protector de los anticuerpos maternos es de interés porque los anticuerpos depositados en el huevo y los niveles transferidos a la descendencia están directamente relacionados con los niveles circulantes de estas inmunoglobulinas en la madre. Se necesita más investigación para refinar la relación entre la composición y la función de los derivados de levadura con el fin de seleccionar una nueva generación de fracciones de levadura con características optimizadas para su aplicación en reproductoras de pollos de engorde. Al igual que muchas evaluaciones de aditivos para piensos, las inconsistencias de las respuestas son a menudo una preocupación importante. La utilidad del modelo experimental de desafío de enfermedades puede ayudar a imitar las condiciones del campo; Sin embargo, la dosificación correcta del ingrediente activo es crítica. Aunque los proveedores y las agencias reguladoras han avanzado en el uso de enzimas alimenticias en la medida en que se puede determinar la dosificación y la recuperación en el alimento, hay escasez de análisis para cuantificar los derivados de levadura en el alimento, lo cual es fundamental para una dosificación precisa. La dosificación precisa es esencial para lograr los beneficios deseados y prevenir la alimentación excesiva de componentes biológicamente activos. Además, la investigación de aditivos para piensos rara vez considera que los piensos comerciales para pollos de engorde contienen una gama de aditivos. Tal vez, las futuras investigaciones sobre aditivos para piensos deberían considerar la suplementación conjunta de para documentar las respuestas y las posibles sinergias.
Contribuciones del autor
El NS y la AR eran estudiantes graduados de EK y participaron en la ejecución de estudios de desafío de coccidiosis y en algunas búsquedas bibliográficas. RP proporcionó conocimientos técnicos sobre enzimas alimenticias y derivados de levaduras y revisó la integración de conceptos. JB es un parasitólogo experto en el modelo de desafío de Eimeria. Todos los autores revisaron el manuscrito. EK buscó en la literatura y escribió una parte significativa de la revisión y tuvo la responsabilidad conceptual y editorial general.
Financiación
Este trabajo fue apoyado por el Consejo de Investigación de Ciencias Naturales e Ingeniería de Canadá (programa Discovery).
Conflicto de intereses
RP es un empleado de Canadian Bio-Systems Inc.
Los autores restantes declaran que la investigación se realizó en ausencia de cualquier relación comercial o financiera que pudiera interpretarse como un posible conflicto de intereses.
Reconocimientos
Se aprecian las contribuciones de los estudiantes graduados actuales y anteriores y los asociados de investigación del Laboratorio de Investigación de Nutrición Monogástrica EK en la Universidad de Guelph.
Referencias
1. Mottet A, Tempio G. Global poultry production: current state and future outlook and challenges. Mundo Poult Sci J. (2017) 73:245–56. doi: 10.1017/S0043933917000071
2. Dalloul RA, Lillehoj HS. Coccidiosis avícola: avances recientes en las medidas de control y desarrollo de vacunas. Experto Rev Vacc. (2006) 5:143–63. doi: 10.1586/14760584.5.1.143
Resumen de PubMed | Texto completo de CrossRef | Google Académico
3. Kadykalo S, Roberts T, Thompson M, Wilson J, Lang M, Espeisse O. El valor de los anticoccidiales para la producción avícola mundial sostenible. Int J Agentes antimicrobianos. (2018) 51:304–10. doi: 10.1016/j.ijantimicag.2017.09.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
4. Barta JR. Coccidiosis. En: eLS. Nueva York, NY: John Wiley and Sons (2001). doi: 10.1038/npg.els.0001947
5. Chapman HD. Hitos en la investigación de la coccidiosis aviar: una revisión. Poult Sci. (2014) 93:501–11. DOI: 10.3382/PS.2013-03634
Resumen de PubMed | Texto completo de CrossRef | Google Académico
6. Chapman HD, Barta JR, Hafeez MA, Matsler P, Rathinam T, Raccoursier M. La epizootiología de las infecciones por Eimeria en pollos de engorde comerciales donde se emplearon programas de medicamentos anticoccidiales en seis parvadas sucesivas para controlar la coccidiosis. Poult Sci. (2016) 95:1774–8. doi: 10.3382/ps/pew091
Resumen de PubMed | Texto completo de CrossRef | Google Académico
7. Chapman HD, Roberts B, Shirley MW, Williams RB. Directrices para evaluar la eficacia y seguridad de las vacunas anticoccidiales vivas y obtener la aprobación para su uso en pollos y pavos. Pathol aviar. (2005) 34:279–90. doi: 10.1080/03079450500178378
Resumen de PubMed | Texto completo de CrossRef | Google Académico
8. Shirley MW, Smith AL, Tomley FM. La biología de la Eimeria aviar con énfasis en su control por vacunación. En: Baker JR, Muller R, Rollinson D, editores. Avances en Parasitología. Amsterdam: Academic Press (2005), p. 285–330.
9. Williams RB. Coccidiosis intercurrente y enteritis necrótica de pollos: manejo racional e integrado de la enfermedad mediante el mantenimiento de la integridad intestinal. Pathol aviar. (2005) 34:159–80. DOI: 10.1080/03079450500112195
Resumen de PubMed | Texto completo de CrossRef | Google Académico
10. Arakawa A, Baba E, Fukata T. La infección por Eimeria tenella aumenta la infección por Salmonella typhimurium en pollos. Poult Sci. (1981) 60:2203–9. DOI: 10.3382/PS.0602203
Resumen de PubMed | Texto completo de CrossRef | Google Académico
11. Williams RB. Vacunas anticoccidiales para pollos de engorde: caminos hacia el éxito. Pathol aviar. (2002) 31:317–53. DOI: 10.1080/03079450220148988
Resumen de PubMed | Texto completo de CrossRef | Google Académico
12. Timbermont L, Haesebrouck F, Ducatelle R, Van Immerseel F. Enteritis necrótica en pollos de engorde: una revisión actualizada sobre la patogénesis. Pathol aviar. (2011) 40:341–7. doi: 10.1080/03079457.2011.590967
Resumen de PubMed | Texto completo de CrossRef | Google Académico
13. Cosby DE, Cox NA, Harrison MA, Wilson JL, Buhr RJ, Fedorka-Cray PJ. Salmonella y resistencia a los antimicrobianos en pollos de engorde: una revisión. J Appl Poult Res. (2015) 24:408–26. doi: 10.3382/japr/pfv038
14. Chapman HD. Una contribución histórica a la ciencia avícola: el control profiláctico de la coccidiosis en aves de corral. Poult Sci. (2009) 88:813–5. DOI: 10.3382/PS.2008-00316
Resumen de PubMed | Texto completo de CrossRef | Google Académico
15. Chapman HD, Jeffers TK. La vacunación de pollos contra la coccidiosis mejora la resistencia a los medicamentos en la producción avícola comercial. Int J Parasitol. (2014) 4:214–7. doi: 10.1016/j.ijpddr.2014.10.002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
16. Joyner LP. Variación inmunológica entre dos cepas de Eimeria acervulina. Parasitología. (1969) 59:725–73. doi: 10.1017/S0031182000031243
Resumen de PubMed | Texto completo de CrossRef | Google Académico
17. Martin AG, Danforth HD, Barta JR, Fernando MA. Análisis de la protección cruzada inmunológica y sensibilidades a fármacos anticoccidiales entre cinco cepas geográficas y temporales de Eimeria maxima. Int. J. Parasitol. (1997) 27:527–33. doi: 10.1016/S0020-7519(97)00027-1
Resumen de PubMed | Texto completo de CrossRef | Google Académico
18. Kogut MH, Klasing K. La perspectiva de un inmunólogo sobre nutrición, inmunidad y enfermedades infecciosas: introducción y visión general. J Appl Poult Res. (2009) 18:103–10. DOI: 10.3382/JAP.2008-00080
19. Quiroz-Castañeda RE, Dantán-González E. Control de la coccidiosis aviar: alternativas naturales futuras y presentes. Biomed Res Int. (2015) 2015:430610. doi: 10.1155/2015/430610
Resumen de PubMed | Texto completo de CrossRef | Google Académico
20. Hervey GW. Un estudio nutricional sobre una enzima de hongo. Ciencia. (1925) 62:247. doi: 10.1126/science.62.1602.247
Resumen de PubMed | Texto completo de CrossRef | Google Académico
21. Masey O’neill HV, Smith JA, Bedford MR. Multicarbohydrase enzymes for non-rumiants. Asiático-australas. J Anim Sci. (2014) 27:290–301. doi: 10.5713/ajas.2013.13261
22. Bedford MR, Schulze H. Enzimas exógenas para cerdos y aves de corral. (1998) 11:91–114. doi: 10.1079/NRR19980007
Resumen de PubMed | Texto completo de CrossRef | Google Académico
23. Slominski BA. Avances recientes en la investigación sobre enzimas para dietas avícolas. Poult Sci. (2011) 90:2013–23. DOI: 10.3382/PS.2011-01372
Resumen de PubMed | Texto completo de CrossRef | Google Académico
24. Kiarie E, Romero LF, Nyachoti CM. El papel de las enzimas alimenticias añadidas en la promoción de la salud intestinal en cerdos y aves de corral. (2013) 26:71–88. doi: 10.1017/S0954422413000048
Resumen de PubMed | Texto completo de CrossRef | Google Académico
25. Kiarie E, Walsh MC, Nyachoti CM. Rendimiento, función digestiva y respuestas de la mucosa a aditivos alimentarios seleccionados para cerdos. J Anim Sci. (2016) 94:169–80. DOI: 10.2527/JAS.2015-9835
26. Kiarie EG, Mills A. Role of feed processing on gut health and function in pigs and poultry: conundrum of optimal particle size and hydrothermal regimens. Front Vet Sci. (2019) 6:19. DOI: 10.3389/fvets.2019.00019
Resumen de PubMed | Texto completo de CrossRef | Google Académico
27. Adeola O, Cowieson AJ. Revisión invitada por la Junta: oportunidades y desafíos en el uso de enzimas exógenas para mejorar la producción animal de no rumiantes. J Anim Sci. (2011) 89:3189–218. doi: 10.2527/jas.2010-3715
Resumen de PubMed | Texto completo de CrossRef | Google Académico
28. Ravindran V. Advances and future directions in poultry nutrition: an overview. Coreano J Poult Sci. (2012) 39:53–62. doi: 10.5536/KJPS.2012.39.1.053
29. Kiarie E, Romero LF, Ravindran V. Growth performance, nutrient utilization, and digesta characteristics in broiler chickens fed corn or wheat diets without or with supplemental xylanase. Poult Sci. (2014) 93:1186–96. DOI: 10.3382/PS.2013-03715
30. Kiarie E, Walsh MC, Romero LF, are not S, Ravindran V. Las respuestas de utilización de nutrientes y fibra de la xilanasa suplementaria en pollos de engorde alimentados con dietas a base de trigo son independientes del período de adaptación para probar las dietas. Poult Sci. (2017) 96:3239–45. doi: 10.3382/ps/pex100
Resumen de PubMed | Texto completo de CrossRef | Google Académico
31. Croom WJ, Brake J, Coles BA, Havenstein GB, Christensen VL, Mcbride BW, et al. ¿La tasa de capacidad de absorción intestinal limita el rendimiento en aves de corral? J Appl Poult Res. (1999) 8:242–52. DOI: 10.1093/JAP/8.2.242
32. Gilbert ER, Li H, Ernmersonj DA, Webb KE, Wong EA. Developmental regulation of nutrient transporter and enzyme mRNA abundance in the small bow of broilers. Poult Sci. (2007) 86:1739–53. DOI: 10.1093/PS/86.8.1739
Resumen de PubMed | Texto completo de CrossRef | Google Académico
33. Friedman A, Elad O, Cohen I, Bar Shira E. El sistema linfoide asociado al intestino en el pollito post-nacimiento: dinámica de la IgA materna. Isr J Vet Med. (2012) 67:75–81. Disponible en línea en: http://www.ijvm.org.il/sites/default/files/friedman.pdf
34. Apajalahti J, Kettunen A, Graham H. Características de las comunidades microbianas gastrointestinales, con especial referencia al pollo. Mundo Poult Sci J. (2004) 60:223–32. doi: 10.1079/WPS20040017
35. Courtin CM, Broekaert WF, Swennen K, Lescroart O, Onagbesan O, Buyse J, et al. La inclusión dietética de arabinoxilooligosacáridos de salvado de trigo induce efectos nutricionales beneficiosos en pollos. Química de cereales. (2008) 85:607–13. doi: 10.1094/CCHEM-85-5-0607
36. Wealleans AL, Walsh MC, Romero LF, Ravindran V. Efectos comparativos de dos combinaciones multienzimáticas y un probiótico Bacillus sobre el rendimiento del crecimiento, la digestibilidad de la energía y los nutrientes, la desaparición de polisacáridos sin almidón y la microflora intestinal en pollos de engorde. Poult Sci. (2017) 96:4287–97. DOI: 10.3382/ps/pex226
Resumen de PubMed | Texto completo de CrossRef | Google Académico
37. Bedford MR. La evolución y aplicación de las enzimas en la industria de la alimentación animal: el papel de la interpretación de datos. Br Poult Sci. (2018) 59:486–93. doi: 10.1080/00071668.2018.1484074
Resumen de PubMed | Texto completo de CrossRef | Google Académico
38. Bennett JW. Micotecnología: el papel de los hongos en la biotecnología1Basado en una conferencia celebrada en el simposio «Progress in US Biotechnology», en el 8º Congreso Europeo de Biotecnología (BCE8) celebrado en Budapest (Hungría) en agosto de 1997.1. J Biotechnol. (1998) 66:101–7. doi: 10.1016/S0168-1656(98)00133-3
39. Barnett JA. Una historia de la investigación sobre levaduras 2: Louis Pasteur y sus contemporáneos, 1850-1880. Levadura. (2000) 16:755–71. doi: 10.1002/1097-0061(20000615)16:8<755::AID-YEA587>3.0.CO;2-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
40. Kurtzman C, Fell JW, Boekhout T. Clasificación de la levadura. En: Kurtzman C, Fell JW, Boekhout T, editores. Las levaduras, un estudio taxonómico. Londres: Elsevier (2011), p. 3–8. doi: 10.1016/B978-0-444-52149-1.00001-X
41. Jansen MLA, Bracher JM, Papapetridis I, Verhoeven MD, De Bruijn H, De Waal PP, et al. Cepas de Saccharomyces cerevisiae para la producción de etanol de segunda generación: de la exploración académica a la implementación industrial. FEMS Levadura Res. (2017) 17:FOX044. DOI: 10.1093/femsyr/fox044
Resumen de PubMed | Texto completo de CrossRef | Google Académico
42. Nielsen J, Larsson C, Van Maris A, Pronk J. Ingeniería metabólica de levadura para la producción de combustibles y productos químicos. Curr Opin Biotechnol. (2013) 24:398–404. doi: 10.1016/j.copbio.2013.03.023
Resumen de PubMed | Texto completo de CrossRef | Google Académico
43. Shurson GC. Levadura y derivados de levadura en aditivos e ingredientes para piensos: fuentes, características, respuestas animales y métodos de cuantificación. Anim Feed Sci Technol. (2018) 235:60–76. doi: 10.1016/j.anifeedsci.2017.11.010
44. Stein HH, Shurson GC. Revisión invitada por la junta: el uso y la aplicación de granos secos de destilería con solubles en dietas porcinas. J Anim Sci. (2009) 87:1292–303. DOI: 10.2527/jas.2008-1290
Resumen de PubMed | Texto completo de CrossRef | Google Académico
45. Shurson GC. El papel de los coproductos de biocombustibles en la alimentación sostenible del mundo. Ann Rev Anim Biosci. (2017) 5:229–54. doi: 10.1146/annurev-animal-022516-022907
Resumen de PubMed | Texto completo de CrossRef | Google Académico
46. Ugalde UO, Castrillo JI. Proteínas unicelulares de hongos y levaduras. En: Khachatourians GG, Arora DK, editores. Micología Aplicada y Biotecnología. Ámsterdam: Elsevier (2002), p. 123–49. doi: 10.1016/S1874-5334(02)80008-9
47. Kollár R, Reinhold BB, Petráková E, Yeh HJC, Ashwell G, Drgonová J, et al. Arquitectura de la pared celular de la levadura: β(1 → 6)-glucano interconecta manoproteína, β(1 → 3)-glucano y quitina. J Biol Chem. (1997) 272:17762–75. DOI: 10.1074/JBC.272.28.17762
Resumen de PubMed | Texto completo de CrossRef | Google Académico
48. Cabib E, Farkas V, Kosík O, Blanco N, Arroyo J, Mcphie P. Assembly of the yeast cell wall: Crh1p and Crh2p act as transglycosylases in vivo and in vitro. J Biol Chem. (2008) 283:29859–72. DOI: 10.1074/JBC. M804274200
Resumen de PubMed | Texto completo de CrossRef | Google Académico
49. Waititu SM, Yitbarek A, Matini E, Echeverry H, Kiarie E, Rodriguez-Lecompte JC, et al. Efecto de la suplementación de microbios alimentados directamente sobre el rendimiento de los pollos de engorde, las digestibilidades de los nutrientes y las respuestas inmunes. Poult Sci. (2014) 93:625–35. DOI: 10.3382/PS.2013-03575
Resumen de PubMed | Texto completo de CrossRef | Google Académico
50. Mohammadigheisar M, Shirley RB, Barton J, Welsher A, Thiery P, Kiarie E. Growth performance and gastrointestinal responses in heavy Tom turkeys fed antibiotic free corn-soybean meal diets supplemented with multiple dose of a single strain Bacillus subtilis probiotic (DSM29784). Poult Sci. (2019) 98:5541–50. DOI: 10.3382/ps/pez305
Resumen de PubMed | Texto completo de CrossRef | Google Académico
51. Neijat M, Habtewold J, Shirley RB, Welsher A, Barton J, Thiery P, et al. Bacillus subtilis DSM29784 modula el microbioma cecal, la concentración de ácidos grasos de cadena corta y la retención aparente de componentes dietéticos en las claras de afeitadora durante las fases de crecimiento, revelado y colocación. Aplicación Environ Microbiol. (2019) 85:E00402–19. doi: 10.1128/AEM.00402-19
52. Neijat M, Kiarie E, Welsher A, Barton J, Shirley RB, Thiery P. Growth performance, apparent retention of components, and excreta dry matter content in Shaver White pollitas (5 to 16 week of age) in response to dietary supplementation of graded levels of a single strain Bacillus subtilis probiotic. Poult Sci. (2019) 98:3777–86. doi: 10.3382/ps/pez080
Resumen de PubMed | Texto completo de CrossRef | Google Académico
53. Neijat M, Shirley RB, Barton J, Thiery P, Welsher A, Kiarie E. Effect of dietary supplementation of Bacillus subtilis DSM29784 on hen performance, egg quality indices, and apparent retention of dietary components in landing hens from 19 to 48 weeks of age. Poult Sci. (2019) 98:5622–35. doi: 10.3382/ps/pez324
Resumen de PubMed | Texto completo de CrossRef | Google Académico
54. Daşkiran M, Önol AG, Cengiz Ö, Ünsal H, Türkyilmaz S, Tatli O, et al. Influencia de la inclusión de probióticos dietéticos en el rendimiento de crecimiento, parámetros sanguíneos y microflora intestinal de pollos de engorde machos expuestos al tiempo de retención posterior a la eclosión. J Appl Poult Res. (2012) 21:612–22. DOI: 10.3382/japr.2011-00512
55. Shim YH, Ingale SL, Kim JS, Kim KH, Seo DK, Lee SC, et al. Una formulación probiótica multimicrobiana procesada a bajas y altas temperaturas de secado: efectos sobre el rendimiento del crecimiento, la retención de nutrientes y la microbiología cecal de pollos de engorde. Br Poult Sci. (2012) 53:482–90. doi: 10.1080/00071668.2012.690508
Resumen de PubMed | Texto completo de CrossRef | Google Académico
56. Rahman M, Mustari A, Salauddin M, Rahman M. Effects of probiotics and enzymes on growth performance and hematobiochemical parameters in broilers. J Bangl Agric Univ. (2013) 11:111–8. doi: 10.3329/jbau.v11i1.18221
57. Bai SP, Wu AM, Ding XM, Lei Y, Bai J, Zhang KY, et al. Efectos de las dietas suplementadas con probióticos sobre el rendimiento del crecimiento y las características inmunes intestinales de pollos de engorde. Poult Sci. (2013) 92:663–70. DOI: 10.3382/PS.2012-02813
Resumen de PubMed | Texto completo de CrossRef | Google Académico
58. Chaucheyras-Durand F, Chevaux E, Martin C, Forano E. Use of yeast probiotics in rumiants: effects and mechanisms of action on rumen pH, fibre degradation, and microbiota according to the diet. En: Rigobelo EC, editor. Probiótico en animales. IntechOpen (2012), p. 119–52. DOI: 10.5772/50192
59. Vohra A, Syal P, Madan A. Probiotic yeasts in livestock sector. Anim Feed Sci Technol. (2016) 219:31–47. doi: 10.1016/j.anifeedsci.2016.05.019
60. Bayrock D, Ingledew WM. Fluidized bed drying of baker’s yeast: moisture levels, drying rates, and viability changes during drying. Comida Res Int. (1997) 30:407–15. doi: 10.1016/S0963-9969(98)00003-9
61. Aguirre-Guzmán G, Ricque-Marie D, Cruz-Suárez LE. Supervivencia de Saccharomyces cerevisiae aglomerado en alimentos para camarones peletizados. Acuicultura. (2002) 208:125–35. doi: 10.1016/S0044-8486(01)00711-6
62. Bayrock D, Ingledew WM. Mecanismo de pérdida de viabilidad durante el secado en lecho fluidizado de levadura de panadería. Comida Res Int. (1997) 30:417–25. doi: 10.1016/S0963-9969(97)00072-0
63. Hatoum R, Labrie S, Fliss I. Antimicrobial and probiotic properties of yeasts: from fundamental to novel applications. Microbiol frontal. (2012) 3:421. DOI: 10.3389/fmicb.2012.00421
Resumen de PubMed | Texto completo de CrossRef | Google Académico
64. Payen A, Persoz JF. Mémoire sur la diastase, les principaux produits de ses réactions et leurs applications aux arts industriels [Memoria sobre la diastasa, los principales productos de sus reacciones y sus aplicaciones a las artes industriales] [en línea]. (1933). Disponible en línea en: https://en.wikipedia.org/wiki/Enzyme#cite_note-9 (consultado el 29 de junio de 2019).
65. Manchester KL. Louis Pasteur (1822–1895)—El azar y la mente preparada. Tendencias Biotechnol. (1995) 13:511–5. doi: 10.1016/S0167-7799(00)89014-9
Resumen de PubMed | Texto completo de CrossRef | Google Académico
66. Klabunde J, Kunze G, Gellissen G, Hollenberg CP. Integration of heterologous genes in several yeast species using vectors containing a Hansenula polymorpha-derived rDNA-targeting element. FEMS Levadura Res. (2003) 4:185–93. doi: 10.1016/S1567-1356(03)00148-X
Resumen de PubMed | Texto completo de CrossRef | Google Académico
67. Schuller D, Casal M. El uso de cepas modificadas genéticamente de Saccharomyces cerevisiae en la industria vitivinícola. Aplicación Microbiol Biotechnol. (2005) 68:292–304. DOI: 10.1007/S00253-005-1994-2
Resumen de PubMed | Texto completo de CrossRef | Google Académico
68. Johnson EA. Biotechnology of non-saccharomyces yeasts—the ascomycetes. Aplicación Microbiol Biotechnol. (2013) 97:503–17. DOI: 10.1007/S00253-012-4497-Y
69. Kim H, Yoo SJ, Kang HA. Biología sintética de levaduras para la producción de proteínas terapéuticas recombinantes. FEMS Levadura Res. (2015) 15:1–16. doi: 10.1111/1567-1364.12195
Resumen de PubMed | Texto completo de CrossRef | Google Académico
70. Waldron C, Lacroute F. Effect of growth rate on the amount of ribosomal and transfer ribonucleic acids in yeast. J Bacteriol. (1975) 122:855–65.
71. Běhalová B, Bláhová M, Šillinger V, Machek F. Comparison of various ways of extraction of nucleic acids and of preparation of yeast extract from saccharomyces cerevisiae and Candida utilis. Acta Biotechnol. (1991) 11:547–52. doi: 10.1002/abio.370110608
72. Chaffin WL, López-Ribot JL, Casanova M, Gozalbo D, Martínez JP. Pared celular y proteínas secretadas de Candida albicans: identificación, función y expresión. Microbiol Mol Biol Rev. (1998) 62:130–80.
73. Chew LY, Toh GT, Ismail A. Capítulo 15 – aplicación de proteasas para la producción de péptidos bioactivos. En Kuddus M, editor. Enzymes in Food Biotechnology, Ámsterdam: Academic Press (2019), p. 247–61. doi: 10.1016/B978-0-12-813280-7.00015-3
74. Kempler GM. Producción de compuestos de sabor por microorganismos. En: Laskin AI, editores. Avances en Microbiología Aplicada. Academic Press (1983), p. 29–51. doi: 10.1016/S0065-2164(08)70353-8
Resumen de PubMed | Texto completo de CrossRef | Google Académico
75. Aimanianda V, Clavaud C, Simenel C, Fontaine T, Delepierre M, Latgé J-P. Pared celular beta-(1,6)-glucano de Saccharomyces cerevisiae: caracterización estructural y síntesis in situ. J Biol Chem. (2009) 284:13401–12. DOI: 10.1074/JBC. M807667200
Resumen de PubMed | Texto completo de CrossRef | Google Académico
76. Goodridge HS, Wolf AJ, Underhill DM. Reconocimiento de beta-glucano por el sistema inmune innato. (2009) 230:38–50. doi: 10.1111/j.1600-065X.2009.00793.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
77. Bzducha-Wróbel A, Błazejak S, Kawarska A, Stasiak-Rózanska L, Gientka I, Majewska E. Evaluación de la eficiencia de diferentes métodos de interrupción en la preparación de la pared celular de levadura para el aislamiento de β-glucano. Moléculas. (2014) 19:20941–61. doi: 10.3390/moléculas191220941
Resumen de PubMed | Texto completo de CrossRef | Google Académico
78. Schiavone M, Vax A, Formosa C, Martin-Yken H, Dague E, François JM. Un método químico y enzimático combinado para determinar cuantitativamente los componentes polisacáridos en la pared celular de las levaduras. FEMS Levadura Res. (2014) 14:933–47. doi: 10.1111/1567-1364.12182
Resumen de PubMed | Texto completo de CrossRef | Google Académico
79. Lesage G, Bussey H. Cell wall assembly in Saccharomyces cerevisiae. Microbiol Mol Biol Rev. (2006) 70:317–43. doi: 10.1128/MMBR.00038-05
Resumen de PubMed | Texto completo de CrossRef | Google Académico
80. Cooper KK, Songer JG. Virulencia de Clostridium perfringens en un modelo experimental de enteritis necrótica de aves de corral. Veterinario Microbiol. (2010) 142:323–8. doi: 10.1016/j.vetmic.2009.09.065
Resumen de PubMed | Texto completo de CrossRef | Google Académico
81. Peek HW, Landman WJM. Coccidiosis en aves de corral: productos anticoccidiales, vacunas y otras estrategias de prevención. Veterinario Q. (2011) 31:143–61. doi: 10.1080/01652176.2011.605247
Resumen de PubMed | Texto completo de CrossRef | Google Académico
82. Elmusharaf MA, Mohamed HE, Alhaidary A, Beynen AC. Eficacia y características de diferentes métodos de infección por coccidiosis en pollos de engorde. Am J Anim Vet Sci. (2010) 5:45–50. doi: 10.3844/ajavsp.2010.45.51
83. Shojadoost B, Vince AR, Prescott JF. La exitosa inducción experimental de enteritis necrótica en pollos por Clostridium perfringens: una revisión crítica. Vet Res. (2012) 43:74. doi: 10.1186/1297-9716-43-74
Resumen de PubMed | Texto completo de CrossRef | Google Académico
84. Kim E, Leung H, Akhtar N, Li J, Barta JR, Wang Y, et al. Rendimiento de crecimiento y respuestas gastrointestinales de pollos de engorde alimentados con harina de maíz y soja sin o con factor de crecimiento epidérmico exógeno al desafiar con Eimeria. Poult Sci. (2017) 96:3676–86. DOI: 10.3382/ps/pex192
Resumen de PubMed | Texto completo de CrossRef | Google Académico
85. Akbari Moghaddam Kakhki R, Lu Z, Thanabalan A, Leung H, Mohammadigheisar M, Kiarie E. El desafío de Eimeria afectó negativamente los atributos de los huesos largos relacionados con el aumento de la reabsorción en pollos de engorde de 14 días de edad. Poult Sci. (2019) 98:1615–21. doi: 10.3382/ps/pey527
Resumen de PubMed | Texto completo de CrossRef | Google Académico
86. Leung H, Patterson R, Barta JR, Karrow N, Kiarie E. Extracto de levadura rico en nucleótidos alimentado a pollos de engorde desafiados con Eimeria: impacto en el rendimiento del crecimiento, histomorfología yeyunal, sistema inmunológico y retención aparente de componentes dietéticos y eficiencia calórica. Poult Sci. (2019) 98:4375–83. DOI: 10.3382/ps/pez213
87. Leung H, Yitbarek A, Snyder R, Patterson R, Barta JR, Karrow N, et al. Las respuestas de los pollos de engorde al desafío de Eimeria cuando se les alimenta con un extracto de levadura rico en nucleótidos. Poult Sci. (2019) 98:1622–33. doi: 10.3382/ps/pey533
88. Thanabalan A, Moats J, Price K, Kiarie E. Omega-3 fatty acids sources fed to groiler breeders and/or their progeny: impact on growth performance and breast yield in 42-day old buriler chickens. En: 2019 Poultry Science Association Annual Meeting. Montreal, QC: Asociación de Ciencias Avícolas (2019).
89. Shirley MW. Especies y cepas de Eimeria en pollos. En: Eckert J, Braun R, Shirley MW, Coudert P, editores. COST89/820, Biotechnology Guidelines on Techniques in Coccidiosis Research. Luxemburgo: Comisión Europea (1995), pp. 1-25.
90. Price KR, Guerin MT, Barta JR. Éxito y fracaso: el papel de los niveles de humedad relativa y la gestión ambiental en la vacunación viva de Eimeria de pollitas de ponedoras de reemplazo criadas en jaula. J. Appl. Poultry Res. (2014) 23:523–35. DOI: 10.3382/japr.2014-00989
91. Johnson J, Reid M. Anticoccidial drugs: injury scoring techniques in battery and floor-pen experiments with chickens. Exp Parasitol. (1970) 28:30–6. doi: 10.1016/0014-4894(70)90063-9
Resumen de PubMed | Texto completo de CrossRef | Google Académico
92. Mayor JR Jr, Ruff MD. Eimeria spp.: influencia de los coccidios en la digestión (actividad amilolítica) en pollos de engorde. Exp Parasitol. (1978) 45:234–40. doi: 10.1016/0014-4894(78)90064-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
93. Su S, Miska KB, Fetterer RH, Jenkins MC, Wong EA. Expression of digestive enzymes and nutrient transporters in Eimeria acervulina-challenged layers and broilers. Poult Sci. (2014) 93:1217–26. DOI: 10.3382/PS.2013-03807
Resumen de PubMed | Texto completo de CrossRef | Google Académico
94. Su S, Miska KB, Fetterer RH, Jenkins MC, Wong EA. Expression of digestive enzymes and nutrient transporters in Eimeria-challenged broilers. Exp Parasitol. (2015) 150:13–21. doi: 10.1016/j.exppara.2015.01.003
Resumen de PubMed | Texto completo de CrossRef | Google Académico
95. Adams C, Vahl HA, Veldman A. Interacción entre la nutrición y la infección por Eimeria acervulina en pollos de engorde: desarrollo de un modelo experimental de infección. H. J Nutr. (1996) 75:867–73. doi: 10.1079/BJN19960192
Resumen de PubMed | Texto completo de CrossRef | Google Académico
96. Sun L, Dong H, Zhang Z, Liu J, Hu Y, Ni Y, et al. La activación de la proliferación epitelial inducida por la infección por Eimeria acervulina en el duodeno puede estar asociada con el metabolismo del colesterol. Oncotarget. (2016) 7:27627–40. doi: 10.18632/oncotarget.8490
Resumen de PubMed | Texto completo de CrossRef | Google Académico
97. Teeter RG, Beker A. Cómo el manejo y la salud intestinal influyen en la eficiencia calórica de las aves de corral. En: WATT Poultry USA. Rockford, IL: WATT Global Media (2011). Disponible en línea en: https://www.wattagnet.com/articles/11083-how-management-intestinal-health-influence-poultry-caloric-efficiency
98. Peek HW, Van der Klis JD, Vermeulen B, Landman WJM. La proteasa dietética puede aliviar los efectos negativos de una infección por coccidiosis en el rendimiento de producción en pollos de engorde. Anim Feed Sci Technol. (2009) 150:151–9. doi: 10.1016/j.anifeedsci.2008.08.006
99. Parker J, Oviedo-Rondón EO, Clack BA, Clemente-Hernández S, Osborne J, Remus JC, et al. Enzimas como aditivo para piensos para ayudar en las respuestas contra especies de eimeria en pollos de engorde vacunados con coccidios alimentados con dietas de harina de maíz y soja con diferentes niveles de proteínas. Poult Sci. (2007) 86:643–53. DOI: 10.1093/PS/86.4.643
Resumen de PubMed | Texto completo de CrossRef | Google Académico
100. Turk DE. Absorción de calcio durante infecciones coccidiales en pollitos. Poult Sci. (1973) 52:854–7. DOI: 10.3382/PS.0520854
Resumen de PubMed | Texto completo de CrossRef | Google Académico
101. Van Der Klis JD, Verstegen MWA, De Wit W. Absorption of minerals and retention time of dry matter in the gastrointestinal tract of broilers. Poult Sci. (1990) 69:2185–94. DOI: 10.3382/PS.0692185
Resumen de PubMed | Texto completo de CrossRef | Google Académico
102. Watson BC, Matthews JO, Southern LL, Shelton JL. Los efectos interactivos de la infección por Eimeria acervulina y la fitasa para pollos de engorde 1. Poult Sci. (2005) 84:910–3. doi: 10.1093/ps/84.6.910
103. Mansoori B, Modirsanei M, Nodeh H, Rahbari S. El efecto interactivo de la fitasa y los coccidios sobre las lesiones macroscópicas, así como la capacidad de absorción del intestino en pollos de engorde alimentados con dietas bajas en calcio y fósforo disponible. Veterinario Parasitol. (2010) 168:111–5. doi: 10.1016/j.vetpar.2009.10.018
Resumen de PubMed | Texto completo de CrossRef | Google Académico
104. Fox MC, Brown DR, Southern LL. Effect of dietary buffer additions on gain, efficiency, duodenal pH, and copper concentration in liver of Eimeria acervulina-infected chicks. Poult Sci. (1987) 66:500–4. DOI: 10.3382/PS.0660500
Resumen de PubMed | Texto completo de CrossRef | Google Académico
105. Kiarie E, Woyengo T, Nyachoti CM. Efficacy of new 6-phytase from Buttiauxella spp. on growth performance and nutrient retention in broiler chickens fed corn soybean meal-based diets Asian-Australas. J Anim Sci. (2015) 28:1479–87. doi: 10.5713/ajas.15.0059
Resumen de PubMed | Texto completo de CrossRef | Google Académico
106. Shaw AL, Van Ginkel FW, Macklin KS, Blake JP. Efectos de la suplementación con fitasa en dietas de pollos de engorde en un desafío natural de Eimeria en aves ingenuas y vacunadas. Poult Sci. (2011) 90:781–90. DOI: 10.3382/PS.2010-01158
Resumen de PubMed | Texto completo de CrossRef | Google Académico
107. Walk CL, Cowieson AJ, Remus JC, Novak CL, Mcelroy AP. Efectos de las enzimas dietéticas sobre el rendimiento y el número de células caliciformes intestinales de pollos de engorde expuestos a una vacuna viva contra el ooquiste de coccidios. Poult Sci. (2011) 90:91–8. DOI: 10.3382/PS.2010-00760
Resumen de PubMed | Texto completo de CrossRef | Google Académico
108. Nyachoti CM, Omogbenigun FO, Rademacher M, Blank G. Performance responses and indicators of gastrointestinal health in early-weaned pigs fed low-protein aminoacid-supplemented diets. J Anim Sci. (2006) 84:125–34. doi: 10.2527/2006.841125x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
109. Ruff MD, Johnson JK, Dykstra DD, Reid WM. Efectos de Eimeria acervulina sobre el pH intestinal en pollos convencionales y gnotobióticos. Avian Dis. (1974) 18:96–104. DOI: 10.2307/1589247
Resumen de PubMed | Texto completo de CrossRef | Google Académico
110. Abbas RZ, Munawar SH, Manzoor Z, Iqbal Z, Khan MN, Saleemi MK, et al. Efectos anticoccidiales del ácido acético sobre el rendimiento y los parámetros patogénicos en pollos de engorde desafiados con Eimeria tenella. Pesqui Vet Brasil. (2011) 31:99–103. doi: 10.1590/S0100-736X2011000200001
111. Bradley GL, Savage TF, Timm KI. Los efectos de complementar las dietas con Saccharomyces cerevisiae var. boulardii en el rendimiento del pollito masculino y la morfología ileal. Poult Sci. 73:1766–70. DOI: 10.3382/PS.0731766
Resumen de PubMed | Texto completo de CrossRef | Google Académico
112. Patterson JA, Burkholder KM. Aplicación de prebióticos y probióticos en la producción avícola. Poult Sci. (2003) 82:627–31. doi: 10.1093/ps/82.4.627
Resumen de PubMed | Texto completo de CrossRef | Google Académico
113. Liu S-Q, Tsao M. Mejora de la supervivencia de bacterias probióticas y no probióticas del ácido láctico por levaduras en leche fermentada en condiciones no refrigeradas. Int J Food Microbiol. (2009) 135:34–8. doi: 10.1016/j.ijfoodmicro.2009.07.017
Resumen de PubMed | Texto completo de CrossRef | Google Académico
114. Suharja AAS, Henriksson A, Liu S-Q. Impacto de Saccharomyces cerevisiae sobre la viabilidad del probiótico Lactobacillus rhamnosus en leche fermentada en condiciones ambientales. J Conserva Proc Alimentaria. (2014) 38:326–337. doi: 10.1111/j.1745-4549.2012.00780.x
115. Xie N, Zhou T, Li B. Las levaduras de kéfir aumentan los potenciales probióticos de Lactobacillus paracasei H9: los efectos positivos de la coagregación entre las dos cepas. Comida Res Int. (2012) 45:394–401. doi: 10.1016/j.foodres.2011.10.045
116. Roto SM, Rubinelli PM, Ricke SC. Una introducción a la microbiota intestinal aviar y los efectos de los compuestos de tipo prebiótico a base de levadura como posibles aditivos para piensos. Front Vet Sci. (2015) 2:28. DOI: 10.3389/FVETS.2015.00028
Resumen de PubMed | Texto completo de CrossRef | Google Académico
117. Foligné B, Dewulf J, Vandekerckove P, Pignède G, Pot B. Probiotic yeasts: anti-inflammatory potential of various non-pathogenic strains in experimental colitis in ratones. Mundo J Gastroenterol. (2010) 16:2134–45. doi: 10.3748/wjg.v16.i17.2134
Resumen de PubMed | Texto completo de CrossRef | Google Académico
118. Van Der Aa Kühle A, Skovgaard K, Jespersen L. Cribado in vitro de las propiedades probióticas de Saccharomyces cerevisiae var. boulardii y cepas de Saccharomyces cerevisiae transmitidas por los alimentos. Int J Food Microbiol. (2005) 101:29–39. doi: 10.1016/j.ijfoodmicro.2004.10.039
Resumen de PubMed | Texto completo de CrossRef | Google Académico
119. Czerucka D, Rampal P. Efectos experimentales de Saccharomyces boulardii en patógenos diarreicos. Microb Infectar. (2002) 4:733–9. doi: 10.1016/S1286-4579(02)01592-7
Resumen de PubMed | Texto completo de CrossRef | Google Académico
120. Czerucka D, Piche T, Rampal P. Artículo de revisión: levadura como probióticos–Saccharomyces boulardii. Aliment Pharmacol Ther. (2007) 26:767–78. doi: 10.1111/j.1365-2036.2007.03442.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
121. Zanello G, Meurens F, Berri M, Salmon H. Saccharomyces boulardii effects on gastrointestinal diseases. Curr emite mol biol. (2009) 11:47–8. DOI: 10.21775/CIMB.011.047
Resumen de PubMed | Texto completo de CrossRef | Google Académico
122. Geyik MF, Aldemir M, Hosoglu S, Ayaz C, Satilmis S, Buyukbayram H, et al. Los efectos de Saccharomyces boulardii en la translocación bacteriana en ratas con ictericia obstructiva. Ann R Coll Surg Engl. (2006) 88:176–80. doi: 10.1308/003588406X94986
Resumen de PubMed | Texto completo de CrossRef | Google Académico
123. Karen M, Yuksel O, Akyürek N, Ofluoglu E, Çaglar K, Sahin TT, et al. El agente probiótico Saccharomyces boulardii reduce la incidencia de lesión pulmonar en ratas inducidas por pancreatitis necrotizante aguda. J Surg Res. (2010) 160:139–44. doi: 10.1016/j.jss.2009.02.008
Resumen de PubMed | Texto completo de CrossRef | Google Académico
124. Line JE, Bailey JS, Cox NA, Stern NJ, Tompkins T. Effect of yeast-supplemented feed on Salmonella and Campylobacter populations in broilers. Poult Sci. (1998) 77:405–10. doi: 10.1093/ps/77.3.405
Resumen de PubMed | Texto completo de CrossRef | Google Académico
125. Dantán-González E, Quiroz-Castañeda RE, Cobaxin-Cárdenas M, Valle-Hernández J, Gama-Martínez Y, Tinoco-Valencia JR, et al. Impacto de Meyerozyma guilliermondii aislado de pollos contra Eimeria sp. protozoo, un análisis in vitro. BMC Vet Res. (2015) 11:278. DOI: 10.1186/S12917-015-0589-0
Resumen de PubMed | Texto completo de CrossRef | Google Académico
126. Lin G-J, Liu T-Y, Tseng Y-Y, Chen Z-W, You C-C, Hsuan S-L, et al. La glicoproteína E2 del virus de la peste porcina clásica expresada por levadura induce una respuesta inmune protectora. Veterinario Microbiol. (2009) 139:369–74. doi: 10.1016/j.vetmic.2009.06.027
Resumen de PubMed | Texto completo de CrossRef | Google Académico
127. Arnold M, Durairaj V, Mundt E, Schulze K, Breunig KD, Behrens S-E. Vacunación protectora contra el virus de la bursitis infecciosa con levadura Kluyveromyces lactis recombinante entera que expresa la subunidad VP2 viral. PLoS ONE. (2012) 7:E42870. doi: 10.1371/journal.pone.0042870
Resumen de PubMed | Texto completo de CrossRef | Google Académico
128. Sathish K, Sriraman R, Subramanian BM, Rao NH, Balaji K, Narasu ML, et al. La planta expresada EtMIC2 es un inmunógeno eficaz para conferir protección contra la coccidiosis del pollo. Vacuna. (2011) 29:9201–8. doi: 10.1016/j.vaccine.2011.09.117
Resumen de PubMed | Texto completo de CrossRef | Google Académico
129. Sun H, Wang L, Wang T, Zhang J, Liu Q, Chen P, et al. Exhibición de la proteína Eimeria tenella EtMic2 en la superficie de Saccharomyces cerevisiae como una potencial vacuna oral contra la coccidiosis del pollo. Vacuna. (2014) 32:1869–76. doi: 10.1016/j.vaccine.2014.01.068
Resumen de PubMed | Texto completo de CrossRef | Google Académico
130. Elaine-Rose M, Long PL. Inmunidad a la coccidiosis: efectos protectores del suero y las células transferidas investigadas en embriones de pollo infectados con Eimeria tenella. Parasitología. (2009) 63:299–313. doi: 10.1017/S0031182000079610
131. Shivaramaiah C, Barta JR, Hernandez-Velasco X, Téllez G, Hargis B. Coccidiosis: recent advances in the immunobiology of Eimeria species, preventive measures, and the importance of vaccination as a control tool against these Apicomplexan parasites. Veterinario Med. (2014) 5:23–34. doi: 10.2147/VMRR. S57839
132. Wallach M. Papel del anticuerpo en la inmunidad y el control de la coccidiosis del pollo. Tendencias Parasitol. (2010) 26:382–7. doi: 10.1016/j.pt.2010.04.004
Resumen de PubMed | Texto completo de CrossRef | Google Académico
133. Carver J. Nucleótidos dietéticos: efectos sobre los sistemas inmunológico y gastrointestinal. Acta Paediatr. (1999) 88:83–8. doi: 10.1111/j.1651-2227.1999.tb01306.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
134. Hess JR, Greenberg NA. El papel de los nucleótidos en los sistemas inmunológico y gastrointestinal. Nutr Clin Pract. (2012) 27:281–94. doi: 10.1177/0884533611434933
Resumen de PubMed | Texto completo de CrossRef | Google Académico
135. Jung B, Batal AB. Efecto de la suplementación con nucleótidos dietéticos sobre el rendimiento y el desarrollo del tracto gastrointestinal de pollos de engorde. Br Poult Sci. (2012) 53:98–105. doi: 10.1080/00071668.2012.659654
Resumen de PubMed | Texto completo de CrossRef | Google Académico
136. Alizadeh M, Rodriguez-Lecompte JC, Yitbarek A, Sharif S, Crow G, Slominski BA. Efecto de los productos derivados de la levadura en la respuesta inmune innata sistémica de pollos de engorde después de un desafío de lipopolisacáridos. Poult Sci. (2016) 95:2266–73. DOI: 10.3382/ps/pew154
Resumen de PubMed | Texto completo de CrossRef | Google Académico
137. Navarro J, Maldonado J, Narbona E, Ruiz-Bravo A, García Salmerón JL, Molina JA, et al. Influencia de los nucleótidos dietéticos en los niveles de inmunoglobulina plasmática y los subgrupos de linfocitos de los lactantes prematuros. BioFactores. (1999) 10:67–76. doi: 10.1002/biof.5520100108
Resumen de PubMed | Texto completo de CrossRef | Google Académico
138. Yu VYH. Justificación científica y beneficios de la suplementación con nucleótidos de la fórmula infantil. J Paediatr Salud Infantil. (2002) 38:543–49. doi: 10.1046/j.1440-1754.2002.00056.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
139. Salaheen S, Kim S-W, Haley BJ, Van Kessel JAS, Biswas D. Alternative growth promoters modulate broiler gut microbiome and enhance body weight gain. Microbiol frontal. (2017) 8:2088. DOI: 10.3389/fmicb.2017.02088
Resumen de PubMed | Texto completo de CrossRef | Google Académico
140. Holck P, Sletmoen M, Stokke BT, Permin H, Norn S. Potentiation of histamine release by microfungal (1 → 3)- and (1 → 6)-β-D-glucans. Clin Pharmacol Toxicol Básico. (2007) 101:455–8. doi: 10.1111/j.1742-7843.2007.00140.x
Resumen de PubMed | Texto completo de CrossRef | Google Académico
141. Volman JJ, Ramakers JD, Plat J. Modulación dietética de la función inmune por β-glucanos. Comportamiento fisiológico. (2008) 94:276–84. doi: 10.1016/j.physbeh.2007.11.045
Resumen de PubMed | Texto completo de CrossRef | Google Académico
142. Elmer GW, Mcfarland LV. Supresión por Saccharomyces boulardii del sobrecrecimiento toxigénico de Clostridium difficile después del tratamiento con vancomicina en hámsteres. Agentes antimicrobianos Chemother. (1987) 31:129. doi: 10.1128/AAC.31.1.129
Resumen de PubMed | Texto completo de CrossRef | Google Académico
143. Ghoneum M, Wang L, Agrawal S, Gollapudi S. Terapia de levadura para el tratamiento del cáncer de mama: un estudio modelo de ratones desnudos. In vivo. (2007) 21:251–8. Disponible en línea en: http://iv.iiarjournals.org/content/21/2/251.long
144. Xue G-D, Wu S-B, Choct M, Swick RA. Efectos de la pared celular de levadura en el rendimiento del crecimiento, las respuestas inmunes y las concentraciones intestinales de ácidos grasos de cadena corta de pollos de engorde en un modelo experimental de enteritis necrótica. Anim Nutr. (2017) 3:399–405. doi: 10.1016/j.aninu.2017.08.002
Resumen de PubMed | Texto completo de CrossRef | Google Académico
145. Hooge DM. Meta-analysis of broiler chicken pen trials evaluating dietary mannan oligosaccharide, 1993-2003. Int J Poult Sci. (2004) 3:163–74. doi: 10.3923/ijps.2004.163.174
146. Shashidhara R, Devegowda G. Effect of dietary mannan oligosaccharide on broiler breeder production traits and immunity. Poult Sci. (2003) 82:1319–25. DOI: 10.1093/PS/82.8.1319
Resumen de PubMed | Texto completo de CrossRef | Google Académico
147. M’sadeq SA, Wu S-B, Choct M, Forder R, Swick RA. Uso de extracto de pared celular de levadura como herramienta para reducir el impacto de la enteritis necrótica en pollos de engorde. Poult Sci. (2015) 94:898–905. doi: 10.3382/ps/pev035
Resumen de PubMed | Texto completo de CrossRef | Google Académico
148. Rose ME. Respuestas inmunes en infecciones con coccidios: actividad de macrófagos. Infectar inmune. (1974) 10:862–71.
149. Lu Z, Thanabalan A, Leung H, Akbari Moghaddam Kakhki R, Patterson R, Kiarie EG. Los efectos de alimentar con bioactivos de levadura a los reproductores de pollos de engorde y / o sus crías en el rendimiento del crecimiento, el desarrollo intestinal y la función inmune en pollos de engorde desafiaron con Eimeria. Poult Sci. (2019) 98:6411–21. doi: 10.3382/ps/pez479
Resumen de PubMed | Texto completo de CrossRef | Google Académico
150. Girard F, Fort G, Yvoré P, Quéré P. Cinética de la producción específica de inmunoglobulina A, M y G en la mucosa duodenal y cecal de pollos infectados con Eimeria acervulina o Eimeria tenella. Int J Parasitol. (1997) 27:803–9. doi: 10.1016/S0020-7519(97)00044-1
151. Yun CH, Lillehoj HS, Lillehoj EP. Respuestas inmunes intestinales a la coccidiosis. Dev Comp Immunol. (2000) 24:303–24. doi: 10.1016/S0145-305X(99)00080-4
Resumen de PubMed | Texto completo de CrossRef | Google Académico
152. Gómez-Verduzco G, Cortes-Cuevas A, López-Coello C, Avila-González E, Nava GM. La suplementación dietética de manano-oligosacárido mejora las respuestas inmunes neonatales en pollos durante la exposición natural a Eimeria spp. Acta Vet Scand. (2009) 51:11. DOI: 10.1186/1751-0147-51-11
Resumen de PubMed | Texto completo de CrossRef | Google Académico
153. Elmusharaf MA, Bautista V, Nollet L, Beynen AC. Effect of a mannanoligosaccharide preparation on Eimeria tenella infection in burler chickens. Int J Poult Sci. (2006) 5:583–8. doi: 10.3923/ijps.2006.583.588
154. Elmusharaf MA, Peek HW, Nollet L, Beynen AC. El efecto de una preparación de mananooligosacáridos (MOS) en el alimento en una infección por coccidiosis en pollos de engorde. Anim Feed Sci Technol. (2007) 134:347–54. doi: 10.1016/j.anifeedsci.2006.11.022
155. Shanmugasundaram R, Sifri M, Selvaraj RK. Efecto de la suplementación con productos de células de levadura (CitriStim) sobre el rendimiento de los pollos de engorde y los parámetros de las células inmunes intestinales durante una infección coccidial experimental1. Poult Sci. (2013) 92:358–63. DOI: 10.3382/PS.2012-02776
Resumen de PubMed | Texto completo de CrossRef | Google Académico
156. Lu HA, Adedokun S, Adeola LM, Ajuwon K. Anti-inflammatory effects of non-antibiotic alternatives in coccidia challenged broiler chickens. J Poult Sci. (2014) 51:14–21. doi: 10.2141/jpsa.0120176
157. Calder PC, Krauss-Etschmann S, De Jong EC, Dupont C, Frick JS, Frokiaer H, et al. Nutrición temprana e inmunidad: progreso y perspectivas. H. J Nutr. (2006) 96:774–90. doi: 10.1079/BJN20061917
158. Reynolds LP, Caton JS. Role of the pre- and post-natal environment in developmental programming of health and productivity. Mol Cell Endocrinol. (2012) 354:54–9. doi: 10.1016/j.mce.2011.11.013
159. Rudrappa SG, Humphrey BD. Energy metabolism in developing chicken lymphocytes is altered during the embryonic to posthatch transition. J Nutr. (2007) 137:427–32. doi: 10.1093/jn/137.2.427
160. Smith NC, Wallach M, Miller CM, Braun R, Eckert J. Maternal transmission of immunity to Eimeria maxima: western blot analysis of protective antibodies induced by infection. Infect Immun. (1994) 62:4811–7.
Palabras clave: Emeria, coccidiosis, enzimas alimenticias, salud y función intestinal, eficiencia alimenticia, derivados de levaduras
Cita: Kiarie EG, Leung H, Akbari Moghaddam Kakhki R, Patterson R y Barta JR (2019) Utilidad de las enzimas alimenticias y derivados de levadura para mejorar los efectos nocivos de la coccidiosis en la salud intestinal y la función en pollos de engorde. Frente. Vet. Sci. 6:473. DOI: 10.3389/fvets.2019.00473
Recibido: 26 Agosto 2019; Aprobado: 05 Diciembre 2019;
Publicado: 20 diciembre 2019.
Editado por:
Youssef A. Attia, Universidad Rey Abdulaziz, Arabia Saudita
Revisado por:
Woo Kyun Kim, Universidad de Georgia, Estados Unidos
Giovanni Piccolo, Universidad de Nápoles Federico II, Italia
Derechos de autor © 2019 Kiarie, Leung, Akbari Moghaddam Kakhki, Patterson y Barta. Este es un artículo de acceso abierto distribuido bajo los términos de la Licencia de Atribución Creative Commons (CC BY).
*Correspondencia: Elijah G. Kiarie, ekiarie@uoguelph.ca
†ORCID: Elijah Kiarie orcid.org/0000-0001-9752-8546
Reza Akbari Moghaddam Kakhki orcid.org/0000-0003-2621-0623
Renuncia: Todas las afirmaciones expresadas en este artículo son únicamente las de los autores y no representan necesariamente las de sus organizaciones afiliadas, o las del editor, los editores y los revisores. Cualquier producto que pueda ser evaluado en este artículo o reclamo que pueda ser hecho por su fabricante no está garantizado ni respaldado por el editor.
Date de alta y recibe nuestro 👉🏼 Diario Digital AXÓN INFORMAVET ONE HEALTH
Date de alta y recibe nuestro 👉🏼 Boletín Digital de Foro Agro Ganadero
Noticias animales de compañía
Noticias animales de producción
Trabajos técnicos animales de producción
Trabajos técnicos animales de compañía